水稻抗褐飞虱主效基因Bph30的分子标记及其应用的制作方法
本发明属于分子遗传学领域,涉及水稻抗褐飞虱主效基因bph30的分子标记及该分子标记在选育水稻抗褐飞虱品种中的应用。
背景技术:
水稻是世界上最主要的粮食作物之一。由于遭到不同病菌和害虫的侵袭,水稻生产受到严重的威胁。其中,稻褐飞虱就是水稻种植过程中危害最严重虫害之一,其成虫和若虫以口针刺吸水稻韧皮部中的汁液,从而引起植株的叶片变黄,严重时整个植株枯死,最后可能导致减产甚至绝收。据中国农业年鉴记载,在1966、1969、1973、1977、1983和2003年这6年中全国性褐飞虱大发生;而在1987、1991、2005、2006和2007年这5年中全国性褐飞虱特大发生,褐飞虱发生面积达到水稻种植总面积的50%以上,给我国水稻生产造成了严重的威胁。由于褐飞虱的危害多发生在水稻的灌浆期,若此时大量使用杀虫剂进行防治,会对环境和稻谷造成较严重的污染。相反,若利用抗褐飞虱基因来培育抗虫水稻品种,这将是褐飞虱综合防治中最为经济有效的方法。迄今为止,在不同的籼稻品种和野生稻材料中已经鉴定并报道了28个抗褐飞虱主效基因(yingwangetal.2015map-basedcloningandcharacterizationofbph29,ab3domain-containingrecessivegeneconferringbrownplanthopperresistanceinrice.journalofexperimentalbotany,vol.66,no.19pp.6035–6045.)。除了bph5和bph8外,其余抗褐飞虱主效基因均已经利用分子标记被定位到水稻不同的染色体上。随着抗褐飞虱基因的发现,应用和推广,褐飞虱出现新的能够致害抗性品种的生物型,而使抗性品种失去抗性。例如,利用早期鉴定的抗性基因bph1、bph2转育的抗性水稻品种经过2~3年(约24~36代)推广种植后,它们就失去对 褐飞虱群体的抗性。而携带bph2和bph3两个抗性基因的水稻品种ptb33对褐飞虱群体表现出更持久的抗性性状。可见,在水稻生产中迫切需要更多的新的抗褐飞虱基因,以利于培育更多携带不同抗性基因的水稻品种。
技术实现要素:
由于水稻材料抗虫性鉴定的复杂性,利用常规育种手段往往难以有效地导入和聚合不同的抗虫基因。本发明是在找到与抗虫基因紧密连锁或共分离的分子标记的基础上,借助分子标记辅助选择(marker-assistedselection,mas)技术可以有目的地进行抗虫基因的导入和聚合,选育持久抗性品种,延缓抗虫品种的退化年限和防止褐飞虱新生物型的发生。
本发明的目的是提供一种水稻抗褐飞虱主效基因bph30的分子标记,通过检测这些与抗褐飞虱主效基因紧密连锁的分子标记,可以预测水稻植株对褐飞虱的抗性,加快抗褐飞虱水稻品种的选择进度。
水稻抗褐飞虱主效基因bph30的分子标记由下述之一的引物对经pcr扩增获得:
1)标记引物rm16290,
左端引物序列,tgcacgttctattcatagcc
右端引物序列,caaactagattgccacaacg;
2)标记引物ssr-28,
左端引物序列,gctcatacgaccacaatc
右端引物序列,cagcactgaagtcccatt;
3)标记引物rm16294,
左端引物序列,ctctagcgatcttctcaatcctttgc
右端引物序列,gagcaggtgaaccgacattatttacc;
4)标记引物ssr-68,
左端引物序列,gagccactggatgagggaaag
右端引物序列,tggtcttgacagtacccgca;
5)标记引物ssr-69,
左端引物序列,ccagacttacaacagcatgcaa
右端引物序列,caatttatgctcgccctcgc;
6)标记引物rm16299,
左端引物序列,cgttgatgtgttccaagattcg
右端引物序列,ggtgaagcaagcacagttaggc;
7)标记引物indel-15,
左端引物序列,gctcactctatccgcacgta
右端引物序列,ttaaaacgaacgttgccggg;
8)标记引物indel-16,
左端引物序列,gacgacgcacatgctctagt
右端引物序列,ccactcgtagcgcacacata;
9)标记引物ssr-35,
左端引物序列,aggtaggtaataactcgatgaa
右端引物序列,gcttgctggtgttttgtc;
10)标记引物rm16303,
左端引物序列,ctcactctgtccacaaatacagg
右端引物序列,gttgggtggatagacagtgg;
本发明还提供水稻抗褐飞虱主效基因bph30的分子标记方法,用上述之一的引物对扩增待检水稻基因组dna,如果rm16290能够扩增出330bp的扩增片段,或者ssr-28能够扩增出390bp的扩增片段,或者rm16294能够扩增出253bp的扩增片段,或者ssr-68能够扩增出440bp的扩增片段,或者ssr-69能够扩增出320bp的扩增片段,或者rm16299能够扩增出265bp的扩增片段,或者indel-15能够扩增出206bp的扩增片段,或者indel-16能够扩增出437bp的扩增片段,或者ssr-35能够扩增出270bp的扩增片段,或者rm16303能够扩增出291bp的扩增片段,均标志着抗性水稻品种ac1613抗褐飞虱主效基因位点bph30的存在。
筛选上述分子标记引物的过程如下:
(1)最早由国际水稻研究所(irri)利用抗虫性筛选发现印度水稻品种ac1613对3种褐飞虱群体(生物型1、2、3)具有抗性(irri,bphthreattoriceproductioninasia,1979)。并且在我国武汉地区通过苗期集团法鉴定ac1613的抗性级别,方法参照huang等介绍的方法对每个单株进行0、1、3、5、7或9级的抗性评价(huangetal.2001identificationandmappingoftwobrownplanthopperresistancegenesinrice.theorapplgenet102,929–934)结果显示该品种高抗我国褐飞虱群体,抗性级别为1级。但到目前为止,抗性水稻品种ac1613含有的抗性基因的定位仍未见报道,这给该抗虫基因在水稻分子辅助育种中的应用带来很大的困难。因此,定位抗性水稻品种ac1613中的抗性基因并且找到与该基因连锁的分子标记是非常必要的,这将为该基因在我国的水稻育种应用中提供技术支持。因此,本发明筛选并开发了基于pcr技术的ssr分子标记。根据gramene网站(http://www.gramene.org)公布的ssr分子标记,按照较均 匀的遗传距离选择一定数目分子标记进行合成。
(2)以高感褐飞虱的籼稻品种9311为母本及轮回亲本,抗褐飞虱品种ac1613为父本,配制杂种,构建f2和bc1f1分离群体,用于分子标记分析;f2和bc1f1分别自交得到f3和bc1f2,以9311为轮回亲本以bc1f1为母本回交得到bc2f1,bc2f1自交得到bc2f2,构建过程如图1。用于重组单株的筛选和基因定位的验证,以上各单株均生长至分蘖期测定表型。
(3)用ctab法(murray&thompson,1980rapidisolationofhigh-molecular-weightplantdna.nucleicacidsres8:4321-4325)提取亲本ac1613、9311及f2、bc1f1群体各单株的基因组dna。用步骤(1)获得的备选标记对两亲本进行多态性筛选,pcr反应在ptc-100扩增仪上进行,扩增产物在6%的聚丙烯酰胺凝胶上进行电泳分析,记录并选择在亲本间具有多态性的ssr标记用于后续分析。
(4)采用分蘖期褐飞虱增重和蜜露分泌量进行抗虫性鉴定,褐飞虱于2006年在武汉田间采集,在tn1稻株上饲养繁殖。分蘖期时按每棵固定三个已知质量的蜡袋,每个蜡袋中放入一头新近羽化的已知质量的短翅雌虫,48h后称量虫的质量及蜡袋的质量,蜡袋大小为30mm×30mm由parafilm膜制作,称量时所用天平为十万分之一精度的天平。感性亲本9311为感性对照,抗性亲本ac1613为抗性对照,用虫增重和蜜露分泌量对f2和bc1f1群体中各单株进行抗虫性鉴定。
(5)根据f2和bc1f1群体中各单株的抗虫性,利用分离体分组混合分析法bsa的方法,从f2群体中选择12个极端抗虫的单株的dna混合构建抗性池r,为了提高bsa的分辨率,从f2群体中分别选出8个和4个极端感虫单株的 dna分别构建感性池a、b,从bc1f1群体中选出8个极端感虫单株dna构建感性池c。同时,利用在亲本间有多态性的引物分别筛选抗、感dna池并获得有多态性的分子标记,该类多态性标记很可能与抗性性状连锁。然后,根据连锁标记所在的染色体,选择该染色体上在亲本间有多态性的引物筛选f2分离群体的各个单株,pcr程序同上,获得群体基因型资料。根据连锁交换规律,利用软件joinmap3.0将群体基因型资料构建水稻的部分遗传图谱。最后,结合f2群体各个单株的分子标记基因型资料和相应的褐飞虱抗性表型,利用mapqtl5.0软件复合区间作图法,对目标染色体进行qtl位点扫描。
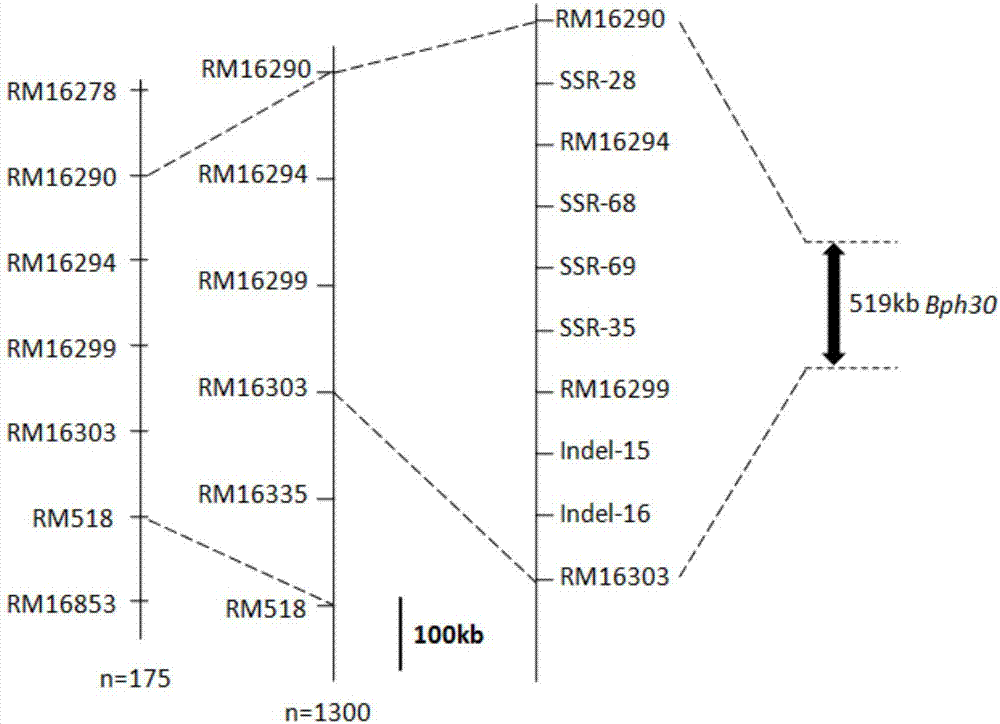
(6)根据初次qtl鉴定的结果,本发明用rm16278和rm16853筛选了175个f3和bc1f2群体的单株,得到10个在标记rm16278和rm16853之间有重组的单株,测得每个重组单株的抗虫性表型,将抗性区间缩小在标记rm16290和rm518之间。再用标记rm16290和rm518筛选1300份f3和bc1f2单株得到8个在标记rm16290和rm518发生重组的单株,测得他们的表型,可以将抗性区间缩小在标记rm16290和rm16303之间。这两个标记在参考序列日本晴基因组中的物理距离为519k。根据已测序水稻品种9311和日本晴在这两个标记间的序列开发了4个ssr标记(ssr-35、ssr-68、ssr-69、ssr-28)和两个插入缺失标记(indel-15、indel-16)。抗性表型的鉴定同(4),dna提取、ssr的pcr扩增及电泳分析同(3)得到与抗褐飞虱主效基因bph30共分离的分子标记,如图2所示。
本发明的有益效果:
1.通过本发明首次用ssr标记定位了水稻品种ac1613中的抗褐飞虱主效基因bph30。
2.通过本发明分子标记定位的主效基因位点位置明确,鉴定方便。通过检测与该基因位点连锁的分子标记,即可以预测水稻植株的褐飞虱抗性,用于水稻品种或品系的基因型检测,以判断该品种或品系是否具有褐飞虱抗性,进而快速筛选抗虫品种或品系用于水稻育种。其检测方便快速,不受环境影响。
3.本发明中抗虫性鉴定方法准确、简单、快速。在传统的抗褐飞虱育种中,材料的表型鉴定大都采用苗期集团鉴定法,该方法在抗虫鉴定前需要准备大量的虫源,且易受环境的影响,重复性差,抗褐飞虱鉴定的可靠性低。因此如果用苗期集团鉴定法准确的鉴定材料的抗褐飞虱表型不仅难度大,而且成本高。然而,通过检测抗褐飞虱主效基因位点,可以在苗期就鉴定出高抗褐飞虱的单株,淘汰其它植株,不仅节约生产成本而且大大提高抗褐飞虱水稻材料的选择效率,极大地缩短水稻品种的育种周期。
附图说明
图1为水稻抗褐飞虱主效基因bph30的克隆技术路线
以9311为母本以ac1613为父本杂交得到f1;f1自交得到f2,以9311为轮回亲本继续回交得到bc1f1;f2自交得到f3;bc1f1自交得到bc1f2,以9311为轮回亲本继续回交得到bc2f1;bc2f1自交得到bc2f2。
图2为水稻品种ac1613抗褐飞虱主效基因bph30定位在第4染色体
水平短线表示染色体上的分子标记,n为群体的单株数目;结合表现型和基因型结果,bph30定位于rm16290和rm16303之间。参照日本晴的基因组序列,这两个标记间的物理距离约为519kb。
图3为银染后读图分析性示意图
1、2、3泳道分别为抗性亲本ac1613、感性亲本9311及后代各单株的基因型,a、b分别为抗性亲本、感性亲本的基因型,后代单株中基因型和抗性亲本 一致的其基因型用a表示,基因型和感性亲本一致的用b表示,基因型为杂合的单株用h表示。
图4以分子标记indel-15表示亲本及bc2f2群体中部分单株的基因型
1、2泳道分别代表抗性亲本ac1613、感性亲本9311,3中各泳道代表bc2f2群体中部分单株的基因型,a、b、h分别表示抗性亲本、感性亲本及杂合基因型。基因型为a、h的单株表现为抗虫表型,基因型为b的单株表现为感虫表型。
图5为bc2f2群体b4-25、b4-28基因型频率分布图
a、b分别表示bc2f2群体b4-25、b4-28基因型频率分布图;纵轴表示单株的个数,横轴表示防虫48h后的褐飞虱增重量(mg);a表示纯合抗性基因型单株,b表示纯合感性基因型单株,h表示杂合基因型单株;64:20、51:24分别表示抗性单株与感性单株的比值。
具体实施方式
通过以下详细说明结合附图可以进一步理解本发明的特点和优点。所提供的实施例仅是对本发明方法的说明,而不以任何方式限制本发明揭示的其余内容。
若未特别指明,所用的技术手段为本领域技术人员所熟知的常规手段;所用的实验方法均为常规方法,并可按照已描述得重组技术(参见分子克隆,实验室手册,第2版,冷泉港实验室出版社,冷泉港,纽约)完成;所用的材料、试剂等,均可从商业途径得到。
【实施例1】与水稻抗褐飞虱主效基因bph30紧密连锁的分子标记的获得
(一)分离群体构建及表型鉴定
(1)在1979年,国际水稻研究所(rri)利用抗虫性筛选发现水稻品种 ac1613对3种褐飞虱群体(生物型1、2、3)具有抗性,2013年利用苗期集团法鉴定该品种对褐飞虱的抗性,发现该品种高抗武汉地区的褐飞虱群体,抗性级别为1级。为了定位该抗性基因并且寻找简单有效且与该基因紧密连锁的分子标记,本发明以感虫品种9311为母本,以抗褐飞虱品种ac1613为父本杂交,得到的f1;f1自交得到f2分离群体,部分f1以9311为轮回亲本回交得到分离群体bc1f1;f2和bc1f1分别自交得到f3和bc1f2;以9311为轮回亲本,以bc1f1为母本回交得到bc2f1,bc2f1自交得到bc2f2,构建过程如图1。用于重组单株的筛选和基因定位的验证,以上各单株均生长至分蘖期测定表型。
(2)采用分蘖期对亲本和f2及bc1f1群体进行抗虫性鉴定。所有供试材料在播种前分别浸种催芽,播种于塑料钵子中,每盆播4棵,播种40天后,待苗子长到分蘖期时,每棵苗子上固定三个已知质量的蜡袋,每个蜡袋中放一头新近羽化的已知质量的褐飞虱雌虫,放虫48h后分别测量相应的蜡袋和褐飞虱的重量,蜡袋及褐飞虱前后之间的差值即为48h褐飞虱分泌的蜜露和体重的增加量,用每棵苗子上的3头褐飞虱的蜜露分泌量和体重增加量的平均值来评价该棵苗子的抗虫性。每个亲本播种3棵,f2播种94棵,bc1f1播种100棵。
(二)f2、bc1f1群体的分子标记分析
(1)用ctab法(murray&thompson,1980rapidisolationofhigh-molecular-weightplantdna.nucleicacidsres8:4321–4325)提取亲本及f2、bc1f1群体各单株的基因组dna。
(2)根据gramene网站(http://www.gramene.org/)公布的ssr分子标记,按照较均匀的遗传距离选择一定数目分子标记进行合成。
(3)ssr标记的分析参照temnykh的方法(temnykhetal,2000mappingand genomeorganizationofmicrosatellitesequencesinrice.theorapplgenet100:697–712)。10μl反应体系包括:10mmtris-hclph8.3,50mmkcl,1.5mmmgcl2,50μmdntps,0.2μm引物,0.5utaqpolymerase和20ngdna模板。扩增反应在ptc-100pcr仪上进行:94℃2min;94℃15sec,55℃30sec,72℃1.5min,35个循环;72℃5min。扩增产物用6%的聚丙烯酰氨凝胶进行分离,通过银染显色(zhuetal,2004identificationandcharacterizationofanewblastresistancegenelocatedonricechromosome1throughlinkageanddifferentialanalyses.phytipathology94:515-519)。扩增的dna条带利用装有荧光灯的灯箱进行观察。记录结果,亲本间有多态的引物在f2群体中进行分析,获取群体基因型资料。
(4)根据f2及bc1f1群体各单株的抗虫性,从f2群体中选择12个极端抗虫的单株的dna混合构建抗性池r,为了提高bsa的分辨率,从f2群体中分别选出8个和4个极端感虫单株的dna构建感性池a、b,从bc1f1群体中选出8个极端感虫单株dna构建感性池c。同时,利用在亲本间有多态性的引物分别筛选抗、感dna池并获得有多态性的分子标记,该类多态性标记可能与抗性性状连锁。然后,根据连锁标记所在的染色体,选择该染色体上在亲本间有多态性的引物筛选f2分离群体的各个单株,pcr程序同上,获得群体中单株的基因型资料。根据连锁交换规律,利用软件joinmap3.0将群体基因型资料构建水稻的部分遗传图谱并获得各分子标记的遗传距离。最后,结合f2群体各个单株的分子标记基因型资料和相应的褐飞虱抗性鉴定的抗虫级别,利用mapqtl5.0软件复合区间作图法,对目标染色体进行qtl位点扫描。
(三)bph30基因的初定位
根据qtl的定位结果,用两侧的ssr标记rm16278和rm16853筛选f3 和bc1f2单株,获得两个标记间发生重组的单株。结合检测各单株的基因型和表型,缩短抗性区间。再利用缩短后的分子标记筛选f3和bc1f2单株,获得在两个标记间发生重组的单株,结合表型和基因型进一步缩短抗性区间。
(四)共分离标记的开发
根据初定位的结果,结合已测序水稻品种9311和日本晴序列开发共分离的分子标记。
(五)结果与分析
分蘖期蜜露和虫增重实验结果表明,水稻品种ac1613对褐飞虱表现出极端抗性,9311表现为极端感性;放虫48h后蜜露分泌量分别为0.15mg和4.94mg,虫增重分别为-0.16mg和1.88mg,94个f2单株对应的褐飞虱蜜露分泌量和虫增重呈连续分布,蜜露量最小为0.02mg,最大为11.92mg,虫增重最小为-0.54mg,最大值为2.82mg。蜜露分泌量和增重呈高度正相关,若以48h褐飞虱增重为0.55mg为界限,增重小于0.55mg的褐飞虱对应的单株认为是抗性单株,增重大于0.55mg的褐飞虱对应的单株认为是感性的单株,我们可以将f2群体分为抗虫、感虫2种表型。f2群体对褐飞虱的抗感分离符合3:1的比例(72:22,p=0.72>0.05)(表1)。
用亲本间有多态性的ssr标记分析f2单株的基因型,同时结合各单株对应放虫48h后褐飞虱蜜露分泌量和增重量进行qtl扫描。结果表明第4染色体短臂rm16278和rm16514间存在一个qtl位点,lod值分别为7.5和11.0,贡献率分别为31.5%和42.7%。
表19311/ac1613f2分离群体94个单株对褐飞虱抗感分离比
ar,抗性单株;s,感性单株;b3r:1s适合性检测值p=0.72>0.05;c放虫48h后褐飞虱增重量:wg,weightgain
由于rm16278和rm16514之间的物理距离较大,为了更加准确的寻找与bph30更紧密连锁的标记,本发明用rm16278作为左端标记,在rm16514的右端找到一个亲本间有多态的标记rm16853作为右端标记筛选了175个f3和bc1f2单株,有10个单株在标记rm16278和rm16853之间存在重组。于是,用这两个标记之间的其它的ssr标记检测重组单株的基因型,并结合重组单株的表型鉴定结果,将bph30定位于rm16290和rm518之间(表2)。利用这两个标记继续筛选了1300f3和bc1f2单株,有8个单株在标记rm16290和rm518之间存在重组,用这两个标记间的其他ssr标记检测重组单株基因型,结合表型鉴定结果,bph30基因定位于rm16290和rm16303之间(表3)。在测序的日本晴品种中,rm16290和rm16303之间的物理距离有519kb。根据已测序水稻品种9311和日本晴在这两个标记间的序列开发了4个ssr标记ssr-35、ssr-68、ssr-69、ssr-28和两个插入缺失标记indel-15、indel-16,经验证这些标记均能和bph30基因共分离。
表2分子标记筛选的f3和bc1f2重组单株的基因型及表现型
注:a、b分别表示抗性亲本、感性亲本基因型。
9311和ac1613是两个亲本材料;通过分析重组单株基因型和抗虫表型,将bph30基因定位于rm16290和rm518之间。
表3分子标记筛选的f3和bc1f2重组单株的基因型及表现型
注:b、h分别表示感性亲本和杂合基因型。
分子标记rm16290和rm518筛选1300份f3和bc1f2单株得到的8个重组单株,通过分析重组单株基因型和抗虫表型,将bph30基因定位于rm16290和rm16303之间。
【实施例2】分子标记的验证
1、材料和方法
1.1材料
抗性亲本ac1613,感性亲本9311,两个在定位区间为杂合的9311/ac1613bc2f2群体(b4-25、b4-28),分别有84棵和75棵。
分子标记引物:rm16290、ssr-28、rm16294、ssr-68、ssr-69、rm16299、indel-15、indel-16、ssr-35、rm16303。
1.2方法
用蜜露量和虫增重实验测得抗性亲本ac1613、感性亲本9311和两个bc2f2群体各单株的表型(方法同实施例1)。
ctab抽提法提取水稻样品基因组dna(方法同实施例1)。分别用引物rm16290、ssr-28、rm16294、ssr-68、ssr-69、rm16299、indel-15、indel-16、ssr-35、rm16303扩增样品dna。反应体系中包括0.10μm的引物、250μmdntp、1×pcr反应缓冲液(50mmkcl、10mmtris-hclph8.3、1.5mmmgcl2)、100ng的dna模板、1utaq酶。反应程序为:94℃预变性5min,循环(94℃1min、 51-61℃1min、72℃1min)41次,最后72℃延伸10min。pcr扩增产物在6%的聚丙烯胺凝胶电泳分离,电泳后用硝酸银银染法染色,读图分析如图3。
2、结果:
(1)用蜜露量和虫增重实验测得两个bc2f2群体的表型结果表明,以褐飞虱增重0.55mg为界限,增重小于0.55mg的为抗性单株,增重大于0.55mg的为感性单株,两个群体(b4-25、b4-28)的抗性单株比感性单株符合3:1分离,分别为64:20(3:1p=0.80>0.05)(表4)、51:24(3:1p=0.16>0.05)(表5)。
用上述方法,用分子标记别对水稻品种ac1613、9311及两个9311/ac1613bc2f2群体进行扩增。结果表明,分子标记rm16290、ssr-28、rm16294、ssr-68、ssr-69、rm16299、indel-15、indel-16、ssr-35、rm16303在抗性单株中均能扩增出与抗性亲本相同大小的片段,而感性单株中均不能扩增出与抗性亲本相同大小的片段,只能扩增出与感性亲本相同大小的片段(图4)。
表4bc2f2分离群体b4-2584个单株对褐飞虱抗感分离比
ar,抗性单株;s,感性单株;b3r:1s适合性检测值p=0.80>0.05;c放虫48h后褐飞虱增重量:wg,weightgain
表5bc2f2分离群体b4-2875个单株对褐飞虱抗感分离比
ar,抗性单株;s,感性单株;b3r:1s适合性检测值p=0.16>0.05;c放虫48h后褐飞虱增重量:wg,weightgain
bc2f2群体b4-25、b4-28基因型频率分布见图5。
由此说明,本发明提供的分子标记方法能够准确筛选出含有抗褐飞虱的主效基因,从而大大提高育种效率。
sequencelisting
<110>武汉大学
<120>水稻抗褐飞虱主效基因bph30的分子标记及其应用
<160>20
<170>patentinversion3.3
<210>1
<211>20
<212>dna
<213>人工序列rm16290左端引物
<400>1
tgcacgttctattcatagcc20
<210>2
<211>20
<212>dna
<213>人工序列rm16290右端引物序列
<400>2
caaactagattgccacaacg20
<210>3
<211>18
<212>dna
<213>人工序列ssr-28左端引物序列
<400>3
gctcatacgaccacaatc18
<210>4
<211>18
<212>dna
<213>人工序列ssr-28右端引物序列
<400>4
cagcactgaagtcccatt18
<210>5
<211>26
<212>dna
<213>人工序列rm16294左端引物序列
<400>5
ctctagcgatcttctcaatcctttgc26
<210>6
<211>26
<212>dna
<213>人工序列rm16294右端引物序列
<400>6
gagcaggtgaaccgacattatttacc26
<210>7
<211>21
<212>dna
<213>人工序列ssr-68左端引物序列
<400>7
gagccactggatgagggaaag21
<210>8
<211>20
<212>dna
<213>人工序列ssr-68右端引物序列
<400>8
tggtcttgacagtacccgca20
<210>9
<211>22
<212>dna
<213>人工序列ssr-69左端引物序列
<400>9
ccagacttacaacagcatgcaa22
<210>10
<211>20
<212>dna
<213>人工序列ssr-69右端引物序列
<400>10
caatttatgctcgccctcgc20
<210>11
<211>22
<212>dna
<213>人工序列rm16299左端引物序列
<400>11
cgttgatgtgttccaagattcg22
<210>12
<211>22
<212>dna
<213>人工序列rm16299右端引物序列
<400>12
ggtgaagcaagcacagttaggc22
<210>13
<211>20
<212>dna
<213>人工序列indel-15左端引物序列
<400>13
gctcactctatccgcacgta20
<210>14
<211>20
<212>dna
<213>人工序列indel-15右端引物序列
<400>14
ttaaaacgaacgttgccggg20
<210>15
<211>20
<212>dna
<213>人工序列indel-16左端引物序列
<400>15
gacgacgcacatgctctagt20
<210>16
<211>20
<212>dna
<213>人工序列indel-16右端引物序列
<400>16
ccactcgtagcgcacacata20
<210>17
<211>22
<212>dna
<213>人工序列ssr-35左端引物序列
<400>17
aggtaggtaataactcgatgaa22
<210>18
<211>18
<212>dna
<213>人工序列ssr-35右端引物序列
<400>18
gcttgctggtgttttgtc18
<210>19
<211>23
<212>dna
<213>人工序列rm16303左端引物序列
<400>19
ctcactctgtccacaaatacagg23
<210>20
<211>20
<212>dna
<213>人工序列rm16303右端引物序列
<400>20
gttgggtggatagacagtgg20
- 还没有人留言评论。精彩留言会获得点赞!