用于高容量离子交换生物分离的接枝海岛型非织造物的制作方法
公开领域
本发明涉及经修改用于生物分离过程的聚合物接枝和官能化非织造膜,以及形成和使用该膜的方法。
背景技术:
作为生物分离的平台,与传统填充床色谱相比,膜色谱提供了几个潜在的优势。与填充床相比时,膜的相互连接的孔允许高的体积通过速率而没有显著的压降。色谱树脂需要填充,它们通常不是一次性的,因此它们需要经过验证的清洁和再生过程才能使用。另一方面,许多膜可以使用可缩放的生产技术由聚合物制成,使得它们能够用作可堆叠、即用的一次性生物分离过滤器。非织造膜对于这些应用特别具有吸引力,因为它们经过高度工程化以使用高速制造技术用低成本材料表现出可控的孔隙率、纤维直径和孔径。到膜的蛋白质结合主要受限于由流动和吸附均可用的孔所产生的表面积。这消除了所有对吸附的扩散限制,但是与色谱树脂相比,它也降低了膜的结合能力。商用非织造物具有色谱树脂表面积的一部分,导致对大多数目标蛋白质捕获应用的结合能力低。通过将聚合物刷束缚在非织造膜中的纤维表面上,可以产生能够显著增加总蛋白质结合能力的三维结合域。在传统的色谱树脂、中空纤维膜、流延膜和非织造膜中,已知聚合物刷接枝将蛋白质吸附能力提高到单层覆盖的蛋白质吸附能力的几倍。
聚合物接枝可以显著改变载体的表面性质。它可以帮助调整表面的极性,以减少或增加生物分子的吸附,并且,在支持界面上引入的三维微环境中,它可以用于引入配体或间隔臂连接用的官能团。在liu等人先前进行的研究中,将甲基丙烯酸缩水甘油酯(gma)单体成功接枝到市售的聚对苯二甲酸丁二醇酯(pbt)非织造织物上。参见h.liu,y.zheng,p.gurgel,r.carbonell,affinitymembranedevelopmentfrompbtnonwovenbyphoto-inducedgraftpolymerization,hydrophilizationandligandattachment(由pbt非织造物通过光诱导接枝聚合、亲水化和配体连接的亲合膜开发),j.membr.sci.428(2013)562-575。使用uv诱导的自由基聚合,在单独的pbt纤维周围实现均匀和共形的聚gma接枝。将二苯甲酮(bp)用作引发剂引发gma聚合,聚gma通过夺氢直接连接到pbt表面。
pbt用作聚gma接枝的起始材料是有利的,因为它不需要将常用于非织造织物生产中的许多聚烯烃接枝所需的单独的表面uv预处理。pbt非织造材料本质上固有地为疏水性,导致高度非特异性蛋白质吸附,使得基材本身成为差的生物分离平台。使用酸性条件在pbt上直接水解聚gma接枝物使得纤维表面完全亲水并且显著降低非特异性疏水蛋白质吸附。gma的每个单体单元含有环氧端基,可用于通过以可用的胺、硫醇和羟基亲核取代来共价连接配体。在liu等人的研究中,还发现共价连接到聚gma刷上的二甘醇通过非特异性疏水相互作用基本上消除了蛋白质吸附。
聚gma接枝的非织造物为开发有效的离子交换膜提供了方便的平台。saito等人成功地将聚gma刷接枝到聚丙烯织物和聚乙烯中空纤维。参见k.saito,t.kaga,h.yamagishi,s.furasaki,t.sugo,j.okamoto,phosphorylatedhollowfiberssynthesizedbyradiationgraftingandcrosslinking(通过辐射接枝和交联合成的磷酸化中空纤维),j.membr.sci.43(1989)131-141。这些接枝的材料用磷酸基团官能化以开发强阳离子交换膜来捕获二价金属阳离子。
在zheng等人的研究中,将聚gma接枝到聚丙烯非织造物上并用二乙胺(dea)官能化,以开发弱阴离子交换剂。参见y.zheng,h.liu,p.gurgel,r.carbonell,polypropylenenonwovenfabricswithconformalgraftingofpoly(glycidylmethacrylate)forbioseparations(用于生物分离的具有聚甲基丙烯酸缩水甘油酯的共形接枝的聚丙烯非织造织物),j.membr.sci.364(2010)362-371。该材料达到了对牛血清白蛋白质(bsa)为120mg/g膜的平衡结合能力。
liu等人研究了在非织造pbt上不同程度的聚gma接枝对于通过阴离子捕获bsa的效果。参见h.liu,surfacemodifiednonwovenmembranesforbioseparations(用于生物分离的表面改性非织造膜),raleighncusa,northcarolinastateuniv.,博士论文,2012。在该研究中,聚gma接枝物用dea转化成弱阴离子交换剂并用bsa检验(challenge)。确定总体蛋白质结合能力随着接枝度而增加(%重量增量)。在12%聚gma重量增量下观察到800mg/g的最大平衡结合能力。该研究还显示需要几个小时到一整天的停留时间以达到最大的结合,并且这些结合时间随着增加的接枝重量%增量而增加。这些长的停留时间排除了使用这些聚gma接枝的非织造pbt膜来开发用于下游加工的高通量、高容量蛋白质捕获装置。
因此,仍然需要能够具有高通量、高容量蛋白质捕获的接枝非织造膜。
发明概述
在本发明的一个实施方案中,平均纤维直径小于约1.5微米的聚对苯二甲酸丁二醇酯(pbt)非织造物,可通过例如紫外线诱导的自由基聚合,容易地用甲基丙烯酸缩水甘油酯(gma)或类似的甲基丙烯酸酯聚合物接枝,以在每根纤维周围产生均匀和共形的聚合物刷网络,其可以进行化学改性以起到阴离子或阳离子交换剂的作用。在根据本发明的平均纤维直径约1μm的海岛型(i/s)pbt非织造物中和在平均纤维直径约3μm的市售pbt非织造物中,测量通过离子交换的蛋白质吸附速率。两种非织造物都成功接枝聚gma,并且在类似的重量%接枝下显示出相似的离子交换平衡蛋白质结合能力。然而,本发明的接枝i/s非织造膜在早期阶段表现出显著更高量的蛋白质结合,并且能够以具有较大纤维直径的接枝商用非织造物所需的一部分时间达到平衡。尽管不受操作理论的束缚,但由本发明的i/spbt非织造物观察到的蛋白质吸附速率较快,这被认为是由于纤维周围的聚gma接枝层的厚度薄于具有相同重量%接枝的商业pbt。
根据一个方面,本发明提供经修改用于生物分离过程的聚合物接枝和官能化的非织造膜,其包含非织造纤维网,所述非织造纤维网包含多根平均纤维直径小于约1.5微米(更通常约1微米或更小)的聚酯纤维(例如聚对苯二甲酸丁二醇酯纤维),所述多根聚酯纤维中的每一根具有接枝在其上的由甲基丙烯酸酯聚合物(例如聚gma)构成的多个聚合物链段,每个聚合物链段带有经修改以结合到目标分子的官能团。在某些实施方案中,甲基丙烯酸酯聚合物由选自甲基丙烯酸缩水甘油酯、甲基丙烯酸、甲基丙烯酸2-(二乙基氨基)乙酯、[2-(甲基丙烯酰氧基)乙基]三甲基氯化铵、甲基丙烯酸2-羟基乙酯、2-丙烯酰氨基-2-甲基丙磺酸、甲基丙烯酸2-(二甲基氨基)乙酯、甲基丙烯酸丁酯、甲基丙烯酸3-氯-2-羟基丙酯、甲基丙烯酸2-乙基己酯以及它们的组合中的一种或多种单体构成。官能团可以根据目标分子而变化,但通常是经修改以与目标分子进行阳离子或阴离子交换的基团。
在某些实施方案中,非织造纤维网包含在去除海岛型双组分纤维的海组分之后保留的岛纤维。非织造纤维网可具有至少约1.5m2/g的示例性bet比表面积。典型地,接枝聚合物链段的重量是非织造纤维网重量的约2至约50%,最优选5-25%重量增量。
另一方面,提供了生物分离方法,其包括使包含目标分子(例如蛋白质)的溶液通过本发明的非织造膜,使得溶液中的目标分子的至少一部分结合到非织造膜。
另一方面,本发明提供了一种制备经修改用于生物分离过程的聚合物接枝和官能化的非织造膜的方法,包括:接收非织造纤维网,所述非织造纤维网包含:多根海岛型纤维,或在去除双组分海岛型纤维的海组分之后剩余的多根岛纤维;任选地,去除双组分海岛型纤维的海组分以暴露其岛纤维;将甲基丙烯酸酯聚合物接枝到岛纤维的表面上以形成与其共价连接的多个聚合物链段,从而形成接枝的岛纤维,接枝步骤包括:使非织造纤维网与包含引发剂和至少一种甲基丙烯酸酯单体的溶液接触,以及将非织造纤维网暴露于紫外光以引发甲基丙烯酸酯单体的聚合;以及任选地,将接枝的岛纤维官能化,以将经修改以与目标分子结合的至少一个官能团连接到接枝的岛纤维的多个聚合物链段中的每一个。接枝溶液中单体的浓度可以变化,但通常为约5至约50%v/v(最优选15至25%v/v),引发剂(如二苯甲酮)通常以引发剂与单体的摩尔比为约1:100至约1:5(例如1:20)存在。紫外光源可以具有200nm至500nm(最优选365nm)的波长以引发聚合。引发聚合的特定波长取决于所使用的光引发剂以及要接枝到pbt表面的单体。此外,取决于接枝所需的能量和要进行接枝以达到所需聚合度的所需速率,紫外光源可以具有1-30mw/cm2(最优选5mw/cm2)的强度。
本发明包括但不限于以下实施方案:
实施方案1:一种经修改用于生物分离过程的聚合物接枝和官能化的非织造膜,其包含非织造纤维网,所述非织造纤维网包含多根平均纤维直径小于约1.5微米的聚酯纤维,所述多根聚酯纤维中的每一根具有接枝在其上的由甲基丙烯酸酯聚合物构成的多个聚合物链段,每个聚合物链段带有经修改以结合到目标分子的官能团。
实施方案2:任何前述或下述实施方案的膜,其中所述甲基丙烯酸酯聚合物由选自以下的一种或多种单体构成:甲基丙烯酸缩水甘油酯、甲基丙烯酸、甲基丙烯酸2-(二乙基氨基)乙酯、[2-(甲基丙烯酰氧基)乙基]三甲基氯化铵、甲基丙烯酸2-羟基乙酯、2-丙烯酰氨基-2-甲基丙磺酸、甲基丙烯酸2-(二甲氨基)乙酯、甲基丙烯酸丁酯、甲基丙烯酸3-氯-2-羟基丙酯、甲基丙烯酸2-乙基己酯及其组合。
实施方案3:任何前述或下述实施方案的膜,其中聚酯纤维由聚对苯二甲酸丁二醇酯构成。
实施方案4:任何前述或下述实施方案的膜,其中平均纤维直径为约1微米或更小。
实施方案5:任何前述或下述实施方案的膜,其中所述官能团经修改以与所述目标分子进行阳离子或阴离子交换。
实施方案6:任何前述或下述实施方案的膜,其中所述非织造纤维网包含在去除海岛型双组分纤维的海组分之后保留的岛纤维。
实施方案7:任何前述或下述实施方案的膜,其中非织造纤维网具有至少约1.5m2/g的bet比表面积。
实施方案8:任何前述或下述实施方案的膜,其中接枝聚合物链段的重量为非织造纤维网重量的约2至约50%。
实施方案9:一种生物分离方法,包括使包含目标分子的溶液通过任何前述或下述的实施方案的非织造膜,使得溶液中的目标分子的至少一部分结合到非织造膜。
实施方案10:任何前述或下述实施方案的方法,其中目标分子是蛋白质。
实施方案11:一种制备经修改用于生物分离过程的聚合物接枝和官能化的非织造膜的方法,包括:i)接收非织造纤维网,所述非织造纤维网包含:多根海岛型纤维,或在去除双组分海岛型纤维的海组分之后剩余的多根岛纤维;ii)任选地,去除双组分海岛型纤维的海组分以暴露其岛纤维;iii)将甲基丙烯酸酯聚合物接枝到岛纤维的表面上以形成与其共价连接的多个聚合物链段,从而形成接枝的岛纤维,接枝步骤包括:使非织造纤维网与包含引发剂和至少一种甲基丙烯酸酯单体的溶液接触,以及将所述非织造纤维网暴露于紫外光或热以引发所述甲基丙烯酸酯单体的聚合;和iv)任选地,将接枝的岛纤维官能化,以将经修改以与目标分子结合的至少一个官能团连接到接枝的岛纤维的多个聚合物链段中的每一个。
实施方案12:任何前述或下述实施方案的方法,其中所述岛纤维由聚对苯二甲酸丁二醇酯构成,并且所述甲基丙烯酸酯聚合物是聚gma。
实施方案13:任何前述或下述实施方案的方法,其中溶液中单体的浓度为约5至约50%v/v,引发剂以约1:100至约1:5的引发剂与单体的摩尔比存在。
实施方案14:任何前述或下述实施方案的方法,其中引发剂是二苯甲酮。
通过阅读以下详细描述以及下面简要描述的附图,本公开的这些和其他特征、方面和优点将变得显而易见。本发明包括上述实施方案中的两个、三个、四个或更多个的任何组合,以及本公开中阐述的任何两个、三个、四个或更多个特征或要素的组合,而不管这类特征或要素是否明确地组合在本文所述的具体实施方案中。本公开旨在从整体上阅读,使得所公开的发明在其各个方面和实施方案中的任何可分离的特征或要素应视为旨在可以组合,除非上下文另有明确规定。本发明的其它方面和优点将从以下变得显而易见。
附图简述
为了提供对本发明的实施方案的理解,参考附图,附图不一定按比例绘制。附图仅是示例性的,不应被解释为限制本发明。
图1a和1b是(a)具有36个岛的i/s纤维和(b)具有108个岛的i/s纤维的横截面图;
图2a和2b是108岛i/spbt非织造物(a)pla去除前和(b)pla去除后的sem图像;
图3说明了在pla去除后,商用pbt非织造物和108i/s非织造物在各种uv暴露时间下的聚gma接枝度;
图4a-4d是(a)接枝之前的商用pbt非织造物、(b)接枝前的108i/spbt非织造物、(c)接枝到20%重量增量的商用pbt非织造物和(d)接枝到20%重量增量的108i/spbt非织造物的sem图像;
图5a-5c是(a)接枝到20%重量增量的商用pbt、(b)接枝到5.9%重量增量的商用pbt非织造物和(c)接枝到20%重量增量的108i/spbt非织造物的纤维直径和干燥接枝物层厚度的横截面示意图;
图6说明了以各种程度接枝并且官能化为阳离子或阴离子交换剂以分别结合igg和bsa的108i/spbt非织造物和商用pbt非织造物的平衡结合能力;
图7说明了阴离子交换官能化接枝非织造物在各种接触时间下的bsa捕获:以20%和5.9%重量增量接枝的商用pbt,以及以20%重量增量接枝的108i/spbt;
图8说明了阳离子交换官能化接枝非织造物在各种接触时间下的higg吸附:以18%和5.3%重量增量接枝的市售pbt,以及以18%重量增量接枝的108i/spbt;
图9说明了对于所有测试bsa和higg结合速率的离子交换官能化接枝非织造物,接触时间15分钟以下的蛋白质结合;
图10是随时间用蛋白质填充的接枝pbt纤维的横截面示意图,随着时间(t)的进展,将可用聚gma“芯”转化为饱和的蛋白质/聚gma壳,r1=pbt纤维半径,rc=芯半径,r2=接枝纤维半径;
图11a和11b示出了(a)由阴离子交换官能化聚gma非织造物的转化的实验数据计算出的ψ值,相对时间标绘,带有最佳拟合线,和(b)由阳离子交换官能化聚gma非织造物的转化的实验数据计算出的ψ值,相对时间标绘,带有最佳拟合线;
图12是作为时间函数的bsa吸附到阴离子交换聚gma的实验和收缩芯模型结果(商用pbt在20%重量增量下:φ=0.75和τ=6716分钟,商用pbt在5.9%重量增量下:φ=0.84和τ=3109分钟,以及108i/spbt在20%重量增量下:φ=0.75和τ=1839分钟);和
图13是作为时间函数的higg吸附到阳离子交换聚gma“芯”的实验和收缩芯模型结果(商用pbt在18%重量增量下:φ=0.71和τ=3789分钟,商用pbt在5.3%重量增量下:φ=0.88和τ=4955分钟,以及108%i/spbt在18%重量增量下:φ=0.74和τ=3189分钟)。
发明详述
现在将参照附图在下文中更全面地描述本发明。本发明可以以许多不同的形式实施,并且不应该被解释为限于在此阐述的实施方案;相反,提供这些实施方案是为了使得本公开将满足适用的法律要求。同样的数字始终指代同样的元素。如在本说明书和权利要求中所使用的,除非上下文另外明确指出,否则单数形式“一”,“一个”和“该”包括复数指示物。
本发明使用非织造纤维网作为基材来构建能够用于生物分离的官能化膜,例如使用离子交换或亲和色谱法从某些溶液中分离蛋白质。用于本发明的非织造纤维网的平均纤维直径处于或接近纳米纤维范围。纳米纤维非织造物潜在商业化生产的解决方案是在纺粘工艺中使用双组分纤维来生产非织造垫。参见例如n.fedorova,b.pourdeyhimi,highstrengthnylonmicro-andnanofiberbasednonwovensviaspunbonding(经由纺粘的高强度尼龙微纤维和纳米纤维基非织造物),j.appl.polym.sci.104(2007)3434-3442。在这种生产方案中,两种聚合物可以从同一喷丝头共挤出,在那里它们结合成为粘合纤维。纤维也可以以桔瓣型或芯鞘型构造挤出。然后可以使这些纤维断裂以释放许多直径小得多的纤维,或者可以选择性地溶解聚合物之一,在非织造基质中留下一组小得多的纤维。参见例如a.durany,n.anantharamaiah,b.pourdeyhimi,highsurfaceareanonwovensviafibrillatingspunbondednonwovenscomprisingislands-in-the-seabicomponentfilaments:structure-process-propertyrelationships(经由将包含海岛型双组份长丝的纺粘非织造物原纤化的高表面积非织造物:结构-工艺-性能关系)j.mater.sci.44(2009)4926-5934。
如本文所用,术语“纤维”被定义为纺织品的基本元素,其具有例如至少约100倍的高纵横比。另外,“长丝/连续长丝”是长度非常长的连续纤维,其具有非常高的纵横比。术语“多组分纤维”是指包含物理或化学性质方面不同的两种或更多种聚合物的纤维,包括双组分纤维。本文所用的术语“非织造物”参照纤维材料、纤维网(web)、垫子、毡或片材,是指其中纤维以不明确或随机取向排列的纤维结构。根据本发明的纤维可以变化,并且包括具有任何类型的横截面的纤维,包括但不限于圆形、矩形、正方形、椭圆形、三角形和多叶形。在某些实施方案中,纤维可具有一个或多个空隙空间,其中空隙空间可具有例如圆形、矩形、正方形、椭圆形、三角形或多叶形横截面。
生产非织造纤维网的手段可以改变。一般而言,非织造纤维网通常以三个阶段生产:纤维网形成、粘合和整理处理。纤维网形成可以通过本领域已知的任何手段来完成。例如,纤维网可以通过干法成网工艺、纺丝成网工艺或湿法成网工艺形成。在本发明的各种实施方案中,非织造纤维网是通过纺粘工艺制成的。纺粘可以使用各种类型的纤维纺丝工艺(例如,湿法、干法、熔化或乳化)。熔融纺丝是最常使用的,其中聚合物被熔化成液态并被迫通过小孔进入冷空气,使得聚合物股线根据孔的形状凝固。然后将由此制得的纤维束拉伸,即机械拉伸(例如以3-5倍)以定向纤维。然后通过将拉伸的纤维沉积到移动的带上来形成非织造纤维网。例如在appel等人的美国专利no.4,340,563、dorschner等人的美国专利no.3,692,618、matsuki等人的美国专利no.3,802,817、kinney的美国专利no.3,338,992和3,341,394、hartmann的美国专利no.3,502,763和dobo等人的美国专利no.3,542,615中描述了一般的纺粘工艺,所有这些都通过引用并入本文。例如,纺粘通常产生比熔喷更大直径的长丝。例如,在一些实施方案中,纺粘产生具有约20微米或更大的平均直径的纤维。
各种方法可用于处理多组分纤维以获得具有较小直径(例如,小于约1.5微米、小于约1.0微米或小于约0.5微米)的纤维。尽管这些方法通常应用于通常具有较大直径的纺粘材料,但是应当指出,它们也可以应用于熔喷材料以及通过其它手段制备的纤维材料。例如,在一些实施方案中,生产可分裂的多组分纤维(例如,包括但不限于桔瓣型、带型、海岛型或多叶形),随后分裂或原纤化以提供具有较小直径的两根或更多根纤维。这种纤维可以被分裂的手段可以变化,并且可以包括向纤维赋予机械能量的各种工艺,例如水刺。例如在pourdeyhimi等人的美国专利no.7,981,226中描述了用于该工艺的示例性方法,该专利通过引用并入本文。
如上所述,在某些实施方案中,产生多组分纤维并随后处理(例如通过使纤维与溶剂接触)以去除一种或多种组分。例如,在某些实施方案中,可以生产和处理海岛型纤维以溶解海组分,将岛作为具有较小直径的纤维留下。例如在kato等人的美国专利no.4,612,228中描述了用于这种类型的工艺的示例性方法,该专利通过引用并入本文。
海岛型(i/s)非织造技术是芯鞘型双组分长丝工艺的延伸。这种类型的非织造物在被称为“海”的牺牲性聚合物鞘中嵌入的被称为“岛”的纤维内具有许多永久性聚合物芯。在本发明中,岛的数量和岛与海组分之比没有特别限制,岛的数目的示例性范围包括约20个至约400个岛(例如,约50个至约200个岛)。图1a和1b是两个示例性i/s结构的横截面图,一个具有36个岛,一个具有108个岛。“岛”与“海”之比可以在例如约25:75(w:w)至约75:25(w:w)的范围内。与许多聚合物(例如,尼龙-6和pbt)相比,聚乳酸(pla)具有较低的熔化温度,并且可以用热苛性碱浴容易地分解,使其成为可溶解的“海”的很好的候选物。与通过熔喷或纺粘技术制造的市售非织造物相比,i/s非织造物能够实现更小的纤维直径,因此具有更高的比表面积,同时仍保持纤维生产的高生产率和尺寸稳定性。
这样制造的纤维网可具有不同的基重。在一些实施方案中,非织造纤维网的基重为约200g/m2或更小、约150g/m2或更小、约100g/m2或更小、或约50g/m2或更小。在某些实施方案中,非织造织物具有约75g/m2至约125g/m2的基重。可以例如使用题为“织物的每单位面积重量的标准测试方法”的astmd3776/d3776m-09ae2中概述的测试方法来测量织物的基重。该测试报告了单位面积质量的测量,并以克每平方米(g/m2)来测量和表示。
非织造纤维网可以具有至少约1.5m2/g,例如至少约2.0m2/g或至少约2.2m2/g的示例性bet比表面积。示例性的bet表面积范围为约1.5m2/g至约3.0m2/g。
非织造纤维网的聚合物可以变化,但通常包含非常适合接枝的热塑性聚合物。示例性的聚合物包括聚烯烃(例如聚乙烯或聚丙烯)、聚酯和聚酰胺。聚酯是特别有用的,包括聚对苯二甲酸丁二醇酯(pbt)、聚对苯二甲酸三亚甲酯(ptt)、聚对苯二甲酸乙二醇酯(pet)、共聚酯及其组合。pbt是特别有用的,因为pbt的聚合物结构对用于接枝反应的某些引发剂(如二苯甲酮)具有天然的亲和性。为此,当使用pbt时,不需要将官能团引入到聚合物的预处理步骤来增强聚合物与引发剂之间的亲和力。
如上所述,非织造纤维网的聚合物纤维经受接枝过程,由此聚合物刷或链段共价连接到纤维。该过程通常需要使非织造纤维网与包含溶解于合适溶剂中的单体以及自由基聚合引发剂(例如光引发剂(例如二苯甲酮))的溶液接触。该过程通常也需要使非织造纤维网经受波长200-500nm(最优选365nm)和强度1-30mw/cm2(最优选5mw/cm2)的紫外光以引发聚合反应。接枝溶液中单体的浓度可以变化,但通常为约5至约50%v/v(最优选15-25%(v/v)),引发剂如二苯甲酮通常以引发剂与单体的摩尔比为约1:100至约1:5(例如1:20)存在。在某些实施方案中,允许聚合反应进行直到接枝聚合物链段的重量为非织造纤维网重量的约2至约50%(最优选5-25%重量增量)。
用于接枝的聚合物可以变化,但通常是丙烯酸酯或甲基丙烯酸酯聚合物。接枝聚合物为非织造纤维网的纤维提供刷状延伸,其可以被官能化以增强对某些目标分子的亲和力。用于接枝聚合物的单体的选择可以变化,并将部分取决于最终膜结构所需的期望结合性能。某些单体将固有地携带可用于亲和或离子交换结合的官能团,而其他单体将需要进一步官能化以添加必要的结合基团。示例性单体及其可能的用途包括:甲基丙烯酸缩水甘油酯(适合进一步官能化)、甲基丙烯酸(弱阳离子交换膜)、甲基丙烯酸2-(二乙氨基)乙酯(弱阴离子交换膜)、[2-(甲基丙烯酰氧基)乙基]三甲基氯化铵(强阴离子交换膜)、甲基丙烯酸2-羟基乙酯(hema、耐蛋白质膜)、2-丙烯酰氨基-2-甲基丙磺酸(强阳离子交换膜)、甲基丙烯酸2-(二甲氨基)乙酯(弱阴离子交换膜)、甲基丙烯酸丁酯(疏水相互作用膜)、甲基丙烯酸3-氯-2-羟基丙酯(适用于进一步官能化)、甲基丙烯酸2-乙基己酯(疏水相互作用膜)及其组合。
如果需要的话,聚合物链段或刷可以被官能化,使得每个聚合物链段带有经修改用于与目标分子结合的官能团。可以在这样的官能团和目标分子(例如蛋白质)之间发生的示例性结合可以包括离子键、氢键和范德华力。示例性官能团包括胺基(包括伯胺、仲胺、叔胺或季胺)、磺酸基团、羧酸基团、磷酸根基团等。连接这种官能团的衍生化反应通常包括使聚合物刷上的环氧基团或其它反应性基团与含有所需官能团的分子反应。
如实验部分中更充分阐述的那样,发明人成功地将完整的和共形的聚gma刷接枝到108岛i/spbt非织造纤维网上。接枝的非织造物分别被成功衍生为弱阴离子和强阳离子交换剂以捕获bsa和higg。对于18-20%聚gma重量增量获得的平衡静态蛋白质结合能力高达1000mg/g,这表明聚gma刷能够比单层覆盖增加数倍的蛋白质捕获。据观察,具有较薄聚gma接枝物的较高表面积108i/spbt非织造物能够降低聚gma/蛋白质层中扩散限制的显著性,导致与商用pbt非织造物相比,到达平衡的时间更短。此外,与商用pbt非织造织物相比,108i/s非织造pbt表现出更高量的初始蛋白质结合,这在需要短停留时间的应用中是有利的。
在某些实施方案中,本发明的聚合物接枝和官能化非织造膜特征可在于非常高的蛋白质结合能力,例如在至少约18%的接枝聚合物重量增量(例如包含18-20重量%接枝聚合物的非织造物,基于该非织造纤维网的重量)下,至少约800mg/g、至少约850mg/g或至少约900mg/g蛋白质的能力。另外,本发明的非织造纤维网特征可在于在短接触时间内结合显著量的蛋白质,例如在至少约18%的接枝聚合物重量增量(例如包含18-20重量%接枝聚合物的非织造物,基于该非织造纤维网的重量)下,不超过约1分钟的接触时间内至少约200mg/g、至少约250mg/g或至少约300mg/g的蛋白质结合。
实验
在非织造物研究所(nwi,北卡罗莱纳州立大学,raleigh,nc)的中试设施上生产了108个岛计数的海岛型非织造pbt织物。岛计数指的是一旦pla“海”被去除,被解放的离散pbt纤维的数量。i/s非织造物以100g/m2基重制造,由50%pla作为“海”聚合物和50%pbt作为“岛”聚合物组成,“海”去除后的基重为50g/m2。macopharma(法国tourcoing)提供市售的基重为52g/m2的熔喷pbt非织造物。甲基丙烯酸缩水甘油酯(gma)购自pflatz&bauer(waterbury,ct)。gma中的抑制剂通过去除氢醌和单甲基醚氢醌的预填充抑制剂去除柱(sigmaaldrich,st.louis,mo)去除。二苯甲酮(bp)购自sigmaaldrich(st.louis,mo)。氢氧化钠、1-丁醇、tris碱、盐酸、氯化钠和三水合乙酸钠购自fisherscientific(fairlawn,nj)。四氢呋喃(thf)、甲醇、硫酸和乙酸购自bdh(westchester,pa)。二乙胺(dea)购自alfaaesar(wardhill,ma)。磷酸(85%)购自acrosorganics(fairlawn,nj)。固相萃取管购自supelco(bellefonte,pa)。牛血清白蛋白质(bsa)购自sigmaaldrich(st.louis,mo)。人免疫球蛋白质g(higg)购自equitek-bioinc.(kerrville,tx)。
pla从i/s非织造物去除
在接枝之前,108i/s非织造物的50%pla“海”必须被去除以解放pbt“岛”。在80-90℃下使用在去离子水中的10%w/w氢氧化钠分解pla。将i/s非织造物在苛性碱浴中浸没5分钟并持续搅拌,直到所有的pla都由非织造织物溶解。然后用去离子水彻底清洗pbt非织造物,直至达到中性ph。然后使样品风干过夜。图2a和2b分别显示了在去除pla之前和之后的108i/spbt非织造物。
在图2a中,pla仍然存在,包封pbt“岛”,显示在pla去除之前直径约15μm的初始较大的纤维。图2b显示在pla去除之后的非织造物,以解放直径约1μm的108根离散的pbt纤维。pla去除后释放的pbt纤维保持原始pla纤维的大体方向。
uv诱导聚gma接枝到pbt非织造物上
gma接枝溶液由在作为溶剂的1-丁醇中的20%v/vgma单体组成。将光引发剂二苯甲酮(bp)以1:20(mol:mol)的bp:gma之比加入到接枝溶液中。将去除pla后的市售pbt非织造物和108i/spbt非织造物切割成75×50mm大小的样品并在接枝前称重,样品对于商用pbt和108i/spbt分别为大约200mg和180mg。通过经由注射器喷洒1.5-2.0ml的接枝溶液使非织造物被接枝溶液饱和,并置于两片硼硅酸盐载玻片(75×50mm)之间。使用具有365nm波长和5mw/cm2强度的uv灯(型号en-180l,spectronicscorporation,westbury,ny)诱导gma自由基聚合到pbt表面上。灯和样品之间的距离是3mm。以各种暴露时间对样品进行照射以实现不同程度的聚gma接枝,具有不同的%重量增量。在聚gma接枝后,将样品置于含有100mlthf的烧瓶中,烧瓶连同thf和样品用超声波浴(bransonic3510r-mt,bransonultrasonicscorporation,danbury,ct)超声处理30分钟以去除任何未反应的接枝溶液或未束缚的聚gma。在thf洗涤之后,将样品从烧瓶中取出并置于含有100ml甲醇的烧瓶中,将含有样品和甲醇的烧瓶用超声波浴超声处理10分钟以从非织造物中去除thf。在甲醇洗涤之后,从烧瓶中取出样品并使其在空气中干燥过夜。测量非织造物的最终重量,并使用关于接枝导致的%重量增量的方程式1确定聚gma接枝度。
在方程式1中,wi是接枝前的初始非织造物重量,wf是聚gma接枝的最终非织造物重量。方程式1中定义的%重量增量在本文给出的图中简写为%wt.增量。
聚gma接枝pbt非织造物的官能化
聚gma接枝的pbt非织造物通过如下进行官能化以产生弱阴离子交换剂:浸入50%v/v二乙胺(dea)水溶液中,从而在聚gma刷上产生叔胺。将180-200mg(75×50mm)的接枝pbt非织造物样品浸入100ml的dea溶液中。使用容纳在培养罩(certomat®hk,b.braunbiotechinternational,melsungen,德国)中的培育摇动器(certomat®rm,b.braunbiotechinternational,melsungen,德国)将反应保持在恒定的30℃并在100rpm下搅动。氨化后,将样品置于含有100ml去离子水的烧瓶中,将烧瓶置于超声波浴(bransonic3510r-mt,bransonultrasonicscorporation,danbury,ct)中5分钟,以去除过量的dea。超声处理之后,用新鲜去离子水置换去离子水洗涤液,重复该过程,直到用ph试纸证实为7.0的中性ph,洗涤10次确保所有dea已经从非织造物中去除。任何未反应的环氧基团通过将样品浸入100ml100mm硫酸过夜而水解。在环氧基团水解之后,将样品置于含有100ml去离子水的烧瓶中,将烧瓶置于超声波浴(bransonic3510r-mt,bransonultrasonicscorporation,danbury,ct)中5分钟以去除过量的硫酸。超声处理后,用新鲜的去离子水置换去离子水洗涤液,重复该过程,直到用ph试纸证实为7.0的中性ph,洗涤10次确保所有的硫酸已经从非织造物中去除。然后将样品风干过夜。
将聚gma接枝的pbt非织造物官能化,以通过将磷酸基团连接到聚gma刷上来产生强阳离子交换剂。将大约20mg(25×15mm)的接枝pbt非织造物样品浸入10ml85%w/w磷酸中并在80℃下孵育过夜(isotemp115,fisherscientific,fairlawn,nj)。官能化后,将样品置于含有100ml去离子水的烧瓶中,将烧瓶置于超声波浴(bransonic3510r-mt,bransonultrasonicscorporation,danbury,ct)中5分钟以去除过量的磷酸。超声处理后,用新鲜的去离子水置换去离子水洗涤液,重复该过程,直到用ph试纸证实为7.0的中性ph,洗涤5次确保所有磷酸已经从非织造物中去除。任何未反应的环氧基团通过将样品浸入10ml100mm硫酸过夜来水解。在环氧基团水解之后,将样品置于含有100ml去离子水的烧瓶中,将烧瓶置于超声波浴(bransonic3510r-mt,bransonultrasonicscorporation,danbury,ct)中5分钟以去除过量的硫酸。超声处理后,用新鲜的去离子水置换去离子水洗涤液,重复该过程,直到用ph试纸证实为7.0的中性ph,洗涤10次确保所有的硫酸已经从非织造物中去除。然后将样品风干过夜。
材料表征
为了确定平均纤维直径并评估uv接枝的有效性,使用hitachis-3200n可变压力扫描电子显微镜(vpsem)(hitachihightechnologiesamerica,inc.,schaumberg,il)获得扫描电子显微镜图像。在氩气中用pd/au溅射涂覆非织造物样品。使用加速电压为20kv,工作距离为33mm的显微镜捕获图像。使用来自4pianalysis,inc.(hillsborough,nc)的revolution软件在sem显微照片上测量横跨纤维直径的距离。通过测量sem显微照片的横跨150个随机纤维的距离来测定108i/spbt和市售pbt非织造物的平均纤维直径。
通过brunauer,emmetandteller(bet)多点分析,使用氮吸附测定去除pla海之后的108i/spbt非织造物和市售的熔喷非织造物的比表面积。将一克非织造材料装载到12mm样品架中,并在autosorbtm-1c化学吸附-物理吸附分析仪(quantachromeindustries,boyntonbeach,fla)上进行分析,测量39个氮分压点。
使用毛细管流动孔隙测定法测定市售pbt熔喷非织造物和去除pla后108i/spbt非织造物的平均孔径。在cfp-1100-ax毛细管流动孔隙度计(porousmaterialsinc.,ithaca,ny)上测试非织造物样品。润湿液是galwicktm(porousmaterialsinc.,ithaca,ny),表面张力为15.9达因/cm。
商用pbt非织造物的平均纤维直径为3000nm±900nm,108i/spbt非织造物的平均纤维直径为916nm±174nm。非织造物的比表面积使用用于氮吸附的bet法测定。根据bet分析,发现商用pbt非织造物具有0.86m2/g的比表面积,去除pla后的108i/spbt具有2.45m2/g的比表面积。平均流动孔径使用毛细管流动孔隙度计测定。商用pbt的平均流动孔径为8.73μm±3.10μm,pla去除后的108i/spbt的平均流动孔径为8.09μm±11.9μm。图2显示了去除pla“海”之前(a)和之后(b)的108i/spbt。可以注意到pla去除后的纤维很大程度上保持了它们原来的定向排列。这使得材料具有宽的孔径分布,保持了原始材料的孔结构,并且一旦pla被去除,就产生了更细孔的增加。
在各种程度的聚gma接枝下的静态平衡蛋白质吸附
以弱阴离子交换形式以及强阳离子交换形式测试市售pbt非织造物和108i/spbt非织造物在各种程度的聚gma覆盖下的平衡静态蛋白质结合能力。以2.5%、5.9%、7.2%、12%和20%重量增量接枝的市售pbt非织造物和以5.6%、12%和20%重量增量接枝的108i/spbt非织造物用dea官能化为弱阴离子交换剂。用纯bsa作为模型蛋白质对这些膜进行检验,以建立这些阴离子交换膜的静态平衡结合能力。bsa的分子量为66.5kda,等电点为4.7[sigmaaldrich,st.louismo]。将约20mg(25×15mm)非织造物样品置于3ml固相萃取(spe)管中,并用3ml低离子强度结合缓冲液(20mmtrishclph7.0)洗涤5次。样品在bsa结合之前在旋转器(tissueculturerotator,glas-col,terrehaute,in)上的结合缓冲液中平衡至少30分钟。一旦平衡,将3ml在20mmtrishclph7.0中的10mg/mlbsa加入到每个样品中并使其结合过夜15小时。ph7.0下的低离子强度缓冲液确保dea官能化的接枝pbt带正电荷且bsa带负电荷,以促进与最小量的会破坏蛋白质结合的离子的结合。结合后,样品用3ml20mmtrishclph7.0洗涤。需要用20mmtrishclph7.0洗涤5次以去除所有未结合的蛋白质,通过在第五次和最后一次洗涤中使用uv-vis光谱在280nm处可忽略量的蛋白质进行验证。使用高离子强度洗脱缓冲液3ml的20mmtrishclph7.0+1mnacl作为洗脱缓冲液洗脱结合的bsa。洗脱缓冲液中高浓度的离子有效破坏离子相互作用,从非织造物中去除蛋白质。收集洗脱级分,并使用uv-vis光谱在280nm处测定蛋白质浓度。静态平衡结合能力(meq,以蛋白质的质量/膜的质量计)值使用方程式2确定。
以类似的方式,通过用磷酸根基团官能化接枝的pbt非织造物来合成强阳离子交换膜。以5.3%、10%和18%重量增量接枝的市售pbt熔喷非织造物和以7%、12%和18%重量增量接枝的108i/spbt非织造物被官能化以产生强阳离子交换剂。用纯的多克隆higg作为模型蛋白质来检验这些膜,以确立这些阳离子交换膜的平衡结合能力。多克隆higg具有150kda的分子量和7-9的等电点[equitek-bio,kerrvilletx]。将大约20毫克(25×15毫米)的非织造物样品置于3mlspe管中,用3ml低离子强度结合缓冲液20mm乙酸盐ph5.5洗涤5次。在higg结合之前,将样品在旋转器(tissueculturerotator,glas-col,terrehaute,in)上的结合缓冲液中平衡至少30分钟。一旦达到平衡,向每个样品中加入3ml在20mm乙酸盐ph5.5中的10mg/mlhigg,并使其结合过夜15小时。在ph5.5下的低离子强度缓冲液确保磷酸官能化的接枝pbt带负电并且higg带正电,以促进与最小量的会破坏蛋白质结合的离子的结合。结合后,样品用3ml20mm乙酸盐ph5.5洗涤。需要用20mm乙酸盐ph5.5洗涤五次以去除所有未结合的蛋白质,通过在第五次和最后一次洗涤中使用uv-vis光谱在280nm处可忽略量的蛋白质进行验证。使用3ml的高离子强度洗脱缓冲液20mm乙酸盐ph5.5+1mnacl洗脱结合的higg。洗脱缓冲液中高浓度的离子有效破坏离子相互作用,从非织造物中去除蛋白质。收集洗脱级分,使用uv-vis光谱在280nm处测定蛋白质浓度。使用方程式2计算静态平衡结合能力。
蛋白质吸附动力学
这些实验目的在于确定接枝离子交换官能化非织造pbt膜上的蛋白质吸附速率。在该实验中,以相同程度的聚gma覆盖(%重量增量)以及相同的干燥聚gma接枝物厚度将市售pbt非织造物和108i/spbt非织造物接枝。假设接枝是均匀的和共形的,可以从样品的聚gma的%重量增量以及聚gma和pbt的密度估算聚gma接枝物的干燥厚度。这些假设允许将要处理的原始pbt纤维和聚gma接枝层的体积视为具有围绕圆柱形pbt内芯的圆柱形外接枝层的同心圆柱体。使用方程式(1)中定义的%重量增量,以及圆柱形聚gma接枝层的外部体积和pbt圆柱形芯的内部体积的表达式,可以推导近似干燥接枝层厚度(δ)的表达式,如方程式3所示。方程式3的完整推导可以在补充信息中找到:干燥聚gma接枝物厚度的推导。
在方程式3中,δ是干燥聚gma接枝物厚度,r1是已经接枝的特定pbt非织造物的平均纤维直径,ρpbt是pbt聚合物的密度(1.30g/cm3),并且ρ聚gma是干燥聚gma聚合物的密度(0.80g/cm3)。表1包含用于分析接触时间对蛋白质结合的影响的实验而制备的样品的列表,具有它们各自的%重量增量、干燥聚gma刷厚度和官能化类型。
表1:在具有特定接枝度、干燥接枝物厚度(δ)和离子交换功能的蛋白质吸附速率研究中使用的聚gma接枝非织造织物。
将约20mg(25×15mm)非织造物样品置于3mlspe管中,用结合缓冲液20mmtrishclph7.0进行充分洗涤,以bsa进行阴离子交换实验,或用20mm乙酸盐ph5.5进行充分洗涤,以higg进行阳离子交换实验。在蛋白质结合之前,将样品在旋转器(tissueculturerotator,glas-col,terrehaute,in)上的结合缓冲液中平衡至少30分钟。一旦样品平衡,它们分别用对阴离子交换非织造物的3ml10mg/mlbsa或对阳离子交换非织造物的3ml10mg/mlhigg进行检验。允许蛋白质以30秒至15小时之间的各种暴露时间进行结合。结合后,结合了bsa的阴离子交换样品用3ml20mmtrishclph7.0洗涤5次,结合了higg的阳离子交换样品用3ml20mm乙酸盐ph5.5洗涤5次,以去除任何未结合的蛋白质。使用3ml高离子强度洗脱缓冲液20mmtrishclph7.0+1mnacl洗脱bsa。使用3ml高离子强度洗脱缓冲液20mm乙酸盐ph5.5+1mnacl洗脱higg。使用uv-vis光谱在280nm处分析洗脱级分,并使用方程式2计算每种材料的蛋白质结合量。
pbt非织造物的接枝
商用pbt和108i/spbt非织造物以各种程度的聚gma覆盖成功地接枝。在增加的紫外线暴露时间下的接枝度的结果如图3所示。
从图3可以看出,108i/spbt非织造物的接枝速率比商用pbt非织造物更快。与商用pbt非织造物相比,108i/spbt非织造物的gma聚合引发可用面积多达2.85倍,导致约2.4倍的更高接枝速率。两种非织造物在不同的最低接枝度下表现出完全共形和均匀的聚gma接枝覆盖:商用pbt熔喷非织造织物为高于3%重量增量,108i/spbt非织造织物为高于6%重量增量。图4显示了108i/spbt非织造物和商用pbt非织造物在20%重量增量下接枝之前和之后的sem图像。
图4c和4d显示了归因于聚gma接枝的pbt纤维表面上的可见的增加的粗糙度,其在图4a和4b中对于未接枝的pbt非织造物不存在。除了提高聚gma接枝速率之外,108i/s纤维的较小直径对接枝层的厚度具有显著的影响。在图4c和d中,商用pbt和108i/spbt都具有相同的20%重量增量的聚gma接枝。然而,在108i/s非织造物的给定样品中存在更多的接枝可用表面积,导致比商用pbt非织造物更薄的接枝层厚度。商用pbt和108i/spbt的各种接枝度的实际干燥接枝物厚度使用方程式3计算,并列于表1中。图5显示了以20%重量增量接枝的商用pbt和108i/spbt与以5.9%重量增量接枝的商用pbt的干燥聚gma接枝物厚度的视觉示意比较。
图5中的视觉表示说明了:对于接枝的相同的%重量增量,相比于108i/spbt非织造物(图5c),如何需要较厚的聚gma接枝物层以在商用pbt非织造物上实现20%的重量增量(图5a)。还可以看出,为了使接枝的商用pbt非织造物与i/spbt非织造物具有相同的接枝层厚度,市售pbt的%重量增量需要小于i/s接枝非织造物的三分之一(比较图5b与图5c)。随后将会看到,接枝层厚度控制蛋白质吸附到膜上的速率。然而,平衡结合能力只是%重量增量的函数。
衍生化pbt非织造物的平衡蛋白质结合离子交换容量
商用pbt非织造物和108i/s非织造物以各种%重量增量接枝,并测定它们对bsa的阴离子交换捕获和higg的阳离子交换捕获两者的平衡蛋白质结合能力。这些实验的结果如图6所示。
图6的结果显示,对于阴离子交换膜和阳离子交换膜两者以及对于具有两种非常不同的分子量和大小的蛋白质,平衡蛋白质结合能力随着聚gma接枝度的增加(%重量增量)线性增加。108i/spbt非织造物的比表面积高达商用pbt非织造物的2.85倍,但它们都达到非常相似的平衡结合能力。这些结果表明,给予足够的时间,中等大小的蛋白质如bsa(66.5kda)和大蛋白质如higg(150kda)能够穿透进样品的接枝层,直到整个接枝层被蛋白质饱和。尽管这些蛋白质的分子量有很大不同,但bsa和higg具有相似的比体积(分别为0.733cm3/g和0.739cm3/g)。当商用pbt非织造物和i/spbt非织造物样品具有相同的原始质量和相同的%重量增量时,它们具有相同的接枝层的质量,因此不难理解基于质量基础的平衡蛋白质结合能力如何是一样的。这些结果还表明,在该实验中使用的结合溶液条件下的接枝层必须是足够柔韧的或足够多孔的,以允许甚至大的分子如具有100nm的回转半径的higg穿透。由于这些接枝层是高度带电的,在低离子强度的结合条件期间,接枝层可能处于延伸的构型,允许蛋白质穿透。
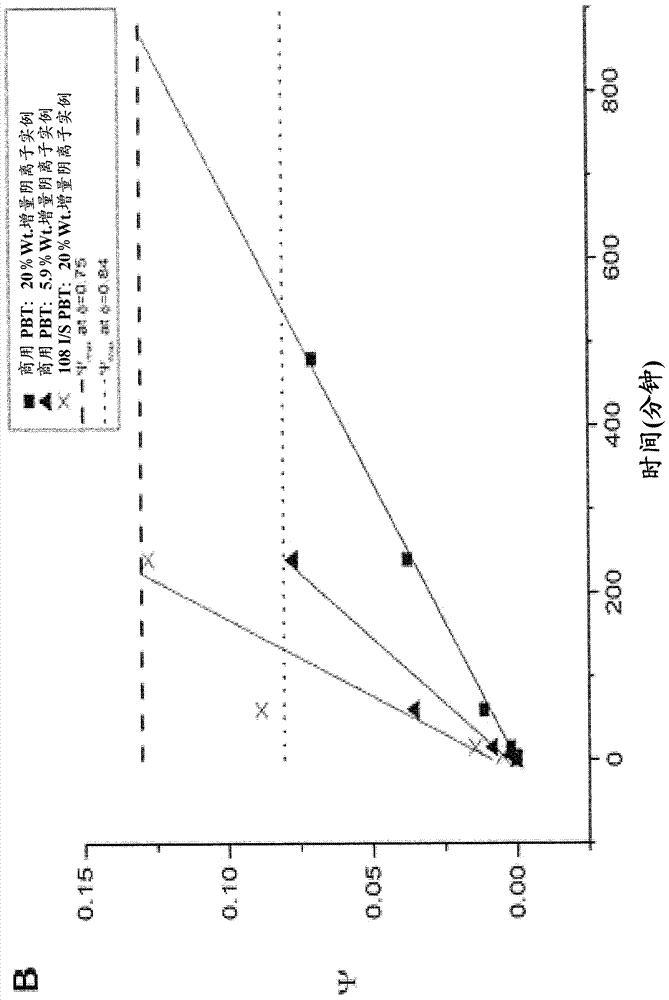
同样重要的是要注意,对于高%重量增量样品,平衡蛋白质结合能力非常大,在20%重量增量下达到每克非织造物超过800mg结合蛋白质的值。如果bsa和higg仅吸附在纤维的外表面上,而不能穿透到接枝层中,则结合能力会低得多。据报道bsa和higg的单层吸附分别在2.5-6mg/m2和2-5.5mg/m2的范围内的某处。考虑到商用pbt非织造物的这些单层覆盖数目和经测量的比表面积,bsa和higg的单层结合能力分别为5.2和4.7mg/g。类似地,108i/spbt非织造物上的bsa和higg的最大单层覆盖分别为14.7和13.5mg/g。图6中报道的所有平衡结合能力高达单层蛋白质吸附的50至200倍。这是非常有力的证据,聚gma刷创造了三维结合的环境,其中,对于单位膜质量的结合量,平衡蛋白质吸附随可用聚合物刷的量而按比例变化。图6显示具有相同聚gma接枝度的两种非织造物显示出相同的平衡蛋白质结合能力。例如,在20%重量增量下,商用pbt非织造物在平衡时捕获823±66mg/g的bsa,并且具有20%重量增量的108i/spbt非织造物通过阴离子交换捕获817±164mg/g的bsa。图5显示,商用pbt非织造物的20%重量增量聚gma干燥接枝物厚度(δ=227nm)比108i/spbt非织造物(δ=69nm)大。%重量增量决定了平衡结合能力,然而在商用pbt非织造物和108i/spbt非织造物之间,平衡时蛋白质的分布是非常不同的。在平衡时,与商用pbt非织造物相比,108i/spbt非织造物的蛋白质分布在更大的表面积上的更薄的层中,商用pbt非织造物由于该材料更低的比表面积而需要更厚的聚gma接枝层结合相同量的蛋白质。
对聚gma接枝阴离子和阳离子交换非织造物的吸附速率
在3.3节中,显示了离子交换官能化的聚gma接枝非织造物表现出非常高的平衡结合能力。然而,对于一些应用,有必要了解%重量增量和接枝层厚度对蛋白质吸附到聚gma刷层的动力学的影响。图7显示了在20%和5.9%wt.增量下的商用接枝pbt以及在20%wt.增量下的108i/s接枝的pbt在各种蛋白质暴露时间下,通过阴离子交换的bsa捕获的结果。选择这些接枝度是因为在20%重量增量下,商用pbt和108i/spbt两者具有相同量的聚gma接枝和图6中显示的平衡结合能力,并且以5.9%接枝的商用pbt具有与108i/spbt相同的聚gma接枝物厚度,如图5所示,并应具有相似的蛋白质吸附速率。没有研究以5.9%重量增量接枝的108i/s,因为它处于提供完全共形接枝所必需的接枝的下限,并且会导致测试蛋白质结合的材料的高度可变性。
这些实验是在静态或非流动条件下分批进行的。在图7中,对于在各种接触时间下的蛋白质捕获,比较了以20%重量增量接枝到相同的聚gma接枝度的市售pbt和108i/spbt非织造物的bsa吸附速率。在20%重量增量下,商用pbt非织造物和108i/spbt非织造物在接触时间充足的情况下会聚到相同的结合能力(〜800mg/g),与之前所示的结果一致。然而,可以看出bsa以比108i/spbt非织造物慢得多的速率吸附到20%重量增量的接枝商用pbt非织造物上。4小时后,以20%重量增量接枝的108i/spbt非织造物已经达到其平衡结合能力,与此相比,以20%重量增量接枝的商用pbt非织造物仅达到平衡结合能力的60%。图7还显示,以5.9%重量增量接枝的商用pbt非织造物在蛋白质接触时间4小时后也达到其平衡结合能力。根据方程式3且如图5所示,接枝到20%重量增量的108i/spbt非织造物和接枝到5.9%重量增量的商用pbt非织造物具有相同的干燥聚gma接枝物厚度70nm。这是良好证据,表明通过离子交换将蛋白质吸附到这些接枝非织造织物上的速率很大程度上取决于通过接枝层的扩散限制,并因此受到接枝层厚度的支配,后者又由非织造物的初始纤维直径和接枝材料的%重量增量决定。
还研究了higg通过阳离子交换吸附到接枝的商用pbt非织造物和108i/spbt非织造物的动力学。图8显示了在18%和5.3%wt.增量下的商用接枝pbt以及18%wt.增量下的108i/s接枝pbt阳离子交换剂在各种接触时间下higg捕获的结果。
图8显示了接枝到相同的聚gma接枝度(18%的重量增量)的商用pbt非织造物和108i/spbt非织造物,当官能化为阳离子交换剂时,如何会聚到类似的平衡higg结合能力。这与图7所示的阴离子交换膜的行为相同。接枝到18%重量增量的108i/spbt非织造物样品在4小时后达到平衡结合,与此相比,以18%重量增量接枝的商用pbt非织造物需要一整天的蛋白质接触。接枝到与以18%重量增量接枝的108i/spbt非织造物相同的接枝厚度(δ=63nm)的、具有5.3%重量增量的商用pbt非织造物在约4小时内也达到平衡。再一次,这些结果与图7中阴离子交换非织造物所见的结果完全类似,进一步证明观察到的动力学现象不依赖于接枝层电荷类型或被吸附的蛋白质的大小。
对于实际应用,重要的是要评估短暂停留时间下蛋白质如何结合。例如,如果这些膜要用于生物制品的下游纯化中的蛋白质捕获,则希望吸附装置中的停留时间为5分钟或更少。对上面讨论的阴离子和阳离子交换接枝非织造物,图9显示了图7和8中的bsa和higg接触时间在15分钟以下的蛋白质结合数据。
在图9中,明显的是在阳离子和阴离子交换模式下,与任何官能化接枝商用pbt非织造物相比,接枝108i/spbt非织造物均能够以非常短的接触时间结合更多的蛋白质。108i/spbt非织造物最初结合更多蛋白质的能力是由于该织物更高的比表面积。回想一下,pla去除后的i/spbt非织造物的比表面积是商用熔喷pbt非织造物的比表面积的2.85倍。将图9中存在的数据外推至时间t=0,以估计在发生任何进入聚gma层的扩散之前,由于蛋白质在非织造物表面区域上的吸附而瞬间发生的蛋白质结合(mi)的初始量。表2中给出了蛋白质吸附的初始量。
表2:由表面区域吸附引起的蛋白质吸附的初始量(mi)。
如果表2中给出的结果是官能化商用pbt非织造物(120mg/g平均初始蛋白质结合)和108i/spbt非织造物(325mg/g平均初始蛋白质结合)的平均值,则观察到108i/spbt非织造物最初相比商用pbt非织造物结合多达2.7倍的蛋白质。这与108i/spbt非织造物的实验表面积高达商用pbt非织造物的2.85倍紧密一致。由于这个原因,当需要非常短的停留时间时,使用接枝108i/spbt非织造物是有利的,因为它们可以严格由于较高的比表面积而获得相当高的结合能力。
阳离子和阴离子交换接枝非织造物两者的吸附速率测量结果表明吸附过程在这些官能化聚gma刷基质中扩散受限,需要几个小时达到平衡。为了使这些扩散问题最小化,对于相同的%重量增量,与较低表面积(较大纤维直径)材料相比,较高表面积(较小纤维直径)非织造织物导致较薄的接枝层厚度,如图5所示。较薄的聚gma层减少扩散距离并缩短达到平衡的时间,同时保持高的平衡结合能力,如图7和8所示。除了对于特定聚gma接枝度减少达到平衡的时间之外,如图9所示,较高比表面积108i/spbt非织造物也有助于较高的蛋白质吸附的初始量。整体平衡蛋白质结合能力将由聚gma接枝度决定,但是蛋白质捕获的有效速率由非织造物的初始比表面积和聚gma刷层的厚度决定。
蛋白质吸附到接枝聚gma非织造物样品的动力学模型
由上述阴离子和阳离子接枝层非织造物的蛋白质吸附的平衡和速率测量两者的结果提供的见解表明,有可能开发出描述吸附过程的数学模型,并且该数学模型应定性和定量地与实验结果一致。与离子交换介质中的传质速率限制相比,带电蛋白质对带电结合位点的吸附速率几乎是瞬时的。已经提出,在扩散到官能化聚合物层中的情况下,蛋白质吸附决定速率的步骤是进入聚合物层的穿透。图10中显示了蛋白质如何随着时间被吸附到pbt纤维周围接枝的官能化聚gma刷层的横截面图示。
由于吸附到接枝层的速率可能比蛋白质通过层的扩散速率快得多,因此以下推论并不是不合理的:一旦蛋白质分子结合到接枝层外表面上的带电基团,进入接枝层的另外的分子将不得不扩散通过含有结合蛋白质的层。一旦未结合的蛋白质扩散通过结合蛋白质层并遇到聚合物上的游离带电基团,它将立即结合,并且结合蛋白质层将随着时间生长直到它到达pbt纤维的表面。刚刚描述的现象与已经应用于色谱树脂中蛋白质扩散的传统收缩芯模型是一致的。可以将聚gma接枝层上的结合蛋白质看作形成“壳”,其对蛋白质传质到未结合聚gma的剩余层中产生显著的扩散阻力。图10显示了壳的厚度随着时间如何增加,直到整个聚gma刷层具有结合蛋白质。未结合的聚gma刷是这个模型中的“芯”,其随着蛋白质穿透“壳”并结合到刷上会缩小。最初,具有吸附的蛋白质的“壳”的厚度将为零,并且蛋白质对外表面的吸附速率将非常快。随着时间的推移,“壳”厚度和传质的扩散阻力增加,直到吸附的蛋白质壳到达pbt表面,“核”收缩到零。我们还假设,由于扩散过程相对于吸附速率较慢,所以扩散过程处于准稳定状态,因此通过扩散进入聚gma刷层的蛋白质传输速率随位置而恒定,从而在壳中任何点,蛋白质到带电聚合物的吸附速率等于通过壳的扩散速率。通过这些假设,在接枝层内的任何径向位置处,蛋白质的吸附速率等于蛋白质径向扩散到聚gma刷层中的速率,由方程式4表达。
在方程式中,ra为蛋白质在给定径向位置处吸附到壳的总速率,de为蛋白质在具有吸附的蛋白质的聚gma刷基质中的有效扩散系数,c蛋白质是扩散通过聚gma层的蛋白质浓度,l是给定纤维的长度,r是径向位置。由于聚gma刷的蛋白质结合速率几乎是瞬时的,因此“芯”和“壳”之间的界面处的蛋白质浓度(r=rc)可以取为零。另外,由于通过壳的扩散是限速步骤,壳外部的蛋白质浓度(r=r2)等于溶液中的本体蛋白质浓度,表示为c本体。将方程式4从接枝层的内径到外径积分得到蛋白质吸附速率的径向变化的表达式,
蛋白质扩散进入芯并与聚gma结合的速率等于可用于结合的聚gma“芯”材料消耗的质量速率,
未被占据的聚gma“芯”的质量消失速率可以用聚gma占据的体积变化的密度和时间速率表示(方程式7),
将方程式6和7替进方程式5给出了聚gma的“芯”与具有结合蛋白质的“壳”之间界面变化的时间速率的方程式,
在时间t=0时,芯半径是r2。方程式8关于时间的积分得到作为时间的函数的吸附到接枝层中的质量的表达式,
在方程式9中,m是在任何给定时间结合的蛋白质的质量,meq是平衡时结合的蛋白质的质量,mi是由于吸附在非织造物的外表面上而在时间t=0时结合的蛋白质的初始质量。收缩芯模型并不预测该初始蛋白质吸附(mi),但其可以从我们的实验结果中估计(见图9)。当使用该模型时,在溶液中的该初始浸入步骤之后,作为时间的函数的吸附到聚gma刷层上的蛋白质平衡量的分数由下式给出:ψ=[(m-mi)/(meq-mi)]。作为时间函数的结合到聚gma刷层上的蛋白质的总量取决于两个参数τ和φ,其对于具有不同纤维直径和接枝层厚度的每种不同非织造物而言是变化的,
在方程式10和11中,r1和r2分别是pbt纤维和被聚gma刷层覆盖的纤维的半径,如图10所示。另外,在方程式10中,de是蛋白质通过壳层的有效扩散系数,c本体是液相中蛋白质溶液的浓度,其对每个进行的实验为10mg/ml。
已知当用带电基团官能化时,由于电荷之间的静电排斥,聚gma刷在含水溶剂中溶胀。因此,由方程式3计算的干燥聚gma刷厚度不能用来确定r2,并且必须计算出溶胀的聚gma刷厚度。如果我们假设聚gma刷在刷层被蛋白质完全填充时完全延伸,我们可以估计聚gma刷的厚度r2。这是一个合理的假设,因为已知在给定足够时间的情况下,蛋白质在聚电解质刷基质中实现高填充效率。平衡时单位质量膜吸附的蛋白质质量为meq(质量蛋白质/质量膜),相当于材料的结合能力。蛋白质结合的质量可以使用该蛋白质的偏微分比容转化为蛋白质结合的体积。蛋白质占据的体积是pbt纤维周围的环形圆柱体积。使用这个几何形状,使用方程式12可以计算出r2,给出溶胀聚gma刷厚度。方程式12的推导可见于补充信息:溶胀聚gma刷半径的推导。
在方程式12中,meq是平衡结合能力,
除了增加观察到的刷厚度之外,聚gma刷的溶胀有效地增加了刷占据的体积,导致“芯”观察到的密度降低。未结合的溶胀聚gma刷组成“芯”,这些刷的密度就是芯的密度。我们从pbt非织造物的重量增量知道聚gma刷的质量,我们可以使用r2的计算值来计算溶胀聚gma刷的体积。聚gma刷的质量和体积可以用来计算由方程式13所表示的“芯”的有效密度,推导可见于补充信息:聚gma芯密度的推导。
在方程式13中,ρ芯是蛋白质吸附到的“芯”的有效密度,%重量增量是从接枝加入非织造物的聚gma的质量,ρpbt是pbt纤维的密度,r1是pbt纤维的半径,r2是溶胀聚gma半径。r2的计算值以及随后的溶胀聚gma刷厚度、有效芯密度和φ值如表3所示。
表3:各种接枝非织造物样品的接枝到pbt的溶胀聚gma层的半径、溶胀聚gma厚度、有效“芯”密度和φ值。
使用表3中给出的φ值,作为时间函数的吸附在膜中的总质量的实验数据可以用于拟合方程式9中仅剩的参数τ。从这些τ值,可以估算扩散通过吸附到聚gma接枝层的蛋白质的“壳”的蛋白质的有效扩散系数的值。
图11a和b显示了对于在动力学结合实验中测试的所有非织造物的ψ相对于蛋白质接触时间(t)的结果的最佳拟合线。在图13中所见的水平点划线是用于表3中获得的各种φ值的官能化聚gma“芯”与蛋白质的平衡吸附的ψ的最大值(ψmax)。图13中所见的最佳拟合线达到ψmax的时间是达到平衡蛋白质结合(teq)所需的时间。此外,图13中所见的最佳拟合线的斜率等于方程式5.1所定义的τ的倒数。一旦τ已知,通过“壳”的蛋白质传输的有效扩散系数(de)的值可以使用方程式5.2计算。表4给出了teq、τ和de的值。
表4:在收缩芯模型中通过“壳”的蛋白质扩散的吸附特征时间(τ)、实现聚gma“芯”完全转化所需的时间(teq)以及计算的有效扩散系数(de)。
表4显示了从阴离子交换和阳离子交换系统的τ的最佳拟合值获得的实验有效扩散系数。对于bsa通过蛋白质填充的阴离子交换官能化聚gma层的扩散,有效扩散系数在2.47x10-13和7.25x10-13cm2/s之间,平均有效扩散系数为4.77×10-13cm2/s。对于higg通过蛋白质填充的阳离子交换官能化聚gma层的扩散,有效扩散系数在1.28x10-13和11.6x10-13cm2/s之间,平均有效扩散系数为5.16x10-13cm2/s。根据斯托克斯-爱因斯坦(stokes-einstein)方程,bsa和igg在室温下在水中的扩散系数分别为6.23×10-7cm2/s和3.41×10-7cm2/s。色谱树脂中蛋白质的典型孔隙扩散系数约为10-9至10-12cm2/s量级。离子交换官能化聚gma接枝非织造物上的蛋白质捕获的有效扩散系数显著低于色谱树脂中的大多数孔隙扩散系数。清楚的是,通过离子交换官能化聚gma刷的蛋白质捕获在图10所示的聚gma/蛋白质“壳”中遭受严重的扩散限制,如表4所示,需要几分钟、否则数小时达到平衡。
收缩芯模型依赖于φ和τ来预测蛋白质在聚gma“芯”中如何随时间吸附。使用表3和4中分别找到的φ和τ的值,可以将收缩芯模型结果与蛋白质吸附到官能化聚gma“芯”的实验数据作为时间的函数进行比较。图12显示了吸附到阴离子交换聚gma“芯”的bsa质量相对于t/τ的使用φ值的结果。
如图12所示,收缩芯模型提供了实验数据的良好近似。在收缩芯模型中,随着聚gma接枝物的厚度围绕给定的pbt纤维直径增加,φ值减少,ψmax值增加,如图11中可见,且吸附特征时间尺度(τ)增加。由于这些原因,对于具有较厚接枝层的样品,需要更长的时间以达到平衡,如表4所示。当比较分别具有溶胀的聚gma刷厚度504nm和281nm的以20%和5.9%重量增量接枝的商用pbt时,可以特别看到这种现象。两个样品使用相同起始材料,具有不同程度的聚gma覆盖,导致φ、ψmax和τ的值显著不同。因此,根据收缩芯模型,具有较薄接枝层的商用pbt将表现出更快的转化速率并且更快达到平衡。以5.9%重量增量接枝的商用pbt在241分钟内达到平衡,相比之下,以20%重量增量接枝的商用pbt在876分钟内达到平衡,观察到该现象。
108i/spbt非织造物具有比商用pbt非织造物更小的pbt纤维直径,并且导致如图5所示的更薄的接枝聚gma刷层。然而,接枝到20%重量增量的商用pbt与108i/spbt非织造物的r1和r2值之间具有比例性,导致在收缩芯模型中出现类似的φ值和类似趋势。尽管接枝到20%重量增量的108i/spbt非织造物和商用pbt非织造物两者的φ值都是相同的,但它们的特征吸附时间尺度基本上不同,其中以20%重量增量接枝的108i/s非织造物的τ为1839分钟,以20%重量增量接枝的商用pbt非织造物则为6716分钟。以20%重量增量接枝的108i/spbt非织造物所观察到的较小特征时间尺度导致在223分钟后达到平衡,与此相比,接枝至20%重量增量的商用pbt非织造物需要876分钟达到平衡。接枝到5.9%重量增量的商用pbt和接枝到20%重量增量的108i/spbt都具有较短的聚gma刷厚度和根据收缩芯模型在220和250分钟之间达到平衡结合所需的显著较短的时间,相比之下,以20%重量增量接枝的商用pbt非织造物需要接近900分钟。这是强有力的证据,表明减少聚gma接枝层的厚度导致聚gma芯的转化速率更快,并且在该扩散限制吸附过程中更短的时间达到平衡。
通过相对于t/τ标绘的higg结合的阳离子交换聚gma“芯”的转化与收缩芯模型拟合到各种φ值的结果示于图13。
如图13所示,收缩芯模型是不同φ值的实验数据的合理近似值。与阴离子交换非织造物类似,对于给定的pbt纤维直径,对于更薄的聚gma层,观察到达到平衡更短的时间。接枝到5.3%重量增量的商用pbt能够在283分钟内达到平衡,相比之下,18%重量增量的聚gma接枝需要556分钟达到平衡。以18%重量增量接枝的108i/spbt非织造物与以18%重量增量接枝的商用pbt相比,也在较短时间内达到平衡结合。然而,如表4所证实,平衡结合时间的差异不如阴离子交换样品那么大。
图12和13中描绘的收缩芯模型证实了蛋白质结合的非常快的初始速率,这可以在蛋白质结合的非常短的时间尺度上看到。由于大量的蛋白质结合,产生了对蛋白质吸附施加扩散限制的“壳”,如图10所示。蛋白质结合的聚gma“壳”随时间变得更厚,并且在该致密层中的扩散距离变得更大,蛋白质扩散需要更长的时间才能到达可用芯以进行结合。这在图12和图13中观察到,随着材料接近平衡,在实验和收缩芯模型中都可见蛋白质吸附速率下降。
很明显,通过离子交换官能化的聚gma刷捕获蛋白质受到严重的扩散限制,需要几分钟、否则数小时才达到平衡。因此,使用更薄的聚gma刷层减少扩散距离并减少完全转化所需的时间是有利的。此外,通过使用高比表面积非织造物(如108i/spbt),聚gma结合体积可以分布在更薄的层中,如图5所示,减少了捕获时间,同时仍保持高蛋白质结合能力。
受益于前述说明中给出的教导,本发明所属领域的技术人员将会想到本发明的许多修改和其他实施方案。因此,应该理解的是,本发明不限于所公开的特定实施方案,并且修改和其他实施方案旨在被包括在所附权利要求的范围内。虽然在此使用了特定的术语,但是它们仅以一般的和描述性的意义使用,而不是为了限制的目的。
- 还没有人留言评论。精彩留言会获得点赞!