一种可高效感染免疫细胞的AAV病毒及其制备方法与应用与流程
本发明涉及一种新的AAV血清型,具体涉及一种可高效感染免疫细胞的AAV病毒及其制备方法与应用。
背景技术:
随着分子生物学的迅速发展,基因治疗已成为人类遗传性疾病或获得性疾病治疗最有效的方法之一。与传统治疗方法相比,基因治疗更接近人体基因表达的自然状态,而其关键是如何将治疗基因安全有效的导入人体内,并使治疗基因能够长期表达。与腺病毒、慢病毒载体相比,腺相关病毒(AAV)凭借着安全性高,免疫原性低,可介导基因长期稳定表达等优势,在许多临床前和临床研究中已表现出显著的潜能,在基因治疗领域受到广泛关注。
腺相关病毒(AAV)属于依赖病毒属的细小病毒科,最初于1965年作为腺病毒实验制备物中的一种污染成分被发现。目前已发现有12个天然存在的AAV血清型和100多种病毒变种。野生型AAV是一类包裹着线性单链DNA基因组的无包膜病毒,在其两端含有145bp的碱基末端重复序列(inverted terminal repeats,ITR)。ITRs序列折叠成发夹结构对于AAV病毒基因组的包装以及起始单链DNA向双链DNA的转变至关重要,且为感染性病毒颗粒所需的唯一已知的顺式作用元件。ITR元件位于两个开放阅读框的两侧,左侧Rep基因编码四个非结构Rep蛋白-Rep78、Rep68、Rep52和Rep40,其中Rep78与Rep68调控病毒的复制与整合,Rep52与Rep40则发挥解ATP酶和螺旋酶的活性作用,他们也与Rep78、Rep68共同调控病毒单链基因组的复制过程;右侧的cap基因编码三个结构蛋白-VP1、VP2和VP3,是装配完整病毒所需的衣壳蛋白,它们在病毒整合、复制和装配中起重要作用。AAV的衣壳为二十面体,由60个病毒壳粒组成,直径大约为20-25 nm,是动物病毒中最小的病毒。AAV是一种缺陷型病毒,不能单独复制,需要在辅助病毒如腺病毒、单纯疱疹病毒存在的条件下,才能在宿主细胞中复制合成和组装蛋白,产生新的病毒颗粒。野生型的AAV感染人的宿主细胞后,可定点整合到宿主细胞的AAVS1位点(19号染色体,19q13.4),这可能与野生型腺相关病毒ITR结构和Rep蛋白的共同作用相关。
目前通用的AAV病毒载体为1982年建立的感染性克隆AAV血清型2(AAV2),由于AAV2载体几乎无致病作用,弱免疫原性,宿主范围比较广,可感染分裂期的细胞和非分裂期细胞,能够介导基因长期稳定的表达等优势已广泛的应用于基因治疗中。重组AAV2载体已在许多疾病临床前研究中进行了测试,如血友病、α1抗胰蛋白酶缺乏性肝病、囊性纤维化、杜氏肌营养不良症和类风湿关节炎。但随着临床研究的增多,AAV2载体的不足也逐渐显现出来,如AAV2对组织的感染效率差异较大,在某些组织中的感染效率很高,而在有些组织中的感染效率又很低。另外,在人体中会对AAV的感染产生中和性抗体,在成人中约50-80%都存在针对各种AAV病毒的中和性抗体,抗AAV2的中和性抗体为主要形式。为了提高AAV病毒载体的感染效率和宿主范围,人们进行了多种尝试,如诱变筛选突变的衣壳蛋白或者构建嵌合的AAV病毒载体等方法。近几年不同血清型AAV病毒载体的组织特异性研究受到广泛的关注,利用各种血清型AAV的天然感染特性,获得对不同组织具有不同转导能力的AAV病毒载体,对AAV在基因治疗的广泛应用至关重要。
迄今为止,从腺病毒或人/灵长类动物组织中已分离出若干种AAV血清型和100多种AAV病毒变种。由于AAV血清型的可选择性,不仅因其潜在的高转导效率可以降低载体负荷量,也有利于规避自然感染或基于AAV病毒治疗产生的中和性抗体。
除了灵长类,AAV的基因组还可以从其他动物中分离获得。由于不同血清型AAV衣壳蛋白的氨基酸组成不同(Cap的同源性较低),导致它们与靶细胞表面的不同受体结合,因而对宿主的各种组织细胞的亲嗜性存在显著差异。无包膜病毒通过衣壳蛋白与细胞表面的糖胺聚糖受体相互作用从而进入细胞。接着病毒衣壳与辅受体的二次相互作用决定了细胞内的运输途径,该阶段称为感染途径(最终转导效率)。AAV血清型的可选择性对最终转导效率具有显著的影响。现有AAV血清型中,AAV2与细胞表面的硫酸肝素蛋白多糖受体相互作用进入宿主细胞,接着AAV2与辅受体人成纤维细胞生长因子受体1(FGFR1),肝细胞生长因子受体和整联蛋白αvβ5/α5β1相互作用对宿主细胞的转导也非常重要。AAV6,仅六个氨基酸残基不同于AAV1以及与AAV2约85%同源的衣壳序列,结合硫酸肝素,并且可以使用肝素亲和层析进行纯化。AAV3与AAV2衣壳具有约87%的同源性,同样可与硫酸肝素受体结合,但亲合力较低。AAV4和AAV5没有硫酸肝素结合为点,通过唾液酸受体与细胞结合并传导,因此其组织特异性与AAV2等有很大区别。近年来,人们已经建立了唾液酸介导的AAV1或AAV6对某些细胞类型的转导。AAV8-10 通过层黏连蛋白受体(LamR)与细胞表面结合,而用于其他AAV血清型和AAV病毒变种结合的细胞表面受体仍有待确定。
各种血清型AAV病毒载体对不同组织转导效率存在很大差异。已知AAV2可转导大部分的组织类型,包括肝,肌肉,肺和中枢神经系统,具有中等转导效率。 AAV9同样可转导多种组织类型,其转导效率比AAV2高。AAV1和AAV7对骨骼肌具有快速感染以及高水平的转导,由于AAV6衣壳仅有六个氨基酸残基不同于AAV1,因此也显示出倾向于对骨骼肌的转导。研究发现在小鼠肝脏和肌肉组织中各种血清型AAV病毒载体感染效率由高到低为AAV1>AAV5>AAV3>AAV2>AAV4;在大鼠视网膜中顺序是AAV5>AAV4>AAV1>AAV2>AAV3。今后,仍需要更广泛的研究来阐明其它血清型的组织趋向性以及转导效率。
一些AAV血清型显示了对神经系统的不同转导模式。一般情况下,从中枢神经系统注射,与AAV2相比, AAV1和AAV5表现出更高的转导效率。然而AAV2在整个中脑中显示了广泛转导效率,AAV4则转导特定的细胞类型,如室管膜下区的室管膜和星形胶质细胞。在肝组织中,AAV8已被证明是一个高转导效率的载体,可实现对肝脏接近100%的转导。此外,AAV6对肝组织的转导效率也比AAV2和AAV1高。值得注意的是,AAV8不仅可高效率的转导肝组织,对其他的器官也有更高的转导效率,高剂量的AAV8转导骨骼肌,对整个身体包括膜片,心脏,胰脏,平滑肌和脑均具有高的转导性。另一方面,AAV8对骨骼肌高的转导效率,需要VEGF同时给药,以便穿过血管屏障实现全身的转基因表达。一般情况下,由于血脑和血管屏障的存在全身注射AAV,很难实现对大脑和骨骼肌的转导,然而,上述的研究表明,AAV8可穿过血管的内皮细胞转导这些组织。因此, AAV血清型的组织亲嗜性非常重要(基因的传递中可导致非靶组织的转导),对于其在人类临床试验中的成功应用,需要严格控制和操纵它们的组织趋向性。另一方面,不同血清型最终的转导效率即转基因表达的动力学变化也是很重要的。如AAV2,6和8对均可有效的转导肝组织;肝门静脉注射AAV2病毒载体后,转基因表达以一个相对较慢的速度增加,通常需要6-8周达到最大的转导水平,与此相反,AAV6和AAV8转导具有快速的转基因表达,一般给药4周后达到最大水平。由于每个组织类型对AAV血清型的细胞摄取和细胞内运输机制不同,AAV血清型衣壳之间的结构性差异引起这种血清型之间的效果差异。
近年来研究发现,AAV2,AAV5和AAV6可有效的转导人单核细胞来源的DCs细胞(Dendritic cells),对DC细胞的形态和功能特性无不利影响,与其他血清型AAV病毒载体相比,AAV5呈现对人DC细胞高的倾向性,但转导效率低;而AAV6对小鼠来源的DC细胞具有高的转导效率。至今没有发现可以高效感染免疫细胞的AAV血清型,为了获得更高转导效率的AAV病毒变种,更好的用于临床基因治疗,很有必要研制其它血清型的AAV病毒。
技术实现要素:
本发明的目的为了提供一种新血清型的AAV病毒,其能够高效感染免疫细胞,为AAV作为基因载体的应用提供新的途径,为基因治疗提供更广泛的选择。
为达到上述发明目的,本发明采用的技术方案是:
一种可高效感染免疫细胞的AAV病毒,包括衣壳蛋白、非结构蛋白以及结构蛋白,其中,所述结构蛋白的氨基酸序列为SEQ ID NO:1所示。
上述技术方案中,氨基酸序列为SEQ ID NO:1的结构蛋白可以由序列为SEQ ID NO:2的DNA编码,也可以由序列为SEQ ID NO:3的DNA编码。
腺相关病毒(AAV病毒)是一种复制缺陷型DNA病毒,由三种包被蛋白(衣壳蛋白、非结构蛋白以及结构蛋白)与一条单链DNA构成,每条DNA单链包括ITR元件以及开放阅读框;本发明的AAV病毒可以装载外源基因,并将其携带至免疫细胞,高效感染后将外源基因注入免疫细胞中,发挥作用,因此可以作为外源基因载体的应用;从而本发明还公开了基于上述可高效感染免疫细胞的AAV病毒的重组腺相关病毒。由上述可高效感染免疫细胞的AAV病毒与外源基因组成,其中外源基因为线性单链结构;外源基因的大小为6bp~6000bp。
本发明还公开了上述重组腺相关病毒的制备方法,包括以下步骤:
(1)合成SEQ ID NO:2或者SEQ ID NO:3的DNA序列;然后将合成的DNA序列克隆至pAAV-RC载体中;
(2)然后取步骤(1)的克隆产物转化至大肠杆菌感受态中,待单克隆菌落长出后,挑单克隆菌落加入SOC培养基,培养后,将菌液转移至LB培养基中,加入终浓度为50ug/mL的氨苄青霉素,进行摇菌扩大培养;最后提取质粒并进行测序验证正确后获得腺相关病毒质粒;
(3)在培养皿中按照3x106个/10cm的密度接种HEK293T细胞;48小时后,在接种HEK293T细胞的培养皿中,利用PEI转染步骤(2)制备的腺相关病毒质粒、携带外源基因的pAAV质粒以及pHelper质粒;转染48~72小时后,收集细胞,离心去掉上清后,加入细胞裂解液,经过3个冻融循环后,离心分离得到腺相关病毒提取液;所述冻融循环的温度为-80度~37度;外源基因为线性单链结构;外源基因的大小为6bp~6000bp;
(4)将质量浓度分别为15%、25%、40%、54%的碘克沙醇溶液依次加入离心管中;然后加入步骤(3)的腺相关病毒提取液;然后离心处理,去除上清,得到重组腺相关病毒。
上述技术方案中,步骤(1)中,利用PacI-AscI双酶切所述合成的DNA序列,回收酶切产物,得到酶切后的DNA片段;利用PacI-AscI双酶切pAAV-RC载体,回收酶切产物,得到酶切载体;将酶切后的DNA片段与酶切载体配制T4连接反应体系,16℃连接过夜;从而将DNA序列克隆至pAAV-RC载体中。
上述技术方案中,步骤(2)中,然后取克隆产物转化至Stbl3大肠杆菌感受态中,待单克隆菌落长出后,挑3-5个单克隆菌落加入5mL SOC培养基,37度180RPM培养1小时后,将所有的液体转移至800mL LB培养基中,加入终浓度为50ug/mL的氨苄青霉素,37度摇瓶培养16小时进行摇菌扩大培养;最后使用质粒大提试剂盒提取质粒并进行测序验证正确后获得腺相关病毒质粒。
上述技术方案中,步骤(3)中,在接种HEK293T细胞的培养皿中,加入步骤(2)制备的腺相关病毒质粒、携带外源基因的pAAV质粒以及pHelper质粒,再按照终浓度为9μg/mL的量添加PEI,进行转染;转染48小时后,收集细胞;加入终浓度为50U/mL全能核酸酶(Benzonase),在37度孵育1小时,然后3000g离心30分钟;得到腺相关病毒提取液;所述细胞裂解液的pH为8.5,包括50mM Tris-HCl和50mM NaHCO3。
上述技术方案中,步骤(4)中,离心管中,15%、25%、40%、54%的碘克沙醇溶液以及腺相关病毒提取液的体积比为5∶7∶5∶5∶15;离心处理时的参数为28000RPM、16度离心5个小时。
本发明的重组腺相关病毒高效感染免疫细胞后将外源基因注入免疫细胞中,发挥作用;因此本发明还公开了所述重组腺相关病毒在制备基因治疗药物中的应用。
与现有技术相比,本发明创造性的制备出具有新结构蛋白的新型AAV病毒,其对免疫细胞具有高效的感染性;可以作为基因载体,与外源基因组成重组腺相关病毒,将外源基因携带至免疫细胞,高效感染后将外源基因注入免疫细胞中,从而发挥外源基因作用;对外源基因的适应性广,并且该AAV病毒不需复杂的制备工艺,极低的免疫原性,对人体安全,为基因治疗提供更广泛的选择。
附图说明
图1为现有AAV病毒以及本发明AAV病毒包装GFP基因感染免疫细胞的效果对比图;
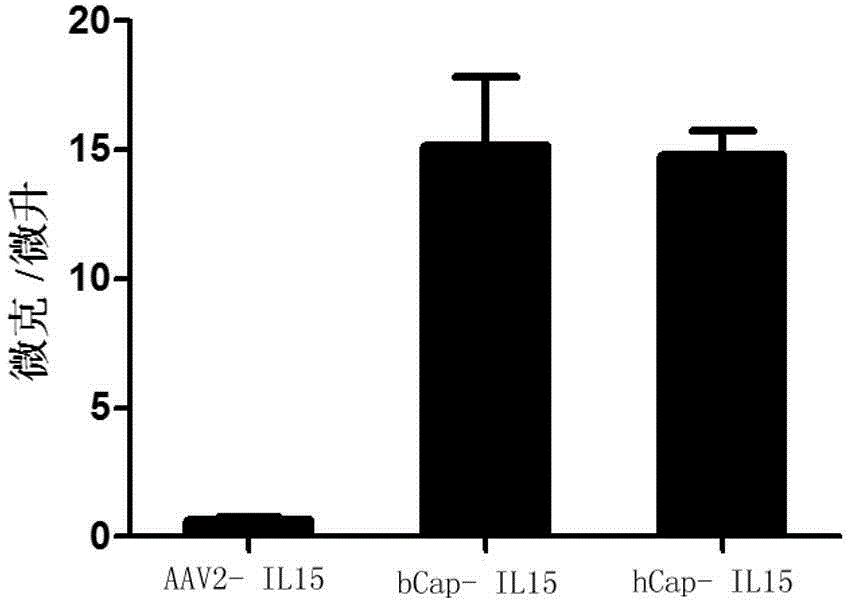
图2为现有AAV病毒以及本发明AAV病毒包装IL15基因感染免疫细胞后,目的蛋白分泌表达的效果对比图。
具体实施方式
下面结合附图以及实施例对本发明作进一步的描述。
实施例一
(1)采用化学合成的方法直接合成SEQ ID NO:2所示的DNA序列,称为hCap基因,并进行PacI-AscI双酶切,得到酶切后的DNA片段;利用PacI-AscI双酶切pAAV-RC载体,回收酶切产物,得到酶切载体;将酶切后的DNA片段与酶切载体配制T4连接反应体系,16℃连接过夜;从而将DNA序列克隆至pAAV-RC载体中,经测序验证正确后,将连接后的产物(克隆产物)转化至30uL Stbl3大肠杆菌感受态中。转化结束后,立即加入1mL预热的SOC培养基,然后将其转至15mL的离心管中,并加入5mLSOC培养基,37度180RPM培养1小时后,将所有的液体转移至800mL LB培养基中,加入终浓度为50ug/mL的氨苄青霉素,37度摇瓶培养16小时。培养结束后,3000g离心收集细菌沉淀,使用质粒大提的试剂盒分离质粒DNA并进行测序验证正确后获得腺相关病毒质粒。即可高效感染免疫细胞的AAV病毒,零下80度保存。
实施例二
(1)采用化学合成的方法直接合成SEQ ID NO:3所示的DNA序列,称为bCap基因,并进行PacI-AscI双酶切,将其克隆至pAAV-RC中,经测序验证正确后,将连接后的产物通过电击的方法转化至30uL Stbl3大肠杆菌感受态中。电转化结束后,立即加入1mL预热的SOC培养基,然后将其转至15mL的离心管中,并加入5mLSOC培养基,37度180RPM培养5小时后,将所有的液体转移至500mL LB培养基中,加入终浓度为50ug/mL的氨苄青霉素,37度摇瓶培养18小时。培养结束后,2000g离心收集细菌沉淀,使用质粒大提的试剂盒分离质粒DNA并进行测序验证正确后获得腺相关病毒质粒。即可高效感染免疫细胞的AAV病毒。
实施例三
(1)采用化学合成的方法直接合成hCap基因,并进行PacI-AscI双酶切,将其克隆至pAAV-RC中,经测序验证正确后,用于后续的AAV病毒包装。将连接后的产物通过电击的方法转化至30uL Stbl3大肠杆菌感受态中。电转化结束后,立即加入1mL预热的SOC培养基,然后将其转至15mL的离心管中,并加入5mLSOC培养基,37度180RPM培养5小时后,将所有的液体转移至500mL LB培养基中,加入终浓度为50ug/mL的氨苄青霉素,37度摇瓶培养18小时。培养结束后,2000g离心收集细菌沉淀,使用质粒大提的试剂盒分离质粒DNA并进行测序验证正确后获得腺相关病毒质粒。
(2)准备20个15cm培养皿,每皿接种4.5x10^6个HEK293免疫细胞。48小时后,使用PEI转染220ug 腺相关病毒质粒及250ug pAAV-IL15和pHelper质粒。48小时后,使用细胞刮将细胞从培养皿底部收集至离心管中,去掉上清后,使用细胞裂解液(50mM Tris-HCl,pH8.5,50mM NaHCO3),经过3个冻融循环(-80度/37度)后,加入50U/mL Benzonase在37度孵育1小时,然后2000g离心20分钟。
(3)制备15%、25%、40%、50%的碘克沙醇梯度。在SW28离心管中,依次加入5mL 54%、7mL 40%、5mL 25%、5mL 15%的碘克沙醇梯度、15mLAAV粗提液。28000RPM、16度离心6个小时,收集40%的碘克沙醇梯,离心处理,去除上清,得到外源基因为IL15基因的重组腺相关病毒,表示为bCap- IL15。
将步骤(1)的hCap基因替换为bCap基因,可得到外源基因为IL15基因的重组腺相关病毒,表示为hCap- IL15。
将步骤(2)的pAAV-IL15替换为pAAV-EGFP,可得到外源基因为GFP基因的重组腺相关病毒,表示为hCap-GFP。
将步骤(1)的hCap基因替换为bCap基因,步骤(2)的pAAV-IL15替换为pAAV-EGFP,可得到外源基因为GFP基因的重组腺相关病毒,表示为bCap-GFP。
以GFP基因、IL15基因为外源基因,以上述可高效感染免疫细胞的AAV病毒为载体,以现有AAV2为对比载体(得到的重组腺相关病毒为AAV2-GFP、AAV2- IL15),制备重组腺相关病毒,并进行荧光测试以及表达浓度测试。分别使用上述制备的含有绿色荧光蛋白GFP、IL15的AAV病毒感染免疫细胞,3天后,在荧光显微镜下观察绿色荧光蛋白的表达情况。使用酶联免疫标记试剂盒检测培养基上清中抗体的表达浓度。
图1为使用AAV2包装GFP及本发明的AAV病毒包装GFP感染免疫细胞的效果对比图,可以看出使用天然的AAV2血清型腺相关病毒感染免疫细胞,感染效率较低,GFP绿色荧光蛋白表达相对较弱;而以本发明的AAV病毒包装GFP感染免疫细胞,感染效率较高,GFP表达的绿色荧光蛋白亮度显著提高。
分别使用本发明的AAV病毒及天然的AAV2携带IL15基因,分别感染免疫细胞,连续培养72小时后,收集培养基上清后,采用ELISA的方法检测IL15的分泌表达量,图2为培养基上清中IL15的表达,结果显示本发明改良后的AAV血清能够有效感染免疫细胞,并高效表达IL15。
综上可以说明本发明提供的新型AAV病毒对免疫细胞具有高效的感染性;可以作为基因载体,与外源基因组成重组腺相关病毒,将外源基因携带至免疫细胞,高效感染后将外源基因注入免疫细胞中,从而发挥外源基因的治疗作用;对外源基因的适应性广,并且该AAV病毒不需复杂的制备工艺,对人体安全,为基因治疗提供更广泛的选择。
SEQUENCE LISTING
<110> 爱康得生物医学技术(苏州)有限公司
<120> 一种可高效感染免疫细胞的AAV病毒及其制备方法与应用
<160> 3
<170> PatentIn version 3.3
<210> 1
<211> 735
<212> PRT
<213> 人工序列
<400> 1
Met Ala Ala Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Thr Leu Ser
1 5 10 15
Glu Gly Ile Arg Gln Trp Trp Lys Leu Lys Pro Gly Pro Pro Pro Pro
20 25 30
Lys Pro Ala Glu Arg His Lys Asp Asp Ser Arg Gly Leu Val Leu Pro
35 40 45
Gly Tyr Lys Tyr Leu Gly Pro Phe Asn Gly Leu Asp Lys Gly Glu Pro
50 55 60
Val Asn Glu Ala Asp Ala Ala Ala Leu Glu His Asp Lys Ala Tyr Asp
65 70 75 80
Arg Gln Leu Asp Ser Gly Asp Asn Pro Tyr Leu Lys Tyr Asn His Ala
85 90 95
Asp Ala Glu Phe Gln Glu Arg Leu Lys Glu Asp Thr Ser Phe Gly Gly
100 105 110
Asn Leu Gly Arg Ala Val Phe Gln Ala Lys Lys Arg Val Leu Glu Pro
115 120 125
Leu Gly Leu Val Glu Glu Pro Val Lys Thr Ala Pro Gly Lys Lys Arg
130 135 140
Pro Val Glu His Ser Pro Val Phe Pro Asp Ser Ser Ser Gly Phe Gly
145 150 155 160
Lys Ala Gly Gln Gln Pro Ala Arg Lys Arg Leu Asn Phe Gly Gln Thr
165 170 175
Gly Asp Ala Asp Ser Val Pro Asp Pro Gln Pro Leu Gly Val Pro Pro
180 185 190
Ala Ala Pro Ser Gly Leu Gly Ser Ser Thr Met Ala Thr Gly Ser Gly
195 200 205
Ala Pro Met Ala Asp Asn Asn Glu Gly Ala Asp Gly Val Gly Asn Ser
210 215 220
Ser Gly Asn Trp His Cys Asp Ser Thr Trp Met Gly Asp Arg Val Ile
225 230 235 240
Thr Thr Ser Thr Arg Thr Trp Ala Leu Pro Thr Tyr Asn Asn His Leu
245 250 255
Tyr Lys Gln Ile Ser Ser Gln Ser Gly Ala Ser Asn Asp Asn His Tyr
260 265 270
Phe Gly Tyr Ser Thr Pro Trp Gly Tyr Phe Asp Phe Asn Arg Phe His
275 280 285
Cys His Phe Ser Pro Arg Asp Trp Gln Arg Leu Ile Asn Asn Asn Trp
290 295 300
Gly Phe Arg Pro Lys Arg Leu Asn Phe Lys Leu Phe Asn Ile Gln Val
305 310 315 320
Lys Glu Val Thr Gln Asn Asp Gly Thr Thr Thr Ile Ala Asn Asn Leu
325 330 335
Thr Ser Thr Val Gln Val Phe Thr Asp Ser Glu Tyr Gln Leu Pro Tyr
340 345 350
Val Leu Gly Ser Ala His Gln Gly Cys Leu Pro Pro Phe Pro Ala Asp
355 360 365
Val Phe Met Val Pro Gln Tyr Gly Tyr Leu Thr Leu Asn Asn Gly Ser
370 375 380
Gln Ala Val Gly Arg Ser Ser Phe Tyr Cys Leu Glu Tyr Phe Pro Ser
385 390 395 400
Gln Met Leu Arg Thr Gly Asn Asn Phe Thr Phe Ser Tyr Thr Phe Glu
405 410 415
Asp Val Pro Phe His Ser Ser Tyr Ala His Ser Gln Ser Leu Asp Arg
420 425 430
Leu Met Asn Pro Leu Ile Asp Gln Tyr Leu Tyr Tyr Leu Ser Arg Thr
435 440 445
Asn Thr Pro Ser Gly Thr Thr Thr Gln Ser Arg Leu Gln Phe Ser Gln
450 455 460
Ala Gly Ala Ser Asp Ile Arg Asp Gln Ser Arg Asn Trp Leu Pro Gly
465 470 475 480
Pro Cys Tyr Arg Gln Gln Arg Val Ser Lys Thr Ser Ala Asp Asn Asn
485 490 495
Asn Ser Glu Tyr Ser Trp Thr Gly Ala Thr Lys Tyr His Leu Asn Gly
500 505 510
Arg Asp Ser Leu Val Asn Pro Gly Pro Ala Met Ala Ser His Lys Asp
515 520 525
Asp Glu Glu Lys Phe Phe Pro Gln Ser Gly Val Leu Ile Phe Gly Lys
530 535 540
Gln Gly Ser Glu Lys Thr Asn Val Asp Ile Glu Lys Val Met Ile Thr
545 550 555 560
Asp Glu Glu Glu Ile Arg Thr Thr Asn Pro Val Ala Thr Glu Gln Tyr
565 570 575
Gly Ser Val Ser Thr Asn Leu Gln Arg Gly Asn Arg Gln Ala Ala Thr
580 585 590
Ala Asp Val Asn Thr Gln Gly Val Leu Pro Gly Met Val Trp Gln Asp
595 600 605
Arg Asp Val Tyr Leu Gln Gly Pro Ile Trp Ala Lys Ile Pro His Thr
610 615 620
Asp Gly His Phe His Pro Ser Pro Leu Met Gly Gly Phe Gly Leu Lys
625 630 635 640
His Pro Pro Pro Gln Ile Leu Ile Lys Asn Thr Pro Val Pro Ala Asn
645 650 655
Pro Ser Thr Thr Phe Ser Ala Ala Lys Phe Ala Ser Phe Ile Thr Gln
660 665 670
Tyr Ser Thr Gly Gln Val Ser Val Glu Ile Glu Trp Glu Leu Gln Lys
675 680 685
Glu Asn Ser Lys Arg Trp Asn Pro Glu Ile Gln Tyr Thr Ser Asn Tyr
690 695 700
Asn Lys Ser Val Asn Val Asp Phe Thr Val Asp Thr Asn Gly Val Tyr
705 710 715 720
Ser Glu Pro Arg Pro Ile Gly Thr Arg Tyr Leu Thr Arg Asn Leu
725 730 735
<210> 2
<211> 2208
<212> DNA
<213> 人工序列
<400> 2
atggccgccg atggctacct gcctgactgg ctggaagata ccctgagcga gggcatccgg 60
cagtggtgga agctgaagcc tggaccccct ccacctaagc ccgccgagag acacaaggac 120
gacagcagag gactggtgct gcccggctac aagtacctgg gccccttcaa cggcctggac 180
aagggcgagc ctgtgaacga agccgatgcc gccgctctgg aacacgacaa ggcctacgac 240
agacagctgg acagcggcga caacccctac ctgaagtaca accacgccga cgccgagttc 300
caggaacggc tgaaagagga caccagcttc ggcggcaatc tgggcagagc cgtgtttcag 360
gccaagaaac gggtgctgga acccctgggc ctggtggaag aacccgtgaa aaccgcccct 420
ggcaaaaagc ggcccgtgga acacagcccc gtgttccctg atagcagcag cggctttggc 480
aaggccggac agcagcccgc cagaaagaga ctgaacttcg gccagaccgg cgacgccgat 540
agcgtgccag atcctcagcc tctgggagtg cctcctgccg ctccttctgg cctgggcagc 600
tctacaatgg ccacaggctc tggcgcccct atggccgaca acaatgaagg cgctgacggc 660
gtgggcaaca gctccggcaa ttggcactgc gacagcacct ggatgggcga cagagtgatc 720
accaccagca ccagaacatg ggccctgccc acctacaaca accacctgta caagcagatc 780
agcagccagt ccggcgccag caacgacaac cactacttcg gctacagcac cccctggggc 840
tacttcgact tcaaccggtt ccactgccac ttcagcccca gagactggca gcggctgatc 900
aacaacaact ggggcttccg gcccaagcgg ctgaacttca agctgttcaa tattcaagtg 960
aaagaagtga cccagaacga cggcaccacc acaatcgcca acaacctgac cagcaccgtg 1020
caggtgttca ccgacagcga gtaccagctg ccctacgtgc tgggatctgc ccaccaggga 1080
tgcctgcctc cctttcccgc cgacgtgttc atggtgcccc agtacggcta cctgaccctg 1140
aacaatggca gccaggccgt gggcagaagc agcttctact gcctggaata cttccccagc 1200
cagatgctgc ggaccggcaa caacttcacc ttcagctaca ccttcgagga cgtgcccttc 1260
cacagcagct acgcccactc tcagagcctg gacagactga tgaaccccct gatcgaccag 1320
tacctgtact acctgagccg gaccaacacc cccagcggca ccacaacaca gagccggctg 1380
cagttttctc aggctggcgc ctccgacatc cgggaccaga gcagaaattg gctgcctggc 1440
ccctgctacc ggcagcagag agtgtctaag accagcgccg ataacaacaa tagcgagtac 1500
tcctggaccg gcgccaccaa gtaccacctg aacggcagag actccctcgt gaaccctgga 1560
cctgccatgg ccagccacaa ggatgacgag gaaaagttct tcccacagtc cggggtgctg 1620
atcttcggca agcagggcag cgaaaagacc aacgtggaca tcgagaaagt gatgatcacc 1680
gacgaggaag agatccggac caccaaccca gtggccaccg agcagtatgg cagcgtgtcc 1740
accaacctgc agcggggcaa tagacaggcc gccaccgccg atgtgaatac tcagggcgtg 1800
ctgccaggca tggtgtggca ggatagggac gtgtacctgc agggccccat ctgggccaag 1860
atccctcaca ccgatggcca cttccacccc agccctctga tgggcggatt cggcctgaag 1920
caccccccac cccagatcct gatcaagaac acccccgtgc ccgccaaccc cagcacaaca 1980
ttttccgccg ccaagttcgc cagcttcatc acccagtaca gcacaggcca ggtgtccgtg 2040
gaaatcgagt gggagctgca gaaagaaaac agcaagcggt ggaaccccga gatccagtac 2100
acctccaact acaacaagag cgtgaacgtg gacttcaccg tggacaccaa cggcgtgtac 2160
agcgagccca gacccatcgg caccagatac ctgacacgga acctgtaa 2208
<210> 3
<211> 2208
<212> DNA
<213> 人工序列
<400> 3
atggccgctg atggctacct gcctgactgg ctggaagata ccctgtccga gggcatccgg 60
cagtggtgga agctgaagcc tggaccccct ccacctaagc ccgccgagag acacaaggac 120
gactccagag gactggtgct gcccggctac aagtacctgg gccctttcaa cggcctggac 180
aagggcgagc ctgtgaacga ggctgatgcc gccgctctgg aacacgacaa ggcctacgac 240
cggcagctgg actctggcga caacccctac ctgaagtaca accacgccga cgccgagttc 300
caggaacggc tgaaagagga cacctccttc ggcggcaatc tgggcagagc cgtgtttcag 360
gccaagaaac gggtgctgga acccctgggc ctggtggaag aacccgtgaa aaccgcccct 420
ggcaaaaagc ggcccgtgga acactccccc gtgttccctg attcctccag cggctttggc 480
aaggccggac agcagcccgc cagaaagaga ctgaacttcg gccagaccgg cgacgccgac 540
tctgtgcctg atcctcagcc tctgggagtg cctcctgctg ctccttctgg cctgggctcc 600
tctaccatgg ctaccggatc tggcgcccct atggccgaca acaatgaagg cgcagacggc 660
gtgggcaact cctccggcaa ttggcactgc gactccacct ggatgggcga cagagtgatc 720
accacctcca cccggacatg ggccctgccc acctacaaca accacctgta caagcagatc 780
agctcccagt ccggcgcctc caacgacaac cactacttcg gctactccac cccctggggc 840
tacttcgact tcaaccggtt ccactgccac ttcagccctc gggactggca gcggctgatc 900
aacaacaact ggggcttccg gcccaagcgg ctgaacttca agctgttcaa tattcaagtg 960
aaagaagtga cccagaacga cggcaccacc acaatcgcca acaacctgac ctccaccgtg 1020
caggtgttca ccgactccga gtatcagctg ccctacgtgc tgggctctgc tcaccaggga 1080
tgtctgcctc ccttccccgc cgacgtgttc atggtgcctc agtacggcta cctgaccctg 1140
aacaacggct ctcaggccgt gggccggtcc agcttctact gcctggaata cttccccagc 1200
cagatgctgc ggaccggcaa caacttcacc ttcagctaca ccttcgagga cgtgcccttc 1260
cactcctcct acgcccactc ccagtctctg gacagactga tgaaccccct gatcgaccag 1320
tacctgtact acctgtcccg gaccaacacc ccctccggca ccacaacaca gtcccggctg 1380
cagttctctc aggctggcgc ctctgacatc cgggaccagt ccagaaattg gctgcctggc 1440
ccttgctacc ggcagcagag agtgtccaag acctccgccg ataacaacaa cagcgagtac 1500
tcttggaccg gcgctaccaa gtaccacctg aacggccggg actctctcgt gaatcctggc 1560
cctgccatgg cctcccacaa ggatgacgag gaaaagttct tcccacagtc tggcgtgctg 1620
atcttcggca agcagggctc cgaaaagacc aacgtggaca tcgagaaagt gatgatcacc 1680
gacgaggaag agatccggac caccaaccct gtggccaccg agcagtatgg ctccgtgtcc 1740
accaacctgc agcggggcaa tagacaggcc gccaccgccg atgtgaatac ccagggcgtg 1800
ctgccaggca tggtgtggca ggatagggac gtgtacctgc agggccccat ctgggccaag 1860
atccctcaca ccgatggcca cttccacccc agccctctga tgggcggatt cggcctgaag 1920
caccccccac cccagatcct gatcaagaac acccccgtgc ccgccaaccc cagcaccacc 1980
ttttctgccg ccaagttcgc ctccttcatc acccagtact ccacaggcca ggtgtccgtg 2040
gaaatcgagt gggagctgca gaaagaaaac tccaagcggt ggaaccccga gatccagtac 2100
accagcaact acaacaagtc cgtgaacgtg gacttcaccg tggacaccaa cggcgtgtac 2160
tccgagccca gacccatcgg caccagatac ctgaccagaa acctgtaa 2208
- 还没有人留言评论。精彩留言会获得点赞!