一种紫茎泽兰来源的倍半萜合酶、基因、载体、工程菌及其应用的制作方法
本发明涉及一种β-榄香烯的制备,特别涉及一种紫茎泽兰来源的倍半萜合酶、基因、载体、工程菌及其制备β-榄香烯的应用。
(二)
背景技术:
榄香烯(Elemene)是从姜科植物温郁金中提取出来的具有抗癌作用的有效成分,是我国自主研发的一类新型倍半萜药物。该药在临床运用时,高效、副作用少;能够改善微循环,有助于化疗、放疗药物进入癌组织发挥作用;具有选择性抑制肿瘤细胞增殖和提高免疫功能的双重效应;还可直接作用于细胞膜,使肿瘤细胞破裂死亡。目前榄香烯正被广泛应用于肺癌、胃癌、大肠癌等恶性肿瘤的治疗。虽然榄香烯系列抗肿瘤植物药的开发和临床应用正如火如荼,市场份额也在不断扩大,但是从天然植物温郁金中提取分离榄香烯仍存在许多技术难点:第一,天然植株仅块根中含有榄香烯,且含量极低,约占千分之一到千分之二;同时榄香烯的含量易受品种,根茎质量,种植环境等诸多因素影响。第二,挥发油中成分复杂,榄香烯与许多结构相似的倍半萜类化合物混杂在一起,必须通过精密分馏、分子蒸馏或超临界二氧化碳法进行初分,再通过薄层色谱和柱层析,经过多次分离才能得到纯品,因此榄香烯的提取工艺要求高难度大。另外,化学合成榄香烯的工艺十分复杂,反应步骤近10步,其中两步反应需在-78℃低温条件进行,且需要用到二氯甲烷、氰化钠、甲苯等剧毒化合物,不利于环境保护;最终合成的产物为外消旋混合物,需要进行拆分才能获得目标产物,产率只有50%,因此化学合成路线经济性差,无法进行商业化生产。这使得榄香烯原料药的生产成本较高,对其市场推广和全球化营销起到一定的阻碍作用。
紫茎泽兰(Eupatorium adenophorum Spreng)是一种菊科泽兰属丛生型的半灌木草本植物,因其茎、叶柄为紫色,因此被称为紫茎泽兰。紫茎泽兰原产于中美洲、墨西哥、牙买加一带,并在1865年作为观赏性植物引入夏威夷群岛,1875年引入澳大利亚,但随后在新西兰、缅甸、越南、泰国等国家泛滥成灾。紫茎泽兰于20世纪50年代入侵我国,目前已在广西、贵州、云南、四川等地蔓延成灾。由于紫茎泽兰具有较强的适应性、抗逆能力和强大的竞争力,因此一旦其侵入林地,草场等地,能够很快形成单种优势群落,给生态环境造成极大危害。通过对紫茎泽兰活性成分的研究,发现其共有49种化合物,包括甾体、倍半萜类、苯丙素酚类、黄酮类、三萜类等。韦会平等人在攀西地区的紫茎泽兰中首次发现了较高含量的β-榄香烯,并且进行测定分析,对β-榄香烯在紫茎泽兰的分布和变化规律有了一定的掌握。
目前国内外研究者通过合成生物学技术,对原料植物进行生物合成途径的改造和优化,或者利用细胞和微生物,探索一条新的生物合成途径。在此领域最著名的例子就是青蒿素的生物合成研究。Keasling等人在大肠杆菌中引入了紫穗槐二烯合酶基因,其能够表达并产生青蒿素合成途径中的重要前体分子紫穗槐二烯(Amorphadiene)。此外,有文献报导在宿主菌内引入了青蒿素天然合成途径中的P450酶,能够把紫穗槐二烯氧化为青蒿素更为直接的前体物质——青蒿酸(Artemisinic acid)。青蒿酸可通过简单化学方法进行转化,变成活性青蒿素衍生物,最终达成了青蒿素的半生物合成的目的。此生物半合成路线的形成,能够使生产青蒿素的成本降低约10倍。
β-榄香烯属倍半萜类化合物,其合成途径同萜类化合物的合成途径一样,存在两条合成途径的,分别是甲羟戊酸途径(MVA途径)和5-磷酸脱氧木酮糖/2-C-甲基-D-赤藓糖醇-4-磷酸途径(DOXP/MEP途径)。这两个途径大体上可分为3个阶段,即中间体异戊烯基焦磷酸(IPP)及其双键异构体二甲基烯丙基焦磷酸的生成(DMAPP)、直接前体物质的生成和萜类生成及其修饰阶段。以萜类物质合成途径中的前体物质FPP为底物,在吉码烯A合酶的催化作用下可形成吉码烯A,吉码烯A可在高温条件下碳原子重排形成形成β-榄香烯。此外,据相关文献报导,紫茎泽兰中的β-榄香烯的含量较温郁金来说相对较高,推测紫茎泽兰中的存在活性较好的吉码烯A合成酶。研究萜类合酶基因的克隆、表达,有助于发掘萜类化合物的合成机制,从而能够进一步的研究代谢调控以及蛋白工程的改造。因此,对紫茎泽兰来源的倍半萜合酶研究具有重要的意义。
(三)
技术实现要素:
本发明的目的是提供一种紫茎泽兰来源的倍半萜合酶、基因及其在催化FPP生成榄香烯中的应用。该倍半萜合酶属于萜类合成酶,具有催化底物FPP生成β-榄香烯的活性。
为实现上述目的,本发明采用的技术方案是:
本发明提供一种紫茎泽兰来源的倍半萜合酶(即EaSQS),所述酶的氨基酸序列为SEQ.ID NO.2所示。经实验证明,上述结构的蛋白质属于萜类合成酶,能催化FPP生成一定量的榄香烯。不难想到的是,在不改变蛋白质特性的情况,适当改变氨基酸序列,仍具有本发明醇脱氢酶特性。比如,氨基酸序列SEQ ID NO.2的保守性变异多肽,或其活性片段、或其衍生物。
本发明提供一种紫茎泽兰来源的倍半萜合酶编码基因(即EaSQS基因),所述编码基因的核苷酸序列为SEQ ID No.1所示。
本发明提供一种由所述紫茎泽兰倍半萜合酶编码基因构建的重组载体。
进一步,所述重组载体按如下方法制备:将倍半萜合酶编码基因与pET28a载体连接,获得连接产物EaSQS-pET28a,即为含有紫茎泽兰倍半萜合酶编码基因的重组载体。将重组载体转化E.coli DH5α感受态细胞中,在含有50mg/mL卡那霉素的LB液体培养基中,37℃、180rpm下摇床培养1h,离心取沉淀后涂布于含有50mg/mL卡那霉素的LB平板,过夜培养后挑取单克隆进行PCR和提取重组质粒,即获得含有紫茎泽兰倍半萜合酶编码基因的重组质粒。
本发明还提供一种含有所述紫茎泽兰倍半萜合酶编码基因或重组载体的重组基因工程菌。
本发明提供一种所述紫茎泽兰倍半萜合酶编码基因在制备重组倍半萜合酶中的应用。具体所述的应用为:将含有所述紫茎泽兰倍半萜合酶编码基因的重组载体转化至大肠杆菌中,获得的重组基因工程菌在含50mg/mL卡那霉素的LB液体培养基上37℃培养12h至OD600=0.6,加入IPTG终浓度为0.2mM,25℃诱导表达15h,将诱导培养液分离纯化,获得含有重组倍半萜合酶基因的菌体细胞,采用镍柱进行亲和层析分离获得倍半萜合酶。
本发明提供一种所述紫茎泽兰来源的倍半萜合酶在制备β-榄香烯中的应用,所述的应用为:以含紫茎泽兰来源的倍半萜合酶编码基因工程菌诱导培养获得的湿菌体超声破碎后的上清液经纯化得到的纯酶为催化剂,以法尼基焦磷酸(FPP)为底物,在二硫苏糖醇(DTT)、MgCl2和稀甘油(即1,2,3-丙三醇)的作用下,于pH为7.0的Tris-HCl缓冲液中构成反应体系,在30℃进行生物转化反应,反应液分离纯化,获得β-榄香烯。所述反应体系中,催化剂的用量为0.1-1mg/L,优选0.681mg/L,所述底物终浓度为1-5μg/mL,优选2μg/mL,所述二硫苏糖醇和MgCl2的终浓度分别为0.1-2mM(优选1mM)和2-20mM(优选10mM),1,2,3-丙三醇体积终浓度为5-20%,优选10%。
进一步,所述催化剂按如下方法制备:将含紫茎泽兰来源的倍半萜合酶编码基因工程菌(优选E.Coil BL21codon plus/pET 28a/EaSQS)接种在含50μg/mL氨苄青霉素的LB液体培养基中,37℃摇床过夜培养;再将培养液以体积浓度1%的接种量接种至含50μg/mL卡那霉素的LB液体培养基,37℃培养,直到菌液OD600达到0.6-0.8时加入终浓度0.5mM的IPTG,28℃诱导16h后,离心收集湿菌体;将湿菌体用pH7.4磷酸缓冲液重悬,超声破碎细胞,离心取上清,用0.22μm醋酸纤维素滤膜过滤,滤液用镍柱亲和层析,收集目标组分,获得纯酶。
与现有技术相比,本发明的有益效果主要体现在:
本发明中的催化剂源自高产β-榄香烯的药用植物紫茎泽兰,体外活性证实,其可以催化底物FPP生成β-榄香烯,最终榄香烯的产量达到7.499μg/mL。
(四)附图说明
图1电泳分析EaSQS基因PCR扩增产物,泳道M为DNA marker,泳道1为EaSQS基因PCR产物。
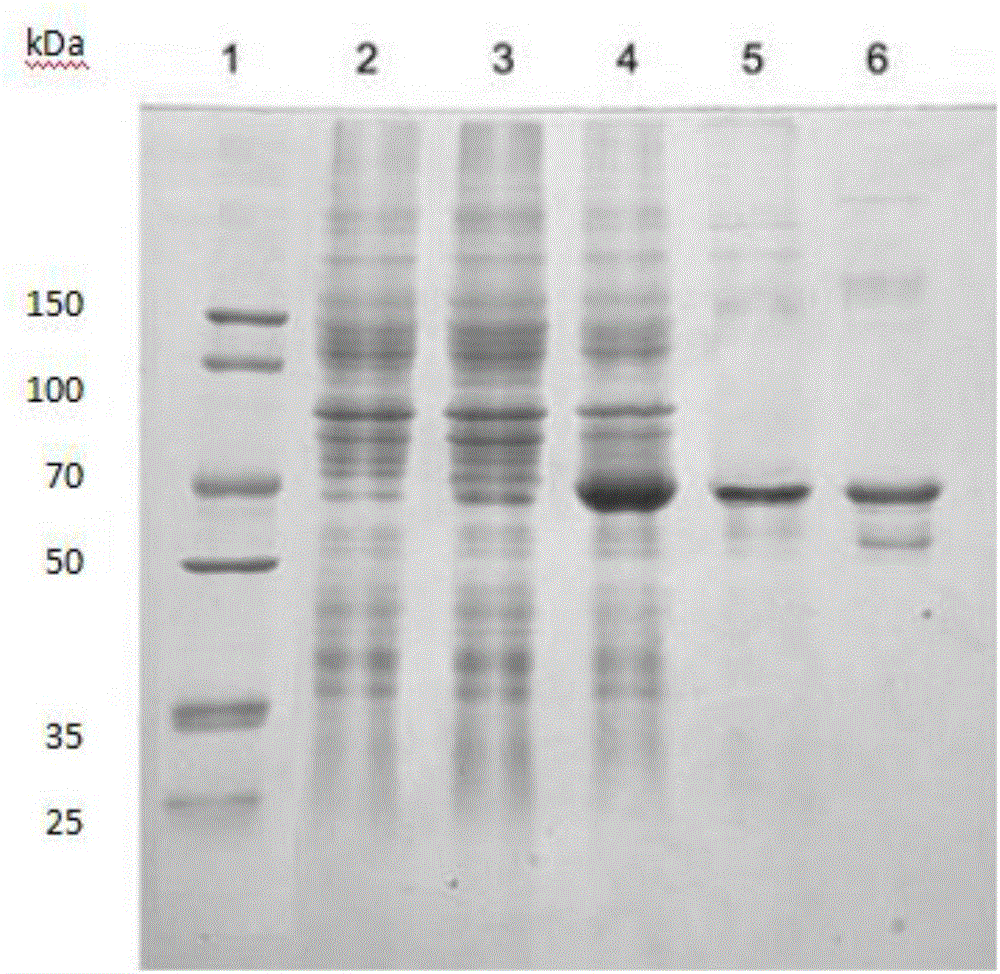
图2 SDS-PAGE分析重组EaSQS工程菌经IPTG诱导表达结果(1:marker;2,诱导前;3,诱导后;4,诱导后上清部分;5,6:EaSQS纯化后)。
图3β-榄香烯标准品(a)与EaSQS催化反应产物的GC-MS分析图(b)。
(五)具体实施方式
下面结合具体实施例对本发明进行进一步描述,但本发明的保护范围并不仅限于此:
实施例1
1、EaSQS基因全长的克隆
本发明首先以植物来源的倍半萜合酶基因的保守氨基酸序列,设计兼并引物,以紫茎泽兰cDNA为模板,进行PCR扩增,获得紫茎泽兰倍半萜合酶的部分序列,继而采用RACE技术扩增到全长序列。利用生物信息学手段进行比对分析,确定了紫茎泽兰来源的倍半萜合酶EaSQS基因的核苷酸序列的编码区。
采用Trizol法提取紫茎泽兰嫩叶RNA,反转录后送上海捷瑞生物技术有限公司进行测序,采用生物信息学BLAST分析方法,与Genbank数据库进行比对,获得一条可能的倍半萜合酶基因,根据该基因的核苷酸序列,设计一对引物(上游引物:5`-ATGGGTTGTAACCAAGTACCT-3`;下游引物:5`-TATGGTCAAAGGATCAATGAA-3`),以cDNA为模板进行PCR,PCR反应参数为:首先95℃变性5min;其次,95℃变性30sec,60℃退火30sec,72℃衍生2min,30个循环;最后72℃延伸10min;待PCR反应结束后将产物进行回收并亚克隆,测序分析,获得SEQ ID NO.1所示的核苷酸序列的EaSQS基因,该基因编码的酶EaSQS氨基酸序列为SEQ ID NO.2所示。
含目的基因的表达载体的构建,根据EaSQS基因编码序列(SEQ ID NO.1),设计扩增出完整编码阅读框的引物,并在上游和下游引物上分别引入限制性内切酶位点(上游为NdeI,下游为XhoI),具体地说,上游引物为:
5`-CATATGATGGGTTGTAACCAAGTACCT-3`,下游引物为:
5`-CTCGAGTATGGTCAAAGGATCAATGAA-3`。PCR反应参数为:首先95℃变性5min;其次,95℃变性30sec,60℃退火30sec,72℃衍生2min,30个循环;最后72℃延伸10min。经PCR扩增后,将EaSQS克隆至中间载体(如pET28a),在保证阅读框架正确的前提下鉴定好的表达载体,再将其转入E.Coil BL21codon plus中,获得工程菌E.Coil BL21codon plus/pET 28a/EaSQS。
2、EaSQS蛋白的诱导表达与纯化。
将第1步中获得的工程菌E.Coil BL21codon plus/pET 28a/EaSQS在含100mL50μg/mL氨苄青霉素的LB液体培养基中,37℃摇床过夜培养。将10ml过夜培养后的菌液倒入1L含50μg/mL卡那霉素的LB液体培养基,37℃培养,直到菌液OD600达到0.6-0.8时加入IPTG终浓度为0.5mM,28℃诱导16h后,离心收集菌体,5g湿菌体用25ml、pH7.4磷酸缓冲液重悬,超声破碎细胞。离心取上清,用0.22μm醋酸纤维素滤膜过滤,根据产品说明书,将滤液进行镍柱(Qiagen,德国)亲和层析纯化获得EaSQS重组酶液,电泳图见图1所示。重组EaSQS纯酶在SDS-PAGE上条带显示约为65kDa,与预期的蛋白分子量63.1kDa相吻合,且以上清形式存在。采用BCA法进行蛋白浓度分析,纯化后的EaSQS酶液浓度达到68.8mg/L(图1)。
实施例2重组酶EaSQS的催化性质分析
1、酶量
以实施例1方法制备的EaSQS重组酶液(浓度为68.8mg/L,体积见表1)为催化剂,加入pH7.0Tris-HCL缓冲液、2μg FPP、500mM MgCl2溶液20μL、500mM DTT2μL、100μL稀甘油(1,2,3-丙三醇)构成反应体系1ml,在30℃下搅拌充分反应,同时采用顶空-固相微萃取技术吸附反应产物,在反应60min时将萃取头取出注入气相色谱仪,对产物进行定量和定性分析。以β-榄香烯标准品为对照,通过分析色谱峰面积的大小来判断该实验所研究的倍半萜酶的催化性质。色谱条件:选用GC-2010岛津气相色谱仪;色谱柱为HP-5;载气:N2,吹扫流量为3mL/min,无分流;柱箱起始温度40℃,保留2分钟,然后以7℃/min的速度升温至220℃,保留5分钟;进样口温度为250℃;检测器温度为250℃。
标准曲线的制备方法为:吸取一定量的β-榄香烯标准品溶液,进行精密称重,加入乙酸乙酯,配置成母液浓度375.2μg/mL;取一定量的β-榄香烯标准品母液,加入一定量的乙酸乙酯,进行4、50、100、200、500倍逐级稀释,配置成186.25、14.9、7.45、3.725、1.863μg/mL的标准品浓度梯度;分别吸取1μL,按上述方法进行色谱分析,峰面积分别为6654519、884323、60439、28991、14002、11295mAu。根据β-榄香烯标准品的浓度(C)和峰面积(S)作出标准曲线:S=18032C+294.25,R2=0.99939。
结果表明:根据β-榄香烯和反应产物的GC-MS分析图谱,确定反应产物是β-榄香烯,即获得的重组酶EaSQS是一种专一性生成β-榄香烯的倍半萜合酶(图2)。
2、反应时间
以实施例1方法制备的EaSQS重组酶液(浓度为68.8mg/L,体积为10μL)为催化剂,分别加入pH7.0Tris-HCL缓冲液、2μg FPP、500mM MgCl2溶液20μL、500mM DTT 2μL、100μL稀甘油构成反应体系1ml,分别在30℃下搅拌反应20min、30min、40min、50min、60min、120min,其它操作同步骤1。
从表1中可见酶量为10μL时,即浓度比在1:3至1:4之间时,产物浓度最大。酶量在5μL时,即反应比例在1:2以下时酶的含量不足,产物产量较少;而在15μL-20μL时,即反应比例大于1:5时,酶的浓度过大,可能对反应产生了抑制,使得反应产物产量下降。
从表2中可以明显看出,产物(榄香烯)浓度随着反应时间的增加而增加。当反应时间在50min以内时,产物含量随着反应时间的增加,其增加的速率并不是很大,但是当反应时间超过50min,其产物增加的速率相对较大。可能在50min-60min或者60min-120min之间存在着一个反应速虑的突变点。
经过上述条件探索后,重组酶EaSQS在浓度为68.8mg/L,体积为10μL,pH7.0缓冲液、反应时间120min、温度30℃的反应条件下,底物FPP与重组酶EaSQS进行反应,最后用GC检测其产物的峰面积,按照上述标准曲线方程,经计算,得到最佳产量为0.75μg。
表1催化剂EaSQS的最佳酶量分析
表2EaSQS的最适反应时间
SEQUENCE LISTING
<110> 杭州师范大学
<120> 一种紫茎泽兰来源的倍半萜合酶、基因、载体、工程菌及其应用
<130>
<160> 2
<170> PatentIn version 3.5
<210> 1
<211> 1641
<212> DNA
<213> unknown
<220>
<223> 人工序列
<400> 1
atgggttgta accaagtacc tttccggccc tttcgtagca cttctccaag tttttgggga 60
gacatccttc ttaactatga aaagaaagca gagctatctg atgctgaacg tttagttgaa 120
gacttgagag aagaagtgag gaaagctata gcaggagctt tagcaaatcc aaaagaacat 180
gtgaatttgc taaaattaat tgatgtaatc caacgccttg gtataccata ttattttgaa 240
gaagagatta caaatgcttt gcaacatgtt tatgacacat atggtgatga ctggaatttc 300
ggtagccctt ccatttggtt taggctccta cgacaacatg gcttctatgt ttcatgtgat 360
atttttaaca agtactttaa ggacgaacat ggagctttca aggaatcctt aaccaatgat 420
gctgaagaaa tgctcgagct gtacgaggca acatgcttga gggtacatgg agaagttgtc 480
ctagacaagg cacttgagtt tacaaaaagt catctggcta acatagctaa ggatcctcat 540
tgtagcaacg ccactttgtc gacctatata caggaggcat tagacacacc tttgtataaa 600
aggattccaa gattagcgac gttaagttac atacgtttct acgaaaaaca agcttctcat 660
gatgaaagtt tacttaaact ggcaaagtta gggttcaatt ggcttcagtc acaacacaag 720
agcgatctta accaaatttc caaatggtgg aaagatgttg acattcaaaa gaatttgcct 780
tatgtgagag atagagtagt tgaatgttac ttttgggcat gtggttcatg ctctgagcct 840
aaatattcac ttgggagagt tttttttgct aaagtgattc aagtggggac gataattgac 900
gacacttttg atgcttatgg tacttatgaa gaacttctaa cttttacaga tgcagttgaa 960
cggtggagca ttacatgctt agatgagctt ccagagtata tgaaattcgc gtaccaagtt 1020
ttactggatt tatacgtaga aatggaacca atcgtggaaa aggaaaaatt aacacctctt 1080
tttaattgtg caaaagcgta tatgaaacaa tttgttagag cctacatggt tgaagcaaaa 1140
tgtctacatg aggggcgcat accaacagtt gatgagcata gatcaattgc atacaaaaca 1200
ggtacttgtg gctttatgat gtcagcatgc tatcttggca tgggtgatat aatcacgaat 1260
gagtcgatca attggattag tggtgaacct cctattttca tagctgcatc taagattgga 1320
agactcctaa atgatattgc cggctataag aaagagcaag aaagagagca tttccaatct 1380
tttgttcaat gctacgagaa gcaatatgat gtgggtgagg aggatgccat taacttggtg 1440
cgcaaggata ttgaagacgt atggaaagat ataacccgag agtccctcat gtgtaaagat 1500
gttccaaggc ctctcataat ggttgtggtc aactatgcac gagcattgta ttgcttgtac 1560
aagtttaatg atagtttcac agaagttggg gaagacatca aagatcatat caaacgcttg 1620
ttcattgatc ctttgaccat a 1641
<210> 2
<211> 547
<212> PRT
<213> unknown
<220>
<223> 人工序列
<400> 2
Met Gly Cys Asn Gln Val Pro Phe Arg Pro Phe Arg Ser Thr Ser Pro
1 5 10 15
Ser Phe Trp Gly Asp Ile Leu Leu Asn Tyr Glu Lys Lys Ala Glu Leu
20 25 30
Ser Asp Ala Glu Arg Leu Val Glu Asp Leu Arg Glu Glu Val Arg Lys
35 40 45
Ala Ile Ala Gly Ala Leu Ala Asn Pro Lys Glu His Val Asn Leu Leu
50 55 60
Lys Leu Ile Asp Val Ile Gln Arg Leu Gly Ile Pro Tyr Tyr Phe Glu
65 70 75 80
Glu Glu Ile Thr Asn Ala Leu Gln His Val Tyr Asp Thr Tyr Gly Asp
85 90 95
Asp Trp Asn Phe Gly Ser Pro Ser Ile Trp Phe Arg Leu Leu Arg Gln
100 105 110
His Gly Phe Tyr Val Ser Cys Asp Ile Phe Asn Lys Tyr Phe Lys Asp
115 120 125
Glu His Gly Ala Phe Lys Glu Ser Leu Thr Asn Asp Ala Glu Glu Met
130 135 140
Leu Glu Leu Tyr Glu Ala Thr Cys Leu Arg Val His Gly Glu Val Val
145 150 155 160
Leu Asp Lys Ala Leu Glu Phe Thr Lys Ser His Leu Ala Asn Ile Ala
165 170 175
Lys Asp Pro His Cys Ser Asn Ala Thr Leu Ser Thr Tyr Ile Gln Glu
180 185 190
Ala Leu Asp Thr Pro Leu Tyr Lys Arg Ile Pro Arg Leu Ala Thr Leu
195 200 205
Ser Tyr Ile Arg Phe Tyr Glu Lys Gln Ala Ser His Asp Glu Ser Leu
210 215 220
Leu Lys Leu Ala Lys Leu Gly Phe Asn Trp Leu Gln Ser Gln His Lys
225 230 235 240
Ser Asp Leu Asn Gln Ile Ser Lys Trp Trp Lys Asp Val Asp Ile Gln
245 250 255
Lys Asn Leu Pro Tyr Val Arg Asp Arg Val Val Glu Cys Tyr Phe Trp
260 265 270
Ala Cys Gly Ser Cys Ser Glu Pro Lys Tyr Ser Leu Gly Arg Val Phe
275 280 285
Phe Ala Lys Val Ile Gln Val Gly Thr Ile Ile Asp Asp Thr Phe Asp
290 295 300
Ala Tyr Gly Thr Tyr Glu Glu Leu Leu Thr Phe Thr Asp Ala Val Glu
305 310 315 320
Arg Trp Ser Ile Thr Cys Leu Asp Glu Leu Pro Glu Tyr Met Lys Phe
325 330 335
Ala Tyr Gln Val Leu Leu Asp Leu Tyr Val Glu Met Glu Pro Ile Val
340 345 350
Glu Lys Glu Lys Leu Thr Pro Leu Phe Asn Cys Ala Lys Ala Tyr Met
355 360 365
Lys Gln Phe Val Arg Ala Tyr Met Val Glu Ala Lys Cys Leu His Glu
370 375 380
Gly Arg Ile Pro Thr Val Asp Glu His Arg Ser Ile Ala Tyr Lys Thr
385 390 395 400
Gly Thr Cys Gly Phe Met Met Ser Ala Cys Tyr Leu Gly Met Gly Asp
405 410 415
Ile Ile Thr Asn Glu Ser Ile Asn Trp Ile Ser Gly Glu Pro Pro Ile
420 425 430
Phe Ile Ala Ala Ser Lys Ile Gly Arg Leu Leu Asn Asp Ile Ala Gly
435 440 445
Tyr Lys Lys Glu Gln Glu Arg Glu His Phe Gln Ser Phe Val Gln Cys
450 455 460
Tyr Glu Lys Gln Tyr Asp Val Gly Glu Glu Asp Ala Ile Asn Leu Val
465 470 475 480
Arg Lys Asp Ile Glu Asp Val Trp Lys Asp Ile Thr Arg Glu Ser Leu
485 490 495
Met Cys Lys Asp Val Pro Arg Pro Leu Ile Met Val Val Val Asn Tyr
500 505 510
Ala Arg Ala Leu Tyr Cys Leu Tyr Lys Phe Asn Asp Ser Phe Thr Glu
515 520 525
Val Gly Glu Asp Ile Lys Asp His Ile Lys Arg Leu Phe Ile Asp Pro
530 535 540
Leu Thr Ile
545
- 还没有人留言评论。精彩留言会获得点赞!