一种能提高耐热脂肪酶产量的重组工程菌及其构建方法和应用与流程
本发明属于酶工程领域,特别涉及一种能提高耐热脂肪酶产量的重组工程菌及其构建方法和应用。
背景技术:
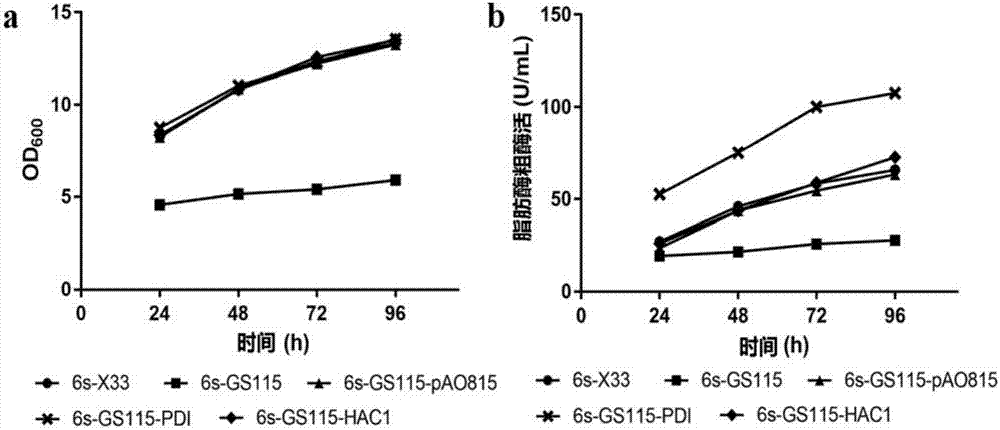
:耶氏解脂酵母脂肪酶2(lip2)对中长链脂肪酸甘油三酯(c12~c16)具有较强偏好性,被广泛应用于油脂水解、污水处理、食品加工、生物能源、化学合成以及胰腺缺乏症的治疗等多个领域,但其热稳定性较差限制了其应用。二硫键能提高蛋白质的热力学和动力学稳定性,增强其刚性,对保持蛋白质的完整结构和生物功能具有重要作用。研究表明,在蛋白质中引入二硫键能大幅度提高其热稳定性,如木聚糖酶,纤维素酶和植酸酶等。在内质网中,蛋白质二硫键的正确形成是折叠的主要限速步骤之一。分子伴侣二硫键异构酶(pdi,proteindisulfideisomerase)能催化新生肽链二硫键的形成和异构化,纠正错误配对的二硫键,减少肽链的聚沉,加快肽链的折叠速度。内质网中错误折叠或未折叠的蛋白质过多时,将产生未折叠蛋白反应(upr,unfoldedproteinresponse),该反应主要为ire1(inositol-requiringenzyme1)介导的hac1(thehomologoustotheactivatingtranscriptfactor/camp-responseelementbingdingprotein)信号分子途径。hac1分子的表达能上调折叠途径的分子伴侣(bip和pdi等)以及分泌途径的分子伴侣的转录和翻译水平。通过采用共表达转录因子hac1和分子伴侣pdi的策略,能显著提高外源蛋白与毕赤酵母的表达水平,已成功应用于人类单克隆抗体、人血清白蛋白融合蛋白、猪胰岛素前体和华根霉脂肪酶等。本实验室已筛选出多对能提高lip2耐热性能的单对二硫键突变体,改善其热稳定性,但其表达量明显下降。目前较少研究关注引入较多二硫键导致的重组蛋白分泌下降问题,并缺乏对此进行优化的报道,因此很有必要缓解该问题,提高外源蛋白质的表达量。技术实现要素:本发明的首要目的在于克服现有技术的缺点与不足,提供一种能提高耐热脂肪酶产量的重组工程菌。本发明的另一目的在于提供所述能提高耐热脂肪酶产量的重组工程菌的构建方法,采取于毕赤酵母gs115中共表达分子伴侣pdi的策略,以优化富含二硫键耶氏解脂酵母脂肪酶2耐热突变体的表达水平,最终提供种产量优化的耐热脂肪酶表达菌株,缓解目前引入二硫键导致的表达水平较低的问题。本发明的再一目的在于提供所述能提高耐热脂肪酶产量的重组工程菌的应用。本发明的目的通过下述技术方案实现:一种能提高耐热脂肪酶产量的重组工程菌,是通过将突变脂肪酶的重组质粒经酶切线性化后转化毕赤酵母gs115感受态细胞,再将线性化的pao815-pdi重组质粒二次转化到带有突变脂肪酶基因的毕赤酵母gs115感受态细胞获得。所述的突变脂肪酶为脂肪酶5s或脂肪酶6s;其中,脂肪酶5s的氨基酸序列如seqidno.1所示;脂肪酶6s的氨基酸序列如seqidno.2所示。所述的脂肪酶5s的编码核苷酸序列如seqidno.3所示。所述的脂肪酶6s的编码核苷酸序列如seqidno.4所示。所述的突变脂肪酶的重组质粒通过如下方法获得:以ppiczαa-lip2为模板,通过反向pcr突变,将所选的氨基酸位点突变为半胱氨酸,并转入大肠杆菌中,进行扩增培养、质粒提取和测序,获得引入五对或六对二硫键的突变脂肪酶重组质粒。所述的引入的五对或六对二硫键分别为二硫键s2-210、s8-214、s60-69、s122-196和s118-177,二硫键s2-210、s8-214、s14-216、s60-69、s122-196和s118-177;其中,二硫键s2-210为突变第2位和第210位氨基酸后,两氨基酸形成的二硫键,其它依此类推。所述的大肠杆菌优选为大肠杆菌top10。所述的pao815-pdi重组质粒为分子伴侣表达载体pao815-pdi,可以由南京金斯瑞生物科技有限公司进行全基因合成和构建。所述的能提高耐热脂肪酶产量的重组工程菌的构建方法,包括如下步骤:(1)将突变脂肪酶的重组质粒以pmeⅰ限制性内切酶进行线性化处理,并电击转化入感受态毕赤酵母gs115中,然后在ypds-zeocin平板上进行筛选,得到带有突变脂肪酶基因的毕赤酵母gs115;(2)将pao815-pdi质粒用sali限制性内切酶进行线性化处理,然后与带有突变脂肪酶基因的毕赤酵母gs115感受态细胞混合进行第二次电击转化,再在md平板上进行筛选,得到能提高耐热脂肪酶产量的重组工程菌。步骤(1)中所述的突变脂肪酶为脂肪酶5s或脂肪酶6s,其中,脂肪酶5s的编码核苷酸序列如seqidno.3所示;脂肪酶6s的编码核苷酸序列如seqidno.4所示。步骤(2)中所述的能提高耐热脂肪酶产量的重组工程菌是通过共表达分子伴侣pdi以优化脂肪酶表达量的菌株,将其命名为5s-gs115-pdi或6s-gs115-pdi。所述的能提高耐热脂肪酶产量的重组工程菌的构建方法,还包括将步骤(2)中得到的重组工程菌进行扩繁培养、补料和诱导的步骤:将获得的重组工程菌在ypds-zeocin液体培养基中进行扩繁培养后,转接至bmgy液体培养基进行去抑制培养,最后接种至bmmy液体培养基进行发酵。所述的ypds-zeocin平板中zeocin的浓度优选为100μg/ml。所述的发酵的时间优选为96h。所述能提高耐热脂肪酶产量的重组工程菌的产量较高,特别适合在工业上进行应用。本发明相对于现有技术具有如下的优点及效果:1、本发明中发现通过迭代组合进化并表达于毕赤酵母x33中获得的脂肪酶5s和6s,相对于lip2热稳定性提高了(tm值分别提高20.23和22.53℃),但其表达量明显下降。因此,本发明采用共表达分子伴侣的策略,以提高富含二硫键脂肪酶的表达水平。与传统提高蛋白表达的水平,更是有针对性地引入分子伴侣以加快蛋白折叠,缓解了由多对二硫键共同引起的表达水平下降等负面效应。2、本发明通过将脂肪酶5s、脂肪酶6s分别与分子伴侣pdi、转录因子hac1共表达于毕赤酵母gs115,获得工程菌株5s-gs115-pdi、5s-gs115-hac1、6s-gs115-pdi和6s-gs115-hac1。5s-gs115-hac1和6s-gs115-hac1菌株的发酵表达上清的脂肪酶粗酶活无显著变化,而5s-gs115-pdi和6s-gs115-pdi菌株发酵表达上清的脂肪酶粗酶活分别提高60%和80%。附图说明图1是表达脂肪酶5s重组工程菌株的生长曲线和发酵上清中粗酶活变化情况图;其中,a为生长曲线,b为粗酶活变化情况。图2是表达脂肪酶6s重组工程菌株的生长曲线和发酵上清中粗酶活变化情况图;其中,a为生长曲线,b为粗酶活变化情况。具体实施方式下面结合实施例及附图对本发明作进一步详细的描述,但本发明的实施方式不限于此。材料与试剂:ppiczαa-lip2、pao815-pdi和pao815-hac1毕赤酵母表达载体由上海金斯瑞生物公司全基因合成与构建;毕赤酵母x33来源于北京大学深圳研究院;毕赤酵母gs115购自lifetechnologies公司;质粒提取试剂盒购自omega贸易有限公司;kod-plus突变试剂盒购自东洋纺公司;pao815载体购自thermo公司;pmeⅰ和salⅰ限制性内切酶购自newenglandbiolabs公司;pcr产物纯化回收试剂盒购自大连宝生物公司;电转仪购自bio-rad公司;llb、llb+zeocin、ypd、ypd+zeocin、md、bmgy、bmmy培养基、均按照invitrogen毕赤酵母表达试剂盒操作手册配制,其余试剂均为国内外购买的分析纯级别。实施例1:分子伴侣和转录因子表达载体的构建以pdi序列(genbankid:eu805807.1)为目的片段,把起始序列atgcaa修改为kozak序列atggaa。该序列具有1个sali酶切位点,1个bglii酶切位点,和1个bamhi酶切位点。为方便线性化和后续研究多目的片段的串联,将第443位氨基酸碱基gtc替换为同义密码子gtt,第78位氨基酸碱基atc替换为同义密码子att,第255位氨基酸碱基tcc替换为同义密码子tct。以hac1序列(genbankid:fr839628.1)为目的片段,把起始序列atgccc修改为kozak序列atggcc。该序列具有1个sali酶切位点,为方便线性化,故将第59位氨基酸碱基gtc替换为同义密码子gtt。以上两段基因序列两端添加限制性酶切位点ecori,以pao815为载体,委托南京金斯瑞生物科技有限公司进行全基因合成和构建pao815-pdi和pao815-hac1质粒,并转化至大肠杆菌top10中。实施例2:突变脂肪酶(脂肪酶5s、脂肪酶6s)表达质粒的构建以ppiczαa-lip2为模板(构建方法参考中国专利申请201610279266.3“一种热稳定的脂肪酶及制备方法与应用”)和表1中引物进行突变,获得2段突变氨基酸。其氨基酸序列如下所示:脂肪酶5s的氨基酸序列如下:vctstetchidqesynffekyarlanigycvgpgtkifkpfncglqcahfpnvelieefcdprlifdvcgylavdhaskqiylvirgthsledvitdirimqapltnfdlaanisstctcdcclvhngfiqsynntynqigpkldsvieqypdyqiavtghslggaaallfginlkcnghdplvvtlgqpivgnacfanwvdklffgqecpdvckvskdrklyrithrgdivpqvpfwdgyqhcsgevfidwplihpplsnvvmcqgqsnkqcsagntllqqvnvignhlqyfvtegvcgi;脂肪酶6s的氨基酸序列如下:vctstetchidqecynffekyarlanigycvgpgtkifkpfncglqcahfpnvelieefcdprlifdvcgylavdhaskqiylvirgthsledvitdirimqapltnfdlaanisstctcdcclvhngfiqsynntynqigpkldsvieqypdyqiavtghslggaaallfginlkcnghdplvvtlgqpivgnacfanwvdklffgqecpdvckcskdrklyrithrgdivpqvpfwdgyqhcsgevfidwplihpplsnvvmcqgqsnkqcsagntllqqvnvignhlqyfvtegvcgi。编码所述耐热脂肪酶的核苷酸序列,如下所示:脂肪酶5s的核苷酸序列如下:gtgtgtacctctaccgagacctgtcacattgaccaggagtcctacaacttctttgagaagtacgcccgactcgcaaacattggatattgtgttggtcccggcactaagatcttcaagcccttcaactgtggcctgcaatgtgcccacttccccaacgttgagctcatcgaggagttctgtgacccccgtctcatctttgatgtttgtggttacctcgctgttgatcatgcctccaagcagatctaccttgttattcgaggaacccactctctggaggacgtcataaccgacatccgaatcatgcaggctcctctgacgaactttgatcttgctgctaacatctcttctacttgtacttgtgattgttgtcttgtccacaatggcttcatccagtcctacaacaacacctacaatcagatcggccccaagctcgactctgtgattgagcagtatcccgactaccagattgctgtcaccggtcactctctcggaggagctgcagcccttctgttcggaatcaacctcaagtgtaacggccacgatcccctcgttgttactcttggtcagcccattgtcggtaacgcttgttttgctaactgggtcgataaactcttctttggccaggagtgtcccgatgtctgtaaggtgtccaaagaccgaaagctctaccgaatcacccaccgaggagatatcgtccctcaagtgcccttctgggacggttaccagcactgctctggtgaggtctttattgactggcccctgatccaccctcctctctccaacgttgtcatgtgccagggccagagcaataaacagtgctctgccggtaacactctgctccagcaggtcaatgtgattggaaaccatctgcagtacttcgtcaccgagggtgtctgtggtatctaataa;脂肪酶6s的核苷酸序列如下:gtgtgtacctctaccgagacctgtcacattgaccaggagtgttacaacttctttgagaagtacgcccgactcgcaaacattggatattgtgttggtcccggcactaagatcttcaagcccttcaactgtggcctgcaatgtgcccacttccccaacgttgagctcatcgaggagttctgtgacccccgtctcatctttgatgtttgtggttacctcgctgttgatcatgcctccaagcagatctaccttgttattcgaggaacccactctctggaggacgtcataaccgacatccgaatcatgcaggctcctctgacgaactttgatcttgctgctaacatctcttctacttgtacttgtgattgttgtcttgtccacaatggcttcatccagtcctacaacaacacctacaatcagatcggccccaagctcgactctgtgattgagcagtatcccgactaccagattgctgtcaccggtcactctctcggaggagctgcagcccttctgttcggaatcaacctcaagtgtaacggccacgatcccctcgttgttactcttggtcagcccattgtcggtaacgcttgttttgctaactgggtcgataaactcttctttggccaggagtgtcccgatgtctgtaagtgttccaaagaccgaaagctctaccgaatcacccaccgaggagatatcgtccctcaagtgcccttctgggacggttaccagcactgctctggtgaggtctttattgactggcccctgatccaccctcctctctccaacgttgtcatgtgccagggccagagcaataaacagtgctctgccggtaacactctgctccagcaggtcaatgtgattggaaaccatctgcagtacttcgtcaccgagggtgtctgtggtatctaataa。脂肪酶5s和脂肪酶6s的制备方法如下:以ppiczαa-lip2为模板进行反向pcr,每引入一对二硫键需要进行两次反向pcr。由于二硫键s14-216和s2-210、s8-214均处于两段loop间(第1位氨基酸至第14位氨基酸和第207位氨基酸至221位氨基酸),而二硫键s14-216和s2-210离酶活中心相对较近,所以选择s8-214、s60-69、s122-196和s118-177作为4s的四条二硫键,s2-210、s8-214、s60-69、s122-196和s118-177作为5s的五条二硫键,s2-210、s8-214、s14-216、s60-69、s122-196和s118-177作为6s的六条二硫键。以表1的122c-f和122-r作为引物,得到脂肪酶4s第一次扩增引物。表1突变引物汇总注:斜线加粗处为突变位点pcr扩增条件为:94℃2min;94℃10s、66℃30s、68℃5min,10个循环。反应体系如下表2所示。表2pcr反应体系上游引物(10μm)1.5μl下游引物(10μm)1.5μlkod-plus高保真酶1μl模板(50ng/μl)1μl双蒸水35μl5×smartpcrbuffer5μl5×dntp(2m)5μl总体系50μl扩增产物以dnpⅰ酶消化模板,在琼脂糖凝胶电泳检测突变条带大小后,用t4连接酶连接环化过夜,随后使用热激法将突变质粒转入top10大肠杆菌感受态细胞,并涂布于llb+zeocin(zeocin浓度为25μg/ml)平板37℃过夜培养,挑选阳性转化子进行质粒的测序。将测序正确的阳性转化子于llb+zeocin(zeocin浓度为25μg/ml)液体培养基过夜扩培后,提取质粒。以该质粒为模板,按照同样的方法,依次以表1的引物196c-f和196c-r,196+118c-f和196+118c-r,177c-f和177c-r,8c-f和8c-r,214c-f和214c-r,60c-f和60c-r,69c-f和69c-r进行7次突变,获得脂肪酶4s的重组质粒。以脂肪酶4s的质粒为模板,依次以表1的引物210+8c-f和210+8c-r,210+214c-f和210+214c-r进行2次突变,获得脂肪酶5s的重组质粒。以脂肪酶5s的质粒为模板,依次以表1的引物210+214+14c-f和210+214+14c-r,210+214+216c-f和210+214+216c-r进行2次突变,获得脂肪酶6s的重组质粒。实施例3:线性化质粒电转化毕赤酵母、转化子筛选与产酶筛选将测序正确的阳性转化子于llb+zeocin(zeocin浓度为25μg/ml)液体培养基过夜扩增培养后,提取质粒,以pmeⅰ线性化处理并纯化回收,以总量为5μg的质粒线性化产物与x33毕赤酵母感受态混合电击转化。毕赤酵母感受态制备参照invitrogen公司操作手册。电转程序按照bio-rad公司推荐参数设置。电转完毕立刻加入1ml1mol/l山梨醇溶液,将菌液在30℃孵育复苏1小时后,均匀涂布于ypds+zeocin(zeocin浓度为100μg/ml)抗性平板上筛选。获得重组工程菌株5s-x33和6s-x33。实施例4:脂肪酶5s、脂肪酶6s与分子伴侣pdi、转录因子hac1的共表达由于以毕赤酵母x33构建的重组工程菌株5s-x33和6s-x33不利于共表达菌株转换子的筛选,于是把表达菌株(毕赤酵母x33)更换为毕赤酵母gs115。扩增含有脂肪酶5s和脂肪酶6s重组质粒的大肠杆菌工程菌,提取质粒并以以pmei将其线性化,将5μg的质粒线性化产物与gs115毕赤酵母感受态细胞混合后以电击法转化。毕赤酵母感受态细胞的制备参照invitrogen公司操作手册。电极法转化的步骤和程序设置按照bio-rad公司推荐参数设置。电转完毕立刻加入1ml1mol/l山梨醇溶液,将菌液在30℃孵育复苏1小时后,均匀涂布于ypd+zecion(zeocin浓度为100μg/ml)平板进行筛选,获得重组工程菌株5s-gs115和6s-gs115。扩增含有pao815空载体质粒、pao815-pdi重组质粒和pao815-hac1重组质粒的大肠杆菌工程菌(实施例1获得),提取质粒并以sali线性化。将5μg的质粒线性化产物分别与5s-gs115和6s-gs115毕赤酵母感受态细胞混合进行第二次电击转化。毕赤酵母感受态细胞的制备参照invitrogen公司操作手册。电极法转化的步骤和程序设置按照bio-rad公司推荐参数设置。电转完毕立刻加入1ml1mol/l山梨醇溶液,将菌液在30℃孵育复苏1小时后,均匀涂布于md平板上进行筛选,获得共表达分子伴侣的重组工程菌株5s-gs115-pao815、6s-gs115-pao815、5s-gs115-pdi、6s-gs115-pdi、5s-gs115-hac1和6s-gs115-hac1。实施例5:优化重组工程菌株的发酵与采样用高压灭菌的牙签,分别挑取实施例4中获得的5s-gs115-pao815、6s-gs115-pao815、5s-gs115-pdi、6s-gs115-pdi、5s-gs115-hac1和6s-gs115-hac1转化子各6株,并接种于ypd-zeocin(zeocin浓度为100μg/ml)液体培养基内过夜纯化培养。以重组工程菌株5s-gs115、6s-gs115、5s-x33和6s-x33为对照组,将其重组单克隆接种至ypd-zeocin(zeocin浓度为100μg/ml)液体培养基中过夜纯化培养。过夜培养后,离心收集菌体,并以50ml的bmgy液体培养基重悬菌体,过夜培养至od600=4,以12000rpm离心收集菌体,并用50ml的bmmy液体培养基(ph=5.5)重悬菌体,在280rpm25℃条件下诱导发酵96h,并每24h测定发酵液od值,并补充甲醇至1%(v/v)的终浓度。实施例6:优化重组工程菌株的生长曲线测定使用分光光度计,每24h测定发酵液的od值。菌株的生长曲线如图1和图2所示,除了未导入pao815空载体的5s-gs115和6s-gs115。其余重组工程菌株的生长曲线基本一致。实施例7:粗酶活测定参照《gb/t23535-2009脂肪酶制剂》并略做修改。(1)取50ml小烧杯若干,空白组和样品组均加入橄榄油乳化剂4.00ml和ph=7.5的磷酸缓冲液5.00ml,于空白组的小烧杯加入95%乙醇15.00ml;(2)将所有小烧杯放进水浴锅于40℃预热10min后,于空白组和样品组各加入待测酶液1.00ml,立刻混匀并计时;(3)精确反应15min后,空白组取出;样品组立刻补加入95%乙醇15.00ml,取出;(4)于空白组和样品组加入15ml蒸馏水,用0.05mol/l氢氧化钠标准溶液滴定,以酚酞为指示剂,记录其消耗体积,利用以下公式计算酶活。u=【(v1-v2)×10-3×m×106×n】/t式中:u:样品中的酶活力,u/ml;v1:滴定样品时消耗氢氧化钠标准溶液的体积,ml;v2:滴定空白时消耗氢氧化钠标准溶液的体积,ml;m:氢氧化钠的标准液的浓度,mol/l;n:样品的稀释倍数;t:反应时间,min。重组工程菌株粗酶活变化情况如图1和图2所示。5s-gs115-pao815与5s-x33菌株发酵表达上清的粗酶活无显著差异,而5s-gs115-pdi菌株的发酵表达上清的粗酶活比5s-gs115-pao815菌株提高60%;6s-gs115-pao815与6s-x33菌株的发酵表达上清粗酶活无显著差异,而6s-gs115-pdi菌株发酵表达上清的粗酶活比6s-gs115-pao815提高80%;5s-gs115-hac1和6s-gs115-hac1菌株的粗酶活分别与5s-gs115和6s-gs115对比,均无显著差异。上述实施例为本发明较佳的实施方式,但本发明的实施方式并不受上述实施例的限制,其他的任何未背离本发明的精神实质与原理下所作的改变、修饰、替代、组合、简化,均应为等效的置换方式,都包含在本发明的保护范围之内。序列表<110>华南农业大学<120>一种能提高耐热脂肪酶产量的重组工程菌及其构建方法和应用<160>28<170>siposequencelisting1.0<210>1<211>301<212>prt<213>人工序列(artificialsequence)<220><223>脂肪酶5s的氨基酸序列<400>1valcysthrserthrgluthrcyshisileaspglnglusertyrasn151015phepheglulystyralaargleualaasnileglytyrcysvalgly202530proglythrlysilephelyspropheasncysglyleuglncysala354045hispheproasnvalgluleuileglugluphecysaspproargleu505560ilepheaspvalcysglytyrleualavalasphisalaserlysgln65707580iletyrleuvalileargglythrhisserleugluaspvalilethr859095aspileargilemetglnalaproleuthrasnpheaspleualaala100105110asnileserserthrcysthrcysaspcyscysleuvalhisasngly115120125pheileglnsertyrasnasnthrtyrasnglnileglyprolysleu130135140aspservalilegluglntyrproasptyrglnilealavalthrgly145150155160hisserleuglyglyalaalaalaleuleupheglyileasnleulys165170175cysasnglyhisaspproleuvalvalthrleuglyglnproileval180185190glyasnalacysphealaasntrpvalasplysleuphepheglygln195200205glucysproaspvalcyslysvalserlysasparglysleutyrarg210215220ilethrhisargglyaspilevalproglnvalprophetrpaspgly225230235240tyrglnhiscysserglygluvalpheileasptrpproleuilehis245250255proproleuserasnvalvalmetcysglnglyglnserasnlysgln260265270cysseralaglyasnthrleuleuglnglnvalasnvalileglyasn275280285hisleuglntyrphevalthrgluglyvalcysglyile290295300<210>2<211>301<212>prt<213>人工序列(artificialsequence)<220><223>脂肪酶6s的氨基酸序列<400>2valcysthrserthrgluthrcyshisileaspglnglucystyrasn151015phepheglulystyralaargleualaasnileglytyrcysvalgly202530proglythrlysilephelyspropheasncysglyleuglncysala354045hispheproasnvalgluleuileglugluphecysaspproargleu505560ilepheaspvalcysglytyrleualavalasphisalaserlysgln65707580iletyrleuvalileargglythrhisserleugluaspvalilethr859095aspileargilemetglnalaproleuthrasnpheaspleualaala100105110asnileserserthrcysthrcysaspcyscysleuvalhisasngly115120125pheileglnsertyrasnasnthrtyrasnglnileglyprolysleu130135140aspservalilegluglntyrproasptyrglnilealavalthrgly145150155160hisserleuglyglyalaalaalaleuleupheglyileasnleulys165170175cysasnglyhisaspproleuvalvalthrleuglyglnproileval180185190glyasnalacysphealaasntrpvalasplysleuphepheglygln195200205glucysproaspvalcyslyscysserlysasparglysleutyrarg210215220ilethrhisargglyaspilevalproglnvalprophetrpaspgly225230235240tyrglnhiscysserglygluvalpheileasptrpproleuilehis245250255proproleuserasnvalvalmetcysglnglyglnserasnlysgln260265270cysseralaglyasnthrleuleuglnglnvalasnvalileglyasn275280285hisleuglntyrphevalthrgluglyvalcysglyile290295300<210>3<211>909<212>dna<213>人工序列(artificialsequence)<220><223>脂肪酶5s的核苷酸序列<400>3gtgtgtacctctaccgagacctgtcacattgaccaggagtcctacaacttctttgagaag60tacgcccgactcgcaaacattggatattgtgttggtcccggcactaagatcttcaagccc120ttcaactgtggcctgcaatgtgcccacttccccaacgttgagctcatcgaggagttctgt180gacccccgtctcatctttgatgtttgtggttacctcgctgttgatcatgcctccaagcag240atctaccttgttattcgaggaacccactctctggaggacgtcataaccgacatccgaatc300atgcaggctcctctgacgaactttgatcttgctgctaacatctcttctacttgtacttgt360gattgttgtcttgtccacaatggcttcatccagtcctacaacaacacctacaatcagatc420ggccccaagctcgactctgtgattgagcagtatcccgactaccagattgctgtcaccggt480cactctctcggaggagctgcagcccttctgttcggaatcaacctcaagtgtaacggccac540gatcccctcgttgttactcttggtcagcccattgtcggtaacgcttgttttgctaactgg600gtcgataaactcttctttggccaggagtgtcccgatgtctgtaaggtgtccaaagaccga660aagctctaccgaatcacccaccgaggagatatcgtccctcaagtgcccttctgggacggt720taccagcactgctctggtgaggtctttattgactggcccctgatccaccctcctctctcc780aacgttgtcatgtgccagggccagagcaataaacagtgctctgccggtaacactctgctc840cagcaggtcaatgtgattggaaaccatctgcagtacttcgtcaccgagggtgtctgtggt900atctaataa939<210>4<211>909<212>dna<213>人工序列(artificialsequence)<220><223>脂肪酶6s的核苷酸序列<400>4gtgtgtacctctaccgagacctgtcacattgaccaggagtgttacaacttctttgagaag60tacgcccgactcgcaaacattggatattgtgttggtcccggcactaagatcttcaagccc120ttcaactgtggcctgcaatgtgcccacttccccaacgttgagctcatcgaggagttctgt180gacccccgtctcatctttgatgtttgtggttacctcgctgttgatcatgcctccaagcag240atctaccttgttattcgaggaacccactctctggaggacgtcataaccgacatccgaatc300atgcaggctcctctgacgaactttgatcttgctgctaacatctcttctacttgtacttgt360gattgttgtcttgtccacaatggcttcatccagtcctacaacaacacctacaatcagatc420ggccccaagctcgactctgtgattgagcagtatcccgactaccagattgctgtcaccggt480cactctctcggaggagctgcagcccttctgttcggaatcaacctcaagtgtaacggccac540gatcccctcgttgttactcttggtcagcccattgtcggtaacgcttgttttgctaactgg600gtcgataaactcttctttggccaggagtgtcccgatgtctgtaagtgttccaaagaccga660aagctctaccgaatcacccaccgaggagatatcgtccctcaagtgcccttctgggacggt720taccagcactgctctggtgaggtctttattgactggcccctgatccaccctcctctctcc780aacgttgtcatgtgccagggccagagcaataaacagtgctctgccggtaacactctgctc840cagcaggtcaatgtgattggaaaccatctgcagtacttcgtcaccgagggtgtctgtggt900atctaataa939<210>5<211>27<212>dna<213>人工序列(artificialsequence)<220><223>122c-f<400>5tgttgtcttgtccacaatggcttcatc27<210>6<211>33<212>dna<213>人工序列(artificialsequence)<220><223>122c-r<400>6atcacaagtagcagtagaagagatgttagcagc33<210>7<211>24<212>dna<213>人工序列(artificialsequence)<220><223>196c-f<400>7acaagcgttaccgacaatgggctg24<210>8<211>31<212>dna<213>人工序列(artificialsequence)<220><223>196c-r<400>8tttgctaactgggtcgataaactcttctttg31<210>9<211>31<212>dna<213>人工序列(artificialsequence)<220><223>196+118c-f<400>9acttgtgattgttgtcttgtccacaatggct31<210>10<211>37<212>dna<213>人工序列(artificialsequence)<220><223>196+118c-r<400>10acaagtagaagagatgttagcagcaagatcaaagttc37<210>11<211>23<212>dna<213>人工序列(artificialsequence)<220><223>177c-f<400>11tgtaacggccacgatcccctcgt23<210>12<211>27<212>dna<213>人工序列(artificialsequence)<220><223>177c-r<400>12cttgaggttgattccgaacagaagggc27<210>13<211>30<212>dna<213>人工序列(artificialsequence)<220><223>8c-f<400>13tgtcacattgaccaggagtcctacaacttc30<210>14<211>29<212>dna<213>人工序列(artificialsequence)<220><223>8c-r<400>14ggtctcggtagaggtgtacacatggtgat29<210>15<211>29<212>dna<213>人工序列(artificialsequence)<220><223>214c-f<400>15tgtaaggtgtccaaagaccgaaagctcta29<210>16<211>22<212>dna<213>人工序列(artificialsequence)<220><223>214c-r<400>16gacatcggggttctcctggcca22<210>17<211>26<212>dna<213>人工序列(artificialsequence)<220><223>60c-f<400>17tgtgacccccgtctcatctttgatgt26<210>18<211>26<212>dna<213>人工序列(artificialsequence)<220><223>60c-r<400>18gaactcctcgatgagctcaacgttgg26<210>19<211>26<212>dna<213>人工序列(artificialsequence)<220><223>69c-f<400>19tgtggttacctcgctgttgatcatgc26<210>20<211>26<212>dna<213>人工序列(artificialsequence)<220><223>69c-r<400>20aacatcaaagatgagacgggggtcac26<210>21<211>30<212>dna<213>人工序列(artificialsequence)<220><223>210+8c-f<400>21tgtcacattgaccaggagtcctacaacttc30<210>22<211>30<212>dna<213>人工序列(artificialsequence)<220><223>210+8c-r<400>22ggtctcggtagaggtacacacatggtgatg30<210>23<211>29<212>dna<213>人工序列(artificialsequence)<220><223>210+214c-f<400>23tgtaaggtgtccaaagaccgaaagctcta29<210>24<211>22<212>dna<213>人工序列(artificialsequence)<220><223>210+214c-r<400>24gacatcgggacactcctggcca22<210>25<211>30<212>dna<213>人工序列(artificialsequence)<220><223>210+214+14c-f<400>25tacaacttctttgagaagtacgcccgactc30<210>26<211>28<212>dna<213>人工序列(artificialsequence)<220><223>210+214+14c-r<400>26acactcctggtcaatgtgacaggtctcg28<210>27<211>25<212>dna<213>人工序列(artificialsequence)<220><223>210+214+216c-f<400>27ccgaatcacccaccgaggagatatc25<210>28<211>30<212>dna<213>人工序列(artificialsequence)<220><223>210+214+216c-r<400>28tagagctttcggtctttggaacacttacag30当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1