事件特异性检测方法与流程
相关申请案的交叉参照本申请要求2014年10月10日所提申的美国临时专利申请第62/062,324号和2015年2月19日所提申的美国临时专利申请第62/118,320号的优先权,其完整内容各自全部结合在此供参考以用于所有目的。用电子方式提交的文本文件的说明用电子方式随同提交的文本文件的内容全部结合在此供参考:序列表的计算机可读格式拷贝(文件名:jrsi_062_02us_seqlist_st25.txt,记录日期:2015年10月04日,文件大小≈26千字节)。本发明涉及鉴别重组马铃薯植物(包括由此类植物制成的食物产品)中的遗传物质的方法。另外,本发明涉及本文所述方法中所用的材料,包括核苷酸引物、探针和非天然存在的核苷酸接合序列本身。
背景技术:
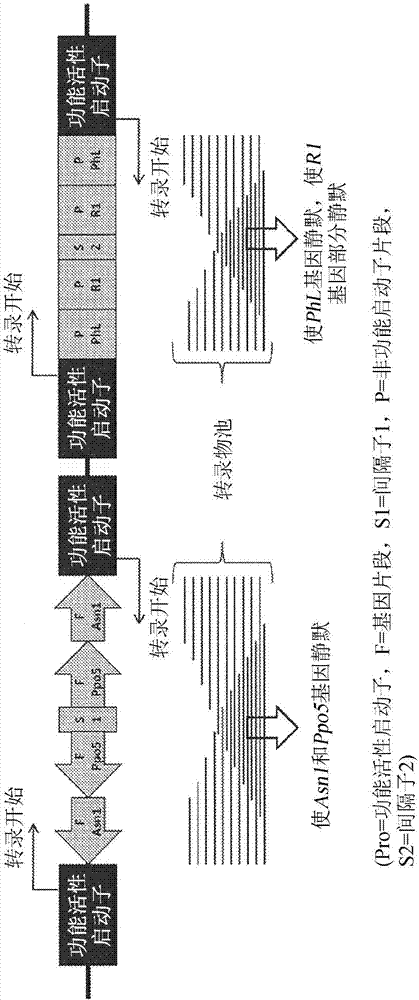
:马铃薯是全世界第四种最重要的食物作物且至今是最重要的蔬菜。目前美国的几乎每个州都商业种植马铃薯。马铃薯年产量在美国超过1800万吨且在全世界超过3亿吨。马铃薯的畅销主要来源于其通用性和营养价值。马铃薯可以新鲜、冷冻或干燥使用,或可以加工成面粉、淀粉或酒精。其含有复杂的碳水化合物且富含钙、烟酸和维生素c。食品工业中的马铃薯品质受两个关键因素影响:(1)马铃薯含有大量的天冬酰胺(一种在油炸或烘烤时会快速氧化形成丙烯酰胺(一种致癌产物)的非必需游离氨基酸);和(2)马铃薯极易发生酶促褐变和变色(当多酚氧化酶从碰损的马铃薯中的损伤质体外渗时发生的非期望事件)。在细胞质中,酶使苯酚发生氧化,其然后快速聚合而产生深色的色素。块茎含有大量的磷酸化淀粉,其中一些在储存期间降解而产生葡萄糖和果糖。这些还原糖当在超过120℃的温度下加热时与氨基酸反应而形成梅纳产物(maillardproducts),包括丙烯酰胺。涉及淀粉磷酸化的两种酶是水二激酶r1和磷酸化酶-l(r1和phl)。褐变也作为淀粉部分降解成葡萄糖和果糖的结果而以非酶促方式触发。为了解决前述两个因素,现在已经研发出马铃薯植物变种,其产生的块茎的丙烯酰胺含量低、黑斑碰伤耐受性增强且还原糖含量降低(美国专利第8,754,303号,“马铃薯栽培品种j3”;美国专利第8,710,311号,“马铃薯栽培品种f10”;美国专利第8,889,963号,“马铃薯栽培品种j55”;美国专利申请第14/072,487号,“马铃薯栽培品种e12”;和美国专利第8,889,964号,“马铃薯栽培品种w8”,其还耐受晚疫病,所述文献各自全部结合在此供参考)。这些被改造的马铃薯植物是通过引入遗传事件而非将来自细菌的外来核酸引入马铃薯中来研发。然而,工业中对用于鉴别这些被改造的马铃薯植物(包括由此类植物制成的食物产品)中所引入的遗传物质的方法和材料存在着巨大的需求。具体地说,对确定所指定的马铃薯或马铃薯产品是否含有所引入的特定遗传转化事件的方法和材料存在着需求。技术实现要素:本发明提供鉴别植物中的遗传转化事件的方法。在一些实施例中,植物是马铃薯。本文阐述的方法广泛地用于检测由植物转化产生的非天然存在的核苷酸接合点。在一些方面中通过农杆菌(agrobacterium)介导载体发生的植物转化事件产生了非天然存在的独特核苷酸接合序列,所述接合序列可以利用本发明方法检测。这些转化事件可以在任何植物物种中检测到,然而,在举例说明的某些实施例中,本文教导的方法教导了检测八种马铃薯栽培品种中的非天然存在的核苷酸接合点。在一些方面中,本发明提供了能够检测已转化的马铃薯的方法和材料,所述马铃薯包含所插入的核酸序列,所述核酸序列是马铃薯植物基因组原生的且不含农杆菌dna(病毒标记)或载体骨架序列。相反,插入马铃薯变种基因组中且被本文所述方法实施例检测的dna可以是非编码聚核苷酸,其是马铃薯原生的或野生马铃薯原生的,或马铃薯性相容植物原生的。所引入的这些核苷酸对于涉及表达黑斑碰伤、天冬酰胺聚积和还原糖聚积的基因起着静默作用。因此,所引入的这些基因引起已转化的马铃薯中的丙烯酰胺含量降低。另外,本文教导的方法能够检测到引入马铃薯中的赋予晚疫病抗性的基因,且在一些方面中能够检测到编码聚核苷酸。所插入的前述dna产生了非天然存在的独特核苷酸接合点,所述核苷酸接合点在自然界中未被发现。因此,本文教导的转化事件促成了非天然核苷酸“接合”序列在已转化马铃薯中的产生。非天然存在的这些核苷酸接合点可以当做一种指示特定遗传转化事件的存在的诊断类型来使用。如前所述,本文教导的方法不限于马铃薯。相反,本文中所利用的八种马铃薯栽培品种表明了本发明方法适用于检测任何植物物种中的由植物转化所产生的非天然存在的核苷酸接合序列。本发明技术能够通过利用特殊的定量pcr方法(包括独特设计的引物和探针)检测到非天然存在的这些核苷酸接合点。在一些方面中,本发明的探针结合到非天然存在的核苷酸接合序列。在一些方面中,利用传统pcr。在其它方面中,利用实时pcr。在一些方面中,利用定量pcr(qpcr)。因此,本发明涵盖利用两种常见方法实时检测pcr产物:(1)插入任何双股dna中的非特异性荧光染料,和(2)由寡核苷酸组成的序列特异性dna探针,所述寡核苷酸经荧光报告基因标记,从而允许只在探针与其互补序列杂交之后检测到。在一些方面中,只有非天然存在的核苷酸接合点将通过所教导的引物扩增,且因此可以通过非特异性染料或通过利用特异性杂交探针检测。另外,通过产生非天然存在的核苷酸接合序列(由所揭露的转化事件产生),本发明人已经创造出在自然界中未发现的独特核苷酸序列。这些序列可以加以分离且包含不存在于自然界中的核苷酸分子,产生此类分子无需人工干预。另外,结合到非天然存在的核苷酸接合序列的所揭露探针序列也是自然界中未发现的新核苷酸分子。因此,本发明的方面涉及非天然存在的核苷酸接合序列分子本身,以及能够在轻度到严格杂交条件下结合到所述非天然存在的核苷酸接合序列的其它核苷酸分子。在一些方面中,能够在轻度到严格杂交条件下结合到所述非天然存在的核苷酸接合序列的核苷酸分子称为“核苷酸探针”。因此,在本发明中,描述了检测e12、f10、j3、j55、v11、w8、x17和y9遗传转化事件的代表性方法。本发明提供了检测非天然存在的核苷酸接合序列的方法,所述核苷酸接合序列是由前述转化事件产生。这些八个实例充当能够检测任何植物物种中的非天然存在的核苷酸接合序列(由转化事件产生)的较大属中的种类。在一个实施例中,提供一种用于检测核酸样品中的植物转化事件的存在的定量pcr方法,所述方法包含:a)将以下组合:i)一对正向和反向核苷酸引物,ii)核苷酸探针,和iii)来自包含待检测的非天然存在的核苷酸接合点的所述样品的目标核苷酸序列;其中核苷酸探针结合到非天然存在的核苷酸接合点,或指示非天然存在的核苷酸接合点的存在的序列;和b)检测来自所述样品的目标核苷酸序列。在一个态样中,非天然存在的核苷酸接合点是由选自由以下组成的群组的植物转化事件产生:e12、f10、j3、j55、v11、w8、x17和y9,或其组合。在一些方面中,非天然存在的核苷酸接合序列发现于食物产品材料中。在特定方面中,食物产品材料是马铃薯食物产品材料。在一个态样中,目标核苷酸序列包含至少一种选自由seqidno:1-48组成的群组的核苷酸序列。在一个态样中,正向和反向核苷酸引物对以及核苷酸探针选自由seqidno:52-90组成的群组。在一个实施例中,正向核苷酸引物包含seqidno:52且反向核苷酸引物包含seqidno:53且核苷酸探针包含seqidno:54且核苷酸探针结合到非天然存在的核苷酸接合点。在一个实施例中,核苷酸探针结合e12事件的左侧或右侧接合点。在一个实施例中,正向核苷酸引物包含seqidno:55且反向核苷酸引物包含seqidno:56且核苷酸探针包含seqidno:57且核苷酸探针结合到非天然存在的核苷酸接合点。在一个实施例中,核苷酸探针结合f10事件的左侧或右侧接合点。在一个实施例中,正向核苷酸引物包含seqidno:58且反向核苷酸引物包含seqidno:59且核苷酸探针包含seqidno:60且核苷酸探针结合到非天然存在的核苷酸接合点。在一个实施例中,核苷酸探针结合j3事件的左侧或右侧接合点。在一个实施例中,正向核苷酸引物包含seqidno:61且反向核苷酸引物包含seqidno:62且核苷酸探针包含seqidno:63且核苷酸探针结合到指示存在于j55事件中的非天然存在的核苷酸接合点的存在的序列。在一个实施例中,正向核苷酸引物包含seqidno:64或67且反向核苷酸引物包含seqidno:65或68且核苷酸探针包含seqidno:66或69且核苷酸探针结合到非天然存在的核苷酸接合点。在一个实施例中,核苷酸探针结合v11事件的左侧或右侧接合点。在一个实施例中,正向核苷酸引物包含seqidno:70且反向核苷酸引物包含seqidno:71且核苷酸探针包含seqidno:72且核苷酸探针结合到指示存在于w8事件中的非天然存在的核苷酸接合点的存在的序列。在一个实施例中,正向核苷酸引物包含seqidno:73且反向核苷酸引物包含seqidno:74且核苷酸探针包含seqidno:75且核苷酸探针结合到非天然存在的核苷酸接合点。在一个实施例中,核苷酸探针结合x17事件的左侧或右侧接合点。在一个实施例中,正向核苷酸引物包含seqidno:76且反向核苷酸引物包含seqidno:77且核苷酸探针包含seqidno:78且核苷酸探针结合到非天然存在的核苷酸接合点。在一个实施例中,核苷酸探针结合y9事件的左侧或右侧接合点。在一个实施例中,正向核苷酸引物包含seqidno:79且反向核苷酸引物包含seqidno:80且核苷酸探针包含seqidno:81且核苷酸探针结合到非天然存在的核苷酸接合点。在一个实施例中,核苷酸探针结合与psim1278有关的内部agp/asn1接合点。在一个实施例中,正向核苷酸引物包含seqidno:82且反向核苷酸引物包含seqidno:83且核苷酸探针包含seqidno:84且核苷酸探针结合到非天然存在的核苷酸接合点。在一个实施例中,核苷酸探针结合与psim1278有关的内部接合点。在一个实施例中,正向核苷酸引物包含seqidno:85且反向核苷酸引物包含seqidno:86且核苷酸探针包含seqidno:87且核苷酸探针结合到非天然存在的核苷酸接合点。在一个实施例中,核苷酸探针结合与psim1678有关的内部接合点。在一个特定方面中,核苷酸探针结合到内部vnt1终止子/pagp接合点。在一个实施例中,正向核苷酸引物包含seqidno:88且反向核苷酸引物包含seqidno:89且核苷酸探针包含seqidno:90且核苷酸探针结合到非天然存在的核苷酸接合点。在一个实施例中,核苷酸探针结合与psim1678有关的内部接合点。在一个实施例中,检测到与psim1278或psim1678有关的非天然存在的构筑体特异性接合序列。在这些实施例中,检测到表6“psim1278和psim1678构筑体接合点”和图5中所示的非天然存在的接合序列。在一个实施例中,检测到与事件e12、f10、j3、j55、v11、w8、x17或y9有关的非天然存在的事件特异性接合序列。在这些实施例中,检测到表6和图6中所示的非天然存在的接合序列。在一个实施例中,核酸样品取自马铃薯植物,或马铃薯植物部分,或马铃薯衍生的食物产品,或食物产品中所用的基于马铃薯的成分。在一个实施例中,马铃薯植物部分是选自由以下组成的群组中的至少一种:马铃薯花、马铃薯花被片、马铃薯花瓣、马铃薯萼片、马铃薯花粉囊、马铃薯花粉、马铃薯种子、马铃薯叶、马铃薯叶柄、马铃薯茎、马铃薯根、马铃薯根茎、马铃薯匍匐茎、马铃薯块茎、马铃薯嫩枝、马铃薯细胞、马铃薯原生质体、马铃薯植物组织,和其组合。在一个实施例中,马铃薯衍生的食物产品是选自由以下组成的群组中的至少一种:马铃薯加工食物产品、马铃薯家畜饲料、法式炸薯条、马铃薯薯片、脱水马铃薯材料、马铃薯薄片、马铃薯颗粒、马铃薯蛋白质、马铃薯粉,和其组合。在一个实施例中,核酸样品取自马铃薯衍生的食物产品且其中能够检测到食物产品中存在至少一种选自由以下组成的群组的植物转化事件:e12、f10、j3、j55、v11、w8、x17、y9或其组合。在一个实施例中,转化事件能够在总食物产品的小于20%、小于10%、小于5%、小于1%和小于0.5%的水平下被检测到。在一个实施例中,转化事件能够在总食物产品的约0.1%到约5%范围内的水平下或在总食物产品的约0.2%到约5.0%范围内的水平下或在总食物产品的约0.1%到约10%范围内的水平下被检测到。在一个实施例中,提供一种分离的非天然存在的核酸接合序列,所述接合序列与选自由seqidno:1-48组成的群组的核酸具有至少75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%序列同源性。在另一个实施例中,提供一种分离的非天然存在的核酸接合序列,所述接合序列与通过将psim1278和/或psim1678插入马铃薯中所产生的非天然存在的核苷酸接合序列具有至少75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%序列同源性。在一些方面中,非天然存在的核苷酸接合序列描绘于图5(构筑体特异性接合点)和图6(事件特异性接合点)和表6中。在另一个实施例中,提供一种分离的非天然存在的核酸接合序列,其选自由seqidno:1-48组成的群组。在一个实施例中,非天然核苷酸接合序列发现于表6中。表6还包括基因插入序列的“左侧”或“右侧”是否含有接合序列的指示。在一些方面中,这些非天然核苷酸接合序列包含于较长核苷酸序列中。举例来说,表6中的序列可以包括于核苷酸序列中,所述核苷酸序列的长度包含30、40、50、60、70、80、90、100、200、300、400、500或更多个核苷酸。在一个实施例中,提供一种分离的非天然存在的核酸序列,其能够在轻度条件下与选自由seqidno:1-48组成的群组的核酸杂交。在一个实施例中,提供一种分离的非天然存在的核酸序列,其能够在严格条件下与选自由seqidno:1-48组成的群组的核酸杂交。在一些方面中,能够与seqidno:1-48杂交的前述核酸序列是核苷酸探针。在一些方面中,核苷酸探针被配置成用于实时pcr。在一些方面中,探针用报告分子标记。在另一个实施例中,提供了一种分离的非天然存在的核酸探针序列,其与选自由以下组成的群组的核酸具有至少75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%序列同源性:seqidno:54、57、60、63、66、69、72、75、78、81、84、87和90。在另一个实施例中,提供了一种分离的非天然存在的核酸探针序列,其选自由以下组成的群组:seqidno:54、57、60、63、66、69、72、75、78、81、84、87和90。如表7中所述,前述探针能够结合到非天然存在的如下事件特异性和构筑体特异性核苷酸接合序列:54(e12)、57(f10)、60(j3)、63(j55)、66(v11)、69(v11)、72(w8)、75(x17)、78(y9)、81(psim1278)、84(psim1278)、87(psim1678)和90(psim1678)。在另一个实施例中,提供了一种分离的非天然存在的核酸引物或探针序列,其与选自由seqidno:52-90组成的群组的核酸具有至少75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%序列同源性。在另一个实施例中,提供了一种分离的非天然存在的核酸引物或探针序列,其选自由seqidno:52-90组成的群组。本文中还教导了试剂盒,其包含根据前述揭露内容的正向和反向核苷酸引物和核苷酸探针,以及pcr和qpcr反应中所用的标准试剂。另外,在一个实施例中,本发明提供一种植物载体,称为psim1278,其包含:含有dna区段的两个拷贝的第一静默盒,所述dna区段按反义取向包含天冬酰胺合成酶-1基因(fasn1)的片段和多酚氧化酶-5基因的3'-未转译序列;和含有dna区段的两个拷贝的第二静默盒,所述dna区段按反义取向包含马铃薯磷酸化酶-l(pphl)基因的片段和马铃薯r1基因的片段。psim1278载体包含9,512bp骨架区域,所述骨架区域在植物转化之前向植物dna提供支撑性维持且在植物细胞转化后不转移到植物细胞中;和10,148bpdna插入区域,所述插入区域包含在转化后稳定整合到植物细胞基因组中的原生dna。另外,在另一个实施例中,本发明提供了一种植物载体,称为psim1678,其包含:含有dna区段的一个拷贝的第一表达盒,所述dna区段按有义取向包含rpi-vnt1晚疫病抗性基因(vnt1);和含有dna区段的两个拷贝的第二静默盒,所述dna区段按反义取向包含液泡酸转化酶(vinv)基因的片段。psim1678载体包含9,512bp骨架区域,所述骨架区域在植物转化之前向植物dna提供支撑性维持且在植物细胞转化后不转移到植物细胞中;和9,090bpdna插入区域,所述插入区域包含在转化后稳定整合到植物细胞基因组中的原生dna。本发明提供了检测前述dna插入区域是否已经引入植物中的方法。正如前述,dna的插入区域导致了独特非天然核苷酸接合序列的形成。这些接合序列可以发现于表6、图5和图6中以及本发明的其它地方中。另外,本发明可以检测psim1278和/或psim1678载体是否用于将遗传物质引入植物中。在一些实施例中,植物是马铃薯。然而,本文中揭露的本发明方法和材料可以用于检测引入任何适合植物中的所教导遗传事件的存在。由于本发明方法可以用于检测psim1278和/或psim1678载体是否用于将遗传物质引入植物中,因此所述方法可以检测任何植物或任何马铃薯植物中的转化,其利用此类载体将dna引入植物中。在实施例中,研发了优化dna提取的方法,以便向生物技术马铃薯食物产品提供事件特异性检测。本文在某些实施例中呈现了所教导的事件特异性或构筑体特异性检测方案中所用的所有设备、试剂和方法的详细说明。另外,在某些方面中,所呈现的数据支持了所教导的pcrdna提取程序的可重复性以及对dna品质的筛选。在所教导的pcr事件特异性或构筑体特异性检测的一些方面中,概述了实时pcr的所有程序以及能够检测生物技术马铃薯食物产品中的低水平(0.2-5.0%)的证据。在本发明的一个方面中,表达植物dna载体的一或多种静默盒的马铃薯植物变种选自由以下转化事件组成的群组:e12(褐色布尔班克(russetburbank))、j3(大西洋(atlantic))、j55(大西洋)、f10(褐色兰吉尔(rangerrusset))、w8(褐色布尔班克)、v11(斯诺登(snowden))、x17(褐色兰吉尔)和y9(大西洋)。本发明教导了适用于检测任一种前述遗传转化事件的存在的方法和材料。能够利用马铃薯植物的一部分检测到这些事件。在特定实施例中,可以利用马铃薯植物的任何部分分离出遗传物质以便并入本文教导的检测方法中。在一些方面中,所教导的方法将使用马铃薯植物的胚、原生质体、分生组织细胞、愈合组织、花粉、叶子、花粉囊、雌蕊、子叶、下胚轴、根、根尖、花、种子、叶柄、块茎、芽眼或茎作为源材料。再者,本发明提供了使用由马铃薯植物产生的任何产物作为源材料的方法。在一些方面中,食物产品是法式炸薯条、马铃薯薯片、脱水马铃薯材料、马铃薯薄片或马铃薯颗粒。在一个特定方面中,四种不同的马铃薯变种(褐色布尔班克、褐色兰吉尔、大西洋和斯诺登)是用psim1278构筑体转化。在一些实施例中,这些马铃薯称为“gen1”或代1或第一代且包括:e12(褐色布尔班克)、j3(大西洋)、j55(大西洋)、f10(褐色兰吉尔)和v11(斯诺登)。在另一态样中,褐色布尔班克、褐色兰吉尔和大西洋马铃薯变种是用psim1278与psim1678构筑体转化。在一些实施例中,这些马铃薯称为“gen2”或代2或第二代且包括:w8(褐色布尔班克)、x17(褐色兰吉尔)和y9(大西洋)。鉴别出展现低丙烯酰胺含量和/或增强的黑斑碰伤耐受性的八种事件:e12(褐色布尔班克)、j3(大西洋)、j55(大西洋)、f10(褐色兰吉尔)、w8(褐色布尔班克)、v11(斯诺登)、x17(褐色兰吉尔)和y9(大西洋)。w8、x17和y9事件也展现增强的晚疫病抗性。所有这些事件均能够被本文中揭露的事件特异性或构筑体特异性检测方法检测到。本发明的一个实施例教导了构筑体特异性和变种/事件特异性引物和探针和qpcr条件以在遗传上鉴别每个转化事件。在一个方面中,本发明教导了用于鉴别转化事件的qpcr方法,所述转化事件选自由以下组成的群组:e12(褐色布尔班克)、j3(大西洋)、j55(大西洋)、f10(褐色兰吉尔)、w8(褐色布尔班克)、v11(斯诺登)、x17(褐色兰吉尔)和y9(大西洋)。本发明的一个实施例教导了一种检查样品中存在或不存在衍生自一或多种转基因植物事件的材料的方法,所述方法包含以下步骤:(a)检测样品中存在或不存在包含具有seqidno1-48的核苷酸序列中的一个、超过一个或全部的核酸;和(b)根据样品中所述seqidnos的存在或不存在来断定样品是否含有来自植物转化事件的遗传物质。这些序列指示由转化事件产生的非天然存在的核苷酸接合点。这些非天然接合序列以及基因插入序列的“左侧”或“右侧”是否含有接合序列的指示尤其概述于表6中。另外,所述事件的可视化描绘尤其可以发现于图5和图6中。在一些方面中,使用pcr扩增法检测样品存在或不存在核酸。在一些方面中,使用实时pcr扩增。在一些方面中,所教导的方法是使用表7中的具有seqidno:52-90的引物/探针组,或所述引物/探针组的变异体。在一些方面中,seqidno:49-51用于检测对照物。在一些实施例中,样品包含存在于事件e12中的psim1278插入区域。在另一个实施例中,除磷酸化酶-l和二激酶r1基因的内源马铃薯启动子的反向重复序列之外,事件e12还含有有效抑制内源天冬酰胺合成酶-1基因和内源多酚氧化酶-5基因表达的马铃薯dna反向重复序列。在一些实施例中,样品包含存在于事件f10中的psim1278插入区域。在另一个实施例中,除磷酸化酶-l和二激酶r1基因的内源马铃薯启动子的反向重复序列之外,事件f10还含有有效抑制内源天冬酰胺合成酶-1基因和内源多酚氧化酶-5基因表达的马铃薯dna反向重复序列。在一些实施例中,样品包含存在于事件j3中的psim1278插入区域。在另一个实施例中,除磷酸化酶-l和二激酶r1基因的内源马铃薯启动子的反向重复序列之外,事件j3还含有有效抑制内源天冬酰胺合成酶-1基因和内源多酚氧化酶-5基因表达的马铃薯dna反向重复序列。在一些实施例中,样品包含存在于事件j55中的psim1278插入区域。在另一个实施例中,除磷酸化酶-l和二激酶r1基因的内源马铃薯启动子的反向重复序列之外,事件j55还含有有效抑制内源天冬酰胺合成酶-1基因和内源多酚氧化酶-5基因表达的马铃薯dna反向重复序列。在一些实施例中,样品包含存在于事件v11中的psim1278插入区域。在另一个实施例中,除磷酸化酶-l和二激酶r1基因的内源马铃薯启动子的反向重复序列之外,事件v11还含有有效抑制内源天冬酰胺合成酶-1基因和内源多酚氧化酶-5基因表达的马铃薯dna反向重复序列。在一些实施例中,样品包含存在于事件w8中的psim1278插入区域和psim1678插入区域。在另一个实施例中,除磷酸化酶-l和二激酶r1基因的内源马铃薯启动子的反向重复序列之外,事件w8还含有有效抑制内源天冬酰胺合成酶-1基因和内源多酚氧化酶-5基因表达的马铃薯dna反向重复序列。在另一个实施例中,除有效表达晚疫病抗性基因rpi-vnt1的马铃薯有义dna之外,事件w8还含有有效抑制内源液泡酸转化酶基因表达的马铃薯dna反向重复序列。在一些实施例中,样品包含存在于事件x17中的psim1278插入区域和psim1678插入区域。在另一个实施例中,除磷酸化酶-l和二激酶r1基因的内源马铃薯启动子的反向重复序列之外,事件x17还含有有效抑制内源天冬酰胺合成酶-1基因和内源多酚氧化酶-5基因表达的马铃薯dna反向重复序列。在另一个实施例中,除有效表达晚疫病抗性基因rpi-vnt1的马铃薯有义dna之外,事件x17还含有有效抑制内源液泡酸转化酶基因表达的马铃薯dna反向重复序列。在一些实施例中,样品包含存在于事件y9中的psim1278插入区域和psim1678插入区域。在另一个实施例中,除磷酸化酶-l和二激酶r1基因的内源马铃薯启动子的反向重复序列之外,事件y9还含有有效抑制内源天冬酰胺合成酶-1基因和内源多酚氧化酶-5基因表达的马铃薯dna反向重复序列。在另一个实施例中,除有效表达晚疫病抗性基因rpi-vnt1的马铃薯有义dna之外,事件y9还含有有效抑制内源液泡酸转化酶基因表达的马铃薯dna反向重复序列。在一个实施例中,本发明提供了qpcr方案,其使用5'端经6-羧基荧光素标记且3'端经黑洞淬灭剂tm(blackholequencherstm)标记的核苷酸探针。然而,所属领域的技术人员将了解,可以建构任何杂交探针和任何报告分子。在一些实施例中,pcr扩增效率是90%到110%。在另一个实施例中,pcr扩增线性度是根据r2值测量。在一些实施例中,r2值大于或等于0.98。在一些实施例中,pcr扩增可以使用60℃的粘接/延长温度、在34个与35个扩增循环之间检测至少24pg的马铃薯叶dna。在一些实施例中,pcr扩增是稳定的。在一些实施例中,pcr扩增的热循环条件包含:(a)在95℃历时600秒的一个pcr循环用于初始变性;(b)四十五个pcr循环:在95℃历时15秒用于变性,然后在60℃历时60秒用于粘接/延长。在一些实施例中,热循环条件包含:(a)在95℃历时600秒的一个pcr循环用于初始变性;(b)四十个pcr循环:在95℃历时15秒用于变性,然后在60℃历时15秒用于粘接,然后在72℃历时10秒用于延长。在一些实施例中,样品包含植物或其一部分,包括花、被片、、瓣片、萼片、花粉囊、花粉、种子、叶子、叶柄、茎、根、根茎、匍匐茎、块茎或嫩枝或其一部分、植物细胞、植物原生质体和/或植物组织,和/或源于植物的材料,优选食物或饲料,包括加工食物或饲料。在一些实施例中,加工食物选自由以下组成的群组:法式炸薯条、马铃薯薯片、脱水马铃薯材料、马铃薯薄片、马铃薯蛋白质、马铃薯面粉和马铃薯颗粒。在一些实施例中,样品包含食物产品。在其它实施例中,食物产品包含衍生自事件e12、f10、j3、j55、v11、w8、x17和/或y9的材料的混合物。在一些实施例中,材料包含马铃薯炸薯条、马铃薯薯片、马铃薯薄片和马铃薯块茎。在一些实施例中,衍生自事件e12、f10、j3、j55、v11、w8、x17和/或y9的马铃薯块茎和/或炸薯条构成总食物产品的约小于1%。在一些实施例中,衍生自事件e12、f10、j3、j55、v11、w8、x17和/或y9的马铃薯块茎和/或炸薯条构成总食物产品的约小于0.5%。在一些实施例中,衍生自事件e12、f10、j3、j55、v11、w8、x17和/或y9的马铃薯块茎和/或炸薯条构成总食物产品的约0.2%。在一些实施例中,衍生自事件e12、f10、j3、j55、v11、w8、x17和/或y9的马铃薯薯片构成总食物产品的约小于10%。在一些实施例中,衍生自事件e12、f10、j3、j55、v11、w8、x17和/或y9的马铃薯薯片构成总食物产品的约5%。在一些实施例中,衍生自事件e12、f10、j3、j55、v11、w8、x17和/或y9的马铃薯薄片构成总食物产品的约小于15%。在一些实施例中,衍生自事件e12、f10、j3、j55、v11、w8、x17和/或y9的马铃薯薄片构成总食物产品的约小于8%。在一些实施例中,衍生自事件e12、f10、j3、j55、v11、w8、x17和/或y9的马铃薯薄片构成总食物产品的约2.5%。本文中教导了一种分离的核苷酸序列,其包含选自seqidno1-90的序列。本文中教导了一种用于检查样品中潜在性存在或不存在衍生自一或多种转化植物事件的材料的试剂盒,所述试剂盒包含一个、超过一个或全部引物/探针组,或所述引物/探针组的变异体。本发明的试剂盒还可以包括所述试剂盒的使用说明/说明书。除上述示例性方面和实施例之外,其它方面和实施例通过研究以下说明将变得显而易见。附图说明图1描绘了psim1278转化载体。位于左侧的载体骨架区域的长度是9,512bp,因为其始于位置10,149bp且终于位置19,660bp。骨架dna主要由细菌dna组成,其在植物转化之前,向dna插入序列提供支撑性维持。包括侧接边界序列的dna插入区域(右侧)的长度是10,148bp(从1bp到10,148bp)。转化后,dna插入序列稳定地整合到马铃薯基因组中。图2提供了psim1278转化载体中所插入的dna插入序列中的两个静默盒的示意图。每个静默盒含有由间隔子分隔的两个基因片段的两个拷贝。包含四种靶向基因(即asn-1、ppo-5、ph1和r1)片段的dna区段的两个拷贝作为反向重复序列插入两个命名为pro的趋同启动子之间,这些启动子主要在块茎中发挥活性。含有所得静默盒的植物在块茎中产生了多种多样的且未发生聚腺苷酸化的rna分子,其动态地且有力地使预定的目标基因静默。由于趋同性转录引起了碰撞性转录,因此rna分子尺寸通常小于所使用的两个启动子之间的距离。图3描绘了本发明的psim1678转化载体。位于左侧的载体骨架区域的长度是9,512bp,因为其始于位置9,091bp且终于位置18,602bp。骨架dna主要由细菌dna组成,其在植物转化之前,向dna插入序列提供支撑性维持。包括侧接边界序列的dna插入区域(右侧)的长度是9,090bp(从1bp到9,090bp)。转化后,dna插入序列稳定地整合到马铃薯基因组中。图4a-d展示了马铃薯事件e12、f10、j3和j55中的dna插入序列的结构图。缩写如下:lb=左侧边界(25个碱基对序列),类似于根癌农杆菌(a.tumefaciens)t-dna边界;agp=adp葡萄糖焦磷酸化酶基因启动子;gbs=颗粒结合的淀粉合成酶基因启动子;rb=右侧边界(25个碱基对序列),类似于根癌农杆菌t-dna边界;asn1=dna凝胶印迹杂交中所用且衍生自天冬酰胺合成酶1(asn1)基因的探针;fasn1=天冬酰胺合成酶1(asn1)基因片段;fppo5=多酚氧化酶5(ppo5)基因片段;pphl=第二反向重复盒中所用的磷酸化酶-l基因启动子的片段;phl=dna凝胶印迹杂交中所用且衍生自磷酸化酶-l基因启动子的探针;prl=第二反向重复盒中所用的水二激酶r1基因启动子的片段;间隔子=每个反向重复序列的各臂之间的序列;rv=限制酶ecorv;hd=限制酶hindiii;r1=限制酶ecori;sc=限制酶scai。深黑色线表示dna凝胶印迹杂交中所用的dna插入序列的多个区域的探针。弯箭头表示每个相应启动子的转录起点。白箭头描绘所指定基因或启动子片段的每个股(有义股或反义股)在每个反向重复盒中的方向。编号描绘dna插入序列中的核苷酸位置。核苷酸位置1是lb之后的agp启动子的起点。表2给出了关于dna插入序列中的各种元件的其它细节。栽培品种描绘如下:图4a-f10插入序列;图4b-e12插入序列;图4c-j3插入序列;图4d-j55插入序列。图5展示了质体构筑体psim1278和psim1678的插入区域中的构筑体特异性接合点。插入区域下方的编号表示构筑体特异性接合点且对应于表6中的seqidno:3-16和seqidno:42-48。psim1278插入区域中的缩写如下:lb=左侧边界(25个碱基对序列),类似于根癌农杆菌t-dna边界;pagp=adp葡萄糖焦磷酸化酶基因启动子;pgbss=颗粒结合的淀粉合成酶基因启动子;rb=右侧边界(25个碱基对序列),类似于根癌农杆菌t-dna边界;asn=天冬酰胺合成酶1(asn1)基因片段;ppo=多酚氧化酶5(ppo5)基因片段;phl=第二反向重复盒中所用的磷酸化酶-l基因启动子的片段;rl=第二反向重复盒中所用的水二激酶r1基因启动子的片段;红盒=每个反向重复序列的各臂之间的间隔序列。psim1678插入区域中的缩写如下:lb=左侧边界(25个碱基对序列),类似于根癌农杆菌t-dna边界;pagp=adp葡萄糖焦磷酸化酶基因启动子;pgbss=颗粒结合的淀粉合成酶基因的启动子;rb=右侧边界(25个碱基对序列),类似于根癌农杆菌t-dna边界;pvnt1=晚疫病抗性基因rpi-vnt1的原生启动子;vnt1=赋予晚疫病(马铃薯晚疫病菌(phytophthorainfestans))抗性的基因;tvnt1=晚疫病抗性基因rpi-vnt1的原生终止子;inv=液泡酸转化酶基因片段。箭头描绘了所指定基因或启动子片段的每个股(有义股或反义股)在每个反向重复盒中的方向或所指定启动子的转录方向。表2和4给出了关于每个插入区域中的每个元件的其它细节。图6a-i展示了天生tm1.0插入型(psim1278)栽培品种e12、f10、v11、j3、j55和e56以及天生tm2.0插入型(psim1278和psim1678)栽培品种w8、x17和y9的事件特异性接合点和构筑体特异性接合点。插入区域下方的编号表示构筑体特异性接合点且对应于表6中所发现的seqidno。插入区域上方的编号表示事件特异性接合点且对应于表6中所发现的seqidno。缩写如图5中所述。栽培品种插入序列描绘如下:图6a-e12结构;图6b-f10结构;图6c-v11结构;图6d-j3结构;图6e-j55结构;图6f-e56结构;图6g-w8结构;图6h-x17结构;图6i-y9结构。图7图解说明了针对褐色兰吉尔事件f10块茎、炸薯条和薄片与大西洋事件j3块茎和薯片的食物混合物进行的所有dna分离均能够在磨碎的天生tm(innatetm)食物产品按最低百分比混合于商业变种食物产品中的情况下放大。在2.5%时,f10薄片中存在一个假阴性。这些结果表明所揭露的方法产生的dna品质足以用于qpcr测试。天生tm是j.r.新普洛特(j.r.simplot)所用的商标,其表示已经用psim1278和/或psim1678转化载体转化的马铃薯植物和由所述植物制成的食物产品。图8图解说明了使用如表1和表2中所述的dna序列构筑质体psim1278的方法。起始载体pcambia1301含有位于最后psim1278骨架中的复制起点。图9图解说明了psim1278中的t-dna表达盒的构造。使用融合pcr扩增元件1a(pagp-第1拷贝)、1b(pagp-第2拷贝)、2(asn1、ppo5)、3(ppo5、asn1)、4(pgbss-第1拷贝)和7(间隔子1、ppo5、asn1)。元件5(phl、r1)和6(间隔子2、r1、phl、pgbss)是由蓝鹭生物技术有限公司(blueheronbiotechnology,inc.)(华盛顿州波塞尔(bothell,wa))基于马铃薯基因组的序列合成。元件8、9和10是通过将图中所示的结构单元连接而产生。最后,跨越所期望的表达盒而产生三个片段10、11和6。将这些三个片段连接且插入图8所示的kpni-saci限制位点中以产生psim1278。图10图解说明了使用如表3和表4中所述的dna序列建构质体psim1678的方法。起始载体psim1278含有最后psim1678骨架。所属领域的技术人员能够使用实例和图9和图10转化任何马铃薯植物,所述马铃薯植物然后含有可根据本文教导的方法检测到的非天然存在的核苷酸接合点。具体实施方式定义在以下说明和表格中,使用许多术语。为了清楚且一致地了解说明书和权利要求书(包括此类术语中所指定的范围),提供以下定义。术语“一(a)”或“一(an)”是指一或多种所述实体;举例来说,“一种引物”是指一或多种引物或至少一种引物。因而,术语“一(a)”(或“一(an)”)、“一或多种”和“至少一种”在本文中可互换地使用。另外,通过不定冠词“一(a)”或“一(an)”提及“一个元件”并不排除存在超过一个元件的可能性,除非上下文明确要求存在一个且仅存在一个元件。如本文中所用,术语“等位基因”是涉及一种性状或特征的基因的一或多种替代形式中的任一种。在二倍体细胞或生物体中,所指定基因的两个等位基因占据一对同源染色体上的相应基因座。如本文中所用,术语“氨基酸序列”包括从植物中分离的、植物原生的或植物中天然存在的寡肽、肽、多肽或蛋白质和其片段,或以合成方式制备,但是包含内源对应物的核酸序列。如本文中所用,术语“人工操纵”是指通过手或通过机械方式或重组方式(如通过基因工程技术)移动、配置、操作或控制植物或植物细胞,以便产生在生物学、生物化学、形态学或生理学表型和/或基因型上不同于未被操纵的天然存在的对应物的植物或植物细胞。如本文中所用,术语“无性繁殖”是指通过由叶插条、茎插条、根插条、块茎芽眼、匍匐茎、单一植物细胞原生质体、愈合组织和类似者产生完整植物来产生后代,其不涉及配偶子融合。如本文中所用,术语“骨架”是指二元载体的核酸序列,其不包括旨在转移的dna插入序列。如本文中所用,术语“回交”是一种方法,其中育种者反复地使杂种后代与亲代之一(例如第一代杂种f1与f1杂种的亲代基因型之一)返交。如本文中所用,术语“黑斑碰伤”描述一种病状,其中发现于碰伤块茎组织中的黑斑是细胞损伤之后产生色素(称为黑色素)的结果且赋予组织褐色、灰色或黑色外观。当酚底物和适当的酶由于细胞损伤而彼此接触时,形成黑色素。损伤无需破坏细胞。然而,底物和酶必须发生混合(通常在组织受到冲击时)。黑斑主要存在于刚好位于脉管环下方的环髓组织中,但可大以足以包括皮层组织的一部分。如本文中所用,术语“边界样序列”是指以下。“边界样”序列是从所选择的被改造的植物物种中分离,或从与被改造的植物物种性相容的植物中分离,且发挥与农杆菌的边界序列类似的功能。也就是说,本发明的边界样序列促进和有助于其所连接的聚核苷酸的整合。本发明的dna插入序列优选含有边界样序列。dna插入序列的边界样序列的长度在5-100bp、10-80bp、15-75bp、15-60bp、15-50bp、15-40bp、15-30bp、16-30bp、20-30bp、21-30bp、22-30bp、23-30bp、24-30bp、25-30bp或26-30bp之间。dna插入序列左侧和右侧边界序列可以从待被改造的植物的基因组分离和/或可以是待被改造的植物的基因组原生的。在核苷酸序列中,dna插入序列边界样序列与任何已知的农杆菌衍生t-dna边界序列不相同。因此,dna插入序列边界样序列可以具有1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20个或更多个与来自农杆菌种(如根癌农杆菌(agrobacteriumtumefaciens)或发根农杆菌(agrobacteriumrhizogenes))的t-dna边界序列不同的核苷酸。即,本发明的dna插入序列边界或边界样序列与来自农杆菌种(如根癌农杆菌或发根农杆菌)的t-dna边界序列具有至少95%、至少90%、至少80%、至少75%、至少70%、至少60%或至少50%序列一致性,但不具有100%序列一致性。如本文中所用,描述性术语“dna插入序列边界”和“dna插入序列边界样”是可交换的。边界样序列可从植物基因组中分离且进行修饰或突变以改变其能够将核苷酸序列整合到另一核苷酸序列中的效率。可以将其它聚核苷酸序列添加到或并入本发明的边界样序列内。因此,dna插入序列左侧边界或dna插入序列右侧边界可以加以修饰以具有5'-和3'-多克隆位点,或额外的限制位点。dna插入序列边界序列可以加以修饰以增加来自伴随载体的骨架dna不整合到植物基因组中的可能性。如本文中所用,术语“薯片”是马铃薯薄切片,其已经深度油炸或烘烤直至松脆。马铃薯薯片通常充当开胃食品、配菜或零食。薯片也称为油炸土豆片。如本文所使用,如本说明书和权利要求书中所用的动词“包含”和其词形变化形式在其非限制性意义上用于意指包括所述词语之后的项目,但不排除未专门提到的项目。如本文中所用,“基本上由某些元件组成”的组合物限于包括那些元件,以及实质上不影响本发明组合物的基本和新颖特征的那些元件。因此,只要组合物不影响本发明的基本和新颖特征,即,不含不来自所选植物物种或与所选植物物种性相容的植物的外来dna,那么所述组合物可以视为以措辞“基本上由……组成”为特征的本发明组合物的组分。如本文中所用,术语“子叶”是种子叶子的一种类型。子叶含有种子的食物储存组织。如本文中所用,术语“简并引物”是一种寡核苷酸,其含有足够的核苷酸变异,使得其当与类似、但不严格同源的序列杂交时可以接纳碱基错配。如本文中所用,术语“双子叶植物”或“双子叶”是一种开花植物,其胚具有两个种子叶子或子叶。双子叶植物的实例包括(但不限于)烟草、番茄、马铃薯、甘薯、木薯、豆科植物(包括苜蓿和大豆)、胡萝卜、草莓、莴苣、橡树、枫树、胡桃、玫瑰、薄荷、南瓜、菊花和仙人掌。如本文中所用,术语“dna插入序列”根据本发明是指待插入植物基因组中的dna插入序列包含那种植物原生的聚核苷酸序列或具有那种原生的基因元件。在一个实例中,举例来说,本发明的马铃薯变种j3的dna插入序列是10,147bp非编码聚核苷酸,其是马铃薯或野生型马铃薯或马铃薯性相容植物原生的,在转化后稳定整合到植物细胞基因组中且使涉及表达黑斑碰伤、天冬酰胺聚积和衰老甜化的基因静默。dna插入序列优选包含两个表达盒且插入称为psim1278转化载体的转化载体中。第一个盒包含天冬酰胺合成酶-1基因(asn1)和多酚氧化酶-5基因(ppo5)的片段,其以反向重复序列形式配置于adp葡萄糖焦磷酸化酶基因(agp)的agp启动子与颗粒结合的淀粉合成酶基因(gbss)的gbss启动子之间。这些启动子主要在块茎中发挥活性。第二个盒的功能是使淀粉相关基因二激酶-r1(r1)和磷酸化酶-l基因(phl)的启动子静默。这个盒包含淀粉相关基因二激酶-r1(r1)和磷酸化酶-l基因(phl)的启动子的片段,其可操作地连接到与第一个盒相同的agp和gbss启动子。这些表达盒含有仅来自所选植物物种或来自与所选植物物种性相容的植物的dna。如本文中所用,术语“非天然核苷酸接合点”或“非天然存在的核苷酸接合点”是指不存在于自然界中的核苷酸序列。相反,这些序列是通过遗传转化事件形成。正如前述,本文所述的遗传转化事件是用不含有非天然马铃薯dna的表达盒产生。因此,这些非天然核苷酸接合点是由马铃薯核苷酸组成,但是这些核苷酸的基因配置不存在于自然界中,而是在马铃薯遗传转化期间由人工操纵产生。表6描述了这些接合序列的实施例。对于事件特异性接合序列来说,表6图解说明:在接合点的一侧上发现来自已经转化的马铃薯的核苷酸,且在接合点的另一侧上发现已经通过转化事件插入的核苷酸。因此,对于事件特异性接合序列来说,非天然核苷酸接合点代表了其中所插入的核苷酸与马铃薯植物的原生核苷酸相遇的边界。表6中还图解说明了构筑体接合点。这些独特的接合序列存在于所有转化事件中且不是特定事件所特有的,而是存在于利用psim1278构筑体和/或psim1678构筑体执行转化的任何事件中。这些接合点代表了构筑体内所含的各种基因元件的序列,例如其中asn和ppo(asn/ppo)元件会合在一起的接合点。参看图5,容易目测到这些构筑体特异性接合点。如本文中所用,术语“效率”是指实时pcr分析的标志。理想的qpcr(定量pcr)反应在-3.32斜率下具有100%效率,其与pcr产物在每个循环期间的完全倍增相关。然而,斜率在-3.1与-3.6之间且效率在90%与110%之间通常视为可接受的(康密辛c.a.(commission,c.a.)(2009).gmo实验室欧洲网(engl)gmo测试分析方法的最小性能要求定义(definitionofminimumperformancerequirementsforanalyticalmethodsofgmotestingeuropeannetworkofgmolaboratories(engl))),(2008年10月),1-8))。效率是根据复制的标准曲线确立。扩增效率是根据标准曲线的对数线性部分的斜率测定且如下计算:e=(10(-1/斜率)-1)*100.(布什丁s.a.(bustin,s.a.)等人(2009).miqe指南:关于定量实时pcr实验的最小公开信息(themiqeguidelines:minimuminformationforpublicationofquantitativereal-timepcrexperiments).《临床化学(clinicalchemistry)》,55(4),1-12.doi:10.1373/clinchem.2008.112797)。如本文中所用,术语“胚”是成熟种子中所含的不成熟植物。如本文中所用,术语“事件”是指发生于一种植物细胞中的独特dna重组事件,其然后用于产生完整的转基因植物。植物细胞用携带所关注的dna插入序列的二元转化载体转化。转化的细胞再生成转基因植物,且每种所产生的转基因植物代表一种独特事件。使用分子技术(如南方印迹杂交或pcr)证实每个转化事件。每个衍生事件通过缩写(例如j3)鉴别。不同事件在细胞基因组中的dna插入序列的拷贝数、dna插入序列拷贝的配置和/或基因组中的dna插入位置方面具有差异。可以进一步分析且研究引起dna插入序列中的基因产生最佳表达和展现性状的事件。如本文中所用,关于核酸的术语“外来”意指所述核酸来源于非植物生物体,或来源于物种与待转化的植物不同的植物,或来源于与待转化的植物不可杂交的植物,或不属于目标植物的物种。根据本发明,外来dna或rna代表天然存在于真菌、细菌、病毒、哺乳动物、鱼或鸟类的基因组成中,而非天然存在于待转化的植物中的核酸。因此,外来核酸是编码例如并非由所转化植物天然产生的多肽的核酸。外来核酸并非必须编码蛋白质产物。根据本发明,所期望的基因内植物是不含任何整合到其基因组中的外来核酸的植物。如本文中所用,术语“薄片”是指马铃薯薄片,其通过烹调、磨碎和脱水工业过程产生,以产生包装便利食物,其可以通过添加热水或牛奶来复原,从而产生紧密近似的马铃薯泥。如本文中所用,术语“炸薯条”是深度油炸的马铃薯条。炸薯条是热吃的油炸马铃薯细长片,其软或脆,且通常作为午餐或晚餐的小菜食用,或作为零食食用。如本文中所使用,术语“基因”是指编码区且不包括那个区域的5'-或3'-的核苷酸序列。功能基因是可操作地连接到启动子或终止子的编码区。可使用转化或各种育种方法将基因引入物种的基因组(无论来自不同物种或来自相同物种)。如本文中所用,术语“所转化的基因”或“转化”是指通过称为回交的植物育种技术研发出的植物,其中除通过回交技术、通过基因工程或通过突变转移到变种中的一或多种基因之外,还回收了变种的基本上全部的所期望形态学和生理学特征。也可以转移一或多个基因座。如本文中所用,术语“基因重排”是指可以在活体内以及活体外自发发生的基因元件再结合,从而引入遗传物质的新组织。举例来说,不同染色体基因座的聚核苷酸拼接在一起可以在植物发育和性重组期间活体内自发进行。因此,在活体外通过非天然基因修饰技术进行的基因元件重组与也可以通过活体内性重组进行的重组事件类似。如本文中所用,术语“下胚轴”是子叶与根之间的胚或幼苗的一部分。因此,其可以视为芽与根之间的过渡区。如本文中所用,术语“同框”是指以下。核苷酸三联体(密码子)转译成植物细胞中的所期望重组蛋白的新生氨基酸序列。具体来说,本发明涵盖在阅读框架中连接到第二核酸的第一核酸,其中第一核苷酸序列是基因且第二核苷酸是启动子或类似调节元件。如本文中所用,术语“整合”是指来自所选植物物种或来自物种与所选植物相同的植物或来自与所选植物物种性相容的植物的核酸序列插入所选植物物种细胞基因组中。“整合”是指仅将原生基因元件并入植物细胞基因组中。为了整合原生基因元件,如通过同源重组,本发明可以“使用”非天然dna作为此方法中的步骤。因此,本发明对特定dna分子的“使用”与将特定dna分子“整合”到植物细胞基因组中进行了区分。如本文中所用,术语“引入”是指通过包括感染、转染、转化或转导的方法将核酸序列插入细胞中。如本文中所用,术语“分离”是指实体上从其正常原生环境中分离出的任何核酸或化合物。被分离的材料可以维持在含有例如溶剂、缓冲液、离子或其它组分的适合溶液中,且可以呈提纯或或未提纯的形式。如本文中所用,术语“晚疫病”是指一种马铃薯疾病,其由卵菌纲马铃薯晚疫病菌引起且还称为‘马铃薯晚疫病’,其可以感染且摧毁马铃薯植物的叶子、茎、果实和块茎。如本文中所用,术语“前导序列”是指在基因之前(或5')且被转录、但不被转译的序列。如本文中所用,术语“检测水平”或“lod”是样品中的分析物的最低量或浓度,其能够可靠地检测到,但不一定能够定量,如根据gmo实验室欧洲网通过单一实验室验证所证明。如本文中所用,术语“线性度”是指优化实时pcr分析的标志且根据通过线性回归分析所得的r2值测定,其应该≥0.98(布什丁等人,2009)。如本文中所用,术语“基因座”赋予一或多种性状,如雄性不育、除草剂耐受性、昆虫抗性、疾病抗性、糯性淀粉、改良的脂肪酸代谢、改良的植酸代谢、改良的碳水化合物代谢和改良的蛋白质代谢。性状可以通过例如引入变种基因组中的天然存在的基因(通过回交、天然或诱发突变来引入)或通过遗传转化技术引入的转基因赋予。基因座可以包含一或多个在单一染色体位置整合的等位基因。如本文中所用,术语“可销售产量”是在直径为2英寸与4英寸之间所收集的全部块茎的重量。可销售产量是按cwt(英担(hundredweight))度量,其中cwt=100磅。如本文中所用,术语“单子叶植物”或“单子叶”是一种开花植物,其胚具有一个子叶或种子叶子。单子叶植物的实例包括(但不限于)草坪草、玉米、稻米、燕麦、小麦、大麦、高粱、兰花、鸢尾、百合、洋葱和棕榈。如本文中所用,术语“原生”基因元件是指天然存在于、起源于或属于待转化的植物的基因组中的核酸。因此,从待转化的植物或植物物种的基因组中分离或从与待转化的植物物种性相容或可杂交的植物或物种中分离的任何核酸、基因、聚核苷酸、dna、rna、mrna或cdna分子是所述植物物种“原生”的,即原产的。换句话说,原生基因元件代表植物育种者可获得的用于通过传统植物育种改良植物的全部遗传物质。根据本发明,原生核酸的任何变异体也视为“原生”。就此而言,“原生”核酸还可以从植物或其性相容物种中分离且进行修饰或突变,使得所得变异体在核苷酸序列方面与从植物中分离的未被修饰的原生核酸具有大于或等于99%、98%、97%、96%、95%、94%、93%、92%、91%、90%、89%、88%、87%、86%、85%、84%、83%、82%、81%、80%、79%、78%、77%、76%、75%、74%、73%、72%、71%、70%、69%、68%、67%、66%、65%、64%、63%、62%、61%或60%相似性。原生核酸变异体的核苷酸序列还可以具有小于约60%、小于约55%或小于约50%相似性。从植物中分离的“原生”核酸还可以编码从核酸转录和转译的天然存在的蛋白质产物的变异体。因此,原生核酸可以编码一种蛋白质,所述蛋白质的氨基酸序列与核酸从中分离的植物中所表达的未修饰的原生蛋白质具有大于或等于99%、98%、97%、96%、95%、94%、93%、92%、91%、90%、89%、88%、87%、86%、85%、84%、83%、82%、81%、80%、79%、78%、77%、76%、75%、74%、73%、72%、71%、70%、69%、68%、67%、66%、65%、64%、63%、62%、61%或60%相似性。如本文中所用,术语“天然存在的核酸”发现于所选植物物种的基因组内且可以是dna分子或rna分子。植物物种的基因组中通常存在的限制位点的序列可以工程改造成外源性dna分子,如载体或寡核苷酸,即使限制位点在实体上未与那个基因组分离。因此,本发明允许合成产生核苷酸序列,如限制酶识别序列,只要那个序列天然存在于所选植物物种的基因组中或存在于与待转化的所选植物物种性相容的植物中即可。如本文中所用,术语“可操作地连接”是指两种或超过两种分子的组合,以使得其组合时在植物细胞中正确发挥功能。举例来说,当启动子控制结构基因的转录时,启动子可操作地连接到结构基因。如本文中所用,术语“植物”包括(但不限于)被子植物和裸子植物,如马铃薯、番茄、烟草、苜蓿、莴苣、胡萝卜、草莓、甜菜、木薯、甘薯、大豆、玉米、草坪草、小麦、稻米、大麦、高粱、燕麦、橡树、桉树、胡桃和棕榈。因此,植物可以是单子叶植物或双子叶植物。如本文中所用,词语“植物”还涵盖植物细胞、种子、植物后代、繁殖体(无论以有性或无性方式产生),和其中任一种的后代,如插条或种子。植物细胞包括悬浮培养物、愈合组织(callus)、胚、分生组织区、愈合组织(callustissue)、叶、根、嫩枝、配子体、孢子体、花粉、种子和花粉粒。植物可以处于成熟期的各个阶段且可以在液体或固体培养物中或在罐、温室或旷野中的土壤或适合培养基中种植。植物中所引入的前导序列、尾序列或基因序列的表达可以是暂时性或永久性的。“所选植物物种”可以是(但不限于)这些“植物”中的任一种的物种。如本文中所用,术语“植物部分”(或马铃薯植物,或其一部分)包括(但不限于)原生质体、叶子、茎、根、根尖、花药、雌蕊、种子、胚、花粉、胚珠、子叶、下胚轴、花、块茎、芽眼、组织、叶柄、细胞、分生组织细胞和类似物。如本文中所用,术语“植物物种”是属于呈现至少一些性相容性的各种正式命名的植物物种的植物群组。如本文中所用,术语“植物转化”和“细胞培养”泛指借以对植物细胞进行遗传改造且转移到适当植物培养基中供维持、进一步生长和/或进一步研发的方法。如本文中所用,术语“精确育种”是指通过将核酸(如从所选植物物种中或从与所选植物相同的物种中的另一植物中或从与所选植物物种性相容的物种中分离出的原生基因和调节元件)稳定引入个别植物细胞中且随后使这些被遗传改造的植物细胞再生为完整植物来改良植物。由于不存在未知的或外来核酸永久性并入植物基因组中,因此本发明技术利用也可以通过传统植物育种获得的相同遗传物质。如本文中所用,术语“引物”是与所关注的核酸序列粘接的寡核苷酸。引物充当核酸合成的起点。dna聚合酶(一种催化此过程的酶)使新核苷酸相加到dna引物的3'端且拷贝反向股。举例来说,与所关注的dna序列互补的正向和反向引物在聚合酶链反应(pcr)分析中用于扩增所关注的dna区域。如本文中所用,术语“探针”是已用可检测分子(如放射性标记、生物素、洋地黄毒苷(digoxygenin)或荧光素)标记且与所关注的核酸序列互补的寡核苷酸。举例来说,在5'端经6-fam(6-羧基荧光素)标记且在3'端经bhq1(黑洞淬灭剂tm1)标记的探针在实时pcr中用于检测所关注的核酸序列。然而,术语探针还可以更广泛地用于指能够结合到非天然存在的核苷酸接合序列的核苷酸序列,不论探针上是否附接r标记。如本文中所用,术语“后代”包括两个马铃薯植物杂交所产生的f1马铃薯植物,且后代进一步包括(但不限于)与复现亲代品系杂交的后续f2、f3、f4、f5、f6、f7、f8、f9和f10代。如本文中所用,术语“定量性状基因座”(qtl)是指在一定程度上控制通常连续地分布的可用数值表示的性状的遗传基因座。如本文中所用,术语“重组”广泛地描述借以能够克隆基因、能够对dna定序且能够产生蛋白质产物的各种技术。如本文中所用,所述术语还描述在基因转移到植物宿主系统细胞中之后已经产生的蛋白质。如本文中所用,术语“再生”是指从组织培养物研发出植物。如本文中所用,术语“调节序列”是指所属领域的技术人员已知的那些标准序列,其可以包含于表达载体中以在植物系统中增强和/或最大化所关注基因的转录或所得rna的转译。这些序列包括(但不限于)启动子、肽输出信号序列、内含子、聚腺苷酸化和转录终止位点。修饰核酸构筑体以增加植物中的表达量的方法在所属领域中通常也已为人知(参见例如罗格(rogers)等人,260《生物化学杂志(j.biol.chem.)》3731-38,1985;科尔内霍(cornejo)等人,23《植物分子生物学(plantmol.biol.)》567:81,1993)。在工程改造植物系统以影响蛋白质转录速率时,所属领域中已知的多种因素(包括调节序列(如正或负作用序列)、增强子和静默子以及染色质结构)可以产生影响。本发明规定这些因素中的至少一个可以用于工程改造植物以表达所关注的蛋白质。本发明的调节序列是原生基因元件,即从待改造的所选植物物种中分离出来的。如本文中所用,术语“可选标记”典型地是一种编码蛋白质的基因,其将一些种类的抗性赋予抗生素、除草剂或毒性化合物,且用于鉴别转化事件。可选标记的实例包括编码链霉素(streptomycin)抗性的链霉素磷酸转移酶(spt)基因、将甘露糖-6-磷酸酯转化成果糖-6-磷酸酯的磷酸甘露糖异构酶(pmi)基因;编码卡那霉素(kanamycin)和遗传霉素(geneticin)抗性的新霉素(neomycin)磷酸转移酶(nptii)基因、编码潮霉素(hygromycin)抗性的潮霉素磷酸转移酶(hpt或aphiv)基因、编码磺酰脲型除草剂抗性的乙酰乳酸合成酶(als)基因、编码用于抑制谷氨酰胺合成酶作用的除草剂(如草铵膦(phosphinothricin)或巴斯特(basta)抗性的基因(例如bar基因)),或所属领域中已知的其它类似基因。如本文中所用,术语“有义抑制”是内源基因表达的减少,这是通过在转基因植物中表达那种基因的全部或一部分的一或多个额外拷贝来实现。如本文中所用,术语“比重”是密度的表达并且是马铃薯品质的度量。块茎比重和淀粉含量与干物质或总固体百分比之间存在高度相关性。比重越高,则回收率越高且所加工产品的品质越好。本文中所用的术语“严格条件”是指借以形成特定杂种、而不形成或极不可能形成非特定杂种的条件。举例来说,严格条件可以是与非天然存在的核苷酸接合点dna具有高度同源性(90%或更大,或95%或更大)的dna(例如探针)(例如序列与表6中的序列具有高度同源性的探针序列)借以与所述序列杂交的条件。严格条件可以指借以在比完全杂种的解链温度(tm)低约5℃到约30℃(在某些方面中为约10℃到约25℃)的温度下发生杂交的条件。举例来说,j.萨布鲁克(j.sambrook)等人(《实验室手册·分子选殖(molecularcloning,alaboratorymanual)》第二版,冷泉港实验室出版社(coldspringharborlaboratorypress)(1989))所述的条件(具体地说,§11.45“寡核苷酸探针杂交条件(conditionsforhybridizationofoligonucleotideprobes)”中所述的条件)可以用作严格条件。因此,两个核酸序列基本上同源表示两个分子在严格条件下彼此杂交。严格条件与序列相关且在不同环境参数下是不同的。一般来说,在限定的离子强度和ph值下,所选的严格条件比特定序列的热解链点(tm)低约5℃到20℃。tm定义为所指定dna序列的全部分子中50%杂交形成双股且50%以单股形式存在时的温度(摄氏度)。如本文中所用,术语“t-dna样”序列是从所选植物物种中或从与所选植物物种性相容的植物中分离出的核酸序列,且所述核酸序列与农杆菌种t-dna具有至少75%、80%、85%、90%或95%、而非100%的序列一致性。t-dna样序列可以含有一或多个边界或边界样序列,其各自能够将核苷酸序列整合到另一个聚核苷酸中。如本文中所用,术语“总产量”是指所收获的全部块茎的总重量。如本文中所用,术语“尾序列”是指位于基因之后(或基因的3')的被转录、但未被转译的序列。如本文中所用,术语“转录的dna”是包含基因与未转译的前导序列和尾序列的dna,所述前导序列和尾序列均与那个基因结合。基因是作为单一mrna通过前面启动子的作用转录。如本文中所用,术语“植物细胞的转化”是一种借以将dna稳定整合到植物细胞基因组中的方法。“稳定”是指聚核苷酸永久性或非暂时性滞留和/或表达于细胞基因组中和被细胞基因组表达。因此,稳定整合的聚核苷酸是所转化的细胞基因组内的固定物并且可以通过细胞或所得转化植物的连续后代来复制和繁殖。转化可以在天然或人工条件下使用所属领域中众所周知的多种方法进行。转化可以凭借用于将核酸序列插入原核或真核宿主细胞中的任何已知方法,包括农杆菌介导的转化方案、病毒感染、晶须、电致孔、热休克、脂质体转染、聚乙二醇处理、微注射和粒子轰击。如本文中所用,术语“转基因”是将插入宿主基因组中的基因。如本文中所用,术语“转基因植物”是含有至少一种转基因的经遗传改造的植物。如本文中所用,术语“块茎”是指一种类型的被改造植物结构,其经扩大以储存营养。植物利用其经受住冬季或旱月,以在接下来的生长季节期间向再生长提供能量和营养,且作为无性繁殖的手段。其可以来源于茎或根。马铃薯是块茎。如本文中所用,术语“变异体”应理解成意指偏离特定基因或蛋白质的标准或所指定核苷酸或氨基酸序列的核苷酸或氨基酸序列。术语“同功异型物”、“同型”和“类似物”也指核苷酸或氨基酸序列的“变异体”形式。通过添加、移出或取代一或多个氨基酸或改变核苷酸序列而改变的氨基酸序列可以视为“变异体”序列。变异体可以具有「保守」变化,其中被取代的氨基酸具有类似的结构或化学特性,例如用异白氨酸置换白氨酸。变异体可以具有“非保守”变化,例如用色氨酸置换甘氨酸。类似的较小变异还可以包括氨基酸缺失或插入或两者。确定哪个氨基酸残基可以被取代、插入或缺失的指南可以使用所属领域中众所周知的计算机程序(例如vectornti程序组(马里兰州因弗马斯公司(informax,md))软件)发现。如本文中所用,术语“藤本植物成熟度”是指植物继续使用碳水化合物和光合作用的能力。藤本植物成熟度按照1到5的等级评分,其中1=死亡的藤本植物且5=绿色、仍然开花的藤本植物。天生tm技术将理想的性状插入马铃薯植物的基因组中存在特定的困难,因为马铃薯是四倍体、高杂合的且对近交衰退敏感。因此,在使用传统育种进行处理期间,极难以有效地研发出转基因马铃薯植物,其产生较少的丙烯酰胺和不太有害的梅纳反应产物,包括n-亚硝基-n-(3-酮基-1,2-丁二醇)-3'-硝基酪胺酸(王(wang)等人,《毒理学文献(archtoxicol)》70:10-5,1995)、5-羟基甲基-2-糠醛(扬佐夫斯基(janzowski)等人,《食品与化学品毒理学(foodchemtoxicol)》38:801-9,2000)和具有突变诱发特性的其它梅纳反应产物(柴本(shibamoto),《临床与生物学研究进展(progclinbiolres)》304:359-76,1989)。已经测试若干种方法且正在进行研究以通过工艺变化、减少右旋糖以及添加剂(如天冬酰胺酶、柠檬酸盐和竞争氨基酸)来减少丙烯酰胺。整个马铃薯工业中实施工艺变化所需的资本费用将花费数百万美元。除费用以外,这些工艺变化具有明显的缺陷,包括与添加剂(如天冬酰胺酶或柠檬酸盐)有关的潜在负面风味。典型地,炸薯条制造商在法式炸薯条加工期间添加右旋糖以呈现所期望的金褐色,但右旋糖也通过梅纳反应而增加丙烯酰胺的形成。仅仅从工艺中省去右旋糖便使丙烯酰胺明显减少;然而,必须接着利用一些其它方式来呈现标志性的金褐色(如通过添加颜料,如胭脂树红(annatto))。替代颜料的使用导致通过那些褐变反应而呈现的典型风味缺失。使用添加剂减少反应物(如天冬酰胺)的另一项挑战是在冷冻储存期间发生的水分迁移,导致天冬酰胺返回表面和丙烯酰胺增加。最终,在马铃薯碰伤之后发生的变黑影响了法式炸薯条和薯片加工中的品质和回收。损伤或碰伤的马铃薯必须在加工之前修整或丢弃,导致品质难题或经济损失。天生tm技术的说明概述了在一起协作产生植物的植物生物学系统。这些包括性状鉴别、载体设计、载体并入农杆菌中、受体马铃薯变种选择、转化植物,和新马铃薯含有预期dna插入序列的确认。天生tm方法允许将非编码dna插入马铃薯中,以呈现具有所期望性状的新马铃薯事件。本发明的“原生技术”策略解决了马铃薯工业对改进马铃薯的农学特征和营养价值的需求,这是通过减少多酚氧化酶-5(ppo5)的表达(引起黑斑碰伤)、天冬酰胺合成酶-1(asn1)的表达(引起天冬酰胺(形成丙烯酰胺的前驱物)聚积)和/或磷酸化酶-l和二激酶-r1(其是与还原糖聚积有关的酶,所述还原糖通常与氨基酸(如天冬酰胺)反应且形成毒性梅纳产物,包括丙烯酰胺)的表达来实现。块茎中这些基因的部分或完全静默使得产生丙烯酰胺的潜在性降低。使用本发明的天生tm技术允许将所期望的性状并入有商业价值的马铃薯植物变种的基因组中,这是通过仅用包含非编码调节区的“原生”遗传物质(即从马铃薯植物或与马铃薯植物性相容的植物获得的遗传物质)转化马铃薯,而非将任何外来遗传物质整合到植物基因组中。所期望的性状包括高度耐受冲击诱发黑斑碰伤、减少丙烯酰胺前驱物天冬酰胺的形成和减少还原糖的聚积,随之而来的是毒性梅纳产物(包括丙烯酰胺)的聚积减少、品质改善和食物颜色控制。这些期望性状并入现有马铃薯变种中不可能通过传统育种实现,因为马铃薯是四倍体、高杂合并且对近交衰退敏感。本发明中使用的非编码马铃薯植物dna插入序列是马铃薯植物基因组原生的且不含任何农杆菌dna。dna插入序列优选包含两个表达盒且插入称为psim1278转化载体的转化载体中(描述于美国专利第8,754,303号,“马铃薯栽培品种j3”;美国专利第8,710,311号,“马铃薯栽培品种f10”;美国专利第8,889,963号,“马铃薯栽培品种j55”;和美国专利申请第14/072,487号,“马铃薯栽培品种e12”;和美国专利第8,889,964号,“马铃薯栽培品种w8”,其还具有psim1678载体,这些专利和申请各自全文结合在此供参考)。第一个盒包含天冬酰胺合成酶-1基因(asn1)和多酚氧化酶-5基因(ppo5)的片段,其以反向重复序列形式配置于adp葡萄糖焦磷酸化酶基因(agp)的agp启动子与颗粒结合的淀粉合成酶基因(gbss)的gbss启动子之间。这些启动子主要在块茎中发挥活性。第二个盒的功能是使淀粉相关基因二激酶-r1(r1)和磷酸化酶-l基因(phl)的启动子静默。这个盒包含淀粉相关基因二激酶-r1(r1)和磷酸化酶-l基因(phl)的启动子的片段,其可操作地连接到与第一个盒相同的agp和gbss启动子。这些表达盒不含外来dna,并且仅由来自所选植物物种或与所选植物物种性相容的植物的dna组成。第二dna插入序列来自称为psim1678的转化载体(描述于美国专利第8,889,964号中,“马铃薯栽培品种w8”,所述专利全文结合在此供参考),所述载体包含rpi-vnt1表达盒和植物液泡转化酶基因vinv的静默盒。rpi-vnt1基因盒由vnt1蛋白质编码区组成,所述编码区由其原生启动子和终止序列调节以赋予针对晚疫病的广泛抗性,而静默盒由来自马铃薯vinv基因的反向重复序列、侧接的反向植物启动子pgbss和pagp组成。第一个盒的功能是赋予针对晚疫病的抗性,而第二个盒的功能是使液泡转化酶基因静默,从而减少葡萄糖和果糖。用原生dna进行的靶向基因静默降低了靶向基因的rna转录物在马铃薯块茎事件中的含量。一般来说,所插入的dna含有静默盒,所述静默盒当表达时产生了大小可变且未加工的转录物。这些转录物触发了通常编码酶(如天冬酰胺合成酶)的mrna的降解。由此使得靶向“静默”的酶含量大大降低。在不进一步抑制淀粉相关基因激酶-r1(r1)和磷酸化酶-l(phl)的情况下,asn1和ppo5基因静默足以使丙烯酰胺形成明显减少两倍到四倍。因此,马铃薯块茎事件并有高度期望的性状,包括游离酰胺氨基酸天冬酰胺与谷氨酰胺的比率降低,其与油炸或烘烤后的丙烯酰胺形成减少有关。具体地说,本发明的马铃薯变种的特征是游离天冬酰胺含量降低两倍到超过四倍。此外,本发明的马铃薯变种显示淀粉在储存期间延迟降解成还原糖葡萄糖和果糖。阻碍淀粉转化成糖进一步减少了衰老甜化和丙烯酰胺形成且限制了热诱发褐变。另外,事件w8、x17和y9也展示出存在于事件j3、f10、j55和e12中的针对晚疫病的抗性,这是除psim1278载体之外,额外使用psim1678载体的结果。因此,本发明的马铃薯变种在马铃薯工业和食品市场中极有价值,因为其块茎在热加工后产生的丙烯酰胺明显更少且不携带任何潜在有害的外来基因。产生具有上文提及的有利特征的组合的马铃薯变种的研究在很大程度上是凭经验的。这种研究需要投入大量时间、劳动力和金钱。从温室到商业用途,马铃薯栽培品种的研发往往可以耗费长达八年或更长时间。育种从谨慎选择优良亲代开始,以将最重要的特征并入后代。由于全部所期望的性状通常不会在仅一次杂交后呈现,因此育种必须是累积的。现有育种技术继续使用亲代克隆的可控授粉。花粉典型地收集于明胶胶囊中以便后来用于对雌性亲代授粉。在温室中播种杂交种子并且从数千个个别幼苗收获和保留块茎。次年,在田间种植来自每个所得幼苗的一到四个块茎,其中密切注意避免病毒和疾病的传播。从这种第一年幼苗作物开始,保留在选择过程中存活的来自每个杂交个体的若干“种子”块茎用于次年的种植。在第二年之后,取得样品用于密度测量和油炸测试,以便确定块茎用于商业用途的合适性。历经选择过程到此时仍存活的植物接着在第三年按扩大的容量种植,用于一系列更全面的油炸测试和密度测定。在第四年研发期,存活选择在若干状态下经历田间试验以测定其对不同生长条件的适应性。最终,将具有优良品质的变种转移到其它农场且将种子增加到商业规模。一般来说,到这时,已经投入八年或更长时间的种植、收获和测试以尝试研发出新颖的和改良的马铃薯栽培品种。随着允许对编码特定蛋白质产物的基因进行分离和表征的分子生物技术的出现,植物生物学领域的科学家对工程改造植物基因组产生了强烈的兴趣,以含有和表达外来基因或原生或内源基因的其它或经修饰形式(可能由不同启动子驱动),从而以特定方式改变植物性状。这类外来的其它和/或经修饰基因在本文中统称为“转基因”。在最近十五到二十年,已研发出若干种用于产生转基因植物的方法,并且本发明在特定实施例中还涉及了所要求的变种或品系的转化形式。植物转化涉及建构在植物细胞中发挥功能的表达载体。此类载体包含含有受调节元件(例如启动子)控制或可操作地连接到调节元件(例如启动子)的基因的dna。表达载体可以含有一或多个此类可操作地连接的基因/调节元件组合。载体可以呈质体形式且可以单独或与其它质体组合使用,以便使用如下文所述的将转基因并入马铃薯植物遗传物质中的转化方法提供被转化的马铃薯植物。传统植物育种典型地依赖于植物染色体的随机重组,以产生具有新颖和改良特征的变种。根据标准、众所周知的技术,将包含基因和调节元件的基因“表达盒”插入农杆菌分离的转移dna(“t-dna”)的边界内并且整合到植物基因组中。农杆菌介导的t-dna物质转移典型地包含以下标准程序:(1)基因元件的活体外重组,其中至少一个基因元件具有外来来源,以便产生表达盒用于转化选择;(2)将这个表达盒(往往连同至少一个含有外来dna的其它表达盒)一起插入二元载体的t-dna区域中,所述t-dna区域通常由侧接t-dna边界序列的农杆菌dna的数百个碱基对组成;(3)将位于t-dna边界之间的序列(往往伴随来自农杆菌的一些或全部其它二元载体序列)转移到植物细胞中;和(4)选择经稳定转化的植物细胞,所述植物细胞呈现所期望的性状,如产量增加、活力改良、对疾病和昆虫的抗性增强或在压力下存活的能力更大。因此,遗传工程方法可以依赖于外来的非内源性核酸的引入,所述核酸包括调节元件,如启动子和终止子,以及涉及新性状表达或充当标记物用于鉴别和选择来自病毒、细菌和植物的转化体的基因。标记基因典型地来源于细菌来源并且赋予抗生素或除草剂抗性。经典育种方法是费力且耗时的,并且新颖的变种典型地仅呈现相对适度的改良。在“反义”技术中,颠倒原生基因的序列以静默转基因植物对基因的表达。马铃薯转化用的表达载体:标记基因表达载体包括至少一种可操作地连接到调节元件(例如启动子)的基因标记物,其允许通过负向选择(即,抑制不含可选标记基因的细胞生长)或正向选择(即,筛选由基因标记物编码的产物)回收含有所述标记物的已转化细胞。植物转化用的许多常用可选标记基因在转型技术中众所周知,并且包括例如编码以代谢方式使选择性化学试剂(其可以是抗生素或除草剂)解毒的酶的基因,或编码对抑制剂不敏感的变异目标的基因。所属领域中还知悉几种正向选择方法。植物转化用的一种常用可选标记基因是新霉素磷酸转移酶ii(nptii)基因,其在植物调节信号的控制下赋予卡那霉素抗性。弗雷利(fraley)等人,《美国国家科学院院刊(proc.natl.acad.sci.u.s.a.)》,80:4803(1983)。另一种常用的可选标记基因是潮霉素磷酸转移酶基因,其赋予抗生素潮霉素抗性。王登艾尔争(vandenelzen)等人,《植物分子生物学(plantmol.biol.)》,5:299(1985)。赋予抗生素抗性的细菌来源的其它可选标记基因包括健大霉素(gentamycin)乙酰基转移酶、链霉素磷酸转移酶和氨基糖苷-3'-腺苷转移酶(博莱霉素(bleomycin)抗性决定子)。海福德(hayford)等人,《植物生理学(plantphysiol.)》86:1216(1988);琼斯(jones)等人,《分子遗传学与普通遗传学(mol.gen.genet.)》,210:86(1987);斯瓦伯(svab)等人,《植物分子生物学》14:197(1990);希尔(hille)等人,《植物分子生物学》7:171(1986)。其它可选标记基因赋予针对除草剂(如草甘膦(glyphosate)、草铵膦(glufosinate)或溴苯腈(bromoxynil))的抗性。科麦(comai)等人,《自然(nature)》317:741-744(1985);戈登-卡姆(gordon-kamm)等人,《植物细胞(plantcell)》2:603-618(1990)和斯德克(stalker)等人,《科学(science)》242:419-423(1988)。非细菌来源的用于植物转化的可选标记基因包括例如小鼠二氢叶酸还原酶、植物5-烯醇丙酮酰莽草酸-3-磷酸合成酶和植物乙酰乳酸合成酶。艾科茨(eichholtz)等人,《体细胞分子遗传学(somaticcellmol.genet.)》13:67(1987);沙阿(shah)等人,《科学》233:478(1986);查尔斯特(charest)等人,《植物细胞报告(plantcellrep.)》8:643(1990)。另一类用于植物转化的标记物基因需要筛选假设转化的植物细胞,而非对具有针对毒性物质(如抗生素)的抗性的转化细胞进行直接基因选择。这些基因特别适用于对特定组织中的基因表达空间模式进行定量或可视化且通常称为报告基因,因为其可以与基因或基因调节序列融合以便研究基因表达。用于筛选假设转化细胞的常用基因包括β-葡糖醛酸酶(gus)、β-半乳糖苷酶、荧光素酶和氯霉素乙酰转移酶。杰佛森r.a.(jefferson,r.a.),《植物分子生物学报告(plantmol.biol.rep.)》5:387(1987);泰里(teeri)等人,《欧洲分子生物学杂志(emboj.)》8:343(1989);孔茨(koncz)等人,《美国国家科学院院刊》84:131(1987);德布劳克(deblock)等人,《欧洲分子生物学杂志》3:1681(1984)。用于目测gus活性的活体内方法无需破坏可利用的植物组织。分子探针公司(molecularprobes)公告2908,imagenegreen,第1-4页(1993)和奈尔维(naleway)等人,《细胞生物学杂志(j.cellbiol.)》115:151a(1991)。然而,由于灵敏度低、荧光背景高和与使用荧光素酶基因作为可选标记物相关的局限性,因此用于目测gus活性的这些活体内方法尚未证实适用于回收所转化的细胞。在一些方面中,已经使用编码绿色荧光蛋白(gfp)的基因作为原核和真核细胞中的基因表达标记物。查尔菲(chalfie)等人,《科学》263:802(1994)。gfp和gfp突变体可以作为可筛选的标记物使用。马铃薯转化用的表达载体:启动子表达载体中包括的基因必须由包含调节元件(例如启动子)的核苷酸序列驱动。若干类型的启动子以及可以单独或与启动子组合使用的其它调节元件在转化技术中是众所周知的。如本文中所用,“启动子”包括提及转录起点上游的起始转录的dna区域,其涉及rna聚合酶和其它蛋白质的识别和结合。“植物启动子”是能够在植物细胞中起始转录的启动子。在发育控制下的启动子的实例包括优先在某些组织(如叶子、根、种子、纤维、木质部导管、管胞或厚壁组织)中起始转录的启动子。这类启动子称为“组织优先”。仅在某一组织中起始转录的启动子称为“组织特异性”。“细胞类型”特异性启动子主要驱动一或多种器官中某些细胞类型(例如根或叶中的维管细胞)中的表达。“诱导型”启动子是受环境控制的启动子。可以通过诱导型启动子实现转录的环境条件的实例包括厌氧条件或存在光。组织特异性、组织优先、细胞类型特异性和诱导型启动子构成“非组成性”启动子类别。“组成性”启动子是在大部分环境条件下具有活性的启动子。a.诱导型启动子诱导型启动子可操作地连接到用于在马铃薯中表达的基因。任选地,诱导型启动子可操作地连接到编码信号序列的核苷酸序列,所述信号序列可操作地连接到用于在马铃薯中表达的基因。在诱导型启动子情况下,转录速率回应于诱导剂而增加。本发明可以使用任何诱导型启动子。参见沃德(ward)等人,《植物分子生物学》22:361-366(1993)。例示性诱导型启动子包括(但不限于)来自acei系统的对铜有反应的启动子(米特(mett)等人,《美国国家科学院院刊》90:4567-4571(1993));来自玉米的对苯磺酰胺除草剂安全剂有反应的in2基因(赫胥(hershey)等人,《分子基因遗传学(mol.gengenetics)》227:229-237(1991)和加茨(gatz)等人,《分子基因遗传学》243:32-38(1994))或来自tn10的tet抑制因子(加茨等人,《分子基因遗传学》227:229-237(1991))。特别优选的诱导型启动子是对诱导剂有反应的启动子,而植物对所述诱导剂通常无反应。示例性诱导型启动子是来自类固醇激素基因的诱导型启动子,其转录活性是由糖皮类固醇激素诱导。斯赫纳(schena)等人,《美国国家科学院院刊》88:0421(1991)。b.组成型启动子组成型启动子可操作地连接到用于在马铃薯中表达的基因或组成型启动子可操作地连接到编码信号序列的核苷酸序列,所述信号序列可操作地连接到用于在马铃薯中表达的基因。本发明中可以使用许多不同的组成型启动子。示例性组成型启动子包括(但不限于)来自植物病毒的启动子,如来自camv的35s启动子(奥德尔(odell)等人,《自然》313:810-812(1985))和来自如稻米肌动蛋白的基因的启动子(麦克尔罗伊(mcelroy)等人,《植物细胞(plantcell)》2:163-171(1990));泛素(克里斯滕森(christensen)等人,《植物分子生物学》12:619-632(1989)和克里斯滕森等人,《植物分子生物学》18:675-689(1992));pemu(拉斯特(last)等人,《理论应用遗传学(theor.appl.genet.)》81:581-588(1991));mas(费尔滕(velten)等人,《欧洲分子生物学杂志(emboj.)》3:2723-2730(1984))和玉米h3组蛋白(勒珀蒂(lepetit)等人,《分子基因遗传学(mol.gen.genetics)》231:276-285(1992)和安塔索瓦(atanassova)等人,《植物杂志(plantjournal)》2(3):291-300(1992))。als启动子,甘蓝型油菜(brassicanapus)als3结构基因5'的xba1/ncol片段(或与所述xba1/ncol片段相似的核苷酸序列),代表特别适用的组成型启动子。参见pct申请wo96/30530。c.组织特异性或组织优先型启动子组织特异性启动子可操作地连接到用于在马铃薯中表达的基因。任选地,组织特异性启动子可操作地连接到编码信号序列的核苷酸序列,所述信号序列可操作地连接到用于在马铃薯中表达的基因。被可操作地连接到组织特异性启动子的所关注基因转化的植物独占地或优先地在特定组织中产生转基因的蛋白质产物。本发明中可以使用任何组织特异性或组织优先型启动子。示例性组织特异性或组织优先型启动子包括(但不限于)根优先型启动子,如来自菜豆蛋白基因的启动子(穆雷(murai)等人,《科学》23:476-482(1983)和森古普塔-格帕兰(sengupta-gopalan)等人,《美国国家科学院院刊》82:3320-3324(1985));叶子特异性和光诱导启动子,如来自叶绿素a/b结合蛋白(cab)或核酮糖二磷酸羧化酶(rubisco)的启动子(辛普森(simpson)等人,《欧洲分子生物学杂志》4(11):2723-2729(1985)和蒂姆科(timko)等人,《自然》318:579-582(1985));花药特异性启动子,如来自lat52的启动子(特维尔(twell)等人,《分子基因遗传学》217:240-245(1989));花粉特异性启动子,如来自zm13的启动子(格雷罗(guerrero)等人,《分子基因遗传学》244:161-168(1993))或小孢子优先型启动子,如来自apg的启动子(特维尔(twell)等人,《有性植物繁殖(sex.plantreprod.)》6:217-224(1993))。用于使蛋白质靶向亚细胞隔室的信号序列借助于使编码信号序列的核苷酸序列可操作地连接到编码所关注蛋白质的基因的5'和/或3'区域来将通过转基因产生的蛋白质输送到亚细胞隔室(如叶绿体、液泡、过氧化体、乙醛酸循环体、细胞壁或线粒体)或分泌到非原质体(apoplast)中。结构基因5'和/或3'端的靶向序列可以在蛋白质合成和加工期间决定所编码蛋白质最终在何处隔室化。信号序列的存在将多肽引导到细胞内的细胞器或亚细胞隔室中或分泌到非原质体中。许多信号序列是所属领域中已知的。参见例如贝克尔(becker)等人,《植物分子生物学》20:49(1992);克罗斯p.s.(close,p.s.),《硕士论文(master'sthesis)》,爱荷华州立大学(iowastateuniversity)(1993);克诺克斯c.(knox,c.)等人,《植物分子生物学》9:3-17(1987);莱内尔(lerner)等人,《植物生理学(plantphysiol.)》91:124-129(1989);方特斯(frontes)等人,《植物细胞(plantcell)》3:483-496(1991);松冈(matsuoka)等人,《美国国家科学院院刊》88:834(1991);古尔德(gould)等人,《细胞生物学杂志(j.cell.biol.)》108:1657(1989);克里森(creissen)等人,《植物杂志(plantj.)》2:129(1991);卡尔德隆(kalderon)等人,《细胞(cell)》39:499-509(1984);斯特非尔(steifel)等人,《植物细胞》2:785-793(1990)。茄属的分类学茄科含有众所周知的若干栽培农作物,如西红柿(番茄(solanumlycopersicum),也称为番茄(lycopersiconesculentum))、茄子(solanummelogena)、烟草(nicotianatabacum)、胡椒(capsicumannuum)和马铃薯(solanumtuberosum)。在茄属内,已经识别出逾千种物种。马铃薯与不具有块茎的茄属(西红柿、茄子等)物种(包括商业马铃薯田中和周围通常发现的野草)不杂交(洛夫(love)1994)。茄属分成若干子目,其中马铃薯子目含有具有块茎的所有马铃薯。马铃薯子目按系列划分,其中块茎与本文相关。在块茎序列内发现大约54个野生和栽培马铃薯物种。其中之一是马铃薯。马铃薯分成两个亚种:块茎种(tuberosum)和安第斯种(andigena)。亚种块茎种是在例如北美洲和欧洲中广泛用作农作物植物的栽培马铃薯。亚种安第斯种也是栽培物种,但是栽培限于中美洲和南美洲(汉莱曼(hanneman)1994)。美国的野生马铃薯在美国边界内种植且其试样存在于基因库中的仅两种野生马铃薯种包括四倍体物种芬德瑞茄(s.fendleri)(最近作为匐枝茄(s.stoloniferum)重新分类);然而,一些来源(包括基因库间马铃薯数据库(inter-genebankpotatodatabase))仍然使用芬德瑞茄名称);和二倍体物种詹姆斯茄(s.jamesii)(班伯格(bamberg)等人,2003;ipd2011;班伯格和德尔里奥(delrio)2011a;班伯格和德尔里奥,2011b;斯普勒(spooner)等人,2004)。洛夫(love)(1994)报道了第三物种裂叶茄(s.pinnatisectum)也是美国的一种原生物种。然而,斯普勒等人(2004)确定了先前被认为是裂叶茄的实际上是詹姆斯茄。通过超过10年的田间工作和现有记录的评估,班伯格等人(2003)和斯普勒等人(2004)确认了美国仅存在这两种物种:芬德瑞茄和詹姆斯茄。这些研究人员还尝试检验先前记录的地点,且通过此方法更新了这些物种的当前已知地点的地图,从而向每种记录的群体提供经度和纬度位置(班伯格等人,2003)和分布图(斯普勒等人,2004)。这些物种主要存在于干燥森林、沙漠灌木丛,和海拔高度5,000到10,000英尺的多沙地区中,与大多数商业生产地区良好隔离(班伯格和德尔里奥,2011a)。虽然用于商业生产的面积与野生物种在县级的出现率之间存在一些重叠,但美国大部分的马铃薯生产不发生于野生马铃薯地区。然而,存在着几种野生马铃薯植物可能在马铃薯田地附近生长的可能性(洛夫,1994)。斯普勒等人(2004)描述詹姆斯茄在美国栖息于山坡上的大石头之间、砂质冲积流底部、沿着小径或道路的砾石中、冲积山谷的富有机质土壤、砂质闲田、草原、灌木丛沙漠上的杜松、橡树丛、海拔在4,500到9,400英尺之间的针叶和落叶森林。其类似地描述了海拔在4700到11,200英尺之间的芬德瑞茄栖息地。马铃薯遗传学茄属中的基本染色体数目是十二。马铃薯亚种块茎种可以二倍体(2n=2x=24)或四倍体(2n=4x=48)。二倍体的范围限于南美洲部分,而四倍体最广泛地栽培于全世界。四倍体起源于马铃薯中的方式不清楚。栽培的马铃薯亚种块茎种可以是同源四倍体(二倍体物种中的染色体倍增)或异源四倍体(两个相关物种之间的二倍体杂种的染色体倍增)。虽然几乎所有的二倍体物种是自我不相容的,但是栽培的四倍体马铃薯亚种块茎种能够自我授粉(自花授粉)。普雷斯特公司(plaisted)(1980)已经表明在田间条件下,四倍体马铃薯最可能发生自花授粉,其中80-100%的种子由于自花授粉而形成。康纳(conner)和黛尔(dale)(1996)从若干个田间实验收集了远交数据,所述实验是在新西兰、英国和瑞典使用被遗传改造的马铃薯进行。在各项研究中,当接收植物与被遗传改造的植物相隔超过20公尺时,远交率是零。虽然多种茄种是可育的,但是似乎大量的栽培四倍体马铃薯亚种块茎种栽培品种的可育性降低。马铃薯变种花费多年研发出了马铃薯变种。创立新变种的决定是基于许多因素,如市场需求、潜在的消费者认可,和害虫耐受性或抗性。相较于一些其它农作物(如田地玉米或大豆),马铃薯变种的引入和中断频率不高。由于马铃薯被克隆繁殖,因此杂交授粉所致的变种稀释的风险降低。本发明所用的马铃薯事件来源于四个马铃薯变种。褐色布尔班克是事件e12和w8的亲代变种。卢瑟·布尔班克(lutherburbank)在十九世纪七十年代早期研发出了此变种。植物在整个季节茁壮地且持续地按藤本植物方式生长。茎较厚,有明显的倾角并且有细斑。小叶的宽度是长到中等并且颜色是浅绿色到中等绿色。花朵较少、呈白色并且不可育。栽培品种耐受常见的疮痂病,但对种枯萎病和黄萎病、卷叶和净坏死和病毒y敏感。植物需要高度且均一的土壤水分和可控的氮肥条件以产生不含节、尖端和哑铃状物的块茎。当植物经历压力时,块茎中产生胶状端和糖端。所产生的块茎有较大褐皮和白色肉质,显示良好的长期储存特征,并且代表极佳烘烤和加工品质的标准。所述变种是不育的并且在西北部和中西部广泛种植,特别是用于生产法式炸薯条。褐色兰吉尔是事件f10和x17的亲代变种。这种全季变种发行于1991年。褐色兰吉尔对黄萎病、病毒x和y、卷叶和净坏死以及镰孢干腐病的抗性强于褐色布尔班克。其高度耐受空心病。植物大且直立展开。茎厚,呈绿色,其在全日照下可以是浅褐色到浅紫色。叶子宽大且呈中等绿色。大量开花且产生有活力的花粉。芽呈绿色,基部和花梗是淡红色-紫色,有中等量的短柔毛。花冠呈中等偏大,红色-紫色且花粉囊呈亮黄色。其产生了品质良好、比重高、产量高的块茎,所述块茎长且稍微扁平,且较适合烘烤和加工成法式炸薯条。块茎对常见的疮痂病和黑斑碰伤敏感。褐色兰吉尔成熟早于褐色布尔班克且被视为中长期储存变种。所述变种是可育的并且主要在西北部种植,特别是用于生产法式炸薯条。大西洋是事件j3和j55和y9的亲代变种。植物中等偏大,茎厚且笔直,且稍微膨胀,有软毛的结节稀少。叶子呈中等亮绿色,平滑且有中等柔毛,突出的翼,有不对称的较大初代小叶和许多次生和三次小叶。花富有绿色、锥状、柔毛萼瓣,浅紫色花冠,橙色花粉囊和充裕的有活力的花粉。栽培品种耐受疮痂病和黄萎病,耐受红眼,高度耐受a种金线虫、病毒x、块茎净坏死,且针对黑斑碰伤展示一些抗性。块茎对内部热坏死敏感,特别是在砂质土壤中、在温热的干燥季节。在一些种植地区,直径较大的块茎(直径>4英寸)可能具有严重的的空心病。块茎呈椭圆形到圆形,具有浅到深的鳞片状网状皮,中等浅芽眼和白色果肉。块茎冬眠是中长期的。潜在产量高、比重大且块茎尺寸和形状均一的大西洋是来自田间或来自极短期储存的用于油炸薯片的标准变种(韦布(webb)等人,1978)。所述变种是可育的并且主要种植于东北部和东南部,特别是用于生产薯片。斯诺登是事件v11的亲代变种。斯诺登(w855)是在二十世纪七十年代晚期、在威斯康星州(wisconsin)选自wischip与b5141-6之间的杂交且在1990年命名。选择和早期测试是由斯坦·皮诺昆(stanpeloquin)博士和唐纳德·奇彻夫斯基先生(mr.donaldkichefski)在威斯康星州的拉兰德的uw-lelah斯达克马铃薯育种农场(uw-lelahstarkspotatobreedingfarm,rhinelander,wi)进行。藤本植物大、直且适中。叶子呈浅绿色且闭合。花呈白色,具有黄色花粉囊且倾向于异常中止。雄性不育是常见的且很少出现果实。芽眼适中,在顶端较深且均一地分布。块茎具有白色果肉,而皮呈浅褐色、轻微网状。块茎呈均一、圆形和稍微扁平状,直径始终为2.5到3.5英寸。其对早疫病和晚疫病和常见疮痂病敏感且对于科罗拉多(colorado)马铃薯甲虫具有吸引力。所述变种耐受空心病和中心褐变。其产量稍微小于或大约等同于大西洋。其现在在北方中心地区试验中是标准物。这非常像大西洋,但其在不进行重新调节的情况下在45℉储存后碎裂。其用于生产薯片。马铃薯转化方法已经研发用于植物转化的许多方法,包括生物学和物理学植物转化方案。参见例如三木等人,“用于将外来dna引入植物中的程序(proceduresforintroducingforeigndnaintoplants)”,于《植物分子生物学和生物技术中的方法(methodsinplantmolecularbiologyandbiotechnology)》中,格里克b.r.(glick,b.r.)和汤普森j.e.(thompson,j.e.)编(crc出版公司(crcpress,inc.),波卡拉顿(bocaraton),1993),第67-88页。此外,可利用表达载体和活体外培养方法进行植物细胞或组织的转化和植物的再生。参见例如格鲁伯(gruber)等人,“用于植物转化的载体(vectorsforplanttransformation)”,于《植物分子生物学和生物技术中的方法》中,格里克b.r.和汤普森j.e.编(crc出版公司,波卡拉顿,1993),第89-119页。a.农杆菌介导的转化一种将表达载体引入植物中的方法是基于农杆菌的天然转化系统。参见例如霍施(horsch)等人,《科学》227:1229(1985)。根癌农杆菌(a.tumefaciens)和发根农杆菌(a.rhizogenes)是植物病原性土壤细菌,其以遗传方式转化植物细胞。根癌农杆菌和发根农杆菌的ti和ri质体分别携带负责遗传转化植物的基因。参见例如卡多c.i.(kado,c.i.),《植物科学重要评论(crit.rev.plantsci.)》10:1(1991)。用于农杆菌介导式基因转移的农杆菌载体系统和方法的说明由格鲁伯等人(同上)、三木等人(同上)和莫洛尼(moloney)等人,《植物细胞报告(plantcellreports)》8:238(1989)提供。还参见美国专利第5,563,055号(唐森德(townsend)和托马斯(thomas)),1996年10月8日颁布。关于农杆菌介导式转化和特定的dna递送质体(为用于农杆菌而专门设计)的专利有许多,例如us4536475、ep0265556、ep0270822、wo8504899、wo8603516、us5591616、ep0604662、ep0672752、wo8603776、wo9209696、wo9419930、wo9967357、us4399216、wo8303259、us5731179、ep068730、wo9516031、us5693512、us6051757和ep904362a1,这些专利皆全文结合在此供参考。农杆菌介导式植物转化包括将克隆于质体上的dna片段放置于活的农杆菌细胞内作为第一步骤,随后接着用于转化成个别植物细胞。农杆菌介导式植物转化因此是间接的植物转化方法。农杆菌介导式转化是通过使用属于农杆菌属的基因工程改造土壤细菌来实现。若干农杆菌种介导了可以经基因工程改造以携带任何期望的dna片断的特定dna(称为“t-dna”)转移到许多植物物种中。t-dna介导式发病过程中标志的主要事件是:毒性基因的诱导、t-dna的加工和转移。这种方法是许多评论的主题(莱恩(ream),1989;霍华德(howard)和西托斯基(citovsky),1990;卡多(kado),1991;胡卡斯(hooykaas)和斯奇普特(schilperoort),1992;温兰斯(winnans),1992;詹布斯基(zambryski),1992;格尔文(gelvin),1993;宾斯(binns)和霍维茨(howitz),1994;胡卡斯(hooykaas)和贝焦波庚(beijersbergen),1994;莱索(lessl)和南卡(lanka),1994;朱潘(zupan)和詹布斯基(zambryski),1995)。农杆菌介导植物发生遗传转化包括若干个步骤。第一步骤通常称为“接种”,其中首先使农杆菌与植物细胞彼此接触。接种步骤之后,通常使农杆菌和植物细胞/组织一起在适于生长和t-dna转移的条件下生长数小时到数日。这个步骤称为“共培养”。共培养和t-dna递送之后,植物细胞往往用杀菌剂和/或抑菌剂处理以杀死农杆菌。若这是在缺乏任何选择剂的情况下进行以促进转基因植物细胞相对于非转基因植物细胞的优先生长,则此典型地称为“延时”步骤。若在选择性压力有利的转基因植物细胞存在下进行,则其称为“选择”步骤。使用“延时”时,其后为一或多个“选择”步骤。“延时”与“选择”步骤典型地包括杀菌剂和/或抑菌剂以杀死任何残余的农杆菌细胞,因为在感染(接种和共培养)过程之后不期望农杆菌细胞生长。虽然通过农杆菌介导式转化所产生的转基因植物通常含有简单的整合模式(相较于微粒介导式遗传转化),但是拷贝数和插入模式存在广泛的变化(琼斯(jones),1987;乔根森(jorgensen),1987)。此外,即使在单一植物基因型内,也可以基于外植体的类型和所用的转化系统存在不同的t-dna整合模式(格莱维丁(grevelding)等人,1993)。调节t-dna拷贝数的因素尚不明确。特定的天生tm转化方法将阐述于随附实例中。然而,前述针对植物的“通用”农杆菌介导式植物转化方法还产生了非天然存在的核苷酸接合序列,其可以通过所教导的方法检测。b.直接基因转移使用dna的植物直接转化方法也已经报道。历史上最先报道的其中之一是电穿孔,其使用施加于含有植物细胞的溶液的电流(m.e.弗罗姆(m.e.fromm)等人,《自然》,319,791(1986);h.琼斯(h.jones)等人,《植物分子生物学(plantmol.biol.)》,13,501(1989)和h.杨(h.yang)等人,《植物细胞报告(plantcellreports)》,7,421(1988))。另一种直接方法(称为“基因枪轰击”)是使用涂有dna的超细颗粒(通常为钨或金)且然后用足够的力喷射到植物组织表面上,以促使颗粒穿透植物细胞,包括厚的细胞壁、膜和核被膜,但是不杀死其中至少一些(us5,204,253、us5,015,580)。第三种直接方法是使用纤维形式的金属或陶瓷,其由几乎刺穿细胞并且还刺穿细胞核被膜的尖锐、多孔或中空的针样突出组成。碳化硅和硼酸铝晶须均已经用于植物转化(水野(mizuno)等人,2004;派托尼诺(petolino)等人,2000;us5302523us申请20040197909)并且还已经用于细菌动物转化(凯普勒(kaepler)等人,1992;拉罗夫(raloff),1990;王(wang),1995)。其它方法已有报道,且无疑将研发额外的方法。本文教导的方法能够检测到由任何植物转化方法产生的非天然存在的核苷酸接合点。马铃薯育种前述转化方法典型地用于产生转基因变种。转基因变种然后可以与另一种(未转化或转化)变种杂交以产生新的转基因变种。或者,已使用前述转化技术工程改造成特定马铃薯品系的基因性状可以使用植物育种技术中众所周知的传统回交技术移入另一品系中。举例来说,回交方法可以用于使经工程改造的性状从大众化非精华变种移入精华变种内,或从基因组中含有外来基因的变种移入不含有那个基因的变种内。如本文中所用,视上下文而定,“杂交”可指简单的x与y杂交或回交过程。所属领域的一般技术人员将认识到,当本发明的上下文中使用术语马铃薯植物时,这也包括保留相关事件的基本区别性特征的衍生物变种,如那个变种的基因转化植物或其中并有一或多个附加值基因(如除草剂或害虫抗性)的转基因衍生物。回交方法可以联合本发明使用以改进变种的特征或将特征引入变种中。如本文所用,术语“回交”是指杂种后代返回与再现亲代重复杂交1、2、3、4、5、6、7、8、9或更多次。向基因贡献一或多种所期望特征的亲代马铃薯植物称为非再现或供体亲代。这个术语是指非再现亲代在回交方案中使用一次并且因此不再现的事实。来自非再现亲代的基因所转移的亲代马铃薯植物称为再现性亲代,因为其在回交方案中使用若干轮次。在典型回交方案中,所关注的最初变种(再现性亲代)与携带待转移的所关注基因的第二变种(非再现亲代)杂交。得自此杂交的后代然后再次与再现性亲代杂交并且重复所述过程直到获得马铃薯植物,其中除从非再现亲代转移的一或多种基因之外,所转化的植物中还恢复了再现性亲代的基本上全部的所期望形态学和生理学特征。选择适合的再现性亲代是成功回交程序中的重要步骤。回交方案的目标是改变或取代最初变种中的一或多种性状或特征。为此,用来自非再现亲代的所期望基因修饰、取代或补充再现性变种的一或多种基因,同时保留基本上全部的其余所期望基因,并且因此保留最初变种的所期望生理学和形态学组成。特定非再现亲代的选择将取决于回交的目的。主要目的之一是使植物增添一些商业上所期望、农艺学上重要的性状。确切回交方案将取决于为了确定适当测试方案而改变或增添的特征或性状。尽管回交方法在所转移的特征是显性等位基因时简化,但也可以转移隐性等位基因。在这种情况下,可能需要对后代引入测试以确定所期望的特征是否已成功地转移。同样,可以使用所属领域的技术人员众所周知的多种已确立重组方法中的任一种将转基因引入植物中,如:格里希尔(gressel),1985,赋予农作物除草剂抗性的生物技术:当前现状(biotechnologicallyconferringherbicideresistanceincrops:thepresentrealities),《植物基因组的分子形式和功能(molecularformandfunctionoftheplantgenome)》;范沃登-道亭(l.vanvloten-doting)(编),纽约普莱南出版社(plenumpress,newyork);赫特纳s.l.(huttner,s.l.)等人,1992,遗传改造植物的修正监管(revisingoversightofgeneticallymodifiedplants),《生物/技术(bio/technology)》;克里h.(klee,h.)等人,1989,植物基因载体和遗传转化:基于使用根癌农杆菌的植物转化系统(plantgenevectorsandgenetictransformation:planttransformationsystemsbasedontheuseofagrobacteriumtumefaciens),《植物的细胞培养和体细胞遗传学(cellcultureandsomaticcellgeneticsofplants)》;孔茨c.(koncz,c.)等人,1986,tl-dna基因5启动子控制新型农杆菌二元载体所携带的嵌合基因的组织特异性表达(thepromoteroft.sub.l-dnagene5controlsthetissue-specificexpressionofchimericgenescarriedbyanoveltypeofagrobacteriumbinaryvector),《分子和通用遗传学(molecularandgeneralgenetics)》;劳森c.(lawson,c.)等人,1990,商业马铃薯栽培品种对混合病毒感染的工程化抗性:转基因褐色布尔班克对马铃薯病毒x和马铃薯病毒y的抗性(engineeringresistancetomixedvirusinfectioninacommercialpotatocultivar:resistancetopotatovirusxandpotatovirusyintransgenicrussetburbank),《生物/技术》;米茨基t.a.(mitsky,t.a.)等人,1996,植物对plrv感染的抗性(plantsresistanttoinfectionbyplrv);美国专利第5,510,253号;纽厄尔c.a.(newell,c.a.)等人,1991,农杆菌介导马铃薯栽培种褐色布尔班克的转化(agrobacterium-mediatedtransformationofsolanumtuberosuml.cv.russetburbank),《植物细胞报告(plantcellreports)》;佩尔拉克f.j.(perlak,f.j.)等人,1993,遗传改良的马铃薯:避免科罗拉多马铃薯甲虫的伤害(protectionfromdamagebycoloradopotatobeetles),《植物分子生物学(plantmolecularbiology)》;所述文献均结合在此供参考以用于此目的。已经鉴别出多种性状,其未经常性地选用于研发新变种,但可以通过回交和遗传工程改造技术得到改良。这些性状可以是或可以不是转基因;这些性状的实例包括(但不限于):除草剂抗性;对细菌、真菌或病毒疾病的抗性;昆虫抗性;淀粉和其它碳水化合物浓度的均一性或增加;营养品质增强;块茎碰伤的倾向降低;以及淀粉转化成糖的速率降低。这些基因通常通过细胞核遗传。寄存信息以下寄存信息仅为了提供实例和所要求方法中所用的代表性植物材料而包括在内。上文所公开的j.r.新普洛特公司(j.r.simplotcompany)专有马铃薯栽培品种j3的块茎已经寄存于美国菌种保藏中心(americantypeculturecollection,atcc)(10801,弗吉尼亚州马纳萨斯大学大道(universityboulevard,manassas,va.),20110)。寄存日期是2013年5月23日。atcc寄存编号是pta-120371。参见美国专利第8,754,303号,“马铃薯栽培品种j3”,其结合在此供参考。上文所公开的j.r.新普洛特公司专有马铃薯栽培品种f10的块茎已经寄存于美国菌种保藏中心(atcc)(10801,弗吉尼亚州马纳萨斯大学大道,20110)。寄存日期是2013年5月23日。atcc寄存编号是pta-120373。参见美国专利第8,710,311号,“马铃薯栽培品种f10”,其结合在此供参考。上文所公开的j.r.新普洛特公司专有马铃薯栽培品种w8的块茎已经寄存于美国菌种保藏中心(atcc)(10801,弗吉尼亚州马纳萨斯大学大道(universityboulevard,manassas,va.),20110)。寄存日期是2014年3月11日。atcc寄存编号是pta-121079。参见美国专利第8,889,964号,“马铃薯栽培品种w8”,其结合在此供参考。上文所公开的j.r.新普洛特公司专有马铃薯栽培品种j55的块茎已经寄存于美国菌种保藏中心(atcc)(10801,弗吉尼亚州马纳萨斯大学大道,20110)。寄存日期是2013年9月25日。atcc寄存编号是pta-120601。参见美国专利第8,889,963号,“马铃薯栽培品种j55”,其结合在此供参考。上文所公开的j.r.新普洛特公司专有马铃薯栽培品种e12的块茎已经寄存于美国菌种保藏中心(atcc)(10801,弗吉尼亚州马纳萨斯大学大道,20110)。寄存日期是2013年5月23日。atcc寄存编号是pta-120372。参见美国专利申请第14/072,487号,“马铃薯栽培品种e12”,其结合在此供参考。上文所公开的j.r.新普洛特公司专有马铃薯栽培品种x17的块茎已经寄存于美国菌种保藏中心(atcc)(10801,弗吉尼亚州马纳萨斯大学大道,20110)。寄存日期是2015年6月17日。atcc寄存编号是pta-122248。上文所公开的j.r.新普洛特公司专有马铃薯栽培品种y9的块茎已经寄存于美国菌种保藏中心(atcc)(10801,弗吉尼亚州马纳萨斯大学大道,20110)。寄存日期是2015年6月17日。atcc寄存编号是pta-122247。上文所公开的j.r.新普洛特公司专有马铃薯栽培品种v11的块茎已经寄存于美国菌种保藏中心(atcc)(10801,弗吉尼亚州马纳萨斯大学大道,20110)。寄存日期是。atcc寄存编号是。实例实例1:psim1278转化载体骨架质体psim1278是用于转化马铃薯的19.7kb二元转化载体。这个实例展示基因元件的来源、骨架的克隆步骤和t-dna序列,以及质体中的元件顺序。质体骨架(图1和表1)含有两个经充分表征的细菌性复制起点。pvs1(pvs1sta和rep)能够使质体维持于农杆菌中,且pbr322(pbr322bom和ori)能够使质体维持于大肠杆菌中。农杆菌dna过驱动序列使rb处的裂解增强,且大肠杆菌nptii基因是细菌卡那霉素可选标记物。骨架含有表达盒,所述表达盒包含农杆菌异戊烯基转移酶(ipt)基因、侧接的褐色兰吉尔马铃薯聚泛素(ubi7)启动子和褐色兰吉尔马铃薯聚泛素(ubi3)终止子。ipt盒是可筛选的表型,其用于选择整合到宿主植物中的质体骨架dna。当存在于所转化的植物组织中时,ipt的过度表达引起植物激素细胞分裂素的过度产生,从而产生表型转轨、叶子异常且根失能的植物。骨架部分未转移到植物细胞中。骨架的各种元件描述于表1中。表1.psim1278骨架的基因元件1http://www.cambia.org/daisy/cambia/585.html-(pcambia载体的通用结构图)实例2:psim1278转化载体t-dnapsim1278中使用的psim1278dna插入区域(包括侧接边界序列)的长度是10,148bp(从1bp到10,148bp)。psim1278dna插入序列仅由原生dna组成并且稳定整合到马铃薯基因组中。psim1278dna插入序列或其功能部分是整合到本发明的马铃薯植物变种中的载体psim1278的唯一遗传物质。psim1278dna插入序列描述于:图1(与载体骨架区域一起)、图2、图5和下文表2中。lb和rb序列(各25bp)是以合成方式设计以与根癌农杆菌的t-dna边界类似且发挥类似功能。修订基因库(genbank)寄存号ay566555以阐明用于边界区域的dna的来源。查瓦拉(chawla)等人(2012)将描述为基因元件5和10的asn1称为stast1。质体psim1278t-dna含有两个表达盒:第一个盒(元件4到12,表2)引起所转化马铃薯变种中的asn1和ppo5下调。其包含asn1的两个相同405bp片段和ppo5的两个相同144bp片段。asn1和ppo5的片段是作为被非编码157bp褐色兰吉尔马铃薯核苷酸间隔元件分隔的反向重复序列配置。asn1和ppo5片段配置于两个趋同马铃薯启动子之间;adp葡萄糖焦磷酸化酶基因(agp)的agp启动子和颗粒结合的淀粉合成酶基因(gbss)的gbss启动子,这些启动子主要在块茎中发挥活性。这些启动子驱动反向重复序列表达以产生双股rna且下调asn1和ppo5。第二个盒(元件14到21,表2)引起所转化马铃薯变种中的phl和r1下调。其包含phl启动子区(pphl)的两个相同509bp片段和r1启动子区(pr1)的两个相同532bp片段。pphl和pr1片段是作为被褐色兰吉尔马铃薯聚泛素基因的非编码258bp片段分隔的反向重复序列配置。如第一盒,pphl和pr1片段配置于马铃薯agp和gbss启动子之间且通过马铃薯agp和gbss启动子转录。表2.psim1278t-dna的基因元件(从左侧边界位点到右侧边界)因此,正如从表1和表2可以看出,psim1278质体是针对马铃薯植物转化所设计的二元载体。载体骨架含有用于在大肠杆菌和农杆菌中复制的序列,以及ipt标记,所述ipt标记用于筛选以排除具有载体骨架dna的植物。t-dna区域由两个表达盒、侧接的lb和rb序列组成。含有psim1278载体的农杆菌接种宿主植物组织后,将psim1278的t-dna区域转移到宿主基因组中。表2中所述的用于产生本发明马铃薯品系的dna插入序列不使相邻基因活化并且对马铃薯植物变种的表型不会产生不利的影响。实例3:psim1678转化载体骨架质体psim1678是用于转化马铃薯的18.6kb二元转化载体。这个实例展示基因元件的来源、骨架的克隆步骤和t-dna序列,以及质体中的元件顺序。质体骨架(图3;表3)含有两个经充分表征的细菌性复制起点。pvs1(pvs1sta和rep)能够使质体在农杆菌中维持,且pbr322(pbr322bom和ori)能够使质体在大肠杆菌中维持。农杆菌dna过驱动序列使rb处的裂解增强,且大肠杆菌nptii基因是细菌卡那霉素可选标记物。骨架含有表达盒,所述表达盒包含农杆菌异戊烯基转移酶(ipt)基因、侧接的褐色兰吉尔马铃薯聚泛素(ubi7)启动子和褐色兰吉尔马铃薯聚泛素(ubi3)终止子(加巴里诺(garbarino)和贝尔纳普(belknap),1994)。ipt盒是可筛选的表型,其用于选择整合到宿主植物中的质体骨架dna。当存在于所转化的植物组织中时,ipt的过度表达引起植物激素细胞分裂素的过度产生,从而产生表型转轨、叶子异常且根失能的植物。骨架部分未转移到植物细胞中。骨架的各种元件描述于表3中。表3.psim1678骨架的基因元件实例4:psim1678转化载体t-dnapsim1678中使用的psim1678dna插入区域(包括侧接边界序列)的长度是9,090bp(从1bp到9,090bp)。psim1678dna插入序列仅由原生dna组成并且稳定整合到马铃薯基因组中。psim1678dna插入序列或其功能部分是整合到本发明的马铃薯植物变种中的载体psim1678的唯一遗传物质。psim1678dna插入序列描述于图3(与载体骨架区域一起)、图5和下文表4中。在表4中,lb和rb序列(各25bp)是以合成方式设计以与根癌农杆菌的t-dna边界类似且发挥类似功能。修订基因库寄存号ay566555以阐明用于边界区域的dna的来源。质体psim1678t-dna是1bp到9,090bp且含有两个表达盒(图3):第一个盒(元件4到6,表4)含有来源于土癫茄(solanumventurii)的2,626bprpi-vnt1(vnt1)基因。基因产物vnt1是r-蛋白质,其涉及的植物免疫反应保护马铃薯以防马铃薯晚疫病菌的晚疫病感染。基因是在原生vnt1启动子pvnt1和终止子tvnt1下表达。第二个盒(元件8到14,表4)使得液泡转化酶(vinv)在转化的马铃薯变种中下调。其包含作为所分隔的反向重复序列配置的vinv的两个片段(元件10和12,表4)。vinv片段配置于两个趋同的马铃薯启动子之间;adp葡萄糖焦磷酸化酶基因(agp)的agp启动子和颗粒结合的淀粉合成酶基因(gbss)的gbss启动子,这些启动子主要在块茎中发挥活性。这些启动子驱动反向重复序列表达以产生双股rna且下调vinv。表4.psim1678t-dna的基因元件(从左侧边界位点到右侧边界)因此,正如从表3和表4可以看出,psim1678质体是针对马铃薯植物转化所设计的二元载体。载体骨架含有用于在大肠杆菌和农杆菌中复制的序列,以及ipt标记,所述ipt标记用于筛选以排除具有载体骨架dna的植物。t-dna区域由两个表达盒、侧接的lb和rb序列组成。含有psim1678载体的农杆菌接种宿主植物组织后,将psim1678的t-dna区域转移到宿主基因组中。实例5.农杆菌菌株和转染通过准确地使超毒性质体ptibo542的转移dna缺失来产生c58衍生的农杆菌菌株agl1(拉佐(lazo)等人,1991)。转座子插入通用重组基因(reca)中使重组质体载体(如psim1278)稳定(图1)。agl1呈现针对羧苄青霉素(carbenicillin)和利福平(rifampicin)的抗性,且使用特美汀(timentin)从转化的马铃薯组织中排除。选择之后,植物不含抗生素和农杆菌,其中马铃薯来源的表达盒插入植物的基因组中。储备植物在洋红色盒中维持,所述洋红色盒具有含有3%蔗糖和2g/l结冷胶(gelrite)的40ml半浓度m516(植物技术公司(phytotechnology))培养基(繁殖培养基)。从四周龄植物切下四到六毫米的马铃薯节间区段,用携带psim1278的农杆菌agl1菌株感染,并且转移到含有3%蔗糖和6g/l琼脂的组织培养基(共培养基)中。两天之后,将所感染的外植体转移到含有3%蔗糖、6g/l琼脂和150mg/l特美汀(以排除农杆菌)的m404(植物技术公司)培养基(无激素培养基)中。拉克尔(richael)等人(2008)详细描述了所述方法。一个月之后,将所感染的外植体转移到缺乏任何合成激素的新鲜培养基中且在珀西瓦尔(percival)生长箱中、在24℃、在16小时光周期下培育,在生长箱中,其开始形成嫩枝。许多嫩枝表达ipt且呈现细胞分裂素过度产生表型;这些嫩枝不考虑用于进一步分析。pcr基因分型表明剩余嫩枝中约0.3%到1.5%含有p-dna的至少一部分,同时缺乏ipt基因。因此,不使用标记物选择所转化的植物。拉克尔等人(2008)公开了基于ipt的无标记植物转化的细节。外植体感染之后两天开始执行排除农杆菌的方法。为此目的,使组织暴露于抗生素特美汀(150mg/l)直到证实不含活的农杆菌。通过在28℃下将转化事件的茎片段在营养肉汤-酵母提取物(nby培养基)上培育2周来获得证据(重复两次)。根据97cfr部分340,所转化的植物仅当不含活的农杆菌时运输且种植于田间。褐色布尔班克w8事件含有使用不同质体进行两次单独转化而产生的插入序列。第一插入序列,质体psim1278,含有两个由反向重复序列组成的盒,所述反向重复序列经设计以使块茎中的多达四种马铃薯基因asn1、ppo5、r1和phl静默。类似地,第二质体psim1678含有由使块茎中的vinv基因静默的反向重复序列组成的盒,同时还含有其原生马铃薯启动子下的rpi-vnt1基因拷贝。通过dna凝胶印迹分析法来分析马铃薯植物变种,以测定所整合的dna插入序列的结构和拷贝数且确认载体骨架序列的不存在。另外,利用分子特征来确定侧接dna插入序列的接合点的序列且展示所插入dna的稳定性。接合点的定序信息为研发出针对基因内马铃薯植物变种的特定pcr测试提供了依据。因此,所揭露的方法广泛地适用于其它植物物种和其它马铃薯栽培品种(这在所述领域的技术人员的技能范围内),以对已经经历转化事件的植物物种的dna进行定序且鉴别由其所得的非天然存在的核苷酸接合点。所述技术人员然后能够研发出结合到所述非天然存在的接合序列的适当探针和为了扩增此类序列而优化的引物。实例6.载体骨架dna不存在的证据不同于多种商业转基因农作物,本发明的马铃薯栽培品种通过以下三种不同方法证实不含转化所用的来源于农杆菌的dna序列,如载体骨架dna:1)首先,确定负向选择性异戊烯基异构酶(ipt)标记基因存在或不存在于载体骨架中,因为包含来自农杆菌的ipt基因表达盒的骨架dna无意中转移到植物细胞中会触发ipt基因表达且因此触发细胞分裂素型激素异戊烯基腺苷形成;2)然后对所转化的马铃薯植物使用南方印迹杂交,通过第一筛选方法确认骨架dna的不存在;和3)然后设计pcr以扩增指示dna插入序列边界区域与侧接骨架dna或侧接dna插入序列的骨架dna内的区域之间的接合点的片段。使用psim1278dna作为阳性对照来证实所述方法的功效。本发明的马铃薯栽培品种未产生指示载体骨架dna存在的pcr色带。实例7.所插入的dna的稳定性使用dna凝胶印迹杂交和性状评价法评价dna插入序列在最初转化体中的稳定性以及另外在繁殖植物材料中的稳定性。进行这些研究是为了确保基因内事件按一贯且可靠的方式表达所并入的性状。不稳定性可能由稀少的重组事件触发或还可以由甲基化引起。由于马铃薯通常以克隆方式繁殖,因此不能直接使用性繁殖农作物的标准评估,且使用块茎而非种子来定义后代。dna印迹杂交结果表明多个代存在一致的色带,由此指明稳定性。通过证实第一代和第二代块茎种子的性状功效来获得稳定性的其它证据。最初转化材料(g0)中的dna插入序列稳定性是通过提取和评价已经活体外繁殖且未曾在土壤中种植的植物叶子中的dna来证明。在第1代(g1)分析中,在温室中种植两株由每种基因内变种繁殖的植物和一株来自每种对照的植物;种植从每种植物收获的块茎之一以获得g1植物的叶子,所述叶子用于分离dna且评价g1代。来自这一代的块茎再次种植,且所得g2植物的叶子允许对那一代进行表征。使用南方印迹法分析w8马铃薯经历三代(g0-g3)所分离的基因组dna表明插入序列的结构是稳定的,同时通过测量第二代田间种植的块茎中的多酚氧化酶活性来评估表型稳定性。这种方法以可见的证据表明了在将儿茶酚向所切割的马铃薯表面之后存在ppo静默。进行这些研究是为了确保w8中的所期望基因变化在多次克隆循环中保持稳定,同时维持性状。通过使用南方印迹法比较三个连续克隆代(g1、g2和g3)与最初转型体(g0)来评价dna插入序列的稳定性。预期稳定的dna插入序列在植物的多个代中维持相同结构并且由此产生相同的消化模式。为了测试w8事件中插入序列的稳定性,使用与来自psim1278和psim1678的插入序列的区域杂交的两个探针(gbs1和agp)和对psim1678插入序列具有特异性的两个探针(inv和vnt1)比较其消化模式。由于与这些探针杂交的dna序列包含于马铃薯基因组中以及dna插入序列内,因此预期南方印迹中存在内源性和插入序列特异性色带。所有基因组dna样品由限制酶ecorv消化并且与对agp或gbs1具有特异性的探针杂交。这些研究中选择ecorv,原因是其在两种插入序列内消化以提供独特显带图案,其中psim1278插入序列具有所预测尺寸的内部色带(例如2.3kb)。对于两种探针来说,w8的所有样品之间的显带图案彼此相同。w8中还发现存在于褐色布尔班克对照物中的多个色带,但w8还含有对应于psim1278和psim1678插入序列的色带。这些色带在所分析的w8的所有代之间类似地一致,表明两种插入序列的遗传稳定性。使用对psim1678插入序列具有特异性的两种探针进行第二次分析。在此分析中,基因组dna样品用限制酶xbai消化并且与vnt1和inv探针杂交。这些研究中选择xbai作为限制酶,因为其在内部消化psim1678并且产生具有已知尺寸的色带(例如对于inv探针而言,4.6kb)。再次检测到内源性和插入序列特异性色带,其中所分析的三代之间的色带图案一致。基因和表型分析表明psim1278和psim1678转化所产生的插入序列在三代内是稳定的。鉴于三代内的稳定性已得到证明,因此稳定性将在后续无性繁殖循环期间维持是可能的。实例8:接合点分析和变种特异性检测使用接附子连接介导式pcr或热不对称交织式pcr对dna插入序列/侧接植物dna接合点进行定序。接合序列用于设计本发明的马铃薯栽培品种的引物,且这些引物应用于基于变种特异性pcr的检测方法中。引物可以用于扩增变种特异性dna片段dna片段,从而产生用于所述变种的品系特异性测试方法。所研发出的方法是用于监测田间和储存中的植物和块茎,以确认块茎或所加工食物中不存在基因内材料,以及确保有机种子的纯度。实例9:基因静默的功效和组织特异性基因静默方法是用于降低asn1、ppo5、phl、r1和vinv原生蛋白质的活性,且评价转录物含量而非蛋白质量以使新表型性状与分子量变化关联。由于涉及叶子和茎中形成asn(天冬酰胺)的asn1基因的强烈静默对生长的影响可能不利,因此agp启动子和gbss启动子(其是块茎特异性和匍匐茎特异性启动子且在光合作用具有活性的组织和根中具有小得多的活性)用于驱动块茎和匍匐茎中的基因静默。五种靶向基因在植物变种以及其未转化的对应物的各种组织中的转录物含量是通过北方印迹分析进行测定。引入褐色布尔班克中以产生w8事件的三个基因静默盒中的两个在rnai介导式静默中极有效地静默其目标转录物。这两种构筑体有效地使w8块茎中的asn1、ppo5和vinv静默。对块茎的静默特异性表明rnai机制所产生的sirna很少(如果存在)传播到其它组织中或其含量不足以激发那些组织中的rnai反应。块茎外部静默的唯一证据是在花朵中,其中观察到较低含量的asn1,但变化量级比块茎低得多。用phl和r1进行的启动子静默策略的效果最小,这与含有相同psim1278构筑体的其它事件一致(科林奇(collinge)和克拉克(clark),2013)。若干种基因内马铃薯栽培品种的特定组织内下调的转录水平的概述示于表5中。表5中的每个字母(a、p、l、r)表明静默得到确认,但是静默的量根据基因和组织而变。表5.不同组织内下调的基因的概述事件块茎1匍匐枝1根1茎1叶子1花1,2f10apralraprpaae12aplralrappaj3aplraraapaj55aplralrapa1a=asn1,p=ppo5,l=phl,r=r1。表格中的字母表示组织的基因表达下调。2部分下调的asn1基因表达可能改变花的氨基酸组成。此类效果将局限于asn的减少和gln的增加。由于asn和gln是类似的非必需氨基酸,因此预期这些化合物含量的变化不会影响作为昆虫或其它生物体饲料的花瓣、花蜜和花粉的品质。实例10:从马铃薯植物叶子组织中分离dna从3g马铃薯植物叶子组织中提取dna,在液氮中磨碎且与20ml提取缓冲液(0.35m山梨糖醇、0.1mtris-hclph8.0、0.5medtaph8.0)混合。样品以3,000rpm离心5分钟且用2ml提取缓冲液冲洗。将离心块再悬浮于4ml提取缓冲液和4μl100mg/ml核糖核酸酶a中。向各样品中添加四毫升核溶解缓冲液(1mtris-hclph8.0、0.5medtaph8.0、5mnacl、20mg/mlctab)和1.6ml5%十二烷基肌氨酸钠(sarcosyl)。混合样品且在65℃下、在搅拌下培育20分钟。将样品和等体积的氯仿:异戊醇(24:1)一起振荡且以3,000rpm离心5分钟。保留水相且重复氯仿提取步骤2-3次。dna沉淀于等体积的异丙醇中且以3,000rpm离心10分钟。离心块用70%乙醇冲洗,干燥且再悬浮于te中。实例11:基于高产量ctab的dna提取方法马铃薯和马铃薯产物含有高含量的多醣,所述多醣能干扰pcr,尤其是qpcr。因此,建议使用产生高品质dna的方法进行dna分离,所述方法使用为了防止pcr抑制剂(如多醣)干扰而设计的预混液(mastermixes)进行qpcr。为了满足这些要求,使用十六烷基三甲基溴化铵(ctab)分离方法和获自量子公司(quanta)的qpcr(实例16)。使用事件特异性引物测试从马铃薯叶子材料中提取的dna的这些pcr方法已产生高pcr效率、良好线性度、目标特异性和稳固性。提取方法的流程1.每天制备新鲜ctab缓冲液(1mtris-hclph8.0、0.5medtaph8.0、5mnacl和20mg/mlctab)且预温热到65℃。2.向薯片和块茎中添加10mlctab/向薄片和炸薯条中添加30mlctab缓冲液。向叶子中添加1mlctab。3.将以下量的起始物质添加到适当的试管中:薄片:3g冻干的炸薯条:3g(若使用新鲜炸薯条,则使用6g)薯片(研磨薯片直到平滑的糊状物):1g冻干的块茎:1g冻干的叶子(研磨到细粉):0.5g4.向每个试管和混合物中每毫升添加3μl蛋白酶k以除去凝块。5.在65℃下,在培育箱中,在210rpm下振荡培育2-3小时(对于块茎和叶子来说,45分钟是足够的)6.样品以14,000rpm离心40分钟(对于块茎来说,20分钟足够且对于叶子来说,10分钟足够)7.将上清液转移到清洁的试管中且在冰上培育20分钟8.添加等体积的冰冷氯仿9.任选:每1g起始物质添加200μlphytopuredna提取树脂(通用电气医疗生活科学部(gehealthcarelifesciences))。使用之前将树脂涡旋以确保均匀。10.将样品在室温下剧烈混合。11.样品以14,000rpm离心40分钟(对于块茎来说,离心20分钟且对于叶子来说,离心10分钟)12.将上层水相转移到清洁的试管中。13.重复进行氯仿提取直到界面清晰(不需要额外的树脂)。14.添加1/10体积的3m乙酸钠(ph5.3)15.添加等体积的异丙醇。16.将试管倒置直到dna沉淀(在4℃过夜是最佳的)17.以14,000rpm离心20分钟,得到dna离心块(对于叶子来说,5分钟足够)。此时,有些样品可能不具有明显的离心块。在这种情况下,小心地吸移出上面的透明层且保留下面的较深色层,这将在步骤19期间离心。18.用冰冷的70%乙醇洗涤离心块。19.离心块以14,000rpm离心5-10分钟。20.离心块风干10分钟或直到离心块边缘看起来透明。21.将dna再悬浮于200-400μlte缓冲液中。22.添加核糖核酸酶直到20μg/ml的浓度。在37℃培育30分钟。23.添加5个体积的qiagen缓冲液pb到dna中。每个柱不超过10μgdna。24.使dna/pb溶液通过柱进行离心。25.用缓冲液aw2或pe洗涤2次26.使柱以14,000rpm离心2分钟以使柱干燥。27.将适量的te(50μl)施加到柱中且在65℃搁置5分钟,随后洗提。28.应该使用荧光嵌入染料(例如高灵敏度qubit)测量dna浓度且应该通过连续稀释分析测试样品中的pcr抑制剂,尤其是在执行定量分析时。实例12:使用qiaamp粪便dna快速提纯小型试剂盒(qiaampfastdnastoolminikit)从块茎、薄片、薯片和炸薯条中分离dna执行以下方案来分离dna。使用这种技术所分离的dna样品的纯度和产量倾向于低于前述章节中所述的基于ctab的方法。由于pcr抑制剂的存在,因此必须注意以确保dna的品质足以用于任何定量分析。任何基质特异性说明均为粗体。称取约300mg的薄片、薯片或炸薯条组织且放入含有1.7mlqiageninhibitex缓冲液(预热缓冲液以将任何沉淀物再溶解)的2.0ml微量离心试管中。对于冻干的块茎来说,在1.2mlinhibitex缓冲液中使用150mg组织。炸薯条和薄片材料需要将两个300mg样品分别溶解且在下游合并于单柱上。对于薯片样品来说,可以将任选的两个样品溶解且合并以增加产量。通过涡旋充分混合30-60秒。将薄片、薯片和块茎样品在加热块(优选)或水浴中、在95℃±5℃下培育30分钟。对于炸薯条样品来说,改在70℃下培育30分钟。将试管在微量离心机中以14,000rpm离心5分钟。从每个试管中转移600-650μl上清液到个别的2.0ml微量离心管中,且添加25μl于qiagen试剂盒中提供的蛋白酶k。通过涡旋充分混合。向每个试管中添加600-650μl缓冲液al且通过涡旋混合。在70℃下培育样品10分钟。添加600-650μl95-100%乙醇(未提供于试剂盒中)且通过涡旋混合。使用真空歧管每次传送650μl溶液通过qiaamp离心柱或以21,000×g脉动离心。对于炸薯条和薄片材料来说,将两个管中的溶液传送通过单个柱以浓缩。最后以14,000rpm离心1分钟以除去所有缓冲液。柱用500μl缓冲液aw1洗涤一次且用500μl缓冲液aw2洗涤一次,使用真空歧管或14,000rpm离心1分钟。将柱移到2ml收集管中,以14,000rpm离心3分钟且抛弃流过物。将柱转移到一个新的1.7ml微量离心管中且吸移50μlte(用于块茎和薯片)或吸移30μlti(用于炸薯条和薄片样品)直接置于膜上。在室温下培育3分钟。以14,000rpm离心1分钟以洗提dna。对dna(在-20℃储存)进行定量和qa。应该记录所有样品的(od260/od280)和(od230/od260)比率。应该使用荧光嵌入染料(例如生命技术公司(lifetechnologies)的qubitdsdnahs分析试剂盒)测量dna浓度且应该通过连续稀释分析测试样品中的pcr抑制剂,尤其在执行定量分析时。实例13:dna插入序列和非天然存在的接合序列在马铃薯事件中的分子特征通过dna凝胶印迹分析法来分析所有马铃薯事件,以测定所整合的dna插入序列的结构和拷贝数且确认载体骨架序列的不存在。这些研究是作为所述事件的表征和生物安全评估的一部分进行。利用分子特征来确定侧接dna插入序列的接合点的序列且展示所插入dna的稳定性。接合点的定序信息为研发出所有事件的事件特异性pcr测试提供了依据(表6)。图5展示表6中的seqidnos,其中每个非天然存在的构筑体特异性接合点存在于psim1278和psim1678的dna插入区域,且图6a-i展示表6中的seqidnos,其中每个非天然存在的构筑体特异性和事件特异性接合点存在于e12、f10、j3、j55、v11、w8、x17和y9事件中。就表6来说,前两个序列(seqidno:1和2)代表存在于插入序列的最外侧边缘上的接合点,其是25个核苷酸的合成序列且不是马铃薯基因组的一部分。其存在于左侧边界和右侧边界上。这些接合序列作为合成lb/马铃薯lb和合成rb/马铃薯rb包括于表6中。另外,表6用粗体字和突显字提供:1)与染色体dna具有微同源性的序列,这是与染色体dna共同以及与边界序列共同的用于插入的序列。这些序列在其所存在的序列内用粗体和突显字列出。表6还提供:2)存在于接合点之间的间插序列。这些序列加有下划线且接合点任一侧均已存在15bp序列。表6.非天然存在的构筑体特异性和事件特异性接合序列马铃薯事件e12、f10、j3、j55、v11、w8、x17和y9中的dna插入序列的结构图示于图4和图6中。事件e12和f10含有1个拷贝(其中“拷贝”意味着存在至少一个asn1/ppo5基因静默盒)。事件j3和j55含有2个拷贝。在较高拷贝事件与仅有一个拷贝的事件之间,静默活性的程度持久性不存在差异。事件j55含有作为反向重复序列定位的两个所连接dna插入序列(图4和图6)。莱齐坦伯格(lechtenberg)等人(2003)表明第二基因拷贝以串联或反向配置的细菌t-dna不产生静默。因此,j55中的反向连接dna插入序列拷贝可能不会促成静默且因此,靶向基因的静默如所预期是基于定位于趋同启动子之间的反向重复序列来发挥作用。与用psim1278和psim1678转化褐色布尔班克以产生事件w8有关的插入序列的基因和结构表征表明两种转化均使得每个质体产生单一整合位点。psim1278转化所产生的dna的结构相对于最初插入序列的结构来说是复杂的。所插入的dna似乎在转化期间经历重排,产生了由以下组成的结构:串联重复的asn1/ppo5静默盒,继之为几乎完整的psim1278构筑体,和反向重复序列,所述反向重复序列含有复制的pr1/pph1静默盒和串联复制的gbss启动子以及间插的ph1序列(图6)。尽管这种结构比预期更复杂,但所复制的静默盒是完整的并且仍然处于组织特异性启动子的控制下。所述结构对产物的安全性或性状功效不会有不利的影响。w8还含有来自psim1678的dna的单个拷贝,其存在于整合的单一基因座(图6)。psim1678的dna插入序列含有几乎完整的dna插入序列,其中缺失330bp,其除去了整个t-dna左侧边界和137bp的rpi-vnt1启动子。启动子中的这种小缺失不影响基因赋予晚疫病抗性的能力。另外,已经使用rt-pcr证明与rpi-vnt1基因相关的rna表达。插入序列偶尔侧接来源于植物基因组的短dna序列或dna插入序列。这些插入似乎是整合过程的一部分且发生的频率相当高(温德尔(windels)等人,2003)。具有此类序列的事件的实例包括j55的两个dna插入序列之间的49bp序列。使用基因库对这种短dna序列进行的碱基局部比对检索工具(blast)检索使马铃薯的已知序列得到部分的匹配,证实来源最可能来自植物基因组或dna插入序列。大多数所转移的dna插入序列短于左侧与右侧边界序列之间的全距离,如根据边界附近的短缺失所表明。此类缺失还与t-dna整合有关且假设起因于双股断裂修复(戈森(gheysen)等人,1991)。不消弱两个静默盒的功能活性的短缺失发现于例如事件f10中(右侧边界缺失38bp)。实例14:qpcr引物和探针研发设计事件特异性引物是为了扩增基因组中的对所关注事件具有特异性的区域(马铃薯基因组侧接区域)或对psim1278构筑体或psim1678构筑体本身中的接合点具有特异性的区域。这些引物扩增了70到200个碱基对的区域,在此区域内,目标特异性荧光探针的结合允许对产物进行实时检测和定量。每个荧光探针在5′端经6-fam(6-羧基荧光素)部分标记且在3′端经bhq1(黑洞淬灭剂tm1)部分标记。6-fam的荧光是由相同寡核苷酸上存在的bhq1淬灭。在pcr期间,与所扩增的目标股粘接的探针将通过taqdna聚合酶的5'到3'核酸酶活性裂解,从而分离6-fam和bhq1部分。由此消除bhq1对6-fam的淬灭,引起6-fam荧光发射。探针设计为锁核酸(lna)探针以增强热稳定性和特异性(彼得森m.(petersen,m.)和温吉尔j.(wengel,j.)(2003).lna:治疗学和基因组学的通用工具(lna:aversatiletoolfortherapeuticsandgenomics),《生物技术趋势(trendsinbiotechnology)》,doi:10.1016/s0167-7799(02)00038-0)。使用引物和对马铃薯的腺嘌呤磷酸核糖基转移酶(aprt)具有特异性的双重标记(fam和tam或bhq1)探针扩增aprt作为内源对照。在使用实时pcr对马铃薯进行各种生物和非生物压力研究期间,基于稳定性来选择aprt(尼科特n.(nicot,n.),霍斯曼j.-f.(hausman,j.-f.),霍夫曼l.(hoffmann,l.),和伊佛d.(evers,d.),(2005),选择管家基因用于对生物和非生物压力期间的马铃薯进行实时rt-pcr标准化(housekeepinggeneselectionforreal-timert-pcrnormalizationinpotatoduringbioticandabioticstress),《实验植物学杂志(journalofexperimentalbotany)》,56(421),2907-14.doi:10.1093/jxb/eri285)。然而,可以使用任何适当的对照基因。表7中列出了正向和反向引物以及对aprt对照或对每个事件具有特异性的双重标记探针。表7.qpcr分析中所用的寡核苷酸序列实例15:分析的效率和线性度被优化的qpcr分析的两个标志是高pcr效率(e)和根据相关系数r2度量的线性标准曲线。理想的qpcr反应在-3.32的斜率下具有100%的效率,这与pcr产物在每个循环期间的完全倍增相关。然而,斜率在-3.1与-3.6之间且效率在90%与110%之间通常视为可接受的(康密辛c.a.(commission,c.a.)(2009)。gmo实验室欧洲网(engl)gmo测试分析方法的最小性能要求定义(definitionofminimumperformancerequirementsforanalyticalmethodsofgmotestingeuropeannetworkofgmolaboratories(engl))),(2008年10月),1-8))。效率是根据复制的标准曲线确立。扩增效率是根据标准曲线的对数线性部分的斜率测定且计算如下:e=(10(-1/斜率)-1)*100(布什丁s.a.(bustin,s.a.)等人(2009)。miqe指南:关于定量实时pcr实验的最小公开信息(themiqeguidelines:minimuminformationforpublicationofquantitativereal-timepcrexperiments)。《临床化学(clinicalchemistry)》,55(4),1-12.doi:10.1373/clinchem.2008.112797)且r2值是根据线性回归分析来确定,其应该≥0.98(布什丁(bustin)等人,2009)。表8展示了每个事件特异性和构筑体特异性qpcr分析的效率和线性度。各分析表明高效率的建议范围是90到110%且r2值大于或等于0.98。所呈现的数据来自使用60℃的组合式粘接/延长温度对叶子dna进行的分析。表8.事件特异性qpcr分析的效率和线性度*使用具有seqidno79-81的引物/探针组测定^使用具有seqidno82-84的引物/探针组测定1使用具有seqidno85-87的引物/探针组测定(检测vnt1终止子与agp启动子之间的接合点)实例16:马铃薯叶子dna分析中的检测水平(lod)根据gmo实验室欧洲网,“检测极限是样品中的分析物的最低量或浓度,其可以可靠地检测,但不一定能够定量,如单一实验室验证所证明”(康密辛(commission),2009)。lod应该在至少95%的时间内检测到分析物,从而得到≤5%的假阴性结果。所报道的分析各自能够可靠地在大于95%的时间内检测到至少24pg的总目标dna。由于dna可以片段化或含有抑制剂,尤其是多醣,因此lod将根据所用dna的来源而变,如从基于马铃薯的食物产品所分离的dna中频繁地看到。定性和定量分析可以达到<0.1%gmo,限制条件为有足够的dna用于pcr反应。表9展示了每个事件特异性和构筑体特异性qpcr分析的检测水平。各分析利用34到35个循环可靠地扩增至少24pg的马铃薯叶子dna。所呈现的数据来自使用60℃的粘接/延长温度进行的分析。表9.事件特异性qpcr分析的检测水平分析lod下的平均cqe1235.29f1035.21j334.62j5535.32w835.99v1135.04x1735.96y932.85psim1278构筑体*34.42psim1278构筑体^34.78psim1678构筑体134.17*使用具有seqidno79-81的引物/探针组测定^使用具有seqidno82-84的引物/探针组测定1使用具有seqidno85-87的引物/探针组测定实例17:事件特异性qpcr分析的稳固性方法的稳固性是其在分析中不受实验条件微小变化影响的能力的度量。举例来说,pcr分析应该能够由不同使用者对不同热循环仪模型进行,其中温度特征曲线或dna聚合酶的偏差较小。人们普遍接受的是,在这些条件下,分析的偏差不应超过±30%(康密辛(commission),2009)。我们在组合式粘接/延长温度为58℃到61℃的四度范围内测定了各分析的效率和线性度(表10)。分析的进一步稳固性是通过外部实验室使用不同使用者、热循环仪和聚合酶在外部验证分析性能来确定。表10展示了各分析在组合式粘接/延长温度的四度范围内的效率和线性度。除58℃下的f10之外,所有分析均得到良好的执行,在整个范围内,效率为90-110%且r2值≥0.98。表10.事件特异性qpcr分析在4°温度范围内的效率和线性度*使用具有seqidno79-81的引物/探针组测定^使用具有seqidno82-84的引物/探针组测定1使用具有seqidno85-87的引物/探针组测定实例18:品系特异性引物和探针的特异性通过使用25ngdna,在dna的三种浓度(25ng、250pg和50pg)下,对来自各事件(e12、f10、j3、j55、w8、v11、x17和y9)和各种商业变种(大西洋、褐色兰吉尔、褐色布尔班克和斯诺登)的dna执行qpcr来评估每个引物和探针组的特异性。用于检测e12、j3、j55、w8、v11、x17和y9的引物在所测试的任一种浓度下未展现假阳性或假阴性。39个循环之后,用于f10的引物扩增了代表单一技术的25nge12样品,这表示2.3%的假阳性率在5%的可接受范围内。41个循环之后,用于psim1278构筑体的引物扩增了代表单一技术的两个不同野生型变种。为了改进这个分析,调整pcr循环参数且对样品进行再测试。psim1278或psim1678的新参数未产生假阳性。事件(e12、f10、j3、j55、w8、v11、x17和y9)和构筑体(psim1278和psim1678)的qpcr反应配置和qpcr循环条件示于表11-13中。表11.pcr反应配置和条件(20μl总体积)*预混液必须含有减少pcr抑制的组分表12.e12、f10、j3、j55、w8、v11、x17和y9的pcr热循环条件表13.psim1278和psim1678的pcr热循环条件所述方法允许使用qpcr检测所有天生tm事件(包括gen1和gen2)共有的psim1278构筑体、使用来源于冻干的天生tm参考材料的标准曲线检测马铃薯和马铃薯产物中的天生tm产物。另外,提供了一组引物和探针,其在<0.1%gmo的检测极限下独特地、定性地检测各事件。另外,所述方法允许使用qpcr检测所有gen2天生tm事件(即w8、x17和y9)共有的psim1678构筑体、使用来源于冻干的天生tm参考材料的标准曲线检测马铃薯和马铃薯产物中的天生tm产物。实例19:内部使用食物混合物验证所述方法制备多种食物混合物以所研发的dna分离方法。我们的内部测试集中于从来自大西洋j3(薯片变种)或褐色兰吉尔f10(炸薯条变种)的所有食物基质中分离dna。共同之处是,这两种事件涵盖考虑中的所有基质(块茎、炸薯条、薯片和薄片)。将磨碎的天生tm食物产品混入如表14中所述的商业变种食物产品中。预期所有事件的结果类似。表14.所产生的食物混合物品系食物产品天生tm食物百分比j3冻干的块茎0.2/0.5/1.0f10冻干的块茎0.2/0.5/1.0f10冻干的炸薯条0.2/0.5/1.0j3薯片5.0/10.0f10薄片2.5/8.0/15.0使用来自qiagen的qiaamp粪便dna快速提取小型试剂盒,对各种食物混合物进行两次到三次独立的dna分离,如实例12中所述。表15展示了600mg炸薯条、薄片或薯片混合物或300mg块茎中的总体dna浓度和产量。为了确保dna的品质足以用于qpcr,使用适当的引物和探针组、三重复进行各dna分离。各反应使用2μl的各dna分离且内源参考基因aprt用作阳性对照。表15.针对各种食物类型获得的平均dna浓度和产量在每种上述食物产品中,dna分离方法产生的dna的浓度和产量足以用于后续的qpcr分析。后续qpcr分析的结果可以发现于图7中。结合在此供参考本文中所引用的所有参考文献、论文、公开、专利、专利公开以及专利申请全部结合在此供参考以用于所有目的。然而,提及本文引用的任何参考文献、论文、公开、专利、专利公开以及专利申请不是并且不应该认为是承认或以任何形式暗示其构成有效的现有技术或形成世界上任何国家的公共常识的一部分。应了解上述说明仅代表说明性实施例和实例。为了读者方便,上述说明集中于所有可能实施例的有限数目个代表性实例、教导本发明原理的实例。所述说明未试图穷尽性枚举所有可能的变化形式或甚至那些所述变化形式的组合。那些替代实施例可以不是作为本发明的特定部分来呈现,或那些未进一步描述的替代实施例可以作为一部分利用,不是认为不承认那些替代实施例。一般技术人员将了解,那些未描述的实施例中有许多涉及技术和材料的差异,而非本发明原理应用上的差异。因此,本发明不希望局限于比以下权利要求书和等效物中所述的范围更小的范围。参考文献奥尔·查瓦拉r.(allchawla,r.),沙基亚r.(shakya,r.)和罗门斯c.m.(rommens,c.m.),(2012),天冬酰胺合成酶-1的块茎特异性静默使田间种植的马铃薯的丙烯酰胺形成潜在性减少而不影响块茎形状和产量(tuberspecificsilencingofasparaginesynthetase-1reducestheacrylamide-formingpotentialofpotatoesgrowninthefieldwithoutaffectingtubershapeandyield),《植物生物技术杂志(plantbiotechnologyjournal)》10,913-924.加巴里诺j.e.(garbarino,j.e.)和贝尔纳普w.r.(belknap,w.r.),(1994),从马铃薯中分离泛素-核糖体蛋白质基因(ubi3)和其启动子在转基因植物中的表达(isolationofaubiquitin-ribosomalproteingene(ubi3)frompotatoandexpressionofitspromoterintransgenicplants),《植物分子生物学(plantmolecularbiology)》24,119-127.加巴里诺j.e.(garbarino,j.e.),乌苏密t.(oosumi,t.)和贝尔纳普w.r.(belknap,w.r.),(1995),聚泛素启动子的分离和其在转基因马铃薯植物中的表达(isolationofapolyubiquitinpromoteranditsexpressionintransgenicpotatoplants),《植物生理学(plantphysiol.)》109,1371-1378.辛普森j.(simpson,j.),蒂姆科m.p.(timko,m.p.),卡西莫a.r.(cashmore,a.r.),斯切尔j.(schell,j.),凡梦古m.(vanmontagu,m.)和赫莱娜-俄斯特娜l.(herrera-estralla,l.),(1985),嵌合基因在豌豆叶绿素a/b-结合蛋白基因的5'-侧接序列的控制下的光诱导性和组织特异性表达(light-inducibleandtissue-specificexpressionofachimaericgeneundercontrolofthe5'-flankingsequenceofapeachlorophylla/b-bindingproteingene),《欧洲分子生物学杂志(emboj.)》4,2723-2729斯米戈奇ac(smigockiac),欧文斯ld(owensld),(1988),与强启动子融合的细胞分裂素基因增强了芽器官形成和所转化植物细胞中的玉米素含量(cytokiningenefusedwithastrongpromoterenhancesshootorganogenesisandzeatinlevelsintransformedplantcells),《美国国家科学院院刊(pnatlacadsciusa)》85:5131-5135.凡哈仁m.j.j.(vanhaaren,m.j.j.),舍蒂n.j.a(sedee,n.j.a),德波尔h.a.(deboer,h.a.),斯奇普特r.a.(schilperoort,r.a.)和胡卡斯p.j.j.(hooykaas,p.j.j.),(1989),农杆菌膨大真管螺的t-区域边界重复序列的保守域的突变分析(mutationalanalysisoftheconserveddomainsofat-regionborderrepeatofagrobacteriumtumafaciens),《植物分子生物学(plantmolecularbiology)》13,523-531.序列表<110>杰.尔.辛普洛公司叶劲松杰弗里·w·哈比希雅内·莱恩杰弗里·w·海因马修·g·彭斯斯特凡妮·胡敦<120>事件特异性检测方法<130>jrsi-062/02us317577-2415<150>us62/118,320<151>2015-02-19<150>us62/062,324<151>2014-10-10<160>90<170>patentinversion3.5<210>1<211>40<212>dna<213>人工序列<220><223>psim1278构筑体接合点lb合成-lb马铃薯<400>1tggcaggatatataccggtgtaaacgaagtgtgtgtggtt40<210>2<211>40<212>dna<213>人工序列<220><223>psim1278构筑体接合点rb合成-rb马铃薯<400>2gtttacagtaccatatatcctgtcagaggtatagaggcat40<210>3<211>36<212>dna<213>人工序列<220><223>psim1278构筑体接合点lb-agp<400>3ttgtggaggagtaagggtaccaagtgtctgagacaa36<210>4<211>30<212>dna<213>人工序列<220><223>psim1278构筑体接合点agp-asn<400>4aacaagcttgttaacgaactctttatccag30<210>5<211>30<212>dna<213>人工序列<220><223>psim1278构筑体接合点asn-ppo<400>5tggtgagccctcgagataattgtaactgat30<210>6<211>36<212>dna<213>人工序列<220><223>psim1278构筑体接合点ppo-间隔子1<400>6attcaatagagactatctagagtgtatgggtgatcc36<210>7<211>30<212>dna<213>人工序列<220><223>psim1278构筑体接合点间隔子1-ppo<400>7ttatggaggaatcagtagtctctattgaat30<210>8<211>30<212>dna<213>人工序列<220><223>psim1278构筑体接合点ppo-asn<400>8atcagttacaattattctcgagggctcacc30<210>9<211>36<212>dna<213>人工序列<220><223>psim1278构筑体接合点asn-gbss<400>9ctggataaagagttcgaattcgtgatgtgtggtcta36<210>10<211>46<212>dna<213>人工序列<220><223>psim1278构筑体接合点gbss-agp<400>10tgagatgcatggttcactagtgattggtaccaagtgtctgagacaa46<210>11<211>30<212>dna<213>人工序列<220><223>psim1278构筑体接合点agp-phl<400>11aacaagcttgttaacgtgctctctatgcaa30<210>12<211>30<212>dna<213>人工序列<220><223>psim1278构筑体接合点phl-r1<400>12ttttgctaaaacatcggacacgtatatttt30<210>13<211>36<212>dna<213>人工序列<220><223>psim1278构筑体接合点r1-间隔子2<400>13atatttattatataactgcagcgttgttttgatgaa36<210>14<211>30<212>dna<213>人工序列<220><223>psim1278构筑体接合点间隔子2-r1<400>14ttttgatgaaaaagcttatataataaatat30<210>15<211>30<212>dna<213>人工序列<220><223>psim1278构筑体接合点phl-gbss<400>15ttgcatagagagcaccgtgatgtgtggtct30<210>16<211>39<212>dna<213>人工序列<220><223>psim1278构筑体接合点gbss-rb<400>16aaatcataatatctggagctcatcgttatgctataaatt39<210>17<211>30<212>dna<213>人工序列<220><223>e12特异性接合点lj-植物-插入序列<400>17tattggtgagaataataaacgaagtgtgtg30<210>18<211>30<212>dna<213>人工序列<220><223>e12特异性接合点rj插入序列-植物<400>18acggtcagtcactttgtactagtaaagatc30<210>19<211>36<212>dna<213>人工序列<220><223>f10特异性接合点lj-植物-插入序列<400>19cagcagccatcaactggtccagcaggatatataccg36<210>20<211>30<212>dna<213>人工序列<220><223>f10特异性接合点rj插入序列-植物<400>20gtgggcatcaatttagtccaccagccaaaa30<210>21<211>35<212>dna<213>人工序列<220><223>j3特异性接合点lj-植物-插入序列<400>21aacaggacaaccacaagctaggaaactcacatgct35<210>22<211>30<212>dna<213>人工序列<220><223>j3特异性接合点rj插入序列-植物<400>22aaaaagccctagaattaattttttttggaa30<210>23<211>30<212>dna<213>人工序列<220><223>j3特异性内部接合点<400>23gaactgaaaccgatatacaaaatggtataa30<210>24<211>30<212>dna<213>人工序列<220><223>j3特异性内部接合点<400>24cctctatacctctgacagaggtatagaggc30<210>25<211>30<212>dna<213>人工序列<220><223>j55特异性接合点lj-植物-插入序列<400>25aaatcaacaagcaattgaagaactacctat30<210>26<211>30<212>dna<213>人工序列<220><223>j55特异性接合点rj插入序列-植物<400>26cataagtgagactatgcgtgaaactttcag30<210>27<211>30<212>dna<213>人工序列<220><223>j55特异性内部接合点<400>27cctctatacctctgataaattaaactgact30<210>28<211>30<212>dna<213>人工序列<220><223>j55特异性内部接合点<400>28actaaaaatctcagctaaaacaataaaaat30<210>29<211>30<212>dna<213>人工序列<220><223>v11特异性接合点lj植物-插入序列<400>29tcatctttcattccggtgtaaacgaagtgt30<210>30<211>30<212>dna<213>人工序列<220><223>v11特异性接合点rj插入序列-植物<400>30atgcctctatacctctgataataaagaact30<210>31<211>30<212>dna<213>人工序列<220><223>w8特异性接合点lj植物-psim1678插入序列<400>31caacaattaaaaaaagcctaattttcatgg30<210>32<211>30<212>dna<213>人工序列<220><223>w8特异性接合点rjpsim1678插入序列-植物<400>32acggtcagtcactttcaagccctaggagcc30<210>33<211>30<212>dna<213>人工序列<220><223>w8特异性接合点lj植物-psim1278插入序列<400>33agccctacaaaaggcccaaactctaagtca30<210>34<211>30<212>dna<213>人工序列<220><223>w8特异性接合点rj植物-psim1278插入序列<400>34agtcctagaactacgaaaattatattcggt30<210>35<211>30<212>dna<213>人工序列<220><223>w8特异性内部接合点<400>35atgcctctatacctcagaggtatagaggca30<210>36<211>30<212>dna<213>人工序列<220><223>w8特异性内部接合点<400>36tgcctctatacctctgacgcaacacaacac30<210>37<211>30<212>dna<213>人工序列<220><223>x17特异性接合点lj植物-插入序列<400>37gtattcgctgcaggatatataccggtgtaa30<210>38<211>30<212>dna<213>人工序列<220><223>x17特异性接合点rj插入序列-植物<400>38gcctctatacctctgactggaaaattgacg30<210>39<211>30<212>dna<213>人工序列<220><223>y9特异性接合点lj植物-插入序列<400>39caaaatgtgcaccaatgatgtctcacgacc30<210>40<211>30<212>dna<213>人工序列<220><223>y9特异性接合点rj插入序列-植物<400>40ctcaacaaccagtcaccggaagtcccttaa30<210>41<211>30<212>dna<213>人工序列<220><223>y9特异性内部接合点<400>41atgcctctatacctctgaaagtgactgacc30<210>42<211>36<212>dna<213>人工序列<220><223>psim1678构筑体接合点lb-pvnt1<400>42ttgtggaggagtaagggtaccagttatacaccctac36<210>43<211>30<212>dna<213>人工序列<220><223>psim1678构筑体接合点pvnt1-vnt1<400>43accagctaacaaaagatgaattattgtgtt30<210>44<211>30<212>dna<213>人工序列<220><223>psim1678构筑体接合点vnt1-tvnt1<400>44tcacaggtactataaataattatttacgtt30<210>45<211>36<212>dna<213>人工序列<220><223>psim1678构筑体接合点tvnt1-pagp<400>45agcctcttttcaaaggggccccaagtgtctgagaca36<210>46<211>36<212>dna<213>人工序列<220><223>psim1678构筑体接合点pagp-inv<400>46aacaagcttgttaacggatccacattcctcccggat36<210>47<211>36<212>dna<213>人工序列<220><223>psim1678构筑体接合点inv-inv<400>47tcaaggactttagaggaattcagcggacccagtcca36<210>48<211>36<212>dna<213>人工序列<220><223>psim1678构筑体接合点inv-pgbss<400>48gaggaatgtgtaatgactagtcgtgatgtgtggtct36<210>49<211>20<212>dna<213>人工序列<220><223>腺嘌呤磷酸核糖基转移酶aprt正向引物<400>49gaaccggagcaggtgaagaa20<210>50<211>20<212>dna<213>人工序列<220><223>腺嘌呤磷酸核糖基转移酶aprt反向引物<400>50gaagcaatcccagcgatacg20<210>51<211>17<212>dna<213>人工序列<220><223>腺嘌呤磷酸核糖基转移酶aprt探针<220><221>misc_feature<222>(1)..(1)<223>可以经6-fam6-羧基荧光素部分标记<220><221>misc_feature<222>(4)..(4)<223>可以是锁核苷酸<220><221>misc_feature<222>(7)..(7)<223>可以是锁核苷酸<220><221>misc_feature<222>(10)..(10)<223>可以是锁核苷酸<220><221>misc_feature<222>(13)..(13)<223>可以是锁核苷酸<220><221>misc_feature<222>(17)..(17)<223>可以被bhq1黑洞淬灭剂1部分标记<400>51cgcctcatgatccgatt17<210>52<211>18<212>dna<213>人工序列<220><223>e12正向引物<400>52gggagtaggctttatacc18<210>53<211>24<212>dna<213>人工序列<220><223>e12反向引物<400>53ccttgaatgatttgatataagaag24<210>54<211>18<212>dna<213>人工序列<220><223>e12探针<220><221>misc_feature<222>(1)..(1)<223>可以经6-fam6-羧基荧光素部分标记<220><221>misc_feature<222>(5)..(5)<223>可以是锁核苷酸<220><221>misc_feature<222>(8)..(8)<223>可以是锁核苷酸<220><221>misc_feature<222>(11)..(11)<223>可以是锁核苷酸<220><221>misc_feature<222>(14)..(14)<223>可以是锁核苷酸<220><221>misc_feature<222>(18)..(18)<223>可以被bhq1黑洞淬灭剂1部分标记<400>54tcagtcactttgtactag18<210>55<211>19<212>dna<213>人工序列<220><223>f10正向引物<400>55aaggcaatctttcataagc19<210>56<211>18<212>dna<213>人工序列<220><223>f10反向引物<400>56cacacacttcgtttacac18<210>57<211>17<212>dna<213>人工序列<220><223>f10探针<220><221>misc_feature<222>(1)..(1)<223>可以经6-fam6-羧基荧光素部分标记<220><221>misc_feature<222>(4)..(4)<223>可以是锁核苷酸<220><221>misc_feature<222>(7)..(7)<223>可以是锁核苷酸<220><221>misc_feature<222>(10)..(10)<223>可以是锁核苷酸<220><221>misc_feature<222>(13)..(13)<223>可以是锁核苷酸<220><221>misc_feature<222>(17)..(17)<223>可以被bhq1黑洞淬灭剂1部分标记<400>57tatatcctgctggacca17<210>58<211>19<212>dna<213>人工序列<220><223>j3正向引物<400>58ccggtactcaaatttgcaa19<210>59<211>23<212>dna<213>人工序列<220><223>j3反向引物<400>59ggtgttcttaatatcgagtgttc23<210>60<211>19<212>dna<213>人工序列<220><223>j3探针<220><221>misc_feature<222>(1)..(1)<223>可以经6-fam6-羧基荧光素部分标记<220><221>misc_feature<222>(4)..(4)<223>可以是锁核苷酸<220><221>misc_feature<222>(7)..(7)<223>可以是锁核苷酸<220><221>misc_feature<222>(10)..(10)<223>可以是锁核苷酸<220><221>misc_feature<222>(13)..(13)<223>可以是锁核苷酸<220><221>misc_feature<222>(19)..(19)<223>可以被bhq1黑洞淬灭剂1部分标记<400>60ccacaagctaggaaactca19<210>61<211>23<212>dna<213>人工序列<220><223>j55正向引物<400>61catggaattatttagaaacaaaa23<210>62<211>20<212>dna<213>人工序列<220><223>j55反向引物<400>62gacgatcaagaatctcaata20<210>63<211>19<212>dna<213>人工序列<220><223>j55探针<220><221>misc_feature<222>(1)..(1)<223>可以经6-fam6-羧基荧光素部分标记<220><221>misc_feature<222>(4)..(4)<223>可以是锁核苷酸<220><221>misc_feature<222>(7)..(7)<223>可以是锁核苷酸<220><221>misc_feature<222>(10)..(10)<223>可以是锁核苷酸<220><221>misc_feature<222>(13)..(13)<223>可以是锁核苷酸<220><221>misc_feature<222>(19)..(19)<223>可以被bhq1黑洞淬灭剂1部分标记<400>63tcatagtcaaacagtcagc19<210>64<211>18<212>dna<213>人工序列<220><223>v11正向引物<400>64gtgggcatcaatttagtg18<210>65<211>20<212>dna<213>人工序列<220><223>v11反向引物<400>65ggagagtaatagccttagta20<210>66<211>19<212>dna<213>人工序列<220><223>v11探针<220><221>misc_feature<222>(1)..(1)<223>可以经6-fam6-羧基荧光素部分标记<220><221>misc_feature<222>(7)..(7)<223>可以是锁核苷酸<220><221>misc_feature<222>(10)..(10)<223>可以是锁核苷酸<220><221>misc_feature<222>(13)..(13)<223>可以是锁核苷酸<220><221>misc_feature<222>(16)..(16)<223>可以是锁核苷酸<220><221>misc_feature<222>(19)..(19)<223>可以被bhq1黑洞淬灭剂1部分标记<400>66cctctatacctctgataat19<210>67<211>18<212>dna<213>人工序列<220><223>v11正向引物<400>67tagccaagcatgacttac18<210>68<211>18<212>dna<213>人工序列<220><223>v11反向引物<400>68cacacacttcgtttacac18<210>69<211>18<212>dna<213>人工序列<220><223>v11探针<220><221>misc_feature<222>(1)..(1)<223>可以经6-fam6-羧基荧光素部分标记<220><221>misc_feature<222>(4)..(4)<223>可以是锁核苷酸<220><221>misc_feature<222>(7)..(7)<223>可以是锁核苷酸<220><221>misc_feature<222>(10)..(10)<223>可以是锁核苷酸<220><221>misc_feature<222>(13)..(13)<223>可以是锁核苷酸<220><221>misc_feature<222>(18)..(18)<223>可以被bhq1黑洞淬灭剂1部分标记<400>69atgtatgaatggcagaag18<210>70<211>18<212>dna<213>人工序列<220><223>w8正向引物<400>70aggctttataccgagttg18<210>71<211>22<212>dna<213>人工序列<220><223>w8反向引物<400>71tcagcttaatcgaataagaaac22<210>72<211>17<212>dna<213>人工序列<220><223>w8探针<220><221>misc_feature<222>(1)..(1)<223>可以经6-fam6-羧基荧光素部分标记<220><221>misc_feature<222>(4)..(4)<223>可以是锁核苷酸<220><221>misc_feature<222>(7)..(7)<223>可以是锁核苷酸<220><221>misc_feature<222>(10)..(10)<223>可以是锁核苷酸<220><221>misc_feature<222>(13)..(13)<223>可以是锁核苷酸<220><221>misc_feature<222>(17)..(17)<223>可以被bhq1黑洞淬灭剂1部分标记<400>72actacggtcagtcactt17<210>73<211>18<212>dna<213>人工序列<220><223>x17正向引物<400>73gtccgaaggttgagaaaa18<210>74<211>20<212>dna<213>人工序列<220><223>x17反向引物<400>74cgtgagacatcatcataatg20<210>75<211>17<212>dna<213>人工序列<220><223>x17探针<220><221>misc_feature<222>(1)..(1)<223>可以经6-fam6-羧基荧光素部分标记<220><221>misc_feature<222>(4)..(4)<223>可以是锁核苷酸<220><221>misc_feature<222>(7)..(7)<223>可以是锁核苷酸<220><221>misc_feature<222>(10)..(10)<223>可以是锁核苷酸<220><221>misc_feature<222>(13)..(13)<223>可以是锁核苷酸<220><221>misc_feature<222>(17)..(17)<223>可以被bhq1黑洞淬灭剂1部分标记<400>75cgcttcagttacggctt17<210>76<211>25<212>dna<213>人工序列<220><223>y9正向引物<400>76gacagctatcaatatactttagtaa25<210>77<211>19<212>dna<213>人工序列<220><223>y9反向引物<400>77ttgccatactcatacagag19<210>78<211>19<212>dna<213>人工序列<220><223>y9探针<220><221>misc_feature<222>(1)..(1)<223>可以经6-fam6-羧基荧光素部分标记<220><221>misc_feature<222>(4)..(4)<223>可以是锁核苷酸<220><221>misc_feature<222>(7)..(7)<223>可以是锁核苷酸<220><221>misc_feature<222>(10)..(10)<223>可以是锁核苷酸<220><221>misc_feature<222>(13)..(13)<223>可以是锁核苷酸<220><221>misc_feature<222>(19)..(19)<223>可以被bhq1黑洞淬灭剂1部分标记<400>78aatgatgtctcacgaccaa19<210>79<211>19<212>dna<213>人工序列<220><223>psim1278正向引物<400>79tcaccagtatgactgttta19<210>80<211>18<212>dna<213>人工序列<220><223>psim1278反向引物<400>80ctaccacacactctctag18<210>81<211>19<212>dna<213>人工序列<220><223>psim1278探针<220><221>misc_feature<222>(1)..(1)<223>可以经6-fam6-羧基荧光素部分标记<220><221>misc_feature<222>(4)..(4)<223>可以是锁核苷酸<220><221>misc_feature<222>(7)..(7)<223>可以是锁核苷酸<220><221>misc_feature<222>(10)..(10)<223>可以是锁核苷酸<220><221>misc_feature<222>(13)..(13)<223>可以是锁核苷酸<220><221>misc_feature<222>(19)..(19)<223>可以被bhq1黑洞淬灭剂1部分标记<400>81aagcttgttaacgaactct19<210>82<211>18<212>dna<213>人工序列<220><223>psim1278正向引物<400>82gcatcatccatcaaagtg18<210>83<211>21<212>dna<213>人工序列<220><223>psim1278反向引物<400>83gtcggtataataagagaagga21<210>84<211>19<212>dna<213>人工序列<220><223>psim1278探针<220><221>misc_feature<222>(1)..(1)<223>可以经6-fam6-羧基荧光素部分标记<220><221>misc_feature<222>(4)..(4)<223>可以是锁核苷酸<220><221>misc_feature<222>(7)..(7)<223>可以是锁核苷酸<220><221>misc_feature<222>(10)..(10)<223>可以是锁核苷酸<220><221>misc_feature<222>(13)..(13)<223>可以是锁核苷酸<220><221>misc_feature<222>(19)..(19)<223>可以被bhq1黑洞淬灭剂1部分标记<400>84tagagactatctagagtgt19<210>85<211>18<212>dna<213>人工序列<220><223>psim1678正向引物<400>85gaacggagggagtatgaa18<210>86<211>19<212>dna<213>人工序列<220><223>psim1678反向引物<400>86ccactttcagttttggttg19<210>87<211>17<212>dna<213>人工序列<220><223>psim1678探针<220><221>misc_feature<222>(1)..(1)<223>可以经6-fam6-羧基荧光素部分标记<220><221>misc_feature<222>(5)..(5)<223>可以是锁核苷酸<220><221>misc_feature<222>(8)..(8)<223>可以是锁核苷酸<220><221>misc_feature<222>(11)..(11)<223>可以是锁核苷酸<220><221>misc_feature<222>(14)..(14)<223>可以是锁核苷酸<220><221>misc_feature<222>(17)..(17)<223>可以被bhq1黑洞淬灭剂1部分标记<400>87tcttttcaaaggggccc17<210>88<211>18<212>dna<213>人工序列<220><223>psim1678正向引物<400>88gaagcctcttttcaaagg18<210>89<211>18<212>dna<213>人工序列<220><223>psim1678反向引物<400>89acgggttaatcgataacc18<210>90<211>16<212>dna<213>人工序列<220><223>psim1678探针<220><221>misc_feature<222>(1)..(1)<223>可以经6-fam6-羧基荧光素部分标记<220><221>misc_feature<222>(4)..(4)<223>可以是锁核苷酸<220><221>misc_feature<222>(7)..(7)<223>可以是锁核苷酸<220><221>misc_feature<222>(10)..(10)<223>可以是锁核苷酸<220><221>misc_feature<222>(13)..(13)<223>可以是锁核苷酸<220><221>misc_feature<222>(16)..(16)<223>可以被bhq1黑洞淬灭剂1部分标记<400>90ccccaagtgtctgaga16当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1
- 用于治疗HBV感染和相关病症的三特异性结合分子的制造方法与工艺
- 对磺胺类药物具有特异性的固相微萃取纤维的制备方法与流程
- 一种检测猪初乳中抗PEDV特异性IgA抗体的夹心ELISA试剂盒的制造方法与工艺
- 用于施用CD19xCD3双特异性抗体的给药方案的制造方法与工艺
- 双特异性抗‑VEGF/抗‑ANG‑2抗体及其在治疗眼血管疾病中的应用的制造方法与工艺
- 特异性人EWS‑FLI1融合蛋白抗体的制备方法及在制备尤文肉瘤诊断抗体试剂的应用与制造工艺
- 双特异性四价抗体及其制造和使用方法与制造工艺
- 利用特异性挥发物筛选昆虫相关嗅觉蛋白的方法与制造工艺
- 使用与表面偶联的单特异性四聚抗体复合物组合物从样品中分离出靶实体的方法与制造工艺
- 抗-WT1/HLA双特异性抗体的制造方法与工艺