一种新型阿魏酸酯酶及其制备方法和应用与流程
本发明属于生物工程领域,包括一种新型阿魏酸酯酶及其制备方法和应用,具体涉及到阿魏酸酯酶的宏基因筛选技术、新型阿魏酸酯酶基因的重组、表达,阿魏酸酯酶的制备及其应用。
背景技术:
阿魏酸(ferulicacid),化学名称3-甲氧基-4-羟基肉桂酸,具有抗氧化、抗菌消炎、抗癌、抗血栓、抗动脉粥样硬化等良好的生理功能,目前阿魏酸已广泛应用于医药和化妆品等行业,其中反式阿魏酸在美国、日本已被允许作为食品添加剂。阿魏酸广泛存在于食品原料中,如麦麸、玉米芯、甜菜渣、啤酒糟等,但阿魏酸大多以单体或者聚体的形式结合在植物细胞壁的木质素和多糖之间。
阿魏酸酯酶(feruloylesterase,fae,ec3.1.1.73),别名肉桂酸酯酶,属于水解酶家族中的羧酸酯酶亚类,可将阿魏酸类酯、低聚糖阿魏酸酯和多糖阿魏酸酯之间的酯键水解并释放出阿魏酸。阿魏酸酯酶来源广泛,存在于植物、真菌和细菌中。大多数分泌的阿魏酸酯酶来源于微生物,已发现的菌株有黑曲霉(aspergillusniger),米曲霉(aspergillusoryzae),梭菌(clostridiumthermocellum)、乳酸杆菌(lactobacilli)等。研究表明阿魏酸酯酶与其他水解酶(纤维素酶、木聚糖酶等)之间存在协同作用,能提高植物细胞壁降解率和木质纤维素的糖化效率。
现阶段阿魏酸酯酶的研究方法有发酵提取法和异源表达法。前者是以可溶性的羟基肉桂酸酯类化合物(如阿魏酸乙酯等)或者富含阿魏酸的农业原料(如甜菜浆、燕麦木聚糖)为底物和酶诱导剂,筛选产阿魏酸酯酶的活性菌株并从该菌株的发酵产物中得到阿魏酸酯酶,然而,这种阿魏酸酯酶活性响应度低,酶活通常小于1u/ml,极大限制了该类酶在工业中的广泛应用。后者是通过分子克隆的方法,重组阿魏酸酯酶基因使其能够在合适的异源宿主中表达(如大肠杆菌、毕赤酵母、酿酒酵母、丝状真菌等),经分离纯化得到重组酶。两种方法相比,异源表达法能够明显提高酶的产量和酶的活性。
然而有研究指出,环境中有99%以上的微生物是难以利用实验室常规微生物培养方式获得。随着生物信息学和分子生物学的发展,宏基因组学逐渐兴起,这种科学技术可广泛应用于不可培养微生物基因簇的发掘,有效绕开微生物传统分离培养的环节。宏基因组学技术不依赖于传统分离培养,直接提取环境中所有微生物的基因组dna,克隆到载体并转化至可培养的宿主,构建宏基因组文库。文库包含了样本中可培养和不可培养微生物的全部核酸信息,利用序列筛选或功能筛选的方式可从文库中获得有用的目标产物,如酶、抗生素和活性物质等。宏基因组技术已成功地从荷斯坦牛瘤胃、污泥渗滤液、白蚁肠道菌群及土壤等不同样本中鉴定了新型的阿魏酸酯酶,这充分说明宏基因组学是挖掘和发现新型阿魏酸酯酶的一种有力工具。进一步,生物信息学技术的发展为提高酯酶的性能提供了更多的可能,如同源建模来进行虚拟筛选,设计定点诱变实验或理性的改造氨基酸序列。
技术实现要素:
本发明基于土壤宏基因组策略筛选技术得到一种新型阿魏酸酯酶基因,并对该基因克隆及异源表达。重组酶具有较高的酶活,对大多数金属离子和有机溶剂有耐受性等特点,使其在食品、制药与饲料等领域中具有应用潜力。
本发明采取的技术方案如下:
本发明的第一个目的是提供一种新型阿魏酸酯酶bds4,所述阿魏酸酯酶bds4的氨基酸序列如seqidno.2所示:
mpyisteakkilalmsesgapefgapplsvarqvyaglgsklggeviemasvedlsmagpgstlpmriyrplensgqngaliyfhgggwilggiethdrlcrqiamrsscvvisigyrlapehplpaaaddaiaavrwvvdkadflrisgalavggdsaggglaayaalaardeqlpvraqvliypsvdnresaheryasrktnaevppltvqamrnvaaylahekalnedirlspilgvqsrtsvpaliltagadvlrdeglrygcelldagasvlirnypgaihgflempaalqvaqdaheliglflrqqliqkp。
进一步的,所述阿魏酸酯酶bds4的三级结构为具有一个典型的α/β水解酶折叠,由帽子结构域和催化结构域组成,所述新型阿魏酸酯酶bds4的催化三联体位于结合口袋,催化三联体由丝氨酸(s158),天冬氨酸(d256)和组氨酸(h286)组成。
进一步的,是本发明新型阿魏酸酯酶bds4酶催化三联体结构的确定方法为:假定的催化三联体通过多序列比对分析及三维结构模拟得到,并通过定点突变技术确认。
具体的:确定丝氨酸(s158),天冬氨酸(d256)和组氨酸(h286)作为突变位点,构建相应的突变子;依据各自的突变位点设计定点突变引物,以pet-bds4为扩增模板,进行全质粒pcr,将突变引入目的片段中,挑取克隆子测序,经序列分析及活性验证确认突变位点;所述定点突变引物为:
s-fv:5'-cacctgcacactcaccgccaacggctagggcgcccgag-3'(seqidno.5)
s-rv:5'-gcggtgactgtgcaggtggagggttggctgcgtatgct-3'(seqidno.6)
d-fv:5'-ggagctaacgtgttgagggatgaaggattgcgctatgg-3'(seqidno.7)
d-rv:5'-tcaacacgttagctcccgcggtcagtattaaggcag-3'(seqidno.8)
h-fv:5'-ctggtgcaattaagggtttcctcgagatgccagctgccc-3'(seqidno.9)
h-rv:5'-gaaacccttaattgcaccaggatagtttctaattaaaaccga-3'
(seqidno.10)。
本发明的第二个目的是提供编码前述新型阿魏酸酯酶bds4的基因bds4,所述基因bds4的核苷酸序列如seqidno.1所示:
ctatggcttttgaattaattgctgccgaagaaaaagcccaatcaattcgtgcgcatcttgtgcgacttgcagggcagctggcatctcgaggaaaccatgaattgcaccaggatagtttctaattaaaaccgaggcacccgcatcgagcaattcgcaaccatagcgcaatccttcatccctcaacacgtcagctcccgcggtcagtattaaggcaggcaccgatgtccggctttgcacaccaagaataggcgacaggcgaatatcttcgtttaatgctttctcatgcgctagatatgcggcaacgttgcgcattgcctgcaccgtaagaggaggcacttctgcattcgtttttcgtgatgcatatcgctcgtgtgccgattcccggttatcaacgctaggatagattaggacctgagcgcggacgggcaattgttcgtcacgagcagcgagcgcagcatacgcagccaaccctccacctgcactgtcaccgccaacggctagggcgcccgagattcgcaggaaatcagctttgtccacaacccaacgcaccgcagcgatcgcatcgtctgcagccgctggcagcgggtgctcaggcgcaagcctgtaaccgatagagataactacgcaagaggatctcattgcgatttgtcgacagagtctgtcatgagtctctatgccccccagaatccacccgccaccatgaaaataaattaaagctccattttgaccagagttttcaagggggcgatagatccgcatcggcaaagtactacccggcccagccatcgaaagatcctcaacagaggccatctcgataacctccccgcccaactttgagccaaggcctgcatagacctggcgcgcgaccgatagcggcggagctccgaattctggcgcgccgctttcactcatgagagcgagtattttttttgcttcggtggaaatatatggcat。
进一步的,seqidno.1所述的核苷酸序列存在于可培养微生物中和/或不可培养微生物中。
本发明的第三个目的是提供前述编码新型阿魏酸酯酶的基因bds4的宏基因组学功能性筛选方法,包括以下步骤:
s1:提取及纯化土壤总dna,构建fosmid宏基因组文库;
s2:功能性筛选阳性克隆子;
s3:采取亚克隆策略及生物信息学分析获得编码阿魏酸酯酶的基因bds4。
进一步的,所述s1具体步骤为:采用ctab抽提法提取土壤总dna(样品来自表层土壤5-10厘米),利用电泳纯化土壤总dna,利用fosmid(epicenter,usa)试剂盒构建宏基因组文库;按照试剂盒说明,土壤dna经末端修复酶修复后与pcc2fos载体连接,经λ-噬菌体包装后侵染大肠杆菌e.coliepi300,构建文库;将文库以大约5000个克隆子为单位分装,加入15%甘油贮藏,以备筛选;
所述s2具体步骤为:以阿魏酸乙酯作为筛选底物,取适量稀释后的文库菌液进行涂布,经培养后,观察筛选平板上透明圈的产生情况;挑取产透明圈的克隆子,将其接种于含有阿魏酸甲酯的lb液体培养基中过夜摇菌,取部分发酵液上清进行hplc分析确认阿魏酸酯酶活性;
所述s3具体步骤为:提取阳性克隆子质粒,利用sau3ai进行部分酶切,电泳后回收1-5kb大小的dna片段,将其连接至具有相同酶切位点的puc118载体上并转化至大肠杆菌dh5α;使用相同的底物平板筛选法筛选阳性亚克隆,经测序及orffinder在线分析工具预测该酶的开放阅读框;经分析后确认筛选得到seqidno.1所述的阿魏酸酯酶编码基因bds4。
本发明的第四个目的是提供一种重组阿魏酸酯酶bds4的制备方法,采用特异性引物对,扩增权利要求3所述的阿魏酸酯酶基因bds4,所述引物对包括以下两条序列:
上游引物hfv/bamhi:5'-gctggatccatgccatatatttccacc-3'(seqidno.3)
下游引物hrv/hindiii:5'-gctaagctttggcttttgaattaattg-3'(seqidno.4)
其具体步骤为:利用seqidno.3、seqidno.4所述的特异性引物对扩增seqidno.1所述的阿魏酸酯酶基因bds4,pcr扩增产物经bamhi和hindiii双酶切,与相同酶切线性化的表达载体pet-28a(+)连接,构建得到前述的重组表达载体pet-bds4;将重组表达载体pet-bds4转化至宿主菌种大肠杆菌bl21(de3),培养转化子,经iptg诱导,从培养物中分离,镍柱纯化得到所述重组阿魏酸酯酶bds4。
本发明的第五个目的是提供前述新型阿魏酸酯酶bds4在塑化剂降解中的应用。
进一步的,所述塑化剂包括邻苯二甲酸二甲酯、邻苯二甲酸二乙酯、邻苯二甲酸二丁酯等。
本发明的第六个目的是提供前述新型阿魏酸酯酶bds4在降解麦麸中的应用,在木聚糖酶的存在下,阿魏酸酯酶bds4可明显提高从去淀粉麦麸中释放阿魏酸的量。阿魏酸酯酶与木聚糖酶联合作用于植物(如麦麸等),可提高植物木质纤维素的水解率并释放出高价值的阿魏酸。
本发明的第七个目的是提供前述新型阿魏酸酯酶bds4在食品工业、制药工业、造纸工业和饲料工业中的应用。
本发明的有益效果:
本发明提供了一种来源于土壤宏基因文库的阿魏酸酯酶基因,其核苷酸序列和氨基酸序列如seqidno.1和seqidno.2所示。将该酯酶基因插入到质粒pet-28a(+)后,转化至大肠杆菌bl21(de3)中实现异源表达。
纯化后的重组酶(bds4)分子量为38.8kda。bds4可以水解阿魏酸甲酯、阿魏酸乙酯、咖啡酸甲酯、芥子酸甲酯、香草酸甲酯,其中对阿魏酸甲酯的催化能力最强(酶活为57.05u/mg)。对bds4的酶学性质研究表明该酶的最适反应ph8.0,最适反应温度37℃,能耐受大部分的金属离子和有机溶剂。根据氨基酸序列与底物偏好性分析,将bds4鉴定为c型阿魏酸酯酶。bds4还可以水解邻苯二甲酸二甲酯,邻苯二甲酸二乙酯,邻苯二甲酸二丁酯这三种塑化剂。定点诱变实验表明bds4的催化三联体由丝氨酸(s158),天冬氨酸(d256)和组氨酸(h286)组成,三者中任何氨基酸的突变都会导致bds4失去催化能力。
在木聚糖酶的存在下,bds4可明显提高从去淀粉麦麸中释放阿魏酸的量。本发明的新型阿魏酸酯酶,由于其特有的活性及酶学特性,使其可应用于饲料、造纸、食品、制药等领域。
附图说明
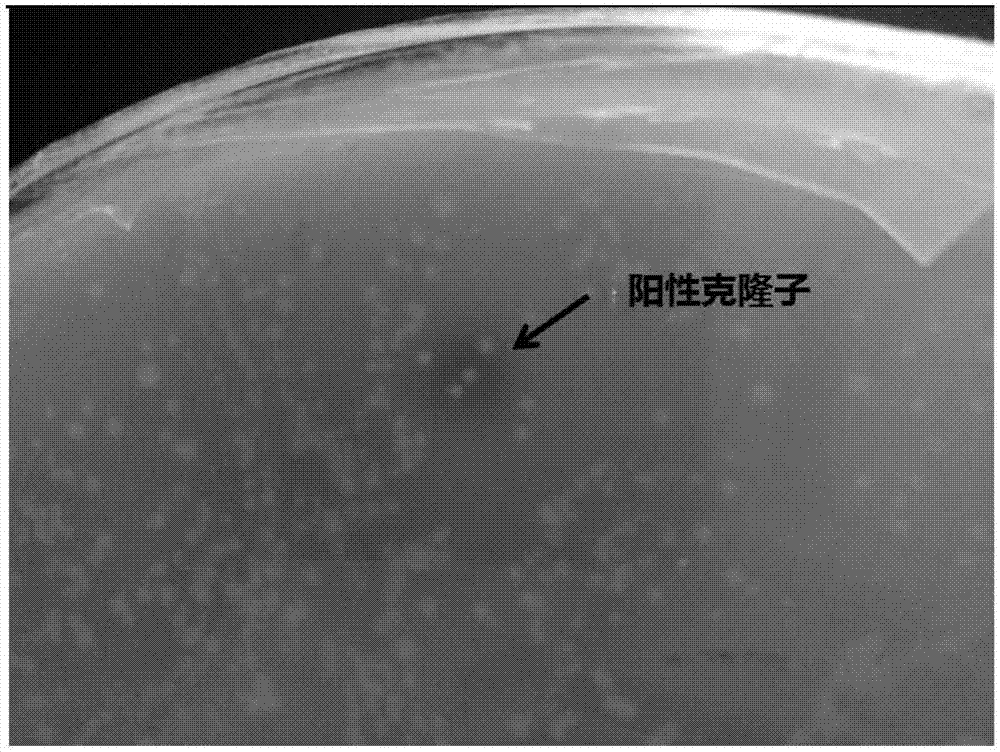
图1.实施例1中土壤宏基因文库中产阿魏酸酯酶的平板初筛。
图2.实施例1中阳性克隆子发酵液高效液相色谱图(a:阿魏酸对照分析品;b:克隆子发酵液)。
图3.实施例2中阿魏酸酯酶的sds-page蛋白分析图谱。
m:蛋白marker;1:粗酶液;2:纯化后的bds4。
图4.实施例3中阿魏酸酯酶最适ph及ph稳定性与最适温度及热稳定性折线图(a,ph对bds4活性的影响;b,bds4的ph稳定性;c,温度对bds4活性的影响;d,bds4的温度热稳定性)。
图5.实施例3中阿魏酸酯酶bds4的系统发育树。
图6.实施例4中阿魏酸酯酶bds4的多序列比对。
图7.实施例4中阿魏酸酯酶bds4的三维结构模拟图。
图8.实施例5中阿魏酸酯酶bds4降解塑化剂的gc-ms分析图(a,由上而下分别为邻苯二甲酸二甲酯降解前和降解后的gc-ms分析图及降解途径;b,由上而下分别为邻苯二甲酸二乙酯降解前和降解后的gc-ms分析图及降解途径;c,由上而下分别为邻苯二甲酸二丁酯降解前和降解后的gc-ms分析图及降解途径)。
图9.实施例6中阿魏酸酯酶bds4降解麦麸柱状图(a,未经处理;b,麦麸经100u的木聚糖酶处理;c,麦麸经20u的bds4处理;d,麦麸经20u的bds4和100u的木聚糖酶共同处理)。
具体实施方式
以下结合具体实施例来进一步阐述本发明。但是这些实施例仅用于详细说明本发明,而不能限制本发明的保护范围。
实施例1:土壤宏基因组文库的构建、阳性克隆子的筛选及相应阿魏酸酯酶基因的鉴定
土壤宏基因组文库的构建:称取土壤样品10g,加入ctab抽提液,37℃振摇混匀培养并振荡混匀培养45min。组分中加入适量溶菌酶和蛋白酶k,裂解细胞及除去蛋白。加入2.5ml20%sds,65℃条件下水浴2h,再加入预冷的氯仿3ml,混匀后离心收集上清。加入等体积预冷的苯酚:氯仿:异戊醇(25:24:1)溶液,颠倒混匀后离心,取上层水相加入等体积的24:1(氯仿:异戊醇),再次离心,取水相加入0.6倍体积的预冷异丙醇,室温水浴醇沉1h,离心收集沉淀。最后用70%乙醇清洗沉淀2次,吹干,te缓冲液溶解粗dna。将提取的粗dna使用1%的琼脂糖凝胶(不含eb),25v电泳11h。割下大小在30-40kb胶条,放入装有1×tae的透析袋中,100v电泳7-8h,收集洗脱液超滤浓缩,异丙醇醇沉,用少量tris-hcl重溶dna并测浓度。依据copycontroltmfosmidlibraryproductionkit构建土壤宏基因文库。将获得的dna片段经过末端修复变成钝末端,然后经过5'-磷酸化、大小筛选,从低融点琼脂糖凝胶回收。将大小选定的dna连接到pcc2fos载体上。用试剂盒附带的高效maxplaxtmλ噬菌体包装蛋白进行包装,接种到大肠杆菌epi300细胞中。菌液离心浓缩均匀涂布在氯霉素抗性的平板上,倒置过夜培养,计算克隆子,并以每管约5000种克隆子为一个亚库,加入15%甘油贮藏,-80℃保种。
阳性克隆子的筛选:利用平板法和高效液相色谱法筛选阳性克隆子。制备阿魏酸酯酶筛选平板。向lb常规培养基中添加阿魏酸乙酯(终浓度为1.5mg/ml),氯霉素(终浓度为12.5μg/ml)、iptg诱导剂(0.5mm),摇匀使得阿魏酸乙酯分布均匀,在超净台倒平板。每块筛选平板上涂布已活化的fosmid文库菌液,37℃培养1天,观察菌落周围透明圈的形成(图1)。
初筛后将产透明圈的克隆子接种至lb液体培养基(阿魏酸甲酯1mg/ml,氯霉素12.5μg/ml,iptg0.5mm)进行发酵,发酵条件为37℃,24h,转速为180r/min。发酵完成后,取过膜后的发酵液进行高效液相色谱检测。色谱条件如下:采用zorbaxsb-c18色谱柱,紫外检测波长320nm,梯度洗脱,流动相为有机相(甲醇)和水相(1%冰醋酸),柱温30℃,进样量20μl,流速为1ml/min。发酵液中检测到降解产物阿魏酸,表明已筛选到具有阿魏酸酯酶活性的阳性克隆子(图2)。
提取阳性克隆子质粒,利用稀释1000倍的sau3ai对阳性克隆子质粒进行部分酶切,酶切体系酶切体系(100μl)为:sau3ai10μl,10×hbuffer10μl,0.1%bsa10μl,dna5μg,反应时间10min。将酶切产物电泳,割胶回收1-5kb大小的dna片段。将回收片段和具有相同酶切位点的puc118载体按照10:1的摩尔比例混合,加入t4dna连接酶,16℃过夜连接。将10μl连接产物加入100μl大肠杆菌dh5α感受态细胞,冰上孵育30min,42℃水浴热激60s,后加入900μllb液体培养基孵育1h(37℃,150rpm)。将菌液离心(4000×g,1min),去掉上清液800ul,剩余培养基将菌体进行重悬,涂布于lb平板(阿魏酸乙酯1.5mg/ml,氨苄100μg/ml,iptg0.5mm),37℃过夜培养,产透明圈的转化子即为含有阿魏酸酯酶的阳性克隆子。对其进行测序,并利用orffinder在线分析工具预测该酶的开放阅读框,进一步分析后确认筛选得到所述的阿魏酸酯酶编码基因bds4。
实施例2:阿魏酸酯酶基因的分子克隆及表达纯化
阿魏酸酯酶基因的分子克隆:利用以下引物扩增阿魏酸酯酶基因bds4hfv/bamhi
5'-gctggatccatgccatatatttccacc-3'(seqidno.3)和hrv/hindiii
5'-gctaagctttggcttttgaattaattg-3'(seqidno.4)(bamhi和hindiii的酶切位点用下划线标注)。pcr反应体系(50μl):超纯水20μl,2×taqmastermix25μl,上下游引物((10μm)各2μl,dna模板1μl。pcr反应条件:95℃5min;95℃5min,56℃30s,72℃1min,35个循环;72℃10min。将pcr产物电泳,割胶回收,得到纯化的阿魏酸酯酶基因pcr产物。pcr产物进行双酶切,酶切时间3h。酶切体系(100μl)为:bamhi5μl,hindiii5μl,10×kbuffer10μl,0.1%bsa10μl,dna5μg,无菌水。割胶回收得到纯化的酶切片段。pet-28a(+)质粒进行双酶切,酶切时间3h。酶切体系(100μl)为:bamhi5μl,hindiii5μl,10×kbuffer10μl,0.1%bsa10μl,dna5μg,无菌水。割胶回收得到纯化的酶切载体。将经过双酶切的dna片段和pet-28a(+)载体按照10:1的摩尔比例混合,加入t4dna连接酶,16℃过夜连接。将10μl连接产物加入100μl大肠杆菌bl21(de3)感受态细胞,冰上孵育30min,42℃水浴热激60s,后加入900μllb液体培养基孵育1h(37℃,150rpm)。将菌液离心(4000×g,1min),去掉上清液800ul,剩余培养基将菌体进行重悬,涂布于lb平板(阿魏酸乙酯1.5mg/ml,卡那霉素50μg/ml,iptg0.5mm),37℃培养1天,产透明圈的转化子即为重组成功的表达菌株。对其进行测序,测序结果显示与bds4的核苷酸序列(如seqidno.1所示)一致,并将已整合bds4基因的pet-28a(+)质粒命名为pet-bds4。
阿魏酸酯酶基因的表达纯化:将重组表达菌株,接种至lb液体培养基(阿魏酸甲酯1mg/ml,卡那霉素50μg/ml,iptg0.5mm)中,37℃过夜培养。取菌液,以1%接种量加入到150ml的lb液体培养基中(阿魏酸甲酯1mg/ml,卡那霉素50μg/ml,iptg0.5mm),于37℃培养24h。离心收集发酵后的菌液。根据ni-nta纯化树脂预装柱(sangon,china)的使用说明处理大肠杆菌菌体,步骤如下:缓冲液重悬,超声破碎,留上清液作为粗酶液,将粗酶液过ni-nta纯化树脂预装柱进行洗脱纯化。洗脱后的蛋白通过聚丙烯酰胺凝胶电泳进行检测分析(图3,图中m:蛋白marker;1:粗酶液;2:纯化后的bds4。)。
实施例3:阿魏酸酯酶bds4的酶学性质表征
酶活测定:取1mlph8.0的tris-hcl缓冲液,分别加入5μmol的阿魏酸甲酯溶液,加入20μg酶液(约10μl),37℃下反应,利用hplc测定320nm下的吸光度值。
酶活定义:37℃、ph8.0的反应条件下,每分钟降解阿魏酸甲酯生成1μmol阿魏酸所需酶量定义为1个酶活力单位(u)。
最适ph及ph稳定性分析:在37℃条件下,测定不同ph(3.0-11.0)条件下的酶活力,根据酶活大小确定酶的最适ph(图4a)。将酶液分别加到不同ph的缓冲液中,4℃放置1h,测定剩余酶活,确定酶的稳定性(图4b)。
最适温度及热稳定性:在ph8.0条件下,测定不同温度(4℃、16℃、25℃、30℃、37℃、45℃、55℃、65℃、75℃)条件下的酶活力,确定该酶发挥催化活性的最适温度(图4c)。将酶液分别放置于37℃、45℃、55℃水浴中4h,期间每隔1h测定残余酶活,确定该酶的热稳定性(图4d)。
底物特异性及酶催化动力学分析:基于酶活测定方法及最适条件下,比较该酶对不同底物的水解能力,包括阿魏酸乙酯(efa)、阿魏酸甲酯(mfa)、咖啡酸甲酯(mca)、对香豆素甲酯(mpca)、香草酸甲酯(mv)、芥子酸甲酯(msa),并计算其相关动力学参数。bds4对底物的催化能力:mfa>efa>mpca>mca>mv>msa(表1)。
表1bds4的底物特异性及相关动力学参数
阿魏酸酯酶bds4活性对金属离子和有机溶剂的耐受性:反应体系内分别加入5mm的k+、mn2+、ca2+、fe2+、fe3+、co2+、cu2+、na+、zn2+、mg2+,1%的丙酮(acetone)、甲醇(methylalcohol)、二甲亚砜(dimethylsulfoxide)、曲拉通(triton-100)、乙腈(acetonitrile)、二甲基甲酰胺(dimethylformamide)、β-巯基乙醇(β-mercaptoethanol)、乙二胺四乙酸二钠(edta-2na)、十二烷基硫酸钠(sds),在最适条件下测定酶活,以不加任何试剂的反应体系作为对照(表2)。除了cu2+、zn2+、β-mercaptoethanol、sds外,该酶对其他的有机溶剂均表现出良好的耐受性。
表2金属离子和有机溶剂对阿魏酸酯酶bds4活性的影响
系统发育树分析:将bds4及其他已知的阿魏酸酯酶通过mega6.0软件构建系统发育树。阿魏酸酯酶分为类型a、b、c、d,bds4与c型阿魏酸酯酶位于同一个进化分支上,表明bds4归于c型阿魏酸酯酶家族(图5)。
实施例4:阿魏酸酯酶bds4的生物信息学分析及催化三联体验证
多序列比对分析:利用在线工具clustalomega(https://www.ebi.ac.uk/tools/msa/clustalo/)对bds4及其同源序列进行多序列比对。
如图6所示,表明bds4含有一个g-d-s-a-g基序,和其他阿魏酸酯酶的保守五肽基序g-x-s-x-g具有一致性。其中,亲核氨基酸s158位于五肽基序中,且与高度保守的d256和h286构成了阿魏酸酯酶中的催化三联体。
三级结构预测:利用在线工具coach网站进行阿魏酸酯酶三维结构模拟。以4re7为模板进行同源建模,预测bds4三级结构。如图7所示,bds4具有一个典型的α/β水解酶折叠,由帽子结构域和催化结构域组成,推测的催化三联体位于结合口袋。
催化三联体验证:设计特异性引物,确定s158、d256和h286作为突变位点,构建相应的突变子。以pet-bds4为扩增模板,进行全质粒pcr。依据各自的突变位点设计特异性引物,通过引物将突变引入目的片段中。引物序列如表3所示,标有下划线部分为互补区域,粗体的三个核苷酸即为编码突变后的氨基酸。定点突变pcr反应体系(50μl):超纯水20μl,
2×high-fidelitymastermix25μl,上下游引物(10μm)各2μl,dna模板1μl。pcr反应程序:98℃2min;98℃10s,58℃10s,72℃1min,10个循环;98℃10s,58℃1min,72℃2min,10个循环;98℃10s,58℃10s,72℃3min,10个循环;72℃10min。将pcr产物电泳,割胶回收目标片段,测定浓度。按照试剂盒说明,构建突变质粒,对线性化pcr产物进行重组,构建突变质粒。其中,连接体系(10μl):线性载体20ng,2×sosoomix5μl,ddh2o。线性载体重组反应条件:50℃反应15min。将连接好的10μl突变质粒加入至100μl大肠杆菌bl21(de3)感受态细胞中,冰上孵育30min,42℃水浴热激60s,后加入900μllb液体培养基孵育1h(37℃,150rpm)。将菌液离心(4000×g,1min),去掉上清液800ul,剩余培养基将菌体进行重悬,涂布于lb平板(阿魏酸乙酯1.5mg/ml,卡那霉素50μg/ml,iptg0.5mm),37℃培养1天,观察有无透明圈产生。挑取克隆子测序,经序列分析及活性验证确认突变位点。
表3:pet-bds4的定点突变引物
实施例5:阿魏酸酯酶bds4对塑化剂的降解反应
实验分为邻苯二甲酸二甲酯组;邻苯二甲酸二乙酯组;邻苯二甲酸二丁酯组,各组分别称取塑化剂邻苯二甲酸二甲酯、邻苯二甲酸二乙酯、邻苯二甲酸二丁酯0.2g,用甲醇作溶剂配制0.2g/ml标准储存液(密封,放至4℃冰箱中保存)。
取标准储存液5ul加入反应体系中,使得塑化剂的终浓度为1mg/ml,加入阿魏酸酯酶bds4酶量为40μg/ml,反应体系1ml,于37℃反应12h。用氮气吹干反应液,并用甲醇复溶。
利用gc-ms联用仪检测反应液中塑化剂降解情况。色谱联用仪gc7890/5975msd条件:色谱柱hp-5;流速1ml/min;进样口温度280℃;载气氦气;进样量1μl;分流方式20:1;溶剂延迟7min。柱温采取程序升温:60℃维持1min;以10℃/min升温至180℃,维持10min;以15℃升温至220摄氏度,维持5min。后运行70℃,维持1min。
如图8所示,图8a为塑化剂邻苯二甲酸二甲酯降解前和降解后的gc-ms分析图及降解途径、图8b为邻苯二甲酸二乙酯降解前和降解后的gc-ms分析图及降解途径、图8c为邻苯二甲酸二丁酯降解前和降解后的gc-ms分析图及降解途径,结果表明,塑化剂邻苯二甲酸二甲酯、邻苯二甲酸二乙酯、邻苯二甲酸二丁酯均可被阿魏酸酯酶bds4降解为邻苯二甲酸,bds4具有用于降解塑化剂的潜力。
实施例6:阿魏酸酯酶bds4在降解麦麸方面的应用
将麦麸粉碎后浸没在0.25%的乙酸钾溶液中,用90℃高温水浴20min,用热水反复洗涤后尽可能去除淀粉,烘干至恒重,过筛获取去淀粉麦麸。
将实验分为:未经处理组;麦麸经100u的木聚糖酶处理组;麦麸经20u的bds4处理组;麦麸经20u的bds4和100u的木聚糖酶共同处理组。
各组称取去淀粉麦麸200mg,往tris-hcl缓冲液加入反应中的酶(bds4、木聚糖酶),反应体系10ml,37℃下水浴反应10h,每过1小时进行颠倒混匀一次。将反应液以4000×g离心5min,取上清过膜,利用hplc检测阿魏酸。
结果如图9所示,重组酶bds4与木聚糖酶协同作用于去淀粉麦麸时,阿魏酸释放量明显增加,即在木聚糖酶的存在下,阿魏酸酯酶bds4能够明显提高从去淀粉麦麸中释放阿魏酸的量。
序列表
<110>南京农业大学
<120>一种新型阿魏酸酯酶及其制备方法和应用
<160>10
<170>siposequencelisting1.0
<210>1
<211>954
<212>dna
<213>阿魏酸酯酶(feruloylesterase)
<400>1
ctatggcttttgaattaattgctgccgaagaaaaagcccaatcaattcgtgcgcatcttg60
tgcgacttgcagggcagctggcatctcgaggaaaccatgaattgcaccaggatagtttct120
aattaaaaccgaggcacccgcatcgagcaattcgcaaccatagcgcaatccttcatccct180
caacacgtcagctcccgcggtcagtattaaggcaggcaccgatgtccggctttgcacacc240
aagaataggcgacaggcgaatatcttcgtttaatgctttctcatgcgctagatatgcggc300
aacgttgcgcattgcctgcaccgtaagaggaggcacttctgcattcgtttttcgtgatgc360
atatcgctcgtgtgccgattcccggttatcaacgctaggatagattaggacctgagcgcg420
gacgggcaattgttcgtcacgagcagcgagcgcagcatacgcagccaaccctccacctgc480
actgtcaccgccaacggctagggcgcccgagattcgcaggaaatcagctttgtccacaac540
ccaacgcaccgcagcgatcgcatcgtctgcagccgctggcagcgggtgctcaggcgcaag600
cctgtaaccgatagagataactacgcaagaggatctcattgcgatttgtcgacagagtct660
gtcatgagtctctatgccccccagaatccacccgccaccatgaaaataaattaaagctcc720
attttgaccagagttttcaagggggcgatagatccgcatcggcaaagtactacccggccc780
agccatcgaaagatcctcaacagaggccatctcgataacctccccgcccaactttgagcc840
aaggcctgcatagacctggcgcgcgaccgatagcggcggagctccgaattctggcgcgcc900
gctttcactcatgagagcgagtattttttttgcttcggtggaaatatatggcat954
<210>2
<211>317
<212>prt
<213>阿魏酸酯酶(feruloylesterase)
<400>2
metprotyrileserthrglualalyslysileleualaleumetser
151015
gluserglyalaproglupheglyalaproproleuservalalaarg
202530
glnvaltyralaglyleuglyserlysleuglyglygluvalileglu
354045
metalaservalgluaspleusermetalaglyproglyserthrleu
505560
prometargiletyrargproleugluasnserglyglnasnglyala
65707580
leuiletyrphehisglyglyglytrpileleuglyglyilegluthr
859095
hisaspargleucysargglnilealametargsersercysvalval
100105110
ileserileglytyrargleualaprogluhisproleuproalaala
115120125
alaaspaspalailealaalavalargtrpvalvalasplysalaasp
130135140
pheleuargileserglyalaleualavalglyglyaspseralagly
145150155160
glyglyleualaalatyralaalaleualaalaargaspgluglnleu
165170175
provalargalaglnvalleuiletyrproservalaspasnargglu
180185190
seralahisgluargtyralaserarglysthrasnalagluvalpro
195200205
proleuthrvalglnalametargasnvalalaalatyrleualahis
210215220
glulysalaleuasngluaspileargleuserproileleuglyval
225230235240
glnserargthrservalproalaleuileleuthralaglyalaasp
245250255
valleuargaspgluglyleuargtyrglycysgluleuleuaspala
260265270
glyalaservalleuileargasntyrproglyalailehisglyphe
275280285
leuglumetproalaalaleuglnvalalaglnaspalahisgluleu
290295300
ileglyleupheleuargglnglnleuileglnlyspro
305310315
<210>3
<211>27
<212>dna
<213>人工序列(artificialsequence)
<400>3
gctggatccatgccatatatttccacc27
<210>4
<211>27
<212>dna
<213>人工序列(artificialsequence)
<400>4
gctaagctttggcttttgaattaattg27
<210>5
<211>38
<212>dna
<213>人工序列(artificialsequence)
<400>5
cacctgcacactcaccgccaacggctagggcgcccgag38
<210>6
<211>38
<212>dna
<213>人工序列(artificialsequence)
<400>6
gcggtgactgtgcaggtggagggttggctgcgtatgct38
<210>7
<211>38
<212>dna
<213>人工序列(artificialsequence)
<400>7
ggagctaacgtgttgagggatgaaggattgcgctatgg38
<210>8
<211>36
<212>dna
<213>人工序列(artificialsequence)
<400>8
tcaacacgttagctcccgcggtcagtattaaggcag36
<210>9
<211>39
<212>dna
<213>人工序列(artificialsequence)
<400>9
ctggtgcaattaagggtttcctcgagatgccagctgccc39
<210>10
<211>42
<212>dna
<213>人工序列(artificialsequence)
<400>10
gaaacccttaattgcaccaggatagtttctaattaaaaccga42
- 还没有人留言评论。精彩留言会获得点赞!