测定多肽的脱酰胺和免疫原性的系统和方法与流程
相关申请本申请要求2018年10月24日提交的美国临时申请第62/750,022号的权益,其全部内容通过引用合并于此。
背景技术:
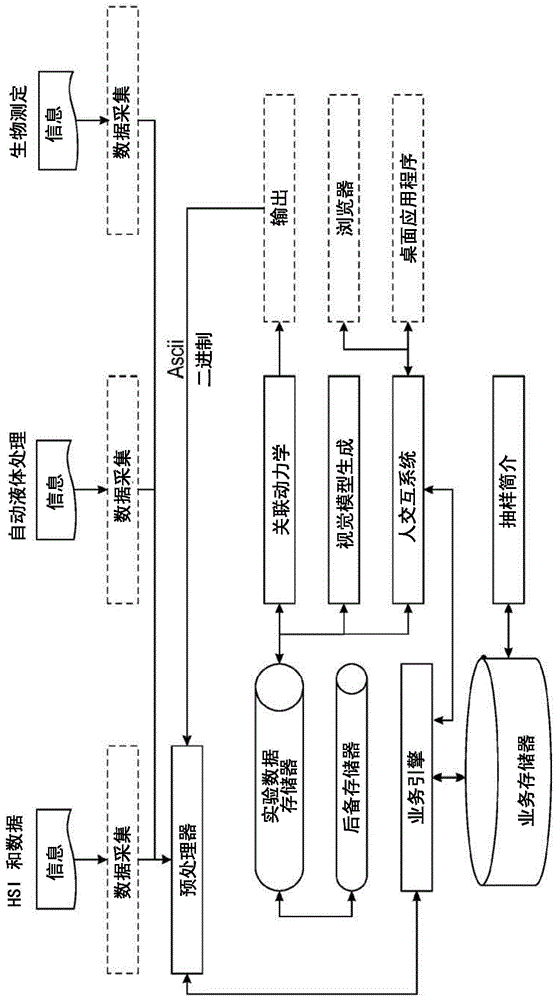
:诸如蛋白质治疗剂的候选药物的高损耗率是药物开发的主要成本驱动因素,并且仍然是生物制药行业的主要挑战。免疫原性、蛋白质聚集、脱酰胺和氧化是监管机构关注的问题,因为它们可能会降低药效和患者的安全性而造成影响。蛋白质是复杂的分子,在不同的条件下,可能会发生非酶促的天冬酰胺转化为天冬氨酸或谷氨酰胺转化为谷氨酸的脱酰胺作用。仅在高ph条件下观察到异构体产物的出现。具体地说,蛋白质中的脱酰胺过程与低和高ph条件以及热应力有关。因此,发生的风险包括:(1)治疗性蛋白质的细胞培养生产过程中的上游加工,和/或(2)纯化,(3)病毒清除以及储存和输送过程中的下游加工,和(4)热应力和/或低/高ph条件。当前以高通量方式评估溶液中蛋白质的脱酰胺作用存在局限性。诸如hplc、nmr和ms的当前技术在可分析的样品数量,脱酰胺作用后蛋白质稳定性的评估以及对功效和安全性的影响方面存在局限性。脱酰胺作用的机理是动力学驱动的,并且要求相邻的残基(n+1)小以防止空间位阻。允许在–nh2基团水解之后形成琥珀酰亚胺中间体,从而导致带负电荷的残基。用于检测脱酰胺作用的当前技术是基于通过高效液相色谱法(“hplc”)如离子交换(iex)或反相来分离电荷变异体。然后,使用核磁共振波谱(“nmr”)来鉴定蛋白质内的结构和一级序列变化。质谱(“ms”)也已经开发出来,仅在高ph下用于天冬酰胺的异天冬氨酸脱酰胺检测。ms技术需要全长电荷变异蛋白的片段化和肽图分析,以专门检测异天冬氨酸质量差异。这是一个复杂且耗时的过程。技术实现要素:例如,根据以下描述的各个方面示出了本主题技术。下面描述本主题技术的方面的各种实例。这些仅作为实例提供,并不限制本主题技术。本主题技术的各方面提供了用于确定和评估在热胁迫下蛋白质样品的脱酰胺作用的系统和方法。特别地,该系统和方法提供用于确定评估谷氨酰胺和天冬酰胺内的脱酰胺作用、确定聚集体大小、同一性、聚集的程度和机制以及稳定性、靶标结合和生物测定的验证。该系统和方法允许治疗性蛋白质的开发性和可比性评估,而与它们的分子量,翻译后修饰和/或配制条件无关,每个样品只有1μl体积。此外,经验结果可以直接影响蛋白质设计和重组工程。根据本主题技术的一方面,本文描述的系统和方法涉及获得和分析蛋白质的光谱数据,包括红外(ir)光谱,诸如使用量子级联激光(“qcl”)显微镜获得的ir光谱。该系统和方法提供实时的高通量高光谱成像(“hsi”),其允许在热应力期间监测溶液中的蛋白质阵列。与某些现有的监测蛋白质的方法不同,该方法不需要分离技术,并且不包含流动通道。该系统使用qcl透射显微镜,该显微镜具有基于第一原理的线性响应检测、精确的热控制和独特的加热单元支架,并带有多路复用阵列载玻片池,可满足固定体积的要求。这提供了一种快速采集系统,其速度比傅立叶变换红外(“ft-ir”)显微镜快200倍,并且具有增强的信噪比(“snr”),能够确定聚集体的大小、特性、程度和机制。如本文所述,使用分析算法处理qcl显微镜光谱数据,以确定脱酰胺的存在和程度。该系统和方法还可以处理光谱数据以监测和评估胶体稳定性,或评估其他应力条件,诸如ph。该系统和方法每天提供对数百个样品的分析。光谱分析中使用的方法用于绘制脱酰胺区域,鉴定易于聚集的区域并建立域稳定性。系统中包括的相关动力学软件,用于实现方法的各个方面,可以关联侧链模式,这些模式用于在应力条件下探测溶液中的蛋白质。结果,数据具有很高的信息量和统计学上的稳健性。根据技术的一方面,系统和方法使用hsi实时监视和分析在热和/或化学应力下蛋白质脱酰胺事件的发生,包括在溶液中使用一系列治疗性蛋白质。此类监视和分析的结果具有预测意义,同时可以绘制容易脱酰胺的位点图。例如,脱酰胺可以预测免疫原性和/或聚集趋势。结果在统计上是可靠的。此外,通过分析蛋白质候选品的变体,可以为发现阶段早期的临床前候选品选择提供全面的证据。此外,本主题技术还能够描述蛋白质聚集机制和展开,从而提供可导致免疫原性的事件的分子细节。基于使用qcl显微镜获得的his,治疗性蛋白质候选品的选择基于本文所述数据处理和分析方法的预测能力。当前用于评估脱酰胺作用发生的其他分析工具(包括hplc、nmr和ms)也可以与本文公开的系统和方法结合使用,从而可以选择稳定的候选品药物,从而降低候选品药物撤出的风险,同时确保疗效和安全。作为蛋白质治疗剂开发的一部分,可以将本文所述的用于处理数据的方法,系统和说明用于评估脱酰胺作用、聚集作用和免疫原性的潜力。使用本主题技术对蛋白质样品进行的研究证明了溶液中一组蛋白质中天冬酰胺和/或谷氨酰胺残基的脱酰胺作用的评估和测定,并通过提供体外或原位检测药品(“ds”)和/或药物产品(“dp”)聚集的直接方法,提供了免疫原性和抗药物抗体(ada)生物测定的验证。本主题技术的附加特征和优点将在下面的描述中阐述,并且部分地从该描述中将变得显而易见,或者可以通过本主题技术的实践而获知。通过在书面描述及其权利要求以及附图中特别指出的结构,将实现和获得本主题技术的优点。应当理解,前面的一般描述和下面的详细描述都是示例性和说明性的,并且旨在提供对所要求保护的主题技术的进一步解释。附图说明被包括以提供对主题技术的进一步理解并且被并入本说明书中并构成其一部分的附图示出了主题技术的各个方面,并且附图与说明书一起用于解释主题技术的原理。图1示出了根据本主题技术的一些方面的示例性计算系统的图。图2a示出了根据本主题技术的一些方面的流程图,该流程图指示验证和准备输入数据的示例性方法的操作。图2b示出了指示根据本主题技术的一些方面的示例性方法的操作的流程图。图3示出了多阶段分析的结果。图4a示出了使用低倍率物镜产生的高光谱图像(hsi),其在28℃和56℃下以1μg/μl的pdsnistmabrm8671的视场为2×2mm2。图4b示出了使用低倍率物镜产生的高光谱图像(hsi),其在28℃和56℃下以2μg/μl的nistmabrm8671的视场为2×2mm2。图4c示出了使用低倍率物镜产生的高光谱图像(hsi),其在28℃和56℃下以2.4μg/μl的pnistmab候选品rm8670的视场为2×2mm2。图5示出了在28℃至56℃的温度范围内以4℃的温度间隔:28℃、32℃、36℃、40℃、44℃、48℃、52℃、56℃在1780至1450cm-1的光谱区域中的具有重叠的l-组氨酸和h2o吸收的酰胺i和ii谱带的qclir光谱叠加。图6a示出了在1780-1450cm-1的光谱区域内对应于1μg/μl的pdsnistmab的28-56℃的温度范围的酰胺i和酰胺ii谱带的qcl光谱叠加。图6b示出了基于图6a所示的qcl光谱叠加数据生成的同步图。图6c示出了基于图6a所示的qcl光谱叠加数据生成的异步图。图7a示出了在1780-1450cm-1的光谱区域内对应于1μg/μl的nistmab的28-56℃的温度范围的酰胺i和酰胺ii谱带的qcl光谱叠加。图7b示出了基于图7a所示的qcl光谱叠加数据生成的同步图。图7c示出了基于图7a所示的qcl光谱叠加数据生成的异步图。图8a示出了在1780-1450cm-1的光谱区域内对应于1.5μg/μl的nistmab候选品的28-56℃的温度范围的酰胺i和酰胺ii谱带的qcl光谱叠加。图8b示出了基于图8a所示的qcl光谱叠加数据生成的同步图。图8c示出了基于图8a所示的qcl光谱叠加数据生成的异步图。图9a示出了在28-56℃的温度范围内在1μg/μl下受到热应力的pdsnistmab的事件的顺序。图9b示出了在28-56℃的温度范围内在1μg/μl下受到热应力的nistmab的事件的顺序。图9c示出了在28-56℃的温度范围内在1.5μg/μl下受到热应力的nistmab候选品的事件的顺序。图10a示出了在1780-1450cm-1的光谱区域内对应于2μg/μl的pdsnistmab的28-56℃的温度范围的酰胺i和酰胺ii谱带的qcl光谱叠加。图10b示出了基于图10a所示的qcl光谱叠加数据生成的同步图。图10c示出了基于图10a所示的qcl光谱叠加数据生成的异步图。图11a示出了在1780-1450cm-1的光谱区域内对应于2μg/μl的nistmab的28-56℃的温度范围的酰胺i和酰胺ii谱带的qcl光谱叠加。图11b示出了基于图11a所示的qcl光谱叠加数据生成的同步图。图11c示出了基于图11a所示的qcl光谱叠加数据生成的异步图。图12a示出了在1780-1450cm-1的光谱区域内对应于2.4μg/μl的nistmab候选品的28-56℃的温度范围的酰胺i和酰胺ii谱带的qcl光谱叠加。图12b示出了基于图12a所示的qcl光谱叠加数据生成的同步图。图12c示出了基于图12a所示的qcl光谱叠加数据生成的异步图。图13a示出了在28-56℃的温度范围内在2μg/μl下受到热应力的pdsnistmab的事件的顺序。图13b示出了在28-56℃的温度范围内在2μg/μl下受到热应力的nistmab的事件的顺序。图13c示出了在28-56℃的温度范围内在2.4μg/μl下受到热应力的nistmab候选品的事件的顺序。图14a示出了在1780-1450cm-1的光谱区域内对应于2.8μg/μl的nistmab样本的28-56℃的温度范围的酰胺i和酰胺ii谱带的qcl光谱叠加。图14b示出了基于图14a所示的qcl光谱叠加数据生成的同步图。图14c示出了基于图14a所示的qcl光谱叠加数据生成的异步图。图15a示出了1780-1450cm-1的光谱区域内对应于10.0μg/μl的nistmab候选品的28-56℃的温度范围的酰胺i和酰胺ii谱带的qcl光谱叠加。图15b示出了基于图15a中所示的qcl光谱叠加数据生成的同步图。图15c示出了基于图15a中所示的qcl光谱叠加数据生成的异步图。图16a示出了在28-56℃的温度范围内在2.8μg/μl下受到热应力的pdsnistmab的事件的顺序。图16b示出了在28-56℃的温度范围内在10.0μg/μl下受到热应力的nistmab候选品的事件的顺序。图17a、图17b和17c是pdsnistmab标准品(rm8671)、nistmab标准品(rm8671)和nistmab候选品(rm8670)在低浓度下的异步图,展示脱酰胺作用的证据。图18a、图18b和图18c是示出与脱酰胺相关的交叉峰内的强度变化的条形图。图19是在热应力期间低浓度下的nistmab候选品的异步图,展示了脱酰胺作用的证据。图20是示出与脱酰胺相关的交叉峰内的强度变化的条形图。图21是天冬酰胺的脱酰胺作用机理以及用于监测热应力期间事件的关键振动模式的示意图。图22是可用于实现本主题技术的系统和方法的示例性平台技术的图示。图23是示出根据本主题技术的一些方面的实验方法的示例性设计的操作的流程图。图24是指示用于ada筛选和免疫原性风险评估的示例性方法的操作的流程图。图25是指示可以使用本文描述的平台技术和方法执行的示例性比较分析的操作的流程图。图26示出了计算系统的示例图。具体实施方式在下面的详细描述中,阐述了具体细节以提供对主题技术的理解。然而,对于本领域的普通技术人员将显而易见的是,可以在没有这些特定细节中的一些的情况下实践本主题技术。在其他情况下,未详细示出公知的结构和技术,以免模糊本技术。蛋白质是由排列成线性链并通过相邻氨基酸残基的羧基和氨基之间的肽键结合在一起的氨基酸组成的大型有机化合物。大多数蛋白质折叠成独特的3维结构。蛋白质自然折叠的形状称为其天然状态。尽管许多蛋白质可以无辅助地,简单地通过氨基酸的化学特性折叠,但其他蛋白质则需要分子伴侣的帮助才能折叠成天然状态。蛋白质结构有四个不同方面:·一级结构:氨基酸序列。·二级结构:通过氢键稳定的有规律地重复的局部结构。因为二级结构是局部的,所以在同一蛋白质分子中可以存在许多不同二级结构的区域。·三级结构:单个蛋白质分子的整体形状;二级结构彼此之间的空间关系。·四级结构:由于一个以上的蛋白质分子(在此情况下通常称为蛋白质亚基)的相互作用而产生的形状或结构,其起较大装配体或蛋白质复合物的作用。蛋白质不是完全刚性的分子。除了这些结构水平外,蛋白质在执行其生物学功能时可能会在几种相关结构之间变换。在这些功能性重排的背景下,这些三级或四级结构通常被称为“构象”,并且它们之间的过渡被称为构象变化。蛋白质聚集被表征为错误折叠的,刚性的蛋白质分组,其被认为是在整个工业生物过程中普遍存在的现象。聚集被认为是蛋白质降解的主要方式,通常导致蛋白质的免疫原性和生物活性的丧失。在各种生物医学情况下,蛋白质聚集至关重要,从阿尔茨海默氏病和帕金森氏病等异常疾病状态到蛋白质药物的生产、稳定性和递送。脱酰胺作用被认为是蛋白质生物合成后蛋白质的翻译后修饰,其可能会影响治疗性蛋白质的稳定性、结构和功效,并可能导致聚集,从而导致不良的免疫反应,诸如免疫原性和抗药物抗体反应(ada)。表现出脱酰胺作用的残基是天冬酰胺和含量较低的谷氨酰胺。脱酰胺作用导致天冬酰胺转化为天冬氨酸和/或谷氨酰胺转化为谷氨酸。在该位点引入的负电荷会导致蛋白质的稳定性下降,从而导致蛋白质聚集,降解,松散的结合选择性以及与其靶标的亲和力,从而导致功效和安全性下降。当天冬酰胺的邻近残基(位置n+1)是甘氨酸时,容易发生天冬酰胺翻译后修饰,从而降低了琥珀酰亚胺中间体形成的空间位阻,产生天冬氨酸或异天冬氨酸。脱酰胺作用发生在不存在任何酶的情况下,并且在高ph和/或高温下会加速脱酰胺作用。脱酰胺作用可能表明细胞内蛋白质的降解,从而降低了细胞内治疗性蛋白质的半衰期,从而潜在地影响pk/pd。本主题技术的各方面提供了一种快速,准确和可再现的技术,用于实时监测溶液中治疗性蛋白质阵列在热和/或化学应力下的脱酰胺作用。例如,本主题技术提供了一种技术,用于评估和监测溶液中的治疗性蛋白质在高和低ph值下在热应力下的天冬酰胺和谷氨酰胺脱酰胺作用。本文所述的系统和方法允许在变化的浓度和热应力条件下对全长单克隆抗体进行可比性评估。实时高通量hsi的比较允许在热应力期间监控溶液中的蛋白质阵列。可以分析来自hsi的光谱数据以生成协方差(差)光谱。然后可以将2dir相关技术应用于协方差谱,生成同步图和异步图。可以分析同步图和异步图中的强度峰值,以及强度的变化。分析了目标光谱区域内强度和峰位移的变化,这些变化代表了蛋白质在热应力下的行为。峰之间的相关性可用于建立样品发生的脱酰胺作用。可以通过确定阵列中每个样品在热应力期间分子事件的顺序来提供对溶液中蛋白质行为的描述。可以绘制发生脱酰胺作用的区域的图,可以根据脱酰胺作用的程度确定被检查蛋白质的稳定性,并根据分子事件的顺序确定热稳定性。本文描述的计算方法和系统相对于蛋白质的现有分析提供了显著的改进。本文所述的计算方法和系统以有助于高效且有意义的分析的形式生成并存储数据,无需使用几台设备。因此,本文描述的计算方法和系统可以提高用于评估候选药物的光谱数据分析的效率。本主题技术的各方面包括使用二维相关光谱法(“2dcos”)和/或二维共分布光谱法(“2dcds”)来提供有关蛋白质治疗剂脱酰胺作用的程度和机理的基本信息。本文所述的方法可包括分析侧链模式作为内部探针,提供证实蛋白质内结构基序或结构域稳定性的信息。已经表明,本文所述的方法通过符合设计质量(“qbd”)的实验设计(“doe”)方法在高通量可开发性和可比性评估(“ht-dca”)中有用。根据一些实施例,光谱分析可以分阶段进行,例如如图3所示。根据一些实施例,溶液样品中的蛋白质被扰动(热、化学、压力或声学),从而引起振动光谱的动态波动。在阶段310,可以收集和/或分析原始光谱数据。可以以规则的温度间隔并以顺序的方式获取光谱数据。根据一些实施例,可以对数据进行基线校正。根据一些实施例,光谱数据可以用于确定脱酰胺事件的存在和程度。为此,从随后的光谱中减去第一低温平均光谱,以生成动态光谱。在阶段320中,可以通过从所有随后的光谱中减去第一低温平均光谱(24℃)来产生协方差(差)光谱。因此,协方差(差)谱包含正峰和负峰;也称为彼此同相和异相。值得注意的是,本文描述的方法不需要从光谱数据中手动减去水或其他参考样品(例如,溶质)。这种手动减法是蛋白质光谱分析中经常发生的高度主观的步骤。取而代之的是,本文所述的方法基于目标样品的扰动产生差异光谱数据集。然后可以将其输出用于进一步分析。通过从所有后续光谱中减去第一低温平均光谱,该光谱具有重叠的水谱带和酰胺i谱带,可自动减去水的光谱贡献。即,减去水和不受热应力扰动的所有蛋白质振动模式的贡献,从而允许仅评估在热应力下在目标光谱区域(1780-1450cm-1)中发生的变化。然后,如步骤330所示,通过应用2dir相关技术获得溶液中蛋白质的详细分子评估。在步骤330中,可以将2dir相关技术应用于生成同步图(步骤340)和异步图(阶段350)。例如,可以对光谱数据进行快速傅立叶变换(“fft”)以生成复杂矩阵,通过互相关积从同步矩阵中获得强度矩阵,该复杂矩阵是通过生成同步图和异步图来获得的。同步图表示在目标光谱区域内的扰动期间发生的总体强度变化。在该图的对角线上是在整个光谱中变化的峰或带(称为自动峰)。在非对角线上是交叉峰,其显示了自动峰之间的相关性,即观察到的二级结构变化之间的关系。同步图可用于关联同相峰值强度的变化或偏移。在同步相关光谱中,对角线位置处的自动峰表示扰动引起的光谱信号动态波动的程度。交叉峰表示在两个不同波数下光谱信号的同时变化,表明强度变化的耦合或相关起源。如果交叉峰的符号为正,则对应波数的强度会一起增加或减少。如果符号为负,则一个在增加,而另一个在减少。异步图仅包含交叉峰,该交叉峰用于确定根据热应力或其他施加的扰动而发生的分子事件的顺序。异步图可用于关联根据热应力发生的异相峰强度变化或位移。例如,可以观察到与δ(nh2)振动模式相关的1612.7cm-1处的降低的天冬酰胺强度以及在1572.0cm-1的v(coo–)振动模式下观察到的天冬氨酸强度的同时增加,其表示脱酰胺。在异步相关光谱中,仅当信号波动的某些傅立叶频率分量的强度彼此异相变化时,才会出现交叉峰。如果在波数v1之前在波数v2处发生强度变化,则交叉峰的符号为正。如果在波数v1之后在波数v2处发生强度变化,则交叉峰的符号为负。如果转换为同步图的相同异步交叉峰位置位于负区域(φ(v1,v2)<0),则上述符号规则将被逆转。2dir相关光谱法可用于解析复杂的谱带,诸如酰胺i和ii谱带。尤其是,2dir相关性通过将峰扩展为二维尺寸,从而提高了宽带潜在峰如酰胺i和ii谱带的光谱分辨率。如前所述,2dir相关技术会生成一个同步图和一个异步图。这些图在本质上是对称的,出于讨论目的,将参考顶部三角形进行分析。同步图(在340处显示)包含两种类型的峰:(a)自动峰,其是对角线上的正峰,并且(b)交叉峰,其是非对角峰,可以是正峰也可以是负峰。异步图(显示为350)仅由与异相峰相关的交叉峰组成。结果,该曲线图显示出更大的光谱分辨率增强。以下规则可用于建立分子事件的顺序:i.如果异步交叉峰值ν2为正,则ν2在ν1之前(ν2→ν1)受到扰动。ii.如果异步交叉峰值ν2为负,则ν2在ν1之后(ν2←ν1)受到扰动。iii.如果同步交叉峰(非对角峰,图3中未显示)为正,则使用异步图(规则i和ii)专门确定事件的顺序。iv.如果同步图包含负的交叉峰,并且相应的异步交叉峰为正,则顺序相反。v.如果同步图包含负的交叉峰,并且相应的异步交叉峰为负,则顺序保持不变。可以为在ν2轴中观察到的每个峰确定事件的顺序。可以提供一个表,总结每个事件的顺序。在阶段360中,使用总结每个事件的顺序的表来生成事件的顺序图。每个步骤(事件)的顶部是交叉峰的光谱信息,ν2,而每个步骤的底部是每个事件的相应峰分配或生化信息,其扰动顺序为函数温度。本文提供了实例。本领域技术人员将注意力吸引到dr.isaonoda,“two-dimensionalco-distributionspectroscopytodeterminethesequentialorderofdistributedpresenceofspecies”,journalofmolecularstructure,vol.1069,pp.51-54,其描述了适用于2dir相关性分析的算法。isaonoda博士开发的2dir相关光谱的摘要如下,使用了样品蛋白质的连续光谱的红外序列。样品蛋白质可包括单克隆抗体(mab)。例如,可以使用qclir光谱作为扰动的函数,在这种情况下为热应力(28-56℃),可通过从所有后续光谱中减去初始光谱来用于获得协方差(差)光谱数据集。可以获得一组离散采样的光谱a(vj,tk)以用于在外部扰动影响下测量的系统,该扰动会引起观察到的光谱强度的变化。光谱变量vj,其中,j=1,2,...,n,例如可以是波数、频率、散射角等,而另一个变量tk,其中,k=1,2,...,m,表示所施加的扰动的效果,例如时间、温度和电势。仅将在t1和tm之间的明确定义的观察间隔内获得的顺序采样的光谱数据集用于2dir相关性分析。为了简单起见,此处使用波数和时间来指定两个变量,但是可以理解,使用其他物理变量也是有效的。在2dir相关光谱法中使用的协方差(差)光谱定义为:其中,是生成协方差光谱的数据集的初始光谱。在没有参考状态的先验知识的情况下,参考光谱还可以设置为在t1和tm之间的观察间隔内的时间平均光谱。协方差光谱数据的同步2d相关强度定义为:协方差光谱数据的异步2d相关强度定义为:术语nij是所谓的希尔伯特-诺达变换矩阵的元素,由下式给出:正是对此差异光谱数据集应用了互相关函数,这导致了两个单独的但对称的2d图。根据两个独立波数轴ν1和ν2得到的相关强度φ(v1,v2)是同步图。根据两个独立波数ν1和ν2得到的相关强度ψ(v1,v2)是异步图。同步图包含对角线上的正峰(称为自动峰),并总结了光谱数据集中观察到的变化,在此同步图中建立的关系与彼此同相(同时发生)的光谱强度变化相关联。异步图是等高线图,它与异相强度变化相关,提高了目标光谱区域的分辨率,并且可以轻松地与同步图区分开,因为它在对角线上没有峰值。这两个图都包含非对角峰,其称为交叉峰,这些峰与观察到的光谱变化相关。观察到的光谱强度变化是由于施加到蛋白质样品的增量热应力引起的。因此,来自同步图和异步图的信息都可以确定分子事件的顺序,这些分子事件是在遵循诺达规则的应力事件或条件期间发生的。同步图和异步图本质上是对称的,出于讨论目的,我们将始终参考顶部三角形进行分析。为了确定分子事件的顺序,我们从分辨率提高最大的图(即异步图)开始:i.如果异步交叉峰值ν2为正,则ν2在ν1之前(ν2→ν1)受到扰动。ii.如果异步交叉峰值ν2为负,则ν2在ν1之后(ν2←ν1)受到扰动。iii.如果相应的同步交叉峰为正,则使用异步图(规则i和ii)确定事件的顺序。iv.但是,如果相应的同步交叉峰值为负,而异步交叉峰值为正,则顺序相反。可以为在v2轴上观察到的定义光谱区域中的每个目标峰建立分子事件的顺序。然后将目标峰用于评估在热应力下蛋白质中的脱酰胺作用,如本文所述。再次参考图xx,在阶段370,共分布相关图提供了溶液中蛋白质群体分布的扰动区域(阈值80%)。共分布相关性分析提供了溶液中蛋白质分布群体的普遍行为。本领域技术人员将注意力吸引到isaonoda,“two-dimensionalco-distributionspectroscopytodeterminethesequentialorderofdistributedpresenceofspecies”,journalofmolecularstructure,vol.1069,pp.54-56,其描述了适用于2dcds分析的算法。对于在t1≤tk≤tm的观察间隔内和按等式(2)给出的时间平均光谱依次获得一组m个随时间变化的光谱a(vj,tk),特征(时间)指标定义为:这里使用的动态光谱与等式(1)中定义的相同。在给定波数为vj时观察到的光谱强度分布的相应特征时间为:再次理解,此处使用的时间是对所施加扰动的代表变量的一般描述,因此可以用任何其他适当的物理变量如温度、浓度和压力代替。根据实验条件选择特定的条件。特征时间是沿着由t1和tm之间的观察间隔界定的时间轴的光谱强度a(vj,tk)的分布密度的第一个矩(大约是时间轴的原点,即t=0)。它对应于观察到的随时间分布的光谱强度的重心位置。给定在两个不同波数v1和v2下测得的光谱强度的时间分布的特征时间和同步和异步共分布光谱定义为:其中,t(v1,v2)是由下式给出的总联合差异:同步共分布强度γ(v1,v2)是沿着时间轴的两个独立光谱强度分布共存或重叠的度量。相反,异步共分布强度δ(v1,v2)是两个光谱信号分布差异的度量。术语“共同分布”表示两个单独分布的比较,将此度量与“相关”的概念区分开,“相关”的概念基于两个变化的比较。通过组合方程式5、6和8,异步共分布光谱的表达式为:δ(v1,v2)的值设置为零,如果遇到或者的条件,这表明在任一波数处都缺乏光谱强度信号。同步共分布光谱可以从以下关系式获得:在异步共分布光谱中,并且对于具有正号(即δ(v1,v2)=0)的交叉峰,相比于v2,在v1处的光谱强度的存在主要分布在沿时间轴的较早阶段。另一方面,如果δ(v1,v2)<0,则顺序相反。在δ(v1,v2)≈0的情况下,在两个波数上随时间变化观察到的光谱强度的平均分布是相似的。同步共分布峰的符号始终为正,这在一定程度上限制了同步光谱的信息内容,超出了对分布图案的重叠程度的明显定性度量。共分布(2dcds)分析能够提供蛋白质的稳定性、蛋白质中的聚集状态或以加权方式研究的任何过程的元素。2dcds可用于直接提供物种在应力(例如温度、浓度、ph等)可变轴上的分布存在顺序。该技术可以用作在直接识别中间物种存在时增强2dcos分析的补充工具。根据一些实施例,在观察间隔期间顺序地获得依赖于扰动的光谱。从光谱变化中得出2d相关光谱(同步光谱和异步光谱)。同步共分布强度是沿扰动轴的两个独立光谱强度分布的共存或重叠测量的。异步共分布强度被测量为两个光谱信号分布的差异。对于具有正号(即,δ(v1,v2)>0)的交叉峰,相比于v2,在v1处的光谱强度的存在主要分布在沿时间轴的较早阶段。另一方面,如果δ(v1,v2)<0,则顺序相反。在δ(v1,v2)≈0的情况下,在两个波数上随时间变化观察到的光谱强度的平均分布是相似的。2dcos分析之间的差异提供了由于扰动过程及其对样品的影响导致的路径的平均描述,而2dcds分析提供了扰动过程中分子(蛋白质)群体中的加权元素。2dcos和2dcds的结果是对由于扰动而在光谱数据中发生变化的元素的直接简化描述。根据一些实施例,例如如图1中所示,用于执行数据分析的系统可以至少包括所示的用于执行本文描述的方法的功能的组件。数据可以从多个来源获取,并且可以包含与使用qcl透射显微镜获取的hsi图像有关的信息,来自自动液体处理系统的信息以及来自生物测定的信息。所获取的数据可以被提供给一个或多个计算单元,包括预处理器和处理器,以进行分析。可以提供模块来执行或管理数据的分析。来自模块的信息也可以在网页浏览器、移动应用程序或桌面应用程序上实现或导出。这样的模块可以包括相关动力学模块,视觉模型生成器模块和/或人机交互模块。人机交互模块可以被提供为例如网络浏览器,移动应用程序或桌面应用程序。模块可以彼此通信。在一些实施例中,模块可以以软件(例如,子程序和代码)实现。例如,模块可以被存储在诸如实验存储器和/或备份存储器之类的存储器和/或数据存储器中,并由处理器执行。处理器可以包括业务引擎,该业务引擎具有预编程的规则和指令以作用于获取的数据。业务引擎可以与业务存储器通信,该业务存储器存储根据本文描述的方法在数据分析中使用的样本简介。在一些方面,一些或所有模块可以以硬件(例如,专用集成电路(asic)、现场可编程门阵列(fpga)、可编程逻辑设备(pld)、控制器、状态机、门控逻辑、分立硬件组件或任何其他合适的设备)、固件、软件和/或其组合来实现。在本公开中进一步描述了根据本主题技术的各个方面的这些模块的附加特征和功能。根据一些实施例,例如如图2a中所示,可以执行用于验证和准备所获取的数据的方法。如图2a所示,加载输入数据200并将其提供给处理器,诸如图1中所示的处理器。识别数据的类型201,并确定该数据是否为有效类型202。如果该数据是有效类型,则在203处理该数据并将其存储204,并且将所获取的数据的验证和准备确定为成功205。但是,如果该数据未被确定为有效类型,则显示错误206,并且在207中将验证和准备确定为失败。当验证成功时,可以转换和/或存储数据,或者在验证失败时,将错误显示给用户以拒绝数据。根据一些实施例,例如如图2b中所示,可以执行用于分析获取的数据的方法。验证数据类型是否具有相对于阈值的足够的信噪比。基于验证,可以在分析之前对数据进行分析或平滑过滤工艺。根据一些实施例,例如如图2b中所示,可以在包括应用基线相关、执行正态分布分析、确定视场的强度、计算集合体大小、选择目标区域、计算平均值、计算协方差、计算相关性和计算共分布的操作中分析数据。数据操作可以包括对目标区域(roi)的自动识别,以区分颗粒和溶液。可以确定颗粒的大小和数量以确定颗粒的总体分布。可以执行数据操作以确保合规性,诸如s/n比确定,基线校正,确定水蒸气含量以及确定所研究光谱区域内目标元素的信号强度。可以使用,尤其是,“实验设计”方法来简化用于统计分析的数据输出。目标元素的强度和光谱位置可以作为逗号分隔文件(*.csv)输出。可以基于目标样本的扰动来生成协方差或动态光谱数据集,该样本的输出可以用于进一步分析。例如,可以以促进与其他生物分析结果合并以进行可比性评估的格式提供数据输出,并通过以下方式提供:扰动类型、赋形剂、蛋白质治疗剂、蛋白质浓度、温度、采集日期和/或生物分析技术。这种方法将允许对在相似条件下进行的所有实验进行统计分析。更重要的是,doe分析的结果将是一个独立的文档,准备进行最终报告并可以做出决策。根据一些实施例,如上所述,本文所述的方法和系统可以将相关函数应用于协方差或动态光谱数据以生成同步图和异步图。光谱数据中彼此同相的变化(例如,峰强度)可以如在同步图中获得的那样相关。可以确定光谱数据中发生变化的元素。可以确定光谱数据中总体上最大的强度变化。可以确定光谱数据中总体上最小的强度变化。可以确定宽谱带(例如蛋白质和肽的酰胺谱带)中基本光谱贡献的最小数量,以进行曲线拟合分析,从而可以确定二级结构组成。可以提高正在研究的光谱区域的分辨率,特别是对于光谱中的宽谱带。此外,通过分析同步图和异步图,可以确定事件的顺序。光谱数据中彼此异相的变化(例如,峰强度)可以如在异步图中获得的那样相关。从异步图中,可以获得描述分子细节蛋白质行为的事件顺序。可以对图进行详细评估,以确定事件的顺序。替代地或组合地,该方法可以是自动化的。可以将联合方差函数应用于协方差或动态光谱数据,以生成合并的异步图,该图可以直接解释以确定事件的顺序。可替代地,该方法可以用于验证上述对于蛋白质分子行为的描述的解释,其是一个复杂的描述。由于热应力而导致的脱酰胺的证据可以通过分析异步图中的峰的异相相关性来获得。当在异步图中存在显示出异相强度变化的峰的相关性时,可以确定发生了脱酰胺。可以将机器学习方法作为对需要关联和解决的属性的复杂性的长期解决方案。实例研究使用三个nistmab(分别为标准品和候选品rm8671和rm8670)在不同浓度范围内进行评估,进行了用于监测和确定蛋白质中脱酰胺作用的系统和方法的可开发性和可比性评估:(低)1.0-1.5μg/μl,(中)2.0-2.4μg/μl,(中-高)2.8-10μg/μl。nistmab(批号14hb-d-002)是分子量为150kda的igg1κ亚型,是由两个重链和两个轻链亚基组成的同型二聚体,含有链间和链内二硫键。另外,该蛋白质具有翻译后修饰(ptm),其涉及位于fc区的n300处的n-链糖基化位点。评估中使用的特定nistmab蛋白质样品为:pdsnistmab(rm8671)、nistmabrm8671的样品,在2017年的玛丽亚飓风期间,当电力和其他基础设施遭到破坏时,其存储在波多黎各的蛋白质动态解决方案中,并且因此承受了极大的热应力;nistmab(rm8671)、nist提供的样品mabrm8671,未暴露于热应力下;nistmab候选品(8670),一种由nist提供的治疗性候选抗体。所研究的蛋白质样品在λmax=280nm处具有理论摩尔消光系数(ε),确定为212,270m-1cm-1。使用12.5mml-组氨酸缓冲液在ph6.00下对nistmab样品进行系列稀释。还对样品进行了浓度测定。稀释的nistmab样品与适当的参考样品12.5mml-组氨酸缓冲液(ph6.00)一起用于通过uv光谱测定浓度。稀释的nistmab(rm8671和8670)样品的紫外光谱是使用jasco(tokyo,jp)型号v-630分光光度计和starna(essex,uk)型号为dmv-bio的可拆卸石英池(光程为0.2mm)在室温(24℃)下获得的。以400nm/min的扫描速率和1.0nm的数据间距在235-320nm的光谱区域内共添加两次扫描。对所有收集的光谱在320nm处进行单点基线校正。microcal的origin7专业软件用于提供所需的图和分析。对于实验设计,将预定量如1μl的每个样品以及相应的参考样品应用于定制设计的caf2载玻片池上的预定义孔中。提供了坐标以用于自动图像采集,同时保持并对载玻片池进行热控制。注意在每个温度下收集背景,以消除量子级联激光器带来的潜在相干效应。使用实时高光谱成像量子级联激光透射显微镜(qcltm)在定制加热载玻片固定器和载玻片池的严格热控制下,对溶液中蛋白质样品阵列进行自动图像采集。已知阵列中每个样品的路径长度允许进行定量分析,例如在pct/us2017/014338中描述的分析,该文献以引用方式并入本文。使用qcltm捕获每种样品蛋白质的hsi原始光谱数据,并hsi数据被评估为存在颗粒/聚集体。此外,对于采样的每种蛋白质溶液,确定平均光谱数据并校正基线,然后生成后续的2dir相关图和共分布图,以进一步评估脱酰胺作用。在检查为样品蛋白质获得的hsi时,没有观察到或观察到少于5个颗粒。观察到的差异是由于脱酰胺影响mab稳定性的程度。例如,评估确定了由于在波多黎各的玛丽亚飓风期间经受了长期高温的nistmab候选品和pdsnistmab内的热应力,定位于低浓度(1.0-1.5μg/μl)的fc结构域的天冬酰胺n318脱酰胺事件。同样,在较高的mab浓度下,nistmab标准品(rm8671)和候选品(rm8670)的胶体稳定性会发生变化,这也可能是脱酰胺的迹象,但需要进一步评估。最后,观察到nistmab标准品比nistmab候选品具有更高的稳定性。使用为阵列中采样的每种蛋白质溶液采集的his,来可视化聚集体。在研究中使用的数据集中,<5或观察不到聚集体。此外,ph6.0的12.5mml-组氨酸缓冲液也没有聚集。根据光学设置,检测到的所有聚集体均在4.3μm-2.0mm尺寸范围内。提供增强的信噪比(snr)的三个量子级联激光器允许使用线性响应微辐射热计焦平面阵列(480×480像素)检测器。对于空间分辨率,使用在2×2mm2的视场(fov)内具有0.3na的数值孔径(na)的低倍率物镜(4×),其提供4.25×4.25μm空间分辨率的像素。对于阵列中的每种蛋白质样品,在1780-1450cm-1的光谱区域内以4cm-1的分辨率收集qclir光谱。为了防止由于qcl波动引起的相干效应,一旦达到热平衡(4分钟),就在每个设定温度下收集背景。阵列中每个样品的典型hsi采集时间为0.4分钟。原始光谱数据被保存为逗号分隔文件(*.csv)。只要需要,就可以从原始数据生成分析和图。根据原始数据,生成了qclir叠加和2dir相关图。在评估研究中用于进行分析的脱酰胺评估模块包括一个光标功能,可实现无偏的交叉峰强度变化和位置,这对于确定分子事件的顺序是有利的。图4a-4c示出了阵列中每个蛋白质样品在1780-1450cm-1的midir光谱区域和28-56℃的温度范围内以4℃的温度间隔采集的高光谱图像。图4a示出了在28℃和56℃下获得的1μg/μl的pdsnistmabrm8671的高光谱图像。图4b示出了在28℃和56℃下获得的2μg/μl的nistmabrm8671的高光谱图像。图4c示出了在28℃和56℃下获得的2.4μg/μl的nistmab候选品rm8670的高光谱图像。在4分钟的平衡期后达到设定温度时,进行hsi和背景采集。每个hsi由223,000个qcl红外光谱组成。在其定义的温度下的每个平均光谱表示在2×2mm2的fov内的223,000个光谱的平均值。对于阵列中的每个样品,fov与孔的直径匹配。然后为阵列中的每个nistmab生成qclir叠加,并且仅进行基线校正。图5示出了在28℃至56℃的温度范围内,以4℃的温度间隔下(28℃、32℃、36℃、40℃、44℃、48℃、52℃、56℃)在1780至1450cm-1的光谱区域中具有叠加的l-组氨酸和h2o吸收的酰胺i和ii谱带的qclir光谱叠加。以两种不同的浓度研究了pdsnistmab标准品(rm8671)、nistmab标准品(rm8671)和nistmab候选品(rm8670)。图5的顶行表示浓度为1-1.5μg/μl的qclir光谱叠加,底行显示浓度为2-2.5μg/μl的qclir光谱叠加。通常,该范围内的低浓度是pdsnist和nistmab标准品(rm8671),该范围内的高浓度是nistmab候选品(rm8670)。每个蛋白质样品具有28℃的低温平均光谱。然后对于每个蛋白质样品,使用28℃下的低温平均光谱生成差异光谱。结果,可以分析目标光谱区域内强度和峰位移的变化,并因此表示蛋白质在溶液中由于热应力的行为。表1总结了评估中使用的主干振动模式和位置:表1、h2o中二级结构谱带分配表2提供了评估中使用的侧链模式和位置的总结:表2、h2o中氨基酸侧链分配项目侧链编码振动模式位置(cm-1)评述1tyryv(c=c)1518周边环境2lyskδs(nh3+)1526ph,h-键,盐桥相互作用,柔性3gluev(coo–)1543-1560ph,h-键,脱酰胺,盐桥,阳离子键合,柔性4aspdv(coo–)1570-1574ph,h-键,脱酰胺,盐桥,阳离子键合,柔性5hishv(c=c)1596-1603ph,h-键,zn2+协调6c-末端v(coo–)1598c-末端的稳定性7ginqδ(nh2)1586-1607h-键,脱酰胺,柔性8asnnδ(nh2)1612-1618h-键,脱酰胺,柔性9lyskδas(nh3+)1625-1629ph,h-键,盐桥相互作用,柔性10argrvs(cn3h5+)1633ph,h-键,盐桥相互作用,柔性11ginqv(c=c)1670h-键,柔性12argrvas(cn3h5+)1673h-键,盐桥相互作用,柔性13asnnv(c=c)1678h-键,柔性14p-arf,y1740-1730疏水的15p-arf,y1720-1715相互作用16p-arf,y1708-1700π-π堆叠用于可比性评估的谱带位置代表用于nistmab标准品rm8671和nistmab候选品rm8670而研究的整个数据集确定的所有2dir相关峰的平均值。对于1700-1600cm-1范围内的酰胺i谱带,主要是由于c=o拉伸,c-n拉伸贡献较小,较小程度的n-h变形模式;对构象变化敏感。通常,对于所有三个mab,qclir光谱均包括:β-转角(1693.1cm-1)、铰链环(1665.2cm-1)、α-螺旋(1665.2cm-1)和β-折叠(1635.6cm-1)。这些二级结构通常在iggs中观察到。对于侧链模式,在酰胺i谱带内叠加有一些振动模式,而在酰胺ii谱带(1600-1500cm-1)内有其他振动模式。以下侧链模式位于酰胺i谱带的正前方:代表苯丙氨酸和酪氨酸侧链的三个对位取代的芳族峰(1748.7、1726.7和1705.0cm-1)、谷氨酰胺v(c=o)(1670.0cm-1)、天冬酰胺v(c=o)(1678.3cm-1)和δ(nh2)(1612.7cm-1),和赖氨酸δas(nh3+)(1621.0cm-1)。位于酰胺ii谱带内的侧链模式为:谷氨酰胺δ(nh2)(1591.0cm-1)、组氨酸v(c=c)(1600.1cm-1)、天冬氨酸盐v(coo–)(1572.0cm-1)和两种不同的谷氨酸盐,其中一种可能涉及到盐桥相互作用v(coo–)(1540.7和1559.0cm-1),赖氨酸δs(nh3+)(1525.0cm-1),以及最后的在(1519.0cm-1)的酪氨酸。也可以考虑精氨酸和色氨酸的振动模式。在从所有后续平均光谱中减去低温平均光谱后,减去h2o的贡献以及不受热应力扰动的所有蛋白质振动模式,从而仅评估热应力作用下在目标光谱区域中(1780-1450cm-1)发生的变化。通过执行2dir相关性分析,可以获得溶液中蛋白质的详细分子评估。应用相关函数来生成两个不同的图:(1)同步图,其提供了目标光谱区域内的整体强度变化;以及(2)异步图,其提供了增强的分辨率和根据热应力发生的分子事件的序列顺序。此外,该异步图提供了显示出异相强度变化的峰的详细相关性,这表明脱酰胺作用。例如,如下所述,该评估观察到由于脱酰胺作用,在与δ(nh2)振动模式相关的1612.7cm-1处,天冬酰胺的强度降低,而在1572.0cm-1v(coo–)振动模式下,天冬氨酸的强度增加。还评估了所研究蛋白质的总体热稳定性。使用热依赖性图,通过评估热转变温度的开始,来确定总体热稳定性。观察到在1780-1450cm-1光谱范围内的qclir峰位置最大值是温度在28-56℃范围内的函数:i)浓度为1-1.5μg/μl、2-2.4μg/μl和2.8或10μg/μl的nisstmabrm8671和rm8670;ii)浓度为1-1.5μg/μl、2-2.4μg/μl和2.8或10μg/μl的nistmab86781和rm8670,以及参考样品;iii)去离子h2o和ph为6.0的12.5mml-组氨酸缓冲液的参考样品。低浓度的pdsnistmab标准品(rm8671)、低浓度的nistmab标准品(rm8671)、nistmab候选品(rm8670)在1-2.8μg/μl之间的范围内,在50℃时表现出相同的峰移发生。但是,10μg/μl的nist候选品(rm8670)表现出较低的稳定性,起始峰位移发生在32.5℃。在去离子h2o的情况下,在整个温度范围内未观察到峰最大值的变化,而对于12.5mml-组氨酸缓冲液,在52.5℃观察到热转变的开始,这表明它比nistmab更稳定。这表明,针对nistmab观察到的变化是由于热应力引起的其固有行为。在评估中还监控了聚集事件,但是,在所检查的条件下,对于三个nistmab(rm8671和8670)样品中的任何一个,在热应力期间均未观察到聚集现象。如下面更详细地描述的,评估还监视光谱数据以检测和分析脱酰胺事件。评估集中分析了与在1678.3cm-1时酰胺内的天冬酰胺侧链羰基拉伸模式v(c=o)和在1612.7cm-1时酰胺弯曲模式δ(nh2)以及在1572.0cm-1时天冬氨酸羧酸盐的拉伸模式v(coo–)相关的交叉峰。确定这些峰之间已证实的相关性,以建立样品(nistmab,rm8671和nistmab候选品8670)的脱酰胺作用。通过确定阵列中每个样品在热应力(28-56℃)期间分子事件的顺序来提供对溶液中蛋白质行为的描述。实验方法不是确定三个nistmab标准品(rm8671和8670)蛋白质样品的总体热转变温度,而是确定所检查的三种蛋白质的稳定性差异以及是否观察到了脱酰胺作用。如果需要,可以使用完善的程序确定热转变温度。在本研究中,数据是根据蛋白质的低(1.0-1.5μg/μl)、中(2.0-2.4μg/μl)和中至高浓度(2.8-10.0μg/μl)分离的,以建立并了解敏感性差异,这是由于对于这样的离散事件,可以导致如上所述的蛋白质稳定性的改变并可能降低功效的浓度引起的。其次,该研究绘制了热应力下溶液中蛋白质发生脱酰胺作用的区域。最后,该研究基于脱酰胺的程度确定了稳定性,并根据分子事件的顺序确定了热稳定性。实施例1使用本文所述的方法和系统对28-56℃的温度范围内的热应力下的ph6.0的12.5mml-组氨酸中的1μg/μl的pdsnistmab、1μg/μl的nistmab和1.5μg/μl的nistmab候选品在1780-1450cm-1的光谱区域内进行了比较2dir相关光谱分析。对于三种研究的样品蛋白中的每一种,产生了在对应于28-56℃温度范围的1780-1450cm-1光谱区域内的酰胺i和酰胺ii谱带的qclir光谱叠加。从这些光谱叠加图中,使用2dir相关性来生成对应于28-56℃温度范围的同步图和异步图。图6a示出了在1780-1450cm-1的光谱区域内对应于pdsnistmab样品的28-56℃的温度范围的酰胺i和酰胺ii谱带的qcl光谱叠加。图6b示出了同步图,和图6c示出了基于图6a所示的qcl光谱叠加数据生成的异步图。图7a示出了在1780-1450cm-1的光谱区域内对应于nistmab样品的28-56℃的温度范围的酰胺i和酰胺ii谱带的qcl光谱叠加。图7b示出了同步图,和图7c示出了基于图7a所示的qcl光谱叠加数据生成的异步图。图8a示出了对应于nistmab候选样品的28-56℃的温度范围的在1780-1450cm-1的光谱区域内的酰胺i和酰胺ii谱带的qcl光谱叠加。图8b示出了同步图,和图8c示出了基于图8a所示的qcl光谱叠加数据生成的异步图。从2dir相关性图的分析解释中得出了在热应力下低浓度范围(1.0-1.5μg/μl)的三个mab样品的行为。如图9a所示,pdsnistmab的事件的顺序是从使用此处所述的noda规则对图6b、6c中所示的同步和异步图的分析中得出的。如图9a所示,对于pdsnistmab(rm8671),分子事件的顺序如下:酪氨酸残基(1519.0cm-1),随后是赖氨酸δs(nh3+)(1525.0cm-1),然后是在1540.7和1559.0cm-1处的两种类型的谷氨酸盐v(coo–),然后是在1580.0cm-1和1572.0cm-1处的两种类型的天冬氨酸盐v(coo–),大概参与了与附近酪氨酸和赖氨酸的氢键或盐桥相互作用,β-折叠(1635.6cm-1),然后是螺旋区域(1653.8cm-1)(在所有低浓度的mab中都观察到),然后是赖氨酸δas(nh3+)(1621.0cm-1),接着是天冬酰胺侧链模式δ(nh2)(1612.7cm-1)和谷氨酰胺δ(nh2)(1591.0cm-1),然后是组氨酸v(c=c)(1600.1cm-1),大概是所有这些残基彼此最接近,然后是铰链环(1665cm-1),然后是谷氨酰胺v(c=o)(1670.0cm-1)和天冬酰胺侧链模式v(c=o)(1678.3cm-1),然后是苯丙氨酸和酪氨酸对位取代的芳环模式(1726.7、1748.7、1705.0cm-1),由于在56℃附近部分展开,表明mab水溶剂可及性发生了变化,最终是β-转角(1693.1cm-1)受到扰动。这些最终分子事件在所有mab之间共享。图9b示出了nistmab的事件的顺序,该顺序是从图7b、7c所示的同步和异步图得出的。如图9b所示,nistmab(rm8671)的事件顺序如下:酪氨酸残基(1519.0cm-1),接着是赖氨酸δs(nh3+)(1525.0cm-1),然后是在1540.7cm-1处的谷氨酸盐v(coo–),接着是在1580.0cm-1处的天冬氨酸盐v(coo–),接着是β-折叠(1635.6cm-1)和螺旋区域(1653.8cm-1),然后是赖氨酸δas(nh3+)(1621.0cm-1),然后是天冬酰胺侧链模式δ(nh2)(1612.7cm-1)和谷氨酰胺δ(nh2)(1591.0cm-1),接着是组氨酸v(c=c)(1600.1cm-1),推测所有这些残基最接近彼此,然后是铰链环(1665cm-1),然后是谷氨酰胺v(c=o)(1670.0cm-1),然后在1559.0cm-1处的谷氨酸盐v(coo–),在1572.0cm-1处的天冬氨酸盐v(coo–)和天冬酰胺侧链模式v(c=o)(1678.3cm-1),然后是苯丙氨酸和酪氨酸对位取代的芳环模式(1726.7、1748.7、1705.0cm-1),由于在56℃附近部分展开,表明mab水溶剂可及性发生了变化,最终是β-转角(1693.1cm-1)受到扰动。对于未经受与飓风玛丽亚相关的应力的nistmab,其具有在1559.0cm-1处的谷氨酸盐v(coo–)和在1572.0cm-1处的天冬氨酸盐v(coo–)的两个稳定的振动模式。这些是与脱酰胺相关的模式。图9c示出了nistmab候选品(rm8670)的事件的顺序,该顺序是从图8b、8c中所示的同步和异步图得出的。如图9c所示,nistmab候选品(rm8670)的事件顺序如下:最不稳定的是酪氨酸残基(1519.0cm-1),然后是赖氨酸δs(nh3+)(1525.0cm-1),然后在1540.7cm-1处的谷氨酸盐v(coo–),然后在1580.0cm-1处的天冬氨酸盐v(coo–),然后是谷氨酰胺δ(nh2)(1591.0cm-1),然后是赖氨酸δas(nh3+)(1621.0cm-1),然后是天冬酰胺侧链模式δ(nh2)(1612.7cm-1),然后是组氨酸v(c=c)(1600.1cm-1),然后二级结构在β-折叠(1635.6cm-1)、螺旋区域(1653.8cm-1)和铰链环(1665cm-1)内受到扰动,然后发生脱酰胺事件谷氨酰胺v(c=o)(1670.0cm-1),然后是1559.0cm-1处的谷氨酸盐v(coo–),以及1572.0cm-1处的天冬氨酸盐v(coo–)和天冬酰胺侧链模式v(c=o)(1678.3cm-1),然后是苯丙氨酸和酪氨酸对位取代的芳族环模式(1726.7、1748.7、1705.0cm-1),由于在56℃附近部分展开,表明mab水溶剂可及性发生了变化,最终是β-转角(1693.1cm-1)受到扰动。分子事件的顺序的差异是由于这些nistmab在其热应力之前的稳定性所致。由于在低浓度下所有三个mab在热应力的初始阶段和最终阶段都观察到重复事件,因此置信度很高。进一步分析了异步图中似乎在不同时间不稳定的交叉峰,因为它们会与脱酰胺过程相关,从而导致mab的结构域稳定性发生变化。实施例2使用本文所述的方法和系统对28-56℃的温度范围内的热应力下的ph6.0的12.5mml-组氨酸中的2μg/μl的pdsnistmab(rm8671)、2μg/μl的nistmab(rm8671)和2.4μg/μl的nistmab候选品(rm8670)在1780-1450cm-1的光谱区域内进行了比较2dir相关光谱分析。对于三种研究的样品蛋白中的每一种,产生了在对应于28-56℃温度范围的1780-1450cm-1光谱区域内的酰胺i和酰胺ii谱带的qclir光谱叠加。从这些光谱叠加图中,使用2dir相关性来生成对应于28-56℃温度范围的同步图和异步图。图10a示出了在1780-1450cm-1的光谱区域内对应于pdsnistmab样品的28-56℃的温度范围的酰胺i和酰胺ii谱带的qcl光谱叠加。图10b示出了同步图,和图10c示出了基于图10a所示的qcl光谱叠加数据生成的异步图。图11a示出了在1780-1450cm-1的光谱区域内对应于nistmab样品的28-56℃的温度范围的酰胺i和酰胺ii谱带的qcl光谱叠加。图11b示出了同步图,和图11c示出了基于图11a所示的qcl光谱叠加数据生成的异步图。图12a示出了对应于nistmab候选样品的28-56℃的温度范围的在1780-1450cm-1的光谱区域内的酰胺i和酰胺ii谱带的qcl光谱叠加。图12b示出了同步图,和图12c示出了基于图12a所示的qcl光谱叠加数据生成的异步图。从2dir相关性图的分析解释中得出了在热应力下中间浓度范围(2.0-2.4μg/μl)的三个mab样品的行为。如图13a所示,pdsnistmab的事件的顺序是从使用此处所述的noda规则对图10b、10c中所示的同步和异步图的分析中得出的。如图13a所示,对于pdsnistmab(rm8671),分子事件的顺序如下:酪氨酸残基(1519.0cm-1),随后是赖氨酸δs(nh3+)(1525.0cm-1),然后是在1540.7cm-1处的谷氨酸盐v(coo–),然后是在1580.0cm-1处的天冬氨酸盐v(coo–),接着是β-折叠(1635.6cm-1),然后是赖氨酸δas(nh3+)(1621.0cm-1),接着是螺旋区域(1653.8cm-1),在1559.0cm-1处的谷氨酸盐v(coo–),和在1572.0cm-1处的天冬氨酸盐v(coo–),以及天冬酰胺侧链模式δ(nh2)(1612.7cm-1)和谷氨酰胺δ(nh2)(1591.0cm-1),表明脱酰胺过程随后是组氨酸v(c=c)(1600.1cm-1),然后是铰链环(1665cm-1)受到扰动,接着是谷氨酰胺v(c=o)(1670.0cm-1),然后是天冬酰胺侧链模式v(c=o)(1678.3cm-1),然后是苯丙氨酸和酪氨酸p-取代的芳环模式(1726.7、1748.7、1705.0cm-1),最后是β-转角(1693.1cm-1)受到扰动。图13b示出了nistmab的事件的顺序,该顺序是从图11b、11c所示的同步和异步图得出的。如图13b所示,nistmab(rm8671)的事件顺序如下:酪氨酸残基(1519.0cm-1),接着是赖氨酸δs(nh3+)(1525.0cm-1),然后是在1540.7cm-1处的谷氨酸盐v(coo–),接着是在1580.0cm-1处的天冬氨酸盐v(coo–),接着是β-折叠(1635.6cm-1),然后是螺旋区域(1653.8cm-1),然后是赖氨酸δas(nh3+)(1621.0cm-1),然后是铰链环(1665cm-1),接着是天冬酰胺侧链模式δ(nh2)(1612.7cm-1),然后是谷氨酰胺v(c=o)(1670.0cm-1),谷氨酰胺δ(nh2)(1591.0cm-1),然后是组氨酸v(c=c)(1600.1cm-1),然后是在1559.0cm-1处的谷氨酸盐v(coo–)和在1572.0cm-1处的天冬氨酸盐v(coo–),接着是天冬酰胺侧链模式v(c=o)(1678.3cm-1),然后是苯丙氨酸和酪氨酸对位取代的芳环模式(1726.7、1748.7、1705.0cm-1),最终是β-转角(1693.1cm-1)受到扰动。图13c示出了nistmab候选品(rm8670)的事件的顺序,该顺序是从图12b、12c中所示的同步和异步图得出的。如图13c所示,nistmab候选品(rm8670)的事件顺序如下:最不稳定的是酪氨酸残基(1519.0cm-1),然后是赖氨酸δs(nh3+)(1525.0cm-1),然后是在1540.7cm-1处的谷氨酸盐v(coo–),然后是β-折叠(1635.6cm-1),然后是螺旋区域(1653.8cm-1),然后在1580.0cm-1处的天冬氨酸盐v(coo–),赖氨酸δas(nh3+)(1621.0cm-1),然后是天冬酰胺侧链模式δ(nh2)(1612.7cm-1),然后是铰链环(1665cm-1),然后是组氨酸v(c=c)(1600.1cm-1),然后是谷氨酰胺δ(nh2)(1591.0cm-1),谷氨酰胺v(c=o)(1670.0cm-1),然后是1559.0cm-1处的谷氨酸盐v(coo–),表明脱酰胺;接着是在1572.0cm-1处的天冬氨酸盐v(coo–),然后是天冬酰胺侧链模式v(c=o)(1678.3cm-1),然后是苯丙氨酸和酪氨酸对位取代的芳环模式(1726.7、1748.7、1705.0cm-1),最后是β-转角(1693.1cm-1)受到扰动。实施例3使用本文所述的方法和系统对28-56℃的温度范围内的热应力下的ph6.0的12.5mml-组氨酸中的2.8μg/μl的nistmab(rm8671)和10μg/μl的nistmab候选品(rm8670)在1780-1450cm-1的光谱区域内进行了比较2dir相关光谱分析。对于2.8μg/μl的nistmab(rm8671)和10μg/μl的nistmab候选品(rm8670),产生了在对应于28-56℃温度范围的1780-1450cm-1光谱区域内的酰胺i和酰胺ii谱带的qclir光谱叠加。从这些光谱叠加图中,使用2dir相关性来生成对应于28-56℃温度范围的同步图和异步图。图14a示出了在1780-1450cm-1的光谱区域内对应于nistmab样品的28-56℃的温度范围的酰胺i和酰胺ii谱带的qcl光谱叠加。图14b示出了同步图,和图14c示出了基于图14a所示的qcl光谱叠加数据生成的异步图。图15a示出了在1780-1450cm-1的光谱区域内对应于nistmab候选样品的28-56℃的温度范围的酰胺i和酰胺ii谱带的qcl光谱叠加。图15b示出了同步图,和图15c示出了基于图15a所示的qcl光谱叠加数据生成的异步图。同步图和异步图中显示的较高浓度可能反映以下一项或多项:(1)由于热应力期间浓度增加,蛋白质的胶体稳定性发生了变化;(2)表明低蛋白浓度下分子间相互作用的频率较低,和/或(3)当谷氨酰胺脱酰胺事件与更容易发生的天冬酰胺残基脱酰胺事件(与谷氨酰胺相比在动力学上有利)结合时,会降低nistmab的稳定性。从2dir相关性图的分析解释中得出了分别处于热应力下的中间浓度和高浓度的2.8和10.0μg/μl的nistmab标准品(rm8671)和nistmab候选品(rm8670)的分子事件的顺序。如图16a所示,pdsnistmab的事件的顺序是从使用此处所述的noda规则对图14b、14c中所示的同步和异步图的分析中得出的。如图16a所示,中间浓度(2.8μg/μl)的nistmab(rm8671)的事件的顺序如下:酪氨酸残基(1519.0cm-1)首先受到扰动,随后是赖氨酸(1525cm-1),然后是在1580.0cm-1处的天冬氨酸盐v(coo–),在1540.7和1559.0cm-1处的两种类型的谷氨酸盐v(coo–)和在1572.0cm-1处的天冬氨酸盐v(coo–),大概参与了氢键和盐桥相互作用;然后是β-折叠(1635.6cm-1),然后是螺旋区域(1653.8cm-1)受到扰动,然后是谷氨酰胺δ(nh2)(1591.0cm-1)和谷氨酰胺v(c=o)(1670.0cm-1),然后是铰链环(1665cm-1)受到扰动,接着是赖氨酸δas(nh3+)(1621.0cm-1),然后是天冬酰胺侧链模式δ(nh2)(1612.7cm-1),然后是组氨酸v(c=c)(1600.1cm-1),然后是天冬酰胺侧链模式v(c=o)(1678.3cm-1),然后是苯丙氨酸和酪氨酸对位取代的芳环模式(1726.7、1748.7、1705.0cm-1),最终是β-转角(1693.1cm-1)受到扰动。图16b示出了nistmab候选品(rm8670)的事件的顺序,该顺序是从图15b、15c中所示的同步和异步图得出的。如图13c所示,高(10μg/μl)浓度的nistmab候选品(rm8670)的事件顺序如下:酪氨酸残基(1519.0cm-1),然后是赖氨酸δs(nh3+)(1525.0cm-1),然后是在1540.7cm-1处和1559.0cm-1处的两种类型的谷氨酸盐v(coo–),然后是在1580.0cm-1处和1572.0cm-1处的两种类型的天冬氨酸盐v(coo–),大概参与了与附近酪氨酸和赖氨酸的氢键或盐桥相互作用,然后是二级结构被β-折叠(1635.6cm-1)扰动,接着是螺旋区域(1653.8cm-1)和铰链环(1665cm-1),然后是谷氨酰胺v(c=o)(1670.0cm-1),然后是赖氨酸δas(nh3+)(1621.0cm-1),然后是天冬酰胺侧链模式δ(nh2)(1612.7cm-1)和谷氨酰胺δ(nh2)(1591.0cm-1),表明这些是不会发生脱酰胺作用的稳定残基,然后是组氨酸v(c=c)(1600.1cm-1),接着是天冬酰胺侧链模式v(c=o)(1678.3cm-1),然后是苯丙氨酸和酪氨酸对位取代的芳族环模式(1726.7、1748.7、1705.0cm-1),最终是β-转角(1693.1cm-1)受到扰动。热应力引起的脱酰胺的测定对图6c、7c和8c中所示的pdsnistmab标准品(rm8671)、nistmab标准品(rm8671)和nistmab候选品(rm8670)的异步图的评估,证明了与蛋白质中脱酰胺作用一致的多元关系。因此,评估中评估的数据表明,使用qcl显微镜和本文公开的方法的hsi对确定由热应力引起的天冬酰胺和谷氨酰胺的脱酰胺具有选择性和敏感性。图17a、17b和17c是对应于图6a、6b和6c所示的异步图的pdsnistmab标准品(rm8671)、nistmab标准品(rm8671)和nistmab候选品(rm8670)的异步图,但带有附加标记以表明观察到的脱酰胺作用的证据。如图17a、17b和17c所示,低浓度范围(1-1.5μg/μl)下的脱酰胺作用是热应力的函数,通过在异步图中用白色圆圈突出显示的异相相关性可以明显看出。在图17a、17b和17c的每个图中白色圆圈表示为(a)并且也可以由左箭头表示,表示天冬氨酸盐v(coo–)在1572.0cm-1处的强度增加。标记为(b)的白色圆圈代表1612.7cm-1处的天冬酰胺δ(nh2)。标记为(c)并且也由上箭头指示的白色圆圈代表在1678cm-1处酰胺侧链模式的天冬酰胺v(c=o)羰基伸缩。对于pdsnistmab标准品(rm8671)和nistmab候选品(rm8670),与nistmah标准品(rm8671)相比,在异步等高线图中观察到的天冬酰胺交叉峰不太明显,在该等高线图中,由于应力源条件,脱酰胺作用已成为一个重要事件。在图17a、17b和17c中确定了关键交叉峰的强度变化,并且这些强度变化被用于脱酰胺分析中。图18a、图18b和图18c提供了分别在图17a、17b和17c中标记为(a)、(b)和(c)的位置处的强度变化的条形图。这些条形图进一步显示了β-折叠/螺旋二级结构(α/β)的相对稳定性。特别地,监测了与脱酰胺事件相关的位于β-折叠中的交叉峰,其中:(a)是表明通过v(coo–)形成天冬氨酸的峰,(b)是表明天冬酰胺通过δ(nh2)损失的峰,(c)是代表在脱酰胺过程中转化为天冬氨酸期间天冬酰胺侧链v(c=o)扰动的峰。在评估中还发现了在热应力期间低浓度的nistmab候选品的谷氨酰胺脱酰胺的证据,这表明谷氨酰胺残基的评估也可用于鉴定和绘制脱酰胺事件。图19示出了热应力期间的低浓度的nistmab候选品在1780-1485cm-1的光谱区域内的异步图。监测图中的关键交叉峰,并确定了β-折叠中与脱酰胺相关的三个关键峰:(a)代表通过v(coo–)形成谷氨酸的峰,(b)代表谷氨酰胺通过δ(nh2)损失的峰,和(c)代表在转化为谷氨酸的过程中谷氨酰胺侧链v(c=o)的扰动的峰。图20是一个柱状图,总结了图19中识别出的关键交叉峰的强度变化率。分析关键交叉峰的强度变化可确认脱酰胺事件。结果证实了nistmab候选品(rm8670)中的谷氨酰胺脱酰胺作用。该过程极大地导致了mab的不稳定。图21是天冬酰胺的脱酰胺作用机理以及用于监测热应力期间事件的关键振动模式的示意图。qclir提供了脱酰胺过程中分子事件的高度选择性和灵敏检测。这些振动模式成为完整蛋白在溶液中应力过程的内部探针。还监测了与脱酰胺事件(天冬酰胺/谷氨酰胺)有关的强度变化和二级结构的相对稳定性。受脱酰胺事件影响的骨干v(c=o)包括:(a)通过v(coo–)形成天冬氨酸,(b)天冬酰胺通过δ(nh2)的损失,以及(c)在生成琥珀酰亚胺中间体并转化为天冬氨酸的过程中,天冬酰胺侧链v(c=o)的扰动。脱酰胺被认为是翻译后修饰,其可以影响治疗性蛋白质的稳定性、结构和功效,并且可以引起聚集,这可以导致不想要的免疫应答。表现出脱酰胺作用的残基是天冬酰胺和较低含量的谷氨酰胺。当天冬酰胺的邻近残基(位置n+1)是甘氨酸时,容易发生天冬酰胺翻译后修饰,从而降低了琥珀酰亚胺中间体形成的空间位阻,产生天冬氨酸或异天冬氨酸。脱酰胺事件发生在不存在任何酶的情况下,并且在高ph和/或高温下会加速脱酰胺。脱酰胺可能表明细胞内蛋白质的降解,从而降低了细胞内治疗性蛋白质的半衰期,从而潜在地影响pk/pd。使用本文所述的系统和方法可以评估脱酰胺作为翻译后修饰在蛋白质溶液中是否普遍存在,以及是否影响溶液中蛋白质的稳定性。为此,将分析重点放在最可能发生脱酰胺的位点上是有益的。对本文所述的nistmab标准品igg1κ一级序列的检查评估表明,在整个蛋白质序列中只有两个天冬酰胺残基满足上文提到的n+1标准:1)n369g370位于也暴露于水性环境的β-转角内,以及2)n318g319位于n300糖基化位点下游的310螺旋内。为了辨别是否存在脱酰胺作用,在蛋白质中鉴定出关键质量属性(cqa)的可能性在于上述鉴定中最容易获得的位点。可能存在其他可能发生脱酰胺作用的天冬酰胺残基,例如n+1,其中相邻残基为丙氨酸(a),也可以使用这些残基。此外,此外,由于脱酰胺事件的负电荷增加,与n369相邻的谷氨酸盐的数量可能会不稳定。这将导致n369所在的β-折叠或铰链环不稳定,即fc结构域。另一个候选品是位于fc结构域中310螺旋的n318,也具有相邻的谷氨酸、天冬氨酸、组氨酸和酪氨酸残基,这可解释在低浓度下观察到的扰动水平。具有最小水平空间位阻的谷氨酰胺残基的脱酰胺分布在重链和轻链中。更重要的是,它们位于可变fab区域内,甚至位于轻链内的cdr内。相反,可能容易脱酰胺的天冬酰胺残基被限制在fc结构域内。检查具有邻近甘氨酸残基(位置q+1)的nistmab标准品(rm8671)谷氨酰胺残基以鉴定周围的邻近残基使得能够作图并随后鉴定引起脱酰胺事件的qg。免疫原性风险生物测定法长期以来一直被用于解决治疗性蛋白质具有免疫原性的潜力。治疗性蛋白质是仅次于疫苗的第二大生物制药产品类别。迄今为止,生物制药行业已经通过使用结合抗体类型筛选(统称为生物测定法)解决了治疗性蛋白质诱导免疫原性或抗药物抗体(ada)反应的潜力。不幸的是,有时这些生物测定法导致产生假阳性或阴性结果。这一直是fda在2016年起草免疫原性指南的动机。总的来说,全世界的监管机构都要求实施正交分析工具来验证生物测定法以评估免疫原性和/或ada风险。蛋白质聚集是免疫原性和ada原位反应的共同因素。在不使用探针的情况下并基于从用于实施本文所述的系统和方法的平台技术获得的第一原理数据直接测量聚集。该平台技术包括一个专用的液体处理系统,一个实时量子级联激光显微镜,该显微镜具有经过改进的载物台,可提供更高的信噪比(snr),载玻片池,加热的载玻片保持架,plc控制器以及实施软件模块的计算机系统来分析数据并存储和传达结果。如图22所示,平台技术可以包括用于样品制备的液体处理系统2201和光谱成像采集系统2202,例如使用qcl显微镜的hsi成像系统,用于在应力期间监视溶液中的蛋白质阵列。可以提供与液体处理系统2201和光谱成像采集系统2202通信的数据管理系统,例如云存储系统。数据管理系统2203也可以与远程计算系统2204通信,从而允许数据的远程或离线分析。平台技术可以在基于阵列的方法中实施,以允许可再现地确定在生理温度范围(37-4l℃)下由人血清中的治疗性蛋白质诱导的聚集。基于阵列的方法只需要最少的样品,就可以在人类临床试验中首先确定结果,并根据性别、既往病情或当前医疗处方对不良事件进行预测。这为后续的临床试验设计提供了一种预测工具。此外,所获得结果的质量和统计稳健性适合大数据分析和机器学习。实施基于阵列的方法的平台技术的正交实施可以为由涉及治疗性蛋白质的生物制药进行的当前生物测定提供验证。设计良好的ada分析应基于试验性新药(ind)申请申报阶段的免疫原性测试范式的基本原理。获得阳性免疫原性结果时,需要进行ada分析。平台技术还提供了对现有免疫原性测定的验证分析方法。验证过程涉及敏感性、特异性、选择性和精密度要求的评估。聚合的评估是此过程的关键,这可以通过统计上可靠的高度选择性和敏感技术来确定。基于动物模型结果和临床试验的结果,人们质疑使用动物模型进行免疫原性筛选是否对人具有可移植性/适用性。提议用于免疫原性和ada风险评估的方法不会对患者或供体产生对治疗性蛋白产品产生免疫应答的风险,因为该分析是在样品血清上进行的,而不是在体内进行的。每一次三份分析测定仅需要100μl血清。该分析被设计为包含适当的阴性和阳性对照。表3总结了用于不同剂量水平的免疫原性可比性的一式三份的典型测定设置:表3注:由同一分析师生成的三份副本实时评估需要在采样之后,在存储样品之前尽快对样品进行分析。根据免疫原性fda草案指南,将人血清样品的等分试样用作阴性对照,另外的对照样品包括制剂,同时将一系列血清样品暴露于不同量的治疗性蛋白质产品。还可以以时间定义的方式执行分析,以评估样品血清中聚集的存在。该平台技术为直接确定聚集体提供了高度选择性和灵敏的方法,从而允许在生物仿制药和标准物质(来源)之间进行可比性评估。如果观察到聚集,则可以确定聚集程度,然后进行滴定度ada测定。已设计用于滴度测定ada测定的平台技术,以通过评估每个血清样品中的聚集程度来测量ada反应的幅度。聚集事件将被视为对患者存在安全隐患的潜力。如果在ada滴度测定期间持续存在,则血清或pbmc中的聚集事件也可能与功效降低有关。每个载玻片池将使用3个独立制备的相同样品来评估ada测定的准确性,方差系数小于10%。评估将涉及低、中和高范围,以验证测定结果。图23是示出根据本主题技术的一些方面的实验方法的示例性设计的操作的流程图。如图23所示,实验技术的设计被应用于获得的生物信息学和序列比较信息。然后在2302处对所得数据进行光谱分析,诸如通过使用关于图3描述的相关性分析技术。然后,可以对光谱分析的结果进行如本文所述的比较分析2303。图24是指示用于ada筛选和免疫原性风险评估的示例性方法的操作的流程图。图25是指示可以使用本文描述的平台技术和方法执行的示例性比较分析的操作的流程图。图26是示出示例性计算机系统的框图,利用该示例性计算机系统可以实现计算装置(例如,图4)。在某些实施例中,计算机系统1900可以使用硬件或者软件和硬件的组合来实现,或者在专用服务器中,或者集成到另一个实体中,或者分布在多个实体中。计算机系统1900包括用于传送信息的总线1908或其他通信机构,以及与总线1908耦合的用于处理信息的处理器1902。举例来说,计算机系统1900可以用一个或多个处理器1902来实现。处理器1902可以是通用微处理器、微控制器、数字信号处理器(dsp)、专用集成电路(asic)、现场可编程门阵列(fpga)、可编程逻辑设备(pld)、控制器、状态机、门控逻辑、离散硬件组件和/或可以执行信息的计算或其他操作的任何其他合适的实体。除了硬件之外,计算机系统1900还可以包括为所讨论的计算机程序创建执行环境的代码,例如,构成处理器固件、协议栈、数据库管理系统、操作系统或存储在包括的存储器1904中的它们的一个或多个的组合的代码、存储器1904诸如随机存取存储器(ram)、闪存、只读存储器(rom)、可编程只读存储器(prom)、可擦除prom(eprom)、寄存器、硬盘、可移动磁盘、cd-rom、dvd和/或任何其他合适的存储设备,其耦合到总线1908用于存储要由处理器1902执行的信息和指令。处理器1902和存储器1904可以补充或并入专用逻辑电路。指令可以存储在存储器1904中,并且可以在一个或多个计算机程序产品中实现,即,在计算机可读介质上编码的计算机程序指令的一个或多个模块,以由计算机系统1900执行或控制计算机系统1900的操作,并且根据本领域技术人员众所周知的任何方法,包括但不限于计算机语言,例如面向数据的语言(例如sql、dbase),系统语言(例如c、objective-c、c++、汇编)、体系结构语言(例如java、.net)和/或应用程序语言(例如php、ruby、perl、python)。指令也可以以计算机语言来实现,例如数组语言、面向方面的语言、汇编语言、创作语言、命令行界面语言、编译语言、并发语言、花括号语言、数据流语言、数据结构化语言、声明性语言、深奥语言、扩展语言、第四代语言、功能语言、交互模式语言、解释语言、迭代语言、基于列表的语言、小语言、基于逻辑的语言、机器语言、宏语言、元编程语言、多范式语言、数值分析、非英语语言、面向对象的基于类的语言、面向对象的基于原型的语言、越位规则语言、过程语言、反射语言、基于规则的语言、脚本语言、基于堆栈的语言、同步语言、语法处理语言、视觉语言、深度语言和/或基于xml的语言。存储器1904还可用于在执行将由处理器1902执行的指令期间存储临时变量或其他中间信息。如本文所讨论的计算机程序不一定对应于文件系统中的文件。程序可以存储在保存其他程序或数据的文件的一部分中(例如,存储在标记语言文档中的一个或多个脚本),专用于所讨论程序的单个文件或多个协调文件(例如,存储一个或多个模块,子程序或部分代码的文件)。可以将计算机程序部署为在一台计算机上执行,或者在位于一个站点上或分布在多个站点上并通过通信网络互连的多台计算机上执行。本说明书中描述的过程和逻辑流程可以由执行一个或多个计算机程序以通过对输入数据进行操作并生成输出来执行功能的一个或多个可编程处理器来执行。计算机系统1900还包括耦接到总线1908的用于存储信息和指令的数据存储设备1906,例如磁盘或光盘。计算机系统1900可以经由输入/输出模块1910耦合到各种设备(例如,设备1914和1916)。输入/输出模块1910可以是任何输入/输出模块。示例性输入/输出模块1910包括数据端口(例如,usb端口),音频端口和/或视频端口。在一些实施例中,输入/输出模块1910包括通信模块。示例性通信模块包括网络接口卡,例如以太网卡,调制解调器和路由器。在某些方面,输入/输出模块1910被配置为连接到多个设备,例如输入设备1914和/或输出设备1916。示例性输入设备1914包括键盘和/或指示设备(例如,鼠标或轨迹球),用户可以通过它向计算机系统1900提供输入。其他类型的输入设备1914也可以用于与用户进行交互,例如触觉输入设备,视觉输入设备,音频输入设备和/或脑机接口设备。例如,提供给用户的反馈可以是任何形式的感觉反馈(例如,视觉反馈、听觉反馈和/或触觉反馈),并且可以以任何形式接收来自用户的输入,包括声音、语音、触觉和/或脑电波输入。示例性输出设备1916包括用于向用户显示信息的显示设备,诸如阴极射线管(crt)或液晶显示器(lcd)监视器。根据某些实施例,响应于处理器1902执行存储器1904中包含的一个或多个指令的一个或多个序列,可以使用计算机系统1900来实现客户端设备和/或服务器。可以从诸如数据存储设备1906之类的另一机器可读介质将这样的指令读入存储器1904。存储器1904中包含的指令序列的执行使处理器1902执行本文所述的处理步骤。也可以采用多处理布置中的一个或多个处理器来执行包含在存储器1904中的指令序列。在一些实施例中,可以使用硬连线电路代替软件指令或与软件指令结合以实现本公开内容的各个方面。因此,本公开的方面不限于硬件电路和软件的任何特定组合。本说明书中描述的主题的各个方面可以在包括后端组件(例如,数据服务器),或包括中间件组件(例如,应用服务器),或包括前端组件(例如,具有图形用户界面和/或网页浏览器的客户端计算机,用户可以通过该图形用户界面和/或网页浏览器与本规范中描述的主题的实现进行交互),或一个或多个此类后端、中间件或前端组件的任意组合的计算系统。系统1900的组件可以通过数字数据通信的任何形式或介质(例如,通信网络)互连。通信网络的实例包括局域网和广域网。如本文所使用的术语“机器可读存储介质”或“计算机可读介质”是指参与向处理器1902提供指令以供执行的任何一种或多种介质。这样的介质可以采取许多形式,包括但不限于非易失性介质、易失性介质和传输介质。非易失性介质包括例如光盘或磁盘,诸如数据存储设备1906。易失性介质包括动态存储器,诸如存储器1904。传输介质包括同轴电缆、铜线和光纤,包括导线、导线包括总线1908。机器可读介质的常见形式包括,例如,软盘、软磁盘、硬盘、磁带、任何其他磁介质、cd-rom、dvd、任何其他光学介质、打孔卡、纸带、带孔图案的任何其他物理介质、ram、prom、eprom、flasheprom、任何其他存储芯片或盒式磁带或计算机可以从中读取的任何其他介质。机器可读存储介质可以是机器可读存储设备、机器可读存储基板、存储设备、影响机器可读传播信号的物质组成或它们中的一个或多个的组合。如本文所使用的,“处理器”可以包括一个或多个处理器,并且“模块”可以包括一个或多个模块。在本主题技术的一方面,机器可读介质是用指令编码或存储的计算机可读介质,并且是计算元件,其定义了指令与系统的其余部分之间的结构和功能关系,允许实现指令的功能。指令可以例如由系统或系统的处理器执行。指令可以是例如包括代码的计算机程序。机器可读介质可以包括一个或多个介质。如本文所使用的,单词“模块”是指体现在硬件或固件中的逻辑,或者指代以诸如c++的编程语言编写的,可能具有入口和出口点的软件指令的集合。可以将软件模块编译并链接到可执行程序中,将其安装在动态链接库中,或者可以用诸如basic之类的解释性语言编写该软件模块。应当理解,软件模块可以从其他模块或者从其自身被调用,和/或可以响应于检测到的事件或中断而被调用。软件指令可以嵌入在固件中,诸如eprom或eeprom。还将意识到,硬件模块可以包括连接的逻辑单元,诸如门和触发器,和/或可以包括可编程单元,诸如可编程门阵列或处理器。本文描述的模块优选地被实现为软件模块,但是可以以硬件或固件来表示。可以设想,模块可以被集成到更少数量的模块中。一个模块也可以分成多个模块。所描述的模块可以被实现为硬件,软件,固件或其任何组合。另外,所描述的模块可以位于通过有线或无线网络或互联网连接的不同位置。通常理解处理器可以包括例如计算机,程序逻辑或表示数据和指令的其他基板配置,其按本文所述进行操作。在其他实施例中,处理器可以包括控制器电路、处理器电路、处理器、通用单芯片或多芯片微处理器、数字信号处理器、嵌入式微处理器、微控制器等。此外理解,在一个实施例中,程序逻辑可以有利地被实现为一个或多个组件。组件可以有利地被配置为在一个或多个处理器上执行。这些组件包括但不限于软件或硬件组件,诸如软件模块的模块,面向对象的软件组件、类组件和任务组件、处理方法、功能、属性、过程、子例程、程序代码段、驱动程序、固件、微码、电路、数据、数据库、数据结构、表、数组和变量。提供前述描述以使本领域技术人员能够实践本文描述的各种配置。尽管已经参考各种附图和配置具体描述了本主题技术,但是应当理解,这些仅是出于说明的目的,而不应被认为是对本主题技术范围的限制。可能有许多其他方式来实现本主题技术。在不脱离本主题技术的范围的情况下,本文所描述的各种功能和元件可以与所示出的功能和元件不同地划分。对这些配置的各种修改对于本领域技术人员将是显而易见的,并且本文定义的一般原理可以应用于其他配置。因此,本领域普通技术人员可以对本主题技术进行多种改变和修改,而不脱离本主题技术的范围。应当理解,所公开的方法中步骤的特定顺序或层次是示例性方法的说明。基于设计偏好,应当理解,可以重新布置方法中步骤的特定顺序或层次。某些步骤可以同时执行。随附的方法权利要求以示例顺序呈现了各个步骤的要素,并不意味着限于所呈现的特定顺序或层次。如本文中所使用的,在一系列项目之前的短语“至少一个”,用术语“和”或“或”来分隔任何一个项目,整体上修改了该列表,而不是整个列表的成员(即每个项目)。短语“至少一个”不需要选择列出的每一项中的至少一项;相反,该短语允许包括任一个项目的至少一个和/或项目的任何组合的至少一个和/或每个项目中的至少一个的含义。举例来说,短语“a、b和c中的至少一个”或“a、b或c中的至少一个”分别仅指a、b或c;a、b和c的任意组合;和/或a、b和c中的至少一个。在本公开中使用的诸如“顶部”、“底部”、“前部”、“后部”等的术语应被理解为是指参照任意参照系,而不是普通引力参照系。因此,在重力参照系中,顶面、底面、前面和后面可以向上、向下、对角或水平地延伸。此外,在一定程度上,说明书或权利要求书中使用的术语“包括”、“具有”等旨在以类似于术语“包括”的方式包括在内,因为“包括”是在权利要求中用作过渡词时解释。词语“示例性”在本文中用来表示“用作示例,实例或说明”。本文中被描述为“示例性”的任何实施例不必被解释为比其他实施例优选或有利。除非特别指出,否则以单数形式提及元件并非旨在表示“一个且仅一个”,而是“一个或多个”。男性代词(例如,他的)包括女性和中性性别(例如,她及其代词),反之亦然。术语“一些”是指一个或多个。带下划线和/或斜体的标题和副标题仅是为了方便起见,不限制本主题技术,并且不与主题技术的描述有关地被提及。本领域普通技术人员已知或以后将知道的,贯穿本公开内容所描述的各种配置的元件的所有结构和功能等同物均通过引用明确地并入本文,并且旨在被本主题技术涵盖。此外,无论在上面的描述中是否明确叙述了这种公开,本文所公开的任何内容都不旨在献给公众。尽管已经描述了本主题技术的某些方面和实施例,但是这些仅以示例的方式给出,并且无意于限制本主题技术的范围。实际上,在不脱离其精神的情况下,可以以各种其他形式来体现本文描述的方法和系统。所附权利要求及其等同物旨在覆盖将落入本主题技术的范围和精神内的这种形式或修改。当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1