一种香蕉枯萎菌milRNA的制作方法
本发明涉及生物技术领域,具体涉及一种香蕉枯萎菌milrna。
背景技术:
1、植物内源mirna在转录后以基因沉默的方式负调控靶标基因的表达,在植物生长发育、抗逆抗病等多个方面发挥重要的作用
真核生物的小rna主要分为micrornas(mirnas)、sirnas和pirnas。mirna和sirna长度约为20~25nt,均由类似rnaseiii的核酸内切酶dicer(或dicer类似蛋白)加工产生,它们的形成既有联系又有区别。sirna途径是由是外源入侵,如病毒,转座子以及转基因等产生变体dsrna引发的;而mirna途径由内源性hprna诱导。sirna和mirna都由dcls(或dicer类似蛋白)剪切形成,其中与mrna互补的链结合rna诱导的沉默复合体(rna-inducedsilencingcomplex,risc),以ptgs的方式作用序列同源的mrna,通过mrna剪切、翻译抑制或者甲基化等方式调控基因的表达。pirna依赖piwi蛋白加工成熟,仅存在动物细胞。
植物内源mirna是由ⅲ型rnase核酸酶dicer从含有茎环结构的内源转录本中切割产生一类21-24nt长度的小分子非编码rna,通过转录后水平以mrna剪切或翻译抑制的方式沉默序列同源的mrna靶标基因。mirna基因广泛分布于植物基因组中,在基因组上通常具有独立的基因座位(1ocus),转录产物自身折叠成不完全配对的发卡结构(hairpin)。已经有大量的报道证实植物内源mirnas参与植物对病原菌的pti和eti免疫,在植物生长发育、抗逆抗病等多个方面发挥重要的作用。
2、真菌中存在与植物mirna在结构、长度、作用方式上类似的小rnas(microrna-likernas,milrnas)。
milrnas最早在粗糙脉孢霉(neurosporacrassa)中发现的,与植物microrna在结构、小rna长度以及作用方式上非常相似,因为与植物的mirna类似而命名microrna-likes(milrnas)。粗糙脉孢霉的milrnas通过dicers,qde-2,核酸外切酶活性的qip效应蛋白以及具有iii型rnase功能域的线粒体核糖体大亚基mrpl3蛋白(mitochondrialribosomalproteinlargesubunit,mrpl3)的不同组合,产生有四种不同的生物形成途径。虽然四种途径中从基因间隔区转录的rna前体序列长度各异,但都具有发卡结构,形成的milrnas具有5'末端u的偏好性,能够特异与argonaute(ago)蛋白结合;此外,实验证明milr-1可以通过基因沉默的方式作用调靶标序列(ncu07311)mrna,主要通过抑制mrna翻译的方式进行,与植物microrna通过mrna剪切和抑制转录的方式不一致(leeetal.,2010)。milr-1是构巢曲霉中丰度最高的mirna-likernas,xueetal.(2012)以milr-1为研究对象构建了依赖ago的milr-1离体生物形成的生物化学研究框架,研究表明milr-1的加工成熟过程可以分为5个步骤,与植物microrna的生物合成途径不一样的是,除了qde-2,dicer等核心蛋白组分外,还需要qip以及rna外来复合体(exosome)的共同参与,加工成为成熟的小rna(xueetal.,2012)。
随后,植物病原真菌的小rna相关研究进展迅速,在稻瘟病(magnaportheoryzae)(nunesetal.,2011)油菜核盘菌(sclerotiniasclerotiorum)(zhouetal.,2012)葡萄灰霉菌(botrytiscinerea)(weibergetal.,2013;wangetal.,2017a)疫霉(phytophthora)(fahlgrenetal.,2013)尖孢镰刀菌番茄专化型(fusariumoxysporumf.sp.lycopersici)(chenetal.,2014)黄曲霉(aspergillusflavus)(baietal.,2015)小麦条锈(pucciniastriiformisf.sp.tritici)(muethetal.,2015;wangetal.,2017b)中陆续都发现存在milrnas。随着技术的进步,深度测序可以预测到低丰度milrna及其骨架序列的存在,但是由于病原真菌中大部分的milrnas表达本底水平太低,常规的同位素32p-atp标记的mirna探针无法检测到低丰度的milrnas,因此,无法对低丰度的milrnas及其发夹序列进行验证。
3、植物病原真菌与寄主之间存在跨界小rna基因沉默,抑制寄主的免疫反应促进病原菌的侵染。
病原真菌利用小rna作为效应因子(effector)进入植物细胞内部抑制植物免疫防卫反应。灰霉菌引发的灰霉病可以侵染200多种植物。weibergetal.(2013)的研究显示灰霉菌能够输送bc-srnas进入寄主细胞通过与ago1特异性结合抑制ago1功能,沉默寄主的免疫相关基因促进病原菌侵染。拟南芥的δago1缺失突变体由于丧失了ago1功能,减少了对灰霉菌的敏感性;相反,灰霉菌的δdcl1dcl2双突变体由于无法加工产生bc-srnas而完全丧失了侵染性。因此,真菌病原物通过转移具有致病力的小rna效应因子进入寄主细胞通过抑制寄主的防卫反应促进病原菌的侵染,该发现阐释了这种自然发生的跨界rnai沉默现象作为病原真菌的一种更加高级的致病机制(weibergetal.,2013)。此外,在葡萄灰霉菌(botrytiscinerea)中还发现一个低丰度表达的bc-sir37,仅侵染时上调表达,而bc-sir37的3个确定的作用靶标都是寄主免疫相关的(wangetal.,2017)。bc-dcl1和dcl2是产生bc-srnas的主要组分,沉默dcls可以显著减少病原菌的致病力。病原菌存在环境rnai可以吸收外部的rna,在水果、蔬菜以及鲜花的表面施用靶向dcls的dsrna和srna可以通过调控bc-srnas表达从而明显减轻灰霉病的症状(wangetal.,2016;wangetal.,2017a)。
小rna是病原真菌一类重要的致病因子。小麦条锈病编码的pst-milr1仅仅在侵染时诱导表达,在pst中通过bsmv基因沉默抑制pst-milr1的表达明显减弱了条锈病的致病性,同时,在小麦中敲除其靶标小麦病程相关基因pr2(sm638)则增强了小麦对无毒菌株的感病性,正反实验证实pst-milr1以跨界基因沉默方式靶向寄主免疫相关基因pr2(sm638)从而抑制寄主的免疫反应促进病原菌侵染(wangetal.,2017b)。
4、香蕉枯萎菌的研究
香蕉枯萎病(fusariumoxysporumf.sp.cubense,foc)是破坏香蕉维管束导致植株死亡的毁灭性土传病害,其病菌腐生能力很强,在土壤中可长期存活,属于典型的潜伏型侵染。早期外部症状表现不明显,中后期才表现症状,因此,给香蕉产业和香蕉种植者造成巨大的经济损失。世界植蕉区已发现其有4个生理小种(还可能存在其它小种),其中生理小种1号侵染粉蕉(abb组)、龙牙蕉(aab组)类及部分aaa组品种;生理小种2号只侵染杂交三倍体棱香蕉(abb),对香蕉栽培品种的危害较小;生理小种3号侵染野生的羯尾蕉属,对香蕉栽培品种不造成危害,在许多地区均有报道;生理小种4号侵染香牙蕉、龙牙蕉、大蕉,其寄主范围大于生理小种1号。4号生理小种又分为亚热带4号(subtropicalrace4,sr4)和热带4号小种(tropicalrace4,tr4),在我国主要是1号和4号生理小种,其中热带4号小种危害最严重。
目前,香蕉没有有效的抗性种质资源,生产上没有有效的化学防治方法,因此,培育抗性品系是解决香蕉枯萎病的根本途径之一。小rna在介导病原物-寄主相互作用过程中发挥着重要的作用,具有提升植物抗病害能力的巨大潜力。香蕉枯萎菌是否编码小rna,是否利用小rna促进侵染等相关研究目前均没有文献报道,为此,我们开展了香蕉枯萎菌内源小rna鉴定及检测验证工作。
技术实现要素:
针对以上现有技术的不足之处,本发明提供了一种香蕉枯萎菌milrna。
本发明采取的技术方案如下:
一种香蕉枯萎菌milrna,所述香蕉枯萎菌milrna的基因序列如seqidno:1所示。
本发明还涉及一种载体,所述载体含有权利要求1所述的milrna。
本发明还涉及一种感受态细胞,所述感受态细胞含有如权利要求1所述的milrna或含有权利要求2所述的载体。
与现有技术相比,本发明的有益效果是:
通过深度测序获得了香蕉枯萎菌milrna,为后续进一步开发、利用小rna在香蕉枯萎菌中的作用机制提供前期工作铺垫。
附图说明
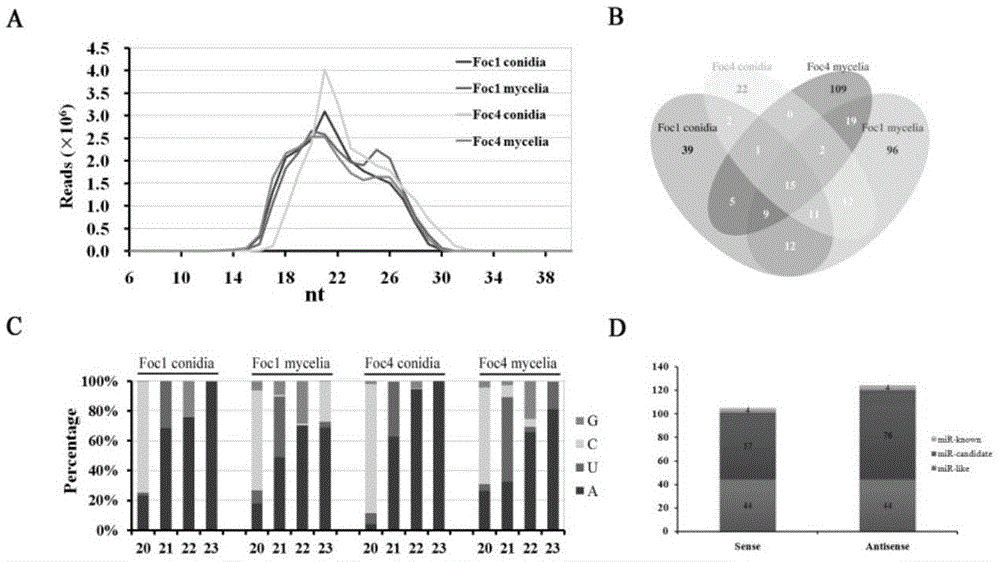
图1:小rna测序结果统计图;
图中,(a)四个小rna文库中小rna的长度统计结果;(b)四个小rna数据库小rna序列的venn图;(c)获得的小rna5末端碱基偏好性分析结果;(d)四个小rna数据库中正链、负链来源的小rna统计;
图2:高丰度milrnas的同位素32p-atp标记的反向互补dna探针杂交结果;
图中,-:孢子;+:菌丝;m:混合样品,*:表示milrna的伴随链小rna。
具体实施方式
下面通过具体实施方式结合附图对本发明作进一步详细说明。
本发明实施例中涉及到的主要试剂:
trizol(trnzolreagent,tiangen,cat#dp405-02)
peg8000沉淀缓冲液:20%(w/v)peg8000,1mnacl。
edc交联缓冲液(12ml):122.5μl12.5m1-甲基咪唑(ph8.0),0.373g1-乙基-3-(3-二甲基氨基丙基)碳酰二亚胺盐酸盐(edc)。
mirnanorthernblotting杂交缓冲液:5×ssc,20mmnahpo4(ph7.2),7%sds,3×denhardt`s溶液。
17%dpage:尿素12.6g,水1.25ml,10×tbe1.5ml,acr/bis(30%)17ml,在微波炉或者热水中溶解尿素,待冷却后加入240μl10%ap,混匀后再加入10μltemed。
washingbuffer:2×ssc,0.1%sds。
1mmes(ph5.6,10ml):称取2.132gmes溶于无菌水中,用koh调ph至5.6,定容至10ml,过滤除菌,分装到离心管中,-20℃保存。
100mmas(10ml):称取0.1962gas溶于dmso,定容至10ml,分装到1.5ml离心管中,-20℃保存。
实施例1:小rna测序及生物信息学分析
分别构建foc1,foc4的菌丝和孢子的小rna文库,送北京华大基因测序,测序获得的数据如图1和表1所示。
表1小rna测序数据一览表
经过生物信息学分析,鉴定得88个具有完整发夹(stem-loopstructure)并且两端均产生小rna的milrnas,(表2给出了其中一个milrna,命名为milrna2,milrna2*表示milrna2的伴随链小rna)。
表2foc测序获得的mirna-likes(milrnas)一览表
实施例2:高丰度milrnas的杂交检测
1、小rna的提取:
1.1样品的获取
菌株pda接种后28℃培养5-7天,用打孔器在pda平板的边缘打菌饼(直径0.5cm),挑取5-6块菌饼接入装有100mlpdb培养液的250ml三角瓶中,于28℃、180rpm震荡培养5-7d后用三层滤纸过滤,分别获得孢子和菌丝进行后续的小rna提取。
1.2、trizol抽提
液氮保护下,将步骤1.1获得的样品研磨成粉后转入离心管,加入等体积的trizol试剂,充分涡旋后室温放置5-10min。加入1/5体积的氯仿:异戊醇(24:1),涡旋后冰上静置3-5min,待自动分层后离心,12000g、4℃条件下离心10min。
1.3、分级沉淀高分子量rna
离心后,上清转入新的离心管,然后加入等体积的65℃孵育的peg8000沉淀缓冲液,颠倒混匀后室温静置直至冷却到室温,然后冰浴30-45min后离心10min沉淀高分子量rna。
1.4、沉淀小rna
离心后的上清液加入1/10体积的3mnaac(ph5.2)和等体积的预冷的异丙醇,-20℃沉淀1-2hr,12000g、4℃离心20min,沉淀用80%的乙醇洗涤1-2次,depc水溶解后放入-80℃备用。
2、小rna的杂交检测:
2.1、17%dpage分离
将步骤1提取的小rna溶入等体积的milrnasloadingbuffer(abi,cat.am8547),100℃变性5min后,立即冰浴5min使rna样品完全变性。变性后的milrnas样品在17%dpage上样,0.5×tbe电泳缓冲液,300v电泳3-6h即可。
2.2、转膜及交联
将含有rna的凝胶切下,先在1×tbe转膜缓冲液中浸润30s,同时准备好nx中性电荷的尼龙膜(hybond-
2.3、小rna杂交
mirna杂交采用mirnanorthernblotting杂交缓冲液,或者是
2.4曝光及信号检测
杂交结束后倒掉带有同位素的杂交液,用washingbuffer洗膜2次,每次15-20min。洗完后用monitor检测信号,阳性区域的信号应比膜边缘地方的信号强,并且将信号控制在5μci以内,如果信号过强,继续洗膜。洗完膜后将膜加载保鲜膜或者拉伸膜上,放在磷屏中曝光30min-3h左右,放入typoon扫描保存图片。结果见图2。
以上内容是结合具体的实施方式对本发明所作的进一步详细说明,不能认定本发明的具体实施只局限于这些说明。对于本发明所属技术领域的普通技术人员来说,在不脱离本发明构思的前提下,还可以做出若干简单推演或替换。
序列表
<110>中国热带农业科学院环境与植物保护研究所
<120>一种香蕉枯萎菌milrna
<160>2
<170>siposequencelisting1.0
<210>1
<211>23
<212>dna/rna
<213>香蕉枯萎菌(fusariumoxysporum)
<400>1
tggatgaatcaagcgtggtatga23
<210>2
<211>23
<212>dna/rna
<213>香蕉枯萎菌(fusariumoxysporum)
<400>2
gtatcatgcgtgattcatccatc23
- 还没有人留言评论。精彩留言会获得点赞!