几丁质脱乙酰酶及其编码基因、重组载体、重组菌株、发酵剂和酶制剂以及它们的应用的制作方法
本发明涉及基因工程领域,具体涉及一种几丁质脱乙酰酶及其编码基因,一种重组载体,一种重组菌株,一种发酵剂,一种几丁质脱乙酰酶的制备方法,一种酶制剂,一种脱除几丁质上的乙酰基的方法,以及它们在生产几丁质脱乙酰酶中的应用和它们在脱除几丁质上的乙酰基中的应用。
背景技术:
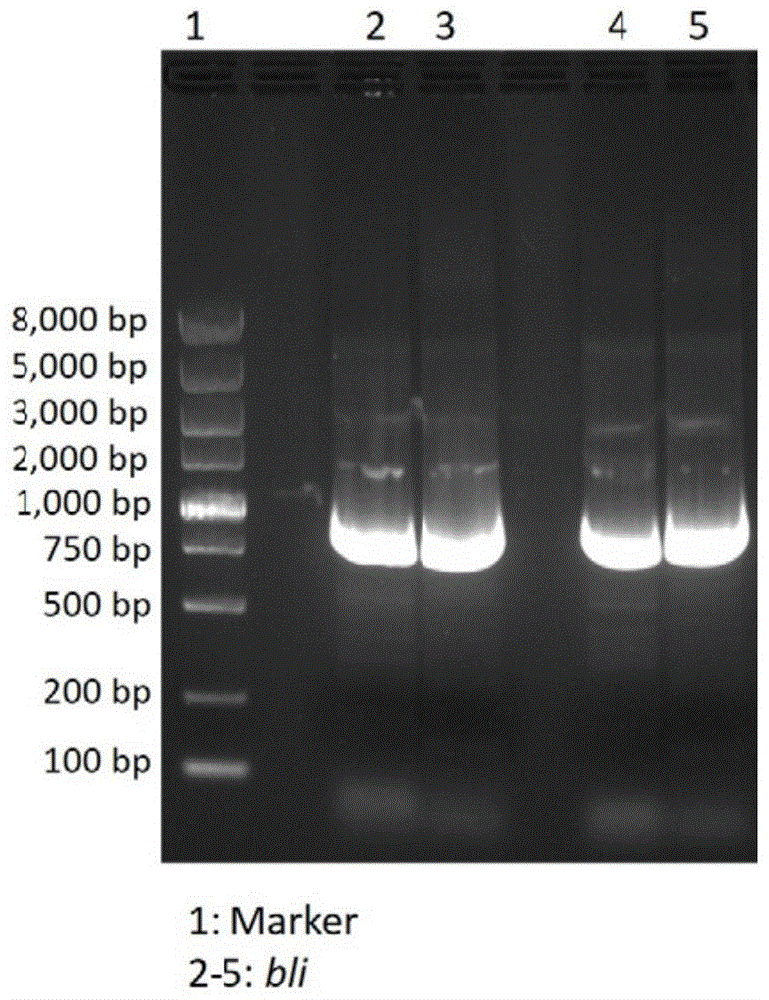
:几丁质(chitin),又名甲壳质、甲壳素,是由n-乙酰氨基-d-葡萄糖单体通过β-1,4-糖苷键连接而成的多糖。它是自然界中含量仅次于纤维素的第二大类天然高分子有机化合物,广泛存在于无脊椎动物(如虾、蟹和昆虫等)的外骨骼以及真菌和藻类的细胞壁中。几丁质由于溶解性差,不溶于水、稀酸、稀碱和有机溶剂等,大大限制了其利用价值。如几丁质脱去乙酰基(脱乙酰度大于55%)便可得到溶解性极大提高的壳聚糖(chitosan)。而且壳聚糖具有生物降解性、生物相容性、无毒性、抑菌、抗癌、降脂、增强免疫等多种生理功能,可广泛应用于医药、食品、纺织、农业、环保和化妆品等领域,有很高的应用价值和开发前景。目前,壳聚糖的工业生产方法主要为化学法。通常使用40%-60%的浓氢氧化钠高温处理几丁质制备壳聚糖。此法存在生产成本高、环境污染大以及产品稳定性和均一性差等缺点。几丁质脱乙酰酶(chitindeacetylase,e.c.3.5.1.41)可以脱去几丁质上的乙酰基生成脱乙酰度稳定、分子质量分布范围窄的壳聚糖产品。此外,酶法生产壳聚糖,反应条件温和,能耗值低且环境友好,为解决化学法制备壳聚糖存在的问题提供了一条新的途径,是壳聚糖生产行业未来的发展方向。自1974年文献首次报道鲁氏毛霉(mucorrouxii)几丁质脱乙酰酶后,研究人员已从真菌、细菌和昆虫中分离获得了多种几丁质脱乙酰酶。然而,目前报道的几丁质脱乙酰酶普遍存在发酵时间长,产酶活力低,催化温度高,脱乙酰效果差等问题,且多数为真菌来源的酶,细菌来源的少。细菌在发酵产酶方面较真菌更有优势,菌株更容易实现规模化发酵培养,产酶更容易分离纯化。因此,通过对产几丁质脱乙酰酶微生物资源的开发和基因工程的方法,构建获得高产酶的工程菌株,有望满足酶法制备壳聚糖的工业化生产需要。技术实现要素:本发明的目的是为了克服现有技术的不足,提供一种几丁质脱乙酰酶及其编码基因,一种重组载体,一种重组菌株,一种发酵剂,一种几丁质脱乙酰酶的制备方法,一种酶制剂,一种脱除几丁质上的乙酰基的方法,以及它们在生产几丁质脱乙酰酶中的应用和它们在脱除几丁质上的乙酰基中的应用,该几丁质脱乙酰酶能够通过细菌表达,具有催化温度低,适用ph广的优点。为了实现上述目的,本发明第一方面提供一种几丁质脱乙酰酶,所述几丁质脱乙酰酶为(a)-(e)中任意一项所述的酶:(a)具有seqidno.2所示氨基酸序列的酶;(b)seqidno:2所示的氨基酸序列经过取代、缺失或添加一个或几个氨基酸残基且仍具有几丁质脱乙酰酶活性的氨基酸序列所示的酶;(c)与seqidno:2所示的氨基酸序列具有80%以上同源性,且具有几丁质脱乙酰酶活性的氨基酸序列所示的酶;(d)在(a)、(b)或(c)所述氨基酸序列的氨基末端和/或羧基末端连接有标签的氨基酸序列所示的酶;(e)在(a)、(b)或(c)所述氨基酸序列的氨基末端连接有信号序列的氨基酸序列所示的酶。本发明第二方面提供一种编码几丁质脱乙酰酶的基因,该基因具有编码如上所述的几丁质脱乙酰酶的核苷酸序列。本发明第三方面提供一种重组载体,所述重组载体含有如上所述的基因。本发明第四方面提供一种重组菌株,所述重组菌株含有如上所述的基因或者如上所述的重组载体。优选地,所述重组菌株为地衣芽孢杆菌工程菌株bl22/pma5-bli,其保藏编号为cgmccno.20933。本发明第五方面提供一种发酵剂,所述发酵剂包含如上所述的重组菌株。本发明第六方面提供一种几丁质脱乙酰酶的制备方法,该方法包括:将如上所述的重组菌株和/或如上所述的发酵剂接种至发酵培养基中进行发酵,对发酵产物进行分离纯化,得到几丁质脱乙酰酶。本发明第七方面提供一种酶制剂,该酶制剂包含如上所述的方法制备得到的几丁质脱乙酰酶。本发明第八方面提供一种脱除几丁质上的乙酰基的方法,该方法包括:将如上所述的重组菌株、如上所述的发酵剂、如上所述的方法制备得到的几丁质脱乙酰酶和如上所述的酶制剂中的至少一种与几丁质接触,以对几丁质的乙酰基进行脱除。本发明第九方面提供如上所述的几丁质脱乙酰酶、如上所述的基因、如上所述的重组载体、如上所述的重组菌株、如上所述的发酵剂和如上所述的方法制备得到的几丁质脱乙酰酶中的至少一种在生产几丁质脱乙酰酶中的应用。本发明第十方面提供如上所述的几丁质脱乙酰酶、如上所述的基因、如上所述的重组载体、如上所述的重组菌株、如上所述的发酵剂、如上所述的方法制备得到的几丁质脱乙酰酶和如上所述的酶制剂中的至少一种在脱除几丁质上的乙酰基中的应用。本发明所述的几丁质脱乙酰酶的最适催化温度相对较低,为40℃左右,在较宽的ph范围内(ph值6-10)均表现出很高的催化活性,工业应用中可以节约能源和辅料等成本,具有广阔的应用前景。本发明的优点还在于在本发明所提供的优选重组菌株(地衣芽孢杆菌工程菌株bl22/pma5-bli)能够利用普通碳源和氮源快速培养并发酵高产几丁质脱乙酰酶。附图说明图1是几丁质脱乙酰酶编码基因bli琼脂糖凝胶电泳检测结果;图2是几丁质脱乙酰酶的聚丙烯酰胺凝胶电泳(sds-page)结果。生物保藏本发明提供的地衣芽孢杆菌(bacilluslicheniformis)基因工程菌株,于2020年10月22日被保藏在中国微生物菌种保藏管理委员会普通微生物中心(地址:北京市朝阳区北辰西路1号院3号,中国科学院微生物研究所,邮政编码:100101)(保藏单位的缩写为cgmcc),保藏编号为cgmccno.20933,简称bl22/pma5-bli。具体实施方式在本文中所披露的范围的端点和任何值都不限于该精确的范围或值,这些范围或值应当理解为包含接近这些范围或值的值。对于数值范围来说,各个范围的端点值之间、各个范围的端点值和单独的点值之间,以及单独的点值之间可以彼此组合而得到一个或多个新的数值范围,这些数值范围应被视为在本文中具体公开。在本发明中,在未作相反说明的情况下,使用的术语“酶活力”的大小即酶含量的多少,用酶活力单位表示,即酶单位(u),本发明中酶单位的定义是:在ph7和40℃的条件下,每小时将对硝基乙酰苯胺脱乙酰产生1μg对硝基苯胺的酶量为一个酶活力单位;“酶的比活力”代表每单位质量蛋白质的催化能力,能够反应酶活性大小,其值越大,表明酶活性越高,比活力的计算公式为:比活力(u/mg)=总酶活力单位数/mg总蛋白;单位m表示mol/l。本发明第一方面提供一种几丁质脱乙酰酶,所述几丁质脱乙酰酶为(a)-(e)中任意一项所述的酶:(a)具有seqidno.2所示氨基酸序列的酶;(b)seqidno:2所示的氨基酸序列经过取代、缺失或添加一个或几个氨基酸残基且仍具有几丁质脱乙酰酶活性的氨基酸序列所示的酶;(c)与seqidno:2所示的氨基酸序列具有80%以上同源性,且具有几丁质脱乙酰酶活性的氨基酸序列所示的酶;(d)在(a)、(b)或(c)所述氨基酸序列的氨基末端和/或羧基末端连接有标签的氨基酸序列所示的酶;(e)在(a)、(b)或(c)所述氨基酸序列的氨基末端连接有信号序列的氨基酸序列所示的酶。组成蛋白质的20种氨基酸残基,按照侧链极性可以分成四类:1、非极性的氨基酸:丙氨酸(ala)、缬氨酸(val)、亮氨酸(leu)、异亮氨酸(ile)、甲硫氨酸(met)、苯丙氨酸(phe)、色氨酸(trp)和脯氨酸(pro);2、极性不带电荷的氨基酸:甘氨酸(gly)、丝氨酸(ser)、苏氨酸(thr)、半胱氨酸(cys)、天冬氨酸(asn)、谷氨酰胺(gln)和酪氨酸(tyr);3、带正电荷的氨基酸:精氨酸(arg)、赖氨酸(lys)和组氨酸(his);4、带负电荷的氨基酸:天冬氨酸(asp)和谷氨酸(glu)(参见“生物化学”(第二版)上册,沈同、王镜岩,第82-83页,高等教育出版社,1990年12月)。蛋白质中如果发生同属一个类别的氨基酸残基取代,例如由arg取代lys或由leu取代ile,所述残基在蛋白质结构域中所起的作用(比如提供正电荷或形成疏水囊袋结构的作用)没有改变,因此对蛋白质的立体结构并不会产生影响,因此仍然可以实现蛋白的功能。所述同属一个类别的氨基酸残基取代可以发生在上述几丁质脱乙酰酶的任意一个氨基酸残基位置上。除上所述的氨基酸残基取代,本发明提供的几丁质脱乙酰酶还包括相比于seqidno.2所示氨基酸序列在1-254位氨基酸残基任意位置发生一个或多个氨基酸残基缺失或添加或兼而有之的蛋白。优选的,本发明中的几丁质脱乙酰酶氨基酸序列可以是相比于seqidno.2所示氨基酸序列可以在第2-7位,第251-254位氨基酸残基发生部分或者全部缺失的序列,或者是在第2-7位氨基酸残基之间任意位置添加1-5个氨基酸残基的序列,或者是在第251-254位氨基酸残基之间任意位置添加1-3个氨基酸残基的序列,或者上述各种情况任意组合形成的氨基酸序列。如前所述,本发明提供的几丁质脱乙酰酶还可以进行修饰或突变,得到衍生的蛋白质。衍生的蛋白质指与具有上述氨基酸序列的几丁质脱乙酰酶具有氨基酸序列上的差异,也可以有不影响序列的修饰形式上的差异,或者兼而有之。这些蛋白包括天然或诱导的遗传变异体。所述诱导变异体可以通过各种技术得到,如辐射或诱变剂等产生的随机突变,也可以通过如定点突变法或其他已知分子生物学的技术。衍生的蛋白质还包括具有天然l型氨基酸的残基的类似物(如d型氨基酸),以及具有非天然存在的或合成的氨基酸(如β-氨基酸、γ-氨基酸等)的类似物。修饰(通常不改变一级结构,即不改变氨基酸序列)形式包括:体内或体外的蛋白的化学衍生形式如乙酰化或羧基化。修饰还包括糖基化,如那些在蛋白的合成和加工中或进一步加工步骤中进行糖基化修饰而产生的蛋白。这种修饰可以通过将蛋白暴露于进行糖基化的酶(如哺乳动物的糖基化酶或去糖基化酶)而完成。修饰形式还包括具有磷酸化氨基酸残基(如磷酸酪氨酸,磷酸丝氨酸,磷酸苏氨酸)的序列。还包括被修饰从而提高了其抗蛋白水解性能或优化了溶解性能的蛋白。在本发明中,所述酶还可以是与seqidno:2所示的氨基酸序列具有80%以上同源性,且具有几丁质脱乙酰酶活性的氨基酸序列所示的酶。优选地,所述几丁质脱乙酰酶为与seqidno:2所示的氨基酸序列具有85%以上,更优选90%以上,进一步优选95%以上,再进一步优选98%以上,最优选99%以上同源性,且具有几丁质脱乙酰酶活性的氨基酸序列所示的酶。为了方便纯化,还可以采用本领域常见的标签对(a)、(b)或(c)进行添加修饰,举例来说,可以通过在(a)的氨基末端和/或羧基末端连接有标签的氨基酸序列(如下表1所示的标签,poly-arg、poly-his、flag、strep-tagⅱ和c-myc中的至少一种)而获得。所述标签不会影响本发明的几丁质脱乙酰酶的活性,在实际应用过程中,可以根据需求选择是否添加标签。表1标签残基数氨基酸序列poly-arg5-6(通常为5个)rrrrr(seqidno:3)poly-his2-10(通常为6个)hhhhhh(seqidno:4)flag8dykddddk(seqidno:5)strep-tagⅱ8wshpqfek(seqidno:6)c-myc10eqkliseedl(seqidno:7)在本发明中,所述几丁质脱乙酰酶的氨基末端还可以连接信号序列。所述信号序列可以来自地衣芽孢杆菌、解淀粉芽孢杆菌和枯草芽孢杆菌,但不限于此。所述信号序列优选seqidno:8所示的氨基酸序列。在本发明中,仍具有酶活力是指在相同的测定条件下,由(a)衍生的酶仍具有酶活力,其酶活力与(a)的酶活力之间的百分比(相对活性)不低于60%,优选不低于70%(或80%,或90%,或95%,或96%,或97%,或98%,或99%,或100%)。上述几丁质脱乙酰酶可以通过人工合成得到,也可以先合成其编码基因,再通过生物表达获得。本发明第二方面提供一种编码几丁质脱乙酰酶的基因,该基因具有编码如上所述的几丁质脱乙酰酶的核苷酸序列。相应地,所述基因可以为如下的(1)或(2):(1)具有编码第一方面所述的几丁质脱乙酰酶的氨基酸序列的dna分子;(2)在严格条件下与(1)限定的dna序列杂交且编码的几丁质脱乙酰酶的酶活性不变的dna分子。其中,所述严格条件可以为:在6×scc,0.5%sds的溶液中,在65℃下杂交,然后用2×scc,0.1%sds和1×scc,0.1%sds各洗膜一次。酶活性不变是指在相同的测定条件下,由(2)编码的蛋白质的酶活力与(1)编码的蛋白质的酶活力之间的百分比(相对活性)不低于95%(或96%,或97%,或98%,或99%,或100%)。本领域公知,组成蛋白质的20种不同的氨基酸中,除met(atg)或trp(tgg)分别为单一密码子编码外,其他18种氨基酸分别由2-6个密码子编码(sambrook等,分子克隆,冷泉港实验室出版社,纽约,美国,第二版,1989,见950页附录d)。即由于遗传密码子的简并性,决定一个氨基酸的密码子大多不止一个,三联体密码子中第三个核苷酸的置换,往往不会改变氨基酸的组成,因此编码相同蛋白的基因的核苷酸序列可以不同。本领域人员根据公知的密码子表,从本发明公开的氨基酸序列,以及由所述氨基酸序列得到的几丁质脱乙酰酶活性不变的氨基酸序列,完全可以推导出能够编码它们的基因的核苷酸序列,通过生物学方法(如pcr方法、突变方法)或化学合成方法得到所述核苷酸序列,因此该部分核苷酸序列都应该包括在本发明范围内。相反,利用本文公开的dna序列,也可以通过本领域公知的方法,例如sambrook等的方法(分子克隆,冷泉港实验室出版社,纽约,美国,第二版,1989)进行,通过修改本发明提供的核酸序列,得到与本发明所述几丁质脱乙酰酶活性一致的氨基酸序列。优选地,所述基因具有编码具有seqidno.2所示氨基酸序列的酶的核苷酸序列。如上所述,相应地,核苷酸序列的5'端和/或3'端还可以连接有上表1所示的标签的编码序列。更优选地,所述基因具有seqidno.1所示的核苷酸序列。本发明提供的核苷酸序列通常可以用聚合酶链式反应(pcr)扩增法、重组法、或人工合成的方法获得。例如,本领域技术人员根据本发明所提供的核苷酸序列,可以很容易得到模板和引物,利用pcr进行扩增获得有关序列。一旦获得了有关核苷酸序列,就可以用重组法大批量的获得有关氨基酸序列。通常将所得核苷酸序列克隆入载体,再转入基因工程菌中,然后通过常规的方法从增殖后的宿主细胞分离得到有关核苷酸序列。此外,还可用公知的人工化学合成的方法来合成有关核苷酸序列。本发明第三方面提供一种重组载体,所述重组载体含有如上所述的基因。重组载体中使用的“载体”可选用本领域已知的各种载体,如市售的各种质粒、粘粒、噬菌体及反转录病毒等,本发明优选的表达载体为pma5质粒。重组载体构建可采用能够在载体多克隆位点具有切割位点的各种核酸内切酶(如对于pma5,可用bamhⅰ、mlui等)进行酶切获得线性质粒,与采用相同核酸内切酶切割的基因片段连接,获得重组质粒。本发明优选采用bamhⅰ和mlui双酶切pma5及与其连接的基因片段,经连接酶连接,构建得到重组载体pma5-bli。本发明第四方面提供一种重组菌株,所述重组菌株含有如上所述的基因或者如上所述的重组载体。可以通过本领域常规的方法将所述重组载体转化、转导或者转染到宿主细胞(菌株)中,如氯化钙法化学转化、高压电击转化,优选电击转化。所述宿主细胞可以为原核细胞或真核细胞,优选为地衣芽孢杆菌(bacilluslicheniformis)、解淀粉芽孢杆菌和枯草芽孢杆菌中的至少一种,更优选地,所述宿主细胞为地衣芽孢杆菌,比如可以为地衣芽孢杆菌bl22。优选地,所述重组菌株为地衣芽孢杆菌工程菌株bl22/pma5-bli,其保藏编号为cgmccno.20933。本发明第五方面提供一种发酵剂,所述发酵剂包含如上所述的重组菌株。所述发酵剂可以以液体形式或固体形式存在。所述发酵剂中可以含有本领域常规制备菌剂时添加的辅料,本领域技术人员可以根据需要选择。优选的,相对于每克所述发酵剂,所述重组菌株的含量为105-1010cfu,更优选为107-109cfu。本发明第六方面提供一种几丁质脱乙酰酶的制备方法,该方法包括:将如上所述的重组菌株和/或如上所述的发酵剂接种至发酵培养基中进行发酵,对发酵产物进行分离纯化,得到几丁质脱乙酰酶。本发明提供的制备几丁质脱乙酰酶的方法包括:培养本发明提供的重组菌株,诱导编码几丁质脱乙酰酶的基因的表达;分离提纯所表达的几丁质脱乙酰酶。其中,所述培养条件为常规的培养条件,如使用lb培养基(溶剂为水,溶质及其终浓度分别为:胰蛋白胨5-15g/l,酵母提取物1-10g/l,nacl5-15g/l),在35-37℃下培养。由于本发明提供的重组菌株中含有编码几丁质脱乙酰酶的基因,其可以高效地表达几丁质脱乙酰酶。培养后经过分离提纯,即可得到高纯度的几丁质脱乙酰酶。可以采用本领域技术人员公知的方法进行分离提纯,在此不再赘述。本发明第七方面提供一种酶制剂,该酶制剂包含如上所述的方法制备得到的几丁质脱乙酰酶。所述酶制剂可以以固体、半固体或液体形式存在,所述酶制剂可以含有制备酶制剂的辅料或添加剂等,本领域技术人员可以根据需要进行选择,在此不再赘述。本发明第八方面提供一种脱除几丁质上的乙酰基的方法,该方法包括:将如上所述的重组菌株、如上所述的发酵剂、如上所述的方法制备得到的几丁质脱乙酰酶和如上所述的酶制剂中的至少一种与几丁质接触,以对几丁质的乙酰基进行脱除。脱除几丁质上的乙酰基的方法可以是本领域常规的脱除方法。其中,所述接触的条件可以是所述几丁质脱乙酰酶适合的条件,优选地,所述接触的条件包括:ph值4-13,更优选为7-10;温度为25-55℃,更优选为30-45℃。相对于每克几丁质样品,所述几丁质脱乙酰酶的用量可以为10-100μg。本发明第九方面提供如上所述的几丁质脱乙酰酶、如上所述的基因、如上所述的重组载体、如上所述的重组菌株、如上所述的发酵剂和如上所述的方法制备得到的几丁质脱乙酰酶中的至少一种在生产几丁质脱乙酰酶中的应用。本发明第十方面提供如上所述的几丁质脱乙酰酶、如上所述的基因、如上所述的重组载体、如上所述的重组菌株、如上所述的发酵剂、如上所述的方法制备得到的几丁质脱乙酰酶和如上所述的酶制剂中的至少一种在脱除几丁质上的乙酰基中的应用。以下将通过实施例对本发明进行详细描述。下述实施例中所使用的实验方法如无特殊说明,均为常规方法。下述实施例中所用的材料、试剂等,如无特殊说明,均可从商业途径得到。几丁质购自wako(日本)公司。实施例1本实施例用于说明目的基因bli的获取。委托上海生工生物工程有限公司合成如seqidno.1所示的基因序列以及seqidno.9和seqidno.10所示的引物序列。其中,seqidno.9:gcgggatccgtgaaccatttttatgtgtg;seqidno.10:gcgacgcgtctactttacttctgaagatt。上下游引物中分别引入bamhi和mlui限制性内切酶酶切位点。以合成的dna为模板,进行pcr扩增。pcr反应体系为:premix(25μl),ddh2o(22μl),上下游引物(各1μl),dna模板(1μl)。反应条件为:94℃5min,1个循环;94℃30s,55℃30s,72℃1min,30个循环;72℃10min,1个循环。pcr产物进行琼脂糖凝胶电泳分析(图1)后,切胶回收目的片段。将目的片段连接到pmd-19t克隆载体上。连接反应体系为:载体1μl,回收片段5μl,t4dna连接酶(neb)1μl,10*buffer1μl,ddh2o2μl。连接反应条件为16℃,2h。将连接液转化大肠杆菌dh5α感受态细胞。方法为:取50ul感受态细胞在冰浴上融化,加入连接液;冰浴静置30min;42℃水浴热击45sec,迅速转移到冰上静置2min,加入500ul无菌lb培养基(无抗),37℃,200rpm培养1h;6000rpm离心3min,去除部分上清,重悬细胞,取100ul涂布在含100μg/ml氨苄青霉素的lb培养基固体平板上,37℃培养过夜。菌落pcr验证。pcr反应体系为:premix(25μl),ddh2o(23μl),上下游引物(各1μl),dna模板(lb平板上单菌落)。反应条件为:94℃5min,1个循环;94℃30s,55℃30s,72℃1min,30个循环;72℃10min,1个循环。扩增产物大小正确的阳性克隆委托上海生工生物工程有限公司测序。测序结果显示,目的基因编码区长765bp,其核苷酸序列如seqidno.1所示。目的基因共编码254个氨基酸和一个终止密码子,其氨基酸序列如seqidno.2所示,预测蛋白质分子量为28kda。实施例2本实施例用于说明地衣芽孢杆菌基因工程菌株bl22/pma5-bli的获取。目的基因双酶切将实施例1获得的目的基因从克隆载体上切下。酶切反应体系为:载体pmd-19t-bli(5μl),限制性内切酶bamhi(1μl),限制性内切酶mlui(1μl),10*buffer(1μl),ddh2o(2μl);反应条件为:37℃,2h。酶切产物进行1%琼脂糖凝胶电泳分析后,切胶回收目的基因片段。表达载体双酶切对表达载体进行双酶切。酶切反应体系为:载体pma5(5μl),限制性内切酶bamhi(1μl),限制性内切酶mlui(1μl),10*buffer(1μl),ddh2o(2μl);反应条件为:37℃,2h。酶切产物进行1%琼脂糖凝胶电泳分析后,切胶回收目的基因片段。目的基因和表达载体连接将目的基因连接到pma5表达载体上。连接反应体系为:载体1μl,目的基因5μl,t4dna连接酶(neb)1μl,10*buffer1μl,ddh2o2μl。连接反应条件为16℃,2h。重组表达载体转化将连接液转化地衣芽孢杆菌bl22感受态细胞。取50ul感受态细胞在冰上融化,加入连接液;静置30min;将混合液转入预冷的电转杯;设置电转参数,按照电击电压2.5kv,电击时间5ms的条件进行电击转化;电击之后,立即将1ml42℃预热的lb培养基加至电转杯;使用移液器反复吹打混匀,然后将电击杯中的液体再转移到离心管中;放入42℃水浴锅,热击6min,之后迅速转移到冰上静置2min;37℃,200rpm振荡孵育2h;6000rpm离心3min,去除部分上清,重悬细胞,取100ul涂布在含5μg/ml氯霉素的lb培养基固体平板上,37℃培养48h。重组菌株筛选菌落pcr验证。pcr反应体系为:premix(25μl),ddh2o(23μl),上下游引物(各1μl),dna模板(lb平板上单菌落)。反应条件为:94℃5min,1个循环;94℃30s,55℃30s,72℃1min,30个循环;72℃10min,1个循环。对pcr扩增产物大小正确的阳性重组菌株进行甘油管保藏,储存于-80℃冰箱。所述地衣芽孢杆菌(bacilluslicheniformis)基因工程菌株,于2020年10月22日被保藏在中国微生物菌种保藏管理委员会普通微生物中心(地址:北京市朝阳区北辰西路1号院3号,中国科学院微生物研究所,邮政编码:100101)(保藏单位的缩写为cgmcc),保藏编号为cgmccno.20933,简称bl22/pma5-bli。实施例3本实施例用于说明几丁质脱乙酰酶的获得和酶活力的测定。重组菌株蛋白表达(1)菌种活化培养方法:取适量-80℃冰箱保藏菌株bl22/pma5-bli于添加5μg/ml氯霉素的lb固体平板划线,置于恒温培养箱培养,温度为35±2℃,时间为20h。活化培养基:酵母提取物5g/l、胰蛋白胨10g/l、nacl10g/l、琼脂粉20g/l,余量为水,ph值6.5-7.2。(2)种子培养培养方法:从活化平板挑取单菌落置于种子培养基(氯霉素浓度为5μg/ml)恒温振荡培养,搅拌速度为150rpm,温度为35±2℃,时间为9h。种子培养基:酵母提取物5g/l、胰蛋白胨10g/l、nacl10g/l,余量为水,ph值6.5-7.2。(3)发酵培养培养方法:将种子液按照1%的接种量转接至发酵培养基(氯霉素浓度为5μg/ml)恒温振荡培养,搅拌速度为150rpm,温度为35±2℃,时间为24h;发酵培养基:葡萄糖40g/l、酵母粉25g/l、胰蛋白胨15g/l、k2hpo43g/l,余量为水,ph值6.5-7.2。重组菌株bl22/pma5-bli几丁质脱乙酰酶粗酶液制备收集10ml发酵液,12000rpm离心5min,倒掉培养基上清,取10ml超声破碎缓冲液(50mmtris-cl、20mmnaoh、ph8.0)上下吸打,充分重悬菌体;将10ml重悬菌液平均分成10份,每份1ml,转移至10个2ml离心管中,将每个离心管置于固定的冰水混合浴中,单独进行超声破碎,设置超声破碎仪振幅杆为2、功率为20%,超声频率为2s/2s(即破碎2s,暂停2s),破碎总时间为5min;所有破碎液经12000rpm离心5min,收集上清至一新的15ml离心管中,即为重组菌株几丁质脱乙酰酶粗酶液。重组蛋白检测用聚丙烯酰胺凝胶电泳(sds-page)检测重组几丁质脱乙酰酶的表达情况(图2)。重组几丁质脱乙酰酶酶活检测取一支15ml离心管,加入1ml200mg/l的对硝基乙酰苯胺水溶液、1ml稀释适当倍数的粗酶液以及3ml预保温的浓度为0.05m的ph值为7.0的磷酸盐缓冲液,使反应液终体积为5ml,于40℃水浴反应0.5h,沸水浴5min,终止酶促反应,加水定容至10ml,混匀,12000rpm离心5min,测定上清液在400nm处的吸光值a400。空白对照体系中添加1ml同样浓度的灭活酶液,其余同上,测定上清液的吸光值a0,每个样品对应一个空白。酶活单位的定义:在上述反应条件下每小时产生1μg对硝基苯胺所需要的酶量定义为一个酶活力单位。酶活力计算公式为:酶活力(u/ml)=((a400-a0)*10*n)/kta400—样品在400nm处的吸光值;a0—空白的吸光值;10—溶液体积10ml;n—酶液稀释倍数;k—线性系数(0.0648);t—反应时间,h经测定,得到的几丁质脱乙酰酶的酶活力结果为6620u。实施例4本实施例用于说明几丁质脱乙酰酶的获得和酶活力的测定。按照实施例1-3的方法进行操作,不同的是,几丁质脱乙酰酶的氨基酸序列不同,具体见表1。上述不同氨基酸序列对应的酶的酶活力结果见表1。表1编号区别酶活力ub1k108r6010b2第2-7位氨基酸缺失5880b3第251-254位氨基酸缺失5310b4第2-7和251-254位氨基酸缺失4560b5第7-8位之间插入rli三个氨基酸残基4980cseqidno.11(与seqidno.2具有83%的同源性)4230d1seqidno.2的氨基末端连接seqidno.34970d2seqidno.2的羧基末端连接seqidno.46480d3seqidno.2的羧基末端连接seqidno.56010d4seqidno.2的氨基末端连接seqidno.66230d5seqidno.2的羧基末端连接seqidno.75870eseqidno.2的氨基末端连接seqidno.86590实施例5本实施例用于说明几丁质脱乙酰酶的获得和酶活力的测定。按照实施例3的方法进行操作,不同的是,分别在ph为4.0、5.0、6.0、7.0、8.0、9.0、10.0、11.0、12.0和13.0的条件下进行酶活力的测定。经测定,得到的几丁质脱乙酰酶的酶活力结果依次分别为2200u、3450u、5860u、6620u、6360u、6320u、6240u、5090u、3310u和2180u。实施例6本实施例用于说明几丁质脱乙酰酶的获得和酶活力的测定。按照实施例3的方法进行操作,不同的是,分别在温度为20、25、30、35、40、45、50和55℃的条件下进行酶活力的测定。经测定,得到的几丁质脱乙酰酶的酶活力结果分别为1390u、3130u、4410u、5470u、6620u、3980u、1970u和810u。通过以上结果可以看出,采用本发明所述的地衣芽孢杆菌基因工程菌株bl22/pma5-bli生产的几丁质脱乙酰酶具有适用ph广和适用温度偏低的特点,且酶活力高,适合工业生产。以上详细描述了本发明的优选实施方式,但是,本发明并不限于此。在本发明的技术构思范围内,可以对本发明的技术方案进行多种简单变型,包括各个技术特征以任何其它的合适方式进行组合,这些简单变型和组合同样应当视为本发明所公开的内容,均属于本发明的保护范围。sequencelisting<110>吉林中粮生化有限公司;中粮营养健康研究院有限公司;中粮生物科技股份有限公司;中粮生化能源(榆树)有限公司<120>几丁质脱乙酰酶及其编码基因、重组载体、重组菌株、发酵剂和酶制剂以及它们的应用<130>65859<160>11<170>patentinversion3.3<210>1<211>765<212>dna<213>artificiallysynthesized<400>1atgaaccatttttatgtgtggcatatcaaacggattaagcagctaatcattattatgata60gccgcttttgcgacagcaagttttttttatgtgcaaaacctgctccctcttcctgtgttt120tctacagaaggcggagcaaaagcggtatatagaggagattcagatacaaatgaagtagcc180cttacatttaatatcagctggggagatcaaaaggcaatgcccattttagacacattaaaa240gcaaacggtattaaagacgcgaccttttttctatcagcttcatgggcagagcgccacccg300gatgtcgtagaaagaatccgtaaagatggtcaccagatcgggagtatgggctatgcttat360aaaaactattcgcaaatgaagaaaagcgagatcaaaaaagacttagcaaaagcacgacac420tcctttcaaaaactcgggcttgacgaccttacgcttttaagaccgccgaccggccagttt480aataaagacgtactcgatgttgctaaacagtacggctacaccgttgttcattatagtatt540aactcggatgactggacgaacccgggggttcaaaagatcgtccaaaacgtaaatgaaacg600gtaaacgccggtgacatcgtgctctttcacgcttcagattccgccaaacaaacaaaagaa660gccctgccagagatcgtgcaccatctcagaagcaaggggctcaaaaacgtaaccgtcagc720gaattaatcgcaaatacggatgcaaaatcttcagaagtaaagtag765<210>2<211>254<212>prt<213>artificiallysynthesized<400>2metasnhisphetyrvaltrphisilelysargilelysglnleuile151015ileilemetilealaalaphealathralaserphephetyrvalgln202530asnleuleuproleuprovalpheserthrgluglyglyalalysala354045valtyrargglyaspseraspthrasngluvalalaleuthrpheasn505560ilesertrpglyaspglnlysalametproileleuaspthrleulys65707580alaasnglyilelysaspalathrphepheleuseralasertrpala859095gluarghisproaspvalvalgluargilearglysaspglyhisgln100105110ileglysermetglytyralatyrlysasntyrserglnmetlyslys115120125sergluilelyslysaspleualalysalaarghisserpheglnlys130135140leuglyleuaspaspleuthrleuleuargproprothrglyglnphe145150155160asnlysaspvalleuaspvalalalysglntyrglytyrthrvalval165170175histyrserileasnseraspasptrpthrasnproglyvalglnlys180185190ilevalglnasnvalasngluthrvalasnalaglyaspilevalleu195200205phehisalaseraspseralalysglnthrlysglualaleuproglu210215220ilevalhishisleuargserlysglyleulysasnvalthrvalser225230235240gluleuilealaasnthraspalalyssersergluvallys245250<210>3<211>5<212>prt<213>artificiallysynthesized<400>3argargargargarg15<210>4<211>6<212>prt<213>artificiallysynthesized<400>4hishishishishishis15<210>5<211>8<212>prt<213>artificiallysynthesized<400>5asptyrlysaspaspaspasplys15<210>6<211>8<212>prt<213>artificiallysynthesized<400>6trpserhisproglnpheglulys15<210>7<211>10<212>prt<213>artificiallysynthesized<400>7gluglnlysleuileserglugluaspleu1510<210>8<211>25<212>prt<213>artificiallysynthesized<400>8metlysserlystrpmetserglyleuleuleuvalalavalglyphe151015serphethrglnvalmetvalhisala2025<210>9<211>29<212>dna<213>artificiallysynthesized<400>9gcgggatccgtgaaccatttttatgtgtg29<210>10<211>29<212>dna<213>artificiallysynthesized<400>10gcgacgcgtctactttacttctgaagatt29<210>11<211>234<212>prt<213>artificiallysynthesized<400>11metilelysleuilevalasnalaaspasppheglyleuthrglugly151015thrasntyrglyileileaspglyhisileasnglyleuvalasnser202530thrthrmetmetmetasnmetproglythrgluhisalavalargleu354045alalysgluhisasnthrleuglyvalglyvalhisleuvalleuthr505560alaglylysproleuleuglyaspvalproserleuvalserserasp65707580glyleuphehislysglnservalvaltrpgluglylysileasnpro859095glugluvalgluargglutrpthralaglnileglulyspheleuser100105110tyrglyleuthrprothrhisleuaspserhishishisvalhisgly115120125leuproileleuhisaspvalleuglulysleualaalathrtyrasn130135140valproileargargcysgluglugluargalavalargpropheser145150155160aspvalphetyrseraspphetyralaaspglyvalthrgluasptyr165170175phevallysleulysgluargvalglnglygluglnthrvalgluile180185190metvalhisproalatyrileaspprogluleuvallysargserser195200205tyrvalmetaspargvallysgluleuargileleuthrgluserglu210215220leuprogluglyilegluleuvallysphe225230当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1