一种玉米II型二酰基甘油酰基转移酶及编码该玉米II型DGAT的基因的制作方法
本发明涉及一种玉米二酰基甘油酰基转移酶及编码该玉米二酰基甘油酰基转移酶的基因。
背景技术:
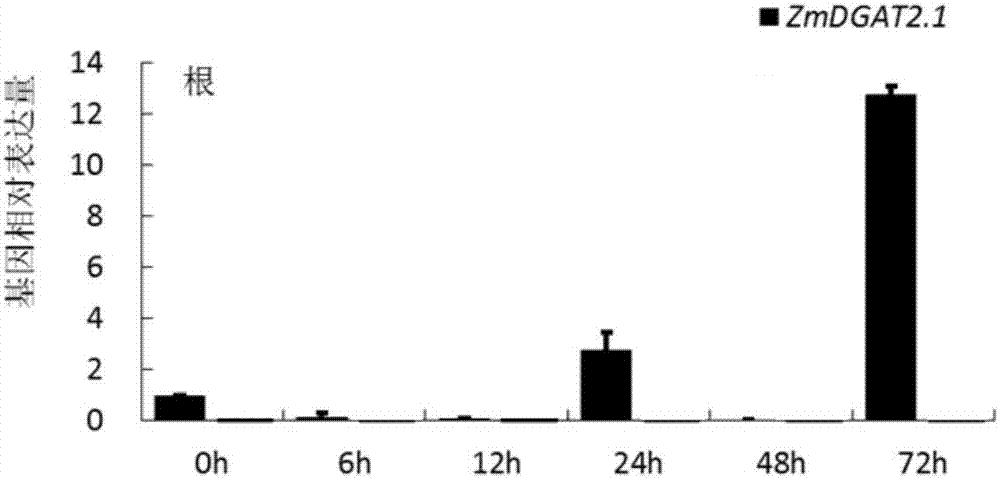
:三酰甘油(triacylglyceroltriacylglycerol,tag)由非极性的甘油和脂肪酸组成,是植物油脂的主要组成部分,是植物体内能量的主要储存形式,同时也参与许多的生理生化过程。植物萌发和幼苗生长的过程中三酰甘油作为碳源和营养物质的主要供应者。在多种植物中的研究表明,三酰甘油tag不但为种子萌发和幼苗生长提供所需能量,在花粉发育和有性生殖过程中也发挥着重要作用。人类日常生活当中的食用油脂和化工产业的用油均以油料作物种子当中所贮存的油类为主要来源。全世界的植物油脂当中大约有75%作为我们日常生活中所用的食用油脂,而且植物油脂包含多数的单不饱和脂肪酸,例如油酸会比饱和脂肪酸的营养价值更高一些。营养学和流行病学的相关研究表明,人们长期的食用富含不饱和脂肪酸的食用油,可增加血液中高密度脂蛋白与低密度脂蛋白的比,从而起到降低人体血液中胆固醇,减少动脉硬化的发生的作用。植物油脂的食用价值之外,植物油脂作为潜在的可再生的生物能源产品,应用前景极其广泛。植物油还是很多化工产品的重要原料,很多的脂肪酸例如月桂酸,斑鸠菊,芥酸在工业生产当中有非常广泛的应用,根据统计,世界上每年应用于化工生产和制药产业当中的植物油量非常巨大;约占植物油脂总产量的1/3。伴随着世界人口的持续增长和经济的飞速发展,使人们对植物油脂的需求量不断提高,这也促使着人们不断的寻找可以增加油料作物产油量的有效方法和便捷的途径。玉米是全世界最重要的粮食作物之一,其品质和产量对全球粮食产业发展具有至关重要的影响。高油玉米和普通玉米相比,具有多方面的优越性。由于玉米油当中含有较高比例的不饱和脂肪酸和维生素等成分,使其具有软化血管和降低血压等作用;高油玉米的蛋白质的含量、赖氨酸含量和类胡萝卜素含也非常的高,因而是一种非常理想的保健用油。玉米油当中的特殊组分和成分不仅对人类和单胃的动物的营养改进具有重要的意义,同时在家禽动物的饲养方面也发挥了重要的作用。其较高的营养成分和重要作用使高油玉米从单纯的粮食或饲料作物逐渐转变成为了兼用作物。与此同时,玉米还有很高的生产潜力,每公顷的高油玉米的产量很可能超过多数的油料作物的产油量,与此同时,高油玉米当中,其除油分以外的产品,无论是数量还是质量都有可能多于其他多数的粮食作物。在我国市场当中销售的玉米油,大多数都是从国外进口的,其高昂的价格导致玉米油在市场竞争中处于劣势。技术实现要素:三酰甘油tag的组装是由kennedy于1961年发现首先发现的,因此被称为kennedy途径。在发育的种子当中,三酰甘油tag以甘油骨架上附连3个脂肪酸的形式而存在。在肯尼迪途径当中,在甘油的sn-1、sn-2和sn-3位点的顺序依次加入脂酰coa最终形成三酰甘油tag:首先,sn-甘油-3-磷酸被甘油-3-磷酸脂酰转移酶(gpat)酰基化,然后由磷脂酸脂酰转移酶(lpaat)催化产生磷脂酸(pa),磷脂酸又被磷脂酸磷酸酯酶(pap)脱磷酸作用生成sn-1,2-甘油二酯(dag),最后由二脂酰甘油酰基转移酶(dgat)酰基化形成tag。三酰甘油生物合成主要在内质网上进行,是膜结合类的催化反应;dgat基因编码的二酰甘油酰基转移酶做为肯尼迪途径之中唯一的一个限速酶;控制着三酰甘油合成的最后一步。因此该基因对提高油料作物的经济效益有重要的作用。为了提高我国的玉米油产量,改善玉米油的市场竞争力,本发明提供了一种玉米ii型二酰基甘油酰基转移酶及编码该玉米ii型dgat的基因。本发明玉米ii型二酰基甘油酰基转移酶命名为zmdgat2.1,zmdgat2.1的氨基酸序列如seqidno:2所示。编码上述玉米ii型二酰基甘油酰基转移酶的基因核苷酸序列如seqidno:1中核苷酸序列所示。本发明提供了一种玉米ii型二酰基甘油酰基转移酶(zmdgat2.1)及其dna片段为调控玉米油脂生成,提高玉米油脂产量和质量奠定了物质基础。本发明有助于揭示玉米dgat基因在玉米生长发育及响应非生物胁迫过程中的作用,深入了解玉米dgat基因的功能,为玉米分子育种提供新的基因资源和理论依据。附图说明图1是具体实施方式一低温处理实验中zmdgat2.1基因在不同时间点叶片中表达分析的实时荧光定量pcr测试图。图2是具体实施方式一低温处理实验中zmdgat2.1基因在不同时间点根系中表达分析的实时荧光定量pcr测试图。图3是具体实施方式一nacl处理实验中zmdgat2.1基因在不同时间点叶片中表达分析的实时荧光定量pcr测试图。图4是具体实施方式一nacl处理实验中zmdgat2.1基因在不同时间点根系中表达分析的实时荧光定量pcr测试图。图5是具体实施方式一nahco3处理实验中zmdgat2.1基因在不同时间点叶片中表达分析的实时荧光定量pcr测试图。图6是具体实施方式一nahco3处理实验中zmdgat2.1基因在不同时间点根系中表达分析的实时荧光定量pcr测试图。图7是具体实施方式一aba处理实验中zmdgat2.1基因在不同时间点叶片中表达分析的实时荧光定量pcr测试图。图8是具体实施方式一aba处理实验中zmdgat2.1基因在不同时间点根系中表达分析的实时荧光定量pcr测试图。图9是具体实施方式一peg处理实验中zmdgat2.1基因在不同时间点叶片中表达分析的实时荧光定量pcr测试图。图10是具体实施方式一peg处理实验中zmdgat2.1基因在不同时间点根系中表达分析的实时荧光定量pcr测试图。图11是具体实施方式一zmdgat2.1基因在不同玉米组织中的qrt-pcr分析结果图。图12是具体实施方式一玉米zmdgat2.1基因酶学功能实验结果图。具体实施方式具体实施方式一:玉米ii型二酰基甘油酰基转移酶命名为zmdgat2.1,zmdgat2.1的氨基酸序列如seqidno:2所示。编码zmdgat2.1的基因核苷酸序列如seqidno:1中核苷酸序列所示。本实施方式zmdgat2.1全长由1314bp组成,含有437个氨基酸,大小为48.6kda,等电点为10.6。zmdgat蛋白的二级结构均包含4种构象,为α-螺旋(alphahelix)、延伸链(extendedstrand)、β-折叠(betaturn)和无规则卷曲(randomcoil),且这4种构象贯穿了整个氨基酸链。zmdgat2.1的α-螺旋包含92个氨基酸,占25.63%;延伸链含有92个氨基酸,占21.05%;β-折叠含有48个氨基酸,占10.98%;无规则卷曲含有185个氨基酸,占42.23%。zmdgat2.1基因的启动子区域包含ltr和myb结合元件;zmdgat2.1包含4个跨膜结构域。本实施方式采用trizol(invitrogen公司)法提取玉米组织部位的总rna,具体步骤如下:a.将研钵及105℃高温2h后的玉米组织,放入-20℃中半小时,向离心管中加入1mltrizolrna提取液,取大约100mg液氮冷冻迅速三次研磨后的玉米组织加入离心管中,颠倒混匀;置于-80℃的冰箱内30min,以使玉米组织部位的rna充分裂解释放。b.将裂解液取出,置于冰上至其融化后,加入200μl的氯仿,充分混匀,静置15min,12000r/min,4℃离心15min;取上清液约300μl移入新的离心管,然后加入等体积的异丙醇,震荡混匀后置于-20℃冰箱内2h,取出后,12000r/min4℃离心15min;c.缓慢倒掉异丙醇,12000r/min4℃离心15min后将液体倒净,可见管底有少量胶状沉淀产生;在沉淀中加入1ml70%的乙醇溶液(含depc)洗涤rna沉淀,将rna沉淀轻轻弹起,充分混匀;d.1200rpm4℃离心10min,倒掉离心管内的乙醇溶液,将离心管置于工作台室温晾数分钟,直到管内乙醇完全挥发;e.用50μl1%depc溶解rna,并用微量分光光度计检测检测rna浓度,利用凝胶电泳检测rna质量。反转录:参照invitrogen公司的试剂说明书进行。反转录得到的cdna用内参基因18srrna进行扩增验证其质量,内参引物序列为:18s-f:agtttgaggcaataacaggtct;18s-r:gatgaaatttcccaagattacc。基因全长pcr扩增:以玉米根系cdna为模板进行pcr扩增,并在c端插入增强子。pcr上游引物f1:5’-agaatggggctggcagtggcgg-3’;下游引物r1:5’-tcaaagaactcttaactgaagatcaggg-3’。pcr反应体系:10×pcrbuffer2μl2mmdntps2μl25mmmgcl21.6μl模板1μlprimerf1μlprimerr1μlkod-plus0.4μlpcr-gradewater11μlpcr反应程序:预变性94℃2min;35个循环:94℃、15s,55℃、30s,68℃、2min。利用基因全长克隆方法,运用相同溶液比例的体系和反应条件,扩大反应体系,进行pcr产物富集,本实施方式采用购自天根生物技术公司的胶回收试剂盒(dp210)进行产物富集。然后测序,获得核苷酸序列如seqidno:1所示。表达载体构建由于kod高保真酶克隆出全长的产物为平末端,连接t载体和酵母表达载体(pyes2.1)的连接方式均是粘性末端连接的方式,因此在构建载体之前对pcr富集产物进行加a,按照以下体系进行混合:pcr产物15μldatp4μltaqdnapolymerase1μl将混合体系置于72℃的条件下反应20~30min。构建表达载体:用克隆到的玉米zmdgat2.1基因的全长编码区构建酵母表达载体在酵母中进行异源表达。玉米zmdgat2.1基因全长cds的pcr产物片段与载体进行连接的方式是通过粘性末端连接。反应体系及反应条件如下:将反应体系温和混合充分置于室温(22~30℃)30min,反应结束立即置于冰上或放于-20℃条件下保存。大肠杆菌转化:a.加入2μl的酵母表达体系混合物于50μl的感受态细胞当中,冰浴30min。b.于42℃水浴当中热激30s,结束后立刻置于冰上。c.加入250μl的室温的soc培养基,置于37℃,200rpm条件下摇1h。d.涂不同体积(10μl,50μl,100μl)的菌液于选择培养基上,37℃条件下过夜培养。e.挑取10个单菌落,利用载体两端引物和基因的引物对菌落进行阳性菌落的验证.将验证成功的阳性菌落进行质粒提取,并进行测序,测序成功进行下一步实验,本实施方式采用购自天根生物技术公司的质粒提取试剂盒(dp105)进行质粒的提取。并且利用载体的正向引物与基因的反向引物,载体的反向引物与基因的正向引物对基因片段插入方向进行验证,并利用基因的正反向引物利用pcr试验方法进一步证实进行验证载体连接是否成功。半乳糖诱导玉米zmdgat2.1基因在酵母当中的表达:由于半乳糖可以诱导包含gal1启动子的目标蛋白的表达,将已经以葡萄糖为碳源生长的菌液离心,去除上清,换新的培养基并加入半乳糖以诱导目标蛋白表达,重组蛋白质可以在半乳糖诱导后不到4h内检测。可以在2h内在含棉子糖由半乳糖诱导培养的细胞中检测到蛋白质的形成。具体操作步骤如下:a.挑取缺陷型酿酒酵母菌株h1246的单菌落,于10ml的ypd培养基当中,30℃,150rpm条件下过夜培养。b.测定过夜培养菌液的od600的值,将过夜培养菌液的od600的值调整为每50ml诱导培养基为0.4,继续在30℃,150rpm条件下摇菌2~4h。c.在2500rpm室温条件下离心使酵母细胞聚于管底,去上清,用40ml1×te重悬细胞,用2ml的1×liac/0.5×te重悬酵母细胞。d.将酵母细胞于室温条件下放置10min。e.每个转化内加入1μgzmdgat2.1/pyes2.1质粒和100μg的变性的鲑鱼精dna和100μl步骤d中的酵母悬浮液。f.在混合液当中加入700μl的1×liac/40%peg-3350/1×te充分混合;将反应体系置于30℃条件下30min后加入88μldmso与反应体系充分混合,在42℃条件下培养7min,离心机瞬离10s,去上清。g.将细胞用1ml的1×te重悬后离心去上清。h.重悬细胞于50~100μl的1×te当中涂板,于30℃条件下进行培养。酶学功能实验:陷型酿酒酵母菌株h1246完全丧失形成脂类的功能,将玉米zmdgat2.1基因转入到缺陷型酿酒酵母菌株h1246菌株中,通过提取转基因菌株的脂类,观察酵母菌株是否具有形成脂类的功能,进而说明玉米zmdgat2.1基因的酶学功能:a.分别挑取玉米zmdgat2.1的转基因菌株的单菌落、拟南芥dgat1基因的转基因菌株的单菌落(阳性对照)、含藻类dgat2基因的转基因菌株的单菌落(阳性对照)、含空载体的转基因菌株的单菌落(阴性对照)、陷型酿酒酵母菌株h1246的单菌落(阴性对照)于5ml的ypd液体培养基当中,30℃,150rpm条件下,过夜培养。b.第二天取一定体积的菌液于室温,2000rpm条件下离心3min,去上清,加入20ml的sc(含葡萄糖和棉籽糖),继续过夜摇菌。c.取一定体积的菌液离心去上清,用1~2ml的sc(含10%棉籽糖和10%半乳糖)冲洗菌液,将菌液加入到50ml的sc(含棉籽糖和半乳糖),摇菌18~36h;离心菌液,去上清,用1ml灭菌的去离子水进行冲洗,离心去上清。d.在收集的菌液当中加入1ml的甲醇溶液,加入0.5ml的玻璃珠,利用细胞破碎仪,力度10,破碎10min。e.将混合液转移到玻璃管当中,加入2ml的氯仿和1ml的氯仿:甲醇(2:1)的混合液,和1ml的0.034%的mgcl2溶液,涡旋震荡充分混匀。f.将体系于室温,2000rpm,室温条件下离心3min。g.取下层有机相于新管当中,加入1ml的2m的kcl:甲醇=4:1的混合液当中,2000rpm,室温条件下离心5min;然后取下层有机相置于一新管中,置于通风橱内利用氮吹仪将其吹干。h.在吹干的管中加入200μl的氯仿:甲醇(2:1)的混合液当中;点样于层析胶板上,半小时后将其置于装有己烷:乙醚:乙酸=70:30:1混合液的层析缸内,跑板45min左右,至层析液距层析板顶端1cm左右处,将板取出,置于通风橱内晾6~8h。i.将充分晾干的层析胶板置于装有固体碘的层析缸内,观看层析条带,获得酶学功能验证结果。实验结果显示,转入了未连接基因片段的空载体的ev(emptyvector)作为阴性对照的菌体当中,未见三酰甘油tag条带生成;作为阳性对照的过表达拟南芥dgat1基因的ad1菌株和过表达藻类dgat2基因的td2菌株,在薄层液相色谱板上可以看到tag条带的形成;过表达玉米zmdgat2.1基因的转基因菌株,出现了清晰可见的tag条带(如图12所示)。因此证明,玉米zmdgat2.1基因具有形成脂类的功能,且弱于拟南芥当中dgat的酶活性,表明玉米zmdgat2.1基因具有在体外催化合成脂类(tag)的功能。非生物胁迫实验:选取玉米自交系合344籽粒饱满,大小一致的种子,用自来水冲洗干净,采用10%naclo3消毒10min,蒸馏水洗净,浸种12h后催芽,将种子放在培养箱中22℃暗培养24h,待种子长出约1.5cm的根后,将种子种在打好孔的泡沫板,悬浮于种植1/2ms营养液当中,在光照培养箱中培养,培养条件为22℃,16h光照/8h黑暗。在玉米幼苗长到三叶一心期,进行非生物胁迫处理,包括低温处理(4℃)、nacl(200mmol·l-1)、nahco3处理(100mmol·l-1)、aba(100μmol·l-1)处理和干旱处理(20%peg6000)。分别在处理后0、12、24、48和72h取样,每种处理选取3棵植株,取根和叶片组织各约200mg,液氮速冻后于-80℃保存。实时荧光定量pcr反应在实时荧光定量pcr仪上进行。反应体系为:实时定量pcr反应程序:预变性95℃3min;40个循环:95℃、60s,95℃、15s,55℃、30s,72℃、60s。每个样品3次重复。用2-δδct方法展开基因的相对表达量分析。上游引物f2:5’-tggcacaccattccccttcc-3’;下游引物r2:5’-ggactcttagatgaaggccaggat-3’。低温处理实验:对玉米自交系合344的根叶进行4℃的低温处理,进行实时定量分析,在玉米的叶部组织和根部组织当中,zmdgat2.1基因表现出上调表达趋势,zmdgat2.1基因在12h的表达量达到峰值,根部组织当中zmdgat2.1基因在各个时间点均上调表达,zmdgat2.1基因在24h和72h上调表达,且在72h达到峰值,在低温的胁迫下的其他时间点呈现下调表达的趋势,zmdgat2.1基因在低温胁迫处理条件下的不同组织部位和不同时期表现出不同的反应,显示了低温胁迫过程的复杂机制(如图1~2所示)。nacl处理实验:玉米的叶片组织和根部组织在nacl的胁迫处理下,在大部分的时间点上,叶片组织和根部组织中的zmdgat2.1基因均有显著上调表达的趋势,其中zmdgat2.1基因呈现先上升后下降的趋势,其表达量在48h达到峰值;在玉米根部组织当中,zmdgat2.1基因在多个时间点呈现上调表达趋势,zmdgat2.1基因呈现波动表达趋势,且在12h的表达量达到了峰值(如图3~4所示)。nahco3处理实验:当玉米的根部组织和叶片组织接受碱胁迫的时候,玉米的zmdgat2.1基因在根和叶中的受诱导机制和程度不同,在叶片组织当中zmdgat2.1基因在48和72h均有上调趋势,但是两个时间点zmdgat2.1基因的表达量存在差异,zmdgat2.1基因在48h和72h的表达量呈现逐渐降低的趋势,72h的表达量约为48h的2倍。在碱胁迫的根部组织当中,zmdgat2.1基因在各时间点呈现上调表达的趋势,zmdgat2.1基因的表达量呈现先生后降的趋势,且在12h时间点表达量达到峰值,显著高于其他时间点上调表达量(如图5~6所示)。aba处理实验:在玉米的叶片组织和根部组织中,经受aba胁迫条件下,zmdgat2.1基因在不同时间点呈现上调表达的趋势,在叶片组织中,受到aba胁迫时,zmdgat2.1基因在12h时间点的上调趋势相比于其他时间点最为明显,在其他时间点表达量较低;且在24h、72h均无上调表达趋势,在24h时间点,zmdgat2.1基因无上调表达趋势,在根部组织当中,zmdgat2.1基因同样被aba胁迫诱导,zmdgat2.1基因在24h表达最高,在6h上调表达量次之,在其他时间点均无上调表达趋势。zmdgat2.1基因在受aba胁迫的条件下受诱导,表明zmdgat2.1基因在参与aba激素胁迫过程当中具有重要的作用(如图7~8所示)。peg处理实验:在玉米的叶片组织和根部组织当中,在经受干旱胁迫下,叶片中zmdgat2.1基因在12和48h有明显上调的趋势,在其他时间点无上调趋势,在根中zmdgat2.1基因的上调明显,且在各个时间点均有上调趋势,zmdgat2.1基因在受胁迫的早期表达量比较高,6h时间点的表达量达到峰值,12h时间点表达量次之,zmdgat2.1基因表达量逐渐降低;zmdgat2.1基因在干旱胁迫当中的表现表明,其在调控作物对干旱逆境环境的响应显著,但响应机制存在差异,对作物抵抗逆境胁迫具有重要作用(如图9~10所示)。不同玉米组织qrt-pcr分析:对玉米不同的生育时期样品进行取样,取样时期分别为幼苗期、拔节期、灌浆前胚乳、大喇叭口期和种子期。样品液氮速冻于-80℃保存,进行rna提取,反转录成cdna,进行qrt-pcr,实验方法同上。实时定量结果显示,玉米dgat2.1基因在灌浆前的胚乳当中表达较高;是幼苗期的28倍;是拔节期的19倍;是大喇叭口期的11倍;是种子时期的24倍(如图11所示)。可见相同基因在不同组织部位的表达水平也存在差异,说明在基因完成其功能的过程当中,是一个极其复杂的过程,其作用机制还有待研究和证实。玉米dgat2.1基因根据玉米数据库下载的cds序列设计全长引物,经过梯度pcr和更换pcr酶等实验方法的尝试并未获得基因全长,因此根据orf(openreadingframe)开放阅读框预测结果,选择第二个翻译起始位点进行基因全长的克隆,克隆出长度为1002bp的基因片段(如seqidno:3所示)。验证结果显示:所选菌落均包含连接成功且方向正确的酵母表达载体。zmdgat2.1验证产物长度为1002bp。选取1-2个阳性zmdgat2.1/pyes2.1酵母表达载体送去测序公司进行测序分析,测序结果正确。经过验证的表达载体转化到陷型酿酒酵母菌株h1246菌株进行异源表达。结果表明,zmdgat2.1基因的表达能够恢复酵母突变体形成脂类的表型,说明zmdgat2.1基因编码的蛋白具有合成脂类的酶学功能。zmdgat2.1在胚乳形成期间发育较高;在本发明的基础上,利用植物中dgat同源基因的时空表达特异性和功能差异,对其进行分别调控,对实现精确调控高等植物种子油脂合成途径具有重要意义。zmdgat2.1对多种非生物胁迫处理有响应,说明其在作物对逆境胁迫的抗性方面有一定的作用,可以丰富作物抗逆育种的基因资源。序列表<110>黑龙江八一农垦大学<120>一种玉米ii型二酰基甘油酰基转移酶及编码该玉米ii型dgat的基因<130>zmdgat2.1<141>2017-06-30<160>9<170>siposequencelisting1.0<210>1<211>1314<212>dna<213>zeamays<400>1atggggctggcagtggcggtgggcaggcggccgacgcagcggtccggtgtctcccgttct60cgctgcgctggcaagagacccactgccgccgagccccgacaccaccggcaccgcacggag120ggaagaaaagaaagaaatcccgggggatcacgtcgttatcagcgaccagattcagacgcg180agccgagcgggggaggcggaccacactagaactagaaccctctctacggctctactacgc240tacgccccatcccgcccggaccggaccggacccaccttcctccacctctctgcccgccgc300cggccgacccgaatgggggcgggaaccaataatggcctgagcaacggcgccgccgcaggg360cagcgcgcggacgacgggaccacggtgttccggggcacggcgtactcgccgctacggacc420acggtggcgctcgcgctgtggctcggggccatccacttcaacgccttcctcgtcctcgcc480tcgctcttcctcttcccgcgccgcgtcgccgcactggtgctggcgacgcagctcttcttc540atgttcctgccgctcagtgataagagcagactgggccgcaagatcgccaggttcataagc600aagtacgtcattgggtattttcccgtcactttgcacgtggaagactatggcgcctttgat660cccaacagggcttatgtgttcggttatgagcctcattctgttttgcccatagctgttggg720atcctcggggaccttgttggattcatgccgctaccaaagatgaagattcttgcaagcagt780gcggtgttctacaccccgttcctaaggcaaatatggacatggttggggttggctcctgcg840tcgagaaagagtttctactcctaccttggagctggttatagctgtattatagtgccagga900ggtgtgcaggaaatacttcatatggatcatgattcagaggttgcttttcttaaaccaaga960aaaggttttgttaagatagctattgagatgggttgccctgtagtccccgtttttgctttc1020ggacagagctatgtttacaaatggtggaggccaggtggcaagttaattgtcaagattgct1080agagcaatcaaattttctccaataatcttctggggaaaactggggactcccatccctttt1140gcaacaccaatgcatgtgattgttggaaggccaattgaggttgtaaagaatcctcaacct1200accattgatgagataaaccaagtccacggacagttcgttgttgcgatgcaagatctgttc1260gagaaatacaagagcagaactggataccctgatcttcagttaagagttctttga1314<210>2<211>437<212>prt<213>zeamays<400>2metglyleualavalalavalglyargargprothrglnargsergly151015valserargserargcysalaglylysargprothralaalaglupro202530arghishisarghisargthrgluglyarglysgluargasnprogly354045glyserargargtyrglnargproaspseraspalaserargalagly505560glualaasphisthrargthrargthrleuserthralaleuleuarg65707580tyralaproserargproaspargthrglyprothrpheleuhisleu859095seralaargargargprothrargmetglyalaglythrasnasngly100105110leuserasnglyalaalaalaglyglnargalaaspaspglythrthr115120125valpheargglythralatyrserproleuargthrthrvalalaleu130135140alaleutrpleuglyalailehispheasnalapheleuvalleuala145150155160serleupheleupheproargargvalalaalaleuvalleualathr165170175glnleuphephemetpheleuproleuserasplysserargleugly180185190arglysilealaargpheileserlystyrvalileglytyrphepro195200205valthrleuhisvalgluasptyrglyalapheaspproasnargala210215220tyrvalpheglytyrgluprohisservalleuproilealavalgly225230235240ileleuglyaspleuvalglyphemetproleuprolysmetlysile245250255leualaserseralavalphetyrthrpropheleuargglniletrp260265270thrtrpleuglyleualaproalaserarglysserphetyrsertyr275280285leuglyalaglytyrsercysileilevalproglyglyvalglnglu290295300ileleuhismetasphisaspsergluvalalapheleulysproarg305310315320lysglyphevallysilealaileglumetglycysprovalvalpro325330335valphealapheglyglnsertyrvaltyrlystrptrpargprogly340345350glylysleuilevallysilealaargalailelyspheserproile355360365ilephetrpglylysleuglythrproileprophealathrpromet370375380hisvalilevalglyargproilegluvalvallysasnproglnpro385390395400thrileaspgluileasnglnvalhisglyglnphevalvalalamet405410415glnaspleupheglulystyrlysserargthrglytyrproaspleu420425430glnleuargvalleu435<210>3<211>1002<212>dna<213>zeamays<400>3atgggggcgggaaccaataatggcctgagcaacggcgccgccgcagggcagcgcgcggac60gacgggaccacggtgttccggggcacggcgtactcgccgctacggaccacggtggcgctc120gcgctgtggctcggggccatccacttcaacgccttcctcgtcctcgcctcgctcttcctc180ttcccgcgccgcgtcgccgcactggtgctggcgacgcagctcttcttcatgttcctgccg240ctcagtgataagagcagactgggccgcaagatcgccaggttcataagcaagtacgtcatt300gggtattttcccgtcactttgcacgtggaagactatggcgcctttgatcccaacagggct360tatgtgttcggttatgagcctcattctgttttgcccatagctgttgggatcctcggggac420cttgttggattcatgccgctaccaaagatgaagattcttgcaagcagtgcggtgttctac480accccgttcctaaggcaaatatggacatggttggggttggctcctgcgtcgagaaagagt540ttctactcctaccttggagctggttatagctgtattatagtgccaggaggtgtgcaggaa600atacttcatatggatcatgattcagaggttgcttttcttaaaccaagaaaaggttttgtt660aagatagctattgagatgggttgccctgtagtccccgtttttgctttcggacagagctat720gtttacaaatggtggaggccaggtggcaagttaattgtcaagattgctagagcaatcaaa780ttttctccaataatcttctggggaaaactggggactcccatcccttttgcaacaccaatg840catgtgattgttggaaggccaattgaggttgtaaagaatcctcaacctaccattgatgag900ataaaccaagtccacggacagttcgttgttgcgatgcaagatctgttcgagaaatacaag960agcagaactggataccctgatcttcagttaagagttctttga1002<210>4<211>22<212>dna<213>artificialsequence<220><223>反转录内参上游引物18s-f。<400>4agtttgaggcaataacaggtct22<210>5<211>22<212>dna<213>artificialsequence<220><223>反转录内参下游引物18s-r。<400>5gatgaaatttcccaagattacc22<210>6<211>22<212>dna<213>artificialsequence<220><223>基因全长pcr扩增上游引物f1。<400>6agaatggggctggcagtggcgg22<210>7<211>28<212>dna<213>artificialsequence<220><223>基因全长pcr扩增下游引物r1。<400>7tcaaagaactcttaactgaagatcaggg28<210>8<211>20<212>dna<213>artificialsequence<220><223>实时定量pcr上游引物f2。<400>8tggcacaccattccccttcc20<210>9<211>24<212>dna<213>artificialsequence<220><223>实时定量pcr下游引物r2。<400>9ggactcttagatgaaggccaggat24当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1
- 一种双酰基双异硫氰酸酯衍生物及其制备方法和应用
- 用于治疗消化不良的单酰基甘油和脂溶性营养物质的制作方法
- 用于治疗具有非机械基础的吸收不良的单酰基甘油和脂溶性营养物质的制作方法
- 含有以单乙酰基二酰基甘油化合物作为有效成分的用于促进红细胞系细胞的分化或者增 ...的制作方法
- 含有以单乙酰基二酰基甘油化合物作为有效成分的哮喘的预防或者治疗用组合物的制作方法
- 一种西替利司他的制备方法
- 含有以单乙酰基二酰基甘油化合物作为有效成分的慢性阻塞性肺病的预防或者治疗用组合物的制作方法
- 一种从含植物油的微生物发酵液中提取脂肪酶的方法
- 含有以单乙酰基二酰基甘油化合物作为有效成分的类风湿性关节炎的预防或者治疗用组合物的制作方法
- 一种合成脂肪酰基甘氨酸的方法