干细胞预防神经元顶梢枯死的用途的制作方法
本申请为分案申请,其原申请的申请日为2009年9月4日,申请号为200980144083.9,名称为“干细胞预防神经元顶梢枯死的用途”。
本发明主要涉及神经元损伤的治疗。具体地,本发明涉及减少因活化的巨噬细胞与营养不良的轴突相互作用而发生的轴突回缩(“顶梢枯死”),所述营养不良的轴突是在神经系统急性或慢性损伤过程中形成的。本发明还涉及促进轴突生长/再生。本发明特别涉及使用干细胞或其分泌的细胞因子,例如会在条件细胞培养基中产生的细胞因子,以改善或预防轴突顶梢枯死和/或促进轴突生长/再生。
背景技术:
轴突回缩
脊髓损伤后形成神经胶质疤痕,其造成cns再生的主要障碍(silverandmiller,2004)。在形成疤痕组织的区域中,再生轴突的末端延伸停止,并且膨胀并扭曲为可在轴突束(axontract)内保持数年的各种奇怪形状的“生长锥”(ramonycajal,1928;liandraisman,1995;houleandjin,2001;kwonetal.,2002)。cns内的损伤轴突在初始损伤后的数小时到数周的时间内从轴索切断的位置回缩。关于轴突回缩的性质、其原因、程度、时间以及对其是被动抑或主动过程的讨论已有不同报道(fayazandtator,2000)。
体外神经胶质疤痕模型
在形成疤痕组织的区域内,多类生长抑制分子被上调,包括被称为软骨素/硫酸角质素蛋白聚糖(pg)的细胞外基质(ecm)分子的家族(fitchandsilver,1997;morgensternetal.,2002;jonesetal.,2003;tangetal.,2003)。pg大致呈梯度分布,在损伤边缘浓度最低而在中心的浓度最高(daviesetal.,1999;fitchetal.,1999)。所述抑制性ecm组分阻断有活性的神经胶质细胞供养轴突通过层粘连蛋白再生的可能性(mckeonetal.,1991)。微移植实验显示成体感觉神经元在远离损伤时具有强再生能力。一旦再生纤维到达所述损伤位置附近,它们就能够艰难进入损伤边缘但最终在深入到最高pg浓度的区域时停止延伸并变得营养不良(daviesetal.,1999;grimpeandsilver,2004)。
在体外神经胶质疤痕模型中,使用尖缘的(即条纹)基底测定来检查pg对轴突的作用,诱导了生长锥转向或萎缩,但未发生营养不良(snowetal.,1990)。然而,在神经胶质疤痕的最新模型中,大致呈梯度的pg足以在再生成体轴突中产生营养不良的末端(tometal.,2004)。此体外系统迫使成体感觉神经元的再生轴突应对与层粘连蛋白混合的pg聚集蛋白聚糖的斑点梯度。在此神经胶质疤痕模型中形成了球状多囊泡末端。pg导致生长锥营养不良和动态营养不良末端。
神经元组织中的炎症和损伤
脊髓损伤的环境极端复杂。神经胶质疤痕的组分,例如高度硫酸化的蛋白聚糖、eph、开口和髓磷脂膜碎片(silverandmiller,2004;yiuandhe,2006;buschandsilver,2007),以及神经性炎症过程均是导致再生障碍的原因。炎性细胞在损伤内积聚(fitchetal.,1999)。星形细胞离开损伤的中心,变大并上调抑制性硫酸软骨素蛋白聚糖(cspg)的产生,cspg转而导致在切断的纤维上形成营养不良的末端球状物(tometal.,2004)。损伤内的少突胶质细胞死亡,引起脱髓鞘,这导致高浓度的抑制性髓磷脂分解产物(yiuandhe,2006;xieandzheng,2008)。
尽管蛋白聚糖和髓磷脂对轴突生长的抑制性作用得到公认,然而神经性炎症在再生和再生障碍中的作用仍然饱受争议(popovichandlongbrake,2008)。然而,研究已经表明巨噬细胞渗入导致损伤大小增加,再生纤维的生长下降,以及幸免于初始损伤的神经元死亡增加(fitchetal.,1999;mcphailetal.,2004;donnellyandpopovich,2007)。活化的巨噬细胞和中性白细胞的负面影响被认为由细胞因子、类二十烷酸、自由基和蛋白酶的分泌介导,这些物质对神经元和神经胶质可能均有毒性(donnellyandpopovich,2007)。其中在脊髓损伤后巨噬细胞被清除、抑制或失活的多项研究已经报道了神经保护、再生增加以及运动、感觉和自主功能的改善(oudegaetal.,1999;popovichetal.,1999;mcphailetal.,2004;stirlingetal.,2004)。
技术实现要素:
本发明部分地基于发明人的以下发现:在体外神经胶质疤痕模型中,轴突回缩(顶梢枯死)ed-1+细胞例如活化的巨噬细胞和小神经胶质可通过外部给予某些类型的细胞或培养过所述细胞的条件细胞培养基而减少。这些体外结果也通过应用于体内脊髓损伤模型的细胞得以证实。
发明人发现ed-1+细胞例如活化的巨噬细胞和小神经胶质粘附于营养不良的轴突,并且这一点是回缩必须的。发明人发现将所述细胞或所述细胞的条件培养基应用于营养不良的轴突降低了或预防了粘附。发明人还发现应用培养所述细胞的条件培养基具有神经刺激作用并且显著地增加神经突向外生长/再生。
因此,本发明主要涉及一种治疗(改善或预防)与轴突回缩相关的神经元损伤的方法。
因此,本发明主要涉及一种通过促进损伤中或损伤周围的轴突生长/再生来治疗(改善或预防)神经元损伤的方法。
本发明还主要涉及一种减少神经元损伤中轴突回缩的方法。
本发明还主要涉及一种促进损伤中或损伤周围的轴突生长/再生的方法。
回缩可由ed-1+细胞例如活化的巨噬细胞和/或小神经胶质引起。
本发明还主要涉及一种减少ed-1+细胞与营养不良的轴突的粘附从而减少轴突回缩的方法。
这些结果通过在足够接近所述损伤的位置、以足够的时间和足够的量对损伤给予细胞以促进损伤中或损伤周围的轴突生长/再生而实现。
这些结果通过在足够接近所述损伤的位置、以足够的时间和足够的量对损伤给予细胞以减少神经元损伤中的轴突回缩并减少与轴突回缩相关的神经元损伤而实现。
这些结果通过在足够接近所述损伤的位置、以足够的时间和足够的量对损伤给予细胞以减少ed-1+细胞与营养不良的轴突的粘附——所述粘附会导致轴突回缩——而实现。
所述细胞被引进至损伤的轴突从而所述细胞减少原有ed-1+细胞与轴突的粘附。
ed-1+细胞包括但不限于巨噬细胞和小神经胶质。
这些结果由所述细胞分泌的因子造成。因此,所述结果还通过使用条件细胞培养基或其级分或源自所述条件培养基的蛋白或其他因子而实现。所述条件培养基通过在细胞培养中培养这样的细胞而产生,即所述细胞可有效减少粘附和轴突回缩并且/或者促进轴突生长。在一个实施方案中,所述条件培养基在用前不冷冻。
这些结果还通过使用细胞裂解物或细胞级分而实现。
可以在对应于轴突回缩及由其导致的损伤的各时间点,例如在急性损伤时给予上述细胞、分泌的因子、级分等,以在初始急性损伤后延续一段时间(例如数周)。
然而,应理解轴突回缩也可在慢性损伤情况下发生,例如下面描述的那些。在慢性损伤中,可按照任何可减少轴突回缩的方案给予所述细胞。
由于所述细胞(及分泌的因子)也会促进轴突生长,因此也可在慢性和急性损伤中给予所述细胞(及分泌的因子),所述损伤不一定与回缩相关。对这些损伤进行治疗从而在损伤中或损伤周围使得和促进轴突生长/再生。
在一个实施方案中,所述细胞为干细胞。干细胞包括但不限于胚胎干细胞和非胚胎干细胞。与胚胎干细胞一样,非胚胎干细胞可具有分化为不止一种胚层的细胞类型和/或表达与分化为不止一种胚层的细胞类型的潜力有关的一个或多个标志物的能力。非胚胎细胞还包括组织特异性干细胞,即具有分化为仅一种胚层的细胞类型的能力的细胞,例如造血干细胞、神经干细胞和间充质干细胞。
在一个具体实施方案中,所述非胚胎干细胞已被命名为“多能成体祖细胞”("mapc")并被记载于u.s.7,015,037。
本发明涵盖形成轴突营养不良的任何神经系统损伤,其中ed-1+细胞例如活化的巨噬细胞或小神经胶质与所述营养不良的轴突相互作用并导致所述轴突回缩。这包括中枢神经系统组织,包括脑和脊髓。与营养不良的轴突有关的病症包括但不限于:影响脊髓的任何类型的外伤(这些包括任何来自脊髓外的力(包括椎间盘突出))或者来自脊髓内的脊髓损伤,例如脊髓空洞症;脑内或脑外的任何类型的外伤影响产生的脑损伤(例如头部创伤);整个中枢神经系统的中风(缺血性或溶血性的);多发性硬化;癫痫;神经变性疾病如阿耳茨海默氏病、帕金森氏病、肌萎缩侧索硬化症(葛雷克氏症)和克罗伊茨费尔特-雅各病(cjd)。
附图说明
图1-脊髓损伤后再生障碍的示意图。
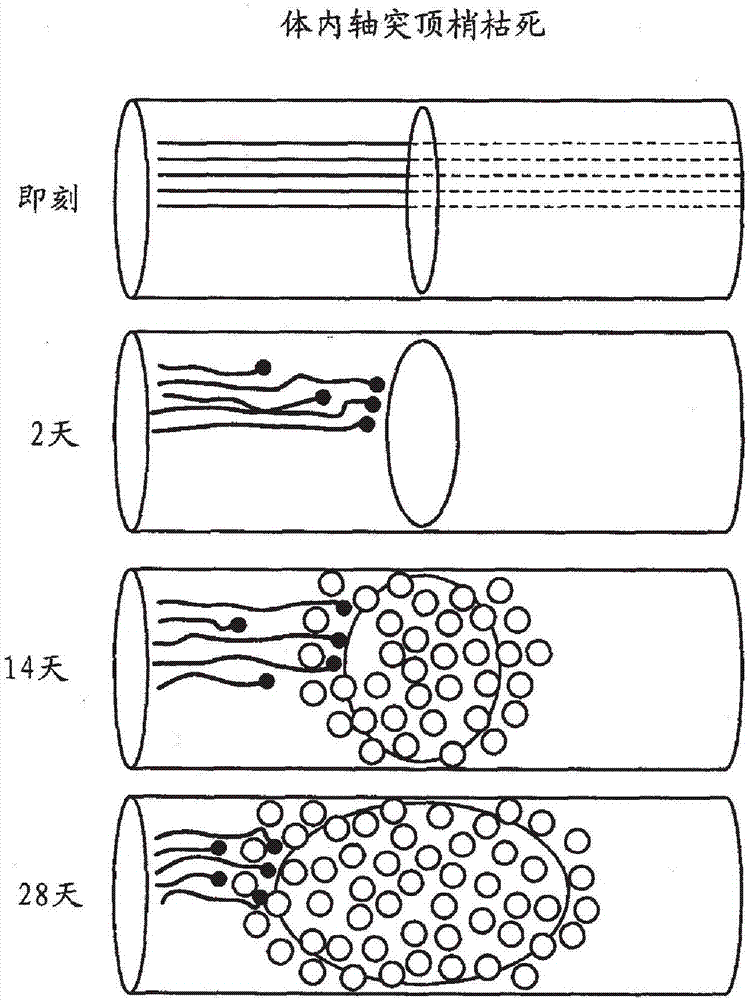
图2-体内轴突顶梢枯死的示意图。
图3-背柱破碎后巨噬细胞渗入和轴突顶梢枯死的真实图和曲线图。巨噬细胞渗入与脊髓损伤后的轴突回缩相关。脊髓损伤后上行感觉轴突随时间大幅回缩。所示a、b为损伤后2天(a)和7天(b)20μm厚的背柱破碎(dcc)脊髓损伤的纵向切片的组合图。dex-tr,texasred缀合的葡聚糖3000mw。切片的方向使得尾端在图像左侧,嘴端在右侧。下面的白色框表示轴突相对损伤中心(点线)的位置,同时具有来自一只动物在每个时间点的多张切片的叠加纤维描线。标尺刻度线表示200μm的增量。a,在损伤后2天,背根神经节轴突(红色)已从损伤中心处的初始切断位置——由有gfap+活性的星形细胞标记(蓝色)——回缩了一段较短的距离。在所述损伤内存在少数ed-1+细胞(绿色),其最可能是活化的小神经胶质。b,到损伤后7天,损伤的轴突(红色)已从损伤中心大幅回缩。现在损伤和周围组织充满了ed-1+细胞(绿色),其主要为渗入的巨噬细胞,而有活性的星形细胞(蓝色)已离开损伤中心。c,表示平均轴突回缩随时间的曲线图。回缩主要发生在损伤后前7天中;然而,损伤后,回缩会一直持续,最长达28天,这是研究的时长。轴突回缩(黑色曲线图)如下:第2天与第7、14和28天显著不同(单因素anova,f(4,40)=6.50,p<0.001;tukey事后检验#p<0.05,*p<0.01,**p<0.001)。巨噬细胞清除(红色曲线图)如下:第2天与第7、14和28天显著不同;第4天与第14和28天显著不同;第7天与第2和14天显著不同(单因素anova,f(4,40)=22.83,p<0.001;tukey事后检验,*p<0.01,**p<0.001,***p<0.0001)。误差棒表示标准误差。比例尺:a,b,250μm。
图4-巨噬细胞体外诱导神经元顶梢枯死的缩时组合图。在神经胶质疤痕的体外模型中,巨噬细胞诱导营养不良的成体背根神经节轴突的大幅回缩。a,缩时动画的6幅单帧图像的组合图,其中将nr8383巨噬细胞加至营养不良的成体背根神经节神经元的培养物,所述神经元在生长促进胞外基质分子层粘连蛋白和强抑制性硫酸软骨素蛋白聚糖聚集蛋白聚糖上培养。每帧的时间在每幅图像的右下角给出,箭头标出生长锥的中心区域。星号标记培养物中的一个固定点,作为变帧过程中的生长锥位置的参照。在30分钟时加入巨噬细胞后巨噬细胞-生长锥接触立即开始。在61分钟时观察到第二个巨噬细胞与该营养不良的轴突之间的物理接触。当在110分钟回缩开始时,其他巨噬细胞物理地改变了轴突轨迹。在150分钟时,生长锥被多个巨噬细胞所遮挡并已几乎回缩到该帧之外。比例尺,20μm。b,追踪a中整个缩时动画的生长锥的位置曲线图。每一点均代表单帧(每30秒)的生长锥中心区域的位置。在接触巨噬细胞后,轴突发生约100μm的大幅回缩。c,另一示例性缩时实验的位置曲线图。
图5-轴突与巨噬细胞之间形成的接触。在神经胶质疤痕的体外模型中,巨噬细胞与营养不良的轴突物理地相互作用。a,从巨噬细胞物理地接触营养不良的轴突的缩时动画中选帧。在回缩发生之前,当轴突从基底上抬起并严重弯曲(箭头)时,生长锥仍然附着。b,图3a的第三幅图像的更高放大率的图像。在巨噬细胞和营养不良的轴突之间发生几处粘附性接触。箭头标示当巨噬细胞移开轴突时,由这些接触形成的膜突起。c,加入巨噬细胞(绿色)后2.5小时,成体drg神经元(红色)的培养物的40×共焦z-stack三维重构。观察到巨噬细胞与营养不良的生长锥直接接触。d,c围绕x轴旋转90°产生的所述三维重构的侧视图。箭头标示已被临近的巨噬细胞(绿色)从基底抬起的神经元突起(红色)。比例尺:a,b,20μm;c,50μm。
图6-mmp9抑制剂阻止巨噬细胞接触所致轴突顶梢枯死的缩时组合图。
图7-评估外加的活细胞(mapc)或条件培养基对巨噬细胞诱导的背根神经节(drg)神经元顶梢枯死的作用的实验设计。
图8-与drg共培养的mapc的缩时组合图,示出mapc的加入阻止巨噬细胞诱导的顶梢枯死。在加入巨噬细胞前的1天给予mapc。
图9-实验缩时组合图,显示mapc条件培养基阻止巨噬细胞诱导的轴突顶梢枯死。在加入条件培养基30分钟后加入巨噬细胞。
图10-显示以mapc条件培养基刺激的巨噬细胞不诱导轴突顶梢枯死的缩时组合图。
图11-mapc阻止巨噬细胞介导的轴突顶梢枯死的图示。
图12-用mapc或条件培养基的体外实验的实验总结。
图13-mapc阻止背柱破碎损伤后巨噬细胞介导的轴突顶梢枯死并促进再生进入损伤中心。移植了媒介(vehicle)对照或mapc的损伤后7天脊髓切片的曲线图和真实图。在神经胶质疤痕的体外模型中,巨噬细胞诱导营养不良的成体背根神经节轴突的大幅回缩。将nr8383巨噬细胞加至营养不良的成体背根神经节神经元的培养物,所述神经元在生长促进胞外基质分子层粘连蛋白和强抑制性硫酸软骨素蛋白聚糖聚集蛋白聚糖的相反斑点梯度上培养。位置图追踪整个缩时动画的生长锥。每一点代表单帧(每30秒)的生长锥中心区域的位置。在接触巨噬细胞后,轴突发生约100μm的大幅回缩。
画面示出了损伤后7天20μm厚的背柱破碎(dcc)脊髓损伤的纵向切片的10×组合图。以texasred缀合的3000mw葡聚糖标记纤维,并且以ed-1+(紫色)可视化巨噬细胞。切片的方向使得尾端在图像左侧,嘴端在右侧。损伤中心标记在下面(黑实线),同时具有来自一只动物在每个时间点的多张切片的3条叠加纤维描线。a,在损伤后7天并且在只注射媒介的情况下,背根神经节轴突(红色)已从损伤中心处的初始切断位置大幅回缩了一段距离。b,到损伤后7天并且在mapc移植物的情况下,损伤的轴突(红色)已大量再生进入损伤。c,表示在接受媒介对照或mapc移植物的动物在损伤后2、4和7天的平均轴突回缩的图表。通过一般线性模型,所述条件、mapc移植物与媒介对照显著不同,*p<0.0001。比例尺:a,b,200μm。
图14-显示ng2+神经胶质细胞不阻止巨噬细胞诱导的顶梢回缩的缩时组合图。ng2+细胞可使轴突稳定,但在体外不阻止巨噬细胞攻击后巨噬细胞介导的回缩。a,显示体外2日后在聚集蛋白聚糖和层粘连蛋白的梯度上β微管蛋白+(红色)轴突与ng2+(绿色)细胞的轴突联系的40×共焦图像。边缘以白色点线标示。b,ng2+细胞表达波形蛋白(红色)。c,ng2+细胞表达巢蛋白(nestin)(红色)。d,显示在成体小鼠脊髓ng2+细胞的存在下在聚集蛋白聚糖/层粘连蛋白的梯度上巨噬细胞/轴突的相互作用的缩时动画中的6帧示例性图像。将nr8383巨噬细胞加至成体drg神经元的2div培养物中。每帧的时间在每幅图像的右下角给出,星号标记培养皿上的一个固定点,作为变帧过程中位置的参照。箭头标示生长锥的中心区域。观察30分钟后加入巨噬细胞,第一次接触发生在103分钟。到110分钟,轴突已经发生长距离的回缩。空心箭头标示回缩纤维的存在。e,d中所示的延缩时动画的每帧(30秒)的生长锥位置的曲线图。红色弧线代表所述斑点的内边缘的位置。箭头表示初始生长轨迹。f,在集聚蛋白聚糖/层粘连蛋白的斑点梯度上与ng2+细胞的共培养的6个营养不良的轴突在与巨噬细胞接触后距离原点的距离。箭头标示轴突已经回缩至ng2细胞的位置。比例尺:a,b,c,50μm。d,20μm。
图15-在斑点梯度上单独培养的mapc的共焦图像以及在斑点梯度上与神经元一起培养的mapc的更高放大率的图像。
在集聚蛋白聚糖——通过cs56(红色)可视化——和层粘连蛋白的双向梯度上培养的gfp+mapc(绿色)的10×共焦图像。与通过β微管蛋白(蓝色)可视化的成体drg神经元共培养的mapc的40×共焦图像。在体外2日后,成体drg和mapc均不通过抑制性斑点边缘。
加至集聚蛋白聚糖斑点梯度的mapc未进入抑制性边缘,但良好地粘附于斑点中心并与成体drg轴突连接。
图16-对照培养基或mapc条件培养基在体外对轴突向外生长的作用的曲线图和真实图。
图17-显示对照培养基不阻止巨噬细胞诱导的顶梢枯死的实验的缩时组合图。在加入条件培养基30分钟后加入巨噬细胞。
虽然存在对照mapc培养基,但是在神经胶质疤痕的体外模型中,巨噬细胞仍诱导营养不良的成体背根神经节轴突的大幅回缩。a,缩时动画的6幅单帧图像的组合图,其中将nr8383巨噬细胞加至营养不良的成体背根神经节神经元的培养物,所述神经元在生长促进胞外基质分子层粘连蛋白和强抑制性硫酸软骨素蛋白聚糖聚集蛋白聚糖的相反斑点梯度上培养。每帧的时间在每幅图像的右下角给出,箭头标出生长锥的中心区域。星号标记培养物中的一个固定点,作为变帧过程中的生长锥位置的参照。比例尺,20μm。b,追踪a中整个缩时动画的生长锥的位置曲线图。每一点代表单帧(每30秒)的生长锥中心区域的位置。在接触巨噬细胞后,轴突发生约80μm的大幅回缩。
直接向所述缩时培养皿中加入mapc条件培养基导致了生长锥形态的改变,从营养不良的、停滞的状态变为能动的、扁平的状态。巨噬细胞仍然接触这些轴突,但接触通常是瞬时的并且通常不导致轴突回缩。对照mapc培养基不阻止轴突回缩。以mapc条件培养基预处理的巨噬细胞也在所述斑点上接触轴突,但不引起回缩(图9-12)。mapc可能作用于巨噬细胞,从而改变它们的受体表达、对损伤细胞的响应或者mmp-9的分泌。
具体实施方式
定义
"a"或"an"在本文中是指一个或多个;至少一个。当本文中使用复数形式时,通常包括单数形式。
本文使用的术语“粘附、附着、粘着”等,是指连接足够长的时间以诱导轴突回缩。如本文进一步描述的,应理解物理接触可发生在巨噬细胞(或其他细胞)与营养不良的轴突之间,所述接触是短暂的并且不导致轴突回缩。在本发明的上下文中,被本发明试剂降低或阻止的附着是持续足够长的时间以诱导轴突回缩的附着。因此,本发明不排除允许营养不良的轴突与ed-1+细胞之间物理接触的试剂。因此,本发明涵盖允许接触(例如短暂的物理接触)但不允许附着足够长的时间以导致轴突顶梢枯死的试剂。
“共给予”是指彼此结合、一起、配合地给予,包括同时或依次给予两种或多种试剂。
在没有其他限制的情况下,“包含”是指必须包括所指事物,但不对其他可包括的事物进行限制或排除。例如,“包含x和y的组合物”包括含有x和y的任何组合物,而不管所述组合物中是否存在其他组分。同样,“包含步骤x的方法”包括其中进行x(不管x是所述方法中的唯一步骤还是只是步骤之一)的任何方法,不管可能存在多少其他步骤,也不管x与这些步骤相比多么简单或复杂。“包括”和使用词根“含”的类似短语在本文中用作“包含”的同意词并具有相同的含义。
“包括”是“包含”的同义词(见上)。
“条件细胞培养基”是本领域公知的术语,指已经培养过细胞的培养基。在本文中,这是指将所述细胞培养足够的时间以分泌对减少活化的巨噬细胞与营养不良的神经元的粘附和/或促进神经突向外生长/轴突再生有效的因子。
条件细胞培养基是指细胞已经在其中培养从而将因子分泌到所述培养基中的培养基。出于本发明的目的,可使细胞生长历经足够数量的细胞分裂以产生有效量的所述因子,使得所述培养基减少巨噬细胞与营养不良的神经元的粘附并因此减少轴突回缩和/或促进神经突向外生长/轴突再生。通过本领域任何已知的方法将细胞从所述培养基中除去,所述方法包括但不限于离心、过滤、免疫清除(例如通过标记的抗体和磁性柱)和facs分选。
“顶梢枯死”是用于指因轴突损伤而发生的轴突回缩的技术术语。在本发明上下文中,轴突回缩是指因ed-1+细胞并且特别是巨噬细胞和小神经胶质的充分粘附而发生的回缩。所述巨噬细胞和小神经胶质(即ed-1+细胞)不处于休眠或不活动状态。它们是被活化的。术语“活化的”是指这些细胞允许其粘附于营养不良的轴突从而导致轴突回缩的状态。导致体外活化的条件的实例在本申请中有进一步描述。然而,应理解所述活化不限于本文公开的具体条件。
尽管本发明通常具体涉及活化的巨噬细胞(并以其举例),然而这些细胞是一类ed-1+细胞并且本发明也适用于其他这类细胞。一个实例是活化的小神经胶质。
“有效量”通常指提供需要的局部或全身作用的量。例如,有效量是足以实现有益或需要的临床效果的量。所述有效量可以单次给药的形式一次全部提供或以多次给药的形式分份提供。对什么量会被认为是有效量的准确确定可基于每个受试者的个人因素,包括他们的身高体重、年龄、损伤和/或待治疗的疾病或损伤以及遭受损伤或疾病开始后的时间。本领域技术人员能够基于这些本领域中惯常的考虑确定给定受试者的有效量。当用于本文时,“有效剂量”与“有效量”所指相同。
“有效途径”通常是指提供用于将一种试剂送递至所需腔室、系统或位置的途径。例如,有效途径是给予试剂以在所需的作用位点提供量足以实现有益或所需的临床效果的所述试剂的途径。
“ec细胞”是从被称为畸胎瘤的一种癌症的分析中发现的。1964年,研究人员注意到畸胎瘤中的单个细胞可被分离并在培养中保持不分化状态。于是这种类型的干细胞被称为胚胎癌细胞(ec细胞,embryoniccarcinomacell)。
“胚胎干细胞(esc)”为本领域所熟知并多年以来已从多种不同哺乳动物种中制得。胚胎干细胞是源自被称为胚泡的早期胚胎的内细胞团的干细胞。它们能够分化为三种原胚层的所有衍生细胞:外胚层、内胚层和中胚层。这些包括成体内超过220种细胞类型的每一种。es细胞可成为体内除胎盘外的任何组织。只有桑椹胚细胞是全能性的,能够成为所有组织和胎盘。
使用术语“包括”并非意图进行限制。例如,述及抗体抑制剂“包括”片段和变体时不表示排除其他形式的抗体抑制剂。
“诱导的多能干细胞(ipsc或ips细胞)”是例如通过引入外源基因被再编程的体细胞,其中所述外源基因赋予所述体细胞分化程度较低的表型。然后这些细胞可被诱导分化为分化程度较低的后代。使用首先在2006年公开的方法的改进方法获得了ips细胞(yamanaka,s.etal.,cellstemcell,1:39-49(2007))。例如,在一个实例中,为获得ips细胞,科学家以皮肤细胞为起始材料,通过用反转录病毒将基因插入到细胞dna中的标准实验室技术随后对所述皮肤细胞进行了改造。在一个实例中,所插入的基因为oct4、sox2、lif4和c-myc,已知这些基因作为天然调控因子一起作用以使细胞保持在与胚胎干细胞相似的状态。文献已对这些细胞进行了描述。参见,例如,wernigetal.,pnas,105:5856-5861(2008);jaenischetal.,cell,132:567-582(2008);hannaetal.,cell,133:250-264(2008);以及brambrinketal.,cellstemcell,2:151-159(2008)。这些参考文献以引用的方式纳入以教导ipsc和制备它们的方法。也可通过特定培养条件(暴露于特定试剂)获得这些细胞。
术语“分离的”指不与一种或多种细胞或者在体内时与所述一种细胞或多种细胞结合的一种或多种细胞组分结合的一种或多种细胞。“富集群落”指合乎需要的细胞相对体内或原代培养物中的一种或多种其他细胞类型的数量上相对增加。
然而,本文所用的术语“分离的”不表示只存在胚胎干细胞。而是,术语“分离的”表示细胞从其天然组织环境中取出并以相对所述正常组织环境更高的浓度存在。因此,“分离的”细胞群落可进一步包括干细胞以外的细胞类型并可包括其他组织组分。这可用例如细胞的倍增来表示。细胞可在体外或离体进行10、20、30、40次或更多次的倍增,从而相对其在体内或在原始组织环境(例如骨髓、外周血、脂肪组织等)中的原始数量被富集。
“mapc”是“多能成体祖细胞”的首字母缩写。其指非胚胎干细胞。mapc中的术语“成体”是非限制性的。其指非胚胎体细胞。与胚胎干细胞一样,mapc可产生多于一种胚层的细胞谱系。其可在分化时产生全部三种胚层(即内胚层、中胚层和外胚层)的细胞类型。与胚胎干细胞类似,人mapc表达端粒末端转移酶、oct3/4(即oct3a)、rex-1、rox-1和sox-2,并可表达ssea-4。(亦见jiang,y.etal.,nature,418:41(2002);exphematol,30:896(2002))。端粒在mapc中延长并且核型正常。由于注射到哺乳动物中的mapc可迁移至多个器官内并在这些器官内被同化,因此mapc是自我更新的干细胞。对于mapc,“多能”指在分化时产生多于一种原始胚层(即内胚层、中胚层和外胚层)例如全部三种胚层的细胞系的能力。
“神经突向外生长”是指损伤位置处的神经元不仅停止回缩而且生长并延长的性质。
“可药用载体”是本发明中所用细胞的任何可药用介质。所述介质可保持等渗性、细胞代谢、ph等。所述介质与对受试者的体内给药相容,并因此可用于细胞递送和治疗。
可培养并刺激“原始胚胎生殖细胞”(pg或eg细胞)以产生许多分化程度较低的细胞类型。
“祖细胞”是在干细胞分化过程中产生的具有其最终分化后代的某些但非全部特征的细胞。限定的祖细胞——例如“心肌祖细胞”——定向到细胞系,但不定向到特定的或最终分化的细胞类型。当用于缩写"mapc"时,术语“祖”不将这些细胞限定到具体的细胞系。
本文使用的术语“减少”是指阻止以及减轻。当用于治疗的语境时,“减少”是阻止或改善一种或多种临床症状。临床症状是一种(或多种)对或将对——如果不进行治疗——受试者的生命质量(健康)有负面影响的症状。
术语“回缩”是指轴突逐渐远离损伤位置例如神经胶质疤痕形成之处。此处,再生轴突的末端停止延长并出现营养不良。然后,这些营养不良的末端可进一步逐渐远离神经胶质疤痕和损伤位置。
“自我更新”指复制出具有与亲代细胞相同的分化潜能的子代干细胞的能力。在本文中使用的相似的术语是“增殖”。
“干细胞”指可进行自我更新(即后代具有相同的分化潜能)并且也可产生分化潜能受到更多限制的后代细胞的细胞。在本发明的上下文中,干细胞还可包括已经例如通过核移植、通过与更原始的干细胞融合、通过引入特定转录因子或通过在特定条件下培养而已去分化的分化程度更高的细胞。参见,例如wilmutetal.,nature,385:810-813(1997);yingetal.,nature,416:545-548(2002);guanetal.,nature,440:1199-1203(2006);takahashietal.,cell,126:663-676(2006);okitaetal.,nature,448:313-317(2007);以及takahashietal.,cell,131:861-872(2007)。
去分化也可通过给予某些化合物或暴露于可导致去分化的体外或体内物理环境下而导致。干细胞还可源自异常组织,例如畸胎瘤,以及某些其他来源,例如拟胚体(虽然拟胚体可被认为是胚胎干细胞,因为它们源自胚胎组织,尽管不直接来自内细胞团)。干细胞还可通过将与干细胞功能相关的基因引入非干细胞中而产生,例如诱导的多能干细胞。
“受试者”是指脊椎动物,例如哺乳动物,例如人。哺乳动物包括但不限于人、狗、猫、马、牛和猪。
术语“治疗有效量”指被确定在哺乳动物中产生任何治疗性反应的量。例如,有效量的治疗性细胞或细胞相关剂可延长患者的存活和/或抑制明显的临床症状。具有治疗效果的治疗,在该术语用于本文时的含义下,包括改善受试者生命质量的治疗,即使所述治疗不会改善疾病本身的后果。这种治疗有效量由本领域普通技术人员容易地确定。因此,“治疗”是指送递所述量。因此,治疗可阻止或缓解因活化的巨噬细胞与营养不良的轴突的粘附而发生的任何病理症状。治疗还指轴突再生的有益临床效果。
“治疗”在本发明中广泛地使用,这一术语包括预防、缓解、抑制或治愈缺陷、功能障碍、疾病或其他有害过程等,包括干扰治疗和/或由治疗引起的过程。
干细胞
可实施本发明,优选地使用脊椎动物物种例如人、非人灵长类、家畜、牲畜和其他非人哺乳动物的干细胞实施本发明。这些细胞包括但不限于下文所述的细胞。
胚胎干细胞
研究最深入的干细胞是胚胎干细胞(esc),因为其具有无限自我更新和多能分化潜能。这些细胞源自胚泡的内细胞团或者可源自着床后胚胎的原始生殖细胞(胚胎生殖细胞或eg细胞)。es和eg细胞已首先从小鼠中得到,后来从许多不同动物得到,最近还从非人灵长类和人中得到。当引入小鼠胚泡或其他动物的胚泡中时,esc可促使所述动物的所有组织的形成。es和eg细胞可通过用抗ssea1(小鼠)和ssea4(人)的抗体进行阳性染色而被识别。参见例如美国专利no.5,453,357;5,656,479;5,670,372;5,843,780;5,874,301;5,914,268;6,110,7396,190,910;6,200,806;6,432,711;6,436,701,6,500,668;6,703,279;6,875,607;7,029,913;7,112,437;7,145,057;7,153,684和7,294,508,所述专利的每一篇都以引用的方式纳入本文以教导胚胎干细胞及其制备和扩增方法。因此,esc及其分离和扩增方法为本领域所熟知。
若干影响体内胚胎干细胞潜力状态的转录因子和外源细胞因子已被鉴定出。将要描述的与干细胞多能性相关的第一种转录因子是oct4。oct4属于pou(pit-oct-unc)家族的转录因子并且是能够活化在启动子或增强子区内的含有被称为“八聚体基序”的八聚体序列的基因转录的dna结合蛋白。oct4在受精卵的卵裂期表达直至卵圆筒形成。oct3/4的功能是抑制诱导分化的基因(即foxad3、hcg)以及活化促进多能性的基因(fgf4、utf1、rex1)。sox2——高迁移率组(hmg)盒转录因子的一员——与oct4共同作用以活化在内细胞团中表达的基因的转录。将oct3/4在胚胎干细胞中的表达维持某些水平之间是必需的。oct4表达水平>50%的过表达或下调会改变胚胎干细胞的命运,分别形成原始内胚层/中胚层或滋养外胚层。在体内,oct4缺陷胚胎发育至胚泡期,但内细胞团的细胞不具有多能性。相反,它们沿胚外滋养层谱系分化。sall4——一种哺乳动物spalt转录因子——是oct4的上游调节子,从而对在胚胎发生早期维持oct4的适当水平起重要作用。当sall4下降到低于某个阈值时,滋养外胚层细胞会错位扩张到内细胞团中。多能性所需的另一转录因子是nanog,以一个凯尔特人部落"tirnanog":永远年轻的土地命名。在体内,nanog从致密桑椹胚期表达,随后被限定于内细胞团并在着床期下调。nanog的下调可对于防止多能性细胞不受控制的扩增以及允许原肠胚形成期间的多谱系分化起重要作用。在5.5天分离的无nanog的胚胎由无组织的胚泡构成,主要包含胚外内胚层,而不含可辨别的上胚层。
非胚胎干细胞
干细胞已在大多数组织中被发现。也许表征最好的是造血干细胞(hsc)。hsc是源自中胚层的细胞,其可使用细胞表面标志物和功能性特征进行纯化。它们已从骨髓、外周血、脐带血、胎肝和卵黄囊中分离。它们发起血细胞生成并产生多个造血谱系。当移植到受到致死辐射的动物中时,它们可重建红血球、中性白细胞-巨噬细胞、巨核细胞和淋巴造血细胞池。它们也可被诱导已进行某些自我更新细胞分裂。参见例如美国专利no.5,635,387;5,460,964;5,677,136;5,750,397;5,681,599和5,716,827。美国专利no.5,192,553报道了分离人新生儿或胎儿造血干细胞或祖细胞的方法。美国专利no.5,716,827报道了为thy-1+祖细胞的人造血细胞和在体外再生所述细胞的合适的生长培养基。美国专利no.5,635,387报道了培养人造血细胞及其前体的方法和设备。美国专利no.6,015,554记载了重构成人淋巴和树突细胞的方法。因此,hsc及其分离和扩增方法为本领域所熟知。
本领域熟知的另一种干细胞是神经干细胞(nsc)。这些细胞可在体内增殖并持续再生至少某些神经细胞。当离体培养时,神经干细胞可被诱导进行增殖以及分化为不同类型的神经元和神经胶质细胞。当移植到脑中时,神经干细胞可定植并产生神经细胞和神经胶质细胞。参见,例如gagef.h.,science,287:1433-1438(2000),svendsens.n.etal,brainpathology,9:499-513(1999),和okabes.etal.,mechdevelopment,59:89-102(1996)。美国专利no.5,851,832报道了从脑组织获取的多能性神经干细胞。美国专利no.5,766,948报道了从新生大脑半球产生成神经细胞。美国专利no.5,564,183和5,849,553报道了哺乳动物神经嵴干细胞的用途。美国专利no.6,040,180报道了从哺乳动物多能性cns干细胞培养物体外产生分化的神经元。wo98/50526和wo99/01159报道了神经上皮干细胞、少突胶质细胞-星形细胞前体和谱系限制性神经元前体的产生和分离。美国专利no.5,968,829报道了从胚胎前脑获取的神经干细胞。因此,神经干细胞及其制备和扩增方法为本领域所熟知。
本领域广泛研究的另一种干细胞是间充质干细胞(msc)。msc来源于胚胎中胚层并可从许多来源分离,包括成体骨髓、外周血、脂肪、胎盘、脐带血等。msc可分化为许多中胚层组织,包括肌肉、骨、软骨、脂肪和腱。有大量关于这些细胞的文献。参见例如美国专利no.5,486,389;5,827,735;5,811,094;5,736,396;5,837,539;5,837,670;和5,827,740。也参见pittenger,m.etal,science,284:143-147(1999)
成体干细胞的另一实例是脂肪来源的成体干细胞(adsc),其已从脂肪分离,通常是通过吸脂术继而使用胶原酶释放adsc。adsc在许多方面类似于来源于骨髓的msc,除了可从脂肪分离更多的细胞之外。这些细胞已被报告可分化为骨、脂肪、肌肉、软骨和神经元。一种分离方法记载于u.s.2005/0153442。
本领域已知的其他干细胞包括胃肠干细胞、表皮干细胞和肝干细胞,其中肝干细胞也被称为“卵圆细胞”(potten,c,etal.,transrsoclondbbiolsci,353:821-830(1998),watt,f.,transrsoclondbbiolsci,353:831(1997);alisonetal.,hepatology,29:678-683(1998))。
被报道能够分化为一种以上胚层细胞类型的其他非胚胎细胞包括但不限于来自脐带血(参见美国专利公开no.2002/0164794)、胎盘(参见美国专利公开no.2003/0181269)、脐带基质(mitchelletal.,stemcells,21:50-60,2003)、小胚胎样干细胞(kuciaetal.,jphysiolpharmacol,57suppl5:5-18,2006)、羊水干细胞(atala,a.,jtissueregenmed1:83-96,2007)、源自皮肤的前体细胞(tomaetal.,natcellbiol3:778-784,2001)以及骨髓(参见美国专利公开no.2003/0059414和2006/0147246)的细胞,上述文献的每一篇都以引用的方式纳入本文中以教导这些细胞。
对体细胞再编程的方法
若干不同方法,例如核移植、细胞融合和培养诱导的再编程,已用于诱导已分化细胞转化为胚胎状态。核移植包括将体细胞核注射到去核卵母细胞中,若将所述卵母细胞转移到代孕母体中,其可长成克隆(“生殖性克隆”),或者,若外植培养可长成遗传匹配的胚胎干(es)细胞("体细胞核移植",scnt)。体细胞与es细胞的细胞融合产生具有多能es细胞全部特征的杂交细胞。体细胞的外植培养选择出可具有多能性的永生细胞系。目前,精原干细胞是可从出生后动物中得到的多能细胞的唯一来源。用限定的因子转导体细胞可启动产生多能状态的再编程。这些实验方法已被广泛地综述(hochedlingerandjaenisch,nature441:1061-1067(2006)和yamanaka,s.,cellstemcell1:39-49(2007))。
核移植
核移植(nt),也称为体细胞核移植(scnt),是指将供体体细胞的细胞核引入去核卵母细胞中以长成克隆动物例如多莉羊(wilmutetal.,nature385:810-813(1997))。通过nt实现的活体动物的产生表明了体细胞(包括最终分化的细胞)的外遗传状态是稳定的,但并非被不可逆地固定,而是可被再编程为胚胎状态,所述胚胎状态能够指导新生物体的发育。除了提供阐明胚胎发育和疾病涉及的基本外遗传机制的令人激动的实验方法之外,核克隆技术还可能引起患者特异性移植医学的兴趣。
体细胞和胚胎干细胞的融合
体细胞核到未分化状态的外遗传再编程已在通过胚胎干细胞与体细胞的融合产生的鼠杂种中被证明。多种体细胞和胚胎癌细胞、(solter,d.,natrevgenet,7:319-327(2006)、胚胎生殖细胞(eg)或es细胞(zwakaandthomson,development,132:227-233(2005))的杂交细胞与亲代胚胎细胞具有许多相同的性质,表明在这些融合产物中多能性表型是显性的。与小鼠(tadaetal.,currbiol,11:1553-1558(2001))一样,人es细胞具有在融合后对体细胞核再编程的潜力(cowanetal.,science,309:1369-1373(2005));yuetal.,science,318:1917-1920(2006))。沉默多能性标志物例如oct4的活化或失活体x染色体的再活化为杂交细胞中体基因组的再编程提供了分子证据。已经表明dna复制对于多能性标志物——其在融合后2天首次观察到——的活化是必需的(doandscholer,stemcells,22:941-949(2004));并且当与神经干细胞融合时,es细胞中nanog的强制过表达促进多能性(silvaetal.,nature,441:997-1001(2006))。
培养诱导的再编程
已从胚胎来源获得了多能细胞,所述胚胎来源例如胚泡的卵裂球和内细胞团(icm)(es细胞)、上胚层(episc细胞)、原始生殖细胞(eg细胞)和出生后精原干细胞("magscsm"“es样”细胞)。多能细胞以及它们的供体细胞/组织如下:单性生殖es细胞源自鼠卵母细胞(narasimhaetal.,currbiol,7:881-884(1997));胚胎干细胞源自卵裂球(wakayamaetal.,stemcells,25:986-993(2007));内细胞团细胞(来源不适用)(egganetal.,nature,428:44-49(2004));胚胎生殖细胞和胚胎癌细胞源自原始生殖细胞(matsuietal.,cell,70:841-847(1992));gmcs、massc和masc源自精原干细胞(guanetal.,nature,440:1199-1203(2006);kanatsu-shinoharaetal.,cell,119:1001-1012(2004);和seandeletal.,nature,449:346-350(2007));episc细胞源自上胚层(bronsetal.,nature,448:191-195(2007);tesaretal.,nature,448:196-199(2007));单性生殖es细胞源自人卵母细胞(cibellietal.,science,295l819(2002);revazovaetal.,cloningstemcells,9:432-449(2007));人es细胞源自人胚泡(thomsonetal.,science,282:1145-1147(1998));mapc源自骨髓(jiangetal.,nature,418:41-49(2002);phinneyandprockop,stemcells,25:2896-2902(2007));脐带血细胞(源自脐带血)(vandevenetal.,exphematol,35:1753-1765(2007));神经球源细胞源自神经细胞(clarkeetal.,science,288:1660-1663(2000))。已知来自生殖细胞系的供体细胞例如pgc或精原干细胞在体内是单能性的,然而已证明可在延长体外培养后分离多能es样细胞(kanatsu-shinoharaetal.,cell,119:1001-1012(2004))或magsc(guanetal.,nature,440:1199-1203(2006))。尽管这些多能细胞类型中的大多数能够在体外分化并形成畸胎瘤,然而按照更严格的标准,只有es、eg、ec和源自精原干细胞的magcs或es样细胞是多能性的,因为它们能够形成出生后嵌合体并形成种系。最近,从成体小鼠的睾丸精原干细胞获得了多能成体精原干细胞(masc),这些细胞的表达谱不同于es细胞(seandeletal.,nature,449:346-350(2007)),但类似于episc细胞,episc细胞源自着床后小鼠胚胎的上胚层(bronsetal.,nature448:191-195(2007);tesaretal.,nature,448:196-199(2007))。
通过限定的转录因子进行的再编程
takahashi和yamanaka已经报道了将体细胞再编程回es样状态(takahashiandyamanaka,cell,126:663-676(2006))。在四种转录因子oct4、sox2、c-myc和klf4的病毒介导的转导继而就oct4靶向基因fbxl5的活化进行筛选后,他们成功地将小鼠胚胎成纤维细胞(mef)和成体成纤维细胞再编程为多能es样细胞(图2a)。已活化fbxl5的细胞被称为ips(诱导的多能干)细胞并通过其形成畸胎瘤的能力显示具有多能性,尽管它们不能产生活嵌合体。此多能性状态倚赖转导的oct4和sox2基因的持续病毒表达,而内源oct4和nanog基因不表达或以较es细胞中为低的水平表达,并且它们各自的启动子被发现在很大程度上甲基化。这与fbxl5-ips细胞与es细胞不相应但可表现不完全再编程状态的结论一致。尽管遗传实验已证明oct4和sox2对于多能性是必需的(chambersandsmith,oncogene,23:7150-7160(2004);ivanonaetal.,nature,442:5330538(2006);masuietal.,natcellbiol,9:625-635(2007)),然而两种致癌基因c-myc和klf4在再编程中的作用尚不清楚。实际上,这些致癌基因中的某些可能对于再编程是非必需的,因为小鼠和人ips细胞均已在不进行c-myc转导的情况下得到,尽管效率较低(nakagawaetal.,natbiotechnol,26:191-106(2008);werningetal.,nature,448:318-324(2008);yuetal.,science,318:1917-1920(2007))。
mapc
mapc是“多能成体祖细胞”(非es、非eg、非生殖细胞)的首字母缩写。mapc具有分化为至少两种例如全部三种原始胚层(外胚层、中胚层和内胚层)的细胞类型的能力。在mapc中也发现了在es细胞中发现的基因(例如端粒末端转移酶、oct3/4、rex-1、rox-1、sox-2)。oct3/4(在人中为oct3a)似乎是es和生殖细胞特异的。mapc代表一种比msc更原始的祖细胞群体,并且具有包括上皮、内皮、神经、肌原、生血、生骨、肝原、软骨形成和生脂的细胞系的分化能力(verfaillie,cm.,trendscellbiol12:502-8(2002),jahagirdar,b.n.,etal.,exphematol,29:543-56(2001);reyes,m.andcm.verfaillie,annnyacadsci,938:231-233(2001);jiang,y.etal.,exphematol,30896-904(2002);和jiang,y.etal.,nature,418:41-9.(2002))。
美国专利7,015,037和美国专利申请no.10/467,963记载了人mapc。在其他哺乳动物中已鉴定出mapc。例如,美国专利7,015,037和美国专利申请no.10/467,963还记载了鼠mapc。美国专利申请no.10/467,963还记载了大鼠mapc。
这些文献记载由catherineverfaillie分离的mapc的内容以引用的方式纳入本文。
mapc的分离和生长
mapc分离方法为本领域所知。参见,例如美国专利7,015,037和美国专利申请no.10/467,963,这些方法连同mapc的表征(表型)以引用的方式纳入本文。mapc可从多种来源中分离,所述来源包括但不限于骨髓、胎盘、脐带和脐带血、肌肉、脑、肝脏、脊髓、血液或皮肤。因此,可得到骨髓抽出物、脑或肝活组织以及其他器官,并使用本领域技术人员可采用的阳性或阴性选择技术、根据在这些细胞中表达(或不表达)的基因分离所述细胞(例如通过功能或形态测定例如上面引用的申请中公开的那些,所述申请以引用的方式纳入本文)。
如美国专利7,015,037所述的来自人骨髓的mapc
mapc不表达共同白细胞抗原cd45或成红细胞特异性血型糖蛋白-a(gly-a)。对混合的细胞群体进行聚蔗糖-泛影葡胺分离。然后使用抗cd45和抗gly-a的抗体对所述细胞进行阴性选择,清除cd45+和gly-a+细胞群体,然后回收剩余的大约0.1%的骨髓单核细胞。也可将细胞铺板于纤连蛋白包被的孔中并如下所述培养2-4周以清除cd45+细胞和gly-a+细胞。在贴壁骨髓细胞的培养中,许多贴壁基质细胞在细胞倍增约30次发生复制衰老,更纯系的细胞群体继续扩增并保持长端粒。
或者,阳性选择可用于通过细胞特异性标志物的结合来分离细胞。阳性和阴性选择技术对本领域技术人员都是可用的,适于阴性选择目的的多种单克隆和多克隆抗体也是本领域可获得的(参见,例如leukocytetypingv,schlossman,etal.,eds.(1995)oxforduniversitypress)并可从若干来源购得。
从细胞群体的混合物中分离哺乳动物细胞的技术还在schwartz等人的美国专利no.5,759,793(磁性分离)、baschetal.,1983(免疫亲和色谱)以及wysockiandsato,1978(荧光活化的细胞分选)中有所记载。
如美国专利7,015,037所述培养mapc
如本文所述分离的mapc可使用本文和美国专利7,015,037公开的方法培养,该专利以引用的方式纳入本文以教导这些方法。
其他培养方法
在另外的实验中,mapc的培养密度可从约100个细胞/cm2到约150个细胞/cm2到约10,000个细胞/cm2变化,包括约200个细胞/cm2到约1500个细胞/cm2到约2000个细胞/cm2。所述密度可因物种不同而变化。此外,最佳密度可根据培养条件和细胞来源而变化。为给定的培养条件和细胞确定最佳密度在本领域普通技术人员的能力范围之内。
此外,在培养中mapc的分离、生长和分化过程中的任意时间都可使用低于约10%,包括约1-5%,特别是3-5%的有效大气氧浓度。
细胞可在不同血清浓度下培养,例如约2-20%。可使用胎牛血清。较高的血清——例如约15-20%——可结合较低的氧压使用。在附着于培养皿之前不需选择细胞。例如,在聚蔗糖梯度离心后,可将细胞直接铺板,例如250,000-500,000/cm2。可挑取、可能合并并且扩增附着的克隆。
在实施例的实验过程中使用的一个实施方案中,使用了高血清(约15-20%)和低氧(约3-5%)的条件进行细胞培养。具体地,将来自集落的附着细胞在18%血清和3%氧气(含有pdgf和egf)中以约1700-2300细胞/cm2的密度铺板并传代。
在针对mapc的实施方案中,添加物为可使mapc保留分化为全部三种细胞谱系的能力的细胞因子或组分。这可通过未分化状态的特异性标志物的表达来指示。例如,mapc组成型地表达oct3/4(oct3a)并保持高水平的端粒末端转移酶。
细胞培养
通常,用于本发明的细胞可在本领域可获得并熟知的培养基中维持和扩增。这些培养基包括但不限于dulbecco'smodifiedeagle's
也考虑用哺乳动物血清补充细胞培养基。血清通常含有对存活和扩增必需的细胞因子和组分。血清的实例包括胎牛血清(fbs)、牛血清(bs)、小牛血清(cs)、小胎牛血清(fcs)、新生小牛血清(ncs)、山羊血清(gs)、马血清(hs)、人血清、鸡血清、猪血清、绵羊血清、兔血清、血清替代物和牛胚胎液(embryonicfluid)。应理解,如果认为必须灭活补体级联的组分,那么可在55-65℃加热灭活血清。
也可有利地使用其他补充物来为细胞提供必需的微量元素以优化生长和扩增。这些补充物包括胰岛素、转铁蛋白、硒化钠及其结合物。这些组分可包含在盐溶液中,例如但不限hanks'balancedsalt
激素也可有利地用于细胞培养,包括但不限于d-醛甾酮、乙酚(des)、地塞米松、β-雌二醇、氢化可的松、胰岛素、催乳素、孕酮、生长抑素/人生长激素(hgh)、促甲状腺素、甲状腺素和l-甲状腺素。
根据细胞类型和分化细胞的命运,还可使用脂质和脂质载体来补充细胞培养基。这类脂质和载体可包括但不限于环糊精(α、β、γ)、胆固醇、与白蛋白缀合的亚油酸、与白蛋白缀合的亚油酸和油酸、非缀合的亚油酸、与白蛋白缀合的亚油酸-油酸-花生四烯酸、非缀合的与白蛋白缀合的油酸等。
也考虑使用饲养细胞层。饲养细胞用于支持需要复杂营养的培养细胞特别是es细胞的生长。饲养细胞是通过γ辐射灭活的正常细胞。在培养中,饲养层作为其他细胞的基底层并供给细胞因子,而它们自身不再生长或分裂(lim,j.w.andbodnar,a.,2002)。饲养层细胞的实例一般有人二倍体肺细胞、小鼠胚胎成纤维细胞、瑞士小鼠胚胎成纤维细胞,也可以是任何能够供给对实现干细胞最佳的生长、活力和扩增有利的细胞成分和因子的有丝分裂期后细胞。在许多情况下,饲养细胞层不必使es细胞保持在未分化的、增殖性状态下,因为白血病抑制性因子(lif)具有抗分化性质。因此,补充lif可用于将某些物种的mapc保持在未分化状态下。
细胞可在低血清或无血清的培养基中培养。美国专利7,015,037记载了用于培养mapc的无血清培养基。已在低血清或无血清的培养基中培养了许多细胞。在这种情况下,在所述培养基中添加一种或多种生长因子。常用的生长因子包括但不限于成骨蛋白、碱性成纤维细胞生长因子、血小板衍生生长因子和表皮生长因子。参见例如美国专利no.7,169,610;7,109,032;7,037,721;6,617,161;6,617,159;6,372,210;6,224,860;6,037,174;5,908,782;5,766,951;5,397,706和4,657,866;所述全部专利都以引用的方式纳入本文中以教导在无血清培养基中培养细胞。
培养中的细胞可维持在悬浮液中或贴壁于固体支持物,例如胞外基质成分。干细胞经常需要促进其附着于固体支持物的另外的因子,例如i型和ii型胶原、硫酸软骨素、纤接蛋白、“超纤连蛋白(superfibronectin)”和纤接蛋白样聚合物、明胶、聚-d-和聚-l-赖氨酸、血小板反应蛋白和玻璃粘连蛋白。本发明的一个实施方案利用纤连蛋白。参见,例如ohashietal.,naturemedicine,13:880-885(2007);matsumotoetal.,jbioscienceandbioengineering,105:350-354(2008);kirouacetal.,cellstemcell,3:369-381(2008);chuaetal.,biomaterials,26:2537-2547(2005);drobinskayaetal.,stemcells,26:2245-2256(2008);dvir-ginzbergetal.,fasebj,22:1440-1449(2008);turneretal.,jbiomedmaterrespartb:applbiomater,82b:156-168(2007);和miyazawaetal.,journalofgastroenterologyandhepatology,22:1959-1964(2007)。
细胞也可在"3d"(聚集的)培养中培养。一个实例为2008年1月18日提交的美国临时专利申请no.61/022,121。
一旦在培养中形成,细胞可新鲜使用或冷冻使用,并使用例如含有40%fcs和10%dmso的dmem作为冷冻原种保存。本领域技术人员也可使用制备培养细胞的冷冻原种的其他方法。
药物制剂
在某些实施方案中,纯化的细胞群体存在于适应和适于送递即生理上相容的组合物中。因此,所述干细胞群体的组合物通常还包括一种或多种缓冲剂(例如中性缓冲盐水或磷酸缓冲盐水)、碳水化合物(例如葡萄糖、甘露糖、蔗糖或葡聚糖)、甘露醇、蛋白、多肽或氨基酸例如甘氨酸、抗氧化剂、抑菌剂、螯合剂例如edta或谷胱甘肽、佐剂(例如氢氧化铝)、使所述制剂与受者的血液相比等渗、低渗或稍微高渗的溶质、助悬剂、增稠剂和/或防腐剂。
在其他实施方案中,纯化的细胞群体存在于适应或适于冷冻或保存的组合物中。
在许多实施方案中,用于给予受试者的细胞(或条件培养基)的纯度为约100%。在其他实施方案中,其为95-100%。在某些实施方案中,其为85-95%。特别地,对于与其他细胞的混合物,所述百分比可为约10%-15%、15%-20%、20%-25%、25%-30%、30%-35%、35%-40%、40%-45%、45%-50%、60%-70%、70%-80%、80%-90%或90%-95%。或者,分离/纯度可以细胞倍增的形式表示,其中细胞已发生例如10-20、20-30、30-40、40-50或更多次的细胞倍增。
给定体积的细胞数量可通过广为人知的和常规的方法和仪器测定。给定体积的细胞混合物中的细胞的百分数可通过几乎一样的方法测定。可手动或通过使用自动细胞计数器容易地对细胞进行计数。给定体积的特定细胞可使用特异性染色和目测以及通过使用特异性结合试剂(一般为抗体)、荧光标记和荧光激活细胞分选器的自动化方法而测定。
倚赖多种因素选择用于为给定用途给予所述细胞的制剂。在这些因素中,重要的是受试者的物种,待治疗的病症、功能障碍或疾病的性质及其在受试者中的状态和分布,其他待给予的疗法和试剂的性质,给药的最佳途径,经过该途径的残存性,给药方案以及其他本领域技术人员明了的因素。具体地,例如,合适载体和其他添加剂的选择会倚赖给药的确切途径和特定剂型的性质。
例如,细胞存活可为基于细胞的疗法的效力的一个重要决定因素。这对于主要治疗和辅助治疗都是成立的。目标位点不适于细胞接种和细胞生长时会产生另一个问题。这可能会阻止治疗性细胞接近该位点和/或移植到那里。本发明的多个实施方案包括增加细胞存活和/或克服因接种和/或生长屏障而引起的问题的措施。
细胞/培养基的水性悬液的最终配制一般包括将所述悬液的离子强度调节到等渗(即约0.1-0.2)以及将ph调节到生理ph(即约ph6.8-7.5)。所述最终制剂通常还含有液体润滑剂例如麦芽糖,其必须为身体所耐受。示例性润滑剂组分包括甘油、糖原、麦芽糖等。基于有机聚合物的材料,例如聚乙二醇和透明质酸以及非纤维状胶原(优选琥珀酸化的胶原),也可用作润滑剂。这些润滑剂通常用于在注射位置上改善所注射生物材料的注射能力、润透性和分散性,以及通过改变所述组合物的粘性来降低堵塞(spiking)的量。最后的制剂必然是可药用载体中的细胞。
随后将所述细胞置于注射器或其他注射装置中以精确注射到组织缺损的位点上。术语“可注射”指所述制剂可在正常压力和正常条件下以带有低至25号针头的注射器中给予,而没有显著堵塞。堵塞可导致组合物从注射器溢出而非注射到组织中。对于这一精确布置,需要细至27号(200μi.d.)、甚或30号(150μi.d.)的针头。可通过这些针头挤出的最大颗粒大小将因至少以下参数复杂变化:颗粒最大尺寸、颗粒长宽比(长:宽)、颗粒刚性、颗粒表面粗糙度和影响颗粒之间粘性的有关因素、悬浮液的粘弹性,以及通过针头的流速。悬浮于牛顿流体中的刚性圆珠代表最简单的情况,而在粘弹性流体中的纤维性或有分支的颗粒可能更复杂。
本发明的所述组合物的理想等渗性可使用氯化钠或其他可药用剂例如右旋糖、硼酸、酒石酸钠、丙二醇、或其他无机或有机溶质实现。氯化钠特别优选用于含有钠离子的缓冲液。
如果需要,可使用可药用的增稠剂将所述组合物的粘性保持在选定水平上。优选甲基纤维素,因为它可方便和经济地获得并且易于使用。其他合适的增稠剂包括例如黄原胶、羧甲基纤维素、羟丙基纤维素,卡波姆等。所述增稠剂的优选浓度取决于所选择的试剂。重要的是使用实现选定粘度的量。粘性组合物通常是通过在溶液中加入所述增稠剂而制备的。
可药用的防腐剂或稳定剂可用于增加细胞/培养基组合物的寿命。如果加入所述防腐剂,那么选择不影响所述细胞活力或效力的组合物完全在本领域技术人员的能力范围之内。
本领域技术人员会认识到所述组合物的组分在化学上应为惰性的。这在化学和药学原理方面不对本领域技术人员构成问题。通过参考标准教科书,或者通过使用本公开内容、本文引用文献和本领域通常可获得的信息的简单实验(不涉及过多试验)可以容易地避免问题。
无菌可注射溶液可通过以下方式制备:将用于实施本发明的细胞/培养基与不同量的其他所需成分掺入所需量的合适溶剂中。
在某些实施方案中,细胞/培养基被制成单位剂量的可注射形式,例如溶液、悬液或乳液。适于细胞/培养基注射的药物制剂通常为无菌水性溶液和分散系。用于可注射制剂的载体可为含有如下物质的溶剂或分散介质:例如,水、盐水、磷酸缓冲盐水、多元醇(例如甘油、丙二醇、液体聚乙二醇等)以及它们的合适混合物。
本领域技术人员可容易地确定将在本发明方法中给予的组合物中的细胞和可选添加剂、媒介(vehicle)和/或载体的量。通常,任何添加剂(除所述细胞以外)都以0.001-50重量%的量存在于溶液例如磷酸缓冲盐水中。所述活性组分以微克至毫克量级存在,例如约0.0001到约5重量%,优选约0.0001到约1重量%,最优选约0.0001到约0.05重量%,或者约0.001到约20重量%,优选约0.01到约10重量%,最优选约0.05到约5重量%。
在某些实施方案中,将细胞封装(胶囊化)给药,特别是当胶囊化可提高治疗效力时或者提供处理和/或贮藏寿命上的优点时。在某些实施方案中,当胶囊化增加细胞介导的免疫抑制的效力时,它也因此降低对免疫抑制药物治疗的需要。
此外,在某些实施方案中,胶囊化提供可进一步降低受试者对所述细胞(通常在同种异体移植中具有免疫原性或者只有微弱免疫原性)的免疫反应的针对受试者免疫系统的屏障,从而减轻由于给予所述细胞而可能发生的任何移植排斥或炎症。
细胞可在植入前通过膜以及胶囊进行封装。应考虑到,可使用可用于细胞胶囊化的多种方法中的任意一种。在某些实施方案中,细胞被单独封装。在某些实施方案中,许多细胞被封装在同一膜内。在其中所述细胞在植入后被取出的实施方案中,将许多细胞封装在例如单一膜内的较大尺寸的结构可提供方便的取出方法。
在微囊化细胞的多个实施方案中可使用多种材料。这些材料包括例如聚合物胶囊,藻酸盐-聚-l-赖氨酸-藻酸盐微胶囊、聚-l-赖氨酸藻酸钡胶囊、藻酸钡胶囊、聚丙烯腈/聚氯乙烯(pan/pvc)中空纤维以及聚醚砜(pes)中空纤维。
可用于细胞给药的细胞微囊化技术为本领域技术人员已知,在例如chang,p.,etal.,1999;matthew,h.w.,etal.,1991;yanagi,k.,etal.,1989;caiz.h.,etal.,1988;chang,t.m.,1992以及美国专利no.5,639,275(其描述了例如用于长期维持稳定地表达生物活性分子的细胞的生物相容胶囊)中描述了这些技术。胶囊化的其他方法见欧洲专利公开no.301,777和美国专利no.4,353,888;4,744,933;4,749,620;4,814,274;5,084,350;5,089,272;5,578,442;5,639,275和5,676,943。所有上述文献与细胞胶囊化有关内容都以引用的方式纳入本文。
某些实施方案将细胞加入聚合物中,例如生物聚合物或合成聚合物。生物聚合物的实例包括但不限于纤连蛋白、纤维蛋白(fibin)、纤维蛋白原(fibrinogen)、凝血酶、胶原和蛋白聚糖。其他因子例如上面讨论的细胞因子也可被加入所述聚合物中。在本发明的其他实施方案中,细胞可被加入三维凝胶的间隙中。大的聚合物或凝胶通常通过手术植入。可配制成足够小的颗粒或纤维的聚合物或凝胶可通过其他常规的、更方便的、非手术的途径给予。
给药
组合物可根据诸如具体患者的年龄、性别、体重和病症,以及将给药的制剂(例如固体对液体)等因素,按剂量并通过医药和兽医领域技术人员公知的技术给药。人类或其他哺乳动物的剂量可由技术人员根据本公开内容、本文引用的文献以及本领域的知识,不需过度实验而确定。
适用于本发明多个实施方案的细胞/培养基的剂量取决于多种因素。其对于不同的情况可发生相当大的变动。确定主要和辅助治疗的最佳给药剂量的参数一般包括下面的一些或全部:要治疗的疾病及其阶段;受试者的物种、它们的健康、性别、年龄、体重和代谢速率;受试者的免疫活性;其他正在给予的疗法;以及根据受试者的历史或基因型预测的可能并发症。所述参数还可包括:所述细胞是同基因的、自体的、同种异体的还是异种的;它们的效能(具体活性);使所述细胞/培养基有效而必须靶向的位点和/或分布;以及所述位点的这些特性例如细胞/培养基的可达性和/或细胞的植入性。其他参数包括与其他因子(例如生长因子和细胞因子)的共给药。给定情况下的最佳剂量还要考虑将细胞/培养基配制的方式、给予它们的方式、给药后细胞/培养基定位到靶标位点的程度。最后,最佳剂量的确定必须提供这样的有效剂量,即所述剂量既不低于最大有益效果的阈值也不高于剂量相关的有害作用超过增加剂量的益处的阈值。
对于某些实施方案,细胞的最佳剂量处在用于自体、单核骨髓移植的剂量范围内。对于非常纯的细胞制剂,在不同实施方案中,每次给药的最佳剂量可为104-108细胞/kg接受者质量。在某些实施方案中,每次给药的最佳剂量为105-107细胞/kg。在许多实施方案中,每次给药的最佳剂量为5×105-5×106细胞/kg。用于参考,前述较高的剂量与用于自体单核骨髓移植的有核细胞的剂量相似。某些较低的剂量与用于自体单核骨髓移植的cd34+细胞/kg的数量相似。
应理解,单份剂量可一次全部、分份或在一段时间内连续递送。也可将整份剂量递送到单个位置或分份分散到若干个位置。
在各个实施方案中,细胞/培养基可以以初始剂量给药,然后通过后续给药保持。细胞/培养基可在开始通过一种方法给药,然后通过相同方法或者一种或多种不同方法给药。水平可通过继续给予所述细胞/培养基而保持。各个实施方案通过静脉内注射在开始时给予所述细胞/培养基和/或保持它们在受试者中的水平。在多个实施方案中,可根据患者的病情和本文其他地方讨论的其他因素使用其他给药形式。
应注意,人类受试者接受治疗的时间通常长于实验动物,然而,治疗的时间通常与疾病进展的时间和治疗效果成比例。本领域技术人员在使用在人和/或动物(诸如大鼠、小鼠、非人灵长类等)中实行的其他方法的结果时会考虑这一因素,以确定用于人的合适剂量。基于这些考虑并根据本公开内容和现有技术所提供的指导,这种确定可使技术人员不需过度实验而确定剂量。
初始给药和后续给药或连续给药的合适方案可以都相同或者可以是变化的。合适方案可由技术人员根据本公开内容、本文引用的文献以及本领域的知识确定。
治疗的剂量、频率和持续时间取决于许多因素,包括疾病的性质、受试者以及可能给予的其他疗法。因此,多种方案可用于给予所述细胞/培养基。
在某些实施方案中,将细胞/培养基以一个剂量给予受试者。在其他一些实施方案中,将细胞/培养基以一系列的两个剂量或多个剂量连续给予受试者。在其中以单个剂量、两个剂量和/或多于两个的剂量给予细胞/培养基的其他一些实施方案中,剂量可相同或不同,并且给药之间的间隔可相同或不同。
细胞/培养基可在各种不同时间内以多个频率给药。在某些实施方案中,细胞在小于1天的时间内给药。在其他实施方案中,它们在2、3、4、5、或6天的时间内给药。在某些实施方案中,细胞在数周的时间内以每周一次或多次给药。在其他实施方案中,细胞在数周的时间内给药持续一到数月。在多个实施方案中,细胞可在数月的时间内给药。在其他实施方案中,细胞可在一或数年的时间内给药。通常治疗时间是与疾病进展的时间、所用疗法的效果以及要治疗的受试者的情况和反应成比例的。
例如,本文所解释的脊髓损伤可分两个阶段。在动物模型中,在第一阶段,巨噬细胞不渗入所述损伤,此阶段持续约24小时。然而,在第二阶段,巨噬细胞渗入所述损伤,当评定治疗方案时可考虑此事件后果。在一个实施方案中,如果预期会与营养不良的轴突相互作用的巨噬细胞或其他细胞会渗入,则甚至在第一阶段过程中即给予细胞,例如在刚损伤后或与损伤尽可能接近。然后,可继续治疗以与巨噬细胞的初始和后续渗入同步,并且可预防性地继续治疗,或者在确定巨噬细胞或其他相关细胞不再渗入所述损伤时可能中断治疗。
实施例
实施例i
"glialscarmodel"aggrecan-lamininopposingspotgradients(tometal..2004;steinmetzetal..2005)。这些参考文献以引用的方式纳入以教导神经胶质疤痕模型。此模型提供了细胞、蛋白、培养基等对在体外降低粘附/回缩的有效性的测定法。
如果抑制性基质存在于比体内损伤后形成的空间组织更相似的空间组织中,那么pg可诱导轴突中所谓的营养不良状态。为此,pg聚集蛋白聚糖和生长促进分子层粘连蛋白的溶液的斑点被置于硝酸纤维素盖玻片上并空气干燥。
干燥的连续人工产物形成了粗略的梯度,其中所述斑点的边缘与中心相比含有越来越高的聚集蛋白聚糖浓度。所述边缘的最外部与任何更接近中心的区域相比含有更低的层粘连蛋白的浓度。最佳ecm浓度(0.7mg/ml聚集蛋白聚糖和5μg/ml层粘连蛋白)导致了良好的细胞贴附。因此,高聚集蛋白聚糖-低层粘连蛋白的外边缘似乎是对再生神经突特别不利的区域。没有神经突通过穿过斑点清晰的外边界从环绕的层粘连蛋白进入所述斑点。从所述斑点的中心内向心生长的纤维能够进入所述边缘的内部,但不能再向前生长。一旦进入所述梯度,轴突就显得被困住。在所述梯度内,在神经突末端处形成了球棒状的、营养不良的端球。为了观察所述“营养不良的末端”的表现,使用了缩时显微术(time-lapsemicroscopy)。营养不良的生长锥经常努力前进较短的距离,但不可避免的,所述艰难行进的生长锥会聚拢成更致密的球并回缩,只是为了再次开始移动。
实施例ii
轴突回缩和巨噬细胞
总述
在体内,在背柱破碎脊髓损伤后,在位于切断的轴突末端的营养不良的回缩球棒状物与ed-1+细胞之间发现了紧密关联(图3)。使用了神经胶质疤痕的体外模型(tometal.,2004;steinmetzetal.,2005)来实时检查轴突与ed-1+细胞之间的相互作用。营养不良的生长锥与ed-1+巨噬细胞之间直接的细胞-细胞接触诱导了长距离轴突回缩(图4、5)。使用氯膦酸盐脂质体(popovichetal.,1999)在体内清除巨噬细胞的结果是经氯膦酸盐处理的动物与对照相比轴突回缩的明显减少。这些数据表明ed-1+细胞通过物理的细胞-细胞相互作用直接引起受损伤脊髓轴突的回缩。
结果
1.脊髓损伤后上行脊柱感觉轴突大幅回缩
发明人认为活化的巨噬细胞的渗入能够在轴突回缩中起直接作用。在亚急性和慢性脊髓损伤中,在活化的巨噬细胞和再生轴突的末端之间发现了紧密关联,使得这两种细胞类型之间可发生直接物理相互作用。然后表征背柱破碎损伤后感觉轴突回缩的程度并且将其与巨噬细胞渗入到损伤中相关。将成体雌性sprague-dawley大鼠的脊柱在节段c8打碎,并通过dextran-texasred标记坐骨神经追踪受损伤神经元子群。在损伤后2、4、7、14和28天收集脊髓组织并测量标记纤维末端与损伤中心之间的距离。到损伤后2天,轴突已经回缩343±46.92um的平均距离(平均值±标准偏差),然而,此早期回缩最可能是因为神经元自身内的固有机制(kerschensteineretal.,2005)。需要强调的是,在损伤后2天,损伤主要由有活性的星形细胞(gfap+)和少数ed-1+细胞构成,所述ed-1+细胞最可能是原有的小神经胶质。在损伤后2天和7天之间损伤内的ed-1+细胞的数量急剧增加,其大部分是最可能渗入的巨噬细胞(popovichetal.,1997;donnellyandpopovich,2007)。脊柱内上行感觉纤维的第二阶段回缩在第一周最快地出现,然后在随后的几周逐步地出现。到损伤后28天,轴突已从损伤中心回缩774±70.26um的平均距离。这些数据表明上行感觉轴突回缩的时间与损伤内ed-1+细胞的渗入和集聚在空间和时间上一致。
2.活化的巨噬细胞的清除减少体内轴突回缩
体内轴突回缩的第二阶段主要在损伤后2天和7天之间——与巨噬细胞渗入的时间一致——发生在上行脊柱感觉轴突中。为进一步弄清巨噬细胞在体内轴突回缩中的作用,用氯膦酸盐脂质体处理动物以清除循环的单核细胞/巨噬细胞(vanrooijenetal.,1997;popovichetal.,1999)。动物每隔一天接受氯膦酸盐脂质体注射,在受伤前一天开始处理,以清除循环的单核细胞/巨噬细胞。然后在损伤后2天、4天和7天对动物评定轴突回缩(图3)。与接受对照脂质体的动物在损伤后4天和7天的回缩(分别为586±42.89um和806±62.71um)比较,接受氯膦酸盐脂质体注射的动物在损伤后4天和7天表现出回缩的明显减少(分别为402±81.85um和439±46.33um)。回缩减少与经氯膦酸盐处理的动物与接受空脂质体对照的动物相比损伤内ed-1+细胞数量的显著减少相关。氯膦酸盐脂质体处理还导致损伤中心的gfap+星形细胞突起(process)的增加,与之前观察到的巨噬细胞清除导致空穴形成减少的现象一致(popovichetal.,1999)。重要的是,在损伤后2天,经氯膦酸盐处理和经对照脂质体处理的动物中显现的回缩量没有差别。此时尚未出现巨噬细胞渗入,表明轴突回缩的第一阶段是不巨噬细胞依赖的,可能是由于内生神经元机制或者可能是由于与活化的原有小神经胶质的相互作用。氯膦酸盐介导的循环巨噬细胞/单核细胞的清除阻止了通常在损伤后4天和7天出现的轴突回缩,表明回缩的第二阶段是由渗入巨噬细胞的作用导致的。在经氯膦酸盐处理的动物中未出现明显的再生(即轴突延长至损伤中心之外)。
3.在神经胶质疤痕的体外模型中营养不良的生长锥在与巨噬细胞接触后大幅回缩
在体内观察到ed-1+细胞与受损伤的轴突的紧密关联表明这些细胞之间的相互作用可能在轴突回缩中起作用。研究了神经胶质疤痕的体外模型中成体感觉神经元轴突与巨噬细胞之间的相互作用。在30分钟的基线观察期后,将nr8383巨噬细胞加入培养物中并监测它们与营养不良的轴突的相互作用。在营养不良的轴突与巨噬细胞之间经常观察到直接的细胞-细胞接触。这些接触持续较长一段时间,并且当与巨噬细胞的迁移结合时,引起对轴突的强烈操控,导致轴突有力地弯曲并从基底抬起。很明显,强的、长时间的附着可在所述两种细胞类型之间发生,因为在巨噬细胞移开轴突之后经常仍然存在连接所述两种细胞的长拖尾突起。然而,巨噬细胞诱导的回缩不永久阻止轴突的再生长,因为某些在回缩后不再与巨噬细胞接触的轴突能够延长直至这些轴突再度营养不良。这两种细胞类型之间直接的细胞-细胞接触最终导致了轴突在全部接触时间的大幅回缩。因此,在神经胶质疤痕的体外模型中巨噬细胞接触诱导了营养不良的轴突的回缩。
4.直接的物理的细胞-细胞相互作用对于巨噬细胞诱导的回缩是必需的
有多次观察到巨噬细胞移动至非常接近但未接触营养不良的轴突处的情况,在这些情况下未观察到轴突回缩。为了确定轴突与巨噬细胞之间的物理相互作用是否是诱导轴突回缩所必需的或者巨噬细胞产生的因子是否足够,用胰蛋白酶处理巨噬细胞以除去胞外蛋白,然后将其加至drg培养物。用胰蛋白酶对巨噬细胞的预处理不妨碍巨噬细胞的大幅迁移和与轴突的多次碰撞。然而,所述处理完全阻止了巨噬细胞物理附着于轴突,因此在不存在长时间的直接细胞-细胞接触的情况下未观察到回缩。然而,巨噬细胞分泌诱导回缩的因子是可能的。为检测这一假设,在体外将巨噬细胞条件培养基加至营养不良的轴突。巨噬细胞条件培养基不诱导回缩。因此,在不存在与营养不良的轴突的物理相互作用的情况下,在轴突附近仅存在巨噬细胞或其分泌的因子均不能诱导轴突回缩。
5.巨噬细胞诱导的回缩中基底的作用
为测定基底在轴突回缩中是否起作用,在均匀生长促进层粘连蛋白基底上培养成体感觉神经元,所述基底不产生营养不良的生长锥。借助多个丝状伪足(filopodia)和板层形伪足(lamellapodia)使层粘连蛋白上的生长锥变平,并且全部轴突延伸均以恒定速度发生。当将巨噬细胞加至这些培养物时,观察到了与轴突的直接细胞-细胞接触。然而,这些接触是短暂的并很快被破坏,幅度不那么大,并且未导致轴突回缩。膜接触点的残留部分被快速重吸收回神经元中,并且生长锥继续延伸穿过无阻碍的基底。因此,巨噬细胞诱导的轴突回缩是基底依赖的;在允许的基底层粘连蛋白上活动生长状态的神经元对巨噬细胞接触不敏感,与cspg梯度诱导的营养不良状态下的神经元不同。
6.活化的原代巨噬细胞也诱导轴突回缩
另一问题是,原代巨噬细胞在体外是否以与nr8383巨噬细胞系同样的方式与营养不良的轴突相互作用。收集成年sprague-dawley大鼠骨髓中的祖细胞并使其在体外分化成巨噬细胞,得到多于80%的ed-1+细胞的培养物。此具体巨噬细胞群体已显示出保留有在脊髓损伤中出现的巨噬细胞的表型、形态和功能特征,而不同于从其他身体来源收集的群体(longbrakeetal.,2007)。然后测定了原代巨噬细胞诱导轴突回缩的能力。未刺激的原代巨噬细胞不能诱导回缩。当加至斑点梯度神经元培养物时,这些巨噬细胞附着于基底但不能动,呈现休眠状态的巨噬细胞特征。仅当将巨噬细胞直接置于营养不良的轴突上时才出现与轴突的接触。这些巨噬细胞及其细胞-细胞相互作用均未表现出之前用细胞系巨噬细胞观察到的任何物理特征,即无牵拉、无经细胞突出的物理附着的迹象等。
巨噬细胞可能必须处于活动状态以与营养不良的轴突相互作用。在培养中用活化细胞因子干扰素γ刺激原代巨噬细胞,然而加至缩时培养皿中。尽管这些巨噬细胞呈现中度活化状态和偏圆的形态,然而它们仍然不能动并且不与营养不良的轴突形成强附着,因此不导致轴突回缩。再用干扰素γ和脂多糖(lps)的结合物刺激原代巨噬细胞,然后加至drg培养物中。这些巨噬细胞表现出活化的巨噬细胞的形态和表现:圆的、噬菌细胞的形状和高能动性。这些活化的巨噬细胞以与细胞系巨噬细胞同样的频率诱导营养不良的轴突回缩。它们显示出与营养不良的轴突的有力物理相互作用,导致细胞之间的强附着以及轴突的物理抓附、牵拉和从基底的抬起。当处于活化状态时,原代巨噬细胞在体外诱导了营养不良的轴突的回缩,认可了在此体外轴突回缩的研究中使用细胞系巨噬细胞。因此,实验主要用nr8383巨噬细胞系进行,因为该细胞系形成了处于恒定活化状态的纯细胞群体,其类似于在脊髓损伤内发现的没有额外刺激的巨噬细胞。
7.活化的小神经胶质在体外能够中度诱导轴突回缩
尽管巨噬细胞通常在损伤后3天之前不会侵入受损伤的脊髓,然而cns内原有的小神经胶质会立即对损伤作出反应(watanabeetal.,1999)。损伤内的小神经胶呈现活化和噬菌细胞的状态,很像活化的巨噬细胞。在损伤后2天,在典型的巨噬细胞渗入之前,在损伤内发现有限数量的ed-1+细胞,这些细胞最可能是原有的小神经胶质。使用体外模型评定了小神经胶质对轴突回缩的可能作用。从p1sprague-dawley大鼠收集了脑皮质小神经胶质并使其在体外成熟,然后将其加入缩时培养物中。类似于原代巨噬细胞,在培养中必须用干扰素γ和lpc刺激原代小神经胶质以使其被活化。未被刺激的小神经胶质不能附着于层粘连蛋白/集聚蛋白聚糖斑点梯度基底,这阻止所述细胞在本发明人的模型中与营养不良的轴突相互作用。然而,被刺激的小神经胶质附着并与轴突物理相互作用,在50%的接触时间内诱导回缩,然而,活化的小神经胶质与营养不良的轴突之间的接触不如巨噬细胞的强。因此,实验活化的小神经胶质也可在轴突回缩的诱导中起作用。
8.另一种cns细胞类型——星形细胞——不能在体外诱导轴突回缩
另一个问题是,在体外诱导营养不良的轴突的回缩是否是对在损伤的脊髓内常见的噬菌细胞类型特异的,而不仅是营养不良的轴突与任何其他细胞类型的相互作用的结果。星形细胞是cns的损伤后的神经胶质疤痕的一个不可或缺的组分。它们大量存在并广泛接触再生的轴突。使脑皮质星形细胞在体外成熟,然后加至drg培养物中。星形细胞附着于基底并广泛接触营养不良的轴突。一旦与基底结合,星形细胞就沿集聚蛋白聚糖梯度迅速迁移,远离边缘。星形细胞突起分布在轴突上,有时导致轴突的侧向位移。然而,这些接触不导致被接触的轴突回缩。因此,回缩的诱导对与ed-1+噬菌细胞的相互作用是特异的,而不仅是与任何其他细胞类型的物理相互作用。
材料和方法
1.背柱破碎损伤模型
使用33只成年雌性sprague-dawley大鼠(250-300g)进行体内研究。对于所有手术步骤,均用吸入的异氟醚气(2%)麻醉大鼠。进行t1椎板切除术来暴露c8脊髓节的背面。用30号针头在距离中线0.75mm的两侧进行硬脑膜切开术。然后通过将dumont#3jeweler镊子在c8处插入背脊至1.0mm的深度并压住镊子,保持压力10秒钟并再重复2次来制造背柱破碎损伤。通过观察到白色物质的清除来确认损伤的完成。然后用凝胶膜覆盖硬脑膜上的洞。用4-0尼龙线缝合肌肉层,然后用外科封缝钉闭合皮肤。在闭合切口后,动物沿切口皮下接受麻卡因(marcaine,1.0mg/kg)并肌内注射丁丙诺啡(0.1mg/kg)。手术后,在麻醉恢复期间用加热灯使动物保暖,并使其自由取食和饮水。将动物在损伤后2、4、7、14或28天处死(每组n=3)。所有动物操作均依照casewesternreserveuniversity的animalresourcecenter的指南和方案进行。
2.巨噬细胞清除
动物在背柱破碎损伤前的一天接受脂质体封装的氯膦酸盐或空脂质体对照的腹膜内注射,并且还在损伤后一天(对于2d时间点)、损伤后1天和3天(对于4d时间点)和损伤后1、3和5天(对于7d时间点)接受上述腹膜内注射(每组n=3)。氯膦酸盐由rochediagnosticsgmbh,mannheim,germany赠予。氯膦酸盐如现有技术所述封装在脂质体中(vanrooijenandsanders,1994)。
3.轴突标记
处死前2天,用texas-red缀合的3000mw葡聚糖单侧标记脊柱。简言之,暴露右后肢的坐骨神经,用dumont#3镊子挤压10秒钟并再重复两次。在挤压位点处用hamilton注射器将1.0ul3000mw葡聚糖-texas-red的10%无菌水溶液注射到坐骨神经中。用4-0尼龙线缝合肌肉层,然后用外科封缝钉闭合皮肤。在闭合切口后,动物沿切口皮下接受麻卡因(1.0mg/kg)并肌内注射丁丙诺啡(0.1mg/kg)。手术后,在麻醉恢复期间用加热灯使动物保暖,并使其自由取食和饮水。将动物在标记后2天用过量异氟醚处死并用pbs继而用4%pfa灌注。收集组织并在4%pfa中后固定并进行免疫组织化学测定。
4.免疫组织化学测定
将组织在4%pfa后固定过夜,然后浸没在30%蔗糖中过夜,在oct封装介质中冷冻,然后在恒冷切片机上切成20um的纵向切片。然后将组织分别用抗gfap(accuratechemicalandscientificcorporation,westbury,ny)、抗ed-1(millipore,billerica,ma)染色,并分别用alexafluor-405或alexafluor-488(invitrogen,carlsbad,ca)孵育,然后在zeissaxiovert510激光扫描共焦显微镜上成像。
5.体内轴突回缩定量
对每只动物分析了始于每只动物的脊髓背面下200um深处的三个连续切片以定量轴突回缩。通过特征性gfap和ed-1染色模式和标记的轴突末端之间的距离确定损伤中心,并使用zeisslsm5imagebrowser软件确定中心位置。将一组中所有动物的所有切片的测量值取平均以得出每个时间点的回缩平均距离。
用于追踪受损伤纤维的技术标记位于脊柱内非常浅处的轴突。同时,标记的纤维的数量可由于注射示踪剂的节段的坐骨神经的肌纤维自发性收缩的程度而变化。出于多种原因,仅定量在该深度的标记的轴突。此深度在所有动物中均含有标记的纤维,而某些动物在更深的深度不具有标记的纤维。损伤的线性程度在所述脊柱的更深节段增加。因此位于脊髓更深处的轴突受到了比在更浅节段的损伤大很多的损伤。回缩距离的定量必须发生损伤的类似位置以允许动物和组之间的准确比较。对标记轴突的整个群体的定量可因标记程度的不同而导致结果的偏差。相反,对标记轴突的具体群体和位置进行定量,从而能够在所有动物中对它们进行一致的检查和准确的定量。
6.drg解离
如现有技术所述收集drg(tometal.,2004;daviesetal.,1999)。简言之,从成体雌性sprague-dawley大鼠切下drg(zivicmiller,harlan)。除去中心根和外周根,并在胶原酶ii(200u/ml,worthington)和dispaseii(2.5u/ml,roche)的hbss溶液中孵育神经节。将消化的drg冲洗并在新鲜hbss-cmf中轻轻研磨三次,然后低速离心。然后将解离的drg重悬浮在补充有b-27、glutamax、青霉素/链霉素(均来自invitrogen)的neurobasal-a培养基中并计数。将drg以3000个细胞/ml的密度置于delta-t培养皿上(fisher,pittsburgh,pa),共6000个细胞/培养皿。
7.缩时培养皿制备
与tometal.,2004所述类似地制备delta-t细胞培养皿(fisher,pittsburgh,pa)。简言之,用2号钻头通过每个培养皿的上半部钻一个洞,以形成在缩时显微术过程中向培养物中加细胞、酶、抑制剂等的进口。然后,将培养皿用无菌水冲洗并在室温下以聚-l-赖氨酸(0.1mg/ml,sigma)包被过夜,然后用无菌水冲洗并使其干燥。通过吸取2.0ul的聚集蛋白聚糖溶液(2.0mg/ml,sigma,溶于hbss-cmf,invitrogen)于培养物表面上并使其干燥,形成聚集蛋白聚糖梯度斑点。每个培养皿中放置6个斑点。在聚集蛋白聚糖斑点完全干燥后,将培养皿的整个表面在层粘连蛋白的hbss-cmf溶液(10ug/ml,bti,stoughton,ma)中于37℃温浴3小时。然后撤除层粘连蛋白浴,立即将细胞铺板。只用层粘连蛋白浴不用聚集蛋白聚糖如上所述制备了只含有层粘连蛋白基底的培养皿。此处所用的基底的浓度不同于tometal.,2004中所用的浓度。显微术的清晰度可通过去除硝酸纤维素而从培养皿制备方案改善。然而,为补偿结合于培养皿表面的基底的不同,将所用的基底浓度重新校准为上面所列的浓度。
缩时成像后,将drg在4%pfa中固定并用抗iii型b-微管蛋白(1:500;sigma,st.louis,mo)和抗硫酸软骨素(cs-56,1:500,sigma)免疫染色。
8.细胞系巨噬细胞培养物
如yinetal.(2003)所述培养成体sprague-dawley肺泡巨噬细胞系nr8383细胞(atcc#crl-2192)。简言之,在未包被的组织培养烧瓶(corning)中,将细胞在补充有15%fbs、glutamax、penn/strep(invitrogen)和碳酸氢钠(sigma)的f-12k培养基(invitrogen)中培养,并每周加料2-3次。此细胞系形成贴壁和悬浮细胞的混合培养物,并在加料时通过收集和重铺板悬浮细胞进行传代。为制备用于缩时显微术实验的细胞系巨噬细胞,将细胞用0.5%胰蛋白酶/edta(sigma)收集并用无血清f-12k洗涤3次,然后以1.0×106/ml的密度铺板于含无血清f-12k的未包被组织培养烧瓶中。在用于缩时实验前的1天,用edta和细胞刮刀收集培养的细胞系巨噬细胞并以2.5×105/70ul的密度重悬浮于添加有hepes(50um,sigma)的如上述补充neurobasal-a中。
9.原代骨髓源巨噬细胞培养物
基于现有技术方案(tobianetal.,2004)收集骨髓祖细胞。简言之,将股骨从成年雌性sprague-dawley大鼠(225-275g,harlan)取下。除去股骨末端,将含有补充有10%fbs、glutamax、penn/strep、β-巯基乙醇和hepes(invitrogen)(d10f)的冷dmem的注射器插入股骨,冲出骨髓并收集。然后使所得细胞混合物通过70微米的滤器并离心。然后除去上清液,将所得细胞片状沉淀物重悬浮于akt裂解缓冲液(biowhitacre)中以裂解红细胞,然后离心。除去上清液并将含有骨髓前体细胞的片状沉淀物重悬浮并铺板于另补充有20%ladmac细胞系条件培养基(cliffordharding赠予)的上述dmem中以诱导分为成巨噬细胞。在培养第5、7、9天对细胞加料,在第10天收集细胞用于实验。缩时实验前的1天,用胰蛋白酶/edta收集原代巨噬细胞,用d10f洗涤3次并以1.0×106/ml的密度铺板于含d10f的未包被陪替氏培养皿(falcon)中。第2天,用edta和细胞刮刀收集原代巨噬细胞并以5.0×105/70ul的密度重悬浮于加hepes的neurobasal-a中,用于缩时显微实验。
10.脑皮质星形细胞制备
通过取出p0-p1大鼠的脑皮质,切碎并用0.5%胰蛋白酶的edta溶液处理来收集脑皮质星形细胞。将细胞接种于以多聚l-赖氨酸包被的t75烧瓶上的含有10%fbs(sigma)和2mmglutamax的dmem/f12(invitrogen)中,并摇动,4小时后除去未贴壁的细胞。星形细胞在培养中成熟至少28天。用edta和细胞刮刀收集星形细胞并以5.0×105/70ul的密度重悬浮于加hepes的neurobasal-a中,用于缩时显微实验。
11.皮质小神经胶质制备
通过取出p0-p1大鼠的脑皮质,切碎并用0.5%胰蛋白酶的edta溶液处理来收集脑皮质小神经胶质。将细胞铺板于以多聚l-赖氨酸包被的t75烧瓶上的含有20%fbs(sigma)和2mmglutamax的dmem/f12(invitrogen)中,维持5-7天。缩时实验前的1天,摇动烧瓶以除去贴壁不牢的细胞并将这些细胞以1.0×106/ml的密度铺板于含d10f的未包被陪替氏培养皿(falcon)中。第2天,用edta和细胞刮刀收集原代小神经胶质并以5.0×105/70ul的密度重悬浮于加hepes的neurobasal-a中,用于缩时显微实验。
12.缩时显微术研究
将drg神经元于37℃孵育48小时,然后进行缩时成像。将含有hepes(50um,sigma)的neurobasal-a培养基加至培养物中,然后转至加热的台架仪器中。使用zeissaxiovert405m显微镜和100×油浸物镜,每30秒取得缩时成像,持续3小时。选择径直延伸到斑点边缘并具有特征性营养不良形态的生长锥。观察神经元30分钟以确定基线表现,然后加入其他细胞类型(原代巨噬细胞n=3,其他组n=6)。加入细胞后,观察生长锥150分钟。本发明人用metamorph软件追踪了延伸/回缩和生长速率。
13.统计学分析
用minitab15软件通过单因素或双因素anova或普通线性模型(如果适用)以及tukey事后检验分析数据。
讨论
这些结果首次表明了巨噬细胞可通过直接的物理接触诱导营养不良的成体轴突回缩的确凿证据。回缩的诱导同时倚赖神经元的生长状态和巨噬细胞的活化状态。在体外,原代骨髓源巨噬细胞必须用干扰素γ和lps刺激以达到与细胞系巨噬细胞和脊髓损伤内的巨噬细胞诱导轴突回缩类似的活化状态。这说明巨噬细胞的体内表现是状态依赖的并与之前表明巨噬细胞渗入仅在髓磷脂变性情况下与轴突回缩相关的研究(mcphailetal.,2004)是一致的。本研究表明成体感觉神经元仅在处于由抑制性cspg梯度诱导的停止生长的营养不良状态时才对巨噬细胞诱导的回缩敏感。在均匀层粘连蛋白基底上处于生长活化状态的成体神经元迅速中断与巨噬细胞的接触并且不发生回缩。
回缩的诱导可能包括多种内部和外部机制。本研究表明如果巨噬细胞与营养不良的轴突之间无直接的细胞-细胞接触则不发生回缩。仅加入经胰蛋白酶处理的巨噬细胞或巨噬细胞条件培养基不足以诱导回缩。巨噬细胞无需特别接触在生长锥处的营养不良的轴突来诱导回缩。与活化的巨噬细胞接触可能触发轴突内远离其营养不良末端的信号传递通路。
存在若干候选结合配偶体,通过所述配偶体巨噬细胞和神经元可物理地相互识别并相互作用。巨噬细胞可使用αv和β1整合素受体来识别和结合轴突玻璃粘连蛋白(sobeletal.,1995),并且巨噬细胞与变性周围神经的附着通过阻断β整合素被部分地减弱(brownetal.,1997)。视神经损伤后,轴突表达ephrinb3,其被巨噬细胞上的ephb3受体识别(liuetal.,2006)。唾液酸粘附素——一种巨噬细胞特异的唾液酸受体——存在于神经元细胞膜上(kelmetal.,1994;tangetal.,1997)。巨噬细胞还识别暴露在发生细胞凋亡的细胞外膜表面上的磷脂酰丝氨酸(deetal.,2002;desimoneetal.,2004),所述磷脂酰丝氨酸可标示受损伤的神经元以进行内吞作用。此外,fractalkine是主要在cns神经元的表面上表达的趋化因子,而其受体cx3cr1存在于巨噬细胞上(zujovicetal.,2000;umeharaetal.,2001)。必须进行进一步的研究来确定,这些分子中的哪些(如果有的话)在营养不良的成体神经元表面上表达或上调,确定它们为巨噬细胞识别的靶标。
本研究证实了上行脊柱通路发生回缩(borgensetal.,1986;mcphailetal.,2004;stirlingetal.,2004;bakerandhagg,2005;bakeretal.,2007)并且时间进程与巨噬细胞的渗入一致。轴突回缩已在脊髓内的其他通路包括下行皮质脊髓通路(fishmanandkelley,1984;iizukaetal.,1987;hilletal.,2001;seifetal.,2007)、延髓通路(houleandjin,2001)和红核脊髓通路(schwartzetal.,2005;caoetal.,2007)中被检测。重要的是认为轴突回缩分为两个不同的阶段。对成年小鼠神经轴索断裂的上行感觉轴突进行体内成像的最近研究表明,损伤前几个小时内轴突回缩约300um,然后轴突在损伤后前3天中稳定(kerschensteineretal.,2005)。因此,本研究的焦点是轴突回缩的第二、后一阶段,这是因为活化的巨噬细胞而不是因为所述神经元的固有性质。
本研究表明,在损伤之前以及整个正常回缩期间用氯膦酸盐脂质体处理阻止了损伤后2天至7天之间的轴突回缩的第二阶段。以前的工作已经表明,氯膦酸盐脂质体介导的巨噬细胞清除导致损伤体积减小和神经元存活增加(popovichetal.,1999;vanrooijenandvankesteren-hendrikx,2002)。尽管这些作者强调巨噬细胞清除对轴突再生的作用,然而本研究的数据表明经氯膦酸盐脂质体处理的动物的损伤的轴突含量增加可能(至少部分)是由于轴突回缩变弱所致。
实施例iii
干细胞可阻止活化的巨噬细胞与drg的粘附
结果
1.模型
背柱破碎损伤后,再生的轴突会遭遇巨噬细胞和小神经胶质并形成营养不良的末端。这一点示意性地示于图1。发明人实验室以前的工作已经表明,巨噬细胞的渗入与背柱破碎损伤后的轴突顶梢枯死相关(图2和3)。在表征损伤后上行脊柱感觉轴突的轴突顶梢枯死的程度后,本发明人建立了顶梢枯死的体外模型,其可用于评估多种治疗方法。所述体外测定法包括在如下基底上培养的成体drg神经元,所述基底具有生长促进蛋白层粘连蛋白和强抑制性硫酸软骨素蛋白聚糖聚集蛋白聚糖的相反梯度(tometal.,2004)。此斑点梯度足以使轴突生长停止并诱导与在受损伤的脊髓中出现的生长锥类似的营养不良的生长锥的形成。
缩时显微术使得发明人能够密切地检查生长锥动力学,例如丝状伪足的数目、板层形伪足的程度和营养不良末端中囊泡的数目。在营养不良的轴突与巨噬细胞之间经常出现导致轴突大幅回缩的直接的细胞-细胞接触(图4和5)。直接的细胞接触是诱导回缩所需的,因为巨噬细胞条件培养基和营养不良轴突附近巨噬细胞的存在均未导致回缩。因此,发明人假设活化的巨噬细胞的清除或调制可成为脊髓损伤中可能的治疗靶标。
发明人已经阐明了巨噬细胞-神经元相互作用导致顶梢枯死的机制。已知巨噬细胞分泌多种蛋白酶,这些蛋白酶辅助碎片的分解和清除,并且发明人已证明巨噬细胞会表达和分泌mmp-9。发明人假设某种蛋白酶能够负责将营养不良的轴突局部地移离基底导致其回缩。巨噬细胞表达的一类蛋白酶是基质金属蛋白酶(mmp)。已知mmp是导致cns再生障碍的原因,因为缺少某些mmp的转基因小鼠在损伤后表现出强的轴突再生长,如用通用mmp抑制剂gm6001处理的动物的表现一样。在加入巨噬细胞时,将gm6001——其在mmp活性位点作为锌螯合剂——施加于缩时培养皿中。在体外模型中用gm6001或特异性mmp-9抑制剂处理(图6)阻止了与巨噬细胞直接的细胞-细胞接触后营养不良的生长锥的回缩,而特异性mmp-2抑制剂没有阻止。gm6001或所述特异性mmp-9抑制剂不阻止巨噬细胞与营养不良的轴突之间直接的细胞-细胞接触。因此,证明mmp和mmp-9在轴突顶梢枯死中起作用。
2.mapc在体外阻止巨噬细胞介导的轴突顶梢枯死
图7示出了为确定mapc是否能调制巨噬细胞的抑制性作用的实验设计的示意图。将mapc加至1divdrg斑点培养物并再孵育1天。这些共培养神经元的生长锥的形态迥异于通常出现于斑点上的营养不良的生长锥。这些生长锥的能动性增加、变平并具有大量的板层形伪足。巨噬细胞接触所述生长锥和轴突,但这些接触经常是短暂的,并且成像的6个轴突中有5个不发生特征性巨噬细胞介导的回缩(图8)。
此结果可能是由于mapc的神经刺激作用或免疫调制作用,或是由于两种作用的结合。为解决此问题,进行了一系列条件培养基实验。用mapc条件培养基将解离的drg神经元处理24小时并与培养基对照进行比较以评定最长的神经突。在组之间未观察到神经突长度的显著差异,表明此作用可能不完全是神经刺激作用。
直接向所述缩时培养皿中加入mapc条件培养基导致生长锥形态的改变,从营养不良的、停滞的状态变为能动的、扁平的状态。巨噬细胞仍然接触这些轴突,但接触通常是瞬时的并且通常不导致轴突回缩。以mapc条件培养基预处理的巨噬细胞也在所述斑点上接触轴突,但不引起回缩(图9-12)。mapc可能作用于巨噬细胞从而改变它们的受体表达、对损伤细胞的响应或者mmp-9的分泌。
3.mapc降低背柱破碎损伤后轴突顶梢枯死的程度
使用脊髓损伤的背柱破碎模型研究了体内mapc对轴突顶梢枯死的免疫调制作用。轴突顶梢枯死的最剧烈阶段发生在损伤后2天至4天之间,这在时空上与活化的巨噬细胞渗入损伤一致。mapc可能会以降低轴突顶梢枯死量的方式调制损伤内活化的巨噬细胞。因此,在损伤后立即将mapc移植到脊髓内并在损伤后2天和4天测量轴突顶梢枯死的程度。将mapc移植到位于损伤尾侧大约500微米和中线侧面500微米的位置。选择此位置是为了将mapc置于接近受损伤轴突的末端,以使上行通道的进一步破坏最小化,并且以防止细胞在损伤位置被血液和csf流直接从脊髓中替换。
所移植的mapc成功地整合到脊髓组织中,这一点被在损伤后2天和4天在注射位点处均存在gfp+细胞所证明。此外,mapc从移植位点大幅移出并且能够占据损伤中心并且还被观察到与受损伤轴突的末端相关连。在损伤后2天,经mapc移植的动物中轴突顶梢枯死程度与对照动物的轴突顶梢枯死程度无显著差别(图13)。mapc移植不阻止在损伤后2天正常出现的轴突顶梢枯死程度。然而,此顶梢枯死的初始阶段最可能是由于固有神经元机制,而不受活化的巨噬细胞的介导,因为此时巨噬细胞尚未渗入损伤。
在损伤后4天,与未注射的对照相比,经mapc移植的动物显示出轴突顶梢枯死程度的明显下降(图13)。mapc的移植几乎完全消除了在此时正常观察到的轴突顶梢枯死,此研究已经表明轴突顶梢枯死是由活化的巨噬细胞的渗入直接导致的。因此,mapc在受损伤的脊髓内的存在足以减少体内巨噬细胞诱导的轴突顶梢枯死。
实施例iv
损伤中心的波形蛋白/ng2+少突胶质细胞前体细胞在巨噬细胞渗入的时间前后开始扩增,并且神经轴索断裂的纤维的末端与此细胞群体相关。这表明cns损伤内的ng2+细胞起稳定轴突作用,这使它们成为阻止巨噬细胞介导的回缩的理想候选者。在体外培养1天后向drg培养物加入来自成年小鼠脊髓的ng2+神经胶质细胞。在第2天,在30分钟的基线观察期后,将nr8383巨噬细胞加入缩时培养皿中并再观察2.5小时。与drg共培养的ng2+神经胶质细胞的存在不足以阻止巨噬细胞诱导的轴突回缩(n=5)。在图14中,在巨噬细胞接触后轴突回缩并在ng2+胶质细胞上稳固。
方法
1.drg解离
如现有技术所述收集drg(tometal.,2004;daviesetal.,1999)。简言之,从成年雌性sprague-dawley大鼠(harlan)上切下drg。除去中心根和外周根,并在胶原酶ii(200u/ml,worthington)和dispaseii(2.5u/ml,roche)的hbss溶液中孵育神经节。将消化的drg冲洗并在新鲜hbss-cmf中轻轻研磨三次,然后低速离心。然后将解离的drg重悬浮在补充有b-27、glutamax和青霉素/链霉素(均来自invitrogen)的neurobasal-a培养基中并计数。将drg以3000个细胞/ml的密度铺板于delta-t培养皿上(fisher,pittsburgh,pa),共6000个细胞/培养皿。
2.缩时培养皿制备
与tometal.,2004所述类似地制备delta-t细胞培养皿(fisher)。简言之,用2号钻头通过每个培养皿的上半部钻一个洞,以形成在缩时显微术过程中向培养物中添加细胞的进口。然后,将培养皿用无菌水冲洗并在室温下用聚-l-赖氨酸(0.1mg/ml,invitrogen)包被过夜。然后用无菌水冲洗培养皿并使其干燥。通过吸取2.0ul的集聚蛋白聚糖溶液(2.0mg/ml,sigma,溶于hbss-cmf,invitrogen)于培养物表面上并使其干燥,形成集聚蛋白聚糖梯度斑点。每个培养皿中放置6个斑点。在集聚蛋白聚糖斑点完全干燥后,将培养皿的整个表面在层粘连蛋白溶液(10ug/ml,bti的hbss-cmf溶液)中于37℃温浴3小时。然后撤除层粘连蛋白浴,立即将细胞铺板。
3.细胞系巨噬细胞培养物
如yinetal.,2003中所述培养成体sprague-dawley肺泡巨噬细胞系nr8383细胞(atcc#crl-2192)。简言之,在未包被的组织培养烧瓶(corning)中,将细胞在补充有15%fbs(sigma)、glutamax、penn/strep(invitrogen)和碳酸氢钠(sigma)的f-12k培养基(invitrogen)中培养,并每周加料2-3次。为制备用于缩时显微术实验的细胞系巨噬细胞,用胰蛋白酶/edta(invitrogen)收集细胞,洗涤3次,然后以1.0×106/ml的密度铺板于含无血清f-12k的未包被组织培养烧瓶中。在用于缩时实验前,用edta和细胞刮刀收集培养的细胞系巨噬细胞并以2.5×105/70ul的密度重悬浮于添加有hepes(50um,sigma)的neurobasal-a中。
4.mapc培养物
在大鼠mapc培养基中培养被gfp标记的sprague-dawley大鼠mapc,所述培养基由低葡萄糖dmem(invitrogen)、0.4×mcdb-201培养基(sigma)、1×its液体培养基添加物(sigma)、1mg/ml亚油酸-白蛋白(sigma)、100u/ml青霉素g钠盐或100μg/ml硫酸链霉素(invitrogen)、100μm2-p-l-抗坏血酸(sigma)、100ng/mlegf(sigma)、100ng/mlpdgf(r&dsystems)、50nm地塞米松(sigma)、1000u/mlesgro(chemicon)和2%胎牛血清(hyclone)组成。以1000个细胞/cm2的初始密度将所述培养物铺板于10ng/ml纤连蛋白(invitrogen)包被的150cm2组织培养烧瓶(corning)中,随后以200个细胞/cm2重新铺板。将细胞在37℃和5.0%co2维持于15ml的培养基/烧瓶中,使用胰蛋白酶/edta(invitrogen)每3-4天进行传代。
5.mapc条件培养基
如上述培养细胞,并在48小时后将条件培养基收集到50ml锥形管(bdbioscience)中。将所述条件培养基在4℃以400×g旋转5分钟,然后将上清液移至新的50ml锥形管中。然后将此条件培养基保存于4℃。
如上所述获得了mapc条件培养基,并将其用amiconmicroconultracelym-33,000mwco离心过滤器浓缩50倍。
6.mapc条件培养基处理的巨噬细胞
如上述培养nr8383大鼠巨噬细胞。在缩时显微术实验前的1天,用胰蛋白酶/edta(invitrogen)收集巨噬细胞,洗涤3次,然后以1.0×106/ml的密度铺板于含无血清f-12k的未包被组织培养烧瓶中。每1ml无血清f12k培养基中加入20ul的50倍所述浓缩mapc条件培养基,终浓度为1×。在用于缩时实验前,用edta和细胞刮刀收集培养的细胞系巨噬细胞并以2.5×105/70ul的密度重悬浮于添加有hepes(50um,sigma)的neurobasal-a中。
7.缩时显微术
将drg神经元于37℃孵育48小时,然后进行缩时成像。将含有hepes(50um,sigma)的neurobasal-a培养基加至培养物中,然后转至加热的台架仪器中。使用zeissaxiovert405m显微镜和100×油浸物镜,每30秒取得缩时成像,持续3小时。选择径直延伸到斑点边缘并具有特征营养不良形态的生长锥,观察神经元30分钟以确定基线表现,然后加入细胞或条件培养基,然后观察3小时。
对于细胞加入实验,从组织培养烧瓶中收集所培养的大鼠源mapc,洗涤3次并重悬浮于neurobasal-a培养基中。对于共培养实验,在24小时后将mapc(100,000个/培养皿)加至背根神经节神经元培养物中并于37℃再孵育24小时,然后进行缩时成像。
对于在缩时成像过程中将mapc条件培养基加至drg培养物中的实验,在观察基线生长锥30分钟后加入90ul的50×mapc-cm。
在30分钟基线成像后,将mapc条件培养基处理的巨噬细胞加至缩时培养物中(500,000个细胞/培养皿)。
使用metamorph软件分析延伸/回缩、生长速率、弯向和分枝。
8.免疫化学检测
在缩时成像后,将drg在4%pfa中固定并用抗iii型b-微管蛋白(1:500;sigma)、抗硫酸软骨素(cs-56,1:500,sigma)和抗gfp(1:500,invitrogen)进行免疫染色。
9.原代骨髓源巨噬细胞培养
如tobianetal.,2004中所述收集骨髓祖细胞。简言之,将股骨从成年雌性sprague-dawley大鼠(harlan)取下。除去股骨末端,将含有补充有10%fbs、glutamax、penn/strep、β-巯基乙醇和hepes(invitrogen)(d10f)的冷dmem的注射器插入股骨,冲出骨髓并收集。然后使所得细胞混合物通过70微米的滤器并离心。然后除去上清液,将所得细胞片状沉淀物重悬浮于akt裂解缓冲液(biowhitacre)中以裂解红细胞,然后离心。除去上清液并将含有骨髓前体细胞的片状沉淀物重悬浮并铺板于另补充有20%ladmac细胞系条件培养基(cliffordharding赠予)的上述dmem中以诱导分化为巨噬细胞。在培养第10天收集细胞进行实验。缩时实验前的1天,用胰蛋白酶/edta收集原代巨噬细胞,用d10f洗涤3次并以1.0×106/ml的密度铺板于含d10f的未包被陪替氏培养皿(falcon)中。第2天,用edta和细胞刮刀收集原代巨噬细胞并以5.0×105/70ul的密度重悬浮于加hepes的neurobasal-a中,用于缩时显微术实验。
10.背柱破碎损伤模型
对于所有手术步骤,均用吸入的异氟醚气(2%)麻醉成年雌性sprague-dawley大鼠(250-300g)。进行t1椎板切除术来暴露c8脊髓节的背面。用30号针头在距离中线0.75mm的两侧进行硬脑膜切开术。然后通过将dumont#3jeweler镊子在c8处插入背脊至1.0mm的深度;压住镊子,保持压力10秒钟并再重复2次来制造背柱破碎损伤。通过观察到白色物质的清除来确认损伤的完成。然后用凝胶膜覆盖硬脑膜上的洞。用4-0尼龙线缝合肌肉层,然后用外科封缝钉闭合皮肤。在闭合切口后,动物沿切口皮下接受麻卡因(1.0mg/kg)并肌内注射丁丙诺啡(0.1mg/kg)。手术后,在麻醉恢复期间用加热灯使动物保暖,并使其自由取食和饮水。将动物在损伤后2、4、7、14或28天处死。
11.细胞移植
从组织培养烧瓶中收集培养的大鼠源mapc或原代骨髓源巨噬细胞(已经用干扰素γ和lpc刺激24小时),在hbss-cmf中洗涤3次并以200,000细胞/ul的密度重悬浮于hbss-cmf中。背柱破碎损伤后,立即将1.0ul细胞悬浮液在单侧以0.5mm深度注射到右侧脊柱中。注射位置在中线侧面0.5mm和损伤边缘尾端0.5mm。通过连接于nanojectii(drummond)的拉制玻璃吸液管以15秒间隔用44次23.0nl的脉冲注射细胞。在最后一次注射后的2分钟从脊髓中抽出玻璃吸液管。在移植后,用凝胶膜覆盖注射位点,用4-0ethicon缝线缝合肌肉层,然后用外科封缝钉闭合皮肤。手术后,在麻醉恢复期间用加热灯使动物保暖,并使其自由取食和饮水。在损伤后2天或4天处死动物。
12.轴突标记
处死前2天,用texas-red缀合的3000mw葡聚糖单侧标记脊柱。简言之,暴露右后肢的坐骨神经,用dumont#3镊子挤压3次,持续10秒钟。在挤压位点处用hamilton注射器将1.0ul3000mw葡聚糖-texas-red的10%无菌水溶液注射到坐骨神经中。用4-0尼龙线缝合肌肉层,然后用外科封缝钉闭合皮肤。在闭合切口后,动物沿切口皮下接受麻卡因(1.0mg/kg)并肌内注射丁丙诺啡(0.1mg/kg)。手术后,在麻醉恢复期间用加热灯使动物保暖,并使其自由取食和饮水。将动物在标记后2天用过量异氟醚处死并用pbs继而用4%pfa灌注。收集组织并在4%pfa中后固定并进行免疫组织化学测定。
13.免疫组织化学测定
将组织在4%pfa后固定过夜,然后浸没在30%蔗糖中过夜,在oct封装介质中冷冻,然后在恒切片机上切成20um的纵向切片。然后将组织用抗gfap/alexafluor-405、抗ed-l/alexafluor-594或-633、抗gfp/alexafluor-488和抗波形蛋白/alexafluor-633染色。然后在zeissaxiovert510激光扫描共焦显微镜上在10×放大率下成像。
14.轴突顶梢枯死定量
为定量轴突顶梢枯死,对每只动物分析了始于脊髓背面下200um深处的三个连续切片。通过特征性gfap和/或波形蛋白染色模式确定损伤中心,然后使用zeisslsm5imagebrowser软件确定中心位置。然后测量最远深入到损伤中的5个标记轴突的末端与损伤中心的距离。将一组中所有动物的所有切片的测量值平均以得出每个时间点的顶梢枯死平均距离。
实施例v
间充质干细胞可商业购得。例如,ratmesenchymalstemcellkit(milliporecatalogno.scr026)提供从成年fisher344大鼠的骨髓分离的即用间充质原代干细胞,以及用于鉴定间充质干细胞群体的一组阳性和阴性选择标志物。阳性细胞标志物包括针对间充质干细胞上存在的两种细胞表面分子(整合素b1和cd45)的抗体。阴性细胞标志物包括针对间充质干细胞不表达的两种特定造血细胞表面标志物(白细胞上存在的cd14以及单核细胞和巨噬细胞上存在的cd45)的抗体。对这些间充质干细胞评定减少回缩的能力,并发现这些间充质干细胞在体外(神经胶质疤痕)减少回缩(减少附着)。
实施例vi
对在5μg/ml层粘连蛋白上培养的成体drg进行mapc条件培养基处理可促进神经突生长。参见图16。对向其中添加有含有neurobasal-a的培养基并添加有mapc条件培养基、对照培养基或没有另外添加培养基的各组测量每个解离的drg的最长轴突。所有条件彼此均显著不同,单因素anova,*p<0.0001。b,表示未处理的drg神经元的平均向外生长量的16×图像。c,表示用mapc条件培养基预处理的drg的平均向外生长量的16×图像。
此结果可能是由于mapc的神经刺激作用或免疫调制作用,或者是由于两种作用的结合。为解决此问题,进行了一系列条件培养基实验。用新鲜mapc条件培养基将解离的drg神经元处理24小时并与培养基对照进行比较。测量每个drg的最长神经突。新鲜mapc条件培养基显著增加了层粘连蛋白上的向外生长。这表明mapc分泌一种或多种对成体神经元具有神经刺激作用的生长因子。预先冷冻的mapc条件培养基对神经元不具有同样的作用,表明所述因子在冷冻过程中改变或失活。
参考文献
arantesetal.,neuroreport11:177-181(2000)
bakeretal.,expneurol205:82-91(2007)
bakerandhagg,jneurotrauma22:415-428(2005)
baschetal.,(1983)
batinic,d.,etal.,(1990)
borgensetal.,jcompneurol250:157-167(1986)
brownetal.,eurjneurosci9:2057-2063(1997)
buschandsilver,curropinneurobiol17:120-127(2007)
caiz.h.,etal.,(1988)
caoetal.,neurorehabilneuralrepair(2007)
chang,p.,etal.,(1999)
chang,t.m.,(1992)
daviesetal.,jneurosci19:5810-5822(1999)
deetal.,jneuropatholexpneurol61:237-244(2002)
desimoneetal.,molneurobio29:197-212(2004)
donnellyandpopovich,expneurol209:378-388(2008)
farretal.,aplysiajneurobiol46:89-96(2001)
fayazandtator,jneuroscimethods102:69-79(2000)
fishmanandkelley,neurology34:1161-1167(1984)
fitchandsilverexpneurol148:587-603(1997)
fitchetal.,jneurosci19:8182-8198(1999)
grimpeandsilver,jneurosci24:1393-1397(2004)
hilletal.,expneurol171:153-169(2001)
hohlfeldetal.,neurology68:s58-s63(2007)
houleandjin,expneurol169:208-217(2001)
iizukaetal.,jneurosurg66:595-603(1987)
jonesetal.,expneurol182:399-411(2003)
kelmetal.,currbiol4:965-972(1994)
kerschensteineretal.,natmed11:572-577(2005)
kwonetal.,procnatlacadsciusa99:3246-3251(2002)
liandraisman,expneurol134:102-111(1995)
liuetal.,jneurosci26:3087-3101(2006)
longbrakeetal.,jneurochem102:1083-1094(2007)
matthew,h.w.,etal.,(1991)
mckeonetal.,jneurosci11:3398-3411(1991)
mcphailetal.,eurj.neurosci20:1984-1994(2004)
morgensternetal.,progbrainres137:313-322(2002)
muschler,g.f.,etal.,(1997)
oudegaetal.,eurj.neurosci11:2453-2464(1999)
prewittetal.,expneurol148:433-443(1997)
popovichetal.,jcompneurol377:443-464(1997)
popovichandlongbrake,natrevneurosci9:481-493(2008)
popovichetal.,expneurol158:351-365(1999)
raffetal.,science296:868-871(2002)
ramonycajal,london:oxfordup(1928)
rapalinoetal.,natmed4:814-821(1998)
schwartzetal.,neurosurgery44:1041-1045(1999b)
schwartzetal.,amjneuroradial26:7-18(2005)
seifctal.,jneurotrauma24:1513-1528(2007)
silverandmiller,natrevneurosci5:146-156(2004)
snowetal.,expneurol109:111-130(1990)
sobeletal.,expneurol54:202-213(1995)
steinmetzetal.,jneurosci25:8066-8076(2005)
stirlingetal,jneurosci24:2182-2190(2004)
tangetal.,jneuroscires71;427-444(2003)
tangetal.,jcellbiol138:1355-1366(1997)
tobianetal.,jimmunol172:5277-5286(2004)
tometal.,jneurosci24:6531-6539(2004)
umeharaetal.,immunolcellbiol79:298-302(2001)
vanrooijenetal.,trendsbiotechnol15:178-185(1997)
vanrooijenandvankesteren-hendrikx,jliposomeres12:81-94(2002)
vanrooijenandsanders,jimmunomethods174:83-93(1994)
watanabeetal.,jneurotrauma16:255-265(1999)
wysockiandsato,(1978)
xieandzheng,expneurol209:302-312(2008)
yanagi,k.etal.,(1989)
yinetal.,j.neurosci23:2284-2293(2003)
yiuandhe,natrevneurosci7:617-627(2006)
zujovicetal.,glia29:305-315(2000)
- 还没有人留言评论。精彩留言会获得点赞!
- SPNP‑IX在制备Kv4.2通道工具试剂中的应用的制造方法与工艺
- SPNP‑IX在制备Kv4.3通道工具试剂中的应用的制造方法与工艺
- SPNP‑27在制备Nav1.5通道抑制剂中的应用的制造方法与工艺
- SPNP‑27在制备Nav1.3通道抑制剂中的应用的制造方法与工艺
- 一种多肽在制备Nav1.3钠通道工具试剂中的应用的制造方法与工艺
- SPKP‑XI在制备Kv4.2通道抑制剂中的应用的制造方法与工艺
- 基于水凝胶溶胀特性的可控自变形神经微电极的制造方法与工艺
- SPKP‑XI在制备Kv4.1钾通道抑制剂中的应用的制造方法与工艺
- 小驳骨提取物作为神经保护剂的应用的制造方法与工艺
- 巴戟天低聚果糖提取物在制备防治阿尔茨海默症药物、保健品或食品中的应用的制造方法与工艺