用于治疗癌症或感染的组合制剂的制作方法
本发明涉及组合制剂和药物组合物以及其作为药物,特别用于治疗癌症或感染的用途,以及用于治疗癌症或感染的方法。
当原初(naive)t细胞从胸腺出现后,其通过淋巴结在血液中循环并寻找由特定抗原呈递细胞(antigen-presentingcell,apc)(通常是树突细胞)呈递的外来(“非自身”)抗原。t细胞不仅可以识别病原体相关抗原,而且可以将异常表达的自身蛋白质(指示突变或转化的致瘤细胞)识别为“非自身”。如果t细胞在适当的共刺激分子的情况下遇到其特异性抗原,则细胞被活化并上调活化和归巢分子。这些t细胞(称为效应t细胞)能够进入发炎组织以寻找感染的细胞或癌细胞。在其他功能中,效应t细胞可产生炎性细胞因子和/或溶细胞颗粒(cytolyticgranule),从而导致感染细胞或肿瘤细胞的凋亡或坏死。
在整个免疫应答期间,局部和全身的下调力(down-regulatoryforce)使对健康细胞和组织的损害最小化。这些可能涉及免疫抑制细胞因子、调节性t细胞(treg)和来自其他细胞的负信号转导。肿瘤抗原特异性t细胞显示出以促炎性细胞因子的产生减少和对抗原再刺激的低应答为特征的效应物功能受损和表型耗竭。这是由细胞外在机制(例如调节性t细胞(treg))和细胞内在机制(例如在耗竭的肿瘤浸润淋巴细胞(tumorinfiltratinglymphocyte,til)上上调的抑制性分子)介导的。
免疫检验点通路强烈地下调t细胞活化,目的是保持新生的t细胞应答在控制中,并降低对正常组织的免疫攻击的可能性。然而,在肿瘤发生期间,癌细胞可以利用这些共抑制通路来抵抗检测或避免被适应性免疫系统消除。程序性细胞死亡蛋白-1(programmedcelldeathprotein-1,pd-1)是由活化后t细胞表达的关键检验点分子。认为pd-1检验点通路主要作用于外周组织以抑制持续的免疫应答和/或防止对自身组织的损害。除t细胞外,pd-1还由b细胞、天然杀伤(naturalkiller,nk)细胞、树突细胞和活化的单核细胞表达。pd-1配体(其包括pd-l1和pd-l2等)由巨噬细胞和单核细胞表达,并且这些可在炎性环境中的许多细胞类型中被诱导。
肿瘤利用非免疫细胞表达pd-1的配体(主要是pd-l1)的能力作为避免免疫攻击的一种方法。肿瘤细胞也可下调抗原表达以避免检测。此外,免疫抑制介体的产生以及肿瘤微环境中treg和免疫抑制细胞的保留可抑制抗肿瘤免疫应答。
图1(取自harvey,clinicalpharmacology&therapeutics,2014,第96卷(2),214-223页)描绘了pd-1通路在肿瘤免疫逃避中的作用和pd-1通路阻断的作用机制:(a)t细胞活化中的pd-1。t细胞通过以下活化:(i)apc上的mhc加肽与tcr的结合,然后(ii)apccd80/86与t细胞cd28的结合。在癌症患者中,肿瘤细胞也可充当apc。在t细胞活化后,诱导pd-1表达;(b)t细胞耗竭(exhaustion)中的pd-1。在慢性感染或持续性刺激的情况下,pd-l1通过t细胞pd-1发出信号以“关闭”t细胞,从而最小化对健康组织的损害(阻断活化信号转导)。肿瘤细胞可上调pd-l1从而“关闭”可能破坏它们的t细胞。(c)阻断pd-1/pd-l1信号转导通路允许t细胞维持其效应物功能。在癌症患者中,活化的肿瘤特异性t细胞可杀伤肿瘤细胞并分泌活化/募集其他免疫细胞的细胞因子以参与抗肿瘤应答。
ishida等(theembojournal(1992),第11卷(11),第3887-3895页)描述了pd-1的克隆。人pd-1cdna的序列在genbank登录号nm_005018下记录。人pd-l1cdna的序列以genbank登录号af233516给出,人pd-l2cdna的序列以genbank登录号nm_025239给出。
2014年9月,美国食品和药物管理局(foodanddrugadministration,fda)加速批准了用于治疗患有晚期或无法切除的黑素瘤患者(其不再对其他药物有响应)的keytruda(派姆单抗(pembrolizumab))。keytruda(merck&co.)是针对pd-1的人源化单克隆igg4抗体。它包含接种到人igg4免疫球蛋白中的非常高亲和力的小鼠抗人pd-1抗体的可变区序列,具有改变以提高稳定性。keytruda阻断pd-1与pd-l1和pd-l2的结合。
2014年12月,美国fda还加速批准了opdivo(纳武单抗(nivolumab)),一种用于患有无法切除或转移性黑素瘤患者(其不再对其他药物有响应)的新治疗。opdivo(bristol-myerssquibb)是针对pd-1的完全人单克隆igg4抗体,其阻断pd-1与pd-l1和pd-l2的结合。
在pd-1通路抑制剂中,纳武单抗已经历了肺癌中最广泛的临床评估。在非小细胞肺癌(non-small-celllungcarcinoma,nsclc)患者中已经证明在鳞状和非鳞状非小细胞肺癌中作为单一疗法以及与常规化学疗法的组合二者的活性的证据。在nsclc患者中正在进行的临床试验(nct01295827)中正在评估派姆单抗。
靶向pd-1通路的许多其他有前景的药剂(pd-1通路抑制剂)正处于临床开发中(参见下表1.1):
表1.1:除派姆单抗和纳武单抗外,临床开发中的pd-1通路抑制剂(取自harvey,clinicalpharmacology&therapeutics,2014,第96卷(2),第214-223页)
adcc,抗体依赖性细胞介导的细胞毒性;igg,免疫球蛋白g;pd-1,程序性死亡-1;pd-l1,pd配体1。
临床开发中的另一种pd-1通路抑制剂是由merckkgaa和pfizer共同开发的avelumab(也称为msb0010718c),一种完全人抗pd-l1igg1单克隆抗体。
尽管最近fda批准了keytruda和opdivo用于治疗晚期黑素瘤,并且从靶向pd-1通路之药剂的临床试验中得到针对nsclc的有前景的结果,但是仍需要提供更有效的癌症治疗,从而为更广泛的癌症患者提供有效的治疗,为另一些癌症提供有效的治疗,以及提供副作用减少的有效的癌症治疗。
淋巴细胞活化基因3(lymphocyteactivationgene3,lag-3)是具有四个胞外免疫球蛋白超家族结构域的cd4同源i型膜蛋白。与cd4类似,lag-3在t细胞的表面处寡聚化并且以比cd4显著更高的亲和力与抗原呈递细胞(apc)上的mhcii类分子结合。lag-3在活化的cd4+和cd8+t淋巴细胞上表达,在那里其与在细胞表面处的cd3/t细胞受体复合体缔合并负调控信号转导。因此,它负调控t细胞的增殖、功能和内稳态。与效应t细胞或记忆t细胞相比,lag-3在耗竭的t细胞上被上调。lag-3还在肿瘤浸润淋巴细胞(til)上被上调,并且使用抗lag-3抗体阻断lag-3可增强抗肿瘤t细胞应答。
blackburn等(natimmunol.2009;10(1):29-37)描述了通过多种抑制性受体在慢性病毒感染期间cd8+t细胞耗竭的共调节。使用慢性淋巴细胞性脉络丛脑膜炎病毒(lymphocyticchoriomeningitisvirus,lcmv)的小鼠模型,作者证明与记忆或原初cd8+t细胞相比,耗竭的抗原特异性cd8+t细胞具有多至七种抑制性受体(pd-1、lag3、2b4、cd160、ctla-4、pir-b和gp49)的表达提高。多种不同抑制性受体的共表达与更大的t细胞耗竭和更严重的感染有关。阻断t细胞抑制性受体pd-1和lag-3(使用抗pd-l1和抗lag-3抗体)改善了t细胞应答并降低了体内的病毒载量。
woo等(cancerresearch2011;72(4):917-927)描述了可移植肿瘤中pd-1和lag-3在肿瘤浸润性cd4+和cd8+t细胞上的共表达。抗lag-3/抗pd-1双重抗体治疗治愈了大多数已建立肿瘤的小鼠(其大部分对单一抗体治疗有抗性)。
在慢性感染和癌症中lag-3对t细胞功能的免疫调节作用的基础上,预测的lag-3特异性单克隆抗体的作用机制是抑制肿瘤特异性效应t细胞的负调控。
lag-3还编码翻译为可溶形式的lag-3(slag-3)的替代剪接变体。作为可溶性分子,lag-3通过mhcii类信号转导来活化抗原呈递细胞(apc),从而导致体内抗原特异性t细胞应答增加(triebel,trendsimmunol.,2003,24:619-622)。
通过活化1型细胞毒性(tc1)cd8t细胞、nk细胞和单核细胞/巨噬细胞介导主要的抗肿瘤免疫应答。在短期的离体测定中,可溶形式的lag-3蛋白(imp321)在外周血单核细胞(peripheralbloodmononuclearcell,pbmc)中诱导适当的细胞毒性型应答(brignone等,journalofimmunology,2007,179:4202-4211)。imp321与pbmc中少数mhcii类+细胞(包括所有骨髓树突细胞和少部分单核细胞)结合。将imp321添加至pbmc后4小时,这些骨髓细胞产生tnf-α和ccl4。在第18小时,1%的cd8+t细胞和3.7%的nk细胞产生tc1细胞因子,例如ifn-α和/或tnf-α。这种tc1型活化需要通过imp321的早期apc活化,因为纯的经分选cd8+t细胞不能被imp321活化。只有经历抗原的完全分化的颗粒酶+cd8t细胞(效应物和效应记忆t细胞,但不是原初或中枢记忆t细胞)才能被imp321诱导为完全tc1活化。
现已发现pd-1通路抑制剂(抗pd-1抗体或抗pd-l1抗体)和作为apc活化剂的lag-3的可溶性衍生物(imp321)在体外一起协同活化t细胞(特别是cd8+t细胞)。
t细胞的这种协同活化是出乎意料的。在woo等(同上)描述的抗lag-3/抗-pd-1双重抗体治疗中,认为抗lag-3抗体是通过lag-3抑制肿瘤特异性效应t细胞的负调节,而认为lag-3的可溶性衍生物(imp321)作为apc活化剂通过不同的机制起作用。
根据本发明,提供了组合制剂,其包含:(a)能够与mhcii类分子结合的lag-3蛋白或其衍生物;和(b)pd-1通路抑制剂。
本文使用的术语“组合制剂”是指“药盒组件(kitofparts)”,在该意义上,如上定义的组合组分(a)和(b)可独立地给药或通过使用具有不同量的组合组分(a)和(b)的不同固定组合给药。所述组分可以同时施用或相继施用。如果组分相继施用,则优选地选择施用之间的时间间隔以使得组合使用所述组分的治疗效果高于仅使用组合组分(a)和(b)中任一种将获得的效果。
组合制剂的组分可以以一个组合单位剂型或者作为组分(a)的第一单位剂型和分开的组分(b)的第二单位剂型存在。在组合制剂中待施用的组合组分(a)与组合组分(b)总量的比可以变化,例如以应对待治疗的患者亚群的需求或单个患者的需求,这可能是由于,例如患者的特定疾病、年龄、性别或体重。
优选地,存在至少一种有益效果,例如增强pd-1通路抑制剂的作用,或增强lag-3蛋白或其衍生物的作用,或相互增强组合组分(a)和(b)的作用,例如与有效剂量的组合组分(a)和(b)的一个或两个相比,大于叠加效应、额外的有利效果、较小的副作用、较低的毒性或组合的治疗效果,以及非常优选地组合组分(a)和(b)的协同作用。
本发明的组合制剂可作为用于向哺乳动物(优选人)施用的药物组合制剂提供。lag-3蛋白或其衍生物可任选地与可药用载体、赋形剂或稀释剂一起提供,和/或pd-1通路抑制剂可任选地与可药用载体、赋形剂或稀释剂一起提供。
lag-3或其衍生物可以以摩尔当量为0.25至30mg、1至30mg或6至30mglag-3衍生物lag-3ig融合蛋白imp321的剂量存在。已示出每次皮下(s.c.)注射6至30mg剂量的imp321是安全的并基于在转移性肾细胞癌患者中获得的药代动力学数据的结果提供可接受的全身性暴露。在注射有剂量超过6mg的imp321的患者中在皮下(s.c)注射后至少24小时获得高于1ng/ml的imp321的血液浓度。
本发明的组合制剂可包含多剂lag-3蛋白或其衍生物。
pd-1通路抑制剂可以是抑制pd-1与pd-l1和/或pd-l2的结合的药剂。特别地,所述药剂可抑制人pd-1与人pd-l1和/或人pd-l2的结合。所述药剂可抑制pd-1与pd-l1和/或pd-l2的结合至少50%、60%、70%、80%或90%。ghiotto等(int.immunol,2010年8月;22(8):651-660)中描述了用于通过表面等离子体共振(surfaceplasmonresonance,spr)分析或流式细胞术分析来确定pd-1与pd-l1或pd-l2的结合之合适的测定。所述药剂可抑制pd-1与pd-l1和/或pd-l2的结合,例如通过使pd-1与pd-l1或pd-l2结合。所述药剂可以是抗体,合适地是单克隆抗体,例如人或人源化单克隆抗体。所述药剂可以是保留抑制pd-1与pd-l1和/或pd-l2结合之能力的抗体的片段或衍生物。
适用于根据本发明的抗pd-1抗体的实例包括:派姆单抗(mk-3475),人源化单克隆igg4抗体;纳武单抗,完全人单克隆igg4抗体;匹地利珠单抗(ct-011),人源化igg1单克隆抗体。与pd-1结合但不是抗体的pd-1通路抑制剂的实例是amp-224。amp-224是pd-l2的胞外结构域和人igg的fc区的重组融合蛋白。amp-224引起pd-1高表达t细胞的消耗。根据本发明适用的抗pd-l1抗体的实例包括:bms-936559,完全人igg4单克隆抗体;medi4736(durvalumab),完全人单克隆抗体;mpdl3280a,含有改造的iggfc结构域以防止adcc的人单克隆抗体;aveumab(也称为msb0010718c),完全人抗pd-l1igg1单克隆抗体。
pd-1通路抑制剂的剂量将取决于所使用的具体的pd-1通路抑制剂。一般来说,用于人对象的pd-1通路抑制剂的常用处方剂量可以是0.1至10mg/kg,例如0.1至1mg/kg或1至10mg/kg。术语“常用处方剂量(typicallyprescribeddose)”在本文中用于包括这样的剂量,其与作为单一疗法向对象(合适的是人对象)施用的安全和治疗有效的,或由适当的管理机构批准的用于作为单一疗法向对象(合适的是人对象)施用的剂量相同或在所述剂量范围内。当用作单一疗法时,已知pd-1通路抑制剂的常用处方人剂量的实例包括:
派姆单抗(mk-3475):每两周或三周2至10mg/kg。例如,美国fda已批准每3周在30分钟内以静脉内输注施用2mg/kgkeytruda(派姆单抗);
纳武单抗:每两周0.1至10mg/kg。例如,美国fda已批准每2周在60分钟内以静脉内输注施用3mg/kgopdivo(纳武单抗);
bms-936559:每两周0.3至10mg/kg。
pd-1通路抑制剂可通过任何合适的途径施用,例如肠胃外(包括通过皮下、静脉内或肌内注射)。目前批准的或开发中的pd-1通路抑制剂以静脉内输注施用。
本发明的组合制剂可包含多剂pd-1通路抑制剂。
lag-3蛋白可以是分离的天然或重组lag-3蛋白。lag-3蛋白可包含来自任何合适物种的lag-3蛋白质(例如灵长类动物或鼠lag-3蛋白,但优选人lag-3蛋白)的氨基酸序列。在huard等(proc.natl.acad.sci.usa,11:5744-5749,1997)的图1中提供了人和鼠lag-3蛋白的氨基酸序列。在下面的图15中重复了人lag-3蛋白的序列(seqidno:1)。在huard等的图1中还鉴定了人lag-3的四个胞外ig超家族结构域(d1、d2、d3和d4)的氨基酸序列,其在以下氨基酸残基处:1-149(d1);150-239(d2);240-330(d3);和331-412(d4)。
lag-3蛋白的衍生物包括能够与mhcii类分子结合的lag-3蛋白的可溶性片段、变体或突变体。已知lag-3蛋白的几种衍生物能够与mhcii类分子结合。在huard等(proc.natl.acad.sci.usa,11:5744-5749,1997)中描述了这些衍生物的许多实例。该文件描述了在lag-3蛋白上mhcii类结合位点的特征。描述了用于制备lag-3突变体的方法以及用于确定lag-3突变体与ii类阳性daudi细胞结合的能力的定量细胞粘附测定。测定了lag-3的几个不同突变体与mhcii类分子的结合。一些突变能够减少ii类结合,而其他突变则提高lag-3对ii类分子的亲和力。对于结合mhcii类蛋白所必需的许多残基在lag-3d1结构域中大的30个氨基酸额外环结构的碱基处簇集。人lag-3蛋白d1结构域之额外环结构的氨基酸序列为图15中加下划线的序列:gppaaapghplapgphpaapsswgprprry(seqidno:2)。
lag-3蛋白衍生物可包含人lag-3d1结构域的30个氨基酸的额外环序列或具有一个或更多个保守氨基酸替换的这样的序列的变体。所述变体可包含与人lag-3d1结构域的30个氨基酸的额外环序列具有至少70%、80%、90%或95%氨基酸同一性的氨基酸序列。
lag-3蛋白的衍生物可包含lag-3蛋白(优选人lag-3蛋白)的结构域d1和任选的结构域d2的氨基酸序列。
lag-3蛋白的衍生物可包含与lag-3蛋白(优选人lag-3蛋白)的结构域d1或结构域d1和d2具有至少70%、80%、90%或95%氨基酸同一性的氨基酸序列。
lag-3蛋白的衍生物可包含lag-3蛋白(优选人lag-3蛋白)的结构域d1、d2、d3和任选的d4的氨基酸序列。
lag-3蛋白的衍生物可包含与lag-3蛋白(优选人lag-3)的结构域d1、d2和d3或结构域d1、d2、d3和d4具有至少70%、80%、90%或95%氨基酸同一性的氨基酸序列。
氨基酸序列之间的序列同一性可通过比较序列的比对来确定。当被比较的序列中的等同位置被相同的氨基酸占据时,那么该分子在该位置是相同的。将比对评分为同一性百分比,其是在被比较序列共有位置上相同氨基酸数目的函数。当比较序列时,最佳比对可能需要将空位引入一个或更多个序列中以考虑所述序列中可能的插入和缺失。序列比较方法可采用空位罚分,以使得对于所比较序列中相同数目的相同分子具有尽可能少空位的序列比对(这反映了两个比较序列之间更高的相关性)将比具有许多空位的序列获得更高的得分。考虑到空位罚分,最大百分比同一性的计算包括产生最佳比对。
用于进行序列比较的合适的计算机程序可从商业和公共部门广泛获得。实例包括matgat(campanella等,2003,bmcbioinformatics4:29;可从http://bitincka.com/ledion/matgat获得程序)、gap(needleman&wunsch,1970,j.mol.biol.48:443-453)、fasta(altschul等,1990,j.mol.biol.215:403-410;可从http://www.ebi.ac.uk/fasta获得程序)、clustalw2.0和x2.0(larkin等,2007,bioinformatics23:2947-2948;可从http://www.ebi.ac.uk/tools/clustalw2获得程序)和emboss成对比对算法(needleman&wunsch,1970,同上;kruskal,1983,在:timewarps,stringeditsandmacromolecules:thetheoryandpracticeofsequencecomparison,sankoff&kruskal(编辑),第1-44页,addisonwesley;可从http://www.ebi.ac.uk/tools/emboss/align获得程序)。所有程序都可以使用默认参数运行。
例如,可以使用emboss成对比对算法的“needle”法进行序列比较,当考虑两个序列的整个长度时,其决定了这两个序列的最佳比对(包括空位)并提供百分比同一性得分。氨基酸序列比较(“蛋白质分子”选项)的默认参数可能是空位延伸罚分:0.5,空位开放罚分:10.0,矩阵:blosum62。
序列比较可以在参照序列的全长上进行。
lag-3蛋白衍生物可任选地通过接头氨基酸序列与免疫球蛋白fc氨基酸序列(优选人igg1fc氨基酸序列)融合。
可使用如huard等(同上)中所述的定量细胞粘附测定来确定lag-3蛋白的衍生物与mhcii类分子结合的能力。lag-3蛋白的衍生物对mhcii类分子的亲和力可以是人lag-3蛋白对ii类分子亲和力的至少20%、30%、40%、50%、60%、70%、80%、90%或100%。优选地,lag-3蛋白的衍生物对mhcii类分子的亲和力是人lag-3蛋白对ii类分子的亲和力的至少50%。
能够与mhcii类分子结合的合适的lag-3蛋白衍生物的实例包括包含以下的衍生物:
人lag-3序列的23至448位氨基酸残基;
lag-3的结构域d1和d2的氨基酸序列;
lag-3的结构域d1和d2的氨基酸序列,其中在一个或更多个以下位置处具有氨基酸替换:73位,其中arg被替换为glu;75位,其中arg被替换为ala或glu;76位,其中arg被替换为glu;30位,其中asp被替换为ala;56位,其中his被替换为ala;77位,其中tyr被替换为phe;88位,其中arg被替换为ala;103位,其中arg被替换为ala;109位,其中asp被替换为glu;115位,其中arg被替换为ala;
54至66位氨基酸残基缺失的lag-3之d1结构域的氨基酸序列;
重组可溶性人lag-3ig融合蛋白(imp321)-在转染有编码与人igg1fc融合的hlag-3的胞外结构域之质粒的中国仓鼠卵巢细胞中产生的200-kda二聚体。imp321的序列在us2011/0008331的seqidno:17中给出。
根据本发明,还提供了药物组合物,其包含(a)能够与mhcii类分子结合的lag-3蛋白或其衍生物;(b)pd-1通路抑制剂;和(c)可药用载体、赋形剂或稀释剂。
根据本发明,还提供了本发明的组合制剂或药物组合物,其用作药物。
本发明还提供了本发明的组合制剂或药物组合物,其用于预防、治疗或改善癌症。
根据本发明还提供了本发明的组合制剂或药物组合物在制造用于预防、治疗或改善癌症的药物中的用途。
根据本发明还提供了预防、治疗或改善癌症的方法,其包括向有此预防、治疗或改善需要的对象施用能够与mhcii类分子结合的lag-3蛋白或其衍生物,和pd-1通路抑制剂。
我们已经了解,本发明的组合制剂和组合物还可用于预防、治疗或改善感染,尤其是慢性或持续性感染。
在急性感染期间,活化的病原体特异性细胞毒性cd8t淋巴细胞(ctl)增殖并获得效应物功能(例如细胞因子产生和细胞毒性能力),这使得它们能够有效地清除感染。清除后,仍然存在一小部分的病原体特异性记忆t细胞,其在再次暴露于相同病原体后具有非常快速地重新活化并获得其杀伤功能的能力。然而,在慢性感染期间,这种情况不会发生,因为发现病原体特异性ctl在功能上有缺陷并且不能消除感染。这些耗竭的ctl受其受损的增殖能力、细胞因子产生和细胞毒性能力丧失所限定(参见图1(b),以及hofmeyer等,journalofbiomedicineandbiotechnology,第2011卷,文章id451694的综述)。
这种现象最初是使用小鼠慢性病毒感染的建立良好的小鼠模型(淋巴细胞性脉络丛脑膜炎病毒(lymphocyticchoriomeningitisvirus,lcmv))所限定(zajac等,thejournalofexperimentalmedicine,第188卷,第12期,第2205-2213页,1998;gallimore等,thejournalofexperimentalmedicine,第187卷,第9期,第1383-1393页,1998)。lcmv的阿姆斯特朗毒株(armstrongstrain)导致被免疫系统清除的急性感染,从而产生稳健的ctl记忆。另一方面,lcmv的克隆13毒株在小鼠中产生慢性感染,这使ctl耗竭并且不能清除感染。此外,与正常t细胞相比,耗竭的ctl具有代谢缺陷和参与趋化性、粘附和迁移之改变的基因表达(wherry等,immunity,第27卷,第4期,第670-684页,2007)。
在进行为了揭示导致耗竭的机制的研究中,将来自慢性lmcv感染的耗竭的ctl的遗传谱与对急性lcmv感染应答的功能性ctl的遗传谱进行比较(barber等,nature,第439卷,第7077期,第682-687页,2006)。发现耗竭的ctl具有pd-1的显著过表达,而功能性lcmv特异性ctl没有pd-1的明显表达。发现pd-1的表达与在耗竭的t细胞中看到的限定的功能受损相关,进而与较高的病毒载量相关。在慢性感染的小鼠中用抗pd-l1抗体阻断pd-1/pd-l1通路导致ctl应答增强,其引起病毒载量降低。通过耗竭的ctl的pd-1表达依赖于持续的抗原特异性刺激,因为慢性感染期间特异性表位呈递的丧失导致功能恢复以及降低的表位特异性ctl上的pd-1表达(blattman等,journalofvirology,第83卷,第9期,第4386-4394页,2009)。慢性病毒感染期间的持续性抗原刺激对ctl功能的丧失和pd-1表达的相关提高具有累进效应(progressiveeffect),这意味着更多耗竭的ctl(pd-1hi)与另一些(pd-1int)相比对pd-1阻断的功能性挽救不那么易感(blackburn等,proceedingsofthenationalacademyofsciencesoftheunitedstatesofamerica,第105卷,第39期,第15016-15021页,2008)。
根据本发明,还提供了用于预防、治疗或改善感染的本发明的组合制剂或药物组合物。
根据本发明,还提供了本发明的组合制剂或药物组合物在制造用于预防、治疗或改善感染的药物中的用途。
根据本发明,还提供了预防、治疗或改善感染的方法,其包括向有此预防、治疗或改善需要的对象施用能够与mhcii类分子结合的lag-3蛋白或其衍生物,和pd-1通路抑制剂。
在一些具体实施方案中,所述感染是慢性或持续性感染。术语“慢性或持续性感染”在本文中用于指在感染的对象中已诱导经典的ctl应答的病原体的感染,但感染尚未被清除,从而导致存在耗竭的pd-1表达,具有增殖能力受损、细胞因子产生和细胞毒性能力丧失的病原体特异性ctl。
可根据本发明治疗的感染的实例包括病毒、细菌、真菌或原生动物感染,尤其是慢性或持续性病毒、细菌、真菌或原生动物感染。
病毒感染可由以下引起:例如腺病毒、腺相关病毒、b病毒(i型恒河猴疱疹病毒,macacineherpesvirusi)、bk病毒、布尼亚病毒、基孔肯雅病毒、柯萨奇病毒、冠状病毒、巨细胞病毒、东方马脑炎病毒、埃博拉病毒、肠病毒、eb病毒、汉坦病毒、甲型肝炎病毒、乙型肝炎病毒、丙型肝炎病毒、丁型肝炎病毒、戊型肝炎病毒、疱疹病毒、1型单纯疱疹病毒、2型单纯疱疹病毒、人泡沫病毒、3型人疱疹病毒、5型人疱疹病毒、6型人疱疹病毒、7型人疱疹病毒、人免疫缺陷病毒、人乳头瘤病毒、人嗜β-淋巴细胞病毒、i型人t细胞白血病病毒、ii型人t细胞白血病病毒、流感病毒、jc病毒、jev、卡波西肉瘤相关疱疹病毒、拉沙热病毒(lassavirus)、淋巴细胞性脉络丛脑膜炎病毒、马尔堡病毒、麻疹病毒、腮腺炎病毒、尼帕病毒、诺如病毒、诺沃克病毒、正呼肠孤病毒(orthoreovirus)、副流感病毒、细小病毒、脊髓灰质炎病毒、狂犬病病毒、呼肠孤病毒、呼吸道合胞病毒、鼻病毒、裂谷热病毒、轮状病毒、风疹病毒、天花病毒、圣路易斯脑炎病毒、重型天花病毒、轻型天花病毒、水痘-带状疱疹病毒、西尼罗病毒、西方马脑炎病毒或黄热病病毒。
在一些具体实施方案中,病毒感染由以下引起:肝炎病毒(例如,乙型肝炎病毒、丙型肝炎病毒)、慢病毒(例如,人免疫缺陷病毒)、或疱疹病毒(例如,1型单纯疱疹病毒毒,2型单纯疱疹病毒)。
细菌感染可由以下引起:例如大肠杆菌(escherichiacoli)、艰难梭菌(clostridiumdifficile)、鼠伤寒沙门氏菌(salmonellathyphimurium)、铜绿假单胞菌(pseudomonasaeruginosa)、霍乱弧菌(vibriocholerae)、淋病奈瑟氏球菌(neisseriagonorrhoeae)、幽门螺杆菌(helicobacterpylori)、流感嗜血杆菌(hemophilusinfluenzae)、痢疾志贺菌(shigelladysenteriae)、金黄色葡萄球菌(staphylococcusaureus)、结核分枝杆菌(mycobacteriumtuberculosis)、肺炎链球菌(streptococcuspneumonia)或沙眼衣原体(chlamydiatrachomatis)。
真菌感染可由以下引起:例如假丝酵母属(candida)、曲霉属(aspergillus)、隐球菌属(cryptococcus)、球孢子菌属(coccidioides)、组织胞浆菌属(histoplasma)、肺孢子菌属(pneumocystis)或葡萄穗霉属(stachybotrys)。
原生动物感染可由以下引起:例如变形虫界(amoebozoa)、古虫界(excavata)、囊泡藻界(chromalveolata)、内阿米巴属(entamoeba)、疟原虫属(plasmodium)、贾第虫属(giardia)、锥虫属(trypanosoma)、球虫目(coccidia)、贝诺孢子虫属(besnoitia)、双腔吸虫属(dicrocoelium)或利什曼虫属(leishmania)。
根据本发明还提供了本发明的组合制剂或药物组合物,其用于预防、治疗或改善疾病、病症或状况,所述疾病、病症或状况可通过活化t细胞(特别是通过活化cd8阳性t细胞)预防、治疗或改善。
根据本发明还提供了本发明的组合制剂或药物组合物在制造用于预防、治疗或改善疾病、病症或状况的药物中的用途,所述疾病、病症或状况可通过活化t细胞(特别是通过活化cd8阳性t细胞)预防、治疗或改善。
根据本发明还提供了预防、治疗或改善疾病、病症或状况的方法,所述疾病、病症或状况可通过活化t细胞(特别是通过活化cd8阳性t细胞)预防、治疗或改善。所述方法包括向有此预防、治疗或改善需要的对象施用能够与mhcii类分子结合的lag-3蛋白或其衍生物,和pd-1通路抑制剂。
在一些实施方案中,可通过活化t细胞预防、治疗或改善的疾病、病症或状况可排除癌症。
根据本发明还提供了本发明的组合制剂或药物组合物,其用于增强t细胞介导的免疫应答,特别是cd8阳性t细胞介导的免疫应答。
本发明还提供了本发明的组合制剂或药物组合物在制造用于增强t细胞介导的免疫应答(特别是cd8阳性t细胞介导的免疫应答)的药物中的用途。
根据本发明还提供了增强t细胞介导的免疫应答(特别是cd8阳性t细胞介导的免疫应答)的方法,其包括向有此增强t细胞介导的免疫应答需要的对象施用能够与mhcii类分子结合的lag-3蛋白或其衍生物,和pd-1通路抑制剂。
在一些实施方案中,t细胞介导的免疫应答或cd8阳性t细胞介导的免疫应答的增强可排除癌症的预防、治疗或改善。
可向对象依次施用lag-3蛋白或其衍生物和pd-1通路抑制剂,即可在pd-1通路抑制剂之前、同时或之后施用lag-3蛋白或其衍生物。
lag-3蛋白或其衍生物和pd-1通路抑制剂可在间隔96小时、72小时、48小时、24小时或12小时内向对象施用。
或者,可向对象共施用lag-3蛋白或其衍生物和pd-1通路抑制剂,例如作为包含lag-3蛋白或其衍生物和pd-1通路抑制剂的组合物,或通过同时施用分开剂量的lag-3蛋白或其衍生物和pd-1通路抑制剂。
根据一些实施方案,向对象施用多剂lag-3蛋白或其衍生物和/或多剂pd-1通路抑制剂。
根据一些实施方案,在每次施用两剂或更多剂pd-1通路抑制剂之前、同时或之后施用一剂lag-3蛋白或其衍生物。
例如,可在每次施用两剂或更多剂pd-1通路抑制剂的96小时、72小时、48小时、24小时或12小时内施用一剂lag-3蛋白或其衍生物。
根据本发明在组合疗法中使用的组分之合适剂量的选择可由技术人员来确定和优化,例如,通过对患者进行观察,包括患者的整体健康和对组合疗法的响应。例如,如果确定患者没有表现出所期望的治疗效果或相反地,如果患者正在经历数量过多或引起麻烦的严重程度的不期望的或不良副作用,则可能需要优化。
应该对根据本发明在组合疗法中使用的组分的剂量进行选择以提供治疗有效量的组合中的组分。
组合疗法的“有效量”可以是导致与癌症相关的至少一种病理参数降低的量。例如,在一些实施方案中,组合疗法的有效量是与没有进行组合疗法的癌症相关参数的预期降低相比,有效实现病理参数降低至少约10%、20%、30%、40%、50%、60%、70%、80%或90%的量。例如,所述病理参数可以是肿瘤生长或肿瘤生长速率。
或者,组合疗法的“有效量”可以是导致与癌症治疗相关的临床益处提高的量。例如,在一些实施方案中,组合疗法的“有效量”是与没有进行组合疗法的预期临床益处相比,有效实现临床益处提高至少约10%、20%、30%、40%、50%、60%、70%、80%或90%的量。例如,临床益处可以是肿瘤响应速率、无进展的存活、总的存活或提高对随后治疗的敏感性。
或者,组合疗法的“有效量”可以是导致与癌症治疗相关的至少一个有益参数的变化的量。例如,在一些实施方案中,组合疗法的“有效量”是与没有组合疗法的癌症治疗相关参数的预期变化相比,有效实现参数变化至少约10%、20%、30%、40%、50%、60%、70%、80%或90%的量。例如,该参数可以是循环肿瘤抗原特异性cd8+t细胞数目的增加,或者肿瘤抗原特异性调节性t细胞数目的减少,或者活化的t细胞(特别是活化的cd8+t细胞)数目的增加,耗竭的抗原特异性cd8+t细胞数目的减少,或者循环功能性(即,非耗竭的)抗原特异性cd8+t细胞数目的增加。
在与感染治疗有关的一些实施方案中,组合疗法的“有效量”可以是导致与感染相关的至少一个病理参数降低的量。例如,在一些实施方案中,组合疗法的有效量是与没有组合疗法的感染相关参数的预期降低相比,有效实现病理参数降低至少约10%、20%、30%、40%、50%、60%、70%、80%或90%的量。例如,病理参数可以是病毒载量(例如,每ml血液的病毒颗粒的数目或病毒dna的量)、细菌载量(例如,每ml血液的细菌dna的量或者在不同琼脂平板上1-21天生长期后的细菌菌落的数目)。
测量病毒和细菌载量的合适方法是本领域普通技术人员所公知的。例如,在goldschmidt等(clinicalanddiagnosticlaboratoryimmunology,1998年7月,第513-518页)中比较了通过elisa测量病毒载量的方法。在holguin等(eurjclinmicrobiolinfectdis.1999年4月;18(4):256-9)和swenson等(j.clin.microbiol,2014年2月;52(2):517-523)中比较了使用用于检测病毒核酸的不同商业测定来测量病毒载量的方法。描述通过实时pcr测量细菌载量的论文的一个实例是nadkarni等(microbiology(2002),148,257-266)。该论文引用了bergey的manualofdeterminativebacteriology,现已被bergey的manualofsystematicbacteriology,第2版代替。honeyborne等(j.clin.microbiol,201149:3905-3911和j.clin.microbiol.2014年8月;52(8):3064-7)描述了分子细菌载量测定。fda批准的测量病毒和细菌载量的筛选测定的列表可在fda网站:www.fda.gov/biologicsbloodvaccines/bloodbloodproducts/approvedproducts/licensedproductsblas/blooddonorscreening/infectiousdisease/ucm080466.htm上找到。
或者,组合疗法的“有效量”可以是导致与感染治疗相关的临床益处增加的量。例如,在一些实施方案中,组合疗法的“有效量”是与没有组合疗法的预期临床益处相比,有效实现临床益处增加至少约10%、20%、30%、40%、50%、60%、70%、80%或90%的量。
或者,组合疗法的“有效量”可以是导致与感染治疗相关的至少一个有益参数的变化的量。例如,在一些实施方案中,组合疗法的“有效量”是与没有组合疗法的治疗相关的参数预期变化相比,有效实现参数变化至少约10%、20%、30%、40%、50%、60%、70%、80%或90%的量。例如,该参数可以是活化的t细胞(特别是活化的cd8+t细胞)数目的增加,循环功能性(即,非耗竭的)抗原特异性cd8+t细胞数目的增加,或者耗竭的抗原特异性cd8+t细胞数目的减少,或者抗原特异性调节性t细胞数目的减少。
根据本发明,与作为单一疗法的pd-1通路抑制剂或lag-3蛋白质或其衍生物的效果相比,可采用组合治疗来提高pd-1通路抑制剂或lag-3蛋白或其衍生物的治疗效果,或者降低所得组合中单个组分的剂量,同时防止或进一步降低单个组分的不期望的或有害的副作用的风险。
在一个实施方案中,lag-3蛋白或其衍生物和pd-1通路抑制剂的处方剂量各自在各化合物作为单一疗法的常用处方剂量范围内。可以规定化合物为单独剂量或组合剂量。与任一化合物作为单一疗法的效果相比,这样的组合提供了提高的效力。
在另一个实施方案中,lag-3蛋白或其衍生物和pd-1通路抑制剂的处方剂量各自低于各组分作为单一疗法的常用处方剂量,但为在组合中具有治疗效力的剂量。可以规定组分为单独剂量或组合剂量。可以选择组合中成分的剂量以提供与lag-3蛋白或其衍生物或者pd-1通路抑制剂作为单一疗法之类似水平的治疗效力,但是与各化合物作为单一疗法的处方剂量相比,其优点是较低剂量的lag-3蛋白或其衍生物和pd-1通路抑制剂降低了不利的副作用的风险。
在另一个实施方案中,pd-1通路抑制剂的处方剂量在单一疗法的常用处方剂量范围内,并且lag-3蛋白或其衍生物的处方剂量低于单一疗法的常用处方剂量。
在另一个实施方案中,pd-1通路抑制剂的处方剂量低于单一疗法的常用处方剂量,并且lag-3蛋白或其衍生物的处方剂量在以单一疗法的常用处方剂量范围内。
对于单一疗法,低于常用处方剂量的优选剂量是这样的剂量,其为常用处方剂量的至高50%或至高25%。例如,低于单一疗法常用处方剂量的剂量可以是这样的剂量,其为pd-1通路抑制剂和/或lag-3蛋白质或其衍生物的常用处方剂量的1-50%、1-25%、1-10%、2-50%、2-25%、2-10%。
在人对象中单一疗法的lag-3蛋白或其衍生物的常用处方剂量可以是摩尔当量为0.25-30mg、1-30mg或6-30mglag-3衍生物lag-3ig融合蛋白imp321的剂量。
人对象中单一疗法的pd-1通路抑制剂的常用处方剂量可以是0.1至10mg/kg,0.1至1mg/kg或1至10mg/kg。例如,人对象中单一疗法的派姆单抗的常用处方剂量可以是2至10mg/kg,例如2mg/kg,在人对象中单一疗法的纳武单抗的常用处方剂量可以是0.1至10mg/kg,例如3mg/kg,并且在人对象中单一疗法的bms-936559的常用处方剂量可以是0.3至10mg/kg。
在本发明的组合制剂或组合物的一些具体实施方案中,pd-1通路抑制剂的处方剂量低于单一疗法的常用处方剂量,例如pd-1通路抑制剂常用处方剂量的1-50%、1-25%、1-20%、1-10%、2-50%、2-25%、2-20%、2-10%、0.1-50%、0.1-25%、0.1-20%、0.1-10%、<20%、<10%、0.1-<10%、0.1-<10%、0.01-<20%或0.01-<10%。
根据本发明的pd-1通路抑制剂和lag-3蛋白或其衍生物的合适剂量的实例列于下表1.2中:
表1.2:根据本发明的组合制剂或组合物的一些实施方案的pd-1通路抑制剂和lag-3蛋白或其衍生物的剂量的实例
lag-3衍生物可以是上述的或图7所示的任何lag-3衍生物。在一些具体实施方案中,lag-3衍生物是imp321。
当以单独剂量施用时,lag-3蛋白或其衍生物和pd-1通路抑制剂可以基本上同时施用(例如,在间隔约60分钟、约50分钟、约40分钟、约30分钟、约20分钟、约10分钟、约5分钟或约1分钟内)或者在时间上间隔约1小时、约2小时、约4小时、约6小时、约10小时、约12小时、约24小时、约36小时、约72小时或约96小时或更长时间。
技术人员将能够确定并优化依次施用的合适时间过程,这取决于lag-3蛋白或其衍生物和pd-1通路抑制剂的具体组合。优选地选择所述时间过程以使得存在至少一种有益效果,例如增强lag-3蛋白或其衍生物或者pd-1通路抑制剂的效果,或者相互增强组合组分的效果,例如与非有效剂量的组合组分的一个或两个相比,大于叠加效应、额外的有利效果、较小的副作用、较低的毒性或者组合治疗效果,以及非常优选组合组分的协同作用。
应理解,最佳时间过程将取决于这样的因素,例如在施用后达到化合物的峰血浆浓度所花费的时间以及每个化合物的消除半衰期。优选地,时间差小于待施用的第一组分的半衰期。
技术人员还将能够确定用于施用的适当时机。在某些实施方案中,pd-1通路抑制剂可以在早晨施用,而lag-3蛋白或其衍生物在当天晚些时候施用至少一次。在另一些实施方案中,pd-1通路抑制剂和lag-3蛋白或其衍生物可以在基本上相同的时间施用。
在一些实施方案中,可以(例如,由医务人员)向对象施用pd-1通路抑制剂,以及可以为对象提供一定剂量的lag-3蛋白或其衍生物(例如以预填充注射器的形式)以便随后施用(例如在同一天的晚些时候或第二天)。
pd-1通路抑制剂和lag-3蛋白或其衍生物可以每天、每周、每两周、每三周、每月、每2个月、每3个月、每4个月、每5个月、每6个月、每7个月、每8个月、每9个月、每10个月、每11个月、每1年、每2年、每3年、每4年、每5年或更长时间施用。
对象可以在数周、数月或数年的时间内接受pd-1通路抑制剂和lag-3蛋白或其衍生物的剂量。例如,1周、2周、3周、1个月、2个月、3个月、4个月、5个月、6个月、7个月、8个月、9个月、10个月、11个月、1年、2年、3年、4年、5年或更长时间。
对象可以是哺乳动物对象,适当地是人对象。
根据本发明可治疗的癌症包括其中癌症的肿瘤细胞表达pd-l1和/或pd-l2的癌症(即pd-l1和/或pd-l2阳性癌症)。
已经在肺、卵巢、肾和结肠癌以及恶性黑素瘤中检测到pd-l1表达,但在正常组织(包括肺、子宫、肾、结肠或皮肤)中没有检测到pd-l1表达(benson等,blood116,2286-2294(2010);blank等,int.j.cancer119,317-327(2006);dong等,nat.med.8,793-800(2002))。肿瘤细胞的pd-l1表达与乳腺癌、胃癌、食道癌、肝细胞癌、恶性黑素瘤、卵巢癌、胰腺癌、肾细胞癌和尿道上皮癌的预后较差相关(zou&chen,nat.revimmunol.8,467-477(2008))。
还有证据表明人肿瘤可以表达pd-l2(rozali等,clin.dev.immunol.2012,656340(2012);karim等,clin.cancerres.15,6341-6347(2009))。非小细胞肺癌(nsclc)相关的成纤维细胞组成型表达pd-l1和pd-l2二者。还描述了pd-l2阳性(相对于pd-l2阴性)、食道癌、卵巢癌或肝细胞癌患者存活降低。
根据本发明可治疗的癌症还包括其中肿瘤浸润淋巴细胞(tumor-infiltratinglymphocyte,til)(尤其是cd8+til)表达pd-1的癌症或其中与循环淋巴细胞相比til表达更高水平pd-1的癌症。
在nsclc和黑素瘤患者二者中,在til上比在循环淋巴细胞上观察到更高水平的pd-1(blank等,int.j.cancer119,317-327(2006);zhang等,cell.mol.immunol.7,389-395(2010))。在接种疫苗的黑素瘤患者的外周血中,黑素瘤抗原特异性细胞毒性淋巴细胞和treg二者表达pd-1(wang等,int.immunol.21,1065-1077(2009))。肿瘤pd-l2表达与食道癌中cd8+til的存在之间也存在负相关(rozali等,clin.dev.immunol.,2012,656340(2012))。
与循环cd8+t细胞或来自健康志愿者的cd8+t细胞相比,从nsclc分离的cd8+til具有提高表达的pd-1和受损的功能应答(体外增殖和炎性细胞因子产生)。抗pd-l1抗体的添加显著提高了cd8+til在体外增殖和产生干扰素-γ的能力(zhang等,cell.mol.immunol.7,389-395(2010))。在使用来自卵巢癌患者的肿瘤衍生树突细胞和til的培养物的类似研究中,添加抗pd-l1抗体显著提高了响应肿瘤抗原之通过til的干扰素-γ产生。当这些til转移到携带卵巢肿瘤的免疫缺陷小鼠时,与对照组中的小鼠的肿瘤生长相比,观察到肿瘤生长减少(curiel等,nat.med.9,562-567(2003))。
特别地,根据本发明可治疗的癌症包括皮肤癌、肺癌(尤其是鳞状或非鳞状nsclc)、卵巢癌、肾癌、结肠癌、结肠直肠癌、乳腺癌、胃癌、食道癌、胰腺癌、膀胱癌、尿道上皮癌和肝癌。
根据本发明可治疗的癌症的其他实例包括黑素瘤(例如转移性恶性黑素瘤)、前列腺癌(例如激素难治性前列腺腺癌)、头颈癌(例如头颈部鳞状细胞癌)、宫颈癌、甲状腺癌、成胶质细胞瘤、神经胶质瘤、白血病、淋巴瘤(例如b细胞淋巴瘤)、肾上腺癌、aids相关癌症、软组织腺泡状肉瘤、星形细胞瘤、骨癌、脑和脊髓癌、转移性脑肿瘤、颈动脉体瘤、软骨肉瘤、脊索瘤、肾嫌色细胞癌(chromophoberenalcellcarcinoma)、透明细胞癌、皮肤良性纤维组织细胞瘤、促结缔组织增生性小圆细胞肿瘤、室管膜瘤、尤因瘤、骨外粘液样软骨肉瘤、骨不完全性纤维发生、骨纤维发育异常、胆囊或胆管癌、妊娠滋养细胞疾病、生殖细胞肿瘤、血液恶性肿瘤、肝细胞癌、胰岛细胞瘤、卡波西肉瘤、肾癌、脂肪瘤/良性脂肪瘤、脂肪肉瘤/恶性脂肪瘤、成神经管细胞瘤、脑膜瘤、默克尔细胞癌、多发性内分泌瘤形成、多发性骨髓瘤、脊髓发育不良综合征、成神经细胞瘤、神经内分泌肿瘤、乳头状甲状腺癌、甲状旁腺瘤、小儿癌症、外周神经鞘瘤、嗜铬细胞瘤、垂体瘤、前列腺癌、后葡萄膜黑素瘤、罕见的血液病、肾转移癌、横纹肌样瘤(rhabdoidtumor)、横纹肌肉瘤、肉瘤、软组织肉瘤、鳞状细胞癌、胃癌、滑囊肉瘤、睾丸癌、胸腺癌、胸腺瘤、甲状腺转移癌或子宫癌。
一般而言,本发明组合的组分或本发明的组合物可通过已知方法,以任何合适的制剂,通过任何合适的途径施用。在一些实施方案中,lag-3蛋白或其衍生物经肠胃外施用(包括通过皮下、静脉内或肌内注射)。在一些实施方案中,pd-1通路抑制剂为静脉内施用。在一些具体的实施方案中,lag-3蛋白或其衍生物为皮下施用,而pd-1通路抑制剂为静脉内施用。
可使用药物制剂领域技术人员已知和在相关文本和文献(例如,在remington:thescienceandpracticeofpharmacy(easton,pa.:mackpublishingco.,1995))中描述的常规方法来制备合适的药物组合物和剂型。
为了便于施用和剂量均匀,以单位剂型配制本发明的组合或组合物是特别有利的。如本文中使用的术语“单位剂型”是指适合以单位剂量(unitarydosage)用于待治疗个体的物理离散单位。换言之,组合物被配制成离散剂量单位,各自含有经计算以产生与所需药物载体相关的期望治疗效果的预定“单位剂量”量的活性剂。本发明的单位剂型的规格取决于待递送的活性剂的独特特征。还可以通过参考成分的常用剂量和施用方式来确定剂量。应当指出,在一些情况下,组合中两个或更多个单独的剂量单位提供治疗有效量的活性剂,例如,一起服用的两个片剂或胶囊剂可提供治疗有效剂量,以使得各片剂或胶囊剂中的单位剂量为治疗有效量的约50%。
用于肠胃外施用的根据本发明的制剂包括无菌水和非水溶液、混悬剂和乳剂。可注射水溶液含有水溶性形式的活性剂。非水溶剂或载剂的实例包括脂肪油,例如橄榄油和玉米油;合成的脂肪酸酯,例如油酸乙酯或甘油三酯;低分子量醇,例如丙二醇;合成的亲水性聚合物,例如聚乙二醇、脂质体等。肠胃外制剂还可含有助剂,例如增溶剂、防腐剂、润湿剂、乳化剂、分散剂和稳定剂,并且水性混悬剂可含有提高混悬剂粘度的物质,例如羧甲基纤维素钠、山梨醇和葡聚糖。可通过并入灭菌剂、通过截留细菌的过滤器过滤、辐照或加热来使可注射制剂无菌。它们也可以使用无菌可注射介质来制造。活性剂也可以是干燥(例如,冻干)形式,其可在即将通过注射施用前用合适的载剂再水化。
除先前描述的制剂之外,活性剂还可被配制成用于活性剂控制释放,优选在延长的时间段内持续释放的贮库制剂。这些持续释放剂型通常通过植入施用(例如,皮下或肌内或通过肌内注射)。
本发明的组合制剂可与施用组合之组分的说明书一起包装。说明书可记录在合适的记录介质或基质上。例如,说明书可被印刷在例如纸或塑料的基质上。说明书可作为在容器或其组件的标签中的包装插页存在(即,与包装或分包装相关)。在另一些实施方案中,说明书作为存在于合适的计算机可读存储介质(例如,cd-rom、磁盘)上的电子存储数据文件存在。组合制剂的一些或所有组分可被包装在合适的包装中以保持无菌性。
以下参考附图仅通过示例的方式来描述本发明的一些实施方案,其中:
图1描绘了pd-1通路在肿瘤免疫逃避中的作用和pd-1通路阻断的作用机制(apc,抗原呈递细胞;ifn-γ,干扰素-γ;mhc,主要组织相容性复合体;pd-1,程序性死亡-1;pd-l1,pd配体1;tcr,t细胞受体);
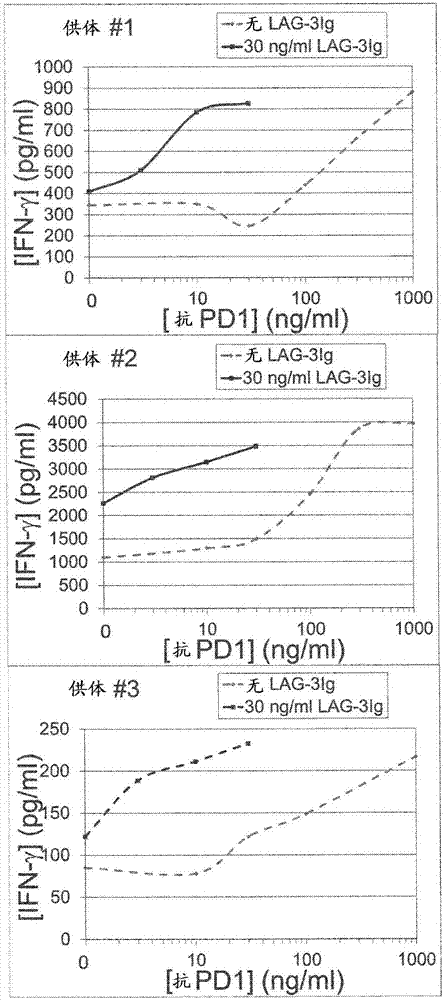
图2示出了lag-3ig和抗pd1抗体对抗原刺激诱导的ifn-γ分泌的作用;
图3示出了lag-3ig和抗pd1抗体对抗原刺激诱导的ifn-γ分泌的作用;
图4示出了lag-3ig和抗pd1抗体对抗原刺激诱导的ifn-γ分泌的作用;
图5示出了lag-3ig和抗pd1抗体对抗原刺激诱导的tnfα、il-6、rantes分泌的作用;
图6示出了lag-3ig和抗pd1抗体对抗原刺激诱导的活化标志物表达的作用。注意,图6的每个图中的最后一列的条件是“1000ng/ml抗pd1+30ng/mllag-3ig”,而不是如图所示的“30ng/ml抗pd1+1000ng/mllag-3ig”;
图7示出了与免疫球蛋白fc(lgfc)序列融合的lag-3蛋白的衍生物的示意图;
图8示出了lag-3衍生物与mhcii类阳性细胞的结合;
图9示出了通过阻断lag-3与mhcii类分子结合的抗体抑制lag-3衍生物与mhcii类阳性细胞的结合;
图10示出了如通过ccl4分泌确定的通过lag-3衍生物活化thp-1细胞;
图11示出了如通过tnf-α分泌确定的通过lag-3衍生物活化thp-1细胞;
图12示出了通过阻断lag-3与mhcii类分子结合的抗体抑制lag衍生物诱导的单核细胞活化;
图13示出了通过lag-3衍生物活化抗原呈递细胞(apc);
图14示出了通过lag-3衍生物活化cd8阳性t细胞;
图15示出了成熟人lag-3蛋白的氨基酸序列。四个胞外lg超家族结构域在以下氨基酸残基处:1-149(d1)、150-239(d2)、240-330(d3)和331-412(d4)。人lag-3蛋白的d1结构域的额外环结构的氨基酸序列以下划线粗体示出;
图16示出了lag-3ig和抗pd-l1抗体对抗原刺激诱导的活化标志物表达的作用;
图17示出了lag-3ig和不同的抗pd-1抗体(ab1和ab2)对抗原刺激诱导的ifn-γ和tnf-α分泌的作用;
图18示出了lag-3ig和不同的抗pd-l1抗体(ab3、ab4、ab5和ab6)对抗原刺激诱导的ifn-γ和tnf-α分泌的作用;以及
图19示出了不同的lag-3衍生物(imp321、imp321r75a、lag3d1d4-接头2-lg)和抗pd-1抗体对抗原刺激诱导的ifn-γ分泌的作用。
在下列实施例、表和附图中,术语“抗pd1抗体”与“抗pd-1抗体”同义使用,以及术语“抗pdl1抗体”与“抗pd-l1抗体”同义使用。
实施例1
lag-3ig和抗pd1抗体对抗原刺激诱导的ifn-γ分泌的作用
本实施例使用ifn-γ分泌测定证明了lag-3的可溶性衍生物(lag-3ig,也被称为imp321)和抗pd1抗体对体外t细胞活化的作用。外周血单核细胞(pbmc)包括淋巴细胞(t细胞、b细胞和nk细胞),单核细胞和树突细胞。ifn-γ主要由活化的cd4+和cd8+记忆和效应t细胞以及活化后的nk细胞分泌。在体外用特定抗原再刺激后,诱导ifn-γ分泌。
在存在或不存在30ng/mllag-3ig和指定浓度的抗pd1mab(克隆eh12.1,bdbiosciences,目录号562138)的情况下,将来自三个健康供体的pbmc(0.2×106个细胞/孔,在完全roswellparkmemorialinstitute(rpmi)+10%胎牛血清(fbs)中1×106m/ml)与覆盖人巨细胞病毒(cmv)pp65(来自miltenyibiotec的
通过使用bd细胞计数珠阵列(cytometricbeadarray)测量刺激后两天的细胞上清液中ifn-γ的浓度来评价t细胞应答。
在下表2中记录了各个供体之合并的一式三份存在的ifn-γ浓度。图2示出了各个供体的ifn-γ浓度对抗pd1mab浓度作图。
表2:在有和没有lag-3ig的情况下在存在抗pd1抗体时抗原诱导的ifn-γ分泌
结果表明,与单独的抗pd1抗体相比,当在存在30ng/mllag-3ig和较低浓度的抗pd1抗体的情况下孵育pbmc时,ifn-γ的分泌显著增加。例如,对于各个供体,在存在30ng/mllag-3ig和30ng/ml抗pd1抗体的情况下,高于背景水平(即,在不存在抗pd1和lag-3ig的情况下的ifn-γ的浓度)的ifn-γ浓度的增加大于在存在单独的30ng/mllag-3ig和单独的30ng/ml抗pd1抗体的情况下的对应增加之和,如下表3所示。因此,对于各个供体lag-3ig和抗pd1抗体的组合的作用是协同的。
表3:在存在30ng/ml抗pd1抗体和/或30ng/mllag-3ig的情况下抗原诱导的高于背景的ifn-γ浓度的增加
结果还表明,由lag-3ig和相对低浓度的抗pd1抗体(30ng/ml)的组合诱导的ifn-γ分泌等同于由单独的高得多浓度(300ng/ml-1000ng/ml,高10至30倍以上)的抗pd1抗体诱导的ifn-γ分泌。对于1和3号供体,与单独的1000ng/ml抗pd1抗体相比,当将pbmc与30ng/ml抗pd1和30ng/mllag-3ig一起孵育时,分泌类似浓度的ifn-γ。对于2号供体,与单独的300ng/ml和1000ng/ml抗pd1抗体相比,当将pbmc与30ng/ml抗pd1和30ng/mllag-3ig一起孵育时,分泌类似浓度的ifn-γ。
从这些结果得出结论,通过可溶性lag-3衍生物协同增加(对于1、2和3号供体分别增加约7.5、1.5和2倍)由相对低剂量的抗pd1抗体诱导的体外t细胞应答(如通过ifn-γ分泌测量)。还得出结论,如果将抗pd1抗体与可溶性lag-3衍生物组合,则使用少约10至30倍的抗pd1抗体获得同等的体外t细胞应答。
实施例2
lag-3ig和抗pd1抗体对抗原刺激诱导的ifn-γ分泌的作用
本实施例使用ifn-γ分泌测定证明了lag-3的可溶性衍生物(lag-3ig)和抗pd1抗体对体外t细胞活化的作用。
在没有任何添加剂(介质)的情况下,在有30ng/ml或1000ng/ml抗pd1mab(克隆eh12.1,bdbiosciences,目录号562138),30ng/mllag-3ig,或者30ng/mllag-3ig和30ng/ml抗pd1mab的情况下,将来自10个健康供体的pbmc(0.2×106个细胞/孔,在完全rpmi+10%fbs中1×106m/ml)与覆盖cmvpp65(来自miltenyibiotec的
通过使用bd细胞计数珠阵列测量刺激后两天的细胞上清液中ifn-γ的浓度来评价t细胞应答。
下表4中记录了各个供体各种刺激条件之合并的一式三份的ifn-γ的浓度。表5中示出了10个供体所获得的结果的平均值。图3和图4中也绘出了各个供体的结果。在图3中以黑色示出统计学差异(*p<0.05)。
表4:在有和没有lag-3ig的情况下在存在抗pd1抗体时抗原诱导的ifn-γ分泌
表5:各种不同刺激条件的平均ifn-γ浓度
结果表明,与单独的30ng/mllag-3ig或30ng/ml抗pd1抗体相比,当在存在30ng/mllag-3ig和30ng/ml抗pd1抗体的情况下孵育pbmc时,各个供体的ifn-γ分泌高得多。表5表明在存在30ng/mllag-3ig和30ng/ml抗pd1抗体的情况下,高于平均背景水平(即,在不存在抗pd1和lag-3ig的情况下的ifn-γ平均浓度)的ifn-γ平均浓度的增加大于在存在单独的30ng/mllag-3ig和单独的30ng/ml抗pd1抗体的情况下的对应增加之和(即765>239+172)。因此,lag-3ig和抗pd1抗体的组合的作用是协同的。
结果还表明,由lag-3ig和相对低浓度的抗pd1抗体(30ng/ml)的组合诱导的ifn-γ分泌等同于由单独的高得多浓度(1000ng/ml,高30倍以上)的抗pd1抗体诱导的ifn-γ分泌。
从这些结果得出结论,通过可溶性lag-3衍生物协同增加(平均增加约2倍)由相对低剂量的抗pd1抗体诱导的体外t细胞应答(如通过ifn-γ分泌测量)。还得出结论,如果将抗pd1抗体与可溶性lag-3衍生物组合,则使用少30倍以上的抗pd1抗体获得同等的体外t细胞应答。
实施例3
lag-3ig和抗pd1抗体对抗原刺激诱导的tnf-α、il-6、rantes分泌的作用
本实施例通过测量tnf-α、il-6和rantes(ccl5)的分泌证明了lag-3的可溶性衍生物(lag-3ig)和抗pd1抗体对体外t细胞活化的作用。
在没有任何添加剂(介质)的情况下,在有30ng/ml或1000ng/ml抗pd1mab(克隆eh12.1,bdbiosciences,目录号562138),30ng/mllag-3ig,或者30ng/mllag-3ig和30ng/ml抗pd1mab的情况下,将来自10个健康供体的pbmc(0.2×106个细胞/孔,在完全rpmi+10%fbs中1×106m/ml)与覆盖cmvpp65(来自miltenyibiotec的
通过使用bd细胞计数珠阵列测量刺激后两天的细胞上清液中tnf-α、il-6和rantes(ccl5)的浓度来评价t细胞应答。
下表6-8中记录了各个供体各种刺激条件之合并的一式三份的细胞因子/趋化因子的浓度。表9中示出了10个供体所获得的结果的平均值,并且表10中示出了高于平均背景的平均值的增加。图5中也绘出了各个供体的结果,并且以黑色示出统计学差异(*p<0.05)。
表6:在有和没有lag-3ig的情况下在存在抗pd1抗体时抗原诱导的tnf-α分泌
表7:在有和没有lag-3ig的情况下在存在抗pd1抗体时抗原诱导的il-6分泌
表8:在有和没有lag-3ig的情况下在存在抗pd1抗体时抗原诱导的rantes(ccl5)分泌
表9:各种不同刺激条件的平均tnf-α、il-6和rantes(ccl5)浓度
表10:各种不同的刺激条件的高于平均背景的平均tnf-α、il-6和rantes(ccl5)浓度的增加
结果表明,与单独的30ng/mllag-3ig或30ng/ml抗pd1抗体相比,当在存在30ng/mllag-3ig和30ng/ml抗pd1抗体的情况下孵育pbmc时,各个供体的il-6分泌高得多。表10表明在存在30ng/mllag-3ig和30ng/ml抗pd1抗体的情况下,高于平均背景水平(即,在不存在抗pd1和lag-3ig的情况下的il-6的平均浓度)的il-6平均浓度的增加大于在存在单独的30ng/mllag-3ig和单独的30ng/ml抗pd1抗体的情况下的对应增加之和(即8594>732+2964)。因此,lag-3ig和抗pd1抗体的组合的作用是协同的。
结果还表明,由lag-3ig和相对低浓度的抗pd1抗体(30ng/ml)的组合诱导的il-6分泌等同于由单独的高得多浓度(1000ng/ml,高30倍以上)的抗pd1抗体诱导的il-6分泌。
从这些结果得出结论,通过可溶性lag-3衍生物协同增加(平均增加2.3倍以上)由相对低剂量的抗pd1抗体诱导的体外t细胞应答(如通过il-6分泌测量)。
实施例4
lag-3ig和抗pd1抗体对抗原刺激诱导的活化标志物表达的作用
本实施例证明了lag-3的可溶性衍生物(lag-3ig)和抗pd1抗体对t细胞活化标志物表达的作用。
在没有任何添加剂(介质)的情况下,在有30ng/ml或1000ng/ml抗pd1mab(克隆eh12.1,bdbiosciences,目录号562138),30ng/mllag-3ig,或者30ng/mllag-3ig和30或1000ng/ml抗pd1mab的情况下,将来自7个健康供体的pbmc(0.2×106个细胞/孔,在完全rpmi+10%fbs中1×106m/ml)与覆盖cmvpp65(来自miltenyibiotec的
通过流式细胞术对细胞刺激后两天的三种活化标志物(lag-3、cd69和cd25)的表达进行表型分析来评价t细胞应答。
下表11-15中记录了对于各种刺激条件之合并的一式三份的表达lag-3、cd69或cd25,三种活化标志物(lag-3、cd69或cd25)中的至少一种或所有三种活化标志物(lag-3、cd69和cd25)的cd8细胞的百分比。表16中示出了7个供体所获得的结果的平均值,表17中示出了高于平均背景的平均值的增加。图6中也绘出了各个供体的结果,并且以黑色示出统计学差异(*p<0.05)。
表11:各种不同的刺激条件表达lag-3的cd8细胞的百分比
表12:各种不同的刺激条件表达cd69的cd8细胞的百分比
表13:各种不同的刺激条件表达cd25的cd8细胞的百分比
表14:各种不同的刺激条件表达三种活化标志物(lag-3、cd69或cd25)中任一种的cd8细胞的百分比
表15:各种不同的刺激条件表达所有三种活化标志物(lag-3、cd69和cd25)的cd8细胞的百分比
表16:各种不同的刺激条件表达lag-3、cd69、cd25,三种活化标志物(lag-3、cd69或cd25)中任一种或所有三种活化标志物(lag-3、cd69和cd25)的cd8细胞的平均百分比
表17:各种不同的刺激条件高于平均背景的表达lag-3、cd69、cd25,三种活化标志物(lag-3、cd69或cd25)中任一种或所有三种活化标志物(lag-3、cd69或cd25)的cd8细胞的平均百分比增加
结果表明,用30ng/ml抗pd-1抗体和30ng/mllag-3ig或者1000ng/ml抗pd-1抗体和30ng/mllag-3ig刺激导致表达任何或所有三种活化标志物的cd8细胞的平均百分比协同增加。
结果还表明,用30ng/ml抗pd-1抗体和30ng/mllag-3ig刺激比用1000ng/ml抗pd-1抗体刺激导致表达任何或所有三种活化标志物的cd8细胞的平均百分比显著更高。
从这些结果得出结论,通过可溶性lag-3衍生物协同增加由相对低剂量的抗pd1抗体诱导的体外cd8+t细胞应答(如通过t细胞活化标志物的表达测量)。还得出结论,如果将抗pd1抗体与可溶性lag-3衍生物组合,则使用少30倍以上的抗pd1抗体获得体外cd8+t细胞应答的显著改善。
由于已知pd-1通路抑制剂(例如keytruda和opdivo)活化cd8+t细胞,并且这种活化与抗癌效果有关,因此上述实施例中呈现的结果提供了如下证据:可通过共同施用pd-1通路抑制剂和能够与mhcii类分子结合的lag-3蛋白或其衍生物获得改善的抗癌效果。或者,与作为单一疗法的pd-1通路抑制剂的施用相比,可通过在较低剂量(例如,低10至30倍的剂量)的pd-1通路抑制剂下共同施用pd-1通路抑制剂和lag-3蛋白(或其能够与mhcii类分子结合的衍生物)来实现类似的抗癌效果。预期这样的共同施用将减少由pd-1通路抑制剂引起的副作用。
类似地,由于还已知cd8+t细胞活化对感染(包括慢性或持续性感染)有效,因此上述实施例中呈现的结果也提供了如下证据:共同施用pd-1通路抑制剂和能够与mhcii类分子结合的lag-3蛋白或其衍生物可用于更有效地预防、治疗或改善感染。或者,与作为单一疗法的pd-1通路抑制剂的施用相比,可通过在较低剂量(例如,低30至100倍的剂量)的pd-1通路抑制剂下共同施用pd-1通路抑制剂与lag-3蛋白(或其能够与mhcii类分子结合的衍生物)来实现类似的针对感染的效果。预期这样的共同施用将减少由pd-1通路抑制剂引起的副作用。
实施例5
lag-3衍生物与mhcii型阳性细胞的结合
测试了lag-3的几种衍生物与mhcii类阳性细胞结合的能力:
i)通过第一接头与免疫球蛋白fc(igfc)序列连接的lag-3的结构域d1-d4(lag-3d1d4-接头1-ig,slag-3d1d4-ig、lag-3ig或imp321);
ii)通过第二接头与igfc序列连接的lag-3的结构域d1-d4(lag-3d1d4-接头2-ig或slag-3d1d4-接头b-ig);
iii)通过第二接头与igfc序列连接的lag-3的结构域d1和d2(lag-3d1d2-接头2-ig或slag-3d1d2接头b-ig);和
iv)通过第一接头与igfc序列连接的lag-3的结构域d1-d4,但在lag-3d1结构域的mhcii类结合位点中于r75位具有突变(r75a),这使与mhcii类分子的结合增强了三倍或更多(huard等,proc.natl.acad.sci.usa,1997,94:5744)(imp321r75a)。
图7中示出了所述衍生物。
用不同浓度的lag-3衍生物或用人igg1抗体(higg1)作为阴性对照将mhcii类+raji细胞在4℃下孵育45分钟。用fitc-缀合的山羊抗小鼠ig(coulter)显示与细胞表面结合的lag-3分子。通过流式细胞术分析细胞。图8中示出了表示为荧光强度单位的结果。结果表明,所有的lag-3衍生物均与mhcii类阳性细胞结合。
实施例6
通过阻断lag-3与mhcii类分子结合的抗体抑制lag-3衍生物imp321与mhcii类阳性细胞的结合
17b4和11e3是已知阻断lag-3与mhcii类分子结合的抗lag-3单克隆抗体。在imp321缀合物(lag-3ig-alexa488)(4μg/ml在4℃下)与17b4或11e3阻断抗体或与同种型匹配的阴性对照单克隆抗体(migg1)预孵育之后,测定该缀合物与mhcii类-阳性b细胞(raji细胞)的结合。使用荧光活化细胞分选(facs)进行细胞结合荧光的分析。图9中示出了结果。
结果表明,通过阻断lag-3与mhcii类分子结合的lag-3特异性单克隆抗体抑制了imp321与raji细胞的结合。
实施例7
通过lag-3衍生物活化单核细胞
将thp-1细胞与图5中所示的lag-3衍生物或与作为阴性对照的人igg1在4℃下孵育4小时。测定了thp-1细胞分泌的趋化因子ccl4和细胞因子肿瘤坏死因子α(tnf-α)的量,并将其用作单核细胞活化的量度。使用细胞计数珠阵列在细胞上清液中定量ccl4和tnf-α分泌。图10中示出了ccl4测定的结果,图11中示出了tnf-α测定的结果。
结果表明,lag-3衍生物均能够活化thp-1单核细胞。
实施例8
通过阻断lag-3与mhcii类分子结合的抗体抑制imp321诱导的单核细胞活化
将imp321(20ng/ml)与17b4或11e3抗体预孵育(在37℃下5分钟),之后将该混合物与thp-1细胞在37℃下孵育4小时。使用thp-1细胞分泌的ccl4的量来确定单核细胞活化的水平。图12中示出了两次实验的结果。
结果证明,imp321诱导的单核细胞活化被阻断性抗lag-3mab17b4和11e3所抑制。这表明imp321活化单核细胞的能力依赖于imp321与mhcii类分子的结合。
实施例9
通过lag-3衍生物活化原代抗原呈递细胞(apc)
在存在布雷菲德菌素(brefeldin)(一种分泌抑制剂)的情况下,将人外周血单核细胞(pbmc)与图7中所示的lag-3衍生物或与作为阴性对照的人igg1在37℃下孵育4小时。通过ccl4(一种已知有利于th1和cd8阳性应答的趋化因子)和tnf-α(一种直接抑制肿瘤发生的多功能细胞因子)的胞内染色来确定存在于pbmc中的apc的细胞因子应答。通过细胞计数法分析结果。由在mhcii类阳性细胞中表达ccl4和/或tnf-α的细胞的百分比所表示的结果示于图13中。
结果表明,所有经测试的lag-3衍生物均在原代apc中诱导了ccl4和tnf-α的产生。
实施例10
通过lag-3衍生物活化cd8+t细胞
将人pbmc与图7中所示的lag-3衍生物,或与作为阴性对照的人igg1孵育18小时。在孵育的最后16小时存在布雷菲德菌素。在暴露于lag-3衍生物18小时之后,通过ccl4、ifn-γ和tnf-α的胞内染色追踪cd8+t细胞的细胞因子应答并通过细胞计数法分析。由在cd3+/cd8+t细胞中表达ccl4、ifn-γ和/或tnf-α的细胞的百分比所表示的结果示于图14中。
结果表明,所有经测试的lag-3衍生物均诱导1型细胞毒性cd8阳性t细胞(tc1细胞)的活化。得出结论,通过与apc表达的mhcii类分子结合,lag-3衍生物诱导了tc1细胞的活化。tc1细胞的活化形成主要的抗肿瘤免疫应答。
实施例11
lag-3ig和抗pd-l1对抗原刺激诱导的活化标志物表达的作用
本实施例证明了lag-3的可溶性衍生物(lag-3ig)和抗pd-l1抗体对t细胞活化标志物表达的作用。
在没有任何添加剂(介质)的情况下,在有30ng/ml或3000ng/ml抗pd-l1人源化抗体(bpsbioscience,目录号71213),30ng/mllag-3ig,或者30ng/mllag-3ig和30ng/ml抗pd-l1抗体的情况下,将来自12个健康供体的pbmc(0.2×106个细胞/孔,在完全rpmi+10%fbs中1m/ml)与覆盖cmvpp35序列的肽库一起一式三份孵育。
通过流式细胞术对细胞刺激后三天的三种活化标志物(lag-3、cd69和cd25)的表达进行表型分析来评价t细胞应答。
下表18-22中记录了各种刺激条件之合并的一式三份的表达lag-3、cd69或cd25,三种活化标志物(lag-3、cd69或cd25)中的至少一种或所有三种活化标志物(lag-3、cd69和cd25)的cd8细胞的百分比。表23中示出了12个供体所获得的结果的平均值,表24中示出了高于平均背景的平均值的增加。图16中也绘出了各个供体的结果,并且以黑色示出统计学差异(*p<0.05)。
表18:各种不同的刺激条件表达lag-3的cd8细胞的百分比
表19:各种不同的刺激条件表达cd69的cd8细胞的百分比
表20:各种不同的刺激条件表达cd25的cd8细胞的百分比
表21:各种不同的刺激条件表达三种活化标志物(lag-3、cd69或cd25)中任一种的cd8细胞的百分比
表22:各种不同的刺激条件表达所有三种活化标志物(lag-3、cd69和cd25)的cd8细胞的百分比
表23:各种不同的刺激条件表达lag-3、cd69、cd25,三种活化标志物(lag-3、cd69或cd25)中任一种或所有三种活化标志物(lag-3、cd69和cd25)的cd8细胞的平均百分比
表24:各种不同的刺激条件高于平均背景的表达lag-3、cd69、cd25,三种活化标志物(lag-3、cd69或cd25)中任一种或所有三种活化标志物(lag-3、cd69和cd25)的cd8细胞的平均百分比的增加
结果表明,用30ng/ml抗pd-l1抗体和30ng/mllag-3ig刺激导致表达任何或所有三种活化标志物的cd8细胞的平均百分比协同增加。
结果还表明,用30ng/ml抗pd-l1抗体和30ng/mllag-3ig刺激比用单独的3000ng/ml抗pd-1抗体刺激导致表达任何或所有三种活化标志物的cd8细胞的平均百分比显著更高。
从这些结果得出结论,通过可溶性lag-3衍生物协同增加由相对低剂量的抗pd-l1抗体诱导的体外cd8+t细胞应答(如通过t细胞活化标志物的表达测量)。还得出结论,如果将抗pd-l1抗体与可溶性lag-3衍生物组合,则使用少100倍的抗pd-l1抗体获得体外cd8+t细胞应答的显著改善。
由于已知pd-1通路抑制剂(例如keytruda和opdivo)活化cd8+t细胞,并且这种活化与抗癌效果有关,因此上述实施例中呈现的结果提供了如下证据:可通过共同施用pd-1通路抑制剂和能够与mhcii类分子结合的lag-3蛋白或其衍生物获得改善的抗癌效果。或者,与作为单一疗法的pd-1通路抑制剂的施用相比,可通过在较低剂量(例如,低30至100倍的剂量)的pd-1通路抑制剂下共同施用pd-1通路抑制剂和lag-3蛋白(或其能够与mhcii类分子结合的衍生物)来实现类似的抗癌效果。预期这样的共同施用将减少由pd-1通路抑制剂引起的副作用。
类似地,由于还已知cd8+t细胞活化对感染(包括慢性或持续性感染)有效,因此上述实施例中呈现的结果也提供了如下证据:共同施用pd-1通路抑制剂和能够与mhcii类分子结合的lag-3蛋白或其衍生物可用于更有效地预防、治疗或改善感染。或者,与作为单一疗法的pd-1通路抑制剂的施用相比,可通过在较低剂量(例如,低30至100倍的剂量)的pd-1通路抑制剂下共同施用pd-1通路抑制剂和lag-3蛋白(或其能够与mhcii类分子结合的衍生物)来实现类似的针对感染的效果。预期这样的共同施用将减少由pd-1通路抑制剂引起的副作用。
实施例12
lag-3ig和多种抗pd-1或抗pd-l1抗体对抗原刺激诱导的ifn-γ和tnf-α产生的作用
本实施例使用ifn-γ和tnf-α分泌测定证明了lag-3的可溶性衍生物(lag-3ig)和多种不同的抗pd-1或抗pd-l1抗体对体外t细胞活化的作用。
在没有任何添加剂(介质)的情况下,在有30ng/ml或1000ng/ml抗pd-1抗体(ab1或ab2)或抗pd-l1抗体(ab3、ab4、ab5或ab6),10或30ng/mllag-3ig,或者10或30ng/mllag-3ig和30ng/ml抗pd-1抗体或抗pd-l1抗体的情况下,将来自健康供体的pbmc(0.2×106个细胞/孔,在完全rpmi+10%fbs中1m/ml)与覆盖cmvpp35序列的肽库一起一式三份孵育。
通过使用bd细胞计数珠阵列测量刺激后3天的细胞培养上清液中ifn-γ和tnf的浓度来评价t细胞应答。
抗pd-1:ab1(来自bdpharmingen的克隆mih4,目录号557823)和ab2(来自bpsbioscience的人源化抗pd-1,目录号71120);
抗pd-l1:ab3(来自ebioscience的克隆mih1,目录号16-5983-82),ab4(来自ebioscience的克隆mih5,目录号16-5982-81),ab5(来自ebioscience的克隆1-111a,目录号14-9971-81)和ab6(来自bpsbioscience的人源化抗pd-l1,目录号71213)。
表25中记录了抗pd-1抗体各种刺激条件之合并的一式三份的ifn-γ和tnf-α的浓度。图17中绘出了结果。
表25:在有和没有lag-3ig的情况下在存在抗pd-1抗体时抗原诱导的ifn-γ和tnf-α分泌
结果表明,对于各抗pd-1抗体,与单独的抗pd-1抗体相比,当在存在lag-3ig和较低浓度的抗pd-1抗体的情况下孵育pbmc时,ifn-γ的分泌增加。例如,在存在30ng/mllag-3ig和30ng/mlab1抗pd-1抗体或者10ng/mllag-3ig和30ng/mlab2抗pd-1抗体的情况下,高于背景水平(即,在不存在抗pd-1和lag-3ig的情况下的ifn-γ的浓度)的ifn-γ浓度的增加大于在存在单独的lag-3ig和单独的30ng/ml抗pd-1抗体的情况下的对应增加之和(即对于ab1,235.1>-53.3+144.6;对于ab2,273.8>176.1+18.2)。因此,lag-3ig和各不同抗pd-1抗体的组合的作用是协同的。
结果还表明,由lag-3ig和相对低浓度的抗pd-1抗体(30ng/ml)的组合诱导的ifn-γ分泌比由单独的高得多浓度(1000ng/ml,高30倍以上)的抗pd-1抗体诱导的ifn-γ分泌高得多(即对于ab1,235.1>107.7;对于ab2,273.8>27.3)。
关于tnf-α分泌,单独的抗pd-1抗体(以相对低或高的浓度)均不对tnf-α分泌具有显著作用。然而,对于各抗pd-1抗体,与单独的抗pd-1抗体相比,当在存在lag-3ig和较低浓度的抗pd-1抗体的情况下孵育pbmc时,tnf-α的分泌增加。例如,在存在30ng/mllag-3ig和30ng/mlab1抗pd-1抗体或者10ng/mllag-3ig和30ng/mlab2抗体的情况下,高于背景水平(即,在不存在抗pd-1和lag-3ig的情况下的tnf-α浓度)的tnf-α浓度的增加大于在存在单独的lag-3ig和单独的30ng/ml抗pd-1抗体的情况下的对应增加之和(即对于ab1,118.7>0.9+82.2;对于ab2,12.778>2.563+9.858)。因此,lag-3ig和各不同抗pd-1抗体的组合的作用是协同的。
结果还表明,由lag-3ig和相对低浓度的抗pd-1抗体(30ng/ml)的组合诱导的tnf-α分泌显著高于由单独的高得多浓度(1000ng/ml,高30倍以上)的抗pd-1抗体诱导的tnf-α分泌(即对于ab1,118.7>1.6;对于ab2,12.778>2.494)。
从这些结果得出结论,通过可溶性lag-3衍生物协同增加由相对低剂量的抗pd-1抗体诱导的体外t细胞应答(如通过ifn-γ和tnf-α分泌测量)。还得出结论,如果将抗pd-1抗体与可溶性lag-3衍生物组合,则使用少30倍以上的抗pd-1抗体获得显著更大的体外t细胞应答。用不同的抗pd-1抗体观察到这些作用。
表26中记录了抗pd-l1抗体各种刺激条件之合并的一式三份的ifn-γ和tnf-α的浓度。图18中绘出了结果。
表26:在有和没有lag-3ig的情况下在存在抗pd-l1抗体时抗原诱导的ifn-γ和tnf-α分泌
结果表明,对于各抗pd-l1抗体,与单独的抗pd-l1抗体相比,当在存在lag-3ig和较低浓度的抗pd-l1抗体的情况下孵育pbmc时,ifn-γ的分泌增加。例如,在存在10或30ng/mllag-3ig和30ng/ml抗pd-l1抗体的情况下,高于背景水平(即,在不存在抗pd-l1和lag-3ig的情况下的ifn-γ的浓度)的ifn-γ浓度的增加大于在存在单独的10或30ng/mllag-3ig和单独的30ng/ml抗pd-l1抗体的情况下的对应增加之和(即对于ab3,226.5>28.5+79.1;对于ab4,126.03>2.31+8.00;对于ab5,180.34>19.84+30.11;对于ab6,95.14>-16.51+19.84)。因此,lag-3ig和各不同抗pd-l1抗体的组合的作用是协同的。
结果还表明,由lag-3ig和相对低浓度的抗pd-l1抗体(30ng/ml)的组合诱导的ifn-γ分泌显著高于由单独的高得多浓度(1000ng/ml,高30倍以上)的抗pd-l1抗体诱导的ifn-γ分泌(即对于ab3,226.5>55.5;对于ab4,126.03>-10.66;对于ab5,180.34>10.89;对于ab6,95.14>-49.61)。
关于tnf-α分泌,对于抗pd-l1抗体ab3、ab4和ab5,与单独的抗pd-l1抗体相比,当在存在lag-3ig和较低浓度的抗pd-l1抗体的情况下孵育pbmc时,tnf-α的分泌增加。例如,在存在10或30ng/mllag-3ig和30ng/mlab3、ab4或ab5抗pd-l1抗体的情况下,高于背景水平(即,在不存在抗pd-l1和lag-3ig的情况下的tnf-α的浓度)的tnf-α浓度的增加大于在存在单独的lag-3ig和单独的30ng/mlab3、ab4或ab5抗pd-l1抗体的情况下的对应增加之和(即对于ab3,9.0>-1.8+2.4;对于ab4,80.34>2.08+58.71;对于ab5,137.84>5.53+84.21)。因此,lag-3ig和这些不同的抗pd-1抗体的组合的作用是协同的。
尽管对于与lag-3ig组合的抗pd-l1抗体ab6未观察到对tnf-α分泌的协同作用,然而这可能是由于在存在单独的这种抗体的情况下高水平的tnf-α分泌。然而,在存在ab6和lag-3ig的组合的情况下,tnf-a的分泌水平高于在存在单独的ab6抗体(30ng/ml和1000ng/ml)的情况下tnf-α的分泌水平。
结果还表明,由lag-3ig和相对低浓度的抗pd-l1抗体(30ng/ml)的组合诱导的tnf-α分泌显著高于由单独的高得多浓度(1000ng/ml,高30倍以上)的抗pd-l1抗体诱导的tnf-α分泌(即对于ab3,9.0>0.6;对于ab4,80.34>2.50;对于ab5,137.84>4.00;对于ab6,100.99>47.81)。
从这些结果得出结论,通过可溶性lag-3衍生物协同增加由相对低剂量的抗pd-l1抗体诱导的体外t细胞应答(如通过ifn-γ和tnf-α分泌测量)。还得出结论,如果将抗pd-l1抗体与可溶性lag-3衍生物组合,则使用少30倍以上的抗pd-l1抗体获得显著更大的体外t细胞应答。用不同的抗pd-l1抗体观察到这些作用。
实施例13
lag-3衍生物和抗pd-1抗体对抗原刺激诱导的ifn-γ产生的作用
本实施例使用ifn-γ分泌测定证明了lag-3的多种不同的可溶性衍生物(实施例5中描述和图7中示出的衍生物(i)、(ii)和(iv))和抗pd-1对体外t细胞活化的作用。
在没有任何添加剂(介质)的情况下,在有30ng/ml或1000ng/ml抗pd-1抗体(eh12克隆),30ng/mllag-3衍生物(imp321、imp321r75a或lag3d1d4-接头2-ig)或者30ng/mllag-3衍生物和30ng/ml抗pd-1的情况下,将来自健康供体的pbmc(0.2×106个细胞/孔,在完全rpmi+10%fbs中1m/ml)与覆盖cmvpp35序列的肽库一起一式三份孵育。
通过使用bd细胞计数珠阵列测量刺激后3天的细胞培养上清液中ifn-γ的浓度来评价t细胞应答。
表27中记录了各种刺激条件之合并的一式三份的ifn-γ的浓度。图19中绘出了结果。
表27:在有和没有lag-3的不同衍生物的情况下在存在抗pd-1抗体时抗原诱导的ifn-γ分泌
结果表明,对于各lag-3衍生物,与单独的30ng/mllag-3衍生物或30ng/ml抗pd-1抗体相比,当在存在30ng/mllag-3衍生物和30ng/ml抗pd-1抗体的情况下孵育pbmc时,ifn-γ的分泌增加。例如,在存在30ng/mllag-3衍生物和30ng/ml抗pd-1抗体的情况下,高于背景水平(即,在不存在抗pd-1和lag-3衍生物的情况下的ifn-γ的浓度)的ifn-γ浓度的增加大于在存在单独的30ng/mllag-3衍生物和单独的30ng/ml抗pd-1抗体的情况下的对应增加之和(即对于imp321,572.3>249.7+22.3;对于imp321r75a,511.2>317.3+22.3;对于lag3d1d4-接头2-ig,520.7>258.0+22.3)。因此,抗pd-1抗体和各不同的lag-3衍生物的组合的作用是协同的。
结果还表明,由各lag-3衍生物和相对低浓度的抗pd-1抗体(30ng/ml)的组合诱导的ifn-γ分泌显著高于由单独的高得多浓度(1000ng/ml,高30倍以上)的抗pd-1抗体诱导的ifn-γ分泌(即对于imp321,572.3>85.9;对于imp321r75a,511.2>85.9;对于lag3d1d4-接头2-ig,520.7>85.9)。
从这些结果得出结论,通过多种不同的可溶性lag-3衍生物(其各自保留与mhcii类阳性细胞结合的能力)协同增加由相对低剂量的抗pd-1抗体诱导的体外t细胞应答(如通过ifn-γ分泌测量)。还得出结论,如果将抗pd-1抗体与任何可溶性lag-3衍生物组合,则使用少30倍以上的抗pd-1抗体获得显著更大的体外t细胞应答。
- 还没有人留言评论。精彩留言会获得点赞!