利用双光子荧光测定法确定蛋白质结构的方法与流程
交叉引用
本申请要求于2017年5月3日提交的美国临时申请号62/500,912的权益,该临时申请通过引用整体并入本文。
背景技术:
所公开的发明涉及分子检测领域,尤其涉及蛋白质检测和结构确定领域。尽管蛋白质结构确定(并且更普遍地,生物分子结构确定)领域已高度发展,但仍需要灵敏且快速的技术来确定蛋白质结构,比较不同样品或不同时间点的蛋白质结构,以及实时和在溶液中检测蛋白质构象变化(conformationalchange)。大多数关于蛋白质结构和动力学的信息主要来自x射线晶体学和nmr研究,但这些技术是相对劳力和材料密集性的、执行缓慢或仅提供蛋白质结构的静态快照。
二次谐波发生(shg)是一种非线性光学过程,其可以被配置为能够标记有二次谐波活性标签来检测蛋白质和其他生物靶标的结合相互作用和构象变化的表面选择性检测技术(参见,例如美国专利号6,953,694和8,497,073)。迄今为止,这些方法已经用于检测多种生物系统中配体诱导的构象变化,并通过它们在结合至靶蛋白时诱导的构象变化的类型来区分配体(salafsky,j.s.(2001),“'shg-labels'fordetectionofmoleculesbysecondharmonicgeneration”,chemicalphysicsletters342,485-491;salafsky,j.s.(2003),“second-harmonicgenerationasaprobeofconformationalchangeinmolecules”,chemicalphysicsletters381,705-709;salafsky,j.s.(2006),“detectionofproteinconformationalchangebyopticalsecond-harmonicgeneration”,journalofchemicalphysics125;moree,b.等人(2015),“smallmoleculesdetectedbysecondharmonicgenerationmodulatetheconformationofmonomericα-synucleinandreduceitsaggregationincells”,j.biol.chem.290(46);27582-27593;moree等人(2015),“proteinconformationalchangesaredetectedandresolvedsitespecificallybysecond-harmonicgeneration”,biophys.j.109:806-815)。使用shg区分不同类型配体的实例包括区分i型与ii型激酶抑制剂,如伊马替尼和达沙替尼,它们分别与蛋白质结合以诱导失活和活性构象。
如本公开内容中所描述的,双光子荧光(不同的非线性光学技术)还可以单独使用或与诸如二次谐波发生(shg)、和频发生(sfg)或差频发生(dfg)的非线性光学技术组合使用,以解决结构生物学和高通量筛选中的问题,包括但不限于,在溶液中以高通量和可选的方式实时进行生物分子结构确定;构象景观映射;鉴别筛选应用中的假阳性和假阴性;等等。因此,本公开内容的第一方面描述了使用双光子荧光(tpf)测量或双光子荧光与其他非线性光学测量的组合来确定蛋白质结构的方法、装置和系统。
在本公开内容的另一方面,描述了使用shg和相关的非线性光学技术来确定附接于蛋白质的标签的绝对取向,从而允许通过系统地改变标签与蛋白质的附接位点来映射蛋白质结构,或在不同时间点比较不同样品之间的或给定样品的蛋白质结构的方法、装置和系统。在一些实施方式中,这些测量可以包括使用偏振光测量shg信号,例如基线shg信号。在一些实施方式中,这些测量可以包括测量shg与tpf信号的比率,例如基线shg与tpf信号的比率,或非线性光学信号的其他比率。
所公开的方法、装置和系统可以在包括诸如结构生物学和药物发现与开发的基础研究领域的多个领域中具有效用。在许多情况下,如果不使用费力、费时且昂贵的技术如x射线晶体学,可能无法获得此信息。
技术实现要素:
本文公开了用于确定附接在拴系的生物分子(tetheredbiomolecule)上的双光子荧光标签的角度参数的方法,所述方法包括:(a)以定向方式将生物分子附接到平面表面上,其中所述生物分子在已知位点用双光子荧光标签标记;(b)使用第一偏振以第一基频的激发光照射所述附接的生物分子;(c)检测由于步骤(b)中的所述照射而由所述双光子荧光标签生成的光的第一物理性质;(d)使用第二偏振通过所述第一基频的激发光照射所述附接的生物分子;(e)检测由于步骤(d)中的所述照射而由所述双光子荧光标签生成的光的第二物理性质;以及(f)比较步骤(e)中检测到的光的所述第二物理性质与步骤(c)中检测到的光的所述第一物理性质,以确定所述双光子荧光标签相对于所述平面表面的角度参数。
在一些实施方式中,所述第一物理性质是p-偏振光强度ip,所述第二物理性质是s-偏振强度is,并且步骤(f)中的所述比较包括求解以下方程以确定角度参数:
在一些实施方式中,所述方法进一步包括对两个或更多个不同生物分子缀合物系列的每一个重复步骤(a)至(f),其中所述系列中的每个所述生物分子缀合物包括用相同的双光子荧光标签在不同位点标签的所述生物分子,并使用对所述两个或更多个不同生物分子缀合物各自确定的所述角度参数来确定所述生物分子的结构。在一些实施方式中,所述生物分子是蛋白质,并且其中所述两个或更多个不同生物分子缀合物系列各自包括单位点半胱氨酸或蛋氨酸取代。在一些实施方式中,所述生物分子在两个或更多个不同位点被两个或更多个不同的双光子荧光标签标记,其中在被所述两个或更多个不同的双光子荧光标签可能相同或不同的基频的光照射后,在步骤(c)和(e)中同时或依次检测由所述两个或更多个不同的双光子荧光标签各自生成的光的第一物理性质和第二物理性质,并且其中对于所述两个或更多个不同的双光子荧光标签中的每一个,比较在步骤(e)中检测到的所述光的所述第二物理性质与在步骤(c)中检测到的所述光的所述第一物理性质,以确定所述两个或更多个不同的双光子荧光标签中的每一个相对于所述平面表面的角度参数生成的光。在一些实施方式中,所述附接的生物分子还在已知位点用二次谐波(sh)-活性标签、和频(sf)-活性标签或差频(df)-活性标签标记。在一些实施方式中,所述双光子荧光标签和第一二次谐波(sh)-活性标签、和频(sf)-活性标签或差频(df)-活性标签是附接到所述生物分子上相同已知位点的相同标签。在一些实施方式中,所述方法还包括在步骤(c)中同时或依次检测由所述二次谐波(sh)-活性标签、和频(sf)-活性标签或差频(df)-活性标签生成的光的第一物理性质,以及在第二基频的激发光照射下,在步骤(e)中由所述二次谐波(sh)-活性标签、和频(sf)-活性标签或差频(df)-活性标签生成的光的第二物理性质,所述第二基频可以与所述第一基频相同或不同。在一些实施方式中,所述方法还包括比较在步骤(e)中检测到的所述光的所述第二物理性质与在步骤(c)中检测到的所述光的所述第一物理性质,以确定相对于所述平面表面的所述二次谐波(sh)-活性标签、和频(sf)-活性标签或差频(df)-活性标签的角度参数。在一些实施方式中,所述方法包括将一个或多个双光子荧光标签、二次谐波(sh)-活性标签、和频(sf)-活性标签或差频(df)-活性标签或其任何组合的所述角度参数的数据全局地拟合到所述生物分子的结构模型,其中所述结构模型包括有关所述生物分子内所述一个或多个标签的所述已知位点信息。在一些实施方式中,所述方法进一步进一步包括结合x射线晶体学数据、nmr数据或其他为所述生物分子的结构建模提供结构约束的实验数据。在一些实施方式中,所述生物分子是蛋白质,并且并且所述双光子荧光标签或二次谐波(sh)-活性标签、和频(sf)-活性标签或差频(df)-活性标签是非线性-活性的非天然氨基酸。在一些实施方式中,所述非线性-活性的非天然氨基酸是l-anap、aladan或萘的衍生物。在一些实施方式中,非线性-活性部分附接至非明显非线性-活性的非天然氨基酸。在一些实施方式中,光的所述第二物理性质与光的所述第一物理性质不同。在一些实施方式中,光的所述第一物理性质和第二物理性质具有相同的偏振,但幅度或强度不同。在一些实施方式中,光的所述第一物理性质和所述第二物理性质具有不同的偏振。在一些实施方式中,所述照射步骤包括调节所述激发光的所述偏振。在一些实施方式中,所述激发光的第一偏振状态包括相对于其入射平面的p-偏振,并且所述激发光的第二偏振状态包括相对于其入射平面的s-偏振。在一些实施方式中,步骤(c)和(e)中的所述检测包括调节由到达检测器的所述双光子荧光标签或二次谐波(sh)-活性标签、和频(sf)-活性标签或差频(df)-活性标签生成的所述光的所述偏振。在一些实施方式中,光的所述第一物理性质和第二物理性质是强度或偏振。在一些实施方式中,使用低数值孔径针孔配置检测由双光子荧光标签生成的光,而不使用会聚透镜。在一些实施方式中,所述低数值孔径针孔放置于所述平面表面上所述激发光入射到所述平面表面上的点的正上方或正下方。在一些实施方式中,所述平面表面包括受支承脂双层,并且所述生物分子附接至或插入所述受支承脂双层。在一些实施方式中,使用全内反射将所述激发光导向所述平面表面。在一些实施方式中,所述双光子荧光标签也是二次谐波(sh)-活性标签、和频(sf)-活性标签或差频(df)-活性,并且还包括通过以下步骤确定标签的角度参数:(g)在用第二基本频率的激发光照射后,同时或依次检测附接在所述附接生物分子上的所述二次谐波(sh)-活性标签、和频(sf)-活性标签或差频(df)-活性标签的光强度,所述第二基频的激发光可以与所述第一基频的所述激发光相同或不同,并且其中使用以下方式执行检测:(i)所述激发光的第一偏振状态;以及(ii)所述激发光的第二偏振状态;(h)通过计算在步骤(c)(i)和(c)(ii)中检测到的所述光强度的比率,确定相对于基底表面法线的二次谐波(sh)活性、和频(sf)-活性或差频(df)-活性标签的角度参数;(i)对与所述二次谐波(sh)-活性标签、和频(sf)-活性标签或差频(df)-活性标签的角度参数有关的方程和计算出的所述二次谐波(sh)、和频(sf)或差频(df)光的所述光强度比进行积分,以确定满足所述二次谐波(sh)、和频(sf)或差频(df)方程的角度参数值对;(j)对与所述二次谐波(sh)-活性标签、和频(sf)-活性标签或差频(df)-活性标签的角度参数有关的方程和计算出的所述二次谐波(sh)、和频(sf)或差频(df)光的所述光强度比进行积分,以确定满足所述二次谐波(sh)、和频(sf)或差频(df)方程的角度参数值对;以及(k)确定在步骤(i)和(j)中鉴别的所述角度参数值对的所述交点,以确定同时满足所述双光子荧光和所述二次谐波(sh)、和频(sf)或差频方程的唯一的角度参数值对。在一些实施方式中,生物分子附接于所述平面表面,使得附接于所述生物分子的所述双光子荧光标签的取向分布的宽度小于或等于35度。在一些实施方式中,所述角度参数包括平均倾斜角、取向分布宽度或其成对组合。
本文还公开了用于检测生物分子构象变化的方法,所述方法包括:a)以定向的方式将所述生物分子附接到平面表面上,其中所述生物分子用双光子荧光标签标记;b)使用第一偏振和第二偏振,以第一基频的激发光照射所述附接的生物分子;c)在步骤(b)中,检测由于所述第一和第二偏振光的所述照射而由所述双光子荧光标签生成的光的第一物理性质和光的第二物理性质;d)使所述附接的生物分子(i)与已知配体接触,(ii)与候选结合配偶体(bindingpartner)接触,或(iii)经受实验条件改变;e)使用所述第一偏振和所述第二偏振,以所述第一基频激发光照射所述附接的生物分子;f)在步骤(e)中,检测由于所述第一和第二偏振光的所述照射而由所述双光子荧光标签生成的光的第三物理性质和光的第四物理性质;以及;以及(f)比较步骤(f)中检测到的光的所述第三物理性质和第四物理性质的比率与步骤(c)中检测到的光的所述第一物理性质和第二物理性质的比率,其中光的所述物理性质的比率的变化表示所述生物分子发生了构象变化。
在一些实施方式中,使用数值孔径小于或等于0.2的针孔检测设备检测双光子荧光的物理性质。在一些实施方式中,所述数值孔径在约0.01至约0.2之间。在一些实施方式中,检测双光子荧光的所述物理性质而无需使用透镜。在一些实施方式中,所述双光子荧光标签也具有二次谐波、和频或差频活性,并且其中由于使用所述第一偏振和第二偏振以第二基频的光进行照射,在检测所述双光子荧光的所述物理性质时,连续或同时检测二次谐波、和频或差频光的物理性质。在一些实施方式中,在步骤(f)中比较的比率包括双光子荧光的所述物理性质与二次谐波、和频或差频光的所述物理性质的比率。在一些实施方式中,所述第二基频与所述第一基频相同。在一些实施方式中,所述第一和第二偏振包括s-偏振和p-偏振。在一些实施方式中,所述生物分子是蛋白质分子。在一些实施方式中,所述蛋白质分子是药物靶标。在一些实施方式中,所述已知配体是已知药物或所述候选结合配偶体是药物候选物。在一些实施方式中,所述双光子荧光标签在一个或多个工程化的半胱氨酸残基处附接于所述蛋白质分子。在一些实施方式中,所述双光子荧光标签是吡啶恶唑(pympo)。在一些实施方式中,所述双光子荧光标签是已被掺入所述蛋白质分子中的非线性-活性的非天然氨基酸。在一些实施方式中,所述非线性的非天然氨基酸是l-anap、aladan或萘的衍生物。在一些实施方式中,使用全内反射将所述激发光传递到所述平面表面。在一些实施方式中,所述生物分子通过插入或拴系在受支承脂双层而附接于所述平面表面。
本文公开了用于筛选候选结合配偶体以鉴别调节靶分子所述构象的结合配偶体的方法,所述方法包括:(a)将所述靶分子拴系在基底表面,其中所述靶分子用双光子荧光标签标记,所述双光子荧光标签附接到与结合配偶体接触时发生构象变化的所述靶分子的一部分,并且其中所述拴系的靶分子在所述基底表面具有净取向(netorientation);(b)以第一基频的激发光照射所述拴系的靶分子;(c)检测由所述双光子荧光标签生成的光的第一物理性质,以生成基线信号;(d)使所述拴系的靶分子与所述一个或多个候选结合配偶体依次并分别接触;(e)对于所述一个或多个候选结合配偶体中的每一个,检测由所述双光子荧光标签响应于所述第一基频所述激发光的照射而生成的光的第二物理性质;以及(f)比较所述一个或多个候选结合配偶体中每一个的所述第二物理性质与所述第一物理性质,其中给定候选结合配偶体的所述第二物理性质的值相对于所述第一物理性质的变化指示所述候选结合配偶体调节所述靶分子的所述构象
在一些实施方式中,光的所述第一物理性质和第二物理性质包括所述激发光在两种不同偏振下的所述光强度,并且其中步骤(f)包括确定所述两种光强度的比率,其中所述比率的变化指示所述候选结合配偶体调节所述靶分子的所述构象。在一些实施方式中,所述靶分子也用二次谐波(sh)-活性标签、和频(sf)-活性标签或差频(df)活性标签进行标记。在一些实施方式中,所述双光子荧光标签和所述二次谐波(sh)-活性标签、和频(sf)-活性标签或差频(df)-活性标签是相同的标签部分。在一些实施方式中,所述方法进一步包括以下步骤:(g)在执行步骤(c)的同时或之后,在用第二基频的激发光照射后,检测由所述二次谐波(sh)-活性标签、和频(sf)-活性标签或差频(df)-活性标签生成的光的第一物理性质,其中所述第二基频可以与所述第一基频相同或不同;(h)在执行步骤(e)的同时或之后,在用所述第二基频的激发光照射后,检测由所述二次谐波(sh)-活性标签、和频(sf)-活性标签或差频(df)-活性标签生成的光的第二物理性质;以及(i)比较所述一个或多个候选结合配偶体中每一个由所述二次谐波(sh)-活性标签、和频(sf)-活性标签或差频(df)-活性标签生成的所述第二物理性质与由所述二次谐波(sh)-活性标签、和频(sf)-活性标签或差频(df)-活性标签生成的所述第一物理性质,其中给定的候选结合配偶体的所述第二物理性质的值相对于所述第一物理性质的值的变化进一步表明,所述候选结合配偶体调节所述靶分子的所述构象。在一些实施方式中,光的所述第一物理性质和第二物理性质包括所述激发光在两种不同偏振下的所述光强度,并且其中步骤(i)包括确定所述两种光强度的比率,其中所述比率的变化指示所述候选结合配偶体调节所述靶分子的所述构象。在一些实施方式中,所述激发光以这样的方式被引导到所述基底表面,使得其从所述表面被全内反射。在一些实施方式中,使用在所述第一基频的所述激发光入射到基底表面的点处位于所述基底表面的正上方或正下方的针孔孔径来收集双光子荧光。在一些实施方式中,收集双光子荧光而无需使用会聚透镜。在一些实施方式中,所述数值孔径的所述数值孔径在0.01至0.2之间。在一些实施方式中,所述非线性-活性标签包括吡啶恶唑(pympo)部分、6-溴乙酰基-2-二甲基氨基萘(badan)部分或6-丙烯酰基-2-二甲基氨基萘(acrylodan)部分。在一些实施方式中,所述靶分子是包括遗传掺入的his标记物的蛋白质。在一些实施方式中,所述his标记物包括6x-his标记物、7x-his标记物、8x-his标记物、9x-his标记物、10x-his标记物、11x-his标记物或12x-his标记物。在一些实施方式中,通过使用全内反射,用所述第一基频的光照射所述拴系的靶分子。
本文公开了用于比较由仿制药物或药物候选物和参考药物在靶蛋白的所述结构中诱导的所述构象变化的方法,其中所述靶蛋白用非线性-活性标签标记并将其拴系在所述界面上,使所述界面有净取向,所述方法包括:a)使所述靶蛋白与所述参考药物接触,其中所述靶蛋白以特定方式与所述参考药物或品牌药物相互作用;b)通过使用表面选择技术测定由所述非线性-活性标签产生的第一信号或信号变化,来检测所述靶蛋白与所述参考药物之间的相互作用,其中所述第一信号或信号变化指示,对所述参考药物具有特异性的所述靶蛋白的所述结构的构象变化;c)使所述靶蛋白与所述仿制药物或药物候选物接触,其中所述靶蛋白以特定方式与所述仿制药物或药物候选物相互作用;以及d)通过使用表面选择技术测定由所述非线性-活性标签产生的第二信号或信号变化,来检测所述靶蛋白与所述仿制药物或药物候选物之间的相互作用,其中所述第二信号或信号变化指示,对所述仿制药物或药物候选物具有特异性的所述靶蛋白的所述结构的构象变化;以及e)比较所述第二信号或信号变化和所述第一信号或信号变化,以确定所述仿制药物或药物候选物在所述靶蛋白中诱导的所述构象变化是否与所述参考药物诱导的变化相同或基本相同。
在一些实施方式中,所述靶蛋白是细胞表面受体或抗原。在一些实施方式中,所述参考药物是单克隆抗体(mab)。在一些实施方式中,所述仿制药物或药物候选物选自小分子化合物、非抗体抑制肽、抗体及其任何组合。在一些实施方式中,所述仿制药物或药物候选物是单克隆抗体(mab)。在一些实施方式中,所述仿制药物是生物仿制药。在一些实施方式中,实时检测所述靶蛋白的所述结构的所述构象变化。在一些实施方式中,所述非线性-活性标签通过所述靶蛋白的所述表面的一个或多个巯基基团与所述靶蛋白结合。在一些实施方式中,所述一个或多个巯基是工程化的巯基。在一些实施方式中,所述非线性-活性标签是二次谐波(sh)-活性标签或双光子荧光标签。在一些实施方式中,所述非线性-活性标签是选自pympo马来酰亚胺、pympo-nhs、pympo琥珀酰亚胺酯、badan和acrylodan的二次谐波(sh)-活性标签。在一些实施方式中,所述非线性-活性标签是非天然氨基酸。在一些实施方式中,所述非天然氨基酸是l-anap、aladan或萘的衍生物。在一些实施方式中,基于诱导的构象变化的所述比较,结合至少从第二结构表征或功能测定技术获得的结构或功能数据,确定生物相似性。在一些实施方式中,所述至少第二结构表征或功能测定技术选自圆二色性、x射线晶体学、生物学测定、结合测定、酶测定、基于细胞的测定、细胞增殖测定、基于细胞的报告测定和动物模型研究。
本文公开了用于比较两个或更多个蛋白质样品的方法,所述方法包括:a)提供从相同的蛋白质生产过程的不同批次(run),或标称(nominally)生产相同蛋白质的不同蛋白质生产过程,在蛋白质生产过程相同步骤的不同时间,在蛋白质生产过程的不同步骤收集的两个或更多个蛋白质样品;b)将来自所述一个或多个蛋白质样品的所述蛋白质拴系在光学界面的一个或多个非连续区域中,其中来自每个样品的所述拴系蛋白质用非线性-活性标签标记,并在所述光学界面处具有净取向;c)测定所述一个或多个拴系蛋白质样品中每一个的基线非线性光学信号,所述信号是在用基频光照射所述非线性-活性标签后生成的;以及d)将所述一个或多个栓系蛋白样品的所述测量的基线非线性光学信号相互比较,或与参考样品的基线非线性光学信号进行比较,其中对于所述一个或多个固定化蛋白质样品测得的所述基线非线性光学信号,或对于所述一个或多个蛋白质样品测得的基线非线性光学信号与参考样品的所述基线非线性光学信号之间的差异小于指定百分比,指示所述一个或多个蛋白质样品或所述参考样品的所述蛋白质具有相同的结构。
在一些实施方式中,在蛋白质生产过程的终点收集所述一个或多个蛋白质样品,并将步骤(d)中的所述比较用于所述蛋白质产物的质量控制。在一些实施方式中,在蛋白质生产过程的一个或多个步骤中收集所述一个或多个蛋白质样品,并将步骤(d)中的所述比较用于优化所述蛋白质生产过程。在一些实施方式中,从标称生产相同蛋白质的不同蛋白质生产过程中收集所述一个或多个蛋白质样品,并将步骤(d)中的所述比较用于证明生物相似性。在一些实施方式中,所述光学界面包括选自玻璃表面、熔融二氧化硅表面或聚合物表面的表面。在一些实施方式中,所述光学界面包括受支承脂双层。在一些实施方式中,所述受支承脂双层还包含ni/nta-脂质分子。在一些实施方式中,所述一个或多个蛋白质样品的所述蛋白质包含his-标记物。在一些实施方式中,实时监测所述基线非线性光学信号或其变化。在一些实施方式中,所述非线性-活性标签通过所述蛋白质的所述表面的一个或多个巯基与所述蛋白质结合。在一些实施方式中,所述一个或多个巯基是工程化的巯基。在一些实施方式中,所述非线性-活性标签是二次谐波(sh)-活性标签。在一些实施方式中,通过使所述固定化或拴系的蛋白质与本身具有shg-活性的肽、拟肽或其他配体接触来对其进行标记,从而使所述shg-活性配体结合到所述固定化或拴系的蛋白质上。在一些实施方式中,所述非线性-活性标签是选自pympo马来酰亚胺、pympo-nhs、pympo琥珀酰亚胺酯、badan和acrylodan的二次谐波(sh)-活性标签。在一些实施方式中,所述非线性-活性标签是已遗传掺入所述一个或多个蛋白质样品的所述蛋白质中的非天然氨基酸。在一些实施方式中,所述非天然氨基酸是l-anap、aladan或萘的衍生物。在一些实施方式中,所述非线性-活性标签既是二次谐波(sh)-活性的又是双光子荧光的,并且其中步骤(c)中的所述测定还包括测定基线二次谐波信号和基线双光子荧光信号。在一些实施方式中,步骤(d)的所述比较还包括将所述一个或多个拴系蛋白样品的二次谐波与双光子荧光基线信号的比率与参考样品的二次谐波和双光子荧光基线信号的比率进行比较,其中差异小于指定百分比指示所述一个或多个蛋白质样品或所述参考样品的所述蛋白质具有相同的结构。本文公开了用于检测附接到拴系的生物分子上的双光子荧光标签的双光子荧光的方法,所述方法包括:(a)以定向方式将生物分子附接到平面表面上,其中所述生物分子在已知位点用双光子荧光标签标记;(b)使用第一偏振以基频的激发光照射所述附接的生物分子;(c)检测由于步骤(b)的所述照射而由所述双光子荧光标签生成的光的物理性质,其中使用低数值孔径针孔配置检测由所述双光子荧光标签生成的所述光,而不使用会聚透镜。在一些实施方式中,所述低数值孔径针孔放置于所述平面表面上所述激发光入射到所述平面表明上的点的正上方或正下方。在一些实施方式中,所述平面表面包括受支承脂双层,并且所述生物分子附接至或插入所述受支承脂双层。在一些实施方式中,使用全内反射将所述激发光导向所述平面表面。在一些实施方式中,所述低数值孔径针孔的数值孔径在0.01和0.2之间。
本文还公开了用于建立生物仿制药药物候选物和参考药物的结构等效性的方法,所述方法包括:a)使用相同的标签反应,以非线性-活性标签标记生物仿制药药物候选物和参考药物;b)将非线性-活性标签的生物仿制药药物候选物和参考药物拴系在界面上,以使其在所述界面上具有净取向;c)在以第一基频的光照射后,测定由非线性-活性标签为生物仿制药药物候选物和参考药物生成的二次谐波的物理性质;d)可选地,在以第二基频的光照射后,测定由非线性-活性标签为生物仿制药和参考药物生成的双光子荧光的物理性质;e)比较对于生物仿制药药物候选物和参考药物的二次谐波光的物理性质,其中生物仿制药药物候选物和参考药物的物理性质在统计学上的显著差异指示它们在结构上不等效;以及f)可选地,计算对于生物仿制药药物候选物和参考药物的二次谐波光的物理性质与所测量的双光子荧光的物理性质的比率,其中对于生物仿制药药物候选物和参考药物计算的比率具有统计学上的显著差异,指示它们在结构上不等效。
在一些实施方式中,所述第一基频与所述第二基频相同。在一些实施方式中,所述标签反应包括所述非线性-活性标签与所述生物仿制药药物候选物和参考药物上的天然官能团的共价缀合。在一些实施方式中,所述天然官能团包括天然胺基、天然羧基或天然巯基。在一些实施方式中,所述标签反应包括所述非线性-活性标签与所述生物仿制药药物候选物和参考药物上的遗传工程官能团的共价缀合。在一些实施方式中,所述遗传工程的官能团包括遗传工程胺基、遗传工程羧基或遗传工程巯基。在一些实施方式中,所述标签反应包括已知与所述参考药物的特定区域结合的非线性-活性标签肽之间的非共价相互作用。在一些实施方式中,所述非线性-活性标签肽包含已知结合单克隆抗体fc区域的肽。在一些实施方式中,所述非线性-活性标签包括吡啶恶唑(pympo)部分、6-溴乙酰基-2-二甲基氨基萘(badan)部分或6-丙烯酰基-2-二甲基氨基萘(acrylodan)部分。在一些实施方式中,所述非线性-活性标签的生物仿制药药物候选物和参考药物被拴系到相同的界面。在一些实施方式中,所述非线性-活性标签的生物仿制药药物候选物和参考药物被拴系到不同的界面。在一些实施方式中,使用固定在界面上的蛋白质a或蛋白质g分子将所述非线性-活性标签的生物仿制药药物候选物和参考药物拴系到界面上。在一些实施方式中,所述界面包括受支承脂双层,并且其中所述非线性-活性标签的生物仿制药药物候选物和参考药物被拴系或嵌入在所述受支承脂双层中。在一些实施方式中,所述界面包括受支承脂双层,并且其中使用结合到包含ni-nta部分的双层脂的遗传掺入的his-标记物将非线性-活性标签的生物仿制药药物候选物和参考药物栓系到受支承脂双层。在一些实施方式中,遗传掺入的his标记物包括6x-his标记物、7x-his标记物、8x-his标记物、9x-his标记物、10x-his标记物、11x-his标记物或12x-his标记物。在一些实施方式中,通过使用全内反射,以第一基频的光照射所述非线性-活性标签的生物仿制药药物候选物和参考药物。在一些实施方式中,使用在第一基频的激发光入射到界面上的点处位于界面正上方或正下方的针孔孔径来收集双光子荧光。在一些实施方式中,收集双光子荧光而无需使用会聚透镜。在一些实施方式中,由p值小于0.05指示生物仿制药药物候选物和参考药物所测得的光的物理性质,或生物仿制药药物候选物和参考药物计算出的比率在统计学上有显著差异。
援引并入
本说明书中提到的所有出版物、专利及专利申请均通过引用以其全文并入本文,其程度如同特别地且单独地指出每个单独的出版物、专利或专利申请均通过引用以其全文而并入。如果本文的术语与并入的参考文献中的术语之间存在冲突,则以本文的术语为准。
附图说明
本发明的新颖特征在所附的权利要求书中具体阐述。通过参考以下对其中利用了本发明原理的说明性实施方式加以阐述的详细描述以及附图,将获得对本发明的特征和优点的更好的理解,在附图中:
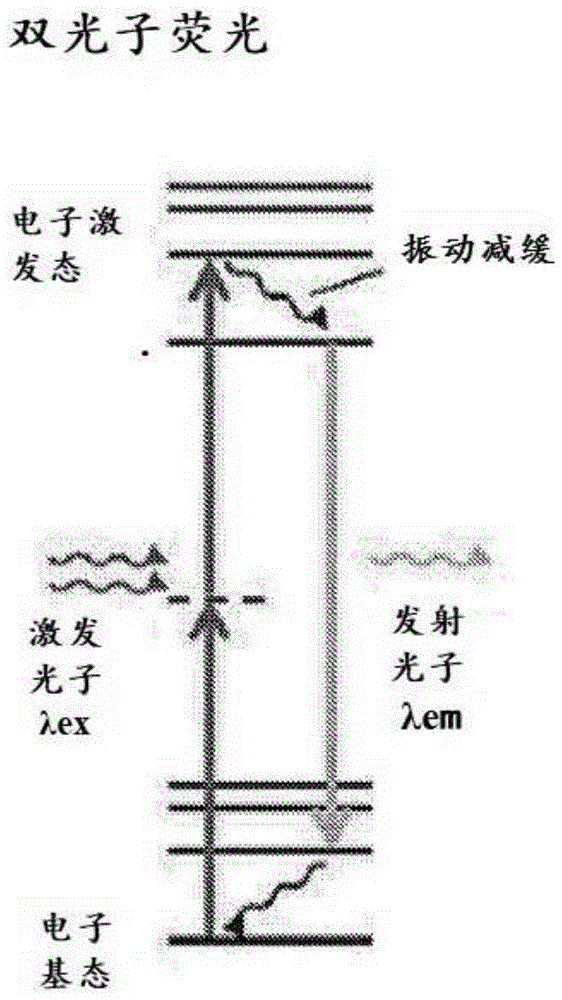
图1a-图1c提供了双光子荧光(图1a)、单光子荧光(图1b)和二次谐波(图1c)的能级图示意图。
图2图示了实验室参照系(由x、y和z轴定义)与分子参照系(由x’、y’和z’轴定义)之间的关系。对于一些非线性-活性分子,超极化率张量(α(2))可以由分子参照系中的单分量来控制,即α(2)=α(2)z’z’z’。
图3图示了由配体的结合诱导的蛋白质(用二次谐波-活性标签、双光子荧光标签或其他非线性-活性部分标记)的构象变化及其对标签相对于蛋白质所附接的光学界面的z轴法线的取向的影响。
图4提供了装置的非限制性示例的示意图,该装置包括围绕基底表面区域的电极的图案化阵列,该基底表面区域用于形成受支承脂双层。
图5提供了使用表面选择性非线性光学技术来执行高通量结构确定的装置的非限制性示例的示意图,其中,以玻璃底微孔板形式结合至基底或与基底成一体的半球形棱镜阵列用于提供激发光与基底顶表面的良好光学耦合。
图6图示了用于根据非线性光学检测来确定生物分子(例如蛋白质或其他生物实体)的结构或构象变化的高通量分析系统的系统架构的一个非限制性示例。
图7示出了低-na检测方案的示意图,其中使用直径0.5mm的纤维来收集发射的荧光。在tpf中,荧光在所有方向上发射(浅阴影),但可以通过使用几何形状来选择总立体角的一小部分(深阴影),其中光纤半径(0.5mm)与光纤(7.5mm)之间的距离的比率很小,从而产生低-na检测器。
图8示出了用于利用非线性光学检测对生物分子的结构或构象变化进行分析的光学设置的一个非限制性示例的示意图。
图9示出了描绘利用棱镜来以适当的入射角引导激发光从而使激发光在基底的顶表面经历全内反射的示意图示。向棱镜右侧的两条虚线指示出反射的激发光的光路以及当非线性-活性物质拴系到基底表面时在该基底表面生成的非线性光学信号。基底可选地连接到x-y平移台的致动器,以便在测量之间重新定位。棱镜的顶表面与基底的底表面之间的曲线指示出用于确保棱镜与基底之间的高光耦合效率的折射率匹配流体薄层(未按比例绘制)的存在。
图10a-图10b图示了具有用于提供激发光与基底顶表面的良好光耦合的集成式棱镜阵列的微孔板。这样的装置对进行蛋白质和其他生物分子的高通量结构确定可能是有用的。图10a:顶轴侧视图。图10b:底轴侧视图。
图11a-图11b示出了图10a-图10b中所示的微孔板装置的分解图。图11a:顶轴侧视图。图11b:底轴侧视图。
图12图示了利用图11a-图11b中图示的设计概念经由全内反射而将激发光耦合到基底表面的入射光路和出射光路。
图13图示了经配置用于控制本文公开的系统的操作的计算机系统。
图14是图示可结合本发明示例实施方式使用的计算机系统的第一示例架构的框图。
图15是示出一个具有多个计算机系统、多个蜂窝电话和个人数字助理以及网络附加存储(nas)的网络实施方式的视图。
图16是根据示例实施方式,利用共享虚拟地址存储器空间的多处理器计算机系统的框图。
图17a-图17b提供了根据所公开的方法用于进行双光子荧光(tpf)和二次谐波发生(shg)测定的光学装置的非限制性示意图。图17a:俯视图。图17b:侧视图。
图18示出了通过拟合高斯取向分布的双光子荧光偏振强度比测量值(绿线)和shg偏振强度比测量值(蓝线)确定的,被拴系到受支承脂双层的蛋白二氢叶酸还原酶(dhfr)的m16c突变体的平均取向倾斜角-取向分布宽度的图。两条曲线的交点定义了唯一的角度参数。
图19示出了在1μmtmp存在下,通过拟合高斯取向分布的双光子荧光偏振强度比测量值(绿线)和shg偏振强度比测量值(蓝线)确定的,被拴系到受支承脂双层的蛋白二氢叶酸还原酶(dhfr)的m16c突变体的平均取向倾斜角-取向分布宽度的图。两条曲线的交点定义了唯一的角度参数。
图20示出了实施例1中使用的所有单半胱氨酸dhfr突变体的平均取向和分布宽度的角度数据。在具有和不具有tmp的情况下,每个突变体的取向平均角均是取向分布的函数,这表明当标签放置在整个蛋白质的不同位置时会采用大范围的角度。
图21示出了在添加tmp后每个单半胱氨酸突变体的取向平均角的变化和取向分布的变化。
图22示出了dhfr的两个晶体结构的叠加。棕褐色结构是载脂蛋白形式,而蓝色结构与mtx结合。在图中标记了可以与半胱氨酸交换的残基,并示出了其侧链相对于肽骨架的投影。可以将在两个晶体结构之间观察到的侧链取向的变化与通过该方法预测的取向平均角和取向分布的变化进行比较。
具体实施方式
本文公开的方法、装置和系统通常涉及使用二次谐波发生(shg)、和频发生(sfg)、差频发生(dfg)和/或双光子荧光(tpf)的生物分子检测和表征领域,包括生物分子的结构和动力学的确定。描述了用于确定在一个或多个位点附接于蛋白质或其他生物分子的非线性-活性标签,例如shg标签或tpf标签(也称为“探针”)的相对和/或绝对取向的方法,以及用于确定分子结构或构象的方法。还描述了在不同时间点比较来自不同样品或相同样品的蛋白质或其他生物分子结构的方法。
在本公开内容的第一方面,描述了单独或与shg、sfg或dfg测量组合使用双光子荧光(tpf)测量来确定蛋白质结构的方法、装置和系统。通常,所公开的使用tpf来确定蛋白质或其他生物分子的结构或检测其构象变化的方法包括:(a)将生物分子附接到平面上,其中该生物分子在已知位点用双光子荧光标签标记;(b)使用第一偏振以第一基频的激发光照射附接的生物分子;(c)检测有步骤(b)的照射而由双光子荧光标签生成的光的第一物理性质;(d)使用第二偏振以第一基频的激发光照射附接的生物分子;(e)检测由于步骤(d)的照射而由双光子荧光标签生成的光的至少第二物理性质;以及(f)比较步骤(e)中检测到的光的至少第二物理性质与步骤(c)中检测到的光的第一物理性质,以确定双光子荧光标签相对于表面平面的取向。在该方法的优选实施方式中,光的第一和第二物理性质是通过低数值孔径(低-na)检测分别从在相对于表面平面的p-偏振和s-偏振的基本激发光下附接于平面表面并经历全内反射(tir)的单-位点-特异性标记生物分子测量的tpf或shg强度;该优选实施方式的步骤(f)包括获取在p-偏振和s-偏振激发下测得的tpf或shg强度的比率,例如,rtpf=ip/is,其中rtpf表示p-偏振或s-偏振激发下tpf强度的比率。
大多数生物分子必须用非线性-活性标签标记,以使其自身具有非线性-活性。对于诸如蛋白质生物分子可以使用双光子荧光(tpf)活性探针、二次谐波发生(shg)-活性探针,或者可选地使用既具有tpf-活性又具有shg-活性的探针在一个或多个胺或巯基位点(例如一个或多个赖氨酸或半胱氨酸残基)处共价标记该蛋白,以赋予非线性光学活性。如下面将更详细描述的,也可以采用替代的标签策略。
如上所述,在本公开内容的方法中,可单独使用双光子荧光(tpf)测量,或作为shg测量技术的正交或互补过程,用于确定蛋白质结构或检测构象变化。与shg不同,tpf不是相干技术,因此不需要标记分子的净平均取向即可产生信号。在一些实施方式中,非线性光学信号测量的比率,例如shg与tpf信号的比率,可以用于蛋白质结构确定或构象变化的检测。
在一些实施方式中,所公开的方法的照射(激发)步骤可以包括调节至少一个基频激发光的偏振。在其他实施方式中,激发光的频率可以在实验之间改变。在一些实施方式中,如将在下面更详细地讨论的,以这样的方式将用于执行tpf和/或shg、sfg或dfg测量的激发光引导至表面,使其从表面被全内反射。在一些实施方式中,激发光的第一偏振状态相对于其入射平面包括p-偏振,并且激发光的第二偏振状态相对于其入射平面包括s-偏振。
在一些实施方式中,使用tpf测量值(单独或与shg、sfg或dfg测量组合使用)确定标签的生物分子中的结构参数、构象状态和/或检测构象变化包括测定非线性光学信号的物理性质或物理性质的变化(例如,信号强度或偏振的变化)或非线性光学信号的物理性质的比率或比率的变化(例如,shg与tpf信号强度的比率)。在一些实施方式中,在使标签的生物分子与配体接触或使其经受一些其他环境变化之前,测定光的第一物理性质,并且在使标签生物分子与配体接触或使其经受其他一些环境变化之后,测定至少第二物理性质。在一些实施方式中,光的至少第二物理性质与光的第一物理性质相同。在一些实施方式中,光的至少第二物理性质与光的第一物理性质不同。在一些实施方式中,可以进行多次测定,其中激发光或检测到的光的偏振、幅度或强度或其任合组合发生改变。在一些实施方式中,该方法进一步包括将提供蛋白质结构限制的x射线晶体学数据、nmr数据或其他实验数据结合到蛋白质分子(或其他生物分子)的结构模型中。
本文公开的基于tpf的方法的第一关键组分是使用全内反射(tir)激发产生tpf信号。tir激发具有两个重要优势:(i)它产生确定的正交偏振状态(p和s),并且(ii)它仅在与标记蛋白质拴系的表面相邻的较弱渐逝区域(约100nm)中生成tpf。这两个优势可以简化理论分析,并显著减少计算探针(或标签)的角度信息时的误差,这对于准确测量标签生物分子(如蛋白质)的准确结构测定。先前描述的绝大多数tpf工作采用了落射荧光显微术,其涉及通过垂直于表面平面的光束进行激发。落射荧光显微术便于成像,但会导致生成背景tpf,并在分析倾斜角和其他方向信息时带来很大的不确定性。对于这样的结构分析,其要求高度自信地知道光学系统对发射光子的收集效率。此外,当使用落射荧光激发时,不可能激发在z方向上具有偏振分量(p-偏振)的栓系标签分子。
本文公开的基于tpf的方法的第二关键组分是收集双光子荧光而无需使用透镜(与显微术的情况不同),因为这将需要详细而精确地了解透镜数值孔径(na),以便将测得的荧光强度与探针取向分布相关联。相反,所公开的方法利用了低-na针孔,该针孔位于激发光聚焦点的正上方或正下方,并且平行于样品平面(即,以垂直于穿过焦点的表面的轴(z-轴)为中心)。然后可以使用光电倍增管或其他合适的检测器检测穿过针孔的光。因此,本发明的主要目的之一是通过采用tir激发和低-na针孔检测,来实现探针取向分布的高精度测定(从而推断关于蛋白质结构和构象的信息),其中针孔在激发点处位于样品的正上方或正下方。
所公开的基于tpf的方法的第三关键组分是使用平面样品形式,其中生物分子基本上被限制在单个平面上,如与单层、受支承脂双层膜等一起出现。与现有技术方法相比,本发明的这一特征既大大简化了分析,又允许以显著更高的精度确定诸如平均倾斜角和取向宽度(例如,假设为高斯分布)的取向信息。
最后,特别是对于需要角度信息的情况,所公开的基于tpf的方法的第四关键组分,是使用至少一个tpf-活性探针,该探针掺入到感兴趣的生物分子中,其相对于表面取向相对狭窄地分布,例如,假设高斯分布,平均倾斜角的标准偏差小于或等于约35度。如下例所示,我们可以通过将蛋白质通过6xhis-标记物拴系到包含捕获剂ni-nta脂质,例如1,2-二油酰基-3-甘油3-[(n-(5-氨基-1-羧基戊基)亚氨基二乙酸)琥珀酰](镍盐)的受支承脂双层来实现探针的相对狭窄的定向分布。
在一些实施方式中,可以通过在两组或更多组不同的实验条件下进行测定来促进基于tpf测量单独或与shg、sfg或dfg测定组合的结构确定。例如,在一些实施方式中,使用his-标记物将蛋白质(或其他生物分子)附接至表面或受支承脂双层。在一些实施方式中,第一组实验条件包括使用附接于蛋白质n端的his-标记物来拴系蛋白质分子,并且至少第二组实验条件包括使用附接于c端的his-标记物来拴系蛋白质分子。或者,在一些实施方式中,第一组实验条件可包括在第一测定缓冲液的存在下拴系蛋白质分子(或将拴系的蛋白质暴露于第一测定缓冲液),并且至少第二组实验条件可包括在不同于第一测定缓冲液的至少第二测定缓冲液的存在下拴系蛋白质分子(或将拴系的蛋白质暴露于至少第二测定缓冲液)。在一些实施方式中,如上所述,第一组实验条件与至少第二组实验条件之间的差异可以包括使拴系的蛋白质分子与至少已知结合并诱导蛋白质分子构象变化的第一配体接触。下面将更详细地描述可用于促进基于单独或与shg、sfg或dfg测量组合使用的tpf测量的结构来确定的不同组实验条件的非限制性示例。使用不同实验条件的目的是产生其中取向分布在实验室框架中发生变化的样品,从而为在给定位点标签的生物分子提供独立的角度测量集,即在表面法线轴(z轴)上具有不同的探针跃迁偶极矩投影。这使更多方程可以确定生物分子参照系中的构象分布(景观)。
在本公开内容的另一方面,描述了使用shg或相关的非线性光学基线信号(例如,sfg和dfg基线信号)来在不同时间点比较来自不同样品或来自给定样品的蛋白质结构。通常,这些方法可以包括:(i)使用相同的标签反应和反应条件,用非线性-活性标签来标记一个或多个样品中的蛋白质,(ii)将标签的蛋白质拴系到界面(例如基底表面)使其在界面上具有净取向,(iii)在用基频光照射后测定非线性-活性标签生成的光的物理性质,以及(iv)比较在不同时间点为不同样品测量的或为给定样品测量的非线性光学信号。例如,可以执行这样的基线信号测量以:(i)比较不同批次纯化蛋白质之间的蛋白质结构,(ii)监测用于表达、生产和/或纯化蛋白质产品的生物反应器或制造过程中不同步骤的蛋白质结构变化,或(iii)在蛋白质与不同的试剂接触或经受不同的实验条件后监测蛋白质的稳定性。该过程利用基线shg或其他非线性光学信号的变化来测定已被非线性-活性部分标签的蛋白质的变性程度。在一些实施方式中,这些比较可以仅依赖于shg、sfg或dfg基线信号的测定。在优选的实施方式中,可以使用shg、sfg或dfg基线信号与针对相同样品测得的tpf基线信号的比率来进行这些比较,其中同时或连续测量shg、sfg或dfg基线信号与tpf基线信号。使用shg、sfg或dfg基线信号与tpf基线信号的比率允许使shg、sfg或dfg基线信号标准化,并校正拴系在基底上的标签蛋白分子的表面密度变化,该变化可能在各孔之间存在,也可能在以表面选择性的方式激发非线性活性标记的实验之间存在。这样的结构比较在各种药物发现和开发应用程序(和其他领域)中具有潜在的实用性,包括但不限于,生物药物的蛋白质稳定性的监测、制造过程监测和质量控制以及生物药物候选物与参考药物之间生物相似性的证明。
除了公开的用于确定生物分子结构/构象的非线性光学方法,以及用于比较在不同时间点的两个或更多个不同样品或相同样品的生物分子结构的非线性光学方法外,还描述了促进以高通量形式执行所公开的方法和/或其实现以供分析分子取向或分子结构的装置和系统。在本公开内容的一些方面,描述了响应于使生物分子与一种或多种测试分子(例如,已知的配体、候选结合配偶体和/或药物候选物)相接触而确定该生物分子的取向、构象、结构或者取向变化、构象变化或结构变化的方法、装置和系统。在本公开内容的一些方面,描述了用于响应于使生物分子经受两组或更多组不同的实验条件而确定该生物分子的取向、构象、结构或者取向变化、构象变化或结构变化的方法、装置和系统。如本文所用,确定生物分子的取向、构象、结构或其变化可以涉及至少一种非线性光学信号的测定,该信号与非线性-活性标签或标记物的平均取向成正比,并且也可以与拴系在表面上的标签生物分子的表面密度成正比。如本文所用,“高通量”是指针对可选地与一个或多个已知配体、候选结合配偶体和/或药物候选物相接触的多个生物分子进行分子取向、构象、结构或其变化的快速分析(相对于例如晶体学结构确定)能力,或者是指针对可选地与大量已知配体、候选结合配偶体和/或药物候选物相接触的一个或多个生物分子进行分子取向、构象、结构或其变化的快速分析能力,或者是指这两种模式的任何组合。
定义:除非另外定义,否则本文中使用的所有技术术语具有与本公开内容所属领域的普通技术人员通常理解的相同含义。如本说明书和所附权利要求书中所使用,除非上下文另外明确指出,否则单数形式“一个”、“一种”和“该”包括复数引用。除非另有说明,否则本文中对“或”的任何引用旨在包含“和/或”。
生物分子:尽管主要在蛋白质样品的表征的背景下进行描述,但是本领域技术人员将认识到,所公开的非线性光学方法可以有利地用于多种其他类型的生物分子的结构和构象表征。如本文所用,短语“生物的分子”、“生物分子”或在一些情况下的“生物实体”包括但不限于蛋白质、蛋白质域或亚域、肽、受体、酶、抗体、抗体片段、dna、rna、寡核苷酸、dna或rna适体、小分子、合成分子、碳水化合物,或在一些情况下的细胞或其任何组合。在一些实施方式中,生物分子可以包含药物靶标或其部分,并且可以被称为“靶蛋白”或“靶分子”。在本发明的一些优选实施方式中,靶分子是蛋白质或其亚基、亚域或片段。在一些优选的实施方式中,靶蛋白是生物药物候选物(生物仿制药药物候选物)和/或参考药物
测试分子:类似地,短语“测试分子”、“测试化合物”、“候选结合配偶体”、“药物候选物”,或在一些情况下的“测试实体”包括但不限于细胞、蛋白质、肽、受体、酶、抗体、dna、rna、dna或rna适体、生物分子、寡核苷酸、缓冲液、溶剂、小分子、合成分子、碳水化合物,或在一些情况下的细胞或其任何组合。在一些实施方式中,测试分子可以包含已知的配体、药物候选物或其部分。在本公开内容的一些优选的实施方式中,药物候选物是其他蛋白质或其亚基、亚域或片段。在一些优选的实施方式中,药物候选物是生物药物候选物(例如,生物仿制药药物候选物)。
生物制品:如本文所用,术语“生物制品”(也称为“生物产品”或“生物治疗学”)是指从多种天然来源(例如,人、动物或微生物)中分离或可以通过遗传工程和其他生物技术方法生产的产品。生物制品可以包含糖、蛋白质、蛋白质片段、核酸或这些物质的复杂组合,或者可以包含具有临床诊断或治疗应用的生命实体如细胞(和组织)。
生物仿制药:如本文所用,术语“生物仿制药”(或生物仿制产品”)是指基于与已获得监管批准的生物制品(称为“参考产品”)高度相似而被批准的生物制品,并在安全性和有效性方面与参考产品无临床意义上的差异。因此,生物仿制药是现有生物药物的仿制版本。生物仿制药药物候选物(或生物药物候选物)是尚未被批准的生物制品。
参考药物:如本文所用,术语“参考药物”(或“参考产品”)是指经过批准的药物产品(例如,小分子药物或生物制品,如治疗性单克隆抗体),将其与新的仿制版本进行比较,以示出它们具有生物等效性。
参考样品:如本文所用,术语“参考样品”是指已在不同时间点制备或已使用来自不同生产过程或生产批次的蛋白质制备的蛋白质(或其他生物分子)样品。标称地,待研究的蛋白质和参考样品的蛋白质具有相同的氨基酸序列。
角度参数:如本文所用,术语“角度参数”是指本领域技术人员已知的探针相对于表面法线的平均倾斜角,围绕该平均倾斜角的取向分布宽度,即,如本文中所定义的(φ,σ)、平均倾斜角和取向分布宽度的成对组合、在两个不同的偏振下测得的tpf、shg、dfg、sfg强度的比率(例如,s-偏振强度和p-偏振强度的比率)或任何其他角度参数、在特定偏振光的激发下测得的光的强度,或在检测到的或激发光的不同频率、偏振或其他物理性质下进行的强度测定的组合,以表征探针的角度参数。
非线性光学技术:如本文所用,短语“非线性光学技术”包括二次谐波发生、和频发生、差频发生和/或双光子荧光。二次谐波发生是非线性光学过程,其中激发光的双光子在基频上与非线性材料或分子相互作用,并作为能量等于激发光子能量的两倍的单个光子重新发射或散射,即频率是激发频率的两倍。和频发生是非线性光学过程,其中激发波长或频率不同的双光子与非线性材料或分子相互作用,并作为能量等于两个激发光子能量之和的单个光子重新发射或散射,即频率等于两个激发频率之和。差频发生是非线性光学过程,其中激发波长或频率不同的双光子与非线性材料或分子相互作用,并作为能量等于两个激发光子的能量之差的单个光子重新发射或散射,即频率等于两个激发频率之差。如本领域技术人员将理解的,在整个本公开内容中,术语shg、sfg和dfg可以互换使用。双光子荧光是一种非线性光学过程,其中具有相同激发波长或频率的双光子与非线性材料或分子相互作用,并被材料或分子吸收,随后作为能量高于激发光子的能量(即具有更高频率和更短波长)的单光子的形式发射。
非线性-活性:如本文所用,短语“非线性-活性”是指二次谐波-活性(sh-活性或shg-活性)、和频-活性(sf-活性或sfg-活性)、差频-活性(df-活性或dfg-活性)或双光子荧光-活性(tpf-活性)的分子、标签或标记物,即当暴露于适当波长、强度和相位的光时,能够分别生成二次谐波光、和频光、差频光或双光子荧光。公开了采用tpf测量以及可选地结合sfg、sfg或dfg测定的多种方法。在一些情况下,分子、标签或标记物可以是非线性-活性的,从而使其在暴露于适当波长、强度和相位的光时,既发射例如二次谐波光,又发射双光子荧光。
使用双光子荧光检测分子取向、构象和结构:与更广泛使用的基于单光子荧光的技术(图1b)相比,双光子荧光(图1a)是非线性光学过程,其中具有相同激发波长或频率的双光子与非线性材料或分子相互作用,并被材料或分子吸收,然后作为能量高于激发光子的能量(即更高频率和更短波长)的单个光子的形式发射(图1a)。如本文所使用的,术语“非线性光学过程”可以指双光子荧光、二次谐波发生、和频发生或差频发生。通常,通过用至少一个基频的激发光照射非线性-活性标签来激发非线性光学过程。
双光子荧光取决于双光子跃迁偶极矩(tdm)附接有双光子活性探针(分子)的表面的法线的角度φ,并且取决于激发光的偏振与表面平面法线之间的角度θ。因此,在低-na针孔检测的极限中可将控制测得的tpf强度的方程写为:
其中io是最大强度,f是表示将激发光耦合到基底表面的棱镜表面上的功率损耗(已知值)的常数,且以下tpf阶次参数定义为:
并且是在所研究样品中探针(tpf-活性)分子的取向分布上发生积分的概率密度函数的三角矩。
通过使用两种不同的激发光偏振进行tpf测量,并采用测得的强度比,可以重新排列方程(1),以在测得的强度比与平均倾斜角φ之间产生以下关系:
其中,ip是使用p-偏振测定的tpf信号,is是使用s-偏振测定的tpf信号,并且括号(<>)表示平均值。
与shg和其他表面选择性技术的情况不同可以简单地从总tpf信号中线性减去tpf背景信号——无论是否存在配体——来确定由与感兴趣的生物分子附接的探针产生的tpf信号,而无需确定它们之间的相位,这是从背景信号解析仅有shg探针信号所需的。这为解决在筛选实验中确定配体是真配体还是假阳性的棘手问题提供了有价值的解决方案,在不存在蛋白质的情况下,配体对背景信号具有显著影响。使用shg,必须将仅有蛋白质信号从总信号和背景信号中解卷积,这需要通常是未知的或不确定的不同信号之间的相对相位的知识。使用tpf,总信号与背景信号之间的差异直接产生纯蛋白质信号。因此,通过使用tpf监测结构或构象变化,可以确定配体在仅蛋白质信号上产生的净变化,以确定配体是否为真阳性。是真阳性的并在与蛋白质特异性结合后诱导构象变化的化合物或配体将相对于它们仅在背景表面上产生的任何变化而在tpf信号中表现出净变化。
由于tpf和shg对取向分布的阶次依赖性不同,因此它们提供了可以用在检测方式(例如,tpf和/或shg)、蛋白质样品、标签位点等会改变的一个实验或单独的实验中的两个独立方程,以获得基础分子取向分布的角度测定。
在这里,在特定的实施方式中公开tpf的使用以获得例如关于标签生物分子的附加取向信息。tpf测量可以单独使用,也可以与shg、sfg或dfg测量组合使用,以获得例如,tpf和/或shg、sfg或dfg标签相对于标签生物分子所拴系表面法线的平均倾斜角度值。与单光子荧光相比,tpf由于其对标签(或探针)的倾斜角的更高阶依赖性而对标签生物分子的取向分布具有增强的角度灵敏度。此外,在使用针孔或低-na检测方案的情况下,将检测器放置在感兴趣样品的正上方或正下方,并沿垂直于表面的轴放置,在没有会聚透镜的情况下,tpf灵敏度提高了系数sin2φ。该额外的灵敏度源自探针分子的发射模式,该模式应以偶极模式辐射。tpf信号的检测取决于两个独立的过程:双光子必须被探针分子吸收,并且必须发射一个光子。在吸收过程中,光子的吸收效率取决于倾斜角φ。同样,在发射过程中,朝向检测器发射的光子的比例也取决于倾斜角φ。在低-na的极限下,检测器与探针分子正上方的表面正交,检测器检测到的光子分数随sin2φ变化。因此,涉及tpf产生和结构或构象变化的检测的本公开内容的优选实施方式使用低-na针孔检测方法。
如果拴系探针分子的取向分布相当宽(例如,大于35度),则tpf对角度变化的灵敏度比shg低得多。例如,以下描述的是对拴系在表面上的tpf和shg-活性探针高斯分布的平均倾斜角度和宽度(标准偏差)的假定值的计算结果,其公开了在没有物镜或会聚透镜的情况下,通过低na检测提供和不提供sin2φ增强的情况下,tpf测量值对平均倾斜的灵敏度。
低-na针孔检测形式用于tpf测量:与称为全内反射荧光显微镜(tirfm)的技术不同,本文公开的方法不需要高-na物镜即可产生tir条件。因此,可以使用棱镜来实现tir激发,并在样品平面上下检测tpf。在优选的实施方式中,棱镜/tir激发光学布置与平面样品(例如,其上附接有tpf-活性探针标签的感兴趣的生物分子的受支承脂双层膜)和中心位于样品平面的焦点的正上方或正下方的低-na针孔检测器组合使用。在该优选实施方式中,将感兴趣的样品拴系或固定在玻璃表面(即,光学界面)上,使用例如浸入油或光学耦合的粘合剂将该玻璃表面本身与下面的棱镜光学耦合,如本领域技术人员所公知的。重要的是,样品平面中的分子发出的荧光不通过透镜,而是由检测器检测到,因为它从样品平面向上穿过可能通过样品体积(如果是液体或空气),或从样品平面向下通过棱镜。在优选的实施方式中,感兴趣的样品包含在样品平面中以各向同性(即,在方位上)的方式分布的或假定为在其取向分布的分析中的拴系或固定化的分子。此外,为了确保高度的偏振纯度,另一优选实施方式包括将激发光聚焦到非常窄的锥角,从而使光基本上以全内反射角准直,并且几乎不存在偏轴偏振。例如,在一个实施方式中,在160mm的距离上将直径为4mm的激光束聚焦到直径为50μm的光斑,从而在临界角上方和下方产生约1.5度或约0.8度的全锥角。
不同检测方案下的双光子荧光灵敏度分析:为了说明tpf对平均角度和分布宽度的灵敏度,进行了计算以确定给定平均角度和分布宽度(假设为高斯分布)下,以相同的分布宽度改变为5°的平均角度时tpf强度变化的幅度。下面的计算估计了当激光的电场垂直于表面法线(s-偏振)时信号的预期变化。对于平行于位于样品正上方或正下方的表面法线的检测器,在高na的情况下,从tpf检测到的光子数应按<sin4φ>标度,而在低-na情况下应按<sin6φ>标度。
可以使用以下方程在高斯分布上对方程(2)(对于tpf)或方程(6)(对于shg)进行积分,以确定满足方程的倾斜角度和分布宽度对。方程(2)和(6)中的括号<f(φ)>指示表达式f(φ)的倾斜角φ从0到π乘以高斯的归一化积分:
其中φ0是平均角度,σ是分布宽度。由于高斯函数具有无限范围,并且积分的求值范围为0到π,因此,按照simpson和rowlen概述的过程(simpson和rowlen(1999),j.am.chem.soc.121:2635-2636),对-4π和4π之间的所有贡献求和,从而将高斯函数“折叠”为积分。这将产生对于至多70°的分布宽度有效的积分。计算结果总结在表1中。
表1.用高na和低-na光学方案收集的tpf信号的比较
初始状态最终状态
从表1总结的结果中可以看出,对于整个可能取值范围内的平均角度值和分布宽度值,低-na检测方法(即,不使用透镜)显著提高了对角度变化的灵敏度。
此外,从这些计算中还发现,如果双光子活性探针的取向分布较窄(≤25°),则tpf测量值对平均倾斜角的灵敏度将接近shg的灵敏度。例如,在平均角度为30°,并且宽度为15°时,shg信号变化10%变为32°的角度,而宽度保持15°不变;对于相同的初始状态和最终状态,没有透镜的低-na检测的tpf变化15%。类似地,对于平均角度为20°,并且宽度为25°的情况,shg变化18%变为25°的平均角度,并且宽度为25°;低-na检测的tpf变化了24%。因此,本发明的关键方面和一个优选的实施方式涉及至少一种样品(例如,蛋白质样品)的使用,该样品包含外源性附接的探针、染料或非天然氨基酸,或遗传掺入的非天然氨基酸或其他标签,其中测得的探针取向分布的宽度小于或等于25°(度)。
tpf跃迁偶极矩(tdm)和可选的shgχ(2)的测量以及与蛋白质结构的关系:通常,所公开的方法、装置和系统可能依赖于使用双光子诱导荧光(tpf),以及可选的二次谐波发生(shg)或者相关的和频发生(shg)或差频发生(dfg)的非线性光学技术,用于确定分子取向、构象、结构或其变化。在这些方法中,偏振依赖性测量用于确定双光子吸收跃迁的跃迁偶极矩(tdm)的分量(或shg和相关非线性光学技术的超极化率χ(2)的分量,如下所述)。tpg强度的值或shg的χ(2)的分量可以使用至少一个基频的偏振激发光在实验室参照系中测量χ(2)的分量。一些光源,例如一些激光器,产生基本上偏振的基频光。在一些实施方式中,可以使用一个或多个光偏振器、波片等来进一步限定并且/或者调节激发光的偏振。通常,偏振光的入射面(即,由激发光的传播方向和垂直于基底平面或反射表面的矢量所定义的平面)将为图2所示的实验室坐标系的x-z平面。具有平行于入射平面的电场矢量的偏振光被称为p-偏振光。具有垂直于入射面的电场矢量的偏振光被称为s-偏振光。在一些实施方式中,还可以使用一个或多个光偏振器、波片等来限定和/或调节由非线性-活性部分的激发所产生的已检测到的二次谐波光的偏振。如上所述,通过使用至少两种不同激发光的偏振来测量双光子荧光强度,可以使用所得到的关于标签的相对取向的信息来开发蛋白质结构的模型,并使用双光子荧光或结合双光子荧光与shg测量或相关的非线性光学测量技术来检测其变化。
在一些实施方式中,tpf与shg测量组合使用,并在激发光的一个或多个偏振下进行检测tpf,以获得与附接在生物分子上的探针有关的附加取向信息,出于获得有关生物分子的结构信息的目的,该探针进而被拴系在表面。例如,由于tpf依赖于探针方向的顺序参数不同于shg,因此它提供了独立的方程,其允许同时求解两个单独的取向参数,例如探针分子的高斯分布的平均值和宽度。在优选的实施方式中,使用如下所述的低-na或针孔检测设备来完成对tpf信号的检测,并且导致对探针取向的甚至更高阶的依赖性,并从而提高了灵敏度。在一些实施方式中,由shg和tpf确定的角度测量的最高置信度发生在探针和生物分子以相对窄的取向分布拴系到表面时,其中对生物分子内的至少一个探针位置,通过shg和tpf组合测量确定的并假设为高斯分布的探针的角度分布,导致取向分布宽度为35度或更小(≤35°)。在一些实施方式中,由shg和tpf确定的角度测量的最高置信度发生在标签生物分子的拴系导致相对窄的取向分布宽度小于或等于30°、小于或等于25°或小于或等于20°时。在一些实施方式中,标签或探针是tpf-活性,并且可选地也是shg-活性、sfg-活性或dfg-活性的,并且可以使用本领域技术人员已知的技术(例如,掺入非线性-活性的非天然氨基酸)将其掺入感兴趣的生物分子如蛋白质内的特定位点。在一些实施方式中,将这样的探针掺入单个生物分子构建体中的单个位点,而在其他实施方式中,将两个或更多个探针掺入单个生物分子构建体中的多个位点。
在一些实施方式中,用标记有tpf-活性探针的生物分子来测量tpf,以确定被标记的生物分子是否附接在表面,这并不总是通过单独使用shg测量就可以看出,因为如果shg探针的净平均取向相对于表面法线相对平坦,则信号将相对较小,而相同样品通常会产生相应的高tpf信号。因此,由同时具有tpf-活性和shg-活性的探针产生的信号量在两种技术中倾向于反相关,下面将在理论背景中进行更详细的描述。
在一些实施方式中,可以用仅有shg-活性或仅有tpf-活性的探针标记生物分子,并且可以将每个实验的测量结果相互比较。在一些情况下,可以在一个位点标记生物分子,而在其他实施方式中,可以创建许多不同版本的生物分子,每个版本在具有shg和tpf-活性的唯一的单个位点带有探针。在其他实施方式中,产生不同版本的生物分子,每个版本在独特的单个位点上带有探针,该位点是i)tpf-活性的,或ii)shg-活性的(或sfg-活性或dfg-活性的)。
从x射线晶体学或nmr方法获得结构信息可能具有挑战性,这些结构信息由于诸如通量、灵敏度、非生理条件的使用、适合该技术的蛋白质大小等因素,在药物发现应用中价值有限。因此,本发明的另一方面是提供蛋白质或其他生物分子中功能相关位点上构象的位点特异性读出。如本文所定义,“功能相关”的蛋白质位点包括通过诸如x射线晶体学、nmr或shg的结构技术确定的与结合配偶体(例如,效应分子)直接或间接结构接触的任何位点。直接结构接触被定义为任何氨基酸或其他结构残基,其一些部分在结合配偶体分子的一些部分的2nm以内。间接结构接触被定义为任何氨基酸或其他结构残基,通过诸如x射线、nmr或shg的结构技术所见,其一些部分在与结合配偶体(例如,效应分子)或模拟或类似物结合后,会相对于其在没有结合配偶体、模拟或类似物的情况下的取向、构象或相对坐标改变其取向、构象或相对坐标。术语“功能相关”还包括已知在通过非结构性手段(例如,诱变或生化数据,表明特定残基对于结合配偶体的结合或调节很重要)的结合分子的结合或调节(例如,活化、抑制、调节等)中重要的残基。
在一些实施方式中,使用所公开的方法获得的tpf结构数据和可选的shg结构数据可以与来自蛋白质晶体学、nmr研究、uv-vis和荧光光谱研究、圆二色性研究、交联实验、小角度x射线散射研究等的结构数据重叠或组合。在一些实施方式中,该方法进一步包括使一个或多个非线性-活性标签的相对取向全局地拟合至蛋白质分子的结构模型,其中该结构模型基于蛋白质分子内一个或多个非线性-活性标签的已知位置。可选地,在确定这样的模型时,可以采用附加的结构测量值或约束条件,例如来自x射线、nmr测量或其他实验测量的数据。
在一些实施方式中,tpf和/或shg信号测量可以在如下所述的多种实验条件下执行,其中不同的实验条件导致拴系到光学界面的标签分子的取向分布发生变化。由于实验系中不同的基础取向分布,导致tpf跃迁偶极矩和可选的shgχ(2)的测量值组不同的每组实验条件均允许独立测量倾斜角φ由tpf和可选的shg确定。通过组合两个或更多个这样的测量,可以更准确地确定蛋白质结构,包括以多种构象状态的平衡存在的蛋白质结构。通过使用本领域技术人员已知的标准分子建模技术,可以使用tpf跃迁偶极矩和可选的shgχ(2)的分量的测量以及φ值的确定来开发结构模型,并且在一些实施方式中,选择适当的简化假设。可以简化分析并开发蛋白质结构模型的假设的一个非限制性示例是,尽管蛋白质表面的tpf-活性和/或shg-活性标签的取向在实验参考系中从一种实验条件到另一实验条件(即,相对于垂直于表面平面的轴)有所不同,但tpg-活性和/或shg-活性标签相对于蛋白质参考系在不同实验条件下保持恒定。实际上,在该假设下,人们以不影响其功能和构象结构的方式改变了蛋白质在表面上的取向分布。每个实验条件都产生至少一个独立的方程,该方程将在不同偏振下测得的tpftdm和可选的shg强度与分子取向分布相关联。可以在每种实验条件下进行适当的控制,如配体诱导构象变化、配体竞争实验、配体结合动力学、剂量-反应测量等,以确保蛋白质仍具有功能并因此具有天然性质。根据本领域技术人员已知方法,平均角度的测量,例如蛋白质中标签或双光子活性或超极化部分的取向分布的其他参数,可以用作全新或整体结构模型构建的约束。在一些实施方式中,例如,蛋白质的载脂蛋白x射线晶体结构可以被包括在模型中,并被由tpf和/或shg测量提供的结构数据覆盖,以改善模型的准确性。
为了简化shg结构数据的分析,可以在所公开的方法的一些实施方式中作出的假设的非限制性示例包括:(i)tpftdm的单个分量,以及可选地,α(2)项(例如,αzzz(2))支配双光子吸收张量(对于shg,可选地是标签的超极化率);(ii)标签在蛋白质中的位置(即,它们所附接的氨基酸残基的身份)是已知的;以及(iii)拴系或固定化的蛋白质分子的取向在x-y平面上是各向同性的(即,它们在基底表面的平面上或受支承脂双层的平面上随机取向)。
在所公开的方法的一个实例中,蛋白质在单个位点——具体改造的半胱氨酸残基——处用双光子活性和可选的shg-活性标签加以标签,该蛋白质进而具有双光子吸收张量和可选地α(2)=α(2)z'z'z'的单一优势元素。经标签蛋白质经由his-标记物与含有附接至脂质头基的ni-nta部分的受支承脂双层膜附接。如本领域技术人员所公知的,以这种方式产生基线tpf信号和可选的shg信号,并且给出了χ(2)的非零分量(salafsky,j.s.(2001),“’shg-labels’fordetectionofmoleculesbysecondharmonicgeneration”,chemicalphysicsletters342,485-491;salafsky,j.s.(2003),“second-harmonicgenerationasaprobeofconformationalchangeinmolecules”,chemicalphysicsletters381,705-709;salafsky,j.s.(2006),“detectionofproteinconformationalchangebyopticalsecond-harmonicgeneration”,journalofchemicalphysics125),由以下方程:
χzzz(2)=ns<cos3φ>αz′z′z′(2)(4)
其中ns和αz’z’z’(2)分别是表面密度和分子超极化率。然后可以由两个不同的偏振依赖性测量值(izzz和izxx,或等同地,ippp和ipss)来确定χ(2)的分量。在这种情况下,可以通过使用p-偏振基本激发光测量p-偏振shg信号来确定χzzz(2)。例如,如果使用800nm的基本激发光(例如,来自ti:蓝宝石锁模激光器),则在400nm处检测到二次谐波信号。一般来说,在p-偏振激发和p-偏振shg检测下观察到的shg信号强度ippp,由非线性极化率的若干个分量所决定。然而,通过使用二氧化硅棱镜测量在全反射几何中的临界入射角处的shg信号来实现在该测量中仅分离χzzz(2)的简化的方法。在全内反射(tir)几何中,由于χ(2)的离轴张量分量消失,仅留下了确定所测量的ipppshg信号强度的χzzz(2)(是指基本光束/shg光束的偏振),因此所测量的shg强度由棱镜和表面拴系的蛋白质泡在其中的缓冲液的折射率而决定。类似地,可以通过使用s-偏振的基本光并测量p-偏振的shg光强测量ipss来确定χzzz(2)。在ns和αz’z’z’未知的情况下,可以使用在不同偏振组合下测量的强度的比例来消除这些参数,只留下作为φ的三角函数的取向分布本身的比例,其中φ被定义为分子系中z-轴和表面法线之间的平均角度。当取向分布较窄时,可以直接确定φ。通过使用在两个或更多个不同位点处标签的蛋白质(例如,在两个或更多个不同的单位点半胱氨酸突变体中,在这些半胱氨酸位点处),在不同的偏振(例如,izzz和izxx)下重复shg强度的测量,例如可以获得针对可以用作结构确定中的约束的每个标签位点处的不同的φ。每次测量都需要经标签蛋白质,优选在蛋白质中已知位置处与标签位点特异性附接(例如,经由定点半胱氨酸诱变的共价附接)。还通过改变实验条件以产生相对于表面平面的不同取向的蛋白质以及两个或更多个不同位点的标签,可以进行独立测量以确定描述取向分布的多个参数,由此为蛋白质结构确定提供重要的约束。本发明的关键步骤是在两个或更多个不同的实验条件下测量在两个或更多个不同位点处(优选在单独的蛋白质-标签缀合物中)标签的蛋白质,这导致不同的χ(2)值,这取决于基本的取向分布,或χ(2)分量的不同比例(例如χzzz(2)/χzxx(2))。通过测量在两个或更多个不同的实验条件下在两个或更多个不同位点处标签的相同蛋白质的χ(2)值,可以获得更准确的φ(在实验系中)的测量值,并且将它们(即在蛋白质系中)之间的差异与蛋白质结构进行关联。
多重构象-平衡取向分布:如果蛋白质(或其他生物分子)以多种构象状态的平衡存在,则所述蛋白质可以通过在每个标签位点处的多模态(或多状态)取向分布加以描述。如果所述分布由具有不同权重、平均角度(φ)和分布宽度(σ)的高斯分布的总和组成,则蛋白质构象景象的完整描述将取决于确定这些参数中的每一个。例如,如果标签位点1处的蛋白质的局部结构采用3个构象,则在这些假设下,局部取向分布可以由待确定的3x3参数或描述每个构象的幅度、平均角度和宽度的9个未知量来描述。标签位点2可以仅采用2个局部构象,并且在这种情况下可以类似地由6个参数来描述。本发明的一个方面是,可以通过改变实验条件,如标签位点(如c-端和n-端)、融合蛋白序列、标签长度(例如6x、8x、10x和12xhis-标记物)、缓冲液条件(例如不同的盐浓度)等,或者通常改变蛋白质在表面上的取向并因此影响χ(2)测量值的任何实验条件来获得附加独立测量值。然后可以使用所有这些独立测量值,例如在全局地拟合方法中确定在这两个位点的蛋白质参照系中的基于溶液的构象景象(即多状态取向分布)。在一些实施方式中,蛋白质的x射线晶体结构坐标系可以可选地用作模型构建中的进一步约束。
使用shg和相关的非线性光学技术检测分子取向、构象和结构:过去已经使用shg和相关技术和频发生(sfg)来研究染料分子在界面处的取向(heinzt.,等人,(1983),"determinationofmolecularorientationofmonolayeradsorbatesbyopticalsecond-harmonicgeneration",physicalreviewa28(3):1883-1885;heinz,t,(1991)second-ordernonlinearopticaleffectsatsurfacesandinterfaces",innonlinearsurfaceelectromagneticphenomena(stegeman,h.p.a.g.ed.),elsevier,amsterdam,第353-416页)。在这些测量中,使用偏振光确定标签界面的非线性磁化率的分量(χ2)。然后可以使用实验确定的χ2值和有关界面平面内染料分子取向程度、染料分子在参考分子系中的超极化性分量(α2)的相对大小等假设,来推断出界面上染料分子的分子取向分布的细节。
在先前的技术出版物和专利申请中已经公开了使用shg以及sfg和dfg的相关的非线性光学技术来检测表面上的生物分子结合事件、测量倾斜角或测量蛋白质的构象变化。(参见,例如,salafsky,j.(2006),“detectionofproteinconformationalchangebyopticalsecond-harmonicgeneration”,j.chem.phys.125:074701;美国专利号6,953,694;和美国专利号8,497,073)。通常,这些公开内容描述了使用shg或其他非线性光学技术来测量靶蛋白与结合配偶体如配体接触后信号的变化。
与更广泛使用的基于单光子荧光的技术(图1b)相比,二次谐波发生(图1c)是一种非线性光学过程,其中同一激发波长或频率的双光子与非线性材料相互作用并作为具有激发光子的两倍能量(即,其两倍的频率和一半的波长)的单个光子进行重新发射。二次谐波发生仅发生在缺乏反演对称性的非线性材料中(即,非中心对称材料中),并且需要高强度激发光源。这是和频发生的一种特殊情况,并且与例如差频发生等其他非线性光学现象相关。
二次谐波发生和其他非线性光学技术可因其对非线性-活性物种的高度相关性而配置成表面选择性检测技术。例如,非线性-活性物种向表面的拴系可以产生在分子可在溶液中经历自由扩散的情况下缺少的净、平均取向度。常用于对非线性-活性物种在界面处的取向相关性建模的公式为:
χ(2)=ns<α(2)>(5)
其中χ2是非线性磁化率,α(2)是非线性磁化率,ns是界面处每单位面积上非线性-活性分子的总数,而<α(2)>是这些分子的非线性超极化率(α(2))所有取向的平均取向。描述二次谐波发生的非线性相互作用的典型方程为:
α2(2ω)=βe(ω)e(ω)或
p2(2ω)=χ2e(ω)e(ω)
其中α和p分别是在2ω频率处振荡的感应分子和宏观偶极子,β和χ2分别是超极化率和二次谐波(非线性)磁化率张量,并且e(ω)是入射辐射在以频率ω振荡的电场分量。宏观非线性磁化率χ2与微观β超极化率的方向平均值为α2。感应的宏观偶极子展开中的下一个阶项描述了其他非线性现象,如三次谐波发生。三阶项复制诸如双光子荧光的非线性现象。为了产生和频或差频,驱动电场(基本)以不同的频率(即,ω1和ω2)振荡,而非线性辐射以和频或差频(ω1±ω2)振荡。
shg的强度与非线性磁化率的平方成正比,并因此既决于界面上定向的非线性-活性物质的数目及其取向分布。可以利用此性质来检测构象变化。例如,受体的构象变化可以使用非线性-活性标签或部分来检测,其中该标签附接于或拴系于表面的受体上;构象变化导致标签相对于表面平面的方向(取向)变化,从而导致非线性光学信号的物理性质(例如,强度)变化。该技术本质上对界面处的标签分子取向分布的变化敏感,无论是在空间上还是时间上。
通过使用两种不同的激发光偏振进行shg测量,并获取测得的强度的比率,可以从测得的强度比与平均倾斜角φ之间的关系得出平均倾斜角:
其中ippp是使用p-偏振测量的shg信号,ipss是使用s-偏振测量的shg信号,f是表示用于将激发光耦合到基底表面的棱镜表面的功率损耗(已知值)的常数,并且括号(<>)表示平均值。
因此,在所公开的方法的一些实施方式中,使用平均倾斜角φ的偏振激发光测量的shg信号强度的强依赖性(如方程(6)所示)被用于在各种检测偏振下,通过使用p-偏振和s-偏振激发光测量shg信号的比率来更灵敏地检测标记的蛋白质或其他生物分子中的构象变化,p-偏振shg检测是具有纯p-偏振和纯s-偏振激发(例如χzzz(2)/χzxx(2))的优选实施方式。在另一实施方式中,可以使用p-偏振光和s-偏振光的混合状态的单个偏振来生成自身在两个正交方向上偏振的shg。通过例如使用偏振光束分裂立方体分裂信号来测量这两个正交偏振的shg信号的相对强度,可以使类似于方程(6)的方程公式化,以中继有关平均倾斜角φ的信息。
通过使用全内反射作为用于将激发光传递到已拴系或固定化了非线性活性物质的光学界面(或表面)的模式,可以附加地使二次谐波发生和其他非线性光学技术(如上所述,包括tpf)成具有表面选择性。入射激发光的全内反射会在界面处产生“迅衰波”,其可以用于选择性地仅激发靠近表面(即,迅衰波的空间衰减距离内,其通常为几十纳米级别)的非线性-活性标签。在本公开内容中,借助于激发光的全内反射生成的迅衰波优选用于激发非线性-活性标签或分子。本文所述的非线性-活性过程中激发非线性-活性物种的效率很大程度上取决于其相对于表面的平均取向。例如,如果不存在非线性-活性物质的净平均取向,则将无shg信号。
可以利用shg和其他非线性光学技术的这种表面选择性质来确定固定于界面的生物分子中平均取向、构象、结构或其变化。例如,由于配体的结合而造成的受体分子中的构象变化可采用非线性-活性标签或部分来检测,其中所述标签附接到或关联于所述受体,以便使构象变化引起标签相对于界面的取向或距离的变化(图3),并从而导致非线性光学信号的物理性质的变化。直到最近,由于相对少数的生物样品本身为非线性-活性,因此对表面选择性非线性光学技术的使用还主要局限于物理和化学应用上。最近,已经介绍了对二次谐波活性标签(“shg标签”)和其他非线性-活性标签的使用,从而事实上使得几乎任何所提供的分子或粒子均具有高非线性-活性。这方面的第一个例子由使用噁唑染料标签蛋白细胞色素c并利用二次谐波发生来检测空气-水界面处的蛋白缀合物所证实[salafsky,j.,“`shg-labels'fordetectionofmoleculesbysecondharmonicgeneration”,chem.phys.lett.342(5-6):485-491(2001)]。下文将更详细地描述标签或以其他方式使靶蛋白、生物药物候选物、参考药物和其他生物实体具有非线性-活性的技术。
表面选择性shg、sfg和dfg非线性光学技术也为相干技术,这意味着基本光束和非线性光束具有以明确定义的空间关系和相位关系通过空间传播的波前。针对分析生物分子或其他生物实体的构象而采用表面选择性非线性光学检测技术相比于其他光学方法具有多种固有优势,包括:i)非线性信号对于非线性-活性物种的取向和/或一个或多个偶极矩的敏感且直接的相关性,赋予其对构象变化的灵敏度;(ii)比基于荧光的检测更高的信噪比(较低的背景),原因在于非线性光学信号仅在创造出非中心对称系统的表面处生成,即,该技术本身具有非常窄的“景深”;(iii)作为窄“景深”的结果,该技术在必须在有覆盖溶液的情况下进行测量时(例如,在结合过程可能被分离或冲洗步骤所消除或扰乱的情况下)有用。该技术的这一方面对于进行需要大块物体存在的平衡结合测量或者在限定时间段内进行测量的动力学测量可能特别有用;(iv)该技术表现出比荧光中发生的光漂白和加热效应更低的光漂白和加热效应,原因在于对于给定分子,双光子截面通常远小于单光子吸收截面,并且shg(以及和频发生或差频发生)涉及散射而非吸收;(v)需要最少的收集光学器件并且预期信噪比较高,原因在于基本光束和非线性光束(例如,二次谐波光)相对于界面具有明确定义的传入和传出方向。这与基于单光子荧光的检测相比特别有利,因为荧光发射是各向同性的,并且对于由焦平面外荧光物产生的所检测信号可能还存在较大的荧光背景成分。shg、sfg或dfg产生的信号提供了研究分子的结构、构象或其变化(诸如例如在配体结合后发生)的即时、实时的手段。该性质可在所公开的用于实时获得经历结构改变的蛋白质的实时“运动”作为其实时功能一部分的方法中非常有用。
如果存在背景shg信号,例如由于基底-缓冲液界面,则可以多种方式“减去”该背景信号。例如,可以在诸如在reider,g.,等人(1999),“coherenceartifactsinsecondharmonicmicroscopy”,appliedphysicsb-lasersandoptics68,343-347描述的实验中,或在clancy和salafsky(2017),“second-harmonicphasedeterminationbyreal-timeinsituinterferometry”,phys.chem.chem.phys.19:3722-3728描述的实验的干涉实验中测量来自蛋白质上标签的shg信号和由于背景引起的shg信号之间的相差。然后可以确定仅由于标签蛋白质引起的shg信号。
可出于结构确定、结构比较和/或构象变化检测的目的而被监测的二次谐波光和相关的非线性光学信号的物理性质的示例包括但不限于强度,偏振,波长,强度、偏振或波长的时间依赖性或其任何组合。
shg信号的归一化:在本文公开的任何实施方式中,用于鉴别感兴趣生物分子内的配体结合位点或配体结合区的位置的方法可以进一步包括同时或连续测量双光子荧光(tpf)信号,及其计算shg:tpf信号比(或sfg:tpf信号比、dfg:tpf信号比或p-偏振/s-偏振tpf比或p-偏振/s-偏振的shg比,如本文所述)的用途,出于将测量的非线性信号归一化为界面的每单位面积所拴系的分子数(即,界面上的拴系分子的表面密度或数密度)的目的。例如,在一些实施方式中,当用与生成二次谐波、和频或差频光的基频相同或不同的基频光照射时,用于在将靶蛋白质拴系到界面之前标记靶蛋白质的非线性-活性(即,shg-活性、sfg-活性或dfg-活性)标签还可以产生双光子荧光。在一些实施方式中,可以用不同于shg-活性、sfg-活性或dfg-活性标签的双光子荧光标签标记靶蛋白。由于双光子荧光信号与被激发的标签分子的数目线性相关,因此双光子荧光信号提供了标准化shg(或sfg或dfg)信号,以校正拴系分子的表面密度变化的方法,并因此便于比较标签蛋白质的不同样品中测得的信号。在一些实施方式中,可以通过使用全内反射将基本光(即,通常由激光器提供的激发光)输送到界面来激发双光子荧光。在一些实施方式中,可以在通过在与界面平面正交的方向上(例如,使用落射荧光光学装置),或以不正交于界面平面的任意角度输送基光来激发双光子荧光。在一些实施方式中,可以使用落射荧光光学装置检测和测量在用基频光照射时激发的双光子荧光,例如,其中使用显微镜物镜会聚发射的双光子荧光。在一些实施方式中,可以使用低-na针孔(即,不使用透镜)来检测和测量双光子荧光,该低-na针孔位于激发光聚焦和定向的点的正上方或正下方,使得其与界面平面平行。然后,通过会聚透镜、显微镜物镜或针孔(使用或不使用任何中间光学元件,如附加透镜、反射镜、二向色反射镜,带通滤光片和/或孔径)的双光子荧光可以随后使用光电倍增管或其他合适的检测器检测到。在适当的反应条件下,对半胱氨酸残基具有特异性的tpf-活性和shg-活性探针的示例为1-(2-马来酰亚胺基乙基)-4-(5-(4-甲氧基苯基)恶唑-2-基)甲基磺酸吡啶鎓。
来自不同样品或在不同时间点的蛋白质结构的结构相似性确定和比较:如上所述,在本公开的第一方面,描述了使用shg或相关的非线性光学基线信号(例如,sfg和dfg基线信号)用于比较来自不同样品或不同时间点的蛋白质结构(或其他生物分子结构)。先前的公开内容与本发明之间的关键区别在于,认识到基线shg信号本身提供了有价值的工具,用于比较不同蛋白质样品或不同时间点的蛋白质结构。由于其对非线性-活性标签的净取向极度敏感,因此在用基频的光在不同时间点照射相同样品中来自不同样品(例如,已使用相同的标记方法标记的样品,以及已使用相同的拴系或固定技术拴系或固定化在光学界面(例如,玻璃基底表面)上)的标签蛋白分子后,基线shg信号的测量提供了方便、灵敏的工具来检测蛋白质结构的细微差异。标称相同的蛋白质产品应具有基本相同的基线shg信号。三级结构略有不同的蛋白质产品,例如,由于生产过程中折叠的细微差异,或由于给定缓冲液配方的稳定性略有不同,应具有不同的基线shg信号。完全变性的蛋白质应具有零个可测量的基线shg信号(即,由于在光学界面没有非线性-活性标签的净取向)。从生产过程中的不同步骤或相同生产过程中生产的不同批次的蛋白质中标记的蛋白质样品标记蛋白样品、或在不同时间点的给定蛋白质样品或通过不同生产过程生产的假定相同的蛋白质产品的基线shg信号的比较应因此提供有用的工具,用于优化和监测生产过程、监测蛋白质在暴露于不同试剂或使其经受不同实验条件后的稳定性、监测生产过程的输出(例如,用于质量控制),以及评估基于结构的相似蛋白质产品(例如,用于证明生物药物候选物与参考药物之间的生物相似性,或用于证明临床诊断测试中使用的单克隆和/或多克隆抗体的结构等效性)。
在一些实施方式中,这些比较可以仅依赖于shg基线信号(或sfg或dfg基线信号)的测量。成功减少所公开方法的实践要求鉴别和消除基线shg信号中所有潜在的错误来源,而非蛋白质三级结构的差异,例如,标记特异性或产量的差异、光学界面上结合位点密度的差异、拴系或固定效率的差异等。
在一些实施方式中,可以使用对相同样品测量的shg基线信号(或sfg或dfg基线信号)与tpf基线信号的比率来进行这些比较,其中,同时或连续测量shg基线信号和tpf基线信号。在优选实施方式中,可以使用具有二次谐波-活性和双光子荧光-活性的单个非线性-活性标签来标记蛋白质样品。由于tpf信号与拴系在表面标签蛋白的表面密度线性成正比(如上所述,shg信号与每单位面积拴系的标签蛋白数的平方成正比),因此使用shg与tpf基线信号比允许使shg基线信号标准化,并校正标签蛋白分子表面密度的变化。
蛋白质样品:所公开的方法、装置和系统可用于监测多种纯化或未纯化蛋白质的任一个的样品中的蛋白质结构变化。适合该方法的蛋白质的示例包括但不限于,酶、受体、抗体、单克隆抗体、多克隆抗体、人源化抗体、igg抗体、igm抗体、iga抗体、igd抗体、ige抗体、融合蛋白质或其他遗传工程蛋白质及其亚基或片段。在一些实施方式中,蛋白质可以是生物药物或候选药物。
在一个优选的实施方式中,要对其三级结构进行监测的蛋白质可以是经过遗传工程化以掺入独特的标签位点以附接非线性-活性部分的蛋白质(即,具有shg和/或tpf活性的非线性活性标签或标记物),和/或经过遗传工程化以掺入本质上具有非线性-活性的非天然氨基酸的标签的蛋白质。可以遗传-掺入的独特标签附接位点的示例包括但不限于在正确折叠蛋白质时已知位于蛋白质表面的氨基酸序列位置处掺入赖氨酸、天冬氨酸、谷氨酸或半胱氨酸残基。在掺入赖氨酸的情况下,然后可以使用本领域技术人员已知的多种缀合化学中的任一种将非线性-活性标签缀合至赖氨酸残基的伯胺。类似地,在掺入天冬氨酸或谷氨酸的情况下,然后可以将非线性-活性标签缀合到天冬氨酸或谷氨酸残基的羧基上。在掺入半胱氨酸的情况下,然后可以将非线性-活性标签缀合至半胱氨酸残基的巯基上。在蛋氨酸标签的情况下,本文描述的一般方法提供了很好的方法,前提是可以获得或合成具有所需化学处理的shg-活性和tpf-活性探针。如所指出的,在一些情况下,标签可包括本质上非线性-活性的非天然氨基酸的遗传掺入。本质上具有非线性-活性的非天然氨基酸的一个非限制性示例是在cohen,等人(2002),“probingproteinelectrostaticswithasyntheticfluorescentaminoacid”,science296(5573):1700-1703中描述的aladan。合适的标签技术的其他示例将在下面更详细地讨论。
在另一优选的实施方式中,要对其三级结构进行监测的蛋白质可以是经过遗传工程化以掺入独特的拴系或固定位点,以将蛋白质附接至光学界面的蛋白质,和/或已被遗传工程化的蛋白质以掺入非天然氨基酸残基的蛋白质,该残基充当独特的拴系或固定位点,用于将蛋白质附接到光学界面或用于以双正交方式附接shg-活性或tpf-活性探针。可以遗传掺入的独特的拴系或固定位点的示例包括但不限于,在正确折叠蛋白质时已知位于蛋白质表面的氨基酸序列位置处掺入赖氨酸、天冬氨酸、谷氨酸、甲硫氨酸或半胱氨酸残基。然后可以使用本领域技术人员已知的多种缀合和接头化学中的任一种将蛋白质拴系或固定在光学界面上。可以遗传-掺入到蛋白质产物中的独特的拴系或固定位点的另一非限制性示例可以是his标记物(例如,1、2、3、4、5、6、7、8、9、10、11、12个或多于12个的组氨酸残基的系列),然后可以提供与结合到光学界面的ni/nta基团结合的附接位点。可以掺入以提供独特附接点的非天然氨基酸的一个非限制性示例是生物素化的非天然氨基酸生物胞素。然后可以使用高亲和力的生物素-链霉亲和素相互作用将蛋白质拴系至或固定在光学界面,以将蛋白质拴系至固定在基底表面上的链霉亲和素分子上。合适的拴系或附接技术的其他示例将在下面更详细地讨论。
在一些优选的实施方式中,要对其三级结构进行监测的蛋白质可以是经过遗传工程化以掺入独特的非线性-活性标签位点或非线性-活性氨基酸残基以及独特的拴系蛋白质或固定位点的蛋白质。
蛋白质结构稳定性的测定:在一些实施方式中,所公开的方法、装置和系统可用于通过使用非线性光学技术来监测蛋白质构象、蛋白质取向分布或其变化随时间和/或在不同实验条件下(例如,在不同的缓冲液中、在不同的储存条件下、在不同的附接接头下等)监测蛋白质稳定性(例如,用于生物制品、基于生物蛋白质的药物或感兴趣的其他蛋白质)。例如,在一些实施方式中,可以通过本领域技术人员已知的多种方式中的任一种,用shg-活性或其他非线性-活性标签标记蛋白分子(例如,单克隆抗体(mab)药物或药物候选物)并拴系于光学界面。然后,将标签蛋白质在被基频照射后产生的shg或其他非线性光学信号强度(或非线性光学信号的其他物理性质)的变化作为时间的函数进行监测,或将标签蛋白质与一种或多种候选稳定化合物、候选破坏性化合物、候选稳定或存储缓冲液等接触。在优选实施方式中,shg与tpf信号强度的比率(或非线性光学信号的其他物理性质的比率)可用于监测蛋白质样品随时间的变化,或将标签蛋白质与一个或多个配体、候选稳定化合物、候选破坏性化合物、候选稳定化合物、温度、缓冲液等接触。然后所得的shg或非线性光学信号或信号比(即“签名”)的比较提供了监测蛋白质构象、取向分布或其变化的方法并在不同实验条件下确定蛋白质随时间变化的稳定性。在一些实施方式中,可以在不同物理条件下(例如,在不同温度下)以及在不同化学条件(例如,不同ph、不同离子强度、不同缓冲剂等)下确定蛋白质的稳定性。稳定性的确定可以基于实时监测shg或其他非线性光学信号或信号比的变化(例如,基于动力学测量),或者可以基于shg或其他非线性光学信号或信号比的端点测量。
因此,基线shg信号(或shg与tpf信号的比率)提供了蛋白质稳定性随时间或在不同组实验条件下的相对量度。更无序的蛋白质可能会导致更低的信号(反之亦然),因为拴系蛋白质的取向分布宽度(但不一定是平均角度),会在不稳定缓冲液中发生变化。shg信号强度与<cos3φ>2成正比,其中φ是分子超极化率轴(假定为单个显性张量元素)与标记签子参考系中表面的法线之间的角度,并且其中括号表示取向平均值。较不稳定并因此更未展开的蛋白质应产生更宽的取向分布和更少的shg信号。在蛋白质完全变性的限制内,信号应接近或等于零,因为非线性-活性染料应相对于光学界面接近完全随机取向或是完全随机取向的。该预测由将在所附示例中描述的实验数据得到证实,在该示例中,将非线性-活性标签的栓系fab片段暴露于浓度增加的尿素中,以生成变性曲线。从这样的曲线还可以获得变性自由能的估计。
在一些实施方式中,所公开的方法可用于在限定的条件组下,例如实验条件或储存条件下,或在限定的条件组变化后,以秒、分钟、小时、天或周为时间尺度来监测蛋白质的稳定性。
蛋白质-蛋白质相互作用和筛选稳定或破坏蛋白质-蛋白质复合物的化合物:在一些实施方式中,公开的方法装置和系统可用于监测蛋白质-蛋白质相互作用和/或筛选稳定或破坏蛋白质-蛋白质复合物的化合物。在一些情况下,可以用非线性-活性标签标记拴系在光学界面的蛋白质,并且可以借助于在拴系分子中诱导的构象变化例如,通过在将拴系分子与一种或多种其他分子接触后,测量基线shg信号或shg与tpf信号的比率的变化来监测一个或多个附加的蛋白质分子的结合。在一些情况下,未标记拴系在光学界面的蛋白质,并且可以借助于通过测量在拴系分子与一个或多个其他分子接触时,基线shg信号或shg与tpf信号的比率的变化来监测一个或多个非线性-活性标签蛋白质与拴系蛋白质的结合。在一些情况下,一个或多个其他蛋白质分子可以与拴系分子相同。在一些情况下,一个或多个额外的蛋白质分子可以与拴系分子不同或彼此不同。在一些情况下,一个或多个其他蛋白质分子中的至少一个可以是拴系分子的天然存在的配体或结合配偶体。在非栓系蛋白分子(或天然存在的配体)为非线性-活性物种的情况下,非线性-活性标签蛋白(或配体)与栓系蛋白的结合可被视为拴系蛋白质或蛋白质-蛋白质复合物的原位标签形式。在一些情况下,待研究的蛋白质或其他生物分子未标记且没有非线性-活性,但与其结合的底物或分子配体是非线性-活性标记的,例如atto390gtp:γ-(6-氨基己基)-gtp-atto-390γ-(6-氨基己基)-鸟苷-5'-三磷酸酯)。
在一些实施方式中,所公开的方法、装置和系统可用于筛选候选化合物文库,以鉴别稳定或破坏由于上文描述的蛋白质-蛋白质结合相互作用而在光学界面上形成的所得蛋白质-蛋白质复合物的化合物,例如,所公开的方法、装置和系统可用于筛选候选化合物文库,以鉴别使复合物与一种或多种候选化合物接触后稳定或破坏蛋白质-蛋白质复合物的化合物。在一些情况下,可以使用下文更详细描述的装置和系统以高通量方式对稳定或破坏蛋白质-蛋白质复合物的化合物执行这样的筛选。
两个或更多个样品的蛋白质结构比较:在一些实施方式中,所公开的非线性光学方法可用于两个或更多个蛋白样品(或其他生物分子样品)例如由相同制造过程在不同时间生产的两个或更多个蛋白质样品,或由不同制造过程生产的两个或更多个蛋白质样品,或由相同制造过程生产但随后经受不同实验条件的两个或更多个蛋白质样品的结构比较。例如,在一些实施方式中,可以使用相同的标签方案用shg-活性或其他非线性-活性标签标记蛋白质分子(例如,单克隆抗体(mab)药物或药物候选物)的两个或更多个样品,并使用相同的拴系方案将其拴系至光学界面,并且可以使用相同的光学仪器(在相同时间或不同时间,前提是光学仪器已根据可靠的参考标准进行了校准)执行shg基线信号(或shg与tpf信号的比率)的测量。然后,所得基线shg信号或shg与tpf信号的比率的比较可提供监测蛋白质结构、构象、取向分布或其差异的方法,并可用于确定两个蛋白质样品包含相同或基本相同的结构和构象的蛋白质。在一些实施方式中,所公开的方法、装置和系统可以用于通过相同制造过程在不同时间生产的至少两个、至少三个、至少四个、至少五个、至少十个、至少二十个、至少三十个、至少四十个、至少五十个、至少一百个或多于一百个的蛋白质样品的结构比较。在一些实施方式中,所公开的方法、装置和系统可以用于通过至少两种或多于两种不同制造过程生产的至少两个、至少三个、至少四个、至少五个、至少十个、至少二十个、至少三十个、至少四十个、至少五十个、至少一百个或多于一百个的蛋白质样品的结构比较。在一些实施方式中,所公开的方法、装置和系统可以用于通过相同的制造过程生产,但随后经受不同实验条件生产的至少两个、至少三个、至少四个、至少五个、至少十个、至少二十个、至少三十个、至少四十个、至少五十个、至少一百个或多于一百个的蛋白质样品的结构比较。
过程优化和质量控制:在一些实施方式中,公开的非线性光学方法可以用于过程优化和/或质量控制目的。通常,使用非线性光学技术进行过程优化和过程输出监测的方法(例如,如在质量控制应用或生物相似性证明中)将涉及:(i)例如,从相同过程的不同批次(例如,不同生产批次)或标称生产相同蛋白质产品的不同生产过程,在流程的同一步骤的不同时间、在流程的不同步骤收集蛋白质的一个或多个等分,(ii)使用标准化标签程序对蛋白质进行标记(如果需要),(iii)在标准的一组实验条件(例如缓冲液、ph、离子强度、洗涤剂浓度、温度等)下,使用标准化的绑定或固定程序,将标记的蛋白质绑定或固定在标准化的光学表面(例如,可进一步包含本领域技术人员已知的多种表面处理或修饰中的任一种的玻璃基板的表面)上,(iv)将光学基底(在一些实施方式中,可以将其结合到包含用于包含缓冲液、确定试剂或其他溶液的孔或腔室的装置中)放置在仪器中,该仪器被配置为以一个或多个基频的光提供照射,并检测由照射导致的非线性光学过程生成的光,(v)测量基线shg(或其他非线性光学)信号(或(shg与tpf信号的比率)以及(vi)将一个或多个样品等分试样彼此比较或与参考样品的基线shg信号或shg与tpf信号进行比较。在一些实施方式中,可以在拴系或固定在基底上之前或之后,将一个或多个蛋白质样品与测试化合物一起温育。在一些实施方式中,可以在拴系或固定在基底上之前或之后,将一个或多个蛋白质样品暴露于不同的实验条件下。在一些实施方式中,用于测量基线shg信号的光学系统还包括荧光检测通道,其可用于监测蛋白质或非线性-活性标签的固有荧光(或附接于蛋白质的附加荧光标签的固有荧光),并用于归一化固定化蛋白质表面密度的孔间(样品间)的变化。与常规结构表征技术(例如,x射线晶体学研究)相比,所公开的测量技术提供了相对快速且容易的方法来监测样品之间的蛋白质结构变化。此外,公开的测量技术提供了监测溶液中样品间蛋白质结构变化的方法。
该方法可用于任何需要监测和/或确认以重复的时间间隔或以不同的处理步骤获取的蛋白质样品的蛋白质结构相似性,或经受不同组实验条件的蛋白质样品的应用。在一些实施方式中,该方法可以包括实时监测蛋白质结构变化。如上所述,在一些实施方式中,该方法可以用于,例如在缓冲液配方的优化期间监测蛋白质的稳定性。在这些实施方式中,研究中的蛋白质保持拴系或固定化在基底表面(即,实验的固定参数),并在操纵其他实验条件(例如,缓冲液条件)以优化蛋白质稳定性的同时测量shg信号(或shg与tpf的信号比)。在其他实施方式中,例如,为了进行过程优化,在过程中的不同时间点或不同步骤(例如,蛋白质样品是实验的可变参数)收集一个或多个样品等分试样,并且在一组标准化的实验条件(例如,缓冲液ph值、离子强度、洗涤剂浓度、温度等)下,基线shg信号测量值(或shg到tpf信号比测量)被用于评估蛋白质的三级结构。例如,该方法可用于在执行给定的处理步骤之前和之后(例如,在冷冻或冻干步骤之前和之后,或在纯化过程中的一个或多个不同步骤中的每个步骤之后)评估蛋白质三级结构。在一些实施方式中,该方法可以用于例如在生物制品生产中出于质量控制目的在过程终点处监视生产过程输出。在这些后面的实施方式中,在相同的一组实验条件下(即,用于进行非线性光学信号测量的实验条件保持固定,并且在两次测量之间操纵蛋白质)。
实验方法的统计设计:在本公开内容的一些实施方式中,例如,所公开的非线性光学方法用于监测蛋白质稳定性和优化缓冲液制剂,或用于优化和监测生物药物制造过程,可以使用实验统计设计(sdoe)方法应用该方法。当所需的结果(例如,蛋白质稳定性在指定时间段内或一致的生物药物生产)构成复杂的“响应表面”的局部最大值时,sdoe可以通过对最少数目的离散实验测试条件进行实验测量来执行复杂的优化程序,该“响应表面”是许多不同实验输入参数(例如缓冲液ph值、离子强度、洗涤剂浓度、添加剂浓度、过程步骤持续时间等)的函数。
生物相似性的证明:在一些实施方式中,所公开的方法提供了直接比较生物仿制药药物候选物和参考药物的结构(或构象)的方法。在一些实施方式中,所公开的方法提供了用于直接比较在与结合生物仿制药药物候选物和参考药物的试剂接触后在生物仿制药药物候选物和参考药物中诱导的构象变化的方法。在一些实施方式中,所公开的方法提供了用于直接比较在与生物仿制药药物候选物和参考药物接触后在靶蛋白或其他生物实体中诱导的构象变化的方法。本文还公开了可以以高吞吐量方式实现这些方法的系统。
如上所述,可以利用shg的表面选择特性和相关的非线性光学技术(或shg与tpf的信号比)来确定非线性-活性部分的平均取向,并从而可以用于比较结构或检测拴系在界面上的生物分子的构象变化。例如,可以通过使用相同的标签反应用非线性-活性部分标记生物药物候选物和参考药物,使用相同的束缚方法将生物药物候选物和参考药物束缚在界面上以使它们在界面处具有净取向,并在用每种基频的光照射(例如,通过测量基线信号)后测量非线性活性标签生成的光的物理性质来执行生物(生物仿制药)药物候选物和参考药物的结构相似性。在一些实施方式中,可以测量基线信号比,例如shg与tpf基线信号强度的比率,并将其用于证明候选生物药与参考药物之间的结构相似性。对于生物仿制药药物候选物和参考药物测得的光的物理性质的统计学显著差异,例如,基线信号强度或基线信号强度比,可能指示其在结构上不等效,而被测光的物理性质的统计学显著差异可以指示其具有基本相同的结构。
shg和其他非线性光学技术的表面选择性特性还可以用来检测拴系在界面的生物分子的构象变化,因此可以用来进一步证明生物相似性。例如,可以使用非线性-活性标签或部分来检测由于配体(例如,生物药物候选物或参考药物)的结合而导致的靶蛋白分子的构象变化,其中标签附接或结合靶蛋白,使得构象变化导致标签相对于界面的取向或距离发生变化(图3),从而导致非线性光学信号的物理性质发生变化。证明靶蛋白在结合生物药物候选物或参考药物后经历了相同的构象变化,如shg信号(或shg与tpf信号的比率)的最终变化所指示的,将因此提供生物相似性的证据。
本文公开的方法和系统提供了用于对生物药物候选物(例如,单克隆抗体(mab))和参考生物药进行实时结构比较的手段,出于建立生物相似性的目的。所公开的方法和系统包括使用shg和相关的非线性光学技术来比较非线性-活性标签的生物药物候选物和参考药物的结构或构象,并在使非线性-活性标签的生物蛋白靶分子(例如,在mab药物或药物候选物的情况下为抗原)与一种或多种药物候选物或参考药物接触时监测蛋白质的构象变化,从而允许比较最终的构象变化(或“构象签名”),以建立等效性或差异性。在相同标签和栓系的药物候选物和参考药物中观察到相同或基本相同的结构或构象可能提供生物相似性的证据。对药物候选物和参考药物的相同或基本相同的构象变化或特征的观察可以表明相似的作用机理和有效性。对药物候选物和参考药物的不同构象变化或特征的观察可以指示不同的作用机制和/或不同的有效性水平。在shg信号强度(或shg与tpf信号强度的比率)的动力学测量中,靶分子的构象变化可以作为时间的函数进行监测,或者可以通过终点测量监测。因此,所公开的非线性光学确定技术使能够实时测定和比较生物药物候选物和参考药物的结构,以及实时测量和比较靶分子与候选生物药物或参考药物接触后引起的生物学靶标构象变化。
所公开的用于将生物药物候选物(例如,仿制药物候选物)与参考药物(例如,品牌药物)进行比较的方法可能比当前使用的许多结构表征技术对结构/构象差异更敏感,并且可以以各种不同的形式执行。例如,在第一实施方式中,可以用shg-活性或非线性-活性标签标记一个或多个生物药物候选物分子和参考药物分子,并通过本领域技术人员已知的多种手段中的任一种将其拴系在光学界面上。例如,对于全长mab,这可以通过与固定在表面上的蛋白质a或g分子的结合来实现。如果药物候选物分子(例如,仿制或生物仿制药)和参考药物分子(例如,品牌药物)在结构上是等效的,并且已经以相同的方式标记和拴系在界面上,则其应具有相同的基线信号。否则,shg或非线性光学特征的差异将为其差异提供结构性证据。在一些实施方式中,可以通过确定标记的生物药物候选物和参考药物的基线shg信号(或其他非线性光学信号)的测量之间的差异(如果有)的统计显著性,来评估结构相似性程度(或相反,结构相似性程度)。例如,在一些实施方式中,对于标记的生物药物候选物和参考药物的基线shg信号测量组,p值小于0.001、小于0.005、小于0.01、小于0.02、小于0.03、小于0.04或小于0.05可以指示测量的基线信号之间的差异显著不同,并且生物药物候选物和参考药物在结构上不等效。
在第二实施方式中,可以用shg-活性或其他非线性-活性标签标记靶分子(例如,mab药物或生物药物候选物的抗原),并通过本领域技术人员已知的各种手段中的任一种将其拴系到光学界面。然后,在使标记的靶标与一个或多个生物药物候选物或参考药物接触后,可以监测由靶分子诱导的构象变化引起的shg或其他非线性光学信号强度的变化(或非线性光学信号的其他物理性质)的变化。比较shg信号或shg与tpf信号的比率(签名)的结果变化,可以提供一种方法,用于确定药物候选物与参考药物在靶分子中的结合相互作用和/或构象变化方面的相似性。例如,在一些实施方式中,可以通过确定靶分子与候选生物药和参考药物接触后标记的靶分子shg信号(或shg与tpf信号的比率)的测量变化之间的差异(如果有)的统计学显著性来评估药物候选物与参考药物之间的结构相似程度(或者相反,结构相异程度)。例如,在一些实施方式中,对于标记靶分子的shg信号(或shg与tpf信号的比率)的测量变化集,p值小于0.001、小于0.005、小于0.01、小于0.02、小于0.03、小于0.04或小于0.05可指示所测信号变化的差异显著不同,并且候选生物药和参考药物在结构上不等效。
在第三实施方式中,可以用非线性-活性部分标记药物候选物分子(例如,mab药物候选物)和参考药物分子(例如,mab药物),并将其拴系到光学界面,并且当拴系的药物候选物和参考药物随后与生物靶分子(例如,在mab药物或药物候选物的情况下为抗原)接触时,可以监测shg信号(或shg与tpf信号的比率)。同样,如果两个药物分子相同(或基本相同),则将其与靶分子(例如,抗原)接触应引起相同的构象反应,如通过shg信号或shg与tpf信号的比率(签名)的相应变化所指示的。例如,在一些实施方式中,可以通过确定标记的药物候选物和参考药物与靶分子接触后,shg(或shg与tpf信号的比率)的测量变化之间的差异(如果有)的统计学显著性来评估药物候选物与参考药物之间的结构相似程度(或者相反,结构相异程度)。例如,在一些实施方式中,对于标记的生物药物候选物和参考药物的shg信号(或shg与tpf信号的比率)的测量变化集,p值小于0.001、小于0.005、小于0.01、小于0.02、小于0.03、小于0.04或小于0.05可能指示所测信号变化的差异显著不同,并且生物药物候选物和参考药物在结构上不等效。
在第四实施方式中,可以将标记的生物药物候选物(例如,mab药物候选物)或参考药物(例如,mab品牌药物)添加到拴系在表面上的未标记的靶蛋白(例如,抗原)。结合应产生标签的净、平均取向,并因此产生基线信号。如果药物候选物与品牌生物药物相同,则两者均应产生相同的基线shg或其他非线性光学信号。在一些实施方式中,可以通过确定标记的生物药物候选物和参考药物的基线shg信号(或其他非线性光学信号)的测量之间的差异(如果有)的统计显著性,来评估结构相似性程度(或相反,结构相似性程度)。例如,在一些实施方式中,对于标记的生物药物候选物和参考药物的基线shg信号测量组,p值小于0.001、小于0.005、小于0.01、小于0.02、小于0.03、小于0.04或小于0.05可以指示测量的基线信号之间的差异显著不同,并且生物药物候选物和参考药物在结构上不等效。
在以上公开的任何实施方式中,用于建立结构等效的方法可以进一步包括同时或连续测量双光子荧光(tpf)信号,以及出于将测得的非线性信号归一化为每单位界面面积的束缚分子数(即界面上拴系分子的表面密度或数密度)的目的,其用于计算shg:tpf信号比(或sfg:tpf信号比或dfg:tpf信号比)的用途。例如,在一些实施方式中,当用与用于产生二次谐波、和频或差频光的基频相同或不同的基频光照射时,在将生物药物候选物和参考药物拴系到界面之前,用于标记生物药物候选物和参考药物的非线性-活性(即shg-活性、sfg-活性或dfg-活性)标签也可能产生双光子荧光。在一些实施方式中,可以用不同于shg-活性、sfg-活性或dfg-活性标签的双光子荧光标签标记生物药物候选物和参考药物。由于双光子荧光信号与被激发的标签分子的数目线性相关,因此双光子荧光信号提供了一种标准化shg(或sfg或dfg)信号,以校正拴系分子表面密度的变化。在一些实施方式中,可以通过使用全内反射将基本光(即,通常由激光器提供的激发光)输送到界面来激发双光子荧光。在一些实施方式中,可以通过在与界面平面正交的方向(例如,使用落射荧光光学装置),或以不正交于界面平面的任意角度输送基光来激发双光子荧光。在一些实施方式中,可以使用落射荧光光学装置检测和测量在用基频光照射时激发的双光子荧光,例如,其中使用显微镜物镜收集发射的双光子荧光。在一些实施方式中,可以使用低-na针孔(即,不使用透镜)来检测和测量双光子荧光,该低-na针孔位于激发光聚焦和定向的点的正上方或正下方,使其与界面平面平行。然后,可以是使用光电倍增管或其他合适的检测器检测到通过会聚透镜、显微镜物镜或针孔(以及任何中间光学元件,例如附加透镜、反射镜、二向色反射镜、带通滤光片和/或光圈)的双光子荧光。pympo染料和类似物是合适的tpf染料,例如,pympo马来酰亚胺,其为1-(2-马来酰亚胺基乙基)-4-(5-(4-甲氧基苯基)恶唑-2-基)甲基磺酸吡啶鎓。
建立生物仿制药“指纹”:所公开的方法和系统可以因此用于建立生物药物候选物(即,生物仿制药药物候选物)相对于靶向多种治疗靶标中的任一种的参考药物的生物相似性。许多最近的出版物都强调了使用多种正交结构和功能表征技术以及收集“指纹状”比较数据组,以证明具有完全生物相似性的要求(参见,例如,greer,(2016),“biosimilarbreakdown”,theanalyticalscientist,issue0916-401;和declerck,(2013),“biosimilarmonoclonalantibodies:ascience-basedregulatorychallenge”,expertopin.biol.ther.13(2):153-156)。fda引入了生物仿制药“指纹”的概念,以确保生物仿制药开发者认真考虑用于证明生物仿制药和参考药物等效的技术。
临床和非临床数据均用于证明生物相似性,并且用于结构和功能表征的技术通常会从一种生物仿生药变化为另一生物仿生药。例如,在尝试证明治疗性单克隆抗体(mab)的生物相似性时,重要的是认识到其包含有助于其作用方式,并影响其临床性质的多个肽域declerck,(2013))。fab区包含负责与靶标的特异性结合相互作用的可变肽域。fc区在抗体依赖性细胞介导的细胞毒性(adcc)、补体依赖性细胞毒性(cdc)中起重要作用,并可能通过触发信号传导途径对细胞周期发挥其他一般调节作用。fc区被糖基化,并且糖基化的类型和程度影响效应子功能和清除率。fab区也被糖基化,其对功能的潜在影响不容忽视。因此,对生物相似mab的评估不仅应包括fab介导的抗原结合的特征,还应包括fc介导的功能(例如,与fcγr、fcrn、补体的结合)的表征。fab相关功能的表征不应局限于抗原结合的确定,还应包括对靶标预期功能作用的测试(例如,中和、受体阻断和受体活化)。由于该复杂性,证明单克隆抗体的生物相似性可能不仅需要体外结构/功能评估,还需要广泛的体内功能评估。根据欧洲药品管理局发布的指南(guidelineonsimilarbiologicalmedicinalproductscontainingmonoclonalantibodies-non-clinicalandclinicalissues,2012),证明mab生物相似性的第一步包括评估特定的结合和功能特征。需要进行体外表征研究,其中在以下方面对生物仿制药和参考药物进行比较:(i)与靶抗原的结合,(ii)与相关的三种fcγ受体(fcγri、fcγrii和fcγriii)、fcrn和补体(c1q)的代表性亚型结合,(iii)fab相关功能(例如,可溶性配体的中和、受体活化或阻断)以及(iv)fc相关功能(例如,adcc、cdc和补体活化)。对体内非临床测试的需求是基于体外表征数据的评估以及相关结构差异(例如,新转译后修饰)或mab药物候选物与参考药物之间的功能差异(例如,结合亲和力,fab-相关功能,或fc-相关功能)被确定。如果在体外表征数据中发现了关键差异,则可能需要进行相关的动物模型研究。
建立生物仿制药指纹可能需要的结构表征数据的示例可以包括,例如,一级结构(如通过质谱法或通过执行核酸测序确定的氨基酸序列)、更高阶的结构(包括二级、三级和四级结构(包括聚合)、酶促转译后修饰(如糖基化和磷酸化)和其他潜在的结构变异(如蛋白质脱酰胺和氧化)。可能还需要对故意的化学修饰(如peg化位点和特征)进行结构表征。
所公开的方法和系统除了将其用于建立生物相似性外,还可以用于建立生物仿制药的质量规格。ich主题q6b是人用药品注册技术要求国际协调会议的指南,该指南定义了制定生物药物产品质量规格的测试程序(greer,(2016),“biosimilarbreakdown”,theanalyticalscientist,issue0916-401)。提及了生物仿制药结构表征的六个规格要求:(i)氨基酸序列,(ii)氨基酸组成,(iii)末端氨基酸序列,(iv)肽图,(v)巯基和二硫键桥以及(vi)碳水化合物结构(如果适用)。生物仿制药的物理化学表征也有六个规格要求:(i)分子量或大小,(ii)亚型图谱,(iii)消光系数,(iv)电泳图谱,(v)液相色谱图以及(vi)光谱特征。表2列出了目前用于确定生物仿制药药物候选物和药物的结构和/或理化性质的不同表征技术和工具的示例。表中指示了本公开内容的非线性光学方法可适用的性质确定的非限制性示例。
表2.证明生物相似性的潜在方法(改编自greer(2016).
可以使用本领域技术人员已知的多种体外和/或体内功能测定法来评价生物药物候选物的药理活性(例如,有效性、效力和不良副作用的发生率)。可以使用的体外测定的示例包括但不限于,生物测定、结合测定、酶测定和基于细胞的测定(例如,细胞增殖确定或基于细胞的报告确定)。体内测定的示例可以包括使用动物模型研究,该动物模型研究使用疾病的动物模型(例如,表现出疾病状态或症状的模型)来评价药物候选物对药效标志物或功效度量的功能作用。使用这些功能测定数据将药物候选物与参考药物进行比较的功能评价是生物相似性证明的重要部分,并且可以进一步用于科学证明对人类患者进行动物和/或临床研究的选择性和针对性方法是合理的。
因此,在一些实施方式中,使用蛋白质结构和构象特征的非线性光学测量来比较生物药物候选物和参考药物的公开方法可以与其他结构和/或功能测定技术相结合,以提供更完整的生物相似性表征。在这些实施方式中,生物药物候选物和参考药物的非线性光学表征和比较可以与上述其他结构或功能测定法平行或串联进行,并可以根据结构等效性、构象特征和效价(例如,响应强度作为药物浓度的函数)、结合亲和力、结合特异性、反应动力学、其他结构表征数据(例如,圆二色性或晶体学数据)、对细胞内信号传导途径和/或基因表达谱的影响等,对药物候选物和参考药物进行比较。在一些实施方式中,药物候选物(或仿制药物)和参考药物(或品牌药物)可能基于一个或多个结构和/或功能特征而似乎是类似的,但基于一种或多种不同的结构和/或功能特性,可能似乎有所不同,并且本公开内容的方法可以允许证实或反驳生物相似性。
使用所公开的方法、装置和系统进行两个或更多个样品的结构比较、过程优化、过程监测或质量控制目的,不仅需要建立标准化的标签、拴系或固定以及测量方案,而且还需要制定标准,以比较在不同时间点、在过程的不同步骤或从不同的蛋白质批次中收集的蛋白质样品的基线shg信号(或shg与tpf信号的比率)并判断为“等效”。在许多情况下,该标准可能是蛋白质特异性的,可能需要通过进行比较研究来建立,这些比较研究使用本文所公开的shg基线信号测量技术(或shg与tpf信号比测量技术)以及其他结构或功能表征方法。
在一些实施方式中,例如,可能需要指定基线shg信号(或基线shg与tpg信号的比率)的可接受变化的范围,以便得出在不同时间,或在一个过程的不同步骤后,或从不同批次收集的蛋白质具有相同的结构的结论。在一些实施方式中,得出两个或更多个蛋白质样品具有等效结构所需的基线shg信号(或其他非线性光学信号)的最大可允许变化可以在约0.1%至约10%的范围内。在一些实施方式中,基线shg信号(或shg与tpf信号的比率)中的最大可允许变化可以为至少0.1%、至少0.25%、至少0.5%、至少1%、至少2%、至少3%、至少4%、至少5%、至少6%、至少7%、至少8%、至少9%或至少10%。在一些实施方式中,基线shg信号(或shg与tpf信号的比率)的最大可允许变化可以是至多10%、至多9%、至多8%、至多7%、至多6%、最多5%、最多4%、最多3%、最多2%、最多1%、最多0.5%、最多0.25%或最多0.1%。可以组合本段中所述的下限值和上限值中的任一个,以形成本公开内容中所包括的范围,例如,基线shg信号(或shg与tpf信号的比率)的最大可允许变化可以在约2%至约6%的范围内。本领域技术人员将认识到,基线shg信号(或shg与tpf信号的比率)的最大可允许变化可以具有在该范围内的任何值,例如,约4.5%。
在一些实施方式中,作为另一示例,可能需要要求收集两个或更多个待比较的蛋白质样品之间的经过时间的允许量在约1分钟至约1周的范围内。在一些实施方式中,收集两个或更多个待比较的蛋白质样品之间经过时间的允许量可以是至少1分钟、至少10分钟、至少20分钟、至少30分钟、至少40分钟、至少50分钟、至少1小时、至少2小时、至少3小时、至少4小时、至少5小时、至少6小时、至少12小时、至少18小时、至少1天、至少2天、至少3天、至少4天、至少5天、至少6天或至少7天。在一些实施方式中,收集两个或更多个待比较的蛋白质样品之间经过时间的允许量可以是至多7天、至多6天、至多5天、至多4天、至多3天、至多2天、至多1天、至多18小时、至多12小时、至多6小时、至多5小时、至多4小时、至多3小时、至多2小时、至多1小时、至多50分钟、至多40分钟、至多30分钟、至多20分钟、至多10分钟或至多1分钟。可以组合本段中所述的下限值和上限值中的任一个,以形成本公开内容中所包括的范围,例如,收集两个或更多个待比较的蛋白质样品之间经过时间的允许量可以在约10分钟至大约2个小时的范围内。本领域技术人员将认识到,收集两个或更多个待比较的蛋白质样品之间经过时间的允许量可以具有在该范围内的任何值,例如,约45分钟。
在一些实施方式中,在蛋白质样品的收集与基线shg信号测量(或其他基线非线性光学信号测量,例如,shg与tpf信号的比率)的执行之间的经过时间的最大量可以在约10分钟至约8小时的范围内。在一些实施方式中,在蛋白质样品的收集与基线shg信号测量(或shg与tpf信号比测量)的执行之间的经过时间的最大量可以是至少10分钟、至少20分钟、至少30分钟。分钟、至少40分钟、至少50分钟、至少1小时、至少2小时、至少3小时、至少4小时、至少5小时、至少6小时、至少7小时或至少8小时。在一些实施方式中,在蛋白质样品的收集与基线shg信号测量(或shg与tpf信号比测量)的执行之间的经过时间的最大量可以是至多8小时、至多7小时、至多6小时、至多5小时、至多4小时、至多3小时、至多2小时、至多1小时、至多50分钟、至多40分钟、至多30分钟、至多20分钟或至多10分钟。可以组合本段中所述的任何下限值和上限值中的任一个,以形成本公开内容中所包括的范围,例如,在蛋白质样品的收集与基线shg信号测量(或shg与tpf信号比测量)的执行之间的经过时间的最大量可以在约20分钟至约2小时的范围内。本领域技术人员将认识到,在蛋白质样品的收集与基线shg信号测量(或shg与tpf信号比测量)的执行之间的经过时间的最大量可以具有在该范围内的任何值,例如,约2.5小时。
在一些实施方式中,每个蛋白质样品获得两个或更多个蛋白质样品的可靠比较所需的数目或重复测量可以在约1个重复至约6个重复的范围内。在一些实施方式中,每个蛋白质样品所需的数目或重复测量可以是至少1次重复、至少2次重复、至少3次重复、至少4次重复、至少5次重复或至少6次重复。在一些实施方式中,每个蛋白质样品所需的数目或重复测量可以是至多6次重复、至多5次重复、至多4次重复、至多3次重复、至多2次重复或至多1次重复。可以组合本段中所述的下限值和上限值中的任一个,以形成本公开内容中包括的范围,例如,每个蛋白质样品所需的数目或重复测量的范围可以在约2个重复至约4个重复的范围内。本领域技术人员将认识到,每个蛋白质样品所需的数目或重复测量可以具有该范围内的任何值,例如,3次重复。
如上所述,在许多情况下,用于建立两个或更多个样品之间的结构等效性的标准可能是蛋白质特异性的,并且可能将需要通过进行比较研究来建立,该比较研究使用本文公开的shg基线信号测量技术和其他结构或功能表征方法。可以与公开的非线性光学测量技术结合执行的合适的结构表征技术的示例包括但不限于,圆二色性研究、核磁共振(nmr)研究、x射线晶体学研究、分子模型研究等。合适的功能表征研究的示例包括但不限于,配体结合测定、酶测定、免疫测定等。
检测到的其他类型的生物相互作用:除了确定或比较蛋白质与其他生物分子的取向或结构之外,本文公开的方法和系统还提供了对生物实体之间,或生物实体与测试实体之间的各种相互作用的检测,这取决于对生物实体、测试实体和所采用的非线性-活性标签技术的选择。一方面,本公开内容提供了结合事件的定性检测,例如,配体与受体的结合,如受体中诱导的构象变化所指示的。在另一方面,本公开内容提供了结合事件的定量分析,例如,通过使用不同浓度的配体分子进行重复测量,并使用观察到的最大构象变化的百分比生成剂量反应曲线,从而确定配体与受体的结合。类似地,本公开内容的其他方面可以提供用于定性或定量测量酶-抑制剂相互作用、抗体-抗原相互作用、生物大分子复合物的形成、受体与变构调节剂的相互作用、药物候选物-药物靶标相互作用、蛋白质-蛋白质相互作用、外周膜蛋白质-外周膜蛋白质相互作用、外周膜蛋白质-完整膜蛋白质相互作用、外周膜蛋白质-磷脂双层相互作用等。
可以通过目前公开的方法将生物实体或生物实体与测试实体之间的相互作用(例如,结合反应、构象变化等)与以下可测量的非线性信号参数相关联:(i)非线性光的强度,(ii)非线性光的波长或光谱,(iii)非线性光的偏振,(iv)(i)、(ii)或(iii)的时程,和/或vi)(i),(ii),(iii)和(iv)的一个或多个组合,以及通过测量信号比(例如,shg与tpf信号强度比)来进行。
绝对极性取向确定:尽管可以确定实验室系中的标签的倾斜角方向,但是这个倾斜角分别在指向和离开表面的两个圆锥中是简并的。本发明还公开了一种用于通过简单实验来获得标签的绝对方向(即标签相对于平面表面指向的方向)的新颖方法。在这个实验中,给定偏振条件下的shg信号使用如下项来测量:i)附接至未经标签表面的经标签蛋白质,ii)附接至经标签表面的未经标签蛋白质,和iii)附接至经标签表面的经标签蛋白质。所述标签表面可以以本领域技术人员公知的多种方式来制备,例如通过羧化的非线性-活性标签与氨基硅烷官能化的玻璃基底表面的共价碳二亚胺偶合。或者,利用受支承脂双层(slb),可以将经由胺或硫醇(例如,对应于赖氨酸或半胱氨酸残基侧链)与蛋白质附接的相同的非线性-活性标签与掺杂有不同摩尔百分比的含胺或硫醇的脂质的双层共价偶合。由于标签与蛋白质标签相同,所以该表面然后提供与蛋白质产生的shg信号同相的自身shg信号。凭借其已知的与表面的定向偶合及其化学结构,附接至受支承双层的标签具有已知的极性取向。确定蛋白质上标签的绝对极性取向(并因此可能是整个蛋白质的极性取向)的实验可以如下进行。首先,测量不存在蛋白质时经标签表面的shg信号(il)。其次,测量附接至未经标签表面的经标签蛋白质的shg信号(ip)。第三,当经标签蛋白质附接至经标签表面时,测量表面的shg信号(itot)。不同shg信号之间的关系如下:
itot=il+ip+2*sqrt(il*ip)*cos(θ)(7)
其中cos(θ)描述了附接至蛋白质分子的标签与第三次测量(itot)中的表面之间的相位关系(其随着朝向或远离表面的绝对极性取向而在符号上翻转)。通过分别测量itot、il和ip并比较所测量的信号强度,可以确定蛋白质上每个标签的绝对极性取向。在一些情况下,例如当il+ip具有大致相当的量级时,如果itot本身小于il,则可以立即确定在蛋白质标签与表面标签之间发生的相消干涉;因此,标签以相反的极性取向定向;如果itot本身大于il,则发生相长干涉,并且标签在相同极性方向上定向。il+ip的大小可以通过调节例如针对染料的受支承双层上附接位点的密度或附接至表面的蛋白质的密度来改变。以类似的方式,可以通过在不同的测量中排列不同比例的标签和未标签生物分子,同时保持每次测量中生物分子的总浓度恒定来确定附接到生物分子的染料探针与不存在生物分子的背景之间的相差。这样,背景与非线性-活性探针之间的干扰将产生强度,该强度取决于背景与染料探针产生的shg波之间的相差。例如,通过在孔中温育相同的总蛋白浓度,但改变标签和未标签蛋白的比例可以将其最简单地实现。每个孔应展示相同的蛋白质表面密度,但标签和未标签分子的比例将反映其在温育过程中的浓度比。因此,总的测量强度itot应取决于从背景信号(例如,表面+水+未标签的蛋白质)生成的shg波与标签的蛋白质信号之间的相差。通过以标签和未标签蛋白质的不同比率进行连续实验并测量itot,可以通过使用方程(7)确定背景与染料标签之间的干扰是相长还是相消来获得染料探针的相对取向。
在一些实施方式中,使用羧基化的非线性-活性标签与氨基硅烷官能化的玻璃基底表面的共价碳二亚胺偶联来制备非线性-活性标签的表面。在一些实施方式中,非线性-活性标签的表面包含受支承脂双层,并且其中受支承脂双层进一步包括与非线性-活性标签共价偶联的含胺或硫醇的脂质。
在一些实施方式中,使用蛋白质的c端与氨基硅烷官能化的玻璃基底表面的共价碳二亚胺偶联,以定向方式将非线性-活性标签的蛋白质拴系在非标签或非线性-活性标签的表面上。在一些实施方式中,将非线性-活性标签蛋白质以定向方式拴系在包含受支承脂双层的非标签或非线性-活性标签的表面上,并且其中将非线性-活性标签蛋白插入受支承脂双层中或附接至被插入受支承脂双层中的锚分子。下面将更详细地描述将标签蛋白质和其他生物分子拴系于基底表面或受支承脂双层的方法。
总之,所公开的用于确定附接于拴系蛋白质的非线性-活性标签的绝对取向的方法可以包括:(a)检测由于至少一个基频的激发光而由非线性-活性表面生成的光的物理性质,其中使用激发光的两种不同的偏振状态进行检测;(b)检测以定向方式拴系在非标签表面上的由非线性-活性标签蛋白生成的光的物理性质,其中光是由至少一个基频的激发光照射而生成的,并且其中使用激发光的两种不同的偏振状态进行检测;(c)检测由以定向方式拴系在非线性-活性标签表面上的非线性-活性标签蛋白生成的光的物理性质,其中光是由至少一个基频的激发光照射而生成的,并且其中使用激发光的两种不同的偏振状态进行检测;以及(d)通过比较步骤(a)中的光的物理性质、步骤(b)中检测到的光的物理性质和步骤(c)中检测到的光的物理性质,来确定附接在拴系蛋白上的非线性-活性标签的绝对取向。如果假定或已知取向宽度很窄,则可以使用在两个正交偏振状态(例如s-偏振和p-偏振)下测得的tpf强度的比率来确定标签的取向,即跃迁偶极矩与表面法线轴之间的角度。同样,可以通过取两个正交偏振下shg强度的比率来确定探针中的主要超极化分量与法线轴之间的角度。为了同时确定分布宽度和平均角度,可以分别求解两个平均角度的交点,即分别满足每个强度比(分别在p-偏振和s-偏振下的tpf和shg)的宽度轨迹,如下例所示。
电场取向、强度和特性:在一些实施方式中,可以施加电场来操纵界面处实验室系中的生物分子的取向。电场方向可以横跨表面,与其垂直,或者通常采取相对于表面平面的任何角度。在一个实施方式中,将一个电极置于脂双层膜或其他表面化学品的下面以将蛋白质与基底(例如玻璃基底)附接。将对电极置于基底表面平面的上方,例如在样品孔中液体的顶部。在另一实施方式中,将两个或更多个电极放置在基底表面平面内,并且电场方向与基底膜的界面平行。
在另一实施方式中,可将电极阵列放置在拴系的或固定的生物分子,例如,蛋白质样品的周围,如图4所示。例如,电极的圆形阵列可以平行于玻璃基底表面上的膜界面放置,每个大约彼此间隔10度。施加到彼此分开180度的一对电极上的电压允许电场的方位角方向以任意及快速的方式改变。例如,电场的方位角方向可以在一秒或几分之一秒内绕整个圆周扫描。
可以使用本领域技术人员已知的多种技术中的任一种在基底表面上对电极进行图案化。实例包括但不限于丝网印刷、光刻图案化、溅射镀膜、化学汽相沉积或其任何组合。
如本领域技术人员公知的,电极可以由多种材料中的任何一种制成。合适的电极材料的实例包括但不限于银、金、铂、铜、铝、石墨、铟锡氧化物(ito)、半导体材料、导电聚合物或其任何组合。
在一些实施方式中,可能需要钝化一个或多个电极的表面,例如,使与含水缓冲液接触的电极表面的腐蚀降至最低,并且/或者防止污染或干扰蛋白质或其他生物组分,并且/或者防止样品中的电流流动。可以使用本领域技术人员已知的多种钝化技术中的任一种,并且通常取决于用于制作电极的材料的选择。例如,玻璃基底上的铟锡氧化物电极可以通过生长或沉积30nm的sio2层来钝化。金属或半导体电极通常暴露于空气时会形成可充当钝化层的惰性“天然氧化物”层。这种惰性表面层通常是氧化物或氮化物,铂的单层厚度为
电场可以是dc或ac,即时间不变的或时间变化的。在后一种情况下,它可以采用任何频率的正弦波,或者可以是由许多频率分量组成的复合波(即阶跃函数、锯齿形等),并且该场在正值或负值之间振荡或者保持全部正值或全部负值。在一些实施方式中可以应用非周期的或脉冲电场。shg信号(或shg与tpf的信号比)可以在将电场施加到样品之前、期间或之后读取。
在一些实施方式中,所述电场强度可以在约零至约106v/cm或更大的范围内。在一些实施方式中,所述电场强度可以是至少零、至少10v/cm、至少102v/cm、至少103v/cm、至少104v/cm、至少105v/cm或至少106v/cm。在一些实施方式中,所述电场强度可以是至多106v/cm、至多105v/cm、至多104v/cm、至多103v/cm、至多102v/cm、至多10v/cm。本领域技术人员将认识到所述电场强度可以是在该范围内的任何值,例如,约500v/cm。
在一些实施方式中,电场改变的频率可以在约0hz至约105hz的范围内。在一些实施方式中,电场改变的频率可以是至少0hz、至少10hz、至少102hz、至少103hz、至少104hz或至少105hz。在一些实施方式中,电场改变的频率可以是至多105hz、至多104hz、至多103hz、至多102hz或至多10hz。本领域技术人员将认识到电场改变的频率可以是在该范围内的任何值,例如,约125hz。
所述电场可用于操纵蛋白质分子或其他生物分子的取向,从而操纵基线shg信号(或基线shg与tpf的信号比)或shg(或shg与tpf)偏振依赖性。在一些实施方式中,如果在不存在施加的电场的情况下在基底表面平面(即xy表面)中发生取向各向同性,并且在施加电场时保持各向同性,则非线性极化率(χ(2))的两个或三个独立的非零分量将存在。在表面平面上存在取向各向异性的其他实施方式中,在施加电场之前、期间或之后,将存在χ(2)的多于两个或三个独立的非零分量,允许用偏振基本光和二次谐波光的不同组合进行附加独立shg测量。在表面平面(例如与经标签蛋白质附接的脂质生物膜)上存在取向各向异性的一些实施方式中,可以在不同方位角进行多个独立的χ(2)的测量。例如,如果平行于表面施加电场,并且这引起蛋白质分子从各向同性平面到各向异性平面的取向分布的变化,则可以在相对于施加的电场方向的许多方位角方向上进行附加独立光学测量以确定分子的取向分布。
具有集成电极的光学多孔板:如将在下面详细讨论的,在一些实施方式中,本文描述的tpf和/或shg测量优选地使用微孔板形式来进行。通常,这些装置包括:(a)包括第一表面的基底,该第一表面进一步包括多个离散的区域,其中每个离散的区域进一步包括电极的图案化阵列和可选的受支承脂双层;以及(b)与基底的第一表面结合或与基底的第一表面集成的孔形成组件,使得每个非连续区域都包含在单个孔内。在使用384孔板(或其他微孔板或多室形式)的一些实施方式中,电极可以在孔壁的内部和附近的基底表面上被图案化,作为盖的一部分,该盖用于对孔、孔内的基底表面(其可以是玻璃)的其他地方,或允许施加电压以在样品上产生电场和光学读取tpf和/或shg信号二者的任何地方加以密封。
在一些实施方式中,多个孔包含受支承脂双层,其进一步包含非线性-活性标签的蛋白质(或其他生物实体)。在一些实施方式中,多个受支承脂双层进一步包括彼此相同的非线性-活性蛋白质。在一些实施方式中,多个受支承脂双层进一步包括两个或更多个受支承脂双层亚组(例如,位于多个孔的两个或更多个亚组中),并且其中每个受支承脂双层的亚组包括不同的非线性-活性蛋白质。在一些实施方式中,基底由选自玻璃、熔融二氧化硅、聚合物或其任何组合的光学透明材料制成。在一些实施方式中,电极的图案化阵列包括两个或更多个电极的阵列,所述电极在围绕受支承脂双层的基底表面被图案化。在一些实施方式中,电极的图案化阵列包括在孔形成单元的每个孔壁上图案化的两个或更多个电极的阵列。在一些实施方式中,电极的图案化阵列包括图案化在密封每个孔的盖上的至少一个电极。在一些实施方式中,孔形成单元包括96个孔。在一些实施方式中,孔形成单元包括384个孔。在一些实施方式中,孔形成单元包括1,536个孔。在一些实施方式中,装置进一步包括棱镜阵列,该棱镜阵列与基底的第二表面集成,并被配置为将激发光递送到基底的第一表面,使其从第一表面被全内反射。
具有半球形棱镜的光学多孔板:在一些实施方式中,特别是在分子的各向异性取向分布存在于表面的情况下,在相对于各向异性轴的不同方位角的方向上光学检测样品将是有用的。作为相对于光轴旋转样品的替代方案,光轴可以相对于固定样品旋转。为了实现这一点,可以使用放置在每个孔的底部或附近的半球形棱镜来将入射到棱镜上的入射光相对于孔以任意角度引导至含有分子的界面区域。在一些实施方式中,半球形棱镜与玻璃底多孔板中的基底结合或集成,如图5所示。所述半球形棱镜与所述多孔板光学接触,从而允许光束以最小损耗传输。在一些实施方式中,所述光学多孔板将包含384孔玻璃底板或其他玻璃底微孔板形式(即,本领域技术人员公知的标准微孔板形式)。
在一些实施方式中,包含与形成孔底部的玻璃基底结合或集成的半球形棱镜阵列的微孔板装置可进一步包含在每个孔内的玻璃基底的上表面上的图案化电极阵列,以便可在施加不同场强的电场时进行偏振tpf和/或shg测量。
构象和配体诱导的构象变化的测量:在一个实施方式中,在蛋白质内的一个或多个标签位点处测量配体诱导的构象变化(例如,局部构象变化)。在一个实施方式中,使用单位点半胱氨酸残基。偏振的基本光和非线性光测量的组合被用于确定添加配体之前或之后的χ(2)分量。例如,如果生物分子在表面上各向同性地定向,并且分子超极化率由单张量元素(例如αz’z’z’,或者在一些文献中等同地为βz’z’z’)控制,然后,可以测量在p-激发偏振和s-激发偏振(zzz和zxz)下产生的shg和tpf的强度比,从而得出两个独立的比例方程,这些方程仅取决于取向角(相对于表面法线或z轴的倾斜角φ)和取向分布。对于使用不同的单位点半胱氨酸突变体的任何数目的标签位点,可以重复该过程,并且可以确定局部或全局配体结合的结构的模型。在一些实施方式中,所述模型可选地并入x-射线晶体结构坐标或其他结构约束(例如来自nmr数据、小角x射线散射数据或本领域技术人员已知的任何其他测量值)。
在不同实验条件下的结构参数的确定和构象变化的检测:在一些实施方式中,可以通过在两组或更多组不同的实验条件下进行测量来促进使用tpf测量和/或使用tpf测量和/或shg、sfg或dfg测量或其比率来确定结构参数,如平均倾斜角或分布宽度,和/或检测标签生物分子的构象变化,其中两组或更多组不同的实验条件影响生物分子的结构参数或构象。
如本文所定义,“实验条件”是指在其下测量shg、tpf和/或其他非线性光学信号的任一组实验参数,其中由于基础分子取向分布的变化,该组实验中的一个或多个实验参数的变化条件导致tdm或χ(2)的测量值发生变化。换言之,在实验系中产生不同取向分布的不同组实验条件将产生不同的基线tpf和/或shg信号强度、不同的偏振依赖性、对相同配体结合事件的不同响应,或上述任一项或全部。在这些实验中,我们可以假定蛋白质结构和构象构图保持恒定,并从而从两个或更多个位置标签的生物分子(以不同角度整体全局地取向在表面上)获得几乎无限数目的独立数据点,以确定结构和构象景观。例如,对附接在受支承脂双层上的蛋白质施加电场可能会改变分子的基本取向分布,并因此可能会改变tdm或χ(2)的测量值。可用于定义一组实验条件的实验参数的其他示例包括但不限于,缓冲条件如ph、离子强度、洗涤剂含量和浓度、拴系附接位点(例如,通过使用n-端或c-端的his标记物)等。具体地,在不同且独立的实验条件下测得的tdm或χ(2)的独立值允许在实验参考系中获得潜在的分子取向分布(图2)。通过将这些不同的实验系测量相互关联,可以确定位于蛋白质参考系中两个或更多个不同标签位点的非线性-活性标签之间的相对角度差,以及取向分布的其他参数,如用于模拟分子取向分布的高斯分布的宽度。
在一些实施方式中,可以使用将标签蛋白质拴系于其上表面的一个或多个组合物来定义不同组的实验条件。例如,如果使用受支承脂双层来拴系和定向标签蛋白质,则双层的两个或更多个不同的脂质组成(例如,具有不同的静电荷密度或不同的摩尔脂质掺杂密度)可用于产生两个或更多个相同蛋白质的不同取向分布。例如,可以使用带有不同净电荷的头基的脂质(例如,两性离子、带正电和带负电的脂质头基)。在一些实施方式中,通过改变例如不同脂质组分的数目和/或其相对浓度来改变受支承脂双层的脂质组成可能是有利的。可用于形成受支承脂双层的脂质分子或可作为受支承脂双层的主要或次要成分插入的脂质分子的示例包括但不限于,二酰甘油、磷脂酸(pa)、磷脂酰乙醇胺(pe)、磷脂酰胆碱(pc)、磷脂酰丝氨酸(ps)、磷脂酰肌醇(pi)、磷脂酰肌醇磷酸(pip)、磷脂酰肌醇二磷酸酯(pip2)、磷脂酰肌醇三磷酸酯(pip3)、神经酰胺磷酸胆碱(鞘磷脂;sph)、神经酰胺磷酸基乙醇胺(鞘磷脂;cer-pe)、神经酰胺磷酸脂或其任何组合。在一些实施方式中,可以将包含镍-三氮三乙酸镍(ni-nta)部分的脂质分子用于通过his标记物的手段拴系蛋白质的目的。例如,双层可以以多种摩尔浓度掺入1,2-二油酰基-sn-甘油-3-[(n-(5-氨基-1-羧基戊基)亚氨基二乙酸)琥珀酰基](镍盐)。
在一些实施方式中,脂质双层的不同脂质组分的数目可以在1至10个或更多个的范围内。在一些实施方式中,不同脂质组分的数目可以为至少1个、至少2个、至少3个、至少4个、至少5个、至少6个、至少7个、至少8个、至少9个或至少10个。在一些实施方式中,不同脂质组分的数目可以为至多10个、至多9个、至多8个、至多7个、至多6个、至多5个、至多4个、至多3个、至多2个或至多1个。
在一些实施方式中,脂质双层的给定脂质组分的相对百分比可以在约0.1%至约100%的范围内。在一些实施方式中,给定脂质组分的相对百分比可以为至少约0.1%、至少约0.2%、至少约0.3%、至少约0.4%、至少约0.5%、至少约1%、至少约5%、至少约10%、至少约20%、至少约30%、至少约40%、至少约50%、至少约60%、至少约70%、至少约80%、至少约90%或至少约100%。在一些实施方式中,给定脂质组分的相对百分比可以为至多约100%、至多约90%、至多约80%、至多约70%、至多约60%、至多约50%、至多约40%、至多约30%、至多约20%、至多约10%、至多约5%、至多约1%、至多约0.5%、至多约0.4%、至多约0.3%、至多约0.2%或至多约0.1%。可以组合本段中所述的下限值和上限值中的任一个,以形成本公开内容中包括的范围,例如,脂质双层的给定脂质组分的相对百分比可以在约0.5%至约20%的范围内。本领域技术人员将认识到,脂质双层中给定脂质组分的相对百分比可以具有在该范围内的任何值,例如,约12.5%。
如上所述,在一些实施方式中,第一组实验条件可包括使用附接至蛋白质n端的his-标记物将蛋白质分子拴系至底物表面或受支承脂双层,并且至少第二组实验条件包括使用附接在c端的his-标记物来拴系蛋白质分子。借助于n端与c端his-标记物的附接通常会产生不同的取向分布,因此可用于定义不同的实验条件组,从而产生不同的tdm或χ(2)测量值。如果使用诸如his标记物的标记物来拴系蛋白质,则不同长度的his-标记物可能会产生不同的取向分布,因此可用于定义不同的实验条件组。在一些实施方式中,第一组实验条件可包括使用选自以下的第一his-标记物对蛋白质分子进行拴系:2xhis、4xhis、6xhis、8xhis、10xhis、12xhis和14xhis,以及至少第二组实验条件可包括使用长度不同于第一his-标记物的至少第二his-标记物来拴系蛋白质分子。
在一些实施方式中,用于将标签蛋白质拴系到包含附接有ni-nta部分的脂质的受支承脂双层上的his-标记物的长度范围可以在约1个his残基至约20个his残基或更多个的范围内。在一些实施方式中,his-标记物的长度可以为至少1个his残基、至少2个his残基、至少3个his残基、至少4个his残基、至少5个his残基、至少6个his残基、至少7个his残基、至少8个his残基、至少9个his残基、至少10个his残基、至少11个his残基、至少12个his残基、至少13个his残基、至少14个his残基、至少15个his残基残基、至少16个his残基、至少17个his残基、至少18个his残基、至少19个his残基或至少20个his残基。在一些实施方式中,his-标记物的长度可以为至多20个his残基、至多19个his残基、至多18个his残基、至多17个his残基、至多16个his残基、至多15个his残基、至多14个his残基、至多13个his残基、至多12个his残基、至多11个his残基、至多10个his残基、至多9个his残基、至多8个his残基、至多7个his残基、至多6个his残基残基、至多5个his残基、至多4个his残基、至多3个his残基、至多2个his残基,或至多1个his残基。
在一些实施方式中,第一缓冲液与至少第二缓冲液之间的差异可用于定义不同组的实验条件。在一些实施方式中,用于定义不同组实验条件的缓冲液之间的差异可以选自缓冲液类型、缓冲液ph、缓冲液粘度、离子强度、洗涤剂浓度、两性离子组分浓度、钙离子(ca2+)浓度、镁离子(mg2+)浓度、碳水化合物、牛血清白蛋白(bsa)、聚乙二醇或其他添加剂浓度、抗氧化剂和还原剂或其任何组合。不同的缓冲条件可能会改变分子的取向分布,并从而改变tdm或χ(2)的测量值。
举例而言,在公开的方法中使用的合适的缓冲液可以包括但不限于,磷酸盐缓冲盐水(pbs)、琥珀酸盐、柠檬酸盐、组氨酸、乙酸盐、tris、taps、mops、pipes、hepes、mes等。合适的缓冲液的选择通常取决于缓冲液的目标ph。通常,缓冲溶液的期望ph将在约ph6至约ph8.4的范围内。在一些实施方式中,缓冲液ph可以为至少6.0、至少6.2、至少6.4、至少6.6、至少6.8、至少7.0、至少7.2、至少7.4、至少7.6、至少7.8、至少8.0、至少8.2或至少8.4。在一些实施方式中,缓冲液ph可以为至多8.4、至多8.2、至多8.0、至多7.8、至多7.6、至多7.4、至多7.2、至多7.0、至多6.8、至多6.6、至多6.4、至多6.2或至多6.0。可以组合本段中所述的下限值和上限值中的任一个,以形成本公开内容中包括的范围,例如,缓冲液的ph可以在约6.2至约8.2的范围内。本领域技术人员将认识到,缓冲液ph可以具有该范围内的任何值,例如,约7.25。在一些情况下,缓冲溶液的ph可以在约4至约10的范围内。
在一些实施方式中,用于定义不同组实验条件的缓冲液的离子强度可包括使用一价盐(例如,nacl,kcl等)、二价盐(例如,cacl2、mgcl2等)、三价盐(例如,alcl3)或其任何组合。在一些实施方式中,用于定义不同组实验条件的缓冲液的离子强度可以在约0.0m至约1m或更高的范围内。在一些实施方式中,缓冲剂的离子强度可以为至少0.0m、至少0.1m、至少0.2m、至少0.3m、至少0.4m、至少0.5m、至少0.6m、至少0.7m、至少0.8m、至少0.9m或至少1.0m。在一些实施方式中,缓冲液的离子强度可以为至多1.0m、至多0.9m、至多0.8m、至多0.7m、至多0.6m、至多0.5m、至多0.4m、至多0.3m、至多0.2m或至多0.1m。可以组合本段中所述的下限值和上限值中的任一个,以形成本公开内容中包括的范围,例如,缓冲剂的离子强度可以在约0.4m至约0.8m的范围内。本领域技术人员将认识到,缓冲剂的离子强度可以具有在该范围内的任何值,例如,约0.15m。
用于缓冲剂制剂的合适的洗涤剂包括但不限于,两性离子洗涤剂(例如,1-十二烷酰基-sn-甘油-3-磷酸胆碱、3-(4-叔丁基-1-吡啶基)-1-丙磺酸盐、3-(n,n-二甲基肉豆蔻酰基铵)丙烷磺酸盐、3-(n,n-二甲基肉豆蔻酰基铵)丙烷磺酸盐、asb-c80、c7bzo、chaps、chaps水合物、chapso、ddmab、二甲基乙基铵丙烷磺酸盐、n,n-二甲基十二烷基胺n-氧化物、n-十二烷基-n,n-二甲基-3-铵-1-丙烷磺酸盐或n-十二烷基-n,n-二甲基-3-铵-1-丙烷磺酸盐)和阴离子、阳离子和非离子型洗涤剂。非离子型洗涤剂的示例包括聚(氧乙烯)醚和相关的聚合物(例如,
在一些实施方式中,缓冲液中洗涤剂的浓度可以在约0.01%(w/v)至约2%(w/v)的范围内。在一些实施方式中,缓冲液中洗涤剂的浓度可以为至少0.01%、至少0.05%、至少0.1%、至少0.5%、至少1.0%、至少1.5%或至少2%。在一些实施方式中,缓冲剂中洗涤剂的浓度可以为至多2%、至多1.5%、至多1.0%、至多0.5%、至多0.1%、至多0.05%或至多0.01%。可以组合本段中所述的下限值和上限值中的任一个,以形成本公开内容中包括的范围,例如,缓冲液中洗涤剂的浓度可以在约0.1%(w/v)至约1.5%(w/v)的范围内。本领域技术人员将认识到,缓冲液中洗涤剂的浓度可以具有该范围内的任何值,例如,约0.12%(w/v)。
在一些实施方式中,也可以使用与界面区结合并根据其浓度产生不同的蛋白质取向分布的缓冲添加剂,例如peg400、乙二醇等。在一些实施方式中,peg400(或任何其他缓冲添加剂如牛血清白蛋白(bsa)、聚乙二醇或其他添加剂的浓度、抗氧化剂和还原剂等)的浓度可以在约0.01%(w/v)至约10%(w/v)的范围内。在一些实施方式中,peg400(或任何其他缓冲添加剂)的浓度可以为至少0.01%、至少0.05%、至少0.1%、至少0.5%、至少1.0%、至少1.5%、至少2%、至少3%、至少4%、至少5%、至少6%、至少7%、至少8%、至少9%或至少10%。在一些实施方式中,peg400(或任何其他缓冲添加剂)的浓度可以为至多10%、至多9%、至多8%、至多7%、至多6%、至多5%、至多4%、至多3%、至多2%、至多1.5%、至多1.0%、至多0.5%、至多0.1%、至多0.05%或至多0.01%。可以组合本段中所述的下限值和上限值中的任一个,以形成本公开内容中包括的范围,例如,peg400(或任何其他缓冲添加剂)的浓度可以在约0.5%(w/v)至约5%(w/v)的范围内。本领域技术人员将认识到,peg400(或任何其他缓冲添加剂)的浓度可以具有该范围内的任何值,例如,约2.25%(w/v)。
在一些实施方式中,可以增加用于shg和/或tpf偏振测量的不同组实验条件的数目,以增加待采样的独立分子取向分布的数目以及可以进行测量的独立偏振测量的数目,从而可以提高角度测量的准确性和由此得出的蛋白质结构模型。在一些实施方式中,所使用的不同组实验条件的数目可以是至少2个、至少3个、至少4个、至少5个、至少6个、至少7个、至少8个、至少9个、至少10个、至少15个、至少20个、至少30个、至少40个、至少50个、至少60个、至少70个、至少80个、至少90个或至少100个。
非线性-活性标签和标签技术:如上所述,大多数生物分子并非本来就是tpf-活性或sh活性的。例外包括胶原蛋白,这是一种存在于大多数结构或承重组织中的结构蛋白质。shg显微术已在例如角膜等含胶原蛋白结构的研究中得到广泛应用。其他生物分子或实体则必须通过引入诸如标记物或标签等非线性-活性部分才成为非线性-活性的。在本发明中采用的标签是指可以共价或非共价结合到分子、粒子或相(例如,脂双层)以便使得由此产生的系统更具非线性光学活性的非线性-活性部分、标记物、分子或粒子。在分子、粒子或相(例如,脂双层)不具非线性-活性的情况下采用标签以使得系统具有非线性-活性,或者在已经具有非线性-活性的系统中采用标签以向该系统中增加额外的表征参数。可以向分子、粒子或其他生物实体预附接外源性标签,并且在用于本文所述方法之前,任何未结合或未反应标签从经标签实体分离出来。在本文公开的方法的特定方面,在将靶分子或生物实体固定在基底表面的非连续区域中之前,将非线性-活性部分体外附接至靶分子或生物实体。在本文公开的方法的另一方面,在将靶分子或生物实体固定在基底表面的非连续区域中之后,将非线性-活性部分附接至靶分子或生物实体。用非线性-活性标签标签生物分子或其他生物实体允许在经标签生物分子或实体与另一分子或实体(即,测试实体)之间的相互作用导致该生物分子或实体的取向或构象变化的情况下,利用表面选择性非线性光学技术以直接光学手段检测该相互作用。
适用于所公开的方法的非线性-活性标记物或标签的示例包括但不限于表3中列出的化合物及其衍生物。
表3.非线性-活性标记物的示例
在评价某一物质是否具有非线性-活性时,以下特性可以表明非线性-活性的潜力:大偶极矩差(分子的基态与激发态之间的偶极矩差)、荧光中的大斯托克斯位移,或者芳族或共轭结合特性。在进一步评价该物质时,实验者可以利用本领域技术人员已知的简单技术来确认非线性-活性,例如,通过检测来自非线性-活性物质所分布于的空气-水界面的shg检测来进行确认。
一旦已针对所进行的实验选择合适的非线性-活性物质,如果需要,可以将该物质缀合到生物分子或实体用于本文公开的表面选择性非线性光学方法。以下参考文献和其中的参考资料描述了用于由合成染料和许多其他分子产生经标签的生物实体的可行技术:gregt.hermanson,bioconjugatetechniques,academicpress,newyork,1996。通常,进行标签反应的重要考虑因素是反应的特异性和产率,应将其最大化以确保获得一致的、可再现的基线shg信号(或相关的非线性光学信号)。
在一些实施方式中,可以使用标准的共价结合化学方法,例如,将非线性-活性标签附接至蛋白质分子。使用与胺基、羧基、硫醇基等具有反应性的非线性-活性部分。在一些实施方式中,可能可选地需要对标签的蛋白质进行质谱分析,以严格鉴别蛋白质中标签的氨基酸残基的位置。
可以使用的合适的胺反应共轭化学的示例包括但不限于,涉及异硫氰酸酯、异氰酸酯、酰叠氮、nhs酯、磺酰氯、醛、乙二醛、环氧化物、环氧乙烷、碳酸盐、芳基卤、亚氨酸酯、碳二亚胺、酸酐和氟苯基酯基团的反应。合适的羧基反应共轭化学的示例包括但不限于,涉及碳二亚胺化合物例如水溶性edc(1-乙基-3-(3-二甲基氨基丙基)碳二亚胺·hcl)的反应。合适的巯基反应共轭化学的示例包括马来酰亚胺、卤代乙酰基和吡啶基二硫化物。
在一些实施方式中,非线性-活性标签是选自吡啶恶唑(pympo)、pympo马来酰亚胺、pympo-nhs、pympo-琥珀酰亚胺基酯(pympo-se)、6-溴乙酰基-2-二甲基氨基萘(badan)和6-丙烯酰基-2-二甲基氨基萘(acrylodan)的双光子荧光和/或二次谐波(sh)-活性标签。为了实现高度的测定可重复性(例如,如使用相同蛋白质样品进行重复测量所得到的紧密变异系数(cv)所示),应使制备用于tpg或shg信号测量的蛋白质样品所需的标签步骤(以及拴系或固定步骤)的数目最小化。例如,理想地,标签反应应对蛋白质的标签附接位点具有高度特异性,应具有较高的产量(即应产生1:1的化学计量标签),并且不需要后标签分离步骤。pympo马来酰亚胺是合适的非线性-活性标签,其对shg和tpf均具有活性。配有蛋氨酸-化学选择性化学的pympo类似物也是合适的。蛋氨酸-化学选择性化学在lin,等人(2017),“redox-basedreagentsforchemoselectivemethioninebioconjugation”,science355(6325):597-602中描述。
在一些实施方式中,非线性-活性标签通过蛋白质表面上的一个或多个巯基、胺基或羧基与蛋白质结合。在一些实施方式中,一个或多个巯基、胺基或羧基是天然巯基、胺基或羧基。在一些实施方式中,一个或多个巯基、胺基或羧基是工程化的巯基、胺基或羧基。用于工程设计标签附接位点的遗传工程和定点诱变技术是本领域技术人员公知的(参见,例如,edelheit,等人(2009),"simpleandefficientsite-directedmutagenesisusingtwosingle-primerreactionsinparalleltogeneratemutantsforproteinstructure-functionstudies",bmcbiotechnology9:61)。使用该方法,例如,在用非线性活性标记物标记之前,可以将包含巯基(即,巯基)、胺基或羧基的氨基酸残基(例如,半胱氨酸、赖氨酸、天冬氨酸或谷氨酸残基)放置于蛋白质中的精确位置。在优选实施方式中,工程化的标签位点包括用半胱氨酸残基取代天然氨基酸残基。通常,被氨基酸残基取代的标签位点可能是在正确折叠蛋白质时已知位于蛋白质表面的氨基酸序列位置处的氨基酸残基。然后可以使用本领域技术人员已知的多种测定中的任一种来测试突变和标签突变蛋白质的类似天然功能,例如,通过使用蛋白质的已知配体进行结合测定。在一些实施方式中,可以制备突变蛋白质系列,其中每个突变体包含附接在蛋白分子内不同位点的非线性-活性标记物。在一些实施方式中,突变蛋白质可以包含用于标记的单个氨基酸取代。在一些实施方式中,突变蛋白质可包含用于标记的两个或更多个氨基酸取代。在一些实施方式中,除工程化标签位点外突变蛋白质还可包含氨基酸取代(例如,赖氨酸、半胱氨酸、蛋氨酸、天冬氨酸或谷氨酸残基),该工程化标签位点用于通过合适的接头分子将标签蛋白拴系于基底表面或受支承脂双层上,如下面将更详细描述的。在一些实施方式中,非线性-活性标签是二次谐波发生(shg)-活性标签、和频发生(sfg)-活性标签、差频(dfg)-活性标签或双光子荧光(tpf)-活性标签。在一些实施方式中,非线性-活性标签是shg-活性和tpf-活性的。
在其他优选的实施方式中,使用本领域技术人员已知的多种体内或无细胞体外技术中的任一种可以将遗传工程技术用于在蛋白质的特定位点掺入非线性-活性的非天然氨基酸。参见,例如,cohen,等人(2002),"probingproteinelectrostaticswithasyntheticfluorescenceaminoacid",science296:1700-1703和美国专利号9,182,406。在一些实施方式中,可以将非线性-活性的非天然氨基酸残基掺入突变蛋白质家族中,该突变蛋白质包含在一个、两个、三个、四个或五个或更多个已知位点上的非线性-活性的非天然氨基酸取代。这样的蛋白质可以是工程化的、天然存在的、使用体外翻译方法制备的、在体内表达的并且通常通过本领域技术人员已知的各种方法中的任一种来产生。在一些实施方式中,非线性-活性的非天然氨基酸是l-anap、aladan或萘的另一衍生物。在一些实施方式中,可以测试将固有非线性-活性的非天然氨基酸残基掺入生物药物候选物中与参考药物相比,对药物候选物的结构或功能是否有任何有害作用,如果没有,则随后可以将其用作生物仿制药生产过程中质量控制的内在标签。
在所公开的方法和系统的一个特定方面,对其金属纳米颗粒和组件进行修饰,以获得生物非线性-活性标签。以下参考文献描述了金属纳米颗粒和组件的修饰:j.p.novak和d.l.feldheim,"assemblyofphenylacetylene-bridgedsilverandgoldnanoparticlearrays",j.am.chem.soc.122:3979-3980(2000);j.p.novak等"nonlinearopticalpropertiesofmolecularlybridgedgoldnanoparticlearrays",j.am.chem.soc.122:12029-12030(2000);vance,f.w.,lemon,b.i.和hupp,j.t.,“enormoushyper-rayleighscatteringfromnanocrystallinegoldparticlesuspensions",j.phys.chem.b102:10091-93(1999)。
在一些实施方式中,可以通过结合非线性-活性肽使靶蛋白(例如,药物靶蛋白、生物药物候选物、生物参考药物、药物靶蛋白等)具有非线性-活性,所述非线性-活性肽通过局部非共价力如静电相互作用、氢键、疏水相互作用和/或范德华相互作用或其任何组合,与蛋白质分子特异性地和/或可逆地结合。可以合成一个或多个标记有shg-活性、sfg-活性、dfg-活性和/或tpf-活性部分的肽,并使其与靶蛋白反应(在将靶蛋白拴系到光学界面之前或之后)并测试其以特定方式与靶蛋白结合的能力(例如,使用shg和/或tpf测量来确定结合肽上的非线性-活性部分的取向角分布的宽度,其中拴系靶蛋白分子的取向分布是从对拴系靶蛋白分子进行shg和/或tpf测量中独立获知的,该拴系靶蛋白分子已通过共价结合或通过非线性-活性非天然氨基酸的遗传掺入直接用非线性-活性标签标记)。在优选实施方式中,非线性-活性肽可以包含已知结合特定蛋白质域的肽序列。例如,文献(sugita,等人(2013),biochem.eng.j.79:33-40)已报道以高亲和力(分别对人igg-fc具有8.9×106m-1和6.5×106m-1)与小鼠或人igg分子的fc部分特异性结合的八聚物肽序列(例如,nkfrgkyk和narkfykg)。
一个非常有用的实施方式是标记肽、拟肽或小分子探针,例如,已知与抗体(可能是抗原的片段)中的互补决定区(cdr)结合的探针。例如,可以使用具有shg和/或tpf-活性的非天然氨基酸(例如,l-anap)或是氨基酸的pympo的变体在固相合成中制备这样的肽;例如,根据本领域技术人员已知的方法,可以使用pympo-nhs或pympo-马来酰亚胺在溶液中标记这样的肽。然后可以将这些标记的肽与本身被拴系在表面的抗体结合,该抗体例如是包含带有ni-nta的脂质的受支承脂双层膜。然后可将shg-活性肽与抗体接触以产生基线信号。例如,可以通过热、光或其他手段对抗体进行“应力处理”,并且可以将该应力样品的基线信号与未应力样品的基线信号进行比较。类似地,可以在生物过程监测设置中比较基线信号,以确保生物制品在由标签肽探测的区域保持恒定的结构。此外,可以以类似的方式比较仿制生物制品与品牌生物制品。在这样的情况下(和/或在其他各种情况下),可依赖于shg和/或tpf对取向和/或结构的高度灵敏度,以及使用本身已赋予shg和/或tpf活性的肽或一些其他配体标记感兴趣的靶蛋白质或分子如生物制品或抗体的能力。
在本文公开的方法和系统的另一方面,还可以通过向分子信标(即已被修饰以并入非线性-活性标签(或其调节剂)而非荧光团或猝灭剂的分子信标探针)引入非线性类似物来操控系统的非线性-活性。分子信标的这些非线性光学相似物在本文中称为分子信标类似物(mb类似物或mba)。要在所描述的方法和系统中采用的mb类似物可以根据本领域普通技术人员已知的规程予以合成。
在一些实施方式中,一个或多个非线性-活性标签可以附接于相同的单个生物分子(例如,蛋白质分子)内的一个或多个已知位置(例如,位点)。在一些实施方式中,可将一个或多个非线性-活性标签附接于相同蛋白质的不同分子中的一个或多个已知位置(例如,位点),即产生包含标签蛋白质的不同单标签形式的蛋白质家族。在一些实施方式中,标签蛋白质(或蛋白质家族)的标签位点的数目可以是至少1个位点、至少2个位点、至少3个位点、至少4个位点、至少5个位点、至少6个位点、至少7个位点、至少8个位点、至少9个位点、至少10个位点或更多。在一些实施方式中,非线性-活性标签可以附接于相同蛋白质的不同单位点半胱氨酸突变体或变体。在一些实施方式中,位于一个、两个、三个或更多个已知位置的非线性-活性标签相同。在一些实施方式中,位于一个、两个、三个或更多个已知位置的非线性-活性标签不同。在一些实施方式中,一个或多个非线性-活性标签是双光子活性标签。在一些实施方式中,一个或多个非线性-活性标签是双光子活性和/或以下中的一个或多个:二次谐波(sh)-活性标签、和频(sf)-活性标签或差频(df)-活性。
在一些实施方式中,shg和/或tpf测量可以包括使用被单个非线性-活性标签标记的蛋白质分子。在一些实施方式中,shg和/或tpf测量可以包括使用被至少2个不同的非线性-活性标签、至少3个不同的非线性-活性标签、至少4个不同的非线性-活性标签、至少5个不同的非线性-活性标签、至少6个不同的非线性-活性标签、至少7个不同的非线性-活性标签、至少8个不同的非线性-活性标签、至少9个不同的非线性-活性标签或至少10个不同的非线性-活性标签标记的蛋白分子。
在本文描述的方法和系统的替代方面,使用至少两个可区分的非线性-活性标签。然后将选择附接的两个或更多个可区分标签的取向,以促进发射相干非线性光束的明确限定方向。两个或更多个可区分的标签可用于这样的测定,其中使用相对于光学界面以一个或多个偏振方向入射的一个或多个频率的多个基本光束,从而产生至少两个非线性光束。在一些实施方式中,所使用的可区分的非线性-活性标签的数目可以是至少两个、至少三个、至少四个、至少五个、至少六个、至少七个、至少八个、至少九个或至少十个。
在一些实施方式中,至少两个可区分的非线性-活性标签可以与以以相对于光学界面的一个或多个偏振方向入射的一个或多个频率的多个基本光束组合使用,以确定至少两个可区分的非线性-活性标签相对于彼此或相对于光学界面的表面法线的相对或绝对倾斜角,从而促进蛋白质结构的映射(前提是蛋白质中每个可区分的非线性-活性标签的标签位点已知),或促进配体与蛋白质结合后局部构象变化的映射。
拴系和固定化学:如本文所公开,下述任何形式的基底被配置用于在全部或部分基底上拴系蛋白质或其他生物分子(或在一些情况下,细胞或其他生物实体)。在许多实施方式中,基底可倍配置用于在基底的指定非连续区域内拴系或固定蛋白质或其他生物分子。生物分子或细胞的拴系(有时在本文中称为“附接”或“固定”)可以通过本领域技术人员已知的多种技术来实现,例如,通过采用氨丙基硅烷化学以胺官能团使玻璃或熔融石英表面官能化,随后使用胺反应性缀合化学来直接与感兴趣的生物分子共价偶合或者经由中间间隔基或连接体分子来共价偶合。还可以直接或间接地例如通过利用bsa-nhs(bsa-n-羟基琥珀酰亚胺)而采用非特异性吸附,具体是首先将bsa分子层附接到表面,继而用n,n'-二琥珀酰亚胺基碳酸酯将其活化。bsa上的活化赖氨酸、天冬氨酸或谷氨酸残基与蛋白质上的表面胺类反应。
受支承脂双层的使用:在本公开内容的一个优选方面,生物分子可以通过拴系到或嵌入于“受支承脂双层”中而拴系到表面上,后者包含通过疏水或静电相互作用约束于硅或玻璃表面的脂双层小片,其中所述双层“浮”在含水缓冲液薄层上的基底表面上方。还可以使用或不使用膜蛋白或其他膜相关成分来制备受支承磷脂双层,例如在以下文献中所描述:salafsky等人“architectureandfunctionofmembraneproteinsinplanarsupportedbilayers:astudywithphotosyntheticreactioncenters",biochemistry35(47):14773-14781(1996);gennis,r.,biomembranes,springer-verlag,1989;kalb等人“formationofsupportedplanarbilayersbyfusionofvesiclestosupportedphospholipidmonolayers”,biochimicabiophysicaacta.1103:307–316(1992);以及brian等人"allogeneicstimulationofcytotoxict-cellsbysupportedplanarmembranes",pnas-biologicalsciences81(19):6159-6163(1984);上述文献的相关部分通过引用而并入于此。受支承磷脂双层是本领域中公知的,并且有多种技术可以用于其制造。使用受支承脂双层将蛋白质或其他生物实体固定在基底表面或光学界面上的潜在优势包括(i)保留通常跨越细胞膜或细胞的其他膜成分并需要与双层的疏水内芯相互作用的那些蛋白质的膜蛋白质结构以稳定二级和三级结构,(ii)保留二维横向和转动扩散迁移率以研究该双层内蛋白质组分之间的相互作用,和(iii)保持分子取向,这取决于诸如研究中的蛋白质类型(即,膜或可溶性蛋白质)、双层膜如何在基底表面形成,以及蛋白质如何拴系在该双层上(在可溶性蛋白质的情况下)。具有或不具有拴系或嵌入的蛋白质的受支承双层通常应当浸没在水溶液中以防止其在暴露于空气时受损。
在公开的方法和装置的一些实施方式中,改变被受支承脂双层的脂质组成,例如不同脂质组分的数目和/或其相对浓度,可能是有利的,以改善蛋白质分子(例如,外周膜蛋白)的结合、保留膜或外周膜蛋白的天然结构和/或模拟体内观察到的生理反应。可用于形成受支承脂双层的脂质分子或可作为受支承脂双层的主要或次要成分插入的脂质分子的示例包括但不限于,二酰甘油、磷脂酸(pa)、磷脂酰乙醇胺(pe)、磷脂酰胆碱(pc)、磷脂酰丝氨酸(ps)、磷脂酰肌醇(pi)、磷脂酰肌醇磷酸(pip)、磷脂酰肌醇二磷酸酯(pip2)、磷脂酰肌醇三磷酸酯(pip3)、神经酰胺磷酸胆碱(鞘磷脂;sph)、神经酰胺磷酸基乙醇胺(鞘磷脂;cer-pe)、神经酰胺磷酸脂、胆固醇或其任何组合。
在一些实施方式中,受支承脂双层的不同脂质组分的数目可以在1至10个或更多个的范围内。在一些实施方式中、不同脂质组分的数目可以为至少1个、至少2个、至少3个、至少4个、至少5个、至少6个、至少7个、至少8个、至少9个或至少10个。在一些实施方式中,不同脂质组分的数目可以为至多10个、至多9个、至多8个、至多7个、至多6个、至多5个、至多4个、至多3个、至多个2或至多1个。
在一些实施方式中,受支承脂双层的给定脂质组分的相对百分比可以在约0.1%至约100%的范围内。在一些实施方式中,给定脂质组分的相对百分比可以为至少约0.1%、至少约0.2%、至少约0.3%、至少约0.4%、至少约0.5%、至少约1%、至少约5%、至少约10%、至少约20%、至少约30%、至少约40%、至少约50%、至少约60%、至少约70%、至少约80%、至少约90%或至少约100%。在一些实施方式中,给定脂质组分的相对百分比可以为至多约100%、至多约90%、至多约80%、至多约70%、至多约60%、至多约50%、至多约40%、至多约30%、至多约20%、至多约10%、至多约5%、至多约1%、至多约0.5%、至多约0.4%、至多约0.3%、至多约0.2%或至多约0.1%。本领域技术人员将认识到,在受支承脂双层中给定脂质组分的相对百分比可以具有在该范围内的任何值,例如,约12.5%。
在其中受支承脂双层包含两个或更多个不同脂质组分的实施方式中,两个或更多个不同脂质组分的相对百分比可以相同也可以不同。在一个非限制性示例中,受支承脂双层可以包含25%的ps、74.5%的pc和0.5%的丽丝胺罗丹明pe。在另一非限制性示例中,受支承脂双层可以包含5%的pip、20%的ps、74.5%的pc和0.5%的丽丝胺罗丹明pe。对于一些类型的脂质,形成稳定的受支承脂双层的要求可能会将双层中该脂质的相对百分比限制为小于100%。在这些情况下,去稳定脂质组分的相对百分比可以通常在约1%至约50%的范围内。在一些实施方式中,在受支承脂双层中去稳定脂质组分的相对百分比可以为至少约1%、至少约5%、至少约10%、至少约20%、至少约30%、至少约40%或至少约50%。在一些实施方式中,在受支承脂双层中去稳定脂质组分的相对百分比可以为至多约50%、至多约40%、至多约30%、至多约20%、至多约10%、至多约5%或至多约1%。本领域技术人员将认识到,受支承脂双层中的去稳定脂质组分的相对百分比可以具有该范围内的任何值,例如,约12.5%。
如上所述,受支承脂双层也可包含靶蛋白或其亚基、亚域或片段。在一些实施方式中,受支承脂双层还可以包括例如,通过共价或非共价偶联至自身插入脂质双层的脂质样或疏水部分而拴系至脂质双分子层的非整合蛋白组分。
在一些实施方式中,包含在受支承脂双层中的不同蛋白质组分(整合或非整合)的数目可以在约1至约10个或更多个的范围内。在一些实施方式中,不同蛋白质组分的数目可以为至少1个、至少2个、至少3个、至少4个、至少5个、至少6个、至少7个、至少8个、至少9个或至少10个。在一些实施方式中,不同蛋白质组分的数目可以为至多10个、至多9个、至多8个、至多7个、至多6个、至多5个、至多4个、至多3个、至多2个或至多1个。
在一些实施方式中,脂质双层的给定蛋白质组分的摩尔分数可以在约0.1至约1的范围内。在一些实施方式中,给定蛋白质组分的摩尔分数可以为至少约0.1、至少约0.2、至少约0.3、至少约0.4、至少约0.5、至少约0.6、至少约0.7、至少约0.8、至少约0.9或至少约1。在一些实施方式中,给定蛋白质组分的摩尔分数可以为至多约1、至多约0.9、至多约0.8、至多约0.7、至多约0.6、至多约0.5、至多约0.4、至多约0.3、至多约0.2或至多约0.1。本领域技术人员将认识到,脂质双层中给定蛋白质组分的摩尔分数可具有该范围内的任何值,例如,约0.15%。
锚分子、接头和附接化学:可以使用许多不同的锚分子、接头和/或附接化学物质,以定向方式将可溶性蛋白质和其他生物实体拴系或附接至受支承脂双层(或直接到达包含光学界面的基底)。如本文所用的,“锚分子”是嵌入至脂双层的分子,并且可以包含脂肪酸、甘油酯、甘油磷脂、鞘脂类或缀合附接部分的其他脂质或非脂质分子。
接头分子是用于提供蛋白质或其他被拴系的生物实体的附接点与嵌入脂双层平面内的锚分子上的附接点之间的空间(“垂直”)分离的分子。在一些实施方式中,接头分子可用于在蛋白质或其他被拴系的生物实体的附接点与直接在包含光学界面的基底上的附接点之间提供空间分离。合适的接头分子的实例包括但不限于ω-氨基脂肪酸、聚乙二醇等。
附接部分(也称为“亲和标签”)是提供两个生物实体之间的共价或非共价结合的特定化学结构或结合配偶体。适用于本文公开的方法的附接部分或亲和标签的实例包括生物素和抗生物素蛋白(或生物素和链霉亲和素)以及his-标记物/ni-nta结合配偶体。
高亲和力、非共价生物素-链霉亲和素相互作用被广泛用于生物确定技术以缀合或固定蛋白质或其他生物实体。蛋白质的生物素化作用使得能够通过自身粘附于表面(例如,载玻片或珠子)或与另一分子缀合的多价抗生物素蛋白或链霉亲和素分子(例如,通过使用生物素-链霉亲和素-生物素桥或接头)捕获。该生物素部分足够小,生物素化作用通常不会干扰蛋白质的功能。生物素与抗生物素蛋白或链霉亲和素之间的结合相互作用的高亲和力(kd为10-14m至10-15m)和高特异性使得即使从复杂样品中也能捕获感兴趣的生物素化的蛋白质。由于极强的结合相互作用,需要苛刻的条件从链霉亲和素涂覆的表面洗脱生物素化的蛋白质(通常用6m,ph1.5的盐酸胍),这通常会使感兴趣的蛋白质变性。使用具有降低的约10-8m的生物素结合亲和力的抗生物素蛋白或链霉亲和素的单体形式可以允许生物素化的蛋白质在必要时用过量的游离生物素洗脱。在本文公开的方法中,为了将生物素化的蛋白质和/或其他生物素化的生物实体经由生物素-抗生物素蛋白-生物素(或生物素-链霉亲和素-生物素)桥固定或拴系在该双层内,可以将包含生物素部分的脂质分子并入到受支承脂双层中。
蛋白质和其他生物实体的生物素化可以通过直接偶合来进行,例如通过使用n-羟基琥珀酰亚胺基生物素(nhs-生物素)将蛋白质表面上的伯胺缀合。或者,使用avitag方法可以方便地对重组蛋白进行生物素化,其中通过使用遗传工程和蛋白质表达技术将avitag肽序列(glndifeaqkiewhe)掺入到蛋白质中。avitag序列的存在允许通过用bira酶处理来进行蛋白质的生物素化。
his-标记物化学是用于纯化重组蛋白和其他生物分子的另一个广泛使用的工具。在该方法中,例如,可以将指定六至九个组氨酸残基链的dna序列并入到用于产生包含与其n-端或c-端融合的6xhis或聚his-标记物的重组蛋白的载体中。在一些实施方式中,靶蛋白或其他生物蛋白可以被工程化以包括与包含ni-nta部分的双层脂质结合的,例如,2x-his标记物、3x-his标记物、4x-his标记物、5x-his标记物、6x-his标记物、7x-his标记物、8x-his标记物、9x-his标记物、10x-his标记物、11x-his标记物或12x-his标记物。
然后可以纯化并检测his-标记物的蛋白质,这是由于在特定的缓冲条件下,组氨酸残基链与若干类型的固定金属离子(包括镍、钴和铜)结合。支持体如琼脂糖小球或磁性颗粒可以用螯合基团衍生以固定所需的金属离子,然后其作为配体用于结合和纯化感兴趣的his-标记物的生物分子。
最常用于产生his-标记物配体的螯合剂是次氮基三乙酸(nta)和亚氨基二乙酸(ida)。一旦制备了nta或ida缀合的支持体,就可以用所需的二价金属(例如,ni、co、cu或fe)来“加载”它们。例如,当使用镍作为金属时,得到的亲和支持体通常称为ni-螯合物、ni-ida或ni-nta支持体。his-标记物的融合蛋白的亲和纯化是蛋白质生物学研究中金属螯合物最常见的应用。通过nta-螯合化学固定的镍或钴金属是针对该应用的选择系统。在本文公开的方法中,可以将包含ni-nta基团(或其他螯合的金属离子)的脂质分子掺入到受支承脂双层中以用于将his-标记物的蛋白质和其他his-标记物的生物实体固定或拴系在该双层中。在一些实施方式中,所述受支承脂双层可以包含1,2-二油酰基-sn-甘油基-3-磷酸胆碱,并且也可以包含各种浓度下的1,2-二油酰基-sn-甘油基-3-[(n-(5-氨基-1-羧基戊基)亚氨基二乙酸)琥珀酰基](镍盐)。
在接近中性的缓冲液条件(生理ph和离子强度)下,聚-his-标记物与螯合的金属离子最佳结合。典型的结合/洗涤缓冲液由含有10-25mm咪唑的ph7.2的tris-缓冲盐水(tbs)组成。低浓度的咪唑有助于防止具有组氨酸簇的内源性蛋白质的非特异性结合。在需要时,从螯合的金属离子支持体上洗脱并回收所捕获的his-标记物的蛋白质通常使用高浓度的咪唑(至少200mm)、低ph(例如0.1mph2.5的甘氨酸-hcl)或过量的强螯合剂(例如edta)来实现。已知免疫球蛋白在其fc区具有多个组氨酸,并可以与螯合的金属离子支持体结合,因此如果免疫球蛋白与感兴趣的his-标记物的蛋白质相比以较高的相对丰度存在于样品中,则严格的结合条件(例如,使用适当浓度的咪唑)是避免高水平的背景结合所必需的。白蛋白如牛血清白蛋白(bsa)也具有多个组氨酸,并可以在不存在丰度更高的his-标记物的蛋白质或在结合/洗涤缓冲液中使用咪唑的情况下产生与螯合的金属离子支持体的高水平的背景结合。
在一些实施方式中,用ni/nta或其他金属离子螯合剂衍生的基底表面可用于固定缺少his-标记物的蛋白质。例如,单克隆抗体(mab)与ni/nta表面接触后将会很容易与其缔合。关于在金属离子螯合剂衍生化的表面上固定蛋白质的附加示例参见block,等人(2009),“immobilized-metalaffinitychromatography(imac):areview”,inmethodsinenzymology,第463卷,第27章,elsevier。
在一些实施方式中,靶蛋白(例如,药物靶蛋白、生物药物候选物、生物参考药物、药物靶蛋白等)可以是已经过遗传工程化以掺入独特的拴系或固定位点以将蛋白质附接至光学界面的蛋白质,和/或已经过遗传工程化以掺入充当将蛋白质附接至光学界面的独特的拴系或固定位点的非天然氨基酸残基的蛋白质。可以遗传-掺入的独特的拴系位点或固定位点的示例包括但不限于,在正确折叠蛋白质时已知位于蛋白质表面上的氨基酸序列位置处掺入赖氨酸、天冬氨酸或谷氨酸残基。然后可以使用本领域技术人员已知的多种共轭和接头化学中的任一种将蛋白质拴系或固定在光学界面上。另一可以遗传-掺入到蛋白质产品中的独特拴系位点或固定位点的非限制性示例可以是可随后提供附接位点以供与附接至光学界面的ni/nta基团结合的his标记物(即,1、2、3、4、5、6、7、8、9、10、11、12个或多于12个的组氨酸残基系列)。可以掺入以提供独特附接点的非天然氨基酸的一个非限制性示例是生物素化的非天然氨基酸生物胞素。然后可以使用高亲和力的生物素-链霉亲和素相互作用将蛋白质拴系到或固定在光学界面上,以将蛋白质拴系至固定在基底表面上的链霉亲和素分子。
拴系后的漂洗/洗涤:在一些实施方式中,避免在蛋白质拴系或固定步骤之后漂洗或洗涤基底表面可能是有利的,即,尚未被拴系或固定的任何残留蛋白质均可留在用于使蛋白质样品与基底表面接触的孔、反应室或隔室中。由于所公开的测量技术的表面选择性,及其对蛋白质分子在光学界面处的净取向的依赖性,因此溶液中任何残留的标签蛋白质向所测量的非线性光学信号提供的贡献很小或完全没有贡献。
基底表面上结合位点的饱和:通常,待分析的一个或多个样品中蛋白质的浓度应足够高,以确保在用于拴系或固定的一组温育条件下基底表面上结合位点的饱和。这是为了使用于基线非线性光信号测量的样品制备的一致性最大。在一些实施方式中,一个或多个待分析样品中蛋白质的浓度相同,并且可以与参考样品中的浓度相同。在一些实施方式中,例如,如果样品等分试样中的蛋白质浓度较低,则可能期望提供具有较低结合位点密度的基底和/或使用更长的温育时间以确保结合位点的饱和。
改变拴系分子的表面密度:在其中期望改变基底表面上蛋白质结合位点的表面密度的实施方式中,可以以本领域技术人员已知的多种方式来实现表面结合位点密度的控制。例如,在其中蛋白质通过使用氨基丙基硅烷化学方法偶联到表面以使具有胺官能团的玻璃或熔融二氧化硅表面官能化,然后使用胺反应性偶联化学试剂和接头分子进行共价偶联的实施方式中,可以改变反应混合物中双官能接头分子(例如,包含伯胺和羧基官能团的接头)与单官能接头分子(例如,仅包含羧基官能团)的比率,以控制可用于与蛋白质偶联的伯胺官能团的表面密度。拴系的(标签的)分子的表面密度也可以通过简单地在溶液中以不同浓度的分子温育来改变。
作为另一示例,在其中使用生物素-链霉抗生物素蛋白结合相互作用将生物素化的蛋白质拴系到掺入受支承脂双层中的生物素化的脂质分子(经由生物素-链霉亲和素-生物素桥)的实施方式中,为了控制可用于结合的生物素基团的表面密度,可以改变形成双层的生物素化脂质分子的摩尔百分比。
作为又一示例,在其中使用包含ni-nta(或其他螯合的金属离子)配体的锚定脂质分子将his-标记物的蛋白固定在受支承脂双层上的实施方式中,为了形成可用于结合的ni-nta配体的表面密度,可以改变形成双层的含ni-nta的脂质分子的摩尔百分比。
在一些实施方式中,可以通过改变包含胺基或硫醇基(或可获得的标准缀合化学的任何其他官能团)的双层的脂质组分的百分比来改变受支承脂双层上附接位点的密度。在一些实施方式中,包含胺基或硫醇基的脂质组分的百分比可在约0%至约100%的范围内。在一些实施方式中,包含胺基或硫醇基的脂质组分的百分比可以为至少0%、至少10%、至少20%、至少30%、至少40%、至少50%。至少60%、至少70%、至少80%、至少90%或至少100%。在一些实施方式中,包含胺基或硫醇基的脂质组分的百分比可以为至多100%、至多90%、至多80%、至多70%、至多60%、至多50%、至多40%、至多30%、至多20%或至多10%。本领域技术人员将认识到,包含胺基或硫醇基的脂质组分的百分比可具有该范围内的任何值,例如,约12%。
在一些实施方式中,可以通过改变与受支承脂双层温育的溶液中标签蛋白质的浓度来改变附接至表面的非线性-活性标签蛋白质的密度。例如,在与受支承脂双层一起温育的溶液中,可以改变his标记的、标签的蛋白质的浓度,该脂质双层包含进一步包含ni-nta部分的脂质组分。在一些实施方式中,溶液中标签蛋白质的浓度可以在约1nm至约100μm的范围内。在优选的实施方式中,溶液中标签蛋白质的浓度可以在约100nm至约5μm的范围内。在一些实施方式中,溶液中标签蛋白质的浓度可以为至少1nm、至少10nm、至少100nm、至少1μm、至少10μm或至少100μm。在一些实施方式中,溶液中标签蛋白质的浓度可以为至多100μm、至多10μm、至多1μm、至多100nm、至多10nm或至多1nm。本领域技术人员将认识到,溶液中标签蛋白质的浓度可以具有该范围内的任何值,例如,约12μm。
在一些实施方式中,可以使用本领域技术人员已知的多种技术中的任一种在约102分子/cm2至约1014分子/cm2的范围内改变表面上的非线性-活性标签蛋白的密度。在一些实施方式中,表面上的非线性-活性标签蛋白的密度可以为至少102分子/cm2、至少103分子/cm2、至少104分子/cm2、至少105分子/cm2、至少106分子/cm2、至少107分子/cm2、至少108分子/cm2、至少109分子/cm2、至少1010分子/cm2、至少1011分子/cm2、至少1012分子/cm2、至少1013分子/cm2或至少1014分子/cm2。在一些实施方式中,表面上的非线性-活性标签蛋白的密度可以为至多1014分子/cm2、至多1013分子/cm2、至多1012分子/cm2、至多1011分子/cm2、至多1010分子/cm2、至多109分子/cm2、至多108分子/cm2、至多107分子/cm2、至多106分子/cm2、至多105分子/cm2、至多104分子/cm2、至多103分子/cm2或至多102分子/cm2。本领域技术人员将认识到,表面上的非线性-活性标签蛋白的密度可以具有该范围内的任何值,例如,约4.0×1012分子/cm2。
高通量方法、装置和系统:本文还公开了基于使用二次谐波或相关的非线性光学检测技术或基于使用双光子荧光或其任何组合,用于实现高通量分析和生物分子(例如,蛋白质或其他生物实体)中结构、构象或构象签名的比较的方法、装置设备和系统。在一些实施方式中,可以使用所公开的用于高通量分析的方法、装置和系统,例如,作为用于比较生物药物候选物和参考药物的筛选工具。如本文所使用,“高通量”是与利用诸如nmr或x射线晶体学等传统技术进行的结构测量相比的相对术语。如下文更详细描述,本文公开的基于shg和/或基于tpf的方法和系统能够以至少比常规技术快一个数量级的速率进行结构确定。
在一些实施方式中,通过使用新颖的装置设计和机制来实现高通量形式的生物分子结构、构象或构象变化,该新颖的装置设计和机制用于相对于用于输送激发光的光学系统快速、精确和可互换地定位基底(包括待分析的拴系的或固定的生物靶标),并同时确保激发光与基底表面之间保持有效的光学耦合。用于生物样品的高通量光学询问的一种优选形式是玻璃底微孔板。本文所公开的系统和方法提供了借助于以与高通量分析系统要求兼容的方式的全内反射(tir),将shg和/或tpf所需的高强度激发光耦合至基底(例如,玻璃底微孔板中的玻璃基底)的机制。
在一个方面,本公开内容提供了一种用于一个或多个生物分子(例如,蛋白质或其他生物实体)中的结构、构象或构象变化的高通量检测的方法,该方法包括(i)用非线性-活性标签或标签使用相同的标签反应或技术,来标签一个或多个靶生物分子,例如,蛋白质分子、生物药物候选物或参考药物分子;(ii)将所述一个或多个经标签的靶生物分子拴系或固定在平面基底表面的一个或多个非连续区域,其中所述基底表面进一步包括光学界面;(iii)通过改变基底相对于外部光源的位置,依次将每个非连续区域暴露于基频的激发光下;(iv)在每个非连续区域暴露于激发光时收集从该非连续区域发射的非线性光学信号(例如,shg、sfg、dfg、tpf或其任何组合);以及(v)处理所述非线性光学信号以确定所述一个或多个靶生物分子中的每一种的取向、结构、构象或构象变化。在另一方面,所述方法进一步包括(vi)继第一次暴露于激发光之后,使所述一个或多个标签生物分子中的每一种与一个或多个测试实体(例如,测试化合物、候选结合配偶体、参考药物、已知配体或其他对照)相接触;(vii)随后将每个非连续区域一次或多次重新暴露于激发光;(viii)在每个非连续区域暴露于激发光时收集来自该非连续区域的非线性光学信号;以及(ix)处理所述非线性光学信号以确定在所述一个或多个生物实体中是否由于与所述一个或多个测试实体相接触而发生取向或构象变化。在一些实施方式中,非线性-活性标签可以附接于拴系或固定在平面基底的非连续区域上的靶蛋白或其他生物实体。在一些实施方式中,非线性-活性标签可以附接于测试实体,该测试实体用于接触拴系的或固定的靶蛋白或其他生物实体。在一些实施方式中,可以用非线性-活性标记物(即,用相同的非线性-活性标记物或用不同的非线性-活性标记物)标记拴系或固定的靶蛋白或其他生物实体和测试实体。在优选实施方式中,标签附接于靶生物分子(例如,蛋白质)表面的多个独特且不同的位点,并据此确定结构、构象或构象变化,即多次进行上述步骤i)至ix)或其一些亚组,在每种情况下均标记了蛋白质或靶生物分子的不同位点,并且这些突变体中的每一个均在多次实验中进行了测量,每个实验均有不同的实验条件。例如,如果蛋白质在10个不同的位置用非线性-活性标签进行标记,则可以在50个不同的实验条件下对蛋白质进行研究,其中每个实验中的基本取向分布均不同。结果,可以确定过渡偶极矩在法向轴上的10x50=500个不同投影,并且这将能够更准确且更完整地确定蛋白质的结构和构象。此外,通过测试无论已知与蛋白质结合或未知的多个配体,可以根据每个配体结合事件生成其他角度信息,即,由于配体结合和构象变化的角度变化数据,并且该信息可以也可用于更完整地确定分子的结构和构象景观。在所述方法的一个方面,在所述一个或多个生物实体(靶蛋白、生物药物候选物、参考药物分子等)与一个或多个测试实体相接触之后仅检测一次非线性光学信号,并继而将其用于确定是否发生构象变化。在另一方面,在所述一个或多个生物实体与一个或多个测试实体相接触之后以限定的时间间隔反复地收集非线性光学信号(即,以动力学模式),并继而将其用于确定所述一个或多个生物实体中的构象变化的动力学。
在该方法的优选方面,基底的每个非连续区域包括受支承脂双层结构,并且借助于脂质双层中的拴系或嵌入,将靶蛋白或其他生物实体固定在每个非连续区域中。在该方法的另一优选方面,借助于全内反射将激发光传递到基底表面,即光学界面,并且沿与反射的激发光相同的光轴收集从基底表面非连续区域发射的非线性光学信号。
为了实现使用非线性光学检测对蛋白质结构构象或构象变化进行高通量分析,本文所述系统需要若干组件(图6中示意图示),其包括(i)至少一个适宜的激发光源,以及用于将所述至少一束激发光束递送到光学界面的光学器件;(ii)可互换基底,其包括光学界面,一个或多个生物实体在所述基底的非连续区域中拴系或固定到该光学界面;(iii)高精度平移台,用于将所述基底相对于所述至少一个激发光源进行定位;以及(iv)光学器件,用于收集由于用激发光照射所述基底的每个所述非连续区域而生成的非线性光学信号并将所述非线性信号递送到检测器;以及(v)处理器,用于分析从所述检测器接收的非线性光学信号数据并确定固定在所述基底上的所述一个或多个生物实体的结构、构象或构象变化。在一些方面,本文公开的系统和方法还包括使用(vi)可编程流体分配系统,用于向所述基底的所述非连续区域中的每一个递送测试实体;以及(vii)使用板装卸机器人,用于在与光学系统的界面处自动化定位和更换基底。在优选实施方式中,相对低-na的检测方案用于检测双光子荧光,其中光学检测器沿着垂直于样品所附接表面的轴位于待测样品(例如,激光焦点)的正上方或正下方。光学检测器应远离表面一定距离,并朝向相对较小的接收角,以使na尽可能低,同时仍能获得足以测量的信号。在一个实施方式中,如图7所示,检测器是纤维半径为0.5mm的多模塑料滤光器,其距滑动表面的距离为7.5mm。如果tpf信号源自包含在过渡到空气之前从光学界面延伸2mm的水性环境的非连续区域,则得到的检测na将为0.054。
本文公开的方法、装置和系统可被配置用于可选地对于多种药物候选物或其他测试实体(例如,参考药物、已知配体、对照等)相接触的单个生物实体(例如蛋白质、生物药物候选物,参考药物或药物靶标)的分析,或者用于与单个测试实体相接触的多个生物实体的分析,或者其任何组合。当使一个或多个生物实体与多种测试实体相接触时,可以依次进行接触步骤,即,通过将固定的生物实体在指定时间段内暴露于单个测试实体,随后是可选的冲洗步骤以去除测试实体溶液并在将固定的生物实体引入到下一测试实体之前使所述固定的生物实体再生,或者接触步骤可以并行进行,即,通过让多个非连续区域包含相同的固定生物实体,并使所述多个非连续区域中的每一个中的生物实体暴露于不同的测试实体。本文公开的方法、装置和系统可被配置用于进行针对至少1种生物实体、至少2种生物实体、至少4种生物实体、至少6种生物实体、至少8种生物实体、至少10种生物实体、至少15种生物实体或至少20种生物实体中的结构、构象或构象变化的分析。在一些方面,本文公开的方法、装置和系统可被配置用于进行针对至多20种生物实体、至多15种生物实体、至多10种生物实体、至多8种生物实体、至多6种生物实体、至多4种生物实体、至多2种生物实体或至多1种生物实体中的结构、构象或构象变化的分析。类似地,本文公开的方法、装置和系统可被配置用于在所述一个或多个生物实体暴露于至少1种测试实体、至少5种测试实体、至少10种测试实体、至少50种测试实体、至少100种测试实体、至少500种测试实体、至少1000种测试实体、至少5000种测试实体、至少10000种测试实体或至少100000种测试实体时进行对结构、构象或构象变化的分析。在一些方面,本文公开的方法、装置和系统可被配置用于在所述一个或多个生物实体暴露于至多100000种测试实体、至多10000种测试实体、至多5000种测试实体、至多1000测试实体、至多500种测试实体、至多100种测试实体、至多50种测试实体、至多10种测试实体、至多5种测试实体或至多1种测试实体时进行对结构、构象或构象变化的分析。
激光光源和激发光学系统:图8图示了本文公开的方法和系统的一个方面,其中通过反射来自包括样品界面(或光学界面)的基底表面上的入射基本激发光来生成二次谐波光(和/或在一些实施方式中为双光子荧光;图8示出了用于检测shg的光学系统)。在一些实施方式中,将基底光学耦合至棱镜,该棱镜用于以适当的角度递送激光,以在基底表面引起全内反射(图9)。在一些实施方式中,通过使用折射率匹配流体的薄膜来提供光耦合。激光器提供在样品界面处生成二次谐波光和荧光所必需的基本光。这通常将会是皮秒或飞秒激光器,其为波长可调式或不可调式,并且可从市场购得(例如,ti:蓝宝石分秒激光器或光纤激光器系统)。处于基频(w)的光射出激光器,并且例如使用适合于该光的频率和强度的半波片来选择其偏振(例如,可从mellesgriot,oriel或newportcorp.购得)。光束继而穿过被设计用于让基本光通过但阻断非线性光(例如,二次谐波光)的谐波分离器。该滤光器用于防止二次谐波光束向激光腔中的背反射——这可能对激光性能造成干扰。继而使用反射镜与透镜的组合,在光束从最终反射镜反射之前对该光束进行转向和塑形,所述最终反射镜经由棱镜引导光束在特定位置并以特定角度θ照射在基底表面上,使得其在该基底表面经历全内反射。如果需要,可以使用电流计控制镜扫描器、旋转多面镜扫描器、布拉格衍射仪、声光偏转器或本领域已知的其它手段来扫描光路中的反射镜中之一,以允许对反射镜位置的控制。包括光学界面和非线性-活性样品表面的基底可以安装在(计算机控制的)x-y平移台上,以选择基底表面上的特定位置用于二次谐波光束和/或双光子荧光的生成。在当前描述的方法和系统的一些方面,期望扫描或旋转一个反射镜以便为了全内反射而略微改变入射角,并从而在无需大幅改变照射的激发光斑的位置的情况下使得从基底表面的非连续区域发射的非线性光学信号达到最大。在一些方面,可以使用具有不同基频的两个(或更多个)激光器在非线性-活性样品所固定于的光学界面处生成和频光或差频光。在一些实施方式中,光激发可以进一步包括附加的光源(例如,激光、弧光灯、卤钨灯或高强度led),该附加的光源可选地用于激发非线性-活性标签的固有荧光或双光子荧光(或附接在固定蛋白质上的其他荧光标签)。
基底形式、光学界面和全内反射:如上所述,本公开内容的方法、装置和系统利用平面基底来将一个或多个生物实体,例如,靶蛋白拴系或固定在该基底的顶表面上,其中顶部基底表面还包括用于激发非线性光学信号的光学界面(或样品界面)。基底可以是玻璃、二氧化硅、熔融石英、塑料,或者任何对基本光束和二次谐波光束透明并且当激发光以适当角度入射时支持基底/样品界面处的全内反射的其他固体材料。在本发明的一些方面,在其内部容纳有生物实体的非连续区域被配置成一维或二维阵列,并且通过疏水涂层或薄金属层彼此隔开。在其他方面,非连续区域可以包括基底表面中的凹坑。在其他方面,非连续区域可以通过孔形成组件而彼此分离,从而使基底形成微孔板(或微板)的底部,并且每个单独的非连续区域形成微孔板中的一个孔的底部。在本公开内容的一个方面,孔形成组件将基底的顶表面分成96个单独的孔。在另一方面,孔形成组件将基底的顶表面分成384个孔。在又一方面,孔形成组件将基底的顶表面分成1536个孔。在所有这些方面中,基底无论配置成平面阵列、凹陷阵列或微孔板形式,都可以包括与测量系统或高通量系统的其他光学和机械组件相接合的一次性或消耗性装置或盒。
本文公开的方法、装置和系统进一步包括对将基底表面划分成的非连续区域或孔的数目加以规定,不论非连续区域或孔之间是如何保持分离的。在基底上具有较大数目的非连续区域或孔在提高所述方法或系统的样品分析通量方面可能是有利的。在本公开内容的一个方面,每基底的非连续区域或孔的数目为10个到1600个之间。在其他方面,非连续区域或孔的数目为至少10个、至少20个、至少50个、至少100个、至少200个、至少300个、至少400个、至少500个、至少750个、至少1000个、至少1250个、至少1500个或至少1600个。在所公开的方法和系统的其他方面,非连续区域或孔的数目为至多1600个、至多1500个、至多1000个、至多750个、至多500个、至多400个、至多300个、至多200个、至多100个、至多50个、至多20个或至多10个。在一个优选方面,非连续区域或孔的数目为96个。在另一优选方面,非连续区域或孔的数目为384个。在又一优选方面,非连续区域或孔的数目为1536个。本领域技术人员将会明白,非连续区域或孔的数目可以在以这些值中的任何一个为界的任何范围内(例如,从约12个到约1400个)。
本文公开的方法、装置和系统还包括对将基底表面划分成的非连续区域或孔的表面积加以规定,不论非连续区域或孔之间是如何保持分离的。具有较大面积的非连续区域或孔可有助于在一些情况下对关联的生物实体的方便进入和操控,而具有较小面积的非连续区域或孔在降低确定试剂量要求和提高所述方法或系统的样品分析通量方面可能有利。在本公开内容的一个方面,非连续区域或孔的表面积为1mm2与100mm2之间。在其他方面,非连续区域或孔的面积为至少1mm2、至少2.5mm2、至少5mm2、至少10mm2、至少20mm2、至少30mm2、至少40mm2、至少50mm2、至少75mm2或至少100mm2。在所公开的方法和系统的其他方面,非连续区域或孔的面积为至多100mm2、至多75mm2、至多50mm2、至多40mm2、至多30mm2、至多20mm2、至多10mm2、至多5mm2、至多2.5mm2或至多1mm2。在一个优选方面,非连续区域或孔的面积约为35mm2。在另一优选方面,非连续区域或孔的面积约为8.6mm2。本领域技术人员将会明白,非连续区域或孔的面积可以在以这些值中的任何一个为界的任何范围内(例如,从约2mm2到约95mm2)。
通过相对于激发光源重新定位基底而将基底表面的非连续区域依次暴露于激发光(用激发光对其进行照射)。入射激发光的全内反射在样品界面产生“迅衰波”,其激发非线性-活性标签并导致二次谐波光和荧光(或者在一些方面,和频光或差频光)的生成。由于迅衰波的强度并且由此所生成的非线性光学信号的强度依赖于激发光束的入射角,因此基底平面相对于激发光束的光轴的精确取向和光束到基底的有效光耦合对于实现整个非连续区域阵列上的最佳shg和/或tpf信号至关重要。在本公开内容的一些方面,通过激发光从基底表面的单一反射来实现全内反射。在其他方面,基底可被配置成波导,使得激发光在其沿着该波导传播时经历多次全内反射。在其他方面,基底可被配置成零模式波导,其中通过纳米工艺结构来创造消逝场。
诸如图8和图9中所示的光学设施中激发光束与基底之间的有效光耦合通常将会通过使用折射率匹配液诸如矿物油、矿物油与氢化三联苯的混合物、全氟化碳液、甘油、丙三醇或具有接近1.5的折射率的类似的流体来实现,其中折射率匹配液被芯吸在棱镜与基底的下表面之间。由于在基底的快速重新定位期间有可能破坏折射率匹配液的静态无气泡膜,因此本文公开的方法、装置和系统包括产生高通量系统中激发光束到基底的有效光耦合的备选方法。
图10a-图10b和图11a-图11b图示了本公开内容的高通量系统的优选方面,其中棱镜或光栅阵列与基底(封装成微孔板形式)的下表面进行集成并且用于替代固定式棱镜,从而完全消除对于折射率匹配液或弹性体层的需求。棱镜(或光栅)阵列以这样的方式与基底的上表面上的非连续区域或孔阵列对准:使得入射激发光被“入口棱镜”(“入口光栅”)以实现激发光束从样品界面全内反射的入射角引导至与入口棱镜(入口光栅)相邻但不位于其正上方的非连续区域或孔(见图12),并且使得反射的激发光束以及在照射的非连续区域生成的非线性光学信号被从所探查的非连续区域再度偏移(与之相邻但不位于其正下方)的“出口棱镜”(“出口光栅”)所收集,并且其中针对每个非连续区域的入口棱镜和出口棱镜(入口光栅和出口光栅)是阵列中不同的、非唯一元件。
总体而言,对于包含m行xn列单个特征的非连续区域阵列,对应的棱镜或光栅阵列将具有m+2行xn列或者n+2列xm行单个棱镜或光栅。在一些实施方式中,对于包含m行×n列的单个特征的非连续区域的阵列,对应的棱镜或光栅阵列将具有m+4行×n列或者n+4列×m行的单个棱镜或光栅的阵列。通常,m≠n。在一些实施方式中,m的值可以为至少2、至少4、至少6、至少8、至少12、至少14、至少16、至少18、至少20、至少25、至少30、至少35、至少40、至少45或至少50行。在一些实施方式中,m的值可以为至多50、至多45、至多40、至多35、至多30、至多25、至多20、至多18、至多16、至多14、至多12、至多10、至多8、至多6行、至多4或至多2行。类似地,在一些实施方式中,n的值可以为至少2、至少4、至少6、至少8、至少12、至少14、至少16、至少18、至少20、至少25、至少30、至少35、至少40、至少45或至少50列。在一些实施方式中,n的值可以为至多50、至多45、至多40、至多35、至多30、至多25、至多20、至多18、至多16、至多14、至多12、至多10、至多8、至多6、至多4或至多2列。对于本领域技术人员显而易见,m和n可以具有相同的值或不同的值,并且可以具有上述指定范围内的任何值,例如,m=15而n=45。
单个棱镜或光栅的几何形状和尺寸,包括棱镜或光栅阵列层的厚度,被优化以确保入射光在基底的选定非连续区域经历全内反射,并且与基底(微孔板)相对于激发光束的位置无关地以高光耦合效率收集在所述选定非连续区域处生成的非线性光学信号。棱镜或光栅阵列可以通过本领域技术人员已知的多种技术来制造,例如,在一个优选方面,它们可以由诸如环烯烃共聚物(coc)或环烯烃聚合物(cop)、丙烯酸、聚酯或类似的聚合物等平滑流动的低双折射材料注射成型。在一些方面,棱镜或光栅阵列可以制造成单独的组件,并于随后与基底的下表面进行集成。在其他方面,棱镜和光栅阵列可以基底自身一体化特征予以制造。
收集光学器件和检测器:图8进一步图示了用于检测在依次照射基底的非连续区域时生成的非线性光学信号的收集光学器件和检测器。由于表面选择性非线性光学技术是相干技术,意味着基本光束和非线性光学光束具有以明确定义的空间关系和相位关系通过空间传播的波前,因此需要的收集光学器件最少。通过棱镜(或者上述微板装置的集成式棱镜或光栅阵列)收集发射的非线性光学信号,并经由分色反射器或反射镜将其引导至检测器。可选地采用附加的光学组件,例如,透镜、光学带通滤光器、反射镜等,在光束到达检测器之前进一步对该光束进行塑形、转向和/或过滤。可以采用多种不同的光检测器,包括但不限于光电二极管、雪崩光电二极管、光电倍增管、cmos传感器或ccd器件。在一些实施方式中,收集光路可进一步包括附加光电检测器,其可选地用于检测蛋白质或非线性-活性标签(或附接在固定蛋白质上的附加荧光标签)的固有荧光或双光子荧光。
同样,双光子荧光信号的收集需要在低-na极限内具有最少数目的光学器件。例如,较小直径的滤光器在距样品合适的距离时可以充当针孔,从而收集从探针分子发出的光的一小部分,并提高技术的灵敏度。通过光纤后,可以使用滤光器选择与荧光相对应的适当带宽,同时拒绝背景光。与shg一样,可以施用多种不同的光电检测器来检测tpf信号,包括但不限于,光电二极管、雪崩光电二极管、光电倍增管、cmos传感器或ccd装置。
x-y平移台:如图6中所示,本文公开的高通量系统的实现方案理想情况下利用高精度x-y(或者在一些情况下,x-y-z)平移台来相对于激发光束重新定位基底(上述任何形式的基底)。适宜的平移台可从多个供应商处商购获得,例如,从parkerhannifin购得。精确平移台系统通常包括若干组件的组合,包括但不限于:线性致动、光学编码器、伺服和/或步进马达,以及马达控制器或驱动单元。本文所公开的系统和方法要求平台移动的高精度和可重复性,以便确保当布置进行反复的光学检测和/或液体分配步骤时对非线性光学信号的准确测量。另外,由于激发光的焦斑的大小[在直径或边长方面为20-200微米]显著小于基底的非连续区域的大小,因此在本公开内容的一些方面,可能还期望在进行重复测量时返回到给定非连续区域内的略微不同的位置,或者在单次测量的过程中跨非连续区域的一部分缓慢扫描激发光束,从而消除关于长时暴露或预先暴露的光漂白效应的潜在担忧。
因此,本文公开的方法和系统进一步包括指定平移台所能够相对于激发光束定位基底的精度。在本公开内容的一个方面,平移台的精度为约1um与约10um之间。在其他方面,平移台的精度为约10um或更低、约9um或更低、约8um或更低、约7um或更低、约6um或更低、约5um或更低、约4um或更低、约3um或更低、约2um或更低或者约1um或更低。本领域技术人员将会明白,平移台的精度可以在以这些值中的任何一个为界的任何范围内(例如,从约1.5um到约7.5um)。
流体分配系统:如图6中所示,本文所公开的高通量系统的一些实施方式还包括用于使固定在基底表面上的生物实体或靶实体与测试实体(或测试化合物)相接触的自动化可编程流体分配(或液体分配)系统,后者通常分配于包含添加或未添加少量有机溶剂成分例如二甲基亚砜(dmso)的水性缓冲液的溶液中。适宜的自动化可编程流体分配系统可从多个供应商处商购获得,例如,从beckmancoulter、perkinelmer、tecan、velocity11以及许多其他供应商购得。在本文所公开的系统和方法的优选方面,流体分配系统还包括多通道分配头,例如,4通道、8通道、16通道、96通道或384通道分配头,用于将可编程体积的液体(例如,范围从约1微升到若干微升)同时递送到基底上的多个孔或位置。
板装卸机器人:在本文公开的高通量系统的其他方面,该系统进一步包括微板装卸(或板装卸)机器人系统(图6),用于相对于光学激发和检测光学器件自动更换和定位基底(上述任何形式的基底),或者用于可选地在光学仪器与流体分配系统之间移动基底。合适的自动化可编程微板装卸机器人系统可从多个供应商处商购获得,包括beckmancoulter、perkinelemer、tecan、velocity11以及许多其他供应商。在本文公开的系统和方法的一个优选方面,自动化微板装卸机器人系统被配置用于从和向冷藏存储单元移动包含固定的生物实体和/或测试化合物的等分试样的多个微孔板。
处理器/控制器和基于约束的调度算法:在本公开内容的另一方面,所公开的高通量系统还包括处理器(或控制器,或者计算机)(图6),该处理器被配置用于运行系统软件,可以可选地将该系统软件存储在存储单元中,并且该系统软件控制所描述的各个子系统(激发和检测光学系统、x-y(或x-y-z)平移台、流体分配系统,以及板装卸机器人)以及对进行高通量shg和/或shg与trpf信号比测量和分析所涉及的不同操作步骤进行同步。除了处置数据采集过程(即,从检测器收集对应于与构象变化相关联的非线性光学信号的输出电子信号)之外,处理器或控制器通常还被配置用于储存数据、执行数据处理和显示功能(包括确定所测试的生物实体或者生物实体与测试实体的组合是否发生了基线信号、取向或构象变化),以及操作图形用户界面以供操作者进行交互控制。处理器或控制器还可以与其他处理器连网,或者连接到因特网以便与远程位置上的其他仪器和计算机通信。
处理器/控制器的典型输入参数可以包括设置参数,诸如所要分析的微孔板总数;每板的孔数;要针对基底的每个非连续区域或微孔板的孔执行激发和检测步骤的次数(例如,以便指定终点确定或动力学确定模式);应当针对每个非连续区域或孔收集动力学数据的总时间过程;要向每个非连续区域或孔递送的测试化合物溶液的顺序、定时和体积;收集和整合非线性光学信号的驻留时间;输出数据文件的一个或多个名称;以及本领域技术人员已知的多种系统设置和控制参数中的任何参数。
在本公开内容的一个优选方面,处理器或控制器进一步被配置用于通过执行基于约束的调度算法来进行系统通量优化。该算法利用上述系统设置参数来确定针对可能彼此相邻或者可能彼此不相邻的非连续区域的穿插激发/检测和液体分配步骤的最佳顺序,从而在每小时分析的生物实体和/或测试实体数目方面使系统的总通量达到最大。系统操作步骤的优化是实现高通量分析的重要方面。在所公开的方法和系统的一些方面,分析系统的平均通量范围可以是从每小时测试约10个测试实体到每小时测试约1000个测试实体。在一些方面,分析系统的平均通量可以是每小时测试至少10个测试实体、每小时测试至少25个测试实体、每小时测试至少50个测试实体、每小时测试至少75个测试实体、每小时测试至少100个测试实体、每小时测试至少200个测试实体、每小时测试至少400个测试实体、每小时测试至少600个测试实体、每小时测试至少800个测试实体或者每小时测试至少1000个测试实体。在其他方面,分析系统的平均通向可以是每小时测试至多1000个测试实体、每小时测试至多800个测试实体、每小时测试至多600个测试实体、每小时测试至多400个测试实体、每小时测试至多200个测试实体、每小时测试至多100个测试实体、每小时测试至多75个测试实体、每小时测试至多50个测试实体、每小时测试至多25个测试实体或者每小时测试至多10个测试实体。
计算机系统和网络:在各个实施方式中,本发明的方法和系统可以进一步包括安装在计算机系统上的软件程序及其用途。相应地,如上文所述,对各个子系统的计算机控制和对进行高通量构象分析所涉及的包括数据分析和显示在内的不同操作步骤的同步都在本发明的范围内。图13中所示的计算机系统500可以理解为能够从介质511和/或网络端口505读取指令的逻辑装置,所述网络端口505能够可选地连接到具有固定介质512的服务器509上。诸如图13中所示的系统可以包括cpu501、磁盘驱动器503、可选的输入设备诸如键盘515和/或鼠标516以及可选的监视器507。通过图中所示通向本地或远程位置上的服务器的通信介质可以实现数据通信。所述通信介质可以包括任何传输和/或接收数据的手段。例如,通信介质可以是网络连接、无线连接或者因特网连接。这样的连接可以提供通过万维网的通信。可以想到,与本公开内容相关的数据可以通过这样的网络或连接来传输,以供如图13中所示的一方522来接收和/或审核。
图14是图示能够与本发明的示例实施方式相组合使用的计算机系统100的第一示例架构的框图。如图14中所描绘,示例计算机系统可以包括用于处理指令的处理器102。处理器的非限制性示例包括:intelxeon.tm处理器、amdopteron.tm处理器、samsung32位riscarm1176jz(f)-sv1.0.tm处理器、armcortex-a8samsungs5pc100.tm处理器、armcortex-a8applea4.tm处理器、marvellpxa930.tm处理器,或者功能上等同的处理器。可以利用多线程执行来进行并行处理。在一些实施方式中,还可以使用多个处理器或者具有多个核的处理器,无论是在单一计算机系统中、集群中,还是分布在包括多个计算机、蜂窝电话和/或个人数字助理装置的网路上的系统之间。
如图14中所示,高速缓存104可以连接到或者并入在处理器102中,以便为处理器102最近使用或频繁使用的指令或数据提供高速存储器。处理器102通过处理器总线108连接到北桥106。北桥106通过存储器总线112连接到随机存取存储器(ram)110,并管理处理器102对ram110的访问。北桥106还通过芯片组总线116连接到南桥114。南桥114转而连接到外围总线118。外围总线例如可以是pci、pci-x、pciexpress或其他外围总线。北桥和南桥通常称为处理器芯片组并且管理处理器、ram和外围总线118上的外围组件之间的数据传输。在一些备选架构中,北桥的功能可以并入到处理器中,而不是采用单独的北桥芯片。
在一些实施方式中,系统100可以包括附接到外围总线118的加速器卡122。加速器可以包括现场可编程门阵列(fpga)或其他用于加速某种处理的硬件。例如,加速器可以用于自适应数据重组,或者用于评价扩展集处理中的代数表达式。
外部存储124中的软件和数据可以加载到ram110和/或缓存104中以供处理器使用。系统100包括用于管理系统资源的操作系统;操作系统的非限制性示例包括:linux、windows.tm、macos.tm、blackberryos.tm、ios.tm和其他功能上等同的操作系统,以及运行于操作系统之上的应用软件用于管理数据存储和根据本发明的示例实施方式的优化。
在本示例中,系统100还包括网络接口卡(nic)120和121,所述nic120和121连接到外围总线,用于提供通向诸如网络附加存储(nas)等外部存储和其他可用于分布式并行处理的计算机系统的网络接口。
图15为示出网络200的视图,该网络200具有多个计算机系统202a和202b、多个蜂窝电话和个人数据助理202c,以及网络附加存储(nas)204a和204b。在示例实施方式中,系统202a、202b和202c可以管理数据存储和优化对储存在网络附加存储(nas)204a和204b中的数据的数据访问。可以针对数据采用数学模型,并且利用跨计算机系统202a和202b以及蜂窝电话和个人数字助理系统202c的分布式并行处理来对其进行评价。计算机系统202a和202b以及蜂窝电话和个人数字助理系统202c还可以提供用于对储存在网络附加存储(nas)204a和204b中的数据进行自适应数据重组的并行处理。图15仅图示了一个示例,而各种各样的其他计算机架构和系统均可结合本发明的各个实施方式使用。例如,可以使用刀片服务器来提供并行处理。处理器刀片可以通过背板连接,以提供并行处理。存储也可以连接到背板,或者通过单独的网络接口作为网络附加存储(nas)而连接。
在一些示例实施方式中,处理器可以保持单独的存储器空间,并通过网络接口、背板或其他连接器传输数据,以供由其他处理器进行并行处理。在其他实施方式中,一些或所有处理器可以使用共享虚拟地址存储器空间。
图16是根据示例实施方式,利用共享虚拟地址存储器空间的多处理器计算机系统的框图。该系统包括可以访问共享存储器子系统304的多个处理器302a-f。系统在存储器子系统304中并入了多个可编程硬件存储器算法处理器(map)306a-f。每个map306a-f可以包括存储器308a-f以及一个或多个现场可编程门阵列(fpga)310a-f。map提供了可配置的功能单元,并且特定算法或算法部分可被提供给fpga310a-f用于密切协同相应的处理器进行处理。例如,在示例实施方式中,map可以用于评价关于数据模型的代数表达式,以及进行自适应数据重组。在该示例中,每个map可由所有处理器出于这些目的而全局访问。在一种配置下,每个map可以使用直接存储器存取(dma)来访问关联的存储器308a-f,从而允许其与相应的微处理器302a-f独立地和异步地执行任务。在该配置下,map可以直接将结果馈送到另一map以供算法的流水线操作和并行执行。
上述计算机架构和系统仅为示例,并且各种各样的其他计算机、蜂窝电话和个人数字助理架构和系统均可结合示例实施方式来使用,包括采用通用处理器、协处理器、fpga和其他可编程逻辑器件、片上系统(soc)、专用集成电路(asic)以及其他处理和逻辑元件的任何组合的系统。在一些实施方式中,整个计算机系统或其部分能够以软件或硬件来实现。可以结合示例实施方式使用任何种类的数据存储介质,包括随机存取存储器、硬盘驱动器、闪速存储器、磁带驱动器、磁盘阵列、网络附加存储(nas)以及其他本地式或分布式数据存储设备和系统。
在示例实施方式中,计算机系统可以利用执行于任何上述或其他计算机架构和系统上的软件模块来实现。在其他实施方式中,系统的功能可以部分地或全部地实现于固件、可编程逻辑器件诸如参照图16的现场可编程门阵列(fpga)、片上系统(soc)、专用集成电路(asic)或者其他处理和逻辑元件中。例如,可以通过利用硬件加速器卡,诸如图14中所示的加速器卡122,以硬件加速来实现集处理器和优化器。
实施例
提供这些实施例仅用于说明性目的,并不限制本文提供的权利要求的范围。
实施例1-二氢叶酸还原酶(dhfr)突变体的结构参数的确定
玻璃器皿清洁:在开始之前用piranha洗液洗涤所有玻璃器皿(20分钟)。小心使用——piranha洗液大量放热并易于爆炸,特别是与有机物接触时。在通风橱中,在耐热的玻璃器皿如pyrex中,通过首先测量h2o2,然后加入乙酸来制备溶液。
超声脂质制备:用氯仿(chcl3)冲洗真空瓶。确定二油酰磷脂酰胆碱dopc脂质与1,2-二油酰基-sn-甘油基-3-[(n-(5-氨基-1-羧基戊基)亚氨基二乙酸)琥珀酰基](镍盐)(dgsnta-ni)的所需摩尔比,同时尽可能地小心避免暴露于空气中。将装有脂质混合物的真空瓶放在旋转蒸发仪上。蒸发至干(约30秒),然后在蒸发的制剂上方用n2吹10min以除去残留的chcl3。在2mldih2o中重悬脂质混合物。剧烈涡旋直至形成混浊的悬浮液(约5分钟)。将悬浮液转移至4ml聚苯乙烯试管中。在冰上超声处理脂质混合物直至溶液清澈。在超声发生器设置为25%功率的情况下,这应该需要约60-90秒。
将超声处理的脂质溶液转移到微量离心管中,并在4℃下以17,000xg离心30分钟。将上清液转移到干净的微量离心管中,并将成品脂质制剂保存在4℃下,该制剂稳定长达约1个月。
载玻片准备与蛋白质加载:临应用dopc/dgsnta(ni)之前,用piranha洗液清洗显微镜载玻片30分钟。在载玻片染色容器中用dih2o洗涤8x。用压缩的氮气干燥载玻片。通过将粘性垫圈与piranha清洗的载玻片附接来组装shg孔(即,每个载玻片含有16个孔,每孔含有1-20μl体积)。使用装配夹具对齐垫圈,小心地将载玻片放入装配夹具中并牢牢按压。用pbs或tbs缓冲液1:1稀释dopc/dgsnta(ni)脂质制剂。需要100mmnacl来减少载玻片的流体静力学电荷,并使受支承脂双层(slb)能够形成。将10-20μl稀释的dopc/dgsnta(ni)脂质移液到载玻片的孔中,并在室温下温育30分钟。通过用20μl新鲜缓冲液交换缓冲液(pbs或tbs)洗涤孔10x,小心在任何时候不得将空气引入到孔中。如有必要,将孔中的缓冲液更换为合适的蛋白质加载缓冲液,并将感兴趣的靶蛋白加载到孔上。在4摄氏度下温育1至24小时,然后在开始试验之前用测定缓冲液彻底冲洗孔。
通过如上所述的超声处理制备小型单层水泡(suv),并将其施加到piranha洗涤的fisher载玻片上以制作slb表面。加入nicl210分钟,并在标签缓冲液中洗涤孔。
将经标签蛋白质以1μm(微摩尔)加载到如上所述制作的slb表面上1至24小时,然后洗涤。如果添加咪唑或edta或者蛋白质与slb表面在一种或两种蛋白质的存在下温育,则shg信号降至基线水平,表明具体地经由蛋白质的his-标记物发生了与表面的附接。
使用本领域技术人员已知的方法创建具有n-端或c-端8xhis-标记物的大肠杆菌蛋白质二氢叶酸还原酶(dhfr)的突变体(rajagopalan,p.等人(2002),“interactionofdihydrofolatereductasewithmethotrexate:ensembleandsingle-moleculekinetics”,proc.nat.acad.ofsci.(usa)99(21):13481-6;goodey,n.(2008),“allostericregulationandcatalysisemergeviaacommonroute”,natchem.biol.4(8):474-82;antikainen,n.(2005),“conformationcoupledenzymecatalysis:asingle-moleculeandtransientkineticsinvestigationofdihydrofolatereductase”,biochemistry44(51):16835-43)。在第一种情况下,制备半胱氨酸最小化的突变体,其中两个天然半胱氨酸残基都被除去(c85a和c152a)。然后将单一的不同残基(例如,m16c、n23c、q65c、k76c等)突变为半胱氨酸,即在该半胱氨酸最小化的背景下产生了单位点突变构建体。为了选择突变的位点,将蛋白质表面上的多个残基突变为半胱氨酸,并确定被shg探针标签的能力。在第二种情况下,标签野生型蛋白质,并且探针与c152的附接通过质谱分析法确认。将野生型或重组蛋白纯化到25mmtrisph7.2,150mmnacl中。按照制造商的说明,用pympo马来酰亚胺标签蛋白质,例如,蛋白质以约50μm的20:1的染料:蛋白质的标签比例在25mmtrisph7.2、150mmnacl、1mmtcep和10%甘油中温育(最终dmso浓度为5%)。将蛋白质在4℃下搅拌过夜,然后凝胶纯化到25mmtris-hclph7.2、150mmnacl、1mmtcep中。然后经由his-标记物将标签并纯化的蛋白质拴系在slb表面。
tpf和shg测量:使用本领域技术人员已知的方法,将shg信号的偏振依赖性测量用于确定针对突变体的χ(2)的独立非零分量。例如,在最简单的光学几何形状和假定方位角的各向同性及超极化率张量的单一优势分量中,确定χ(2)的两个独立非零分量(χzzz和χxzx或χzxx)。这些进而被用于最佳地确定与每个突变体的φ(即φ1和φ2)相关的取向分布。
例如,对于经由his-标记物固定化在c端的m16c突变体,使用图17a-图17b所示的光学装置在p-偏振和s-偏振激发下进行tpf和shg信号的测量。使用针孔检测设备检测tpf,该针孔检测设备由在激发光束(在样品正上方)和光电倍增管的照射下垂直于表面平面并沿轴到达样品的一部分的光纤组成。通过结合tpf和shg测量值,并在两种不同的激发偏振下取各自的强度比,可以得出两个独立的方程,该方程仅为探针角度取向分布的函数。由于具有两个使用tpf和shg测量值的独立方程,因此不必假设零宽度取向分布(即δ函数分布),而是可以将我们的数据拟合到包含平均角度φ和取向宽度σ的分布中。通过假设用于拟合数据的高斯取向分布(即,如上所述,通过对方程(2)和(6)的左侧进行积分,以在强度比的测量值已被替代时,鉴别满足方程的成对平均倾斜角和分布宽度值),由相位空间(φ,σ)的tpf和shg曲线轨迹的交点(tpf=绿色迹线;shg=蓝色迹线)确定m16cdhfr突变体的唯一平均角度和取向分布宽度(图18),其指示与测得的tpf和shg的比率均一致的一对φ和σ值。对于m16c突变体,平均取向倾斜角确定为78°,而取向分布的宽度确定为24°。
配体的添加引起shg基线信号水平的变化(shg%变化),要么是正值要么是负值,其由标签在蛋白质集合上的重新取向所导致。换句话说,shg基线信号的变化是由配体结合引起的标签取向分布的变化所导致。图19(tpf曲线=绿色迹线;shg曲线–蓝色迹线)示出了实验结果,其中1um(微摩尔)甲氧苄氨嘧啶(tmp)(已知可与dhfr结合的配体)对tpf和shg的基线信号产生了差异变化,从而导致配体结合状态的平均角度相差很大,并且取向分布宽度更宽。在存在1μmtmp的情况下(图19),平均取向倾斜角被确定为26°,而取向分布的宽度被确定为33°。
对于标记在残基m16c、n23c、r52c、q65c、t73c、k76c和e118c上的单半胱氨酸dhfr突变体,已经重复了上述确定唯一的取向平均角度和取向分布的过程。在有或没有配体tmp的情况下进行实验,并且实验的结果在表4和图20中详细描述。由于每个单半胱氨酸dhfr突变体,染料均位于蛋白质的不同区域,因此在结合tmp时观察到不同的角度响应。在极端情况下,m16c突变体展示出较大的构象重排,而在q65突变体的情况下,角度变化更适中。将对于所有单半胱氨酸突变体,在配体添加后由于蛋白质的构象重排而引起的染料取向(平均角度)和取向分布的变化总结在表4中并展示在图20中。
表4.每个单半胱氨酸突变体的取向平均角度和分布
图21展示了添加tmp后每个单半胱氨酸突变体的取向平均角度的变化和取向分布的变化。虚线的交点定义无取向变化的原点。数据指示添加tmp后,m16c突变体和n23c突变体在取向平均角度和取向分布上分别展示出最大的变化。
可以通过比较在存在和不存在tmp的情况下公布的晶体结构来定性确认使用所述方法测量的在单半胱氨酸突变位点处添加配体后的取向变化。图22是在药物抑制剂甲氨蝶呤(mtx)存在(蓝色)和不存在(棕褐色)的情况下,dhfr晶体结构的重叠,其在dhfr中产生类似于tmp的构象变化。在图中标记了已被突变为半胱氨酸的残基的位置,并示出了突变前侧链相对于肽主链的角取向。通过将mtx添加至突变蛋白如标记的m16c突变体,在图22观察到侧链取向的较大变化,这是通过所述方法预测的(参见图21)。相反,在一些位置如q65c,侧链取向在图22中展示出的角度变化较小,甚至没有变化,这也可以通过所述的方法预测(参见图21)。
实施例2-用非天然氨基酸标记的蛋白质中结构参数的确定(预示实施例)
具有shg-活性和tpf-活性的非天然氨基酸p17蛋白质是用l-anap作为标签在哺乳动物细胞中制备的(hek293),其中使用本领域技术人员已知的多种遗传工程技术中的任一种将l-anap掺入蛋白质(参见,例如,chatterjee,等人(2013),ageneticallyencodedfluorescentprobeinmammaliancells,jamchemsoc.135(34):12540–12543;lee,等人(2009),thegeneticincorporationofasmall,environmentallysensitive,fluorescentprobeintoproteinsins.cerevisiae,jamchemsoc.131(36):12921–12923)。
使用shg-活性和tpf-活性非天然氨基酸(例如,l-anap或aladan)作为标签而非外源标签来进行实施例1中所述的实验方案。然后根据本领域技术人员已知的方法将非天然氨基酸标签的p17偶联至磷脂酰丝氨酸/dopc受支承脂双层(摩尔百分比分别为25%至75%)(参见,例如,nanda,等人(2010),electrostaticinteractionsandbindingorientationofhiv-1matrixstudiedbyneutronreflectivity,biophysicalj.99(8):2516–2524)。然后,通过在p-偏振和s-偏振激发下在shg和tpf通道中使用低na检测方案(例如,不使用透镜)确定检测到的光的强度,来测量p17中掺入的非天然氨基酸的平均倾斜角和方向分布宽度(进而将其静电附接至受支承脂双层)。
实施例3-在不同实验条件下确定结构参数(预示实施例)
实施例1的实验可以扩展到包括不同浓度的加入到缓冲液中的分子(即添加剂)(例如,不同浓度的peg400(例如,10μm、20μm、40μm和80μm微克分子peg400)),该分子与界面区域关联并为拴系蛋白质产生不同的取向分布。这增加了独立分子取向分布和偏振测量值的数目,并且增加了角度测量和蛋白质结构模型的精度。
实施例4-在不同实验条件下使用靶蛋白的不同突变体确定结构参数(预示实施例)
实施例1或2的实验可以扩展到包括dhfr的10个不同突变体,每一个均在不同的单半胱氨酸位点处标签。这增加了独立偏振测量值的数目,并因此增加了角度测量和相应蛋白质结构模型的精度。尽管本文中已经示出并描述了本发明的优选实施方式,但对于本领域技术人员显而易见的是这些实施方式仅以示例的方式提供。本领域技术人员在不脱离本发明的情况下现将想到多种变化、改变和替代。应当理解本文中所述的本发明实施方式的各种替代方案均可以任何组合用于实施本发明。旨在由所附权利要求来限定本发明的范围,并由此涵盖这些权利要求范围内的方法和结构及其等同物。
- 还没有人留言评论。精彩留言会获得点赞!