湖羊MC4R基因的分子标记SNP732及其应用的制作方法
本发明涉及分子标记领域,具体地涉及一种湖羊mc4r基因的snp分子标记、相关引物对、试剂盒,以及它们在湖羊分子标记辅助育种中的应用,进一步涉及一种筛选湖羊体尺性状的方法及其在湖羊分子标记辅助育种中的应用。
背景技术:
黑素皮质素受体-4(melanocortinreceptor-4,mc4r)是下丘脑腹内侧核分泌的一类肽类物质,参与体重以及能量平衡的调节,由中枢神经系统中的多个神经元群体表达。与mcr家族其它成员不同的是,mc4r不存在于肾上腺皮质、毛囊及胎盘内,而是大量存在于中枢神经系统的各个区域中,包括大脑皮质、丘脑、下丘脑、脑干和脊索。人、猪、牛、羊、犬等哺乳动物的mc4r由332个氨基酸残基组成,具有7次跨膜结构,是跨膜g蛋白耦联受体家族的成员之一。
瘦素和胰岛素是动物摄食调控最重要的外周调节因子,mc4r可作为瘦素、胰岛素摄食调控通路的下游介质,实现对动物摄食的调控。因而该蛋白在中枢神经系统调节葡萄糖稳态过程中发挥着重要作用。哺乳动物mc4r基因的突变将破坏中枢神经系统内的肾上腺皮质通路,增加食欲和减少饱腹感,间接造成肥胖。人、犬、猪、牛、羊等哺乳动物mc4r基因的突变研究均显示,该基因可作为哺乳动物体重或生长性状的候选功能基因,在基因内存在很多与动物生长性状关联的snps。根据mc4r基因突变引起的表型效应,可以将该基因的突变分为五种类型:①引起mc4r蛋白质合成缺陷,或加速其降解,表现为mc4r蛋白量显著降低;②引发蛋白折叠或胞内转运的错误,导致蛋白滞留于细胞内;③虽然蛋白可正确整合于细胞膜,但与配体的结合能力下降;④引发信号传导障碍;⑤突变导致的效应尚不明确。
mc4r基因作为哺乳动物经典的生长性状候选基因,其snp研究受到了广泛关注,并在多物种中得到了可用于后续选育的候选功能性snps。因此,利用家畜群体内的mc4r基因snp对家畜体重、体尺的影响,在该基因内筛选出有利于湖羊体重增长、生长增速及肥育、屠宰性状的snp,并对有利于增加体重和肉质评分的基因型个体进行有目的的选种选配,将有助于培育出生长速度快,胴体品质好的湖羊群体,提高经济效益。
技术实现要素:
本发明以湖羊肉用新类群第三世代核心群的202头雌性个体为研究对象,采取pcr及产物直接测序的方式,确定湖羊肉用新类群第三世代核心群mc4r基因的snps,并与个体的生长性状进行关联分析,筛选与湖羊生长性状关联的snps,为湖羊肉用系分子育种提供有用方法。
为此,本发明的一个目的是提供一种与湖羊体尺性状相关的snp分子标记,其特征在于与seqidno:1比对,该分子标记包括位于相应于seqidno:1的第706位的snp706,其为c或a。优选,其中所述分子标记的序列如seqidno:1所示,所述第706位为c或a。
本发明的另一个目的是提供一种与湖羊体尺性状相关的snp分子标记,其特征在于与seqidno:1比对,该分子标记包括位于相应于seqidno:1的第732位的snp732,其为c或g。优选,其中所述分子标记的序列如seqidno:1所示,所述第732位为c或g。
本发明的另一个目的是提供一种用于检测上述的snp706或snp732的引物对。优选其序列如seqidno:2和seqidno:3所示。
本发明的另一个目的是提供包含上述引物对的试剂盒。
本发明的另一个目的是提供上述的snp分子标记、引物对或试剂盒在筛选湖羊体尺性状或湖羊分子标记辅助育种中的应用。
本发明的另一个目的是提供一种筛选湖羊体尺性状的方法,包括如下步骤:提取湖羊基因组dna,分别利用检测上述的snp706或snp732的引物对进行pcr扩增,分别对扩增产物中相应于seqidno:1的第706位或732位进行检测,从而筛选湖羊体尺性状。优选,其中所述引物对为权利要求4所述的引物对。优选,其中pcr扩增的反应程序为:94℃预变性5min;94℃变性30sec,58℃退火30sec,72℃延伸1min,35个循环;72℃延伸10min;pcr扩增的反应体系如下:
本发明的另一个目的是提供任一项的方法在湖羊分子标记辅助育种中的应用。
本发明的有益效果如下:本发明所述的分子标记、引物对及相关试剂盒可以用于筛选湖羊体尺性状或湖羊分子标记辅助育种。
附图说明
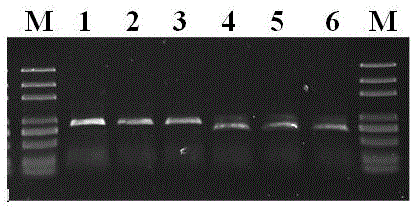
图1是mc4r基因引物1f+1r(泳道1~3)与引物2f+2r(泳道4-6)的pcr扩增(m:markerdl5000)。
具体实施方式
以下结合具体实施方式详细描述本发明,不能将其解释为限制本发明的范围。
大量的研究表明性别这个因素对绵羊与山羊的生长发育都具有显著影响。由于性别对湖羊的体重和体尺性状存在显著影响,因此,本发明在湖羊肉用系雌性个体的mc4r基因内筛选可影响湖羊体重及体尺性状的snp。对样本的性别鉴定可进一步避免个体的生产记录及基因组提取过程中的错误,影响后续mc4r基因snp的关联分析结果。
实施例1试验材料
试验动物样本来源
试验动物均来自杭州庞大农业开发有限公司构建的湖羊肉用新类群核心群g3代群体,共202只雌性个体,饲养管理按照肉羊标准的饲养管理方法进行饲养。每个个体采集颈静脉血10ml,置于edta抗凝管,-20℃保存。
体尺指标的测定
湖羊肉用新类群g3代系核心群个体的体尺测量包括体(斜)长、体高和胸围。每只羊至少测量3次,取平均值作为最终的测量结果。具体测定性状及测定方法见表1。
表1测定性状及测定方法
实施例2试验方法
1.外周血基因组dna提取,按照已知常规方法进行。
2.dna浓度和纯度检测,按照已知常规方法进行。
3.湖羊mc4r基因pcr扩增及snps检测。
湖羊mc4r基因pcr扩增引物
引物由杭州擎科梓熙生物技术有限公司合成,引物序列、扩增片段长度及p退火温度见表2。
表2湖羊mc4r基因pcr扩增引物
pcr扩增
pcr扩増体系为25μl,pcr反应程序见表3。具体成分见表4。
表3湖羊mc4r基因pcr扩增程序
表4湖羊mc4r基因pcr扩增体系
pcr产物的检测
取5μl的pcr产物上样于1.0%的琼脂糖凝胶,使用dl5000为marker标记,200v电压电泳15min左右,eb染色5min,于凝胶成像仪中观察扩增条带的有无,并拍照。
pcr产物的测序
每个样品均按照表2列出的引物分别进行扩增,扩增产物交由杭州擎科梓熙生物技术有限公司,进行正、反向的双向测序,直至双向测序结果一致。
序列分析
使用mutationsurveyor5.02(softgenetics,美国)软件分析每个个体的正、反向测序峰图,以nm_01126370为参照序列,确定每个个体mc4r基因的突变位置和突变方式。测序结果异常或正、反向测序结果不吻合的个体需进行二次测序,直至pcr产物正、反向测序分析结果完全一致。
数据统计与分析
使用haploview4.2软件分析连锁不平衡程度。
使用littleprograme计算pic值。
使用popgen32软件计算各snps的位点杂合度、shannon信息含量、基因频率、基因型频率,并检测各位点是否符合hardy-weinberg平衡。
多态信息含量分析
pic(polymorphisminformationcontent),表示一个后代所获得某个等位基因来自于它的亲本的同一个等位基因的可能性,是衡量等位基因片段多态性的理想指标计算公式为:
pi和pj分别为第i个和第j个等位基因频率;n为等位基因数
位点杂合度
he(heterozygosity)指每个基因座上所存在的杂合个体的平均频率。杂合度能客观地反应群体的遗传变异水平,平均杂合度值越大,表明群体内遗传差异也越大,遗传多样性丰富,遗传潜力大,应用于动物遗传育种研究效果好;其数值越低,表明遗传一致性越高,说明群体内遗传变异小,遗传潜力也小。计算公式为:
pi表示第i个等位基因频率
shannon信息含量
sic(shannoninformationcontent),计算公式为:
sic=-clogpi
其中:pi为第i个等位基因在群体中的频率,c为常数。
基因频率和基因型频率
①基因的型频率=基因型个体数/该群体个体数×100%
②基因的频率=纯合基因型频率+1/2×杂合基因型频率
hardy-weinberg平衡检测
运用卡方检验(χ2)鉴定各位点的基因型频率和基因频率是否符合hardy-weinberg平衡,公式为:
其中,m代表基因型数目;fi表示观察到的第i个基因型的个体数;n代表总的样本数;pi表示第i个基因型的理论基因型频率。
关联分析
使用一般线性模型(generallinearmodel,glm;spss20)挖掘与湖羊肉用新类群核心群体重、体尺性状关联的snps。
由于分析的所有个体均为来自同一饲养场,相同饲养环境和管理条件的雌性羊只,因此数据建模时不包括场效应和性别效应。
具体模型为:y=xβ+e
其中,y:湖羊肉用新类群核心群体尺性状、体重性状表型值向量;
β:表型均值、snp等固定效应向量;
e:残差效应向量;
x为β的关联矩阵。
当y为湖羊肉用新类群核心群初生重表型值向量时,按照模型y=xβ+sα+e分析。
其中y:湖羊肉用新类群核心群体尺性状表型值向量;
α:同胞数的固定效应向量;
β:snp效应向量;
e:残差效应向量;
x、s分别为β、α的关联矩阵。
生物信息学分析
湖羊mc4r蛋白的二、三级结构预测
以dnasta软件预测目的蛋白的二级结构,以phre2(http://www.sbg.bio.ic.ac.uk/phyre2/html/page.cgi?id=index),在线预测目的蛋白的三级结构与mc4r蛋白的跨膜结构。
实施例3试验结果
湖羊mc4r基因片段的pcr扩增
湖羊mc4r基因pcr扩增结果显示,使用引物1f和1r,2f和2r进行扩增时,在750-1000bp间均可出现特异性的扩增条带,与预期分子量(分别对应引物1f和1r产物的862bp,引物2f和2r产物的670bp)一致(图1)。
湖羊mc4r基因pcr扩增产物突变分析
湖羊mc4r基因pcr扩增产物突变分析结果如表5所示。
表5湖羊mc4r基因的snps位点及突变类型、方式
使用引物对1f和1r(扩增产物为862bp,见序列表中的seqidno:6)、2f和2r(扩增产物为670bp,见序列表中的seqidno:7)在湖羊肉用新类群核心群g3个体中共检测到19个突变,其中引物对1f和1r的扩增产物内有11个突变位点,引物对2f和2r的扩增产物有8个突变位点,各位点的突变方式、置换类型、突变类型及突变位置见表3-5。19个突变位点的突变位置来看,其中5'utr占26.3%(5/19),orf区占42.1%(8/19),3'utr占31.6%(6/19);突变方式中转换占52.6%(10/19),颠换占47.4%(9/19);同义突变占75.0%(6/8),错义突变占25.0%(2/8)(表5)。
湖羊mc4r基因多态性与体重、体尺性状的关联分析
利用“关联分析”中的glm模型分别进行湖羊mc4r基因的19个snps与肉用核心群g3代雌性个体的体尺性状、体重性状的关联分析(表10)。
湖羊mc4r基因多态性与湖羊肉用新类群核心群g3代雌性个体体重性状的关联分析
分析结果显示,湖羊mc4r基因的19个snps均不影响湖羊肉用核心群g3代雌性个体的初生重、断奶重、6月龄体重及成年重等体重性状(p>0.05),(表10)。
湖羊mc4r基因多态性与湖羊肉用新类群核心群g3代雌性个体成年体尺性状的关联分析
分析结果显示,检测到的19个snps中有2个snps,分别是位于开放阅读框上的g.706c→a和g.732c→g,可显著影响湖羊肉用核心群g3代雌性个体成年体高性状(p<0.05或p<0.01)(表10)。
其中g.706c→a野生型cc雌性湖羊的成年体高(76.09±3.07cm)显著低于突变纯合型aa个体(79.50±1.91cm)(p<0.05),与ca基因型个体(76.77±2.77cm)显著不差异(p>0.05)。所选育的湖羊肉用新类群核心群g3代雌性个体g.706c→a以野生型为主(186/202)。而突变纯合型个体仅为4/202。
其中g.732c→g野生型cc基因型雌性个体的成年体高(77.57±2.49cm)显著高于突变杂合型cg个体(76.20±2.88cm)(p<0.05),极显著高于突变纯合型gg个体(75.72±3.30cm)(p<0.01),突变杂合型cg雌性个体的成年体高与gg基因型间差异不显著(p>0.05)。所选育的湖羊肉用新类群核心群g3代雌性个体g.732c→g突变杂合型与突变纯合型相同为(86/202)。而野生纯合型个体仅为30/202。
湖羊mc4r蛋白二、三级结构预测结果
发现的mc4r基因222bp、225bp、306bp、706bp、894bp、951bp处的检出突变不引起该蛋白编码氨基酸的变化,与参照序列np_001119842相比,g.548c→t的突变导致112位编码的苏氨酸(thr)转变为蛋氨酸(met),g.732c→g的突变导致173位编码的异亮氨酸(ile)转变为蛋氨酸(met)。
利用dnastar预测上述突变是否导致该蛋白二级结构中的α螺旋(α-helix),β折叠(β-sheet)、β转角(β-turn)和无规则卷曲(coi1)发生变化,上述位点突变可导致蛋白120位β转角、120位的无规则卷曲、1170~175位α螺旋结构消失。
利用phre2预测mc4r蛋白突变前后的三级结构,结果显示错义突变可导致相应位置三级结构的α螺旋、β转角、无规则卷曲发生空间改变,其中深红色的为α螺旋、黄色为β折叠、淡蓝色为β转角(β-turn),白色为其它残基。
对湖羊mc4r蛋白跨膜区预测结果
以phre2在线软件对湖羊mc4r蛋白跨膜区进行预测,湖羊mc4r蛋白具有7个跨膜结构,构成跨膜区的氨基酸序列号为:46~70、80~102、121~145、167~185、193~213、247~267、283~305;蛋白n端在膜外,c端在膜内,g.548c→t(thr112met)位于第2跨膜区与第3跨膜区间的膜外区,g.732c→g(ile173met)位于第4跨膜区。
snp及其检测和分析方法
snp是指基因组水平上某一特定核苷酸的位置发生变化所引起的dna序列多态性。目前,检测snp的方法大致可分为两大类,一类是以单链构象多态性、酶切扩增多态性序列、连接酶检测反应等为代表的以凝胶电泳为基础的检测方法;另一类是以直接测序、变性高效液相色谱、高分辨率溶解曲线等为代表的高通量、自动化程度较高的检测方法。
本发明采用dna直接测序法,结合mutationsurveyor5.02软件,分析了湖羊肉用新类群g3代核心群雌性个体mc4r基因全序列的snps,共发现19个snps,其中5′utr内有5个snps,分别是g.36g→a、g.73g→a、g.79c→t、g.110c→g和g.139g→a;orf区内8个snps,分别是g.222c→t、g.225g→c、g.306g→a、g.548c→t(thr112met)、g.706c→a、g.732c→g(ile173→met)、g.894g→c和g.951g→a;3′utr内8个snps,分别是g.1229g→a、g.1267g→a、g.1291g→c、g.1337c→a、g.1352g→c和g.1382g→c。在绵羊群体中,尚未有mc4r基因g.36g→a、g.73g→a、g.79c→t、g.225g→c、g.1337c→a、g.1352g→c和g.1382g→c突变的报道。
与参照序列相比,在长度为1464bp的扩增产物中,使用dna直接测序,结合mutationsurveyor5.02软件,共检测出19个snps,5′utr区的检测长度为213bp,检出5个snps,平均每43bp出现1个snp;orf区长度为999bp,检出8个snps,平均每125bp出现1个snp;3′utr区检测长度为203bp,检出8个snps,平均每29bp出现1个snp。3′utr区snp的密度高于其他区段,本发明检测出snps无论在数量和密度均高于已发表的绵羊mc4r基因snps研究结果。
湖羊mc4r基因snps
snps的突变形式及位置
snp在群体中的发生频率大于1%。包括单个碱基的转换、颠换、插入和缺失。转换是指同型碱基之间的置换,如嘌呤与嘌呤(a→g)、嘧啶与嘧啶(c→t)间的替换;颠换是指发生在嘌呤与嘧啶(a→c、a→t、g→c和g→t)之间的替换。snp转换发生的频率占多数,约占置换总数的66.7%,且以c→t转换发生的频率高,可能是由于甲基化的cpg可通过自发水解脱氨基形成胸腺嘧啶。
本发明发现,湖羊mc4r基因snps转换比例为52.6%,颠换为47.4%,这目前研究发现“snp转换变异占66.7%”的结论存在一定的差异。且湖羊mc4r基因的转换方式以g→a为主,占70%(7/10),c→t占30%(3/10),与目前研究发现“snp转换中c→t发生频率较高”的结论也不同,可见湖羊mc4r基因的snps的突变形式和数量具有一定特殊性。
根据snp对生物遗传性状的影响不同可分为蛋白编码snp和非蛋白编码snp,其中蛋白编码snp主要分布在基因编码区,这种突变为同义蛋白编码snp,否则称为非同义蛋白编码snp或是错义蛋白编码snp。由于选择压力,整个基因组及种群间的snp分布呈现不均匀状态,在非编码区中snps数量要高于编码区。
本发明发现湖羊mc4r基因snps编码区共发现了8个snps。其中错义突变占25.0%(2/8),分别为g.548c→t(thr112met)与g.732c→g(ile173met),而同义突变则为75.0%(6/8)。其原因是由于错义突变对个体生理功能的影响较大,所以发生频率较低,而同义突变影响较小,发生的概率比较高。
湖羊mc4r基因snps的群体遗传学分析
湖羊mc4r基因snps遗传变异程度的评估
遗传变异是有机体适应环境变化的先决条件。变异杂合度和有效等位基因数等参数可用于衡量群体内的遗传变异程度。对于一个群体来说,变异杂合度和有效等位基因数的数值越大,群体内基因一致性越差,变异性就越高,选择潜力越大,反之,群体变异程度越低,选择的潜力越小。
pic值越大说明有效等位基因数与杂合度越大,该群体在此snp位点的变异性也越高。湖羊mc4r基因的编码区8个snp,其中2个为错义突变:g.548c→t为低度多态,g.732c→g为中度多态,6个为同义突变:g.222c→t、g.225g→c、g.306a→g、g.706c→a为中度多态,g.894g→c、g.951g→a为低度多态,而且群体内属于中度多态突变位点都位于编码区。
g.222t→c、g.225g→c、g.306a→g、g.732c→g位点的pic值均在0.25~0.5之间,为中度多态snp,说明这3个位点具有较高的遗传多样性、选育潜力高。
湖羊mc4r基因snps的评估
hardy-weinberg平衡状态是一种理想的状态。在生物群体里,影响平衡状态的各种因素在不断发生,导致群体遗传组成变化,从而引起生物的进化。若试验群体处于平衡状态,则表明该群体内样本的基因频率可代表正常群体的基因频率。而若试验群体有所偏移,未达到平衡状态,则表明该群体可能存在近亲或小群体婚配、自然选择、随机漂变、突变率升高、迁移等。由于湖羊的育种工作需要根据羊只的生产性能和体型外貌进行连续的选育,因此极易产生hardy-weinberg不平衡的状态。
χ2检验结果表明,g.139g→a、g.706c→a严重偏离了hard-weinberg平衡状态(p<0.01),同群体内的,其余的17个snps均处于hard-weinberg平衡状态。
g.706c→a与湖羊肉用系g3代核心群雌性个体成年羊体重性状、体尺性状的关联分析结果显示,aa基因型个体的体重性状(初生重、断奶重、六月龄重、周岁重)虽然与cc基因型个体相比差异不显著,但aa基因型个体的体高(79.50±1.91cm)显著高于cc基因型个体(76.09±3.07cm)。由于野生型cc和突变纯合型aa的观察个体数均超过相应的理论个体数,说明在湖羊肉用新类群核心群选育过程中,g.706c→a位点可能和某些符合育种需要的基因连锁,导致该位点未处于hard-weinberg平衡状态。当然,这一结论仍需要在湖羊肉用新类群核心群雌性个体中进行大样本量,和多代次的观察与验证。
湖羊mc4r基因多态位点基因型与体重性状关联分析
将该基因的19个snps分别与体重性状进行关联分析,显示检测群体内的19个snps均与体重不相关。本发明组前期研究结果显示,6月龄雌性湖羊与6月龄杜泊×湖羊杂交f1代雌性个体的体重及屠宰率差异不显著,而6月龄雄性湖羊与6月龄杜泊×湖羊杂交f1代雄性个体的体重及屠宰率差异显著。
体重性状为微效多基因控制的qtl性状,鉴于本试验对象均为湖羊肉用新类群核心群雌性个体,而遗传背景相似群体内单个snp对雌性个体体重性状的影响远远达不到品种对体重影响的水平。因此可以解释虽然mc4r基因为重要的生长性状相关候选功能基因,但在该基因内也未检测到与湖羊雌性个体体重相关的位。
湖羊mc4r基因多态位点基因型与体尺性状关联分析
将该基因的19个snps分别与体重性状进行关联分析,显示检测群体内有2个snps与成年雌性个体的体尺性状相关(p<0.05)。
g.706c→a位于编码区,为同义突变,检测发现3种基因型。研究结果显示,g.706c→a仅影响雌性湖羊成年羊的体尺性状,野生型cc雌性湖羊的成年体高(76.09±3.07cm)虽显著低于突变纯合型aa个体(79.50±1.91cm)(p<0.05),但与ca基因型个体(76.77±2.77cm)显著不差异(p>0.05)。
g.732c→g位于编码区,存在cc、cg、gg3种基因型,可导致位于蛋白的第4跨膜区的第173位氨基酸由异亮氨酸突变为蛋氨酸。其中野生型cc基因型雌性个体的成年体高(77.57±2.49cm)显著高于突变杂合型cg个体(76.20±2.88cm)(p<0.05),极显著高于突变纯合型gg个体(75.72±3.30cm)(p<0.01),突变杂合型cg雌性个体的成年体高与gg基因型间差异不显著(p>0.05)。在选育的湖羊肉用新类群核心群g3代雌性个体g.732c→g以突变纯合型为主(86/202)。而野生纯合型个体仅为30/202。在该位点对野生型cc个体进行定向选育,可加快体尺性状的遗传进展。
湖羊mc4r基因连锁不平衡分析
连锁不平衡是一种等位基因的关联现象。连锁不平衡常用r2和d'衡量,如果r2>0.33,d′>0.8,就可以认为2个snps位点之间是紧密连锁的,作为一个整体进行遗传。
连锁不平衡分析结果显示,突变位点g.222c→t与g.225c→g的r2=1,d'=0.97,处于强连锁不平衡状态,但关联分析结果显示g.222c→t、g.225c→g并不影响雌性湖羊的体重性状、体尺性状,似乎这两个位点对于湖羊的生产性状选育并没有太大的意义。但该结论仍需要扩大群体数量,并确定该位点是否同样不会影响湖羊雄性个体的体重和体尺性状。
湖羊mc4r蛋白二、三级结构预测结果及跨膜结构分析
蛋白质的一级结构决定高级结构,而高级结构决定蛋白质的功能。利用生物信息学进行蛋白质结构预测、蛋白跨膜区、进而来预测功能基因的位置,获得与其生物学功能相关的数据。湖羊mc4r蛋白跨膜区的电子预测结果显示,湖羊mc4r蛋白同样具有七次跨膜结构,n端在细胞外,c端在细胞内。
本发明发现的两个错义突变的位点:g.548位c→t可导致第112位苏氨酸(第2跨膜区与第3跨膜区间的膜外区)突变为蛋氨酸。g.732位c→g可导致第173位异亮氨酸(第4跨膜区)突变为蛋氨酸。苏氨酸是一种极性,含羟基类氨基酸。异亮氨酸是一种非极性或疏水性氨基酸,带有脂肪烃侧链的中性氨基酸。蛋氨酸是一种非极性或疏水性氨基酸,是一种含硫氨基酸,同时也是α-氨基酸的一种,在它的分子中,含有一个碱性基团(-nh2)和一个酸性基团(-cooh),它是二性的。若碱基突变发生跨膜结构序列内很可能会导致基因结构发生变化,从而影响该蛋白的活性或者功能。
dnastar对湖羊mc4r蛋白结构预测的结果显示,ile173met可导致湖羊mc4r蛋白1700~175位的α螺旋丢失,继而引起三级结构第四跨膜区α螺旋发生变化。因此,该错义突变可能通过影响蛋白结构,影响其功能,并在体尺性状上表现出差异。
表6湖羊g.732g→a位点与体高性关联分析
注:不同大写字母上标之间差异极显著(p<0.01)
综上,本发明在湖羊肉用新类群g3代核心群群体中检测19个snps,15个位点为低度多态,4个位点为中度多态;g.222c→t与g.222c→g存在强连锁;g.306a→g具有丰富的遗传多态;除g.139g→a、g.706c→a外,其余17个snps均达到hard-weinberg平衡状态。
mc4r基因编码区内存在2个与湖羊雌性个体成年体高显著相关的snps,g.548c→t可导致第112位苏氨酸(第2跨膜区与第3跨膜区间的膜外区)突变为蛋氨酸。g.732位c→g可导致第173位异亮氨酸(第4跨膜区)突变为蛋氨酸。其中g.706c→a野生型cc雌性湖羊的成年体高(76.09±3.07cm)显著低于突变纯合型aa个体(79.50±1.91cm)(p<0.05),与ca基因型个体(76.77±2.77cm)显著不差异(p>0.05)。g.732c→g野生型cc基因型雌性个体的成年体高(77.57±2.49cm)显著高于突变杂合型cg个体(76.20±2.88cm)(p<0.05),极显著高于突变纯合型gg个体(75.72±3.30cm)(p<0.01),突变杂合型cg雌性个体的成年体高与gg基因型间差异不显著(p>0.05)。
利用上述snp进行湖羊肉用系群体分子标记辅助育种的早期选育,可显著提高湖羊肉用新类群雌性个体的肉用性状。
以上以具体实施方式描述了本发明,本领域技术人员在不背离本发明精神的情况下,可以做出等同的修改或变型,其同样在权利要求的范围之内。
表7湖羊mc4r基因snps位点的遗传参数
表8湖羊mc4r基因snps位点的群体遗传学分析
表9湖羊mc4r基因snps位点的hardy-weinberg平衡检验
序列表
<110>浙江省农业科学院
<120>湖羊mc4r基因的分子标记snp732及其应用
<160>7
<170>siposequencelisting1.0
<210>1
<211>1788
<212>dna
<213>湖羊(husheep)
<220>
<221>mutation
<222>(36)..(36)
<223>s为g或a
<220>
<221>mutation
<222>(73)..(73)
<223>s为g或a
<220>
<221>mutation
<222>(79)..(79)
<223>s为c或t
<220>
<221>mutation
<222>(110)..(110)
<223>s为c或g
<220>
<221>mutation
<222>(139)..(139)
<223>s为g或a
<220>
<221>mutation
<222>(222)..(222)
<223>s为c或t
<220>
<221>mutation
<222>(225)..(225)
<223>s为c或g
<220>
<221>mutation
<222>(306)..(306)
<223>s为g或a
<220>
<221>mutation
<222>(548)..(548)
<223>s为c或t
<220>
<221>mutation
<222>(706)..(706)
<223>s为c或a
<220>
<221>mutation
<222>(732)..(732)
<223>s为c或g
<220>
<221>mutation
<222>(894)..(894)
<223>s为g或c
<220>
<221>mutation
<222>(951)..(951)
<223>s为g或a
<220>
<221>mutation
<222>(1229)..(1229)
<223>s为g或a
<220>
<221>mutation
<222>(1267)..(1267)
<223>s为g或a
<220>
<221>mutation
<222>(1291)..(1291)
<223>s为g或c
<220>
<221>mutation
<222>(1337)..(1337)
<223>s为c或a
<220>
<221>mutation
<222>(1352)..(1352)
<223>s为g或c
<220>
<221>mutation
<222>(1382)..(1382)
<223>s为g或c
<400>1
cagcctaagatttccaagtgatgccgaccagagccscacctgaaagagactgaaaacttc60
ctctccagctccsgagctsgggacgtttattcacagcaggcatgccactstccgccgcct120
aactttcgtttggggcaastcaagactggagaaaggcgctgaggctgccggacccgggag180
gttcagtcagtccagaggggacctgaatccaagatgaactcsacscagccccatggaatg240
cacacctctctccactcctggaaccgcagcggccacgggctgcccaccaatgtcagtgag300
tccccsgcaaaaggctactcggacggggggtgctatgagcagctctttgtctctcccgag360
gtgtttgtgactctgggggtcatcagcttgttggagaatattctggtgattgtggccatc420
gccaagaataagaatctgcactcacccatgtactttttcatctgcagcctggctgtggct480
gacatgttggtgagcgtttccaacgggtccgaaaccattgtcatcaccctgctgaacagc540
acagacasggacgcgcagagcttcacggtgaatattgacaacgtcatcgactcggtgatc600
tgcagctccttgctcgcctccatctgcagcttgctgtcgatcgcggtggacaggtacttc660
accatcttctatgcgctccagtaccatagcatcatgacggtgcggsgggtggcgatcacc720
atcagtgccatstgggcggcctgcacggtgtcgggcgtcttgttcatcatttactcagac780
agcagcgctgtcatcatctgcctcatcaccgtgttcttcaccatgctggctctcatggcg840
tctctctacgtccacatgttcctcatggccaggctccacattaagaggatcgcsgtcctg900
ccaggcaccggcgccatccgccagggcgccaacatgaagggggcgatcacsctgaccatc960
ctgatcggggtctttgttgtctgctgggcccccttcttcctgcacctgatattctacatc1020
tcctgcccccagaacccctactgcgtgtgcttcatgtctcactttaacctgtatctcatc1080
ctgatcatgtgtaattctgtcatcgaccctctgatctacgccctgcggagccaggaactg1140
aggaaaaccttcaaagagatcatttgctgctctcctctaggtggcctctgtgatttgtct1200
agcagatattaaatggggacaaactgcgstgccaaacacaagcttaagagaccttctcct1260
tctcatstgtacaatctgaacagtctgtatsagccacagctttttcttctgtgtagggca1320
tggagtgaaaaattctsttgtatcagttgaastttgtgatttttttctgatgtgaagcag1380
tscccagtcttgctgtatttttaatatcatgctactttctggctgtaaaatgtgaatcca1440
catcacaggttataggcactatggatttataaaaaaaaaaaagaaacaaagtccttatga1500
ggagtttaacagtgtttccttcttgtgatttacaaggatgtgacactttgcttgcttttg1560
taacatggaaatcacagcttccttaagtatattctcatgagtggatttaatgttatactt1620
tacaacactggagtataaaatttgattccagtatttaggggagaaatattgagaacatat1680
tgcttaatcataaaaaaaaaaaccaagctgaaatttcaggtaatttaataagacttgctc1740
attcattcttcctgtgccgaagttgaaatgaaacttctactgggacaa1788
<210>2
<211>25
<212>dna
<213>引物(primer)
<400>2
gcctaagatttccaagtgatgctga25
<210>3
<211>24
<212>dna
<213>引物(primer)
<400>3
ggaacatgtggacatagagagacg24
<210>4
<211>24
<212>dna
<213>引物(primer)
<400>4
catctgcctcatcaccgtgttctt24
<210>5
<211>23
<212>dna
<213>引物(primer)
<400>5
ccatagtgcctataacctgtgat23
<210>6
<211>862
<212>dna
<213>湖羊(husheep)
<400>6
cagcctaagatttccaagtgatgccgaccagagccgcacctgaaagagactgaaaacttc60
ctctccagctccggagctcgggacgtttattcacagcaggcatgccactgtcggccacct120
aactttcgtttggggcaagtcaagactggagaaaggcgctgaggctgccggacccgggag180
gttcagtcagtccagaggggacctgaatccaagatgaactctacgcagccccatggaatg240
cacacctctctccactcctggaaccgcagcggccacgggctgcccaccaatgtcagtgag300
tccccagcaaaaggctactcggacggggggtgctgggagcagctctttgtctctcccgag360
gtgtttgtgactctgggggtcatcagcttgttggagaatattctggtgattgtggccatc420
gccaagaataagagtctgcactcacccatgtactttttcatctgcagcctggctgtggct480
gacatgttggtgagcgtttccaacgggtccgaaaccattgtcatcaccctgctgaacagc540
acagacacggacgcgcagagcttcacggtgaatactgacaacgtcatcgactcggtgatc600
tgcagctccttgctcgcctccatctgcagcttgctgtcgatcgcggtggacaggtacttc660
accatcttctatgcgctccagtaccatagcatcatgacggtgcggcgggtggcgatcacc720
atcagtgccatgtgggcggcctgcacggtgtcgggcgtcttgttcatcatttactcagac780
agcagcgctgtcatcatctgcctcatcaccgtgttcttcaccatgctggctctcatggcg840
tctctctacgtccacatgttcc862
<210>7
<211>669
<212>dna
<213>湖羊(husheep)
<400>7
catctgcctcatcaccgtgttcttcaccatgctggctctcatggcgtctctctacgtcca60
catgttcctcatggccaggctccacattaagaggatcgcggtcctgccaggcaccggcgc120
catccgccagggcgccaacatgaagggggcgatcacgctgaccatcctgatcggggtctt180
tgttgtctgctgggcccccttcttcctgcacctgatattctacatctcctgcccccagaa240
cccctactgcgtgtgcttcatgtctcactttaacctgtatctcatcctgatcatgtgtaa300
ttctgtcatcgaccctctgatctacgccctgcggagccaggaactgaggaaaaccttcaa360
agagatcatttgctgctctcctctaggtggcctctgtgatttgtctagcagatattaaat420
ggggacaaactgcgatgccaaacacaagcttaagagaccttctccttctcatgtgtacaa480
tctgaacagtctgtatgagccacagctttttcttctgtgtagggcatggagtgaaaaatt540
ctcttgtatcagttgaagtttgtgatttttttctgatgtgaagcagtgcccagtcttgct600
gtatttttaatatcatgctactttctggctgtaaaatgtgaatcctcatcacaggttaca660
ggcactatg669
- 还没有人留言评论。精彩留言会获得点赞!