恶臭假单胞菌ZJPH1412及其制备(S)‑3‑羟基金刚烷甘氨酸的应用的制作方法
本发明涉及一株微生物新菌种—恶臭假单胞菌(Pseudomonas putida)ZJPH1412,以及在催化2-(3-羟基-1-金刚烷)-2-氧代乙酸不对称转氨制备(S)-3-羟基金刚烷甘氨酸中的应用。
(二)
背景技术:
(S)-3-羟基金刚烷甘氨酸的化学结构式为:
(S)-3-羟基金刚烷甘氨酸是用于合成治疗2型糖尿病DPP-4抑制剂药物沙格列汀的重要手性中间体。沙格列汀是一种高效、选择性、可逆性的DPP-4抑制剂。2013年美国临床内分泌医师学会推出的糖尿病治疗方案最新指南中将DPP-4抑制剂作为2型糖尿病单药和联合治疗的首选药物之一。相比于其他DPP-4抑制剂,在相同抑制浓度为1.3nmol/L DPP-4时沙格列汀的作用约是维格列汀或西他列汀的十倍。其主要代谢产物5-羟沙格列汀也是一个选择性DPP-4抑制剂,其活性为沙格列汀的50%。
目前催化2-(3-羟基-1-金刚烷)-2-氧代乙酸制备光学纯的(S)-3-羟基金刚烷甘氨酸的方法主要有化学法和生物法。采用化学法合成时,需要使用金属雷尼镍催化剂,且步骤繁琐,污染大,产率偏低。生物法则是利用游离酶或全细胞来催化制备。可用于制备(S)-3-羟基金刚烷甘氨酸的酶主要有苯丙氨酸脱氢酶或支链转氨酶,苯丙氨酸脱氢酶在催化还原氨化反应过程中需要添加甲酸铵、二硫苏糖醇、辅酶烟酰胺腺嘌呤二核苷酸,但辅酶价格昂贵,且苯丙氨酸脱氢酶的分离纯化步骤复杂,增加了该药物的制备成本。利用支链转氨酶可一步实现氨化反应,但由于存在副产物抑制,以及反应时所发生的平衡逆向移动的限制,工业化应用存在一定的问题。利用全细胞催化法制备可省去复杂的酶分离纯化过程。
目前文献报道的生物催化合成(S)-3-羟基金刚烷甘氨酸主要涉及以下几种微生物或酶。Ronald L.Hanson等(Hanson R L,Goldberg S L,Brzozowski D B,et al.,Adv.Synth.Catal.,2007,349(8-9):1369-1378)从中间型嗜热放线菌(Thermoactinomyces intermedius)中克隆表达苯丙氨酸脱氢酶基因,经修饰后,转入毕赤酵母中表达。该重组毕赤酵母在甲醇存在下生长并产生一种内源性甲酸脱氢酶,该酶可用于辅酶NAD(H)的再生。从毕赤酵母中提取修饰型的苯丙氨酸脱氢酶和内源性甲酸脱氢酶两种纯酶作为生物催化剂,添加甲酸铵、二硫苏糖醇、辅酶烟酰胺腺嘌呤二核苷酸,以2-(3-羟基-1-金刚烷)-2-氧代乙酸为底物直接还原氨化可制得(S)-3-羟基金刚烷甘氨酸,ee值大于99%,转化率大于80%,但该转化过程需提取并利用两种酶催化制备得到产物;Eun young Hong等(Hong E Y,Cha M,Yun H,et al.,J.Mol.Catal.B:Enzym,2010,66(1-2):228-233)从大肠杆菌(Escherichia coli)K12中分别克隆表达支链转氨酶基因和天冬氨酸转氨酶基因获得两种重组工程菌,并利用这两种工程菌全细胞催化制备(S)-3-羟基金刚烷甘氨酸,产率达90.5%,光学活性大于99%;Ju-Yeon Kim等(Kim J-Y,Lee Y-A,Wittmann C,et al.,Biotechnol.Bioeng.,2013,110(11):2846-2855)从大肠杆菌(Escherichia coli)K12中克隆表达支链转氨酶基因,以野生型谷氨酸棒状杆菌(Corynebacterium Glutamicum)为宿主获得重组工程菌,并利用该重组工程菌进行全细胞催化合成L-HAG,反应80h后,转化率为80%。
(三)
技术实现要素:
本发明目的是提供一株微生物新菌种—恶臭假单胞菌(Pseudomonas putida)ZJPH1412,以及在不对称催化2-(3-羟基-1-金刚烷)-2-氧代乙酸转氨制备(S)-3-羟基金刚烷甘氨酸中的应用。
本发明采用的技术方案是:
本发明提供一株新菌株—恶臭假单胞菌(Pseudomonas putida)ZJPH1412,保藏于中国典型培养物保藏中心,保藏日期为2016年4月11日,保藏编号:CCTCC NO:M 2016187,保藏地址:中国,武汉,武汉大学,邮编430072。
此外,本发明还提供一种所述恶臭假单胞菌ZJPH1412在(S)-3-羟基金刚烷甘氨酸制备中的应用,所述的应用为:以2-(3-羟基-1-金刚烷)-2-氧代乙酸为底物,L-谷氨酸为氨基供体,以恶臭假单胞菌ZJPH1412经发酵培养获得的湿菌体为酶源,于pH为9.0的Tris-HCl缓冲液中构成转化体系,在30-45℃(优选40℃)条件下进行反应,反应结束后,将反应液离心,取上清液,得到含产物(S)-3-羟基金刚烷甘氨酸的转化液。所述底物的初始加入终浓度为10~50mmol/L缓冲液(优选20mmol/L),氨基供体浓度为底物初始浓度的2倍(即终浓度为20~100mmol/L缓冲液),湿菌体的用量以菌体湿重计为50~350g/L缓冲液(优选300g/L)。
进一步,优选所述反应是在40℃条件下转化反应2~50h,优选30~42h。
为更进一步提高产率,所述转化体系中还添加支链转氨酶的辅酶即PLP辅酶。所述辅酶加入终浓度为0.01~0.1mmol/L缓冲液(优选0.02mmol/L)。支链转氨酶是PLP(磷酸吡哆醛)辅酶依赖型的酶,在体系中额外添加辅酶PLP有利于转氨反应的进行。
更进一步,优选所述恶臭假单胞菌ZJPH1412在制备(S)-3-羟基金刚烷甘氨酸中的应用按如下步骤进行:以2-(3-羟基-1-金刚烷)-2-氧代乙酸为底物,L-谷氨酸为氨基供体,以恶臭假单胞菌ZJPH1412经发酵培养获得的湿菌体为酶源,加入辅酶,于pH为9.0的Tris-HCl缓冲液中构成转化体系,在40℃条件下反应42h,反应结束后,获得含有目标产物的转化液;所述底物的初始加入终浓度为20mmol/L缓冲液,L-谷氨酸浓度为40mmol/L缓冲液,湿菌体的用量以菌体湿重计为300g/L缓冲液;所述优选辅酶PLP,在转化体系中的添加终浓度为0.02mmol/L缓冲液。
本发明所述酶源按如下方法获得:
1)斜面培养:挑取恶臭假单胞菌ZJPH1412单菌落接种至斜面培养基,37℃培养1d,4℃冰箱保存;斜面培养基终浓度组成为:蛋白胨10g/L,酵母膏5g/L,NaCl 10g/L,琼脂20g/L,溶剂为水,pH 6.5~7.0;
2)种子培养:从培养成熟的斜面挑一环菌体接入种子培养基中,37℃,200rpm培养16h,获得种子液;所述种子培养基终浓度组成为:蛋白胨10g/L,酵母膏5g/L,NaCl 10g/L,溶剂为水,pH 6.5~7.0;
3)发酵培养:以体积浓度10%的接种量将种子液转接到发酵培养基中,37℃,200rpm培养20h,获得发酵液,将发酵液离心,所得沉淀用pH 8.0的K2HPO4-KH2PO4缓冲液洗涤,获得湿菌体,即为酶源;所述发酵培养基终浓度组成为:30g/L麦芽糖,40.04g/L玉米浆干粉,49.83g/L牛肉膏,0.4g/L MgSO4·7H2O,0.11g/L Li2SO4·H2O,溶剂为水,pH 6.5~7.0。
与现有技术相比,本发明的有益效果主要体现在:本发明提供了一株微生物新菌种—恶臭假单胞菌(Pseudomonas putida)ZJPH1412,其可用于催化转氨制备手性非天然氨基酸,从而在微生物法制备2型糖尿病药物沙格列汀关键手性中间体(S)-3-羟基金刚烷甘氨酸方面提供了有益的参考;采用该新菌种催化制备光学纯(S)-3-羟基金刚烷甘氨酸具有立体选择性好,产物光学纯度高等优点;本发明通过采用恶臭假单胞菌ZJPH1412经发酵获得的湿菌体为催化剂的手性生物催化,当底物浓度为20mmol/L时,目的产物(S)-3-羟基金刚烷甘氨酸的ee达到99.9%,产率达到77.3%。
(四)附图说明
图1为2-(3-羟基-1-金刚烷)-2-氧代乙酸底物标准品液相检测图谱。
图2为(S)-3-羟基金刚烷甘氨酸产物标准品经衍生化后的液相检测图谱。
图3为衍生化试剂2,3,4,6-四-O-乙酰基-β-D-吡喃葡萄糖异硫氰酸酯标准品液相色谱图。
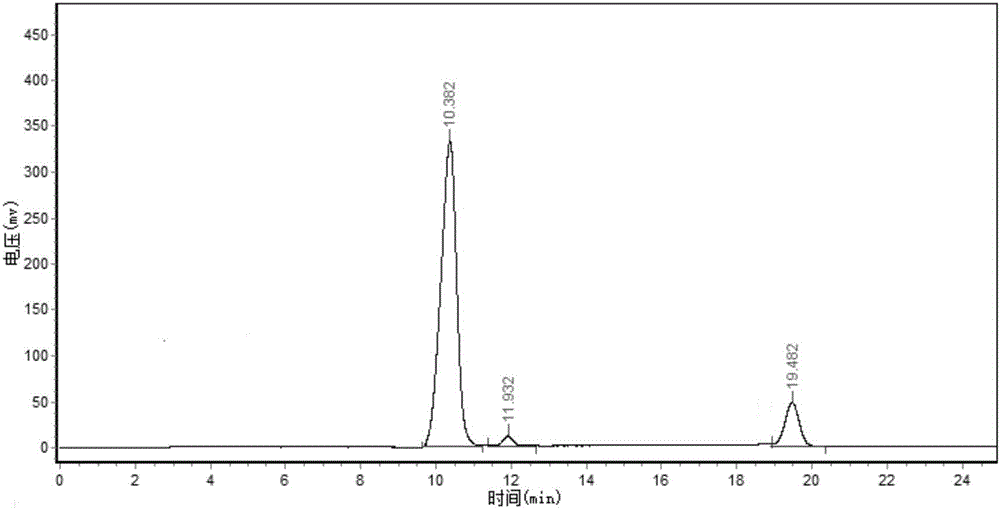
图4为恶臭假单胞菌ZJPH1412转化液的液相色谱图。
(五)具体实施方式
下面结合具体实施例对本发明进行进一步描述,但本发明的保护范围并不仅限于此:
实施例1、菌株筛选与鉴定
1)菌种筛选:
菌种来源:恶臭假单胞菌(Pseudomonas putida)ZJPH1412系从采自沈阳的土样中分离筛选获得。具体筛选方法如下:称1g土样加入到装有50ml无菌水的250ml锥形瓶中,37℃,200rpm,振荡1h,得到土样稀释液。取0.1ml上述稀释液加入到装有10ml富集培养基的50ml锥形瓶中,37℃,200rpm,培养数天,待培养液变浑浊后,取0.2ml培养液加入至新鲜的富集培养基中,同样条件下培养数天,如此重复富集培养3-4次。富集培养基中以L-叔亮氨酸为唯一氮源。富集培养基终浓度组成为:10mmol/L L-叔亮氨酸,100mmol/L甘油,1g/L MgSO4·7H2O,0.2mmol/L CaCl2,0.1mg/L ZnCl2,0.1mg/L MnSO4·4H2O,0.02mg/L H3BO3,0.1mg/L CuSO4·5H2O,0.05mg/L CoCl2,0.1mg/L NiSO4·6H2O,2.0mg/L Na2MoO4,4.0mg/L FeSO4·7H2O,溶剂为0.05mol/L磷酸缓冲液(pH 7.0)。
将富集后的培养液用无菌水稀释至10-5、10-6、10-7,分别取0.2mL涂布筛选平板培养基,37℃培养3d,从平板生长的菌落中挑取形态不同的菌落经平板划线分离得到单菌落。平板培养基组成为富集培养基中加有20g/L的琼脂。
挑取单菌落接种到种子培养基,37℃,200rpm培养24h,取种子液以体积浓度10%的接种量转接至发酵培养基中,37℃,200rpm培养24h,将发酵液离心后,获得湿菌体。种子培养基和发酵培养基配方相同,具体如下:蛋白胨10g/L,酵母膏5g/L,NaCl 10g/L,琼脂20g/L,溶剂为水,pH 6.5~7.0。
以2-(3-羟基-1-金刚烷)-2-氧代乙酸为底物,L-谷氨酸为氨基供体,筛选得到的微生物细胞(即发酵培养后的湿菌体)为催化剂,在pH 8.0的磷酸缓冲液中37℃生物转化48h。将转化液离心后取上清,用薄层色谱法测定转化液中是否有目标产物,并选择显色斑点较深的菌株进行后续复筛。采用高效液相色谱法测定转化液中目标产物(S)-3-羟基金刚烷甘氨酸的对映体过量值(ee值),从而筛选得到具有高对映体过量值(ee值>99%)和较高产率的微生物菌株ZJPH1412。
2)生理生化特征
菌落形态:采用斜面培养基于37℃培养24h,边缘整齐,圆形,隆起,透明。
生理生化特征:革兰氏染色阴性,呈短杆状。可以利用的碳源有:α-D-葡萄糖、D-果糖、D-岩藻糖、甘油、D-果糖-6-磷酸、L-丙氨酸、L-精氨酸、L-天冬氨酸、L-谷氨酸、L-焦谷氨酸、D-半乳糖醛酸、L-半乳糖酸内酯、D-葡糖酸、D-葡糖醛酸、葡糖醛酰胺、粘液酸、奎尼酸、D-砂糖酸、L-乳酸、柠檬酸、α-酮戊二酸、L-苹果酸、γ-氨基丁酸、β-羟基-D,L丁酸、丙酸、乙酸、甲酸。不能利用的碳源有:糊精、D-麦芽糖、明胶、甘氨酰-L-脯氨、D-海藻糖、D-纤维二糖、龙胆二糖、蔗糖、D-松二糖、水苏四糖、D-棉子糖、α-D-乳糖、D-蜜二糖、β-甲基-D-葡萄糖苷、D-水杨苷、N-乙酰-D-葡萄糖胺、N-乙酰-β-D-甘露糖、N-乙酰-D-半乳糖胺、N-乙酰神经氨酸、D-甘露糖、D-半乳糖、3-甲基葡萄糖、L-鼠李糖、肌苷、D-山梨醇、D-甘露醇、D-阿拉伯糖醇、肌醇、甘氨酰-L-脯氨酸、L-组氨酸、果胶、对羟基苯乙酸、D-乳酸甲酯、D-苹果酸、溴丁二酸、吐温40、α-羟基丁酸、α-酮丁酸、乙酰乙酸。
3)16S rDNA序列特性
用Ezup柱式细菌基因组DNA抽提试剂盒提取得到的细胞总DNA为模板,利用通用引物27F(5’-AGAGTTTGATCCTGGCTCAG-3’)和1492R(5’-GGTTACCTTGTTACGACTT-3’)扩增菌株的16S rDNA基因,PCR反应体系为25μl,其中包括基因组DNA 20~25ng 0.5μl,10×Buffer 2.5μl,DNA聚合酶0.2μl,正反向引物10μM各0.5μl,dNTP(各2.5mM)1μl,加双蒸水至25μl;PCR反应条件:94℃预变性4min,然后在94℃保持45s,55℃保持45s,72℃保持1min的条件下进行30个循环,最后在72℃延伸10min,4℃终止反应。经基因组DNA提取和PCR扩增16S rDNA片段后,获得大小为1365bp的DNA片段。将PCR产物纯化后经生工生物工程(上海)股份有限公司测序,得到所述菌株的16S rDNA基因序列如SEQ ID NO.1所示。
该序列已提交GenBank(GenBank登录号为No.KX094933),将菌株ZJPH1412的16S rDNA序列在NCBI网站(http://www.ncbi.nlm.nih.gov)上进行同源性比对(BLAST),结果表明:菌株ZJPH1412与假单胞菌属(Pseudomonas sp.)的部分菌株序列同源性较高,且与Pseudomonas putida R43菌株(GenBank登录号:No.KC990820.1)的序列同源性达到99.85%。
根据生理生化特性并结合分子生物学鉴定,该菌株ZJPH1412被鉴定为恶臭假单胞菌(Pseudomonas putida),命名为恶臭假单胞菌(Pseudomonas putida)ZJPH1412,保藏于中国典型培养物保藏中心,保藏日期为2016年4月11日,保藏编号:CCTCC NO:M 2016187,保藏地址:中国,武汉,武汉大学,邮编430072。
实施例2:湿菌体的获得
1)斜面培养:挑取恶臭假单胞菌ZJPH1412单菌落接种至斜面培养基,37℃培养1d,4℃冰箱保存;斜面培养基终浓度组成为:蛋白胨10g/L,酵母膏5g/L,NaCl10g/L,琼脂20g/L,溶剂为水,pH 7.0。
2)种子培养:从培养成熟的斜面挑一环菌体接入种子培养基中,37℃、200rpm培养16h,获得种子液;种子培养基配方:蛋白胨10g/L,酵母膏5g/L,NaCl 10g/L,溶剂为水,pH 7.0。
3)发酵培养:以体积浓度10%的接种量将种子液转接到发酵培养基中,37℃、200rpm培养20h,获得发酵液,将发酵液离心,所得沉淀用pH 8.0的K2HPO4-KH2PO4缓冲液洗涤,获得湿菌体,即为酶源。发酵液培养基配方:30g/L麦芽糖,40.04g/L玉米浆干粉,49.83g/L牛肉膏,0.4g/L MgSO4·7H2O,0.11g/L Li2SO4·H2O,溶剂为水,pH 6.5。
实施例3~5:
将实施例2所得的湿菌体悬浮于10mLpH 8.5和pH 9.0的Tris-HCl缓冲液,以及pH 9.5的甘氨酸-NaOH缓冲液中,湿菌体以菌体湿重计浓度为100g/L缓冲液;加入终浓度20mmol/L缓冲液的2-(3-羟基-1-金刚烷)-2-氧代乙酸作为底物,终浓度40mmol/L缓冲液的L-谷氨酸为氨基供体,置于37℃、200rpm的摇床中转化24h,反应结束后,将转化液离心,取上清液进行高效液相色谱分析。高效液相色谱条件:日本岛津SPD-10A高效液相色谱仪,浙大N2000色谱工作站,C18色谱柱(250×4.6mm,5μm),检测波长为254nm;进样量为10μL。流动相为甲醇-0.1%三氟乙酸水溶液(58:42,v/v);柱温为30℃;流速为0.8ml/min。根据高效液相色谱检测谱图,计算得出转化液中产物的浓度和ee值。产物(S)-3-羟基金刚烷甘氨酸的浓度和ee值见表1所示。由表1可知,当缓冲液的pH为9.0时,(S)-3-羟基金刚烷甘氨酸的浓度最高,为5.4mmol/L,ee值为99.9%,产率为27.0%。
上清液中的产物和未反应底物的浓度采用高效液相色谱分析。产物需经柱前衍生化反应后再进行检测。衍生化条件为:取200μL上清液用蒸馏水稀释后与200μL的15mmol/L的三乙胺-乙腈水(乙腈-水体积比1:1)溶液混匀,再加入200μL的12mmol/L的2,3,4,6-四-O-乙酰基-β-D-吡喃葡萄糖异硫氰酸酯乙腈水溶液(乙腈-水体积比1:1),涡旋混匀,35℃反应60min。
产率计算方法为:
标准曲线制备方法:用蒸馏水配制浓度分别为4、8、12、16、20mmol/L的底物溶液及经衍生化的0.01、0.05、0.1、0.2、0.3mmol/L产物标准品溶液,分别用高效液相色谱检测。将所得色谱图积分得到峰面积,以底物或产物峰面积(S底物或S产物)为纵坐标,浓度(C底物或C产物)为横坐标,绘制标准曲线。得到底物标准曲线方程:y=28.069x+9.1232,R2=0.9998;产物标准曲线方程:y=46828x+161.32,R2=0.9959。
产物产率计算公式如下:
公式(1)中Cp为(S)-3-羟基金刚烷甘氨酸浓度,C0为2-(3-羟基-1-金刚烷)-2-氧代乙酸初始浓度。
产物的光学纯度由对映体过量值(enantiomeric excess,ee)来表征。计算公式为:
公式(2)中CS和CR分别为S型和R型的3-羟基金刚烷甘氨酸摩尔浓度。
表1产物的浓度及ee值
实施例6~9:
将实施例2所得的湿菌体悬浮于10mL的Tris-HCl缓冲液(pH 9.0)中,湿菌体以菌体湿重计浓度为100g/L缓冲液;加入终浓度20mmol/L缓冲液的2-(3-羟基-1-金刚烷)-2-氧代乙酸作为底物,终浓度40mmol/L缓冲液的L-谷氨酸为氨基供体系,在不同温度、200rpm的摇床中转化24h,反应结束后,将转化液离心,取上清液进行高效液相色谱分析。产物(S)-3-羟基金刚烷甘氨酸的浓度和ee值见表2所示。由表2可知,当温度为40℃时,(S)-3-羟基金刚烷甘氨酸的浓度最高,为5.62mmol/L,ee值为99.9%,产率为28.1%。
表2产物的浓度及ee值
实施例10~12:
将实施例2所得的湿菌体悬浮于10mL Tris-HCl缓冲液(pH 9.0)中,湿菌体以菌体湿重计浓度为100g/L缓冲液;加入终浓度20mmol/L缓冲液的2-(3-羟基-1-金刚烷)-2-氧代乙酸作为底物,终浓度40mmol/L缓冲液的L-谷氨酸为氨基供体,再加入不同终浓度的PLP,置于40℃,200rpm的摇床中转化24h,反应结束后,将转化液离心,取上清液进行高效液相色谱分析。产物(S)-3-羟基金刚烷甘氨酸的浓度和ee值见表3所示。由表3可知,当PLP终浓度为0.02mmol/L时,(S)-3-羟基金刚烷甘氨酸的浓度最高,为6.14mmol/L,ee值为99.6%,产率为30.7%。
表3产物的浓度及ee值
实施例13~15:
将实施例2所得的湿菌体悬浮于10mL Tris-HCl缓冲液(pH 9.0)中,湿菌体以菌体湿重计浓度为100g/L缓冲液;加入不同终浓度的2-(3-羟基-1-金刚烷)-2-氧代乙酸作为底物,终浓度40mmol/L缓冲液的L-谷氨酸为氨基供体,再加入终浓度为0.02mmol/L缓冲液的PLP,置于40℃,200rpm的摇床中转化24h,反应结束后,将转化液离心,取上清液进行高效液相色谱分析。产物(S)-3-羟基金刚烷甘氨酸的浓度和ee值见表4所示。由表4可知,当底物终浓度为20mmol/L时,(S)-3-羟基金刚烷甘氨酸的浓度最高,为6.34mmol/L,ee值为99.6%,产率为31.7%。
表4产物的浓度及ee值
实施例16~18:
将实施例2所得的不同浓度的湿菌体(以湿重计)悬浮于10mL Tris-HCl缓冲液(pH 9.0)中,加入终浓度20mmol/L缓冲液的2-(3-羟基-1-金刚烷)-2-氧代乙酸作为底物,终浓度40mmol/L缓冲液的L-谷氨酸为氨基供体,再加入终浓度为0.02mmol/L缓冲液的PLP,置于40℃,200rpm的摇床中转化24h,反应结束后,将转化液离心,取上清液进行高效液相色谱分析。产物(S)-3-羟基金刚烷甘氨酸的浓度和ee值见表5所示。由表5可知,当湿菌体量为300g/L时,(S)-3-羟基金刚烷甘氨酸的浓度最高,为10.65mmol/L,ee值为99.5%,产率为53.2%。
表5产物的浓度及ee值
SEQUENCE LISTING
<110> 浙江工业大学
<120> 恶臭假单胞菌ZJPH1412及其制备(S)-3-羟基金刚烷甘氨酸的应用
<130>
<160> 1
<170> PatentIn version 3.5
<210> 1
<211> 1365
<212> DNA
<213> Pseudomonas putida
<400> 1
agtcgagcgg atgacgggaa gcttgcttcc ttgattcagc ggcggacggg tgagtaatgc 60
ctaggaatct gcctggtagt gggggacaac gtttcgaaag gaacgctaat accgcatacg 120
tcctacggga gaaagcaggg gaccttcggg ccttgcgcta tcagatgagc ctaggtcgga 180
ttagctagtt ggtgaggtaa tggctcacca aggcgacgat ccgtaactgg tctgagagga 240
tgatcagtca cactggaact gagacacggt ccagactcct acgggaggca gcagtgggga 300
atattggaca atgggcgaaa gcctgatcca gccatgccgc gtgtgtgaag aaggtcttcg 360
gattgtaaag cactttaagt tgggaggaag ggcagtaagt taataccttg ctgttttgac 420
gttaccgaca gaataagcac cggctaactc tgtgccagca gccgcggtaa tacagagggt 480
gcaagcgtta atcggaatta ctgggcgtaa agcgcgcgta ggtggttcgt taagttggat 540
gtgaaagccc cgggctcaac ctgggaactg catccaaaac tggcgagcta gagtacggta 600
gagggtggtg gaatttcctg tgtagcggtg aaatgcgtag atataggaag gaacaccagt 660
ggcgaaggcg accacctgga ctgatactga cactgaggtg cgaaagcgtg gggagcaaac 720
aggattagat accctggtag tccacgccgt aaacgatgtc aactagccgt tggaatcctt 780
gagattttag tggcgcagct aacgcattaa gttgaccgcc tggggagtac ggccgcaagg 840
ttaaaactca aatgaattga cgggggcccg cacaagcggt ggagcatgtg gtttaattcg 900
aagcaacgcg aagaacctta ccaggccttg acatgcagag aactttccag agatggattg 960
gtgccttcgg gaactctgac acaggtgctg catggctgtc gtcagctcgt gtcgtgagat 1020
gttgggttaa gtcccgtaac gagcgcaacc cttgtcctta gttaccagca cgttatggtg 1080
ggcactctaa ggagactgcc ggtgacaaac cggaggaagg tggggatgac gtcaagtcat 1140
catggccctt acggcctggg ctacacacgt gctacaatgg tcggtacaga gggttgccaa 1200
gccgcgaggt ggagctaatc tcacaaaacc gatcgtagtc cggatcgcag tctgcaactc 1260
gactgcgtga agtcggaatc gctagtaatc gcgaatcaga atgtcgcggt gaatacgttc 1320
ccgggccttg tacacaccgc ccgtcacacc atgggaagtg ggttg 1365
- 还没有人留言评论。精彩留言会获得点赞!