稳定表达牛胰蛋白酶原的MDCK细胞系及其用途的制作方法
本发明涉及生物技术领域,具体地,本发明涉及工程细胞系克隆MDCK-S.pro-try及其构建方法,以及所述细胞系在培养流感病毒、制备流感疫苗、抗流感病毒药物筛选、中和抗体测定或者流感其他相关研究等方面的应用。
背景技术:
流感疫苗是目前预防流感发生和传播最为有效的手段。现行市场上人流感疫苗多数以鸡胚为基质进行生产,包含甲型H1N1、H3N2和乙型流感病毒株。传统的鸡胚生产系统已有近70年的历史,疫苗的免疫效果已得到充分证实,但其自身仍存在一些不足之处,比如培养的流感病毒易发生抗原变异,与人群流行株不能完全匹配;残留卵清蛋白易引起过敏反应;尤其当出现大流行时健康鸡胚供应受限不能满足短时间内对疫苗的大量需求等。而哺乳动物细胞培养系统相较鸡胚具有明显的优势,能够避免这些问题,成为未来疫苗生产的发展趋势,1995年WHO建议开展以哺乳动物细胞为生产基质的研究。
目前已探索适用的哺乳动物细胞包括非洲绿猴肾细胞(African green monkey kidney cell,Vero)、犬肾上皮细胞(Madin-Darby canine kidney cells或Madin-Daby canine kidney cell,MDCK)以及人胚胎成视网膜细胞PER.C6。其中MDCK细胞容易培养,胰酶耐受性强,对多种不同流感病毒株易感,可以产生较高病毒滴度,是流感疫苗生产的理想细胞系。MDCK细胞来源的流感疫苗已经获得欧洲药品管理局(European Medicines Agency,EMA)的批准。临床试验表明使用MDCK等细胞生产的流感疫苗是安全的并有高免疫原性,然而较低 的病毒滴度是其大规模生产的瓶颈之一。在生产体系的复杂环境中造成低滴度原因是多方面的,单从细胞基质的角度来讲,这种细胞本身并不表达裂解HA的蛋白酶类,成为病毒产量低的一种限制因素。
流感病毒血凝素HA是病毒主要表面抗原之一,在识别并结合靶细胞受体、介导膜融合、诱导保护性中和抗体方面发挥重要作用。在病毒复制过程中,HA蛋白首先以前体HA0形式合成,经宿主蛋白酶裂解为成熟的HA1和HA2后,才能够介导病毒囊膜与靶细胞膜融合,起始病毒生命周期,因此HA的裂解是病毒感染细胞的首要前提,而MDCK细胞本身并不具备裂解HA的蛋白酶类,研究证明外加一定量的胰蛋白酶能够加速病毒胞间传播,提高病毒滴度,加速病毒多周期生长,现已成为流感病毒细胞培养的常规操作。
胰蛋白酶属于丝氨酸蛋白酶家族,其C端长220aa的丝氨酸肽酶功能域能够特异水解人流感病毒或低致病性禽流感病毒HA0,从而成熟的HA1、HA2发挥作用使病毒基因组释放入细胞内,加速病毒多周期生长,但同时外加的胰酶也加大细胞培养的难度,提高了疫苗生产成本。
在自然状态下人呼吸道上皮细胞中,真正发挥病毒活化作用的蛋白酶种类及作用亚细胞部位一直是研究热点,尚未定论。现已知与胰蛋白酶同家族的多种丝氨酸蛋白酶具有裂解HA的功能,并且作用部位可能位于细胞膜上、胞外和胞内转运过程,然而更为进一步的深入研究却缺失。
技术实现要素:
本发明第一方面,提供一种稳定分泌表达牛胰蛋白酶原的修饰的MDCK细胞系,该细胞系含有整合入其细胞基因组的完整的分泌型牛胰蛋白酶原S.pro-try的编码基因。
在一个具体实施方式中,所述细胞基因组含有分泌型牛胰蛋白酶原编码基因,核苷酸序列如Genbank LOC780933所示。
在一个具体实施方式中,所述分泌型牛胰蛋白酶原的核酸编码序列进行适用于犬类密码子使用的优化。
优选的,所述分泌型牛胰蛋白酶原的核酸编码序列如SEQ ID No.1所示:
ATGAAGACCTTCATCTTCCTGGCCCTGCTGGGCGCCGCCGTGGCCTTCCCCGTGGACGACGACGACAAGATCGTGGGCGGCTACACCTGCGGCGCCAACACCGTGCCCTACCAGGTGAGCCTGAACAGCGGCTACCACTTCTGCGGCGGCAGCCTGATCAACAGCCAGTGGGTGGTGAGCGCCGCCCACTGCTACAAGAGCGGCATCCAGGTGAGGCTGGGCGAGGACAACATCAACGTGGTGGAGGGCAACGAGCAGTTCATCAGCGCCAGCAAGAGCATCGTGCACCCCAGCTACAACAGCAACACCCTGAACAACGACATCATGCTGATCAAGCTGAAGAGCGCCGCCAGCCTGAACAGCAGGGTGGCCAGCATCAGCCTGCCCACCAGCTGCGCCAGCGCCGGCACCCAGTGCCTGATCAGCGGCTGGGGCAACACCAAGAGCAGCGGCACCAGCTACCCCGACGTGCTGAAGTGCCTGAAGGCCCCCATCCTGAGCGACAGCAGCTGCAAGAGCGCCTACCCCGGCCAGATCACCAGCAACATGTTCTGCGCCGGCTACCTGGAGGGCGGCAAGGACAGCTGCCAGGGCGACAGCGGCGGCCCCGTGGTGTGCAGCGGCAAGCTGCAGGGCATCGTGAGCTGGGGCAGCGGCTGCGCCCAGAAGAACAAGCCCGGCGTGTACACCAAGGTGTGCAACTACGTGAGCTGGATCAAGCAGACCATCGCCAGCAACTAA(SEQ ID No.1)。
在一个具体实施方式中,所述细胞系命名为MTY6,保藏在中国微生物菌种保藏管理委员会普通微生物中心,地址在北京市朝阳区北辰西路1号院中科院微生物研究所,其菌种保藏编号为:CGMCC No.12681,保藏日期为2016年7月6日。
本发明第二方面,提供一种构建上述修饰的MDCK细胞系的方法,所述方法包括将编码S.pro-try的开放阅读框cDNA序列与真核表达载体连接,再将所述真核表达载体转染到MDCK细胞系中,经抗生素加压筛选获得稳定表达的细胞系。
在一个具体实施方式中,所述真核表达载体是pcDNA3.1。
在一个具体实施方式中,将分泌型、胞内型和跨膜型酶原或酶的共6种表达方式的目的序列分别与真核表达载体连接,并瞬时转染入MDCK细胞系中,接种人流感病毒比较产量差异,筛选提高病毒滴度的最佳表达模式。
在一个优选的实施方式中,将分泌型牛胰蛋白酶原的真核表达载体连续加压筛选,结合有限稀释法获得稳定表达单克隆细胞。
更优选的,所述连续加压筛选是在高浓度的G418条件下筛选。
更进一步,所述G418的浓度为1000μg/ml。
在一个优选的实施方式中,将分泌型牛胰蛋白酶原的核酸编码序列进行适用于犬类密码子使用的优化。
优选的,所述分泌型牛胰蛋白酶原的核酸编码序列如SEQ ID No.1所示。
更优选的,所述修饰的MDCK细胞系为MTY6,保藏在中国微生物菌种保藏管理委员会普通微生物中心,地址在北京市朝阳区北辰西路1号院中科院微生物研究所,其菌种保藏编号为:CGMCC No.12681,保藏日期为2016年7月6日。
本发明第三方面,提供一种上述修饰的MDCK细胞系的应用,其中,所述应用包括将上述修饰的MDCK细胞系用于培养流感病毒,制备流感疫苗,抗流感药物筛选,中和抗体测定或者流感其他相关研究等领域的应用。
优选的,所述流感为人流感。
本发明的第四方面,提供一种培养流感病毒的方法,其中,所述方法包括利用上述修饰的MDCK细胞系对流感病毒进行培养。
在一个具体实施方式中,上述方法中的流感为人流感。
在一个具体实施方式中,上述方法包括,将人流感病毒H1、H3亚型及B型毒株接种于本发明构建的上述的修饰的MDCK细胞系中。
在一个具体实施方式中,上述方法包括,病毒在所述细胞上的培养条件为37℃、4%CO2、无血清培养48-72h,收获病毒上清液即得流感病毒液。
优选的,所述方法中的增殖人流感病毒的步骤中不需要添加TPCK-胰酶。
优选的,所述方法中的增殖人流感病毒的步骤中也可以进一步添加TPCK-胰酶。
本发明的第五方面,提供一种制备流感疫苗的方法,其中,所述方法包括利用上述修饰的MDCK细胞系对流感病毒进行培养。
在一个具体实施方式中,上述方法中的流感为人流感。
在一个具体实施方式中,上述方法包括,将人流感病毒H1、H3亚型及B型毒株接种于本发明构建的上述的修饰的MDCK细胞系中。
在一个具体实施方式中,上述方法包括,病毒在所述细胞上的培养条件为37℃、4%CO2、无血清培养48-72h,收获病毒上清液即得流感病毒液。
优选的,所述方法中的增殖人流感病毒的步骤中不需要添加TPCK-胰酶。
优选的,所述方法中的增殖人流感病毒的步骤中也可以进一步添加TPCK-胰酶。
本发明采取基因工程手段,模拟自然状态下蛋白酶分泌、胞内和跨膜三种亚细胞分布,将胰蛋白酶基因导入MDCK细胞,接种不同型或亚型人流感病毒,比较病毒产量筛选出分泌型酶原为提升病毒产量的最佳作用模式,并连续加压筛选出能够稳定自主表达的细胞系,一方面用于疫苗规模化生产,简化生产工艺,更加有效地提升病毒产量,为将来大规模流感疫苗生产奠定细胞基础,解决疫苗产能扩大和品质提高的关键问题;另一方面也可以作为实验室有效分离培养和监测流感病毒、抗流感病毒药物的筛选、中和抗体的测定等其他方面的研究工具,具有广泛的应用前景。其中筛选得到的稳定分泌表达牛胰蛋白酶原的MDCK细胞系培养流感病毒生产滴度高,尤其对于B/黑龙江呼兰/116/2010流感病毒的培养滴度,能达到普通MDCK细胞外加TPCK-胰酶培养的病毒滴度,因此利用本发明修饰的MDCK细胞系可以简化病毒培养工艺。
附图说明
图1为分泌、胞内和跨膜型酶原和酶重组质粒瞬时转染MDCK的表达鉴定。
+:TPCK处理的牛胰蛋白酶;-:转染空质粒的细胞样品;S.pro-try:分泌型酶原;S.try:分泌型酶;T.pro-try:跨膜型酶原;T.try:跨膜型酶;C.pro-try:胞内型酶原;C.try:胞内型酶。
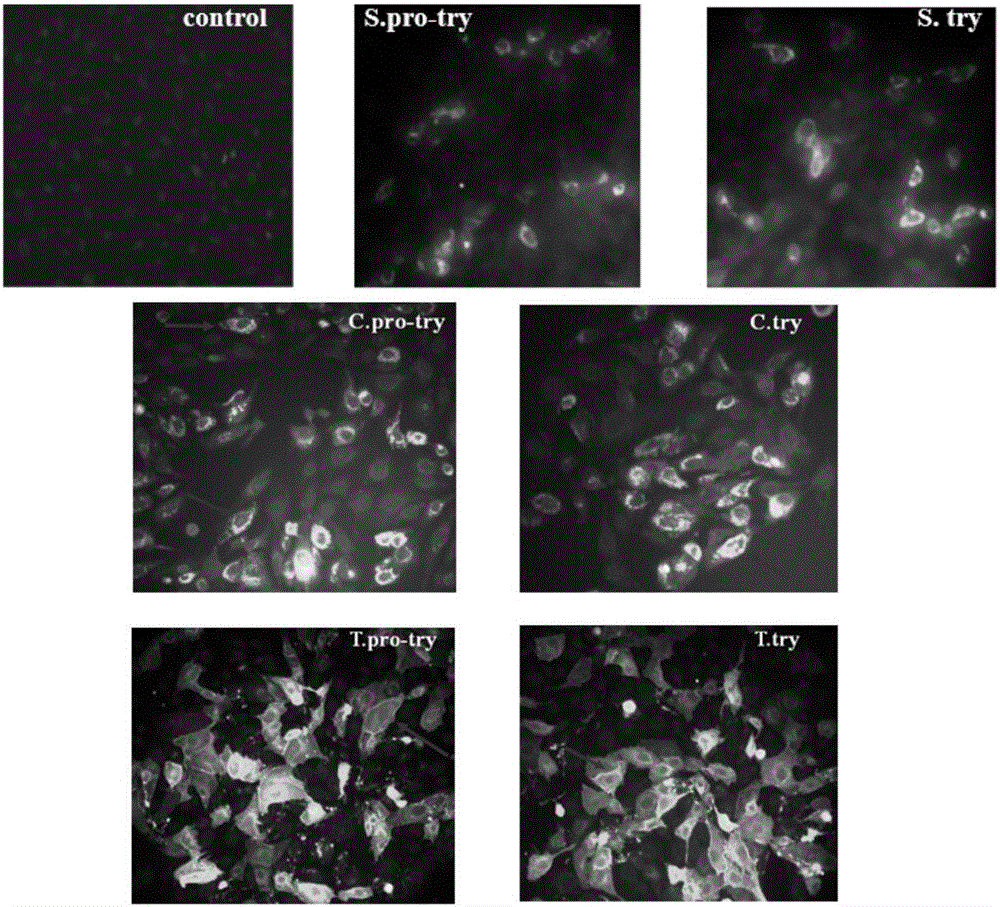
图2为间接免疫荧光鉴定重组质粒瞬时转染MDCK的表达情况。
图3为瞬时转染表达胰酶活性检测。
A:胰酶活性标准曲线;B:6种质粒瞬时转染MDCK表达胰酶活性。
图4为瞬时表达各蛋白酶对流感病毒增殖的影响。
图5为单克隆细胞目的基因的PCR及RT-PCR鉴定。
A:提取基因组DNA的PCR鉴定;B:提取细胞RNA的RT-PCR鉴定。
图6为单克隆细胞株MT21的鉴定。
A:间接免疫荧光鉴定;B:免疫印迹鉴定;C:胰酶活性测定;
D:不同代次单克隆细胞的目的基因的PCR及RT-PCR鉴定;
E:不同代次单克隆细胞的胰酶活性测定。
图7为不同流感病毒株在单克隆细胞的生长情况。
A:检测A/天津津南/15/2009(H1N1)的NA活性及CCID50滴度;
B:检测A/福建同安/196/2009(H3N2)的NA活性及CCID50滴度;
C:检测B/黑龙江呼兰/116/2010的NA活性及CCID50滴度;
D:检测A/California/7/2009(H1N1)的NA活性及CCID50滴度;
图8为优化后单克隆细胞MTY6与MT21表达酶活性比较。
图9为不同病毒在MTY6与MT21的生长情况比较。
A:减毒活疫苗株A1/17/California/2009/38的NA活性检测;
B:减毒活疫苗株A3/Switzerland/9715293/2013-CDC-LA10A的NA活性检测。
具体实施方式
下面结合具体实施例来进一步描述本发明,本发明的优点和特点将会随着描述而更为清楚。但这些实施例仅是范例性的,并不对本发明的范围构成任何限制。本领域技术人员应该理解的是,在不偏离本发明的精神和范围下可以对本发明技术方案的细节和形式进行修改或替换,但这些修改和替换均落入本发明的保护范围内。实施例中所使用到的试剂,除非利用说明,否则为市场上可购得的常规试剂。
本实施例的材料来源:
1、细胞:MDCK购自ATCC,由中国食品药品检定研究院呼吸道病毒疫苗室保存。
2、病毒:A/天津津南/15/2009(H1N1)、A/福建同安/196/2009(H3N2)、B/黑龙江呼兰/116/2010,由中国医学科学院医学生物学研究所廖国阳教授惠赠。甲型流感疫苗病毒株A/California/7/2009(H1N1)由中国食品药品检定研究院呼吸道病毒疫苗室保存。
实施例1不同表达形式的重组牛胰酶基因真核表达载体的构建及瞬时转染功能分析
(1)载体构建:以GenBank提供的牛胰蛋白酶cDNA序列(LOC780933)为模板,设计不同的特异引物(表1和2),扩增得到分泌型酶原表达序列,在此基础上,分别去除N端分泌信号肽扩增得到胞内酶原表达序列,添加HAT跨膜功能域扩增得到跨膜酶原序列;并分别在酶原序列基础上,去除N端APFDDDDK活化肽对应碱基序列扩增得到各自酶表达序列。上游引物引入Nhe I(斜体下滑线所示)、下游引物引入EcoR I(斜体下滑线所示)单一酶切位点,同时在上游引物起始密码子ATG的N端添加Kozak序列(方框标示)提高表达 效率。50μl PCR反应体系中模板5μl、灭菌双蒸水32.8μl,10×PCR缓冲液5μl,dNTP Mix(10mmol/L)1μl,10μmol/L上游引物2μl,10μmol/L下游引物2μl,Taq DNA聚合酶(Invitrogen)0.2μl。PCR扩增条件:94℃预变性1min,94℃变性30s,55℃退火30s,72℃延伸1min,共30个循环,最后72℃保温10min。将6种扩增片段双酶切方式连接入pcDNA3.1载体中,经转化挑选单克隆菌落进行双酶切和测序鉴定,结果序列大小符合预期,各基因读码框正确。
表1.分泌型、胞内型胰蛋白酶原和酶的扩增引物
表2.跨膜型胰蛋白酶原和酶的扩增引物
(2)瞬时转染:对数生长期的MDCK细胞,消化后吹打成单细胞悬液,调整浓度为1×105细胞/ml,接种6孔培养板,每孔2ml,放置37℃、4%CO2培养箱24h。按照LP2000转染试剂操作说明,以1:2.5的最优比例制备脂质体-DNA复合物;待细胞达90%汇合度时逐滴加入,37℃、4%CO2培养6h后换无血清SFM培养液,继续培养48h,分别收集分泌表达酶原和酶的细胞上清液,分泌表达型的细胞以及胞内、跨膜表达细胞分别加入2ml预冷的含0.5%TritonX-100的PBS裂解液,冰浴或4℃作用30min,反复吹打2-3次,5000rpm 4℃低速离心5min去除细胞碎片,吸取上清置于冰上。所有收集样品使用Millipore 3K超滤管4℃5000×g超滤浓缩20min,-70℃保存。
(3)免疫印迹鉴定表达:收集各样品每孔上样20μl,稳压120V进行SDS-PAGE电泳,转PVDF膜后使用PBS(pH7.4)溶解5%脱脂奶37℃低速摇床封闭膜1h,以兔抗牛胰蛋白酶多克隆抗体为一抗、羊抗兔IgG抗体为二抗进行免疫印迹鉴定,结果(图1)三种细胞分布的蛋白均有特异性亮带。分泌型酶原上清和细胞样品的条带较亮,约为24-25kD;分泌型酶上清条带比细胞中明显,分子大小与对照相当;跨膜表达蛋白大小接近30kD,酶原比酶稍大;胞内分布的酶原和酶与分泌型表达相当,约24-25kD。
(4)间接免疫荧光表达定位鉴定:于12孔细胞板各孔中央放置洁净载玻片,接种MDCK细胞1×105/孔,制备细胞爬片;放37℃、4%CO2培养达80%汇合时,分别瞬时转染6种质粒;含2.5%FBS的DMEM继续培养48h,37℃预热的1×PBS(pH7.4)清洗3次;每孔加2ml 4%多聚甲醛室温固定30min;分泌型和胞内型的细胞每孔添加2ml 0.1%TritonX-100,室温通透20min;1ml PBS配制的2%BSA,37℃封闭1h;兔抗牛胰蛋白酶多克隆抗体为一抗,每孔1ml,37℃孵育1h;FITC标记的鼠抗兔IgG单克隆抗体,37℃避光孵育1h;0.5μg/ml的DAPI室温避光染核5min;避光条件下倒置荧光显微镜观察并拍照。结果(图2)分泌型表达的酶原和酶绿色荧光信号明显,主要集中于蓝色荧光标记的细胞核周围;胞内表达的细胞绿色荧光散布于胞核周边,趋向于均质,酶原与酶荧光亮度相当;跨膜型细胞外层边缘亮度聚集,参照自然光镜下同一视野细胞形态,基本可以判定绿色荧光聚集于细胞膜上,各型表达目的蛋白的亚细胞分布符合预期。
(5)胰酶活性检测:以商品化已知酶活性的TPCK-trypsin为阳性标准品, 选择灵敏度高的胰酶特异荧光多肽底物Boc-Leu-Gly-Arg-AMC,参照Eva 的测定方法,每孔添加50mmol/L pH=7.9的Tris-HCl缓冲液进行2倍系列稀释的TPCK-trypsin以及终浓度为25μmol/L的荧光反应底物各50μl,混匀后置于35℃避光反应30min,荧光酶标仪读取各孔值,激发波长355nm,发射波长440nm。以阳性标准品的各稀释度对应酶活力为横坐标,以相应的平均荧光值为纵坐标,绘制酶活性标准曲线,同样方法检测6种表达蛋白样品及对照,代入标准曲线求得表达胰酶活性。结果(图3)分泌型胰酶原活性最高,达10.72×10-2U/ml,相当于实验室使用MDCK培养流感病毒时外加0.5μg/ml TPCK-trypsin活性的1/60,而分泌型上清胰蛋白酶活性与胞内表达的酶原和酶活性相当,约为6×10-2U/ml,跨膜表达的酶原和酶活性最低约为3×10-2U/ml。
(6)对病毒生长的影响:取对数生长期的MDCK细胞参照上述方法瞬时转染6种表达质粒,转染6h后换无血清的DMEM培养基,37℃培养12h后按照moi等于0.001接种A/California/7/2009(H1N1)流感病毒疫苗株,混匀后置37℃4%CO2孵箱待病毒生长。每天镜下观察病毒生长情况,至72h时反复冻融细胞2次使病毒释放,4℃5000rpm离心10min去除细胞碎片,进行病毒NA活性测定,96孔荧光酶标板中每孔MES缓冲液系列稀释样品及NA底物各50μl,混匀室温反应30min,150μl终止液终止反应,荧光酶标仪读取各孔荧光值,激发波长355nm,发射波长460nm,每样品重复3次,数据统计分析。结果(图4)显示分泌型表达胰酶原实验组的病毒NA活性与空转对照组有统计差异(P<0.05),其中,分泌表达胰酶原实验组的病毒NA活性为1340,而空转对照组仅为924,实验组约是对照组的1.5倍;其余5种表达质粒瞬转MDCK培养甲流病毒量与空转对照组相当约为949~1000。因此我们选择表达活性最高的分泌型酶原质粒进行后续稳定转染的细胞筛选。
实施例2 稳定表达分泌型胰酶原MDCK单克隆细胞的筛选及鉴定
(1)筛选培养基中合适的G418浓度的选择:MDCK细胞接种至24孔板 中待汇合度70%时加入G418,使浓度依次为0μg/ml、200μg/ml、400μg/ml、600μg/ml、800μg/ml、900μg/ml、1000μg/ml、1100μg/ml,每浓度复孔,置于37℃、5%CO2培养箱中培养,每日观察记录细胞死亡情况,每隔3-4天换液,培养两周后绘制细胞生存曲线确定G418最小致死浓度,结果当G418浓度在800-900μg/ml之间时,细胞于14天全部死亡,因此选择1000μg/ml为实验中MDCK细胞压力筛选培养基中G418的浓度,维持液浓度为500μg/ml。
(2)G418加压筛选及有限稀释法筛选单克隆细胞:接种约2x105个MDCK细胞至6孔板,培养至细胞密度约为90%时用于转染。24小时后消化细胞,1:10的比例传代至96孔板中,每孔加200μ1 1000μg/ml G418的5%胎牛血清DMEM培养液继续培养,每3-5天换液,持续筛选2周后,可见有抗性细胞生长。对照MDCK细胞不进行空质粒转染,仅使细胞在含有G418的培养液中培养至全部死亡时,选择实验组生长状态良好的抗性细胞集落,消化后继续用含有1000μg/ml G418的压力筛选培养液稀释至0.5细胞/100μl,接种于96孔板中,每孔200μl。继续G418筛选2周后,使用RT-PCR(提取基因组DNA、细胞RNA及RT-PCR方法分别参照选用试剂盒)及胰酶活性检测方法(前面已述)选择表达水平最高、细胞状态较好的克隆再次按有限稀释法进行单细胞克隆筛选。筛选出的细胞胰酶消化后依次于96孔、24孔和6孔细胞培养板扩大培养并按常规方法冻存。结果共得到5株阳性单克隆抗性细胞株,RT-PCR鉴定结果均能扩增出符合预期大小的750bp左右的特异条带,测定序列与目的基因比对一致。结合胰酶活性测定,从中选择了表达酶活性最高、细胞状态较好的21#细胞株进行后续实验,命名为MDCK-S.pro-try(21),简称MT21。
(3)MT21细胞株的鉴定:将单克隆细胞连续传代,提取不同代次细胞基因组DNA进行分泌型牛胰酶原目的基因PCR鉴定,上游引物为Fs:5'-TCCGGCTAGCGCCACCATGAAGACCTTC-3'(SEQ ID No.13)、下游引物为R:5'-GCCGGAATTCTTAGTTGGAGGCGATGG-3'(SEQ ID No.14),使用GAPDH基因作为内参,设计特异上游引物F:5'-GTGATGCTGGTGCTGAGTATGTT-3'(SEQ ID No.15),下游引物R:5'-GGTCTTCTGGGTGGCAGTGAT-3'(SEQ ID No.16),反应体系50μl:Premix Ex TaqTM 2×PCR Solution25μl、模板5μl、引物各2μl、去离子水15μl。扩增条件:94℃预变性1min,94℃变性30s,55℃退火30s,72℃延伸1min,共20个循环,最后72℃保温10min。提取细胞RNA进行RT-PCR鉴定,引物同上,10μl反应体系:RT Master Mix2μl、模板2μl、DEPC处理的水6μl。RT反应条件:37℃15min;85℃5s;以反转录cDNA为模板进行PCR反应体系及条件同上。收集各代细胞表达上清及细胞裂解液,分别进行免疫印迹、间接免疫荧光鉴定,并检测表达上清胰酶活性(方法同实例1)。结果(图5、6):MT21经鉴定证明分泌型胰酶原基因已成功整合至MDCK基因组中,并能正确转录(参见图5);在普通显微镜下观察MT21与正常MDCK细胞未见明显的形态差异,间接免疫荧光显示MT21可见明显绿色荧光,而对照组细胞呈阴性(参见图6A);Western Blot鉴定MT21有单一清晰的特异条带,大小约为24kD,并且上清中条带亮度明显高于细胞裂解液而正常MDCK对照没有任何条带(参见图6B);胰酶活性测定结果上清中胰酶活性较高,约为20×10-2U/ml,细胞中胰酶活性约为17×10-2U/ml,相当于外加0.5μg/ml TPCK-trypsin活性的1/30(参见图6C);单克隆细胞株连续传代至第15代,经PCR及RT-PCR鉴定证实各代细胞间目的基因遗传稳定,检测表达上清胰酶活性没有统计差异,约为20×10-2U/ml,目的蛋白表达稳定(参见图6D和6E,)。
实施例3 流感病毒在稳定表达牛胰酶原的MDCK细胞上的增殖
将3株野生流感病毒A/天津津南/15/2009(H1N1)、A/福建同安/196/2009(H3N2)、B/黑龙江呼兰/116/2010和1株甲型流感毒株分别采用鸡胚尿囊腔对病毒进行大量扩增,收获病毒尿囊液离心后分装,-70℃保存备用。为便于比较,实验设置单克隆细胞MT21实验组(MT21)、正常MDCK对照组(control)、MT21外加0.5μg/mlTPCK-trypsin实验组(MT21+)、正常MDCK 外加0.5μg/mlTPCK-trypsin对照组(control+)。单克隆细胞MT21和正常MDCK分别接种6孔细胞培养板至90%汇合度时,吸去细胞培养液,使用无菌PBS清洗细胞3遍去除FBS,更换含1%双抗的无血清DMEM 1.5mL 37℃孵箱培养24h,待单克隆细胞酶原表达累积。24h后按照moi等于0.001接种各亚型流感病毒,外加胰酶组同时添加终浓度为0.5μg/mlTPCK-胰酶,、每组每孔500μl混匀后置37℃4%CO2孵箱培养。分别于24h,48h,72h时反复冻融细胞2次使病毒释放,4℃5000rpm离心10min去除细胞碎片,收集上清病毒液,分别检测病毒NA活性及CCID50滴度。
结果(图7)所测试的4个毒株中,3株野生病毒对MT21细胞更加敏感,NA活性及病毒滴度均高于在MDCK细胞上测得的结果,尤其是B型病毒株不需外加TPCK处理的胰蛋白酶培养30h便达到与正常MDCK添加0.5μg/mlTPCK-trypsin时一致的病毒产量,而MT21表达上清中胰蛋白酶活性仅为外加0.5μg/mlTPCK-trypsin的1/30,可见构建的稳定细胞系MT21较正常MDCK对流感病毒更加敏感,不同型或亚型作用效果不同,尤其对低生长力的毒株作用显著。另外在外加0.5μg/mlTPCK-trypsin后MT21细胞培养病毒产量显著提升,病毒滴度最高能够达到正常MDCK对照组产量的近4倍,例如,其中接种B型流感病毒株后,MT21细胞培养72h获得病毒滴度为7.9×103TCID50/ml,普通MDCK培养相同时间的病毒滴度为2.0×103TCID50/ml,初步说明我们构建的MDCK-S.pro-try细胞系因分泌表达胰蛋白酶原而比MDCK细胞更适于流感病毒的增殖。
实施例4 密码子优化后单克隆细胞MTY6的鉴定及应用
根据Codon Usage Database数据库提供的canis密码子使用频率表,利用在线密码子优化软件OPTIMIZER,我们将牛胰蛋白酶原基因的编码区的基因序列进行优化。优化只改变基因序列并不改变氨基酸序列。酶原序列共有741个碱基,246个氨基酸,优化后有135个碱基发生了改变,其中75个碱基发生了转 换(嘌呤-嘌呤,嘧啶-嘧啶),60个碱基发生了颠换(嘌呤-嘧啶)。共有101个氨基酸的密码子发生了改变,密码子改变率为40%。具体序列结构如SEQ ID No.1所示:
ATGAAGACCTTCATCTTCCTGGCCCTGCTGGGCGCCGCCGTGGCCTTCCCCGTGGACGACGACGACAAGATCGTGGGCGGCTACACCTGCGGCGCCAACACCGTGCCCTACCAGGTGAGCCTGAACAGCGGCTACCACTTCTGCGGCGGCAGCCTGATCAACAGCCAGTGGGTGGTGAGCGCCGCCCACTGCTACAAGAGCGGCATCCAGGTGAGGCTGGGCGAGGACAACATCAACGTGGTGGAGGGCAACGAGCAGTTCATCAGCGCCAGCAAGAGCATCGTGCACCCCAGCTACAACAGCAACACCCTGAACAACGACATCATGCTGATCAAGCTGAAGAGCGCCGCCAGCCTGAACAGCAGGGTGGCCAGCATCAGCCTGCCCACCAGCTGCGCCAGCGCCGGCACCCAGTGCCTGATCAGCGGCTGGGGCAACACCAAGAGCAGCGGCACCAGCTACCCCGACGTGCTGAAGTGCCTGAAGGCCCCCATCCTGAGCGACAGCAGCTGCAAGAGCGCCTACCCCGGCCAGATCACCAGCAACATGTTCTGCGCCGGCTACCTGGAGGGCGGCAAGGACAGCTGCCAGGGCGACAGCGGCGGCCCCGTGGTGTGCAGCGGCAAGCTGCAGGGCATCGTGAGCTGGGGCAGCGGCTGCGCCCAGAAGAACAAGCCCGGCGTGTACACCAAGGTGTGCAACTACGTGAGCTGGATCAAGCAGACCATCGCCAGCAACTAA(SEQ ID No.1)
按照实施例1和2中同样的方法,将优化后的牛胰酶原编码基因成功瞬时转入MDCK细胞,经G418加压筛选后得到稳定分泌表达的单克隆细胞系,命名为MTY6。测定表达酶活性前统一对样品进行肠激酶激活处理,之后按照实施例1中胰酶活性检测方法,与优化前的MT21单克隆细胞(约9.37U/ml)相比,目的蛋白表达活性提高约3倍(约29.12U/ml)(图8)。
相同条件下培养流感病毒,MTY6单克隆细胞培养流感病毒的产量能达到普通MDCK的5倍左右,如在不外加胰蛋白酶的条件下,培养H3N2亚型流感病毒72h,MTY6单克隆细胞收获病毒NA活性为830,而正常MDCK细胞培养病毒NA活性值仅为173;相较于优化前MT21单克隆细胞,MTY6对流感病毒更加敏感,尤其是当外加0.5μg/mL TPCK-Trypsin时这种差异更加明显,而无论 是否外加TPCK-Trypsin,MTY6培养病毒产量是MT21的2倍左右(参见图9),当外加胰酶培养H1N1流感病毒时,病毒产量随时间不断累积,当72h时出现明显增长,MTY6细胞的病毒产量NA值约为800,MT21细胞的病毒NA值约为220;当不外加胰酶培养H3N2流感病毒时,同样病毒产量随时间累积,至72h时出现明显增长,MTY6细胞的病毒产量NA值约为180,MT21细胞的病毒NA值约为80。总之我们构建的MTY6细胞系因分泌表达胰蛋白酶原而比MDCK细胞更适于流感病毒的增殖,在细胞流感培养及相关疫苗生产上更具有应用价值。同时也使其成为临床或实验研究中分离、培养流感病毒、监测病毒变异、抗病毒药物筛选、中和抗体的测定等的便捷工具,具有广泛应用前景。
序列表
<110> 中国食品药品检定研究院
<120> 稳定表达牛胰蛋白酶原的MDCK细胞系及其用途
<160> 16
<170> Patent In version 3.3
<210> 1
<211> 741
<212> DNA
<213> 人工序列
<400> 1
atgaagacct tcatcttcct ggccctgctg ggcgccgccg tggccttccc cgtggacgac 60
gacgacaaga tcgtgggcgg ctacacctgc ggcgccaaca ccgtgcccta ccaggtgagc 120
ctgaacagcg gctaccactt ctgcggcggc agcctgatca acagccagtg ggtggtgagc 180
gccgcccact gctacaagag cggcatccag gtgaggctgg gcgaggacaa catcaacgtg 240
gtggagggca acgagcagtt catcagcgcc agcaagagca tcgtgcaccc cagctacaac 300
agcaacaccc tgaacaacga catcatgctg atcaagctga agagcgccgc cagcctgaac 360
agcagggtgg ccagcatcag cctgcccacc agctgcgcca gcgccggcac ccagtgcctg 420
atcagcggct ggggcaacac caagagcagc ggcaccagct accccgacgt gctgaagtgc 480
ctgaaggccc ccatcctgag cgacagcagc tgcaagagcg cctaccccgg ccagatcacc 540
agcaacatgt tctgcgccgg ctacctggag ggcggcaagg acagctgcca gggcgacagc 600
ggcggccccg tggtgtgcag cggcaagctg cagggcatcg tgagctgggg cagcggctgc 660
gcccagaaga acaagcccgg cgtgtacacc aaggtgtgca actacgtgag ctggatcaag 720
cagaccatcg ccagcaacta a 741
<210> 2
<211> 28
<212> DNA
<213> 人工序列
<400> 2
tccggctagc gccaccatga agaccttc 28
<210> 3
<211> 81
<212> DNA
<213> 人工序列
<400> 3
tccggctagc gccaccatga agaccttcat ctttctggct ctcttgggag ccgctgttgc 60
tatcgtgggc ggctacacct g 81
<210> 4
<211> 43
<212> DNA
<213> 人工序列
<400> 4
tccggctagc gccaccatgt tccccgtgga cgatgatgac aag 43
<210> 5
<211> 36
<212> DNA
<213> 人工序列
<400> 5
tccggctagc gccaccatga tcgtgggcgg ctacac 36
<210> 6
<211> 27
<212> DNA
<213> 人工序列
<400> 6
gccggaattc ttagttggag gcgatgg 27
<210> 7
<211> 35
<212> DNA
<213> 人工序列
<400> 7
tccggctagc gccaccatgt ataggccagc acgtg 35
<210> 8
<211> 34
<212> DNA
<213> 人工序列
<400> 8
gtcatcatcg tccacgggga aaaaagctaa aaag 34
<210> 9
<211> 34
<212> DNA
<213> 人工序列
<400> 9
ctttttagct tttttccccg tggacgatga tgac 34
<210> 10
<211> 30
<212> DNA
<213> 人工序列
<400> 10
gtgtagccgc ccacgataaa agctaaaaag 30
<210> 11
<211> 30
<212> DNA
<213> 人工序列
<400> 11
ctttttagct tttatcgtgg gcggctacac 30
<210> 12
<211> 27
<212> DNA
<213> 人工序列
<400> 12
gccggaattc ttagttggag gcgatgg 27
<210> 13
<211> 28
<212> DNA
<213> 人工序列
<400> 13
tccggctagc gccaccatga agaccttc 28
<210> 14
<211> 27
<212> DNA
<213> 人工序列
<400> 14
gccggaattc ttagttggag gcgatgg 27
<210> 15
<211> 23
<212> DNA
<213> 人工序列
<400> 15
gtgatgctgg tgctgagtat gtt 23
<210> 16
<211> 21
<212> DNA
<213> 人工序列
<400> 16
ggtcttctgg gtggcagtga t 21
- 还没有人留言评论。精彩留言会获得点赞!