一种金属硫蛋白DaMT3a及其编码基因的应用
本发明属于生物领域,具体涉及一种金属硫蛋白damt3a及其编码基因的应用。
背景技术:
金属硫蛋白(metallothionein,mts)是一类富含半胱氨酸(cys),能够与重金属结合的低分子(6-7kda)多肽,普遍存在于微生物、高等动植物和人体中。根据目前已克隆得到的植物中金属硫蛋白基因cys残基的分布和排列方式,可以分为type1、type2、type3和type4四类。四类金属硫蛋白基因具有组织特异性表达的特点,其中type1主要在根中表达,type2主要在叶子中表达,type3主要在成熟的果实和叶中表达,type4主要在种子中表达对植物中金属硫蛋白基因的功能研究表明,其功能主要体现在:(1)能够结合细胞内的重金属,维持细胞内重金属离子的生态平衡;(2)能够作为活性氧的清除剂,其清除自由基(·oh)的能力约为sod的几千倍,而清除氧自由基(·o)的能力约是谷胱甘肽(gsh)的25倍,而且具有很强的抗氧化活动;(3)参与植物的生长和发育;(4)调控酶与转录因子活性等方面。
参薯(dioscoreaalatal.)又名大薯,也有称其为紫参薯(紫参薯花青素稳定性及合成途径中关键基因的克隆与分析,赵景梅,2018),作为一种药食同源的粮食作物,在我国被主要用于食品、医药、工业及燃料乙醇生产等。近年来,尽管我国种植产业取得了较快发展,但受土壤重金属污染和非生物胁迫,植物的生长发育受到严重的影响。金属硫蛋白,作为一种具有重要生物学功能的蛋白,不但可以有效减轻重金属对植物的伤害,而且能够通过清除植物体内的活性氧提高植物对逆境的耐受性。因此研究参薯中的重金属硫蛋白有助于了解其提高参薯抗逆的作用机制。目前,参薯中金属硫蛋白基因的功能还未见报道。
技术实现要素:
本发明的目的在于克服现有技术中的不足,提供一种金属硫蛋白damt3a及其编码基因的应用。
本发明的第一个方面是提供一种金属硫蛋白基因,其命名为damt3a基因,其核苷酸序列如seqidno:1所示。
本发明的第二个方面是提供一种蛋白质,其为金属硫蛋白damt3a,是本发明第一个方面所述的金属硫蛋白基因编码的蛋白质,其氨基酸序列如seqidno:2所示。
本发明的第三个方面是提供一种重组表达载体,其包含原始载体和本发明第一个方面所述的金属硫蛋白基因或其开放阅读框(seqidno:1所示的第66-254位核苷酸序列)。
其中,所述原始载体可以采用基因重组领域中常用的载体,例如病毒、质粒等。本发明对此不进行限定。在本发明的一个具体实施方式中,所述原始载体采用pgex-4t-1载体质粒或pmd18-t载体质粒,但应当理解的是,本发明还可以采用其他质粒、或者病毒等。
优选地,所述原始载体为pgex-4t-1载体质粒,seqidno:1所示的第66-254位核苷酸序列位于pgex-4t-1载体质粒的ecori和xhoi两限制性内切酶位点之间。
本发明的第四个方面是提供如本发明第一个方面所述的金属硫蛋白基因、或者本发明第二个方面所述的蛋白质、或者本发明第三个方面所述的重组表达载体在提高原核表达菌株生长速度中的应用。
其中,所述原核表达菌株可以是基因重组领域中常用的原核表达菌株,本发明对此不进行限定。在本发明的一个具体实施方式中,所述原核表达菌株为大肠杆菌e.colibl21(de3)。
本发明的第五个方面是提供如本发明第一个方面所述的金属硫蛋白基因、或者本发明第二个方面所述的蛋白质、或者本发明第三个方面所述的重组表达载体在提高原核表达菌株抗镉重金属胁迫、和/或nacl胁迫、和/或过氧化氢胁迫中的应用。
其中,所述原核表达菌株可以是基因重组领域中常用的原核表达菌株,本发明对此不进行限定。在本发明的一个具体实施方式中,所述原核表达菌株为大肠杆菌e.colibl21(de3)。
本发明的第六个方面是提供一种引物对,其核苷酸序列如seqidno:4和seqidno:5所示。
本发明的第七个方面是提供一种引物对,其核苷酸序列如seqidno:6和seqidno:7所示。
本发明的第八个方面是提供一种引物对,其特征在于,其核苷酸序列如seqidno:8和seqidno:9所示。
本发明首次克隆了参薯金属硫蛋白damt3a基因的全长cdna,基因表达分析显示damt3a基因在叶片中特异性表达,受乙烯利及脱落酸诱导上调表达,同时该基因受低温和高度胁迫上调表达,机械伤害胁迫下调表达,因此该基因可能与参薯抗逆密切相关,可作为参薯转基因育种的重要靶基因,有望通过调控该基因的表达来促进参薯的生长发育,从而最大潜力的挖掘参薯生产潜力。进一步将该基因转入原核表达菌株,转基因菌株在正常条件下、重金属离子(cd2+)、nacl胁迫和过氧化氢胁迫下的生长得到明显提高,所以,该基因可作为重要的基因资源,可以在参薯以外的其他植物或微生物的抗逆和抗重金属胁迫基因工程中得到应用。
附图说明
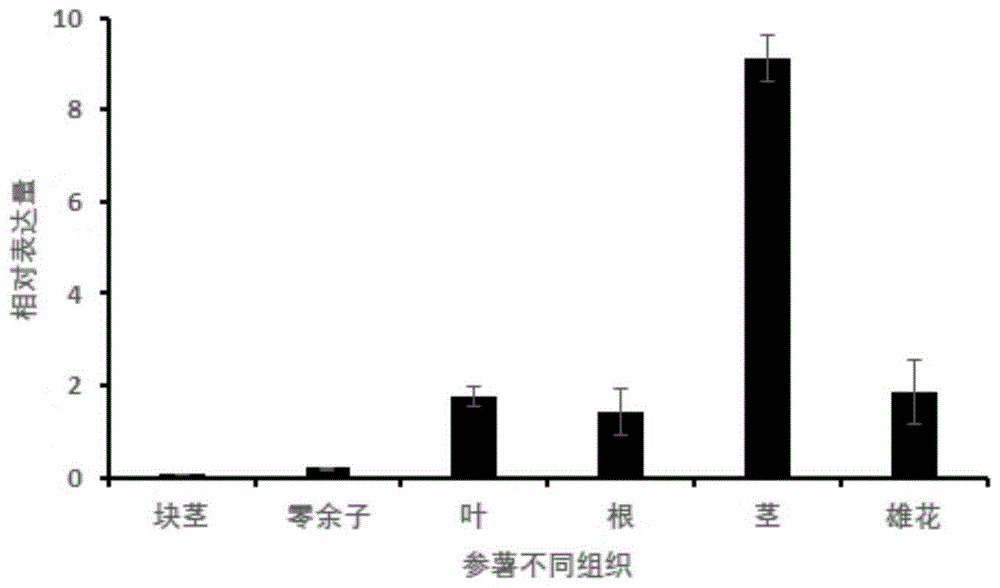
图1为damt3a基因的组织特异性表达分析结果(块茎、零余子、叶、根、茎、花)。
图2为低温处理对damt3a基因表达的影响。
图3为高温处理对damt3a基因表达的影响。
图4为机械伤害处理对damt3a基因表达的影响。
图5为aba处理对damt3a基因表达的影响。
图6为乙烯利处理对damt3a基因表达的影响。
图7为damt3a的原核表达分析(黑框内所示条带为受iptg诱导后所产生的特异表达的目标蛋白)。
图8为转基因大肠杆菌在培养基上生长情况(a:在正常条件下生长的od600测定;b:在重金属镉离子胁迫下生长的od600测定;c:在氯化钠胁迫下生长的od600测定;d:在过氧化氢胁迫下生长的od600测定)。
具体实施方式
我们首次克隆了参薯金属硫蛋白基因的全长cdna,并进行了基因表达分析和功能研究,将该基因命名为damt3a。下面参照附图,结合具体的实施例对本发明作进一步的说明,以更好地理解本发明。实施例中未注明具体技术或条件者,按照本领域内的文献所描述的技术或条件或者按照产品说明书进行。所用试剂或仪器未注明生产厂商者,均为可以通过公司购得的常规产品。
实施例1.金属硫蛋白及其编码基因的获得
分析已在ncbi登陆的拟南芥、木薯和甘薯等的金属硫蛋白的核苷酸序列,通过搜索我们建立的参薯块茎和叶片est序列数据库,筛选并拼接一条400bp左右的参薯金属硫蛋白基因组装后序列(contig),设计一对特异性引物扩增得到包含完整读码框的cdna全长序列。
具体方法如下:
特异性引物设计如下:
f(5’端):5’-acaacacaagccattcaatctat-3’;
r(3’端):5’-ttcattaaaataaaagagaggcag-3’。
以参薯(dioscoreaalatal.;海南大学儋州校区薯蓣种质资源圃,编号da56;以下简称参薯da56)叶片cdna为模板(以随机引物反转录获得),f和r为引物,其终浓度为0.5μmol/l,在25μl反应体系中进行pcr扩增。扩增程序为:94℃预变性5min;94℃变性30s,60℃退火30s,72℃延伸60s,30次循环;72℃延伸10min。
将该获得的一段300bp左右的核苷酸片段连接到pmd18-t载体(takara)进行测序,测序表明上述获得的片段即为本发明的金属硫蛋白基因,该片段具有序列表中seqidno:1的核苷酸序列,序列表中seqidno:1全长为345个核苷酸,包含一个长度为189个核苷酸的开放阅读框(orf,自seqidno:1的5’端第66-254位核苷酸序列)、65个核苷酸的5’-utr(自seqidno:1的5’端第1-65位核苷酸序列)和91个核苷酸的3’-utr(自seqidno:1的5’端第255-345位核苷酸序列),编码一个长度为62个氨基酸(序列表中seqidno:2)、分子量约为7.2kda的蛋白,即为金属硫蛋白,将该金属硫蛋白基因命名为damt3a基因。将上述含有seqidno:1的核苷酸的pmd18-t重组载体命名为pmd18-damt3a。
同时以参薯da56叶片基因组dna为模板,进行pcr扩增(程序同上所述),测序结果表明获得金属硫蛋白基因组序列,该序列具有序列表中seqidno:3的核苷酸序列,共441个核苷酸,包含了1个内含子(该内含子自序列3的5’端第113-208位核苷酸序列)。
实施例2.参薯damt3a基因表达模式分析
<1>参薯damt3a基因组织特异性表达
以参薯da56块茎、零余子、叶片、根、茎及雄花rna随机反转录的cdna为模板,用damt3a基因特异引物(f:5'-atgggatggtgattgttgagg-3';r:5'-cccatcattctcagcagcagt-3')进行实时荧光定量pcr。反应总体系20μl,包含2μl模板、10μl2×sybrpremix和10μmol·l-1的上下游引物各0.3μl;扩增程序为95℃预变性30s;94℃5s,60℃20s,72℃20s,45个循环。结果显示该基因在组织间的表达丰度差异较大,主要集中在茎中表达,其次是叶、根和雄花,在块茎和零余子中基本不表达(图1)。
<2>低温对damt3a基因的表达影响
选取有6-8片完全展开叶、长势基本一致且无病虫害的参薯da56组培苗进行低温(4℃),分别在0h、3h、6h和9h取样,取下后置于液氮冻存,提取rna。以反转录的cdna为模板,用damt3a基因特异引物进行实时荧光定量pcr(同“<1>参薯damt3a基因组织特异性表达”),结果表明,低温处理组培苗,damt3a基因的表达变化在6小时内不明显,只有9h时才有增加(图2)。
<3>高温对damt3a基因的表达影响
选取有6-8片完全展开叶、长势基本一致且无病虫害的参薯da56组培苗进行高温(45℃),分别在0h、3h、6h和9h取样,取下后置于液氮冻存,提取rna。以反转录的cdna为模板,用damt3a基因特异引物进行实时荧光定量pcr(同“<1>参薯damt3a基因组织特异性表达”),结果表明在高温处理下,damt3a基因的表达量有明显的下调表达。(图3)。
<4>伤害处理对damt3a基因的表达影响
选取有6-8片完全展开叶、长势基本一致且无病虫害的参薯da56组培苗进行伤害处理。用镊子对从顶端分生组织开始往下数的第三片叶子,进行一次挤压伤害(rajendran,linetal.2014),在0h、0.5h、6h和12h,取下后置于液氮冻存,提取rna。以反转录的cdna为模板,用damt3a基因特异引物进行实时荧光定量pcr(同“<1>参薯damt3a基因组织特异性表达”),结果显示damt3a基因受伤害胁迫,在初期有上调表达,之后表达量开始明显下调表达(图4)。
<5>植物生长物质对damt3a基因的表达影响
选取植物生长物质脱落酸aba和乙烯利,分别有6-8片完全展开叶、长势基本一致且无病虫害的参薯da56组培苗进行整株喷洒,直到叶片滴水,停止喷洒,分别在0h、3h、6h和9h取样,取下后置于液氮冻存,提取rna。以反转录的cdna为模板,用damt3a基因特异引物进行实时荧光定量pcr(同“<1>参薯damt3a基因组织特异性表达”),结果显示,damt3a基因的表达明显受脱落酸aba和乙烯利诱导,在6h表达量达到峰值,达到初始水平的2倍以上,之后开始下调表达(图5和图6)。
实施例3.原核表达与damt3a基因的功能验证
利用pgex-4t-1表达载体(载体购自transgenbiotech公司)构建了damt3a基因的原核表达载体(本实施例中表达载体仅为举例,本发明也可以采用其他表达质粒和病毒载体等),同时采用大肠杆菌表达菌株e.colibl21(de3)(感受态购自天根生化科技有限公司)诱导重组蛋白,测定重组蛋白对bl21菌株生长的影响,具体方法如下:
<1>含damt3a基因编码区重组载体的获得
设计damt3a基因编码区引物
f:5'-cgcgaattc(ecori酶切位点)atgtctagctgtggcaactgtg-3',
r:5'-cccctcgag(xhoi酶切位点)tcacttgccgcagttgc-3',
以pmd18-damt3a为模版,进行pcr扩增,扩增程序为:95℃预变性4min;95℃变性45s,57℃退火30s,共30次循环;72℃延伸8min。将扩增产物与pgex-4t-1表达载体用限制性内切酶ecori和xhoi进行双酶切,通过t4dnaligase进行连接,得到重组载体,利用载体引物pgex-5(5'-gggctggcaagccacgtttggtg-3')和pgex-3(5'-ccgggagctgcatgtgtcagagg-3')进行pcr鉴定,确保金属硫蛋白编码片段正向克隆到表达载体。重组载体进行测序鉴定,将鉴定表明正确的含有序列表中seqidno:1的第66-254位核苷酸序列、读框准确的重组表达载体命名为pgex-4t-1-damt3a。
<2>damt3a基因的原核表达
将获得的重组载体pgex-4t-1-damt3a导入e.colibl21(de3)中,得到重组表达菌,将鉴定正确的重组菌在含50μg/ml氨苄青霉素的lb培养基中培养至od600=0.4~0.6,添加iptg(异丙基-β-d-硫代半乳糖苷)至终浓度为1mm,30℃下诱导培养4h,以未加iptg的培养基培养的同样重组菌为对照,离心收集菌体,菌体蛋白进行15%sds-page电泳检测。结果表明,在iptg诱导下damt3a这个基因实现了高效异源表达,重组蛋白包含了damt3a以及胞质融合蛋白gst(谷胱甘肽s转移酶),并且表达蛋白的表观分子量与理论分子量相近,大约31kda(图7)。
<3>damt3a重组蛋白对菌株生长的影响
将含有pgex-4t-1-damt3a和pgex-4t-1的菌株培养至相同od600,添加iptg至终浓度1mm,30℃下诱导培养1h、2h、3h、4h、5h、6h和7h后分别测定其od600值。结果如图8a所示:含有pgex-4t-1-damt3a菌株的增长高于含有pgex-4t-1的菌株,说明damt3a基因编码的蛋白可以提高菌株在正常条件下的生长。此外,还测定了含有pgex-4t-1-damt3a和pgex-4t-1的菌株在重金属离子(cd2+,500μm)、盐(nacl,500mm)和过氧化氢(h2o2,1mm)胁迫下培养1h、2h、3h、4h、5h、6h和7h后的od600值。结果显示:含有pgex-4t-1-damt3a菌株的增长高于含有pgex-4t-1的菌株,说明damt3a基因编码的蛋白可以提高菌株在重金属镉离子、氯化钠和过氧化氢胁迫下的生长(图8b-8d)。将damt3a基因导入其他原核表达菌(比如e.colirosetta等)进行上述试验,结果同e.colibl21(de3),说明damt3a基因编码的蛋白可以提高其他原核菌株在正常条件下、重金属胁迫、氯化钠胁迫和过氧化氢胁迫下的生长。
以上对本发明的具体实施例进行了详细描述,但其只是作为范例,本发明并不限制于以上描述的具体实施例。对于本领域技术人员而言,任何对本发明进行的等同修改和替代也都在本发明的范畴之中。因此,在不脱离本发明的精神和范围下所作的均等变换和修改,都应涵盖在本发明的范围内。
序列表
<110>六盘水师范学院
<120>一种金属硫蛋白damt3a及其编码基因的应用
<160>9
<170>siposequencelisting1.0
<210>1
<211>345
<212>dna
<213>artificial
<400>1
acaacacaagccattcaatctattaactctttcttcttcttcttctccacaacatcatta60
caaacatgtctagctgtggcaactgtgactgtgctgacaagaaccagtgtgtgaagaagg120
ggaactcatatgggatggtgattgttgaggagaaggaggttgttgaggttgtggaaatgg180
caactgctgctgagaatgatgggtgcaagtgtgctgctaattgcacttgtgctggttgca240
actgcggcaagtgatcatatattaattaccctttaattatgtagtaattaataagatgta300
agccagagttacactggctttctgcctctcttttattttaatgaa345
<210>2
<211>62
<212>prt
<213>artificial
<400>2
metsersercysglyasncysaspcysalaasplysasnglncysval
151015
lyslysglyasnsertyrglymetvalilevalgluglulysgluval
202530
valgluvalvalglumetalathralaalagluasnaspglycyslys
354045
cysalaalaasncysthrcysalaglycysasncysglylys
505560
<210>3
<211>441
<212>dna
<213>artificial
<400>3
acaacacaagccattcaatctattaactctttcttcttcttcttctccacaacatcatta60
caaacatgtctagctgtggcaactgtgactgtgctgacaagaaccagtgtgtgtaagtag120
ttcttgtgtacttaaaactagatttatatatatatgtttgtgtttaattatgtatatgaa180
atggtgtttttggattaatggaatgcaggaagaaggggaactcatatgggatggtgattg240
ttgaggagaaggaggttgttgaggttgtggaaatggcaactgctgctgagaatgatgggt300
gcaagtgtgctgctaattgcacttgtgctggttgcaactgcggcaagtgatcatatatta360
attaccctttaattatgtagtaattaataagatgtaagccagagttacactggctttctg420
cctctcttttattttaatgaa441
<210>4
<211>23
<212>dna
<213>artificial
<400>4
acaacacaagccattcaatctat23
<210>5
<211>24
<212>dna
<213>artificial
<400>5
ttcattaaaataaaagagaggcag24
<210>6
<211>21
<212>dna
<213>artificial
<400>6
atgggatggtgattgttgagg21
<210>7
<211>21
<212>dna
<213>artificial
<400>7
cccatcattctcagcagcagt21
<210>8
<211>31
<212>dna
<213>artificial
<400>8
cgcgaattcatgtctagctgtggcaactgtg31
<210>9
<211>26
<212>dna
<213>artificial
<400>9
cccctcgagtcacttgccgcagttgc26
- 还没有人留言评论。精彩留言会获得点赞!