用于克隆茶树LOB基因的引物及其克隆方法和应用与流程
本发明涉及生物技术领域,尤其是茶树lob基因、用于克隆茶树lob基因的引物及其克隆方法和应用。
背景技术:
高等植物的顶端生长以及众多侧生器官的相继分化和发育,是高等植物最基本的生长发育问题,直接关系到植株各部分器官的组成、个体的大小和经济价值的高低,因而成为近年来植物功能基因组研究的一个重要热点。
植物的茎尖分生组织(sam,shootapicalmeristem)是位于生长点的一群细胞,它从胚胎的形态发生之初开始启动,一直维持分生状态并贯穿植物生长发育的始终。茎尖分生组织的维持需要在中央区茎细胞和周边区生成细胞之间保持一种平衡,这种平衡的打破往往导致侧生器官原基的启动和形成。
高等植物的侧枝、叶片、花序、花器官等均为侧生器官。植物的侧生器官边界区(lateralorganboundariesdomain,lbd)将叶片等侧生器官(分化细胞)与顶端分生组织(干细胞)分隔开,确保器官的形成和干细胞的维持。此外,器官边界区产成侧生分生组织,进而形成侧芽,影响植物株型的建成。lbd基因是最近在高等植物中发现的为植物所特有的一类新的基因,含有保守的lob(lateralorganboundaries)结构域,lob基因是植物所特有的一个基因。但由于边界区细胞数量较少,表型不易观察,因此对边界区形成的正反向遗传学研究都很困难,使得我们对边界区形成的调控机理知之甚少。
茶叶对现代疾病如高血压、糖尿病、辐射病、心脏病、心血管病、帕金森病、癌症和肥胖症等有一定的药理功效。茶多酚是茶树中主要的次生代谢产物,儿茶素约占茶多酚总量的70%。儿茶素生物合成主要涉及的酶类有pal、c4h、4cl、chs、chi、f3h、f3’h、f3’5’h、dfr、lar、ans和anr等,已有研究结果表明,pal、f3h、lar和dfr的表达量随叶片成熟度增加而逐渐下降:在新芽表达量最高,新叶次之,老叶最少。
研究发现拟南芥、甜橙、亚麻荠等植物的lob基因均对植物地上部、地下部的特定器官的形成与发育具有重要影响,但对茶树中相关lob基因的研究以及lob基因与儿茶素合成的关系的研究还没有报道,因此对茶树lob基因的克隆及其他分子研究尤为重要。
主要的技术文献有:bowmanj.l.,andeshedy.,2000,formationandmaintenanceoftheshootapicalmeristem,trendsplantsci.,5(3):110-115;aida,m.,ishida,t.,tasaka,m.,1999.shootapicalmeristemandcotyledonformationduringarabidopsisembryogenesis,interactionamongthecupshapedcotyledonandshootmeristemlessgenes.development126,1563–1570;clarks.e.,2001,meristems:startyoursignaling,curr.cpin.plantbiol.,4(1):28-32;iwakawah.,uenoy.,semiartie.,onouchih.,kojimas.,tsukayah.,hasebem.,somat.,ikezakim.,machidac.,andmachiday.,2002,theasymmetricleaves2geneofarabidopsisthaliana,requiredforformationofasymmet-ricflatleaflamina,encodesamemberofanovelfamilyofproteinscharacterizedbycysteinerepeatsandaleucinezipper,plantcellphysiol.,43(5):467-478;chalfun-juniora.,frankenj.,mesj.j.,marsch-martinezn.,pereiraa.,andangenentg.c.,2005,asymmetricleaves-like1gene,amemberoftheas2/lobfamily,controlsproximal-distalpatterninginarabidopsispetals,plantmol.biol.,57(4):559-575;liuh.j.,wangs.f.,yux.b.,yuj.,hex.w.,zhangs.l.,shouh.x.,andwup.,2005,arl1,alob-domainproteinrequiredforadventitiousrootformationinrice,plantj.,43(1):47-56;abe,m.,2003.regμulationofshootepidermalcelldifferentiationbyapairofhomeodomainproteinsinarabidopsis.development130,635-643.
技术实现要素:
本发明要解决的第一个技术问题是提供一种用于克隆茶树lob基因保守域的引物及其克隆方法,其核苷酸序列为seqidno.1,氨基酸序列为seqidno.2。为实现上述目的,本发明采用以下技术方案:
首先利用ncbi的entrez检索系统,查找到一条拟南芥lob基因的氨基酸序列,具有seqidno.3所述的氨基酸序列。随后利用这一序列使用blastp(通过蛋白查查找蛋白),在整个nr数据库中查找与之相似的其它物种的氨基酸序列。
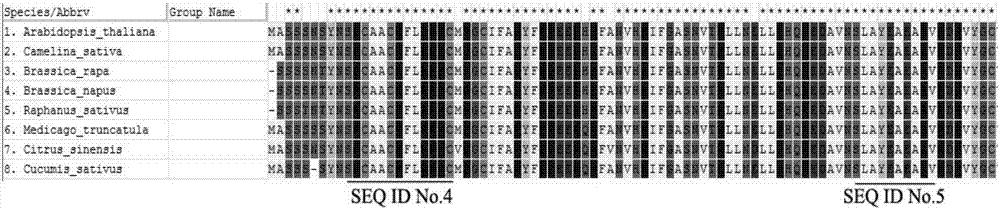
将搜索到的同一基因的不同氨基酸序列进行多序列比对,可选工具clustalw。通过序列比对,确定两个相距47个氨基酸的保守区域,氨基酸序列分别为seqidno.4和seqidno.5所述的氨基酸序列。得到保守区域后,利用primer5.0进行简并引物的设计。将参与多序列比对的序列中的任一条导入primer5.0中,再将其翻译成核苷酸序列,有密码子简并性,其结果是有n多条彼此只相差一个核苷酸的序列群,该群可用一条有简并性的核苷酸链来表示(其中r=a/g,y=c/t,m=a/c,k=g/t,s=c/g,w=a/t,h=a/c/t,b=c/g/t,v=a/c/g,d=a/g/t,n=a/c/g/t),在primer5.0中修改参数,令其在两个距离合适的保守的nt区域内寻找引物对,要保证上下游引物都落在该简并链的保守区域内。
基于上述步骤,本发明提供的克隆茶树lob基因保守域的引物组为:
lob-degenerate-f:5’-tgygcbgcstgcaartt-3’
lob-degenerate-r:5’-gakatdgcdccdacrcagcc-3’
克隆茶树lob基因保守域的方法包括如下步骤:提取龙井长叶rna,按照takaraprimescripttmii1ststrandcdnasynthesiskit的方法将上述rna进行逆转录得到cdna,以该cdna为模板,lob-degenerate-f为上游引物,lob-degenerate-r为下游引物进行pcr。获得茶树lob基因保守域的核苷酸序列,即seqidno.1。
本发明要解决的第二个技术问题是提供扩增茶树lob基因5’末端、3’末端全长的特异性引物组。根据已得到的茶树lob基因保守序列核苷酸序列,设计得到5’和3’末端扩增反应所需的引物组,其特征如下:
5’raceouterprimer:5’-catggctacatgctgacagccta-3’
5’raceinnerprimer:5’-cgcggatccacagcctactgatgatcagtcgatg-3’
cslob-5’race-gsp1:5’-aggagttgactgcgtcttcacgttggtg-3’
cslob-5’race-gsp2:5’-gagcttcgtgacgttgcttgctccaaag-3’
3’raceouterprimer:5’-taccgtcgttccactagtgattt-3’
3’raceouterprimer:5’-cgcggatcctccactagtgatttcactatagg-3’
cslob-3’race-gsp1:5’-tgcaagtttcttcgccggaaatgcatgc-3’
cslob-3’race-gsp2:5’-gagccacacaaattcgccaacgtccac-3’
本发明要解决的第三个技术问题是提供一种用于克隆茶树lob基因全长序列的引物组及该基因的克隆方法,其特征在于用于该基因的引物分别为cslob-f(seqidno.6)和cslob-r(seqidno.7),该基因的核苷酸序列为seqidno.8。为实现上述目的,本发明采用以下技术方案:
(1)茶树lob基因5’末端扩增反应
首先以takara5’-fullracekit提供的5’raceouterprimer和基因特异引物cslob-5’race-gsp1为引物,龙井长叶cdna为模板,进行5’末端outerpcr反应;然后再以5’raceinnerprime和基因特异引物cslob-5’race-gsp2为引物,以0.5μlouterpcr产物为模板,进行茶树lob基因5’末端innerpcr反应。
(2)茶树lob基因3’末端扩增反应
首先以takara3’-fullracekit提供的3’raceouterprimer和基因特异引物cslob-3’race-gsp1为引物,龙井长叶cdna为模板,进行3’末端outerpcr反应;然后再以3’raceinnerprimer和基因特异引物cslob-3’race-gsp2为引物,以0.5μlouterpcr产物为模板,进行茶树lob基因3’末端innerpcr反应。
(3)茶树lob基因全长cdna扩增
根据茶树lob基因5’和3’末端测序结果分别设计了正向引物cslob-f和反向引物cslob-r,以龙井长叶cdna为模板,扩增茶树lob基因全长序列。
本发明要解决的第四个技术问题是提供一种用于lob基因荧光定量rt-pcr的引物组,其特征在于用于定量pcr的引物分别为cslob-qrt-f(seqidno.9)和cslob-qrt-r(seqidno.10),内参基因引物分别为csgapdh-qrt-f(seqidno.11)和csgapdh-qrt-r(seqidno.12)。为实现上述目的,本发明采用以下技术方案:
提取龙井长叶不同组织(芽,第一叶,第二叶,老叶)的rna,按照takaraprimescripttmii1ststrandcdnasynthesiskit的方法将上述rna进行逆转录(20μl逆转录体系加入1μg的totalrna)得到cdna,以该cdna为模板,分别以cslob-qrt-f/cslob-qrt-r和csgapdh-qrt-f/csgapdh-qrt-r引物组,按照sybrpremixextaqii染料法荧光定量试剂盒的方法配制pcr反应液,分析不同组织中lob的表达量。
本发明的有益效果是:本发明根据已知其它物种lob基因的氨基酸及核苷酸序列设计得到扩增茶树lob基因保守序列的引物,再根据扩增得到的茶树lob基因保守序列,扩增茶树lob基因的5’和3’末端序列,最终首次获得茶树lob基因全长序列。通过克隆茶树lob基因的引物组和克隆方法,它能快速准确地克隆茶树lob基因的全序列。已有研究表明,不同发育阶段茶鲜叶中,新芽的儿茶素合成相关基因表达量最高,第一叶次之,成熟叶片中明显低于第一叶。荧光定量pcr检测结果表明,茶树芽中lob的表达量最少,老叶中表达量相对较高,这一结果与儿茶素积累量是相反的,可能是由于lob对儿茶素的合成起负调控作用。本发明为后续茶树lob基因的确切功能和参与的儿茶素代谢调控途径等知识的研究奠定基础。
附图说明
图1为不同植物(拟南芥、亚麻荠、芜菁、甜橙、甘蓝型油菜等)lob氨基酸序列比对图。
图2为cslob3’-race末端扩增的琼脂糖凝胶电泳图。
图3为cslob5’-race末端扩增的琼脂糖凝胶电泳图。
图4为扩增茶树lob基因全长cdna的琼脂糖凝胶电泳图。
图5为茶树不同组织(芽、第一叶、第二叶、老叶)中lob定量pcr图。
具体实施方式
下面结合具体实施方案对本发明做进一步的说明,这些实施例应被理解为仅为本发明的最佳实例,而非以任何方式限制本发明,凡在本发明原则之内所做的任何改进、修饰和等同替换,均应包含在本发明的范围之内。
实施例1:龙井长叶totalrna的提取
取龙井长叶叶片组织2g于液氮中研磨,按照柱式小量植物组织抽提试剂盒说明,提取totalrna,经琼脂糖凝胶电泳检测质量符合要求后,冻存备用。
实施例2:茶树lob基因保守域扩增反应
根据已知其它物种lob基因的氨基酸及核苷酸序列设计得到扩增茶树lob基因保守序列的引物lob-degenerate-f和lob-degenerate-r,以龙井长叶cdna为模板,扩增茶树lob基因保守域的序列。反应程序为94℃3min,94℃30s,58℃30s,72℃30s,35cycles,72℃10min。
根据已知其它物种lob基因的氨基酸及核苷酸序列设计得到扩增茶树lob基因保守序列的引物lob-degenerate-f和lob-degenerate-r具体是:首先利用ncbi的entrez检索系统,查找到一条拟南芥lob基因的氨基酸序列,具有seqidno.3所述的氨基酸序列。随后利用这一序列使用blastp(通过蛋白查查找蛋白),在整个nr数据库中查找与之相似的其它物种的氨基酸序列。
将搜索到的同一基因的不同氨基酸序列进行多序列比对,可选工具clustalw,其结果图如图1所示。通过序列比对,确定两个相距47个氨基酸的保守区域,氨基酸序列分别为seqidno.4和seqidno.5所述的氨基酸序列。得到保守区域后,利用primer5.0进行简并引物的设计。将参与多序列比对的序列中的任一条导入primer5.0中,再将其翻译成核苷酸序列,有密码子简并性,其结果是有n多条彼此只相差一个核苷酸的序列群,该群可用一条有简并性的核苷酸链来表示(其中r=a/g,y=c/t,m=a/c,k=g/t,s=c/g,w=a/t,h=a/c/t,b=c/g/t,v=a/c/g,d=a/g/t,n=a/c/g/t),在primer5.0中修改参数,令其在两个距离合适的保守的nt区域内寻找引物对,要保证上下游引物都落在该简并链的保守区域内。
实施例3:茶树lob基因5’末端扩增反应
a、totalrna去磷酸化处理
按照takara5’-fullracekit说明,使用alkalinephosphatase(ciap)对totalrna中裸露的5’磷酸基团进行去磷酸反应。去磷酸反应液体系为茶树totalrna(1μg/μl)1μl,rnaseinhibitor(40u/μl)1μl,10×alkalinephosphatasebuffer(mgcl2free)5μl,alkalinephosphatase(calfintestine)(16u/μl)0.6μl,rnasefreedh2o46.9μl。50℃反应1小时后。向上述反应液中加入20μl的3mch3coona(ph5.2),130μl的rnasefreedh2o,充分混匀。再加入200μl的苯酚/氯仿/异戊醇(25:24:1),充分混匀后13,000×g室温离心5min,将上层水相转移至新的microtube中。再加入200μl的氯仿,充分混匀后13,000×g室温离心5min,将上层水相转移至新的microtube中。加入2μl的nacarrier均匀混合后,加入200μl的异丙醇,冰上冷却10min,13,000×g4℃离心20min,弃上清,加入500μl70%的乙醇漂洗,13,000×g4℃离心5min,弃上清后干燥,最后加入7μl的rnasefreedh2o溶解沉淀,得到ciap-treatedrna。
b、mrna“去帽子”反应。
使用tobaccoacidpyrophosphatase(tap)去掉mrna的5’帽子结构,保留一个磷酸基团。反应体系为ciap-treatedrna7μl,rnaseinhibitor(40u/μl)1μl,10×tapreactionbuffer1μl,tobaccoacidpyrophosphatase(0.5u/μl)1μl,37℃反应1小时,得到ciap/tap-treatedrna。
c、5’raceadaptor的连接
首先配置下列溶液:ciap/tap-treatedrna5μl,5’raceadaptor(15μm)1μl,rnasefreedh2o4μl。上述溶液65℃反应1小时后,加入20μl的3mch3coona(ph5.2),140μl的rnasefreedh2o,充分混匀。再加入200μl的苯酚/氯仿/异戊醇(25:24:1),充分混匀后13,000×g室温离心5min,将上层水相转移至新的microtube中。再加入200μl的氯仿,充分混匀后13,000×g室温离心5min,将上层水相转移至新的microtube中。加入2μl的nacarrier均匀混合后,加入200μl的异丙醇,冰上冷却10min,13,000×g4℃离心20min,弃上清,加入500μl70%的乙醇漂洗,13,000×g4℃离心5min,弃上清后干燥,最后加入6μl的rnasefreedh2o溶解沉淀,得到ligatedrna。
d、反转录反应。
反应体系为ligatedrna6μl,random9mers(50μm)0.5μl,dntp(10mmeach)1μl,rnaseinhibitor(40u/μl)0.25μl,reversetranscriptasem-mlv(rnaseh-)(200u/μl)0.25μl。反应条件为30℃10min,42℃1hr,72℃15min,即可得到用于扩增5’末端的龙井长叶cdna。
e、outerpcr反应。
以takara5’-fullracekit提供的5’raceouterprimer和基因特异引物cslob-5’race-gsp1为引物,龙井长叶cdna为模板,进行5’末端outerpcr反应;扩增体系为龙井长叶cdna2μl,1×cdnadilutionbufferii8μl,10×lapcrbuffer(mg2+free)4μl,mgcl2(25mm)3μl,takaralataq(5u/μl)0.25μl,cslob-5’race-gsp1(10μm)2μl,5’raceouterprimer(10μm)2μl,dh2o28.75μl。pcr反应条件为94℃3min,94℃30s,55℃30s,72℃1min,20cycles,72℃10min。
f、innerpcr反应。
以5’raceinnerprimer和基因特异引物cslob-5’race-gsp2为引物,以1μlouterpcr产物为模板,进行茶树lob基因5’末端innerpcr反应。反应体系和扩增程序同outerpcr反应。pcr产物经电泳检测,结果如图2所示。
实施例4:茶树lob基因3’末端扩增反应
a、反转录反应。
反应体系为totalrna(500ng/μl)1μl,3’raceadaptor(5μm)1μl,5×primescriptbuffer2μl,dntpmixture(10mmeach)1μl,rnaseinhibitor(40u/μl)0.25μl,primescriptrtase(200u/μl0.25μl)0.25μl,rnasefreedh2o4.5μl。反应条件42℃60min,70℃15min,反应结束即得到用于扩增3’末端的龙井长叶cdna。
b、outerpcr反应。
以takara3’-fullracekit提供的3’raceouterprimer和基因特异引物cslob-3’race-gsp1为引物,龙井长叶cdna为模板,进行3’末端outerpcr反应;扩增体系为龙井长叶cdna2μl,1×cdnadilutionbufferii8μl,10×lapcrbuffer(mg2+free)4μl,mgcl2(25mm)3μl,takaralataq(5u/μl)0.25μl,cslob-3’race-gsp1(10μm)2μl,3’raceouterprimer(10μm)2μl,dh2o28.75μl。pcr反应条件为94℃3min,94℃30s,55℃30s,72℃1min,20cycles,72℃10min。
c、innerpcr反应。
以3’raceinnerprimer和基因特异引物cslob-3’race-gsp2为引物,以1μlouterpcr产物为模板,进行茶树lob基因3’末端innerpcr反应。反应体系和扩增程序同outerpcr反应。pcr产物经电泳检测,结果如图3所示。
实施例5:茶树lob基因全长cdna序列扩增。
根据茶树lob基因5’和3’末端测序结果分别设计了正向引物cslob-f和反向引物cslob-r,以龙井长叶cdna为模板,扩增茶树lob基因全长序列。反应程序为94℃3min,94℃30s,58℃30s,72℃30s,35cycles,72℃10min。pcr产物电泳结果如图4所示。
实施例6:pcr产物的克隆纯化与筛选。
pcr产物经1.5%琼脂糖凝胶电泳后,用胶回收试剂盒对pcr产物进行回收,经纯化后与takara公司t载体pmd18-tvector进行连接,用热激法导入大肠杆菌dh5α感受态中并在氨苄青霉素平板上筛选阳性克隆。分子检测确定后的阳性克隆子送杭州擎科生物公司完成测序。
实施例7:荧光定量pcr检测茶树不同组织中lob的表达量
提取龙井长叶不同组织(芽,第一叶,第二叶,老叶)的rna,按照takaraprimescripttmii1ststrandcdnasynthesiskit的方法将上述rna进行逆转录(20μl逆转录体系加入1μg的totalrna)得到cdna,以该cdna为模板,利用本发明所述的引物cslob-qrt-f、cslob-qrt-r、csgapdh-qrt-f和csgapdh-qrt-r为引物,按照下列组分配制pcr反应液:
pcr扩增程序如下:94℃3min,94℃20s,58℃20s,72℃20s,40cycles,72℃收集荧光,结果如图5所示。
seqidno.1:
tgcgcggcgtgcaagtttcttcgccggaaatgcatgccgggatgcatattcgcgccatattttccaccggaggagccacacaaattcgccaacgtccacaaaatctttggagcaagcaacgtcacgaagctcctcaacgagctcctccctcaccaacgtgaagacgcagtcaactccttagcctatgaggccgaggcacgtgtccgtgaccccgtctatggctgcgtaggcgccatctc
seqidno.2:
caackflrrkcmpgcifapyfppeephkfanvhkifgasnvtkllnellphqredavnslayeaearvrdpvygcvgai
seqidno.3:
masssnsynspcaackflrrkcmpgcifapyfppeephkfanvhkifgasnvtkllnellphqredavnslayeaearvrdpvygcvgaisylqrqvhrlqkeldaanadlahyglstsaagapgnvvdlvfqpqplpsqqlpplnpvyrlsgaspvmnqmprgtggsygtflpwnnghdqqggnm
seqidno.4:spcaackflrrkc
seqidno.5:slayeaearv
seqidno.6:atggcgtcgtcatcaaactcatacaac
seqidno.7:tcaaatatttcctccagaagggctattg
seqidno.8:
atggcgtcgtcatcaaactcatacaactcaacatgcgcggcgtgcaagtttcttcgccggaaatgcatgccgggatgcatattcgcgccatattttccaccggaggagccacacaaattcgccaacgtccacaaaatcttcggcgcaagcaacataagtaagctcctcaatgaaatcctccctcaccaaagagatgatgcagtgagttccctcgcctacgaagccgaagcacgagtgcgagatccagtgtacggttgtgttggtgctatctccttcctccaaagacaggttgagaagctccagaaagatctagaagtcgccaatgccaatttgattcactga
seqidno.9:5’-gaaatcctccctcaccaaagag-3’
seqidno.10:5’-aggagatagcaccaacacaac-3’
seqidno.11:5’-ttggcatcgttgagggtct-3’
seqidno.12:5’-cagtgggaacacggaaagc-3’
sequencelisting
<110>浙江省农业科学院
<120>用于克隆茶树lob基因的引物及其克隆方法
<130>1
<160>12
<170>patentinversion3.3
<210>1
<211>239
<212>dna
<213>人工合成
<400>1
tgcgcggcgtgcaagtttcttcgccggaaatgcatgccgggatgcatattcgcgccatat60
tttccaccggaggagccacacaaattcgccaacgtccacaaaatctttggagcaagcaac120
gtcacgaagctcctcaacgagctcctccctcaccaacgtgaagacgcagtcaactcctta180
gcctatgaggccgaggcacgtgtccgtgaccccgtctatggctgcgtaggcgccatctc239
<210>2
<211>79
<212>prt
<213>人工合成
<400>2
cysalaalacyslyspheleuargarglyscysmetproglycysile
151015
phealaprotyrpheproproglugluprohislysphealaasnval
202530
hislysilepheglyalaserasnvalthrlysleuleuasngluleu
354045
leuprohisglnarggluaspalavalasnserleualatyrgluala
505560
glualaargvalargaspprovaltyrglycysvalglyalaile
657075
<210>3
<211>186
<212>prt
<213>人工合成
<400>3
metalaserserserasnsertyrasnserprocysalaalacyslys
151015
pheleuargarglyscysmetproglycysilephealaprotyrphe
202530
proproglugluprohislysphealaasnvalhislysilephegly
354045
alaserasnvalthrlysleuleuasngluleuleuprohisglnarg
505560
gluaspalavalasnserleualatyrglualaglualaargvalarg
65707580
aspprovaltyrglycysvalglyalailesertyrleuglnarggln
859095
valhisargleuglnlysgluleuaspalaalaasnalaaspleuala
100105110
histyrglyleuserthrseralaalaglyalaproglyasnvalval
115120125
aspleuvalpheglnproglnproleuproserglnglnleupropro
130135140
leuasnprovaltyrargleuserglyalaserprovalmetasngln
145150155160
metproargglythrglyglysertyrglythrpheleuprotrpasn
165170175
asnglyhisaspglnglnglyglyasnmet
180185
<210>4
<211>13
<212>prt
<213>人工合成
<400>4
serprocysalaalacyslyspheleuargarglyscys
1510
<210>5
<211>10
<212>prt
<213>人工合成
<400>5
serleualatyrglualaglualaargval
1510
<210>6
<211>27
<212>dna
<213>人工合成
<400>6
atggcgtcgtcatcaaactcatacaac27
<210>7
<211>28
<212>dna
<213>人工合成
<400>7
tcaaatatttcctccagaagggctattg28
<210>8
<211>342
<212>dna
<213>人工合成
<400>8
atggcgtcgtcatcaaactcatacaactcaacatgcgcggcgtgcaagtttcttcgccgg60
aaatgcatgccgggatgcatattcgcgccatattttccaccggaggagccacacaaattc120
gccaacgtccacaaaatcttcggcgcaagcaacataagtaagctcctcaatgaaatcctc180
cctcaccaaagagatgatgcagtgagttccctcgcctacgaagccgaagcacgagtgcga240
gatccagtgtacggttgtgttggtgctatctccttcctccaaagacaggttgagaagctc300
cagaaagatctagaagtcgccaatgccaatttgattcactga342
<210>9
<211>22
<212>dna
<213>人工合成
<400>9
gaaatcctccctcaccaaagag22
<210>10
<211>21
<212>dna
<213>人工合成
<400>10
aggagatagcaccaacacaac21
<210>11
<211>19
<212>dna
<213>人工合成
<400>11
ttggcatcgttgagggtct19
<210>12
<211>19
<212>dna
<213>人工合成
<400>12
cagtgggaacacggaaagc19
- 还没有人留言评论。精彩留言会获得点赞!
- 一组小熊猫基因的微卫星引物的制作方法与工艺
- 羊肚菌交配型决定基因鉴定专用引物及出菇能力预测方法与流程
- 一种KRAS基因的检测引物组、其构成的反应体系及应用的制作方法与工艺
- MTHFR‑A1298C基因多态性快速检测引物、分子信标、试剂盒及检测方法与流程
- 用于检测人类VKORC1与CYP2C9基因多态性的引物、探针、试剂盒及方法与流程
- 一种牛卷毛基因快速筛选的引物、方法及试剂盒与流程
- KCNQ1OT1基因、aHIF基因及其引物的应用以及用于诊断冠心病的试剂盒的制作方法与工艺
- 用于检测人类ALDH2基因多态性的引物、探针、试剂盒及方法与流程
- 一种检测耳聋基因的引物及其应用的制造方法与工艺
- 海岛棉跨膜蛋白基因、引物及其应用的制造方法与工艺
- 玉米低镁条件下特异性转运镁离子基因ZmMGT10及其应用的制造方法与工艺
- CRISPR‑Cas9特异性敲除猪SLA‑3基因的方法及用于特异性靶向SLA‑3基因的sgRNA与制造工艺
- 毛酸浆的分子特异性标记引物及鉴别方法与制造工艺
- 一种检测CYP2C9基因多态性的ARMS引物的制造方法与工艺
- 构建在海马体区域特异性敲除IKKα基因的小鼠模型的方法及打靶载体和试剂盒与制造工艺
- 使用重叠非等位基因特异性引物和等位基因特异性阻断剂寡核苷酸的组合物进行等位基因特异性扩增的制造方法与工艺
- 一种利用线粒体D-loop区特异区域鉴定鼋的标记引物及方法
- 一种用于检测大豆拟茎点种腐病菌的特异性lamp引物组合物及其应用
- 一种针对膨胀污泥中Eikelboom Type 0092型丝状菌群特异性引物和方法
- 脯氨酸特异性蛋白酶基因及其应用