OsCIPK9蛋白及其编码基因在提高水稻对纹枯病抗性中的应用的制作方法
(一)技术领域
本发明涉及oscipk9蛋白及其编码基因在提高水稻对纹枯病抗性中的应用。
(二)
背景技术:
水稻纹枯病是由立枯丝核菌(rhizoctoniasolanikühn)引起的一种土传真菌病害,为水稻的三大病害之一,也是一种危害遍布全球的水稻病害。该病害在水稻整个生育期均可发生,主要危害叶鞘、叶片,具有危害大、流行性强、寄主范围广等特点,严重影响水稻高产稳产。在农业生产上,水稻纹枯病的防治主要采用药剂防治和栽培管理,但长期使用化学药剂不仅造成环境污染,还会加快病原菌变异,产生耐药性,给纹枯病的防治带来极大的困难,培育稳定的抗病品种才是经济、有效、环保、根本的防治方法。水稻转基因育种技术已经被广泛应用,具有周期短、效率高等特点。目前,通过转基因技术对水稻基因进行操作已经培育了大量的水稻抗病及高产新品种。目前,由于我国抗纹枯病水稻种质资源的缺乏,还未发现对纹枯病高抗或免疫的水稻品种。因此,挖掘水稻纹枯病抗性相关基因,利用转基因育种技术改变抗性相关基因表达对水稻抗纹枯病种质资源创制和纹枯病的有效防控具有重要意义。
植物细胞中,ca2+作为胞内第二信使,参与调控植物多种逆境胁迫响应、植物各器官的形成与发育过程。cbls是植物中广泛存在的一种钙感受器(calciumsensor),通过自身的ef手型(ef-hand)结构域与ca2+结合,导致自身构象改变,从而识别或解码ca2+信号并将信息传递至下游,通过激酶(cipks)的磷酸化作用或转录因子的调控,最终使胞内ca2+信号转化为细胞生理反应。cipks(cblsinteractingproteinkinases)为cbls互作蛋白,是一类植物特有的丝氨酸-苏氨酸蛋白激酶,能将cbls感知的信号通过磷酸化下游互作蛋白的方式,将胞内ca2+信号转化为细胞生理反应,调控植物对逆境的应答反应。水稻oscipk9已报导参与根生长,但是有关oscipk9编码基因与水稻纹枯病抗性间的关系迄今未见报道。
(三)
技术实现要素:
本发明目的是提供oscipk9蛋白及其编码基因在提高水稻对纹枯病抗性中的应用。
本发明采用的技术方案是:
oscipk9蛋白在提高水稻对纹枯病抗性中的应用。
具体的,所述oscipk9蛋白氨基酸序列如seqidno.2所示。
seqidno.2序列如下:
maeaeaeaagagagagparrttrvgryelgktigegsfakvkvardtrtgdtlaikvldrnhvlrhkmveqikreistmklikhpnvvqlhevmaskskiymvleyvdggelfdkivnsgrlgedearryfhqlinavdychsrgvyhrdlkpenllldshgalkvsdfglsafapqtkedgllhtacgtpnyvapevladkgydgmaadvwscgiilfvlmagylpfddpnlmtlyklickakvscphwfssgakkfikrildpnpctritiaqileddwfkkdykpplfeqgedvslddvdaafdcseenlvaekrekpesmnafalisrsqgfnlgnlfekemmgmvkretsftsqctpqeimskieeacgplgfnvrkqnykmklkgdktgrkgylsvatevfevapslhmvelrktggdtlefhnfynnfsselkdivwksesdakaakkr*。
本发明还涉及oscipk9编码基因在提高水稻对纹枯病抗性中的应用。
具体的,所述oscipk9编码基因核苷酸序列如seqidno.1所示。
seqidno.1序列如下:
atggcggaggcggaggcggaggcg.gcgggggcgggggcgggggcggggccggcgaggaggacgacgagggtggggcgctacgagctcggcaagacgatcggggaggggagcttcgccaaggtcaaggtcgcccgcgacacgcgcaccggcgacaccctcgccatcaaggtgctcgaccgcaaccatgtgcttcgccacaagatggtcgagcagatcaaacgggagatttccacaatgaagctaataaaacatccaaatgtggtccagttgcatgaggtcatggctagcaaatcaaagatatacatggttcttgaatatgttgatggaggcgagctttttgataagattgtcaattctgggagactaggagaagatgaagcaagaagatacttccatcaacttataaatgcagtagattattgccacagcagaggagtgtaccatagagatctgaagccagagaatctgcttcttgattcacatggagctcttaaagtttcagattttggccttagtgcgtttgctccacaaacaaaagaggatggacttctgcatactgcttgcggaacaccaaattatgttgcacctgaggtgcttgctgataaaggttatgatggtatggctgctgatgtatggtcctgcggcataatcttatttgtcctcatggctggatatttacctttcgatgaccccaacctaatgaccctttacaaactgatctgcaaggcaaaagtttcttgtccacattggttttcttcaggtgcgaagaagttcattaagcgcatacttgatcctaatccctgcacaagaataacaattgcacaaattttggaagatgattggttcaaaaaggattacaaaccaccactttttgagcaaggtgaagatgtgagccttgatgatgttgatgctgcattcgattgttcagaggagaatctcgtggcagagaagagagaaaagcctgaatccatgaatgcatttgctctaatttcaaggtcacagggattcaaccttggaaatttatttgagaaggaaatgatgggaatggtgaagcgggaaacttccttcacatctcaatgtacaccacaggagatcatgtctaaaatagaggaagcttgtggacctcttggtttcaatgtgcggaaacaaaattataagatgaagttaaaaggtgataagactggaagaaaaggctatttatctgtagcaacggaggttttcgaggttgctccatcactccacatggttgagcttcgtaaaactggaggggacacgctggagtttcacaatttctacaacaatttctcgtcagagttaaaagacatagtgtggaaatctgaatctgacgcaaaagcagcaaagaagaggtga。
具体的,所述应用为:通过敲除oscipk9基因,得到对纹枯病抗性减弱的转基因水稻。相反,通过过表达oscipk9基因,得到对纹枯病抗性增强的转基因水稻。
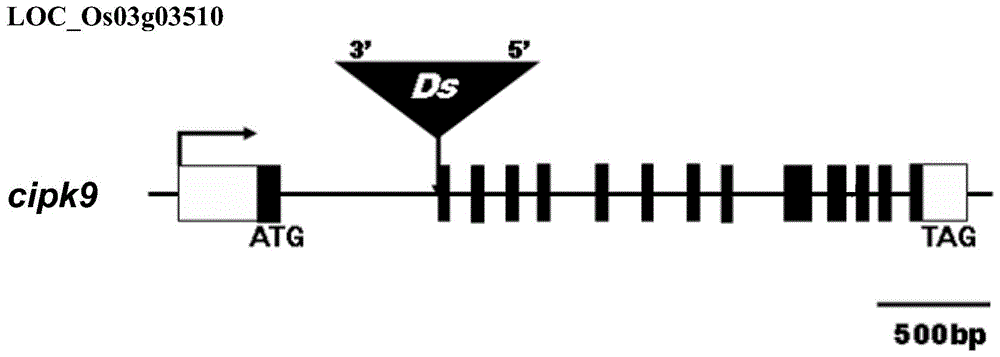
本发明的有益效果主要体现在:本发明通过ds插入水稻中的oscipk9基因第一个内含子中,获得oscipk9基因敲除的转基因水稻。通过纹枯病抗性鉴定发现,与野生型水稻对照相比,cipk9转基因水稻更感病。并通过非特异性启动子ubiquitin表达oscipk9获得过表达植株,通过纹枯病抗性鉴定发现,与野生型相比,oscipk9过表达(oscipk9ox)转基因水稻更抗病,表明cipk9基因参与调控水稻纹枯病抗性。
(四)附图说明
图1为用于敲除oscipk9基因的突变体中染色体的形态示意图;
图2为野生型(wt)和敲除植株(cipk9)基因表达差异图;
图3为野生型(wt)和敲除植株(cipk9)接种纹枯病菌后表型(a)及病斑面积占叶片面积百分比(b)。
图4为用于过表达oscipk9基因的载体结构示意图(a)、野生型(wt)和过表达植株(cipk9ox)中oscipk9的基因表达差异图(b)、野生型(wt)和过表达植株(cipk9ox)接种纹枯病菌后表型(c)及病斑面积占叶片面积百分比(d)。
(五)具体实施方式
下面结合具体实施例对本发明进行进一步描述,但本发明的保护范围并不仅限于此:
实施例1:
敲除oscipk9基因的突变体从水稻ds突变体库中筛选出(chinhg,choems,leesh,park,sh,hancd.1999.molecularanalysisofriceplantsharboringanac/dstransposableelement-mediatedgenetrappingsystem.theplantjournal19:615–623.)。
实施例2:
对上述oscipk9基因敲除突变体cipk9进行分子鉴定,提取水稻根的总rna经反转录后,用如下引物进行rt-pcr方法鉴定:
oscipk9引物为:
oscipk9-f:tgagcttcgtaaaactggag;
oscipk9-r:gcattggatagttggaatac;
内参基因为ubiquitin,内参引物为:
ubiquitin-f:cacggttcaacaacatccag;
ubiquitin-r:tgaagaccctgactgggaag。
结果如图2所示,oscipk9基因敲除突变体cipk9中oscipk9的平均相对表达量明显低于野生型水稻dongjin(wt)中oscipk9的平均相对表达量。
实施例3:野生型(wt)和敲除植株(cipk9)接种纹枯病菌后表型观察
采用纹枯病菌接种水稻离体叶片的方法进行抗性鉴定。将野生型(wt)和敲除植株(cipk9)转基因水稻叶片接纹枯病菌。
将4℃保存的纹枯病菌菌株于超净工作台内接种在pda培养基上,置于26℃培养箱连续培养2~3d,用0.7cm无菌打孔器在菌落边缘取菌饼用于接菌实验。
在灭菌的培养皿中放入两张用无菌水浸湿的已灭菌滤纸,选取健康无病虫害损伤的、叶龄相同的叶片,剪成大小均等的七份,叶背朝上,整齐的摆放在滤纸上,用接种针或牙签在每片叶子的中部位置扎一个小眼,将打孔器提前打好的菌饼接种在叶片打孔位置,盖上盖子后置于27℃培养室培养,将湿度保持在90%左右,三次重复。
接种后96h拍照,结果如图3所示,与野生型对照相比,cipk9转基因水稻更感病,病斑面积占叶片面积80%左右(图3a,b),表明cipk9基因参与调控水稻纹枯病抗性。
实施例4:野生型(wt)和过表达植株(cipk9ox)接种纹枯病菌后表型观察
提取水稻品种日本晴的rna,反转录为cdna。以此cdna为模板,以5′-aagcttatggcggaggcggaggc-3′和5′-ggatcctcacctcttctttgctgct-3′为引物扩增,得到1372bp的pcr产物,该pcr产物具有序列表中序列1自1372位核苷酸。用hindiii和bamhi酶切该pcr产物,得到的酶切产物与经过同样酶切得到的13094bp的pga1611载体(piaohl,xuanyh,parksh,jebi,parksj,parksh,kimcm,huangj,wanggk,kimmj,kangsm,leeij,kwontr,kimyh,yeous,yig,sond,hancd.oscipk31,acbl-interactingproteinkinaseisinvolvedingerminationandseedlinggrowthunderabioticstressconditionsinriceplants.molcells.2010,30:19-27;公众可从韩国国立庆尚大学获得)骨架连接,使该pcr产物插入pga1611载体的hindiii和bamhi位点间,得到终载体。经过测序,该重组载体为将序列表中序列1自5′末端第1位-1372位核苷酸插入pga1611载体的hindiii和bamhi酶切位点间(结构示意图如图4a所示),为cipk9过表达载体。该具有反向重复的重组表达载体用ubiquitin基因的启动子。对上述oscipk9基因过表达植株cipk9ox分子鉴定,提取水稻叶片的总rna经反转录后,用如下引物进行rt-pcr方法鉴定:
oscipk9引物为:
oscipk9-f:tgagcttcgtaaaactggag;
oscipk9-r:gcattggatagttggaatac;
内参基因为ubiquitin,内参引物为:
ubiquitin-f:cacggttcaacaacatccag;
ubiquitin-r:tgaagaccctgactgggaag。
结果如图4b所示,oscipk9过表达植物cipk9ox中oscipk9的平均相对表达量明显高于野生型水稻dongjin(wt)中oscipk9的平均相对表达量。
采用纹枯病菌接种水稻离体叶片的方法进行抗性鉴定。将野生型(wt)和过表达植株(cipk9ox#1,#3)转基因水稻叶片接纹枯病菌。将4℃保存的纹枯病菌菌株于超净工作台内接种在pda培养基上,置于26℃培养箱连续培养2~3d,用0.7cm无菌打孔器在菌落边缘取菌饼用于接菌实验。在灭菌的培养皿中放入两张用无菌水浸湿的已灭菌滤纸,选取健康无病虫害损伤的、叶龄相同的叶片,剪成大小均等的七份,叶背朝上,整齐的摆放在滤纸上,用接种针或牙签在每片叶子的中部位置扎一个小眼,将打孔器提前打好的菌饼接种在叶片打孔位置,盖上盖子后置于27℃培养室培养,将湿度保持在90%左右,三次重复。
接种后96h拍照,结果如图3所示,与野生型对照相比,cipk9ox转基因水稻更抗病,病斑面积占叶片面积30%左右(图4c,4d),表明cipk9基因参与调控水稻纹枯病抗性。
序列表
<110>沈阳农业大学
<120>oscipk9蛋白及其编码基因在提高水稻对纹枯病抗性中的应用
<160>2
<170>siposequencelisting1.0
<210>1
<211>1371
<212>dna
<213>未知(unknown)
<400>1
atggcggaggcggaggcggaggcggcgggggcgggggcgggggcggggccggcgaggagg60
acgacgagggtggggcgctacgagctcggcaagacgatcggggaggggagcttcgccaag120
gtcaaggtcgcccgcgacacgcgcaccggcgacaccctcgccatcaaggtgctcgaccgc180
aaccatgtgcttcgccacaagatggtcgagcagatcaaacgggagatttccacaatgaag240
ctaataaaacatccaaatgtggtccagttgcatgaggtcatggctagcaaatcaaagata300
tacatggttcttgaatatgttgatggaggcgagctttttgataagattgtcaattctggg360
agactaggagaagatgaagcaagaagatacttccatcaacttataaatgcagtagattat420
tgccacagcagaggagtgtaccatagagatctgaagccagagaatctgcttcttgattca480
catggagctcttaaagtttcagattttggccttagtgcgtttgctccacaaacaaaagag540
gatggacttctgcatactgcttgcggaacaccaaattatgttgcacctgaggtgcttgct600
gataaaggttatgatggtatggctgctgatgtatggtcctgcggcataatcttatttgtc660
ctcatggctggatatttacctttcgatgaccccaacctaatgaccctttacaaactgatc720
tgcaaggcaaaagtttcttgtccacattggttttcttcaggtgcgaagaagttcattaag780
cgcatacttgatcctaatccctgcacaagaataacaattgcacaaattttggaagatgat840
tggttcaaaaaggattacaaaccaccactttttgagcaaggtgaagatgtgagccttgat900
gatgttgatgctgcattcgattgttcagaggagaatctcgtggcagagaagagagaaaag960
cctgaatccatgaatgcatttgctctaatttcaaggtcacagggattcaaccttggaaat1020
ttatttgagaaggaaatgatgggaatggtgaagcgggaaacttccttcacatctcaatgt1080
acaccacaggagatcatgtctaaaatagaggaagcttgtggacctcttggtttcaatgtg1140
cggaaacaaaattataagatgaagttaaaaggtgataagactggaagaaaaggctattta1200
tctgtagcaacggaggttttcgaggttgctccatcactccacatggttgagcttcgtaaa1260
actggaggggacacgctggagtttcacaatttctacaacaatttctcgtcagagttaaaa1320
gacatagtgtggaaatctgaatctgacgcaaaagcagcaaagaagaggtga1371
<210>2
<211>456
<212>prt
<213>未知(unknown)
<400>2
metalaglualaglualaglualaalaglyalaglyalaglyalagly
151015
proalaargargthrthrargvalglyargtyrgluleuglylysthr
202530
ileglygluglyserphealalysvallysvalalaargaspthrarg
354045
thrglyaspthrleualailelysvalleuaspargasnhisvalleu
505560
arghislysmetvalgluglnilelysarggluileserthrmetlys
65707580
leuilelyshisproasnvalvalglnleuhisgluvalmetalaser
859095
lysserlysiletyrmetvalleuglutyrvalaspglyglygluleu
100105110
pheasplysilevalasnserglyargleuglygluaspglualaarg
115120125
argtyrphehisglnleuileasnalavalasptyrcyshisserarg
130135140
glyvaltyrhisargaspleulysprogluasnleuleuleuaspser
145150155160
hisglyalaleulysvalserasppheglyleuseralaphealapro
165170175
glnthrlysgluaspglyleuleuhisthralacysglythrproasn
180185190
tyrvalalaprogluvalleualaasplysglytyraspglymetala
195200205
alaaspvaltrpsercysglyileileleuphevalleumetalagly
210215220
tyrleupropheaspaspproasnleumetthrleutyrlysleuile
225230235240
cyslysalalysvalsercysprohistrppheserserglyalalys
245250255
lyspheilelysargileleuaspproasnprocysthrargilethr
260265270
ilealaglnileleugluaspasptrpphelyslysasptyrlyspro
275280285
proleuphegluglnglygluaspvalserleuaspaspvalaspala
290295300
alapheaspcysserglugluasnleuvalalaglulysargglulys
305310315320
proglusermetasnalaphealaleuileserargserglnglyphe
325330335
asnleuglyasnleupheglulysglumetmetglymetvallysarg
340345350
gluthrserphethrserglncysthrproglngluilemetserlys
355360365
ilegluglualacysglyproleuglypheasnvalarglysglnasn
370375380
tyrlysmetlysleulysglyasplysthrglyarglysglytyrleu
385390395400
servalalathrgluvalphegluvalalaproserleuhismetval
405410415
gluleuarglysthrglyglyaspthrleugluphehisasnphetyr
420425430
asnasnphesersergluleulysaspilevaltrplyssergluser
435440445
aspalalysalaalalyslysarg
450455
- 还没有人留言评论。精彩留言会获得点赞!