卷曲蛋白的选择性肽抑制剂的制作方法
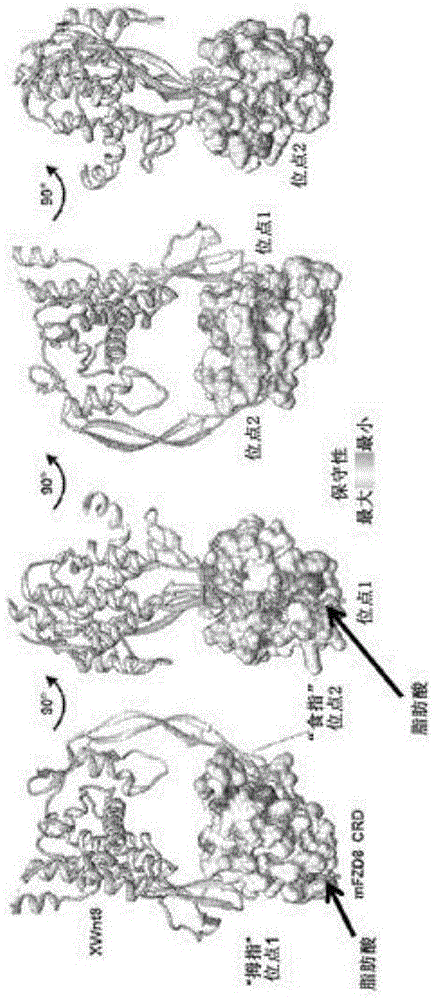
本申请要求2016年9月9日提交的流水号62/385,848的美国临时申请和2016年11月8日提交的流水号62/419,331的美国临时申请的优先权,通过援引将其内容完整收入本文。ascii文本文件上的序列表的提交通过援引将ascii文本文件上的下述提交的内容完整收入本文:计算机可读形式(crf)的序列表(文件名:146392036940seqlist.txt,记录日期:2017年9月6日,大小:50kb)。发明背景wnt信号传导途径与癌发生的关联作为某些鼠乳房肿瘤中的早期观察和实验的结果开始。wnt-1原癌基因(int-1)最初是自由于病毒dna序列的插入由小鼠乳房肿瘤病毒(mmtv)诱发的乳房肿瘤鉴定的(nusseetal.,cell1982;31:99-109)。此类病毒整合的结果是int-1表达失调,导致肿瘤形成(vanooyen,a.etal.,cell1984;39:233-240;nusse,r.etal.,nature1984;307:131-136;tsukamotoetal.,cell1988;55:619-625)。后续序列分析证明int-1是牵涉发育的果蝇基因wingless(wg)的一种哺乳动物同系物,然后组合这些术语以创建“wnt”来鉴定这个蛋白质家族。分泌型配体的人wnt基因家族现在已经生长至至少19个成员(例如wnt-1(refseq.:nm.sub.--005430),wnt-2(refseq.:nm.sub.--003391),wnt-2b(wnt-13)(refseq.:nm.sub.--004185),wnt-3(refseq.:nm.sub.--030753),wnt3a(refseq.:nm.sub.--033131),wnt-4(refseq.:nm.sub.--030761),wnt-5a(refseq.:nm.sub.--003392),wnt-5b(refseq.:nm.sub.--032642),wnt-6(refseq.:nm.sub.--006522),wnt-7a(refseq.:nm.sub.--004625),wnt-7b(refseq.:nm.sub.--058238),wnt-8a(refseq.:nm.sub.--058244),wnt-8b(refseq.:nm.sub.--003393),wnt-9a(wnt-14)(refseq.:nm.sub.--003395),wnt-9b(wnt-15)(refseq.:nm.sub.--003396),wnt-10a(refseq.:nm.sub.--025216),wnt-10b(refseq.:nm.sub.--003394),wnt-11(refseq.:nm.sub.--004626),wnt-16(refseq.:nm.sub.--016087))。每个成员具有不同程度的序列同一性但均含有显示高度保守间隔的23-24个保守半胱氨酸残基(mcmahon,a.p.etal.,trendsgenet.1992;8:236-242;miller,j.r.,genomebiol.2002;3(1):3001.1-3001.15)。wnt蛋白质是在胚胎发生和成熟组织二者中发挥关键作用的小型(即39-46kd)酰化分泌型糖蛋白。在胚胎发育期间,wnt蛋白质的表达在经由控制细胞增殖和决定干细胞命运的样式形成中是重要的。wnt分子还是棕榈酰化的,而且因而比在其它情况中通过单独分析氨基酸序列会预测的更加疏水(willert,k.etal.,nature2003;423:448-52)。还认为一个或多个棕榈酰化位点对于功能是至关紧要的。wnt蛋白质作为配体起作用以活化卷曲蛋白(frizzled,frz)家族七次跨膜受体(ingham,p.w.,trendsgenet.1996;12:382-384;yang-snyder,j.etal.,curr.biol.1996;6:1302-1306;bhanot,p.etal.,nature1996;382:225-230)。frz家族有十个已知成员(例如frz1-frz10),每一个特征在于存在富半胱氨酸域(crd)(huangetal.,genomebiol.2004;5:234.1-234.8)。各种wnt-卷曲蛋白相互作用之间有很大程度的混乱性,但是wnt-frz结合还必须掺入ldl受体相关蛋白(lrp5或lrp6)和膜和胞质蛋白散乱蛋白(dishevelled,dsh)以形成活性信号传导复合物。wnt对卷曲蛋白的结合能经由规范wnt信号传导途径(由此导致β-联蛋白的转录活性升高和稳定化)(peifer,m.etal.,development1994;120:369-380;papkoff,j.etal.,mol.cellbiol.1996;16:2128-2134)或非规范信号传导(诸如经由wnt/平面细胞极性(wnt/pcp)或wnt-钙(wnt/ca2+)途径)(veeman,m.t.etal.,dev.cell2003;5:367-377)激活信号传导。frz受体之一,fzd7在多种多样的人癌症中上调且能够调节wnt信号传导活性,甚至在具有下游信号转导物的突变的癌细胞中。因而,需要为多种治疗应用,诸如癌症开发fzd7的选择性抑制剂。本发明满足这种和其它需求。发明概述在某些实施方案中,提供的是一种包含结合卷曲蛋白7(fzd7)受体之富半胱氨酸域(crd)的非天然发生肽的配体。在依照(或应用于)任何上述实施方案的某些实施方案中,该非天然发生肽特异性结合fzd7之crd。在依照(或应用于)任何上述实施方案的某些实施方案中,该非天然发生肽不结合选自由fzd3,fzd4,fzd5,fzd6,fzd8,fzd9,或fzd10组成的组的fzd受体之crd。在依照(或应用于)任何上述实施方案的某些实施方案中,该非天然发生肽进一步结合fzd1和fzd2。在依照(或应用于)任何上述实施方案的某些实施方案中,该非天然发生肽不结合选自由fzd1,fzd2,fzd3,fzd4,fzd5,fzd6,fzd8,fzd9,或fzd10组成的组的fzd受体之crd。在依照(或应用于)任何上述实施方案的某些实施方案中,该非天然发生肽是线性的。在依照(或应用于)任何上述实施方案的某些实施方案中,该非天然发生肽是环状的。在依照(或应用于)任何上述实施方案的某些实施方案中,该非天然发生肽在长度上介于8-16个氨基酸之间。在依照(或应用于)任何上述实施方案的某些实施方案中,该非天然发生肽在长度上介于11-14个氨基酸之间。在依照(或应用于)任何上述实施方案的某些实施方案中,该fzd7是hfzd7。在依照(或应用于)任何上述实施方案的某些实施方案中,该非天然发生肽特异性结合hfzd7crd的包含至少三个选自由leu81,his84,gln85,tyr87,pro88,phe138,和phe140组成的组的氨基酸的结合区。在依照(或应用于)任何上述实施方案的某些实施方案中,该非天然发生肽包含x1x2x3ddlx4x5wchvmy(seqidno:100)中所列氨基酸序列,其中x1-x3中每个是无氨基酸(noaminoacid),任何氨基酸,或非天然氨基酸,且其中x4-x5中每个是任何氨基酸或非天然氨基酸。在依照(或应用于)任何上述实施方案的某些实施方案中,x1是l,x2是p,x3是s,x4是e,且x5是f。在依照(或应用于)任何上述实施方案的某些实施方案中,该非天然发生肽包含lpsddlefwchvmy(seqidno:13)中所列氨基酸序列。在依照(或应用于)任何上述实施方案的某些实施方案中,x1是无氨基酸,x2是无氨基酸,x3是s,x4是e,且x5是f。在依照(或应用于)任何上述实施方案的某些实施方案中,该非天然发生肽包含sddlefwchvmy(seqidno:99)中所列氨基酸序列。在依照(或应用于)任何上述实施方案的某些实施方案中,该非天然发生肽的n端胺是乙酰化的,该非天然发生肽的c端羧基基团是酰胺化的,或该肽的n端胺是乙酰化的且该肽的c端羧基基团是酰胺化的。在依照(或应用于)任何上述实施方案的某些实施方案中,该非天然发生肽增强wnt对fzd7受体之crd的结合。在依照(或应用于)任何上述实施方案的某些实施方案中,该fzd7受体是hfzd7受体。在依照(或应用于)任何上述实施方案的某些实施方案中,该非天然发生肽包含sddlefwchvxy(seqidno:114)中所列氨基酸序列,其中x是任何氨基酸,或非天然氨基酸。在依照(或应用于)任何上述实施方案的某些实施方案中,该非天然氨基酸选自由2-氨基-3-癸氧基-丙酸,赖氨酸的包含在厄普西隆氨基基团处偶联的辛酸的衍生物,氨基癸酸,2-氨基癸酸,赖氨酸的包含在厄普西隆氨基基团处偶联的癸酸的衍生物,和6-羟基-l-正亮氨酸组成的组。在依照(或应用于)任何上述实施方案的某些实施方案中,该非天然发生肽包含sddxefwchvmy(seqidno:115)中所列氨基酸序列,其中x是任何氨基酸,或非天然氨基酸,且其中该非天然氨基酸选自由l-高亮氨酸,l-高苯丙氨酸,和赖氨酸的包含在厄普西隆氨基基团处偶联的辛酸的衍生物组成的组。在依照(或应用于)任何上述实施方案的某些实施方案中,该非天然发生肽包含seqidno:1-31和39-99任一中所列氨基酸序列。在依照(或应用于)任何上述实施方案的某些实施方案中,该非天然发生肽包含seqidno:32-38任一中所列氨基酸序列。在依照(或应用于)任何上述实施方案的某些实施方案中,该非天然发生肽以120nm或更小的ic50抑制wnt信号传导。在依照(或应用于)任何上述实施方案的某些实施方案中,该非天然发生肽具有90nm或更小的ec50值。在依照(或应用于)任何上述实施方案的某些实施方案中,该非天然发生肽是与脂质缀合的。在依照(或应用于)任何上述实施方案的某些实施方案中,该脂质是长链脂肪酸(lcfa)。在依照(或应用于)任何上述实施方案的某些实施方案中,该脂质是短链脂肪酸(scfa)。在依照(或应用于)任何上述实施方案的某些实施方案中,该脂肪酸包含芳香族尾。在依照(或应用于)任何上述实施方案的某些实施方案中,该配体中的非天然发生肽是二聚化的。在依照(或应用于)任何上述实施方案的某些实施方案中,该非天然发生肽是通过二硫键而二聚化的。在依照(或应用于)任何上述实施方案的某些实施方案中,该非天然发生肽是通过化学接头而二聚化的。在某些实施方案中,提供的是一种包含依照(或应用于)任何上述实施方案的配体和制药学可接受载剂的组合物。在某些实施方案中,提供的是一种抑制细胞中的wnt信号传导的方法,其包含使该细胞与依照(或应用于)任何上述实施方案的配体或组合物接触。在某些实施方案中,提供的是一种抑制干细胞增殖的方法,其包含使干细胞与依照(或应用于)任何上述实施方案的配体或组合物接触。在依照(或应用于)任何上述实施方案的某些实施方案中,该干细胞是肠干细胞。在依照(或应用于)任何上述实施方案的某些实施方案中,该干细胞是癌干细胞。在依照(或应用于)任何上述实施方案的某些实施方案中,该癌干细胞是结肠癌干细胞,胰腺癌干细胞,非小细胞肺癌干细胞,包含rnf43中的突变的癌干细胞,特征在于usp6过表达的癌干细胞,或特征在于牵涉r-spondin(rspo)家族成员的基因融合的癌干细胞在某些实施方案中,提供的是一种杀伤癌细胞的方法,其包含使该癌细胞与依照(或应用于)任何上述实施方案的配体或组合物接触。在依照(或应用于)任何上述实施方案的某些实施方案中,该癌细胞是结肠癌细胞,胰腺癌细胞,非小细胞肺癌细胞,包含rnf43中的突变的癌细胞,特征在于usp6过表达的癌细胞,或特征在于牵涉r-spondin(rspo)家族成员的基因融合的癌细胞。在某些实施方案中,提供的是一种治疗受试者中的癌症的方法,其包含施用有效量的依照(或应用于)任何上述实施方案的配体或组合物。在依照(或应用于)任何上述实施方案的某些实施方案中,该癌症是结肠癌,胰腺癌,非小细胞肺癌(nsclc),特征在于rnf43中的突变的癌症,特征在于usp6过表达的癌症,或特征在于牵涉r-spondin(rspo)家族成员的基因融合的癌症。在某些实施方案中,提供的是依照(或应用于)任何上述实施方案的配体或组合物在制造用于治疗癌症的药物中的用途。在依照(或应用于)任何上述实施方案的某些实施方案中,该癌症是结肠癌,胰腺癌,非小细胞肺癌(nsclc),特征在于rnf43中的突变的癌症,特征在于usp6过表达的癌症,或特征在于牵涉r-spondin(rspo)家族成员的基因融合的癌症。在某些实施方案中,提供的是一种包含依照(或应用于)任何上述实施方案的配体或组合物的组合物,其供治疗受试者中的癌症中使用。在依照(或应用于)任何上述实施方案的某些实施方案中,该癌症是结肠癌,胰腺癌,非小细胞肺癌(nsclc),或特征在于rnf43中的突变的癌症,特征在于usp6过表达的癌症,或特征在于牵涉r-spondin(rspo)家族成员的基因融合的癌症。在某些实施方案中,提供的是一种用于治疗癌症的试剂盒,其包含:(a)依照(或应用于)任何上述实施方案的配体或组合物,和(b)关于将该配体施用于具有癌症的受试者的说明书。在依照(或应用于)任何上述实施方案的某些实施方案中,该癌症是结肠癌,胰腺癌,非小细胞肺癌(nsclc),特征在于rnf43中的突变的癌症,特征在于usp6过表达的癌症,或特征在于牵涉r-spondin(rspo)家族成员的基因融合的癌症。在某些实施方案中,提供的是一种包含lpsddlefwshvmy(seqidno:113)中所列非天然发生肽的配体。要理解的是,可以组合本文中描述的各种实施方案的一项,一些,或所有特性以形成本发明的其它实施方案。本发明的这些和其它方面对本领域技术人员会变得显而易见。下面的详细描述进一步描述本发明的这些和其它实施方案。附图简述图1提供wnt8和小鼠fzd8crd之间的相互作用的结构描绘,将人fzdcrd之间的序列保守性展示到mfzd8crd的表面上。图2a显示为了确定肽fz7-21和fz-21s对用50ng/ml重组wnt3a刺激的hek293-tb细胞中的β-联蛋白信号传导的影响而实施的实验的结果。图2b显示为了确定肽fz7-21和fz-21s对用5ngpcdna3.2-wnt3a或25ngpcdna3.2-wnt3a转染的hek293-tb细胞中wnt刺激的β-联蛋白信号传导的影响而实施的实验的结果。图2c显示为了确定肽fz7-21和fz-21s对用5ngpcdna3.2-wnt1或25ngpcdna3.2-wnt1转染的hek293-tb细胞中wnt刺激的β-联蛋白信号传导的影响而实施的实验的结果。图2d显示为了确定fz7-21衍生的含有位置10处的d-cys立体异构体的肽对用5ngpcdna3.2-wnt1或25ngpcdna3.2-wnt3a转染的hek293-tb细胞中wnt刺激的β-联蛋白信号传导的影响而实施的实验的结果。图2e显示为了确定肽fz7-21和fz-21s对用6-bio处理的hek293-tb细胞中的受体不依赖性β-联蛋白信号传导的影响而实施的实验的结果。图3a提供为了测定5fam-fz7-21或5fam-fz7-21s结合hfzd1crd-fc的ec50值而实施的实验的结果。图3b提供为了测定5fam-fz7-21或5fam-fz7-21s结合mfzd2crd-fc的ec50值而实施的实验的结果。图3c提供为了测定5fam-fz7-21或5fam-fz7-21s结合hfzd4crd-fc的ec50值而实施的实验的结果。图3d提供为了测定5fam-fz7-21或5fam-fz7-21s结合hfzd5crd-fc的ec50值而实施的实验的结果。图3e提供为了测定5fam-fz7-21或5fam-fz7-21s结合hfzd7crd-fc的ec50值而实施的实验的结果。图3f提供为了测定5fam-fz7-21或5fam-fz7-21s结合mfzd7crd-fc的ec50值而实施的实验的结果。图3g提供为了测定5fam-fz7-21或5fam-fz7-21s结合hfzd8crd-fc的ec50值而实施的实验的结果。图3h提供为了测定5fam-fz7-21或5fam-fz7-21s结合mfzd9crd-fc的ec50值而实施的实验的结果。图3i提供为了测定5fam-fz7-21或5fam-fz7-21s结合hfzd10crd-fc的ec50值而实施的实验的结果。图4a提供在通过fsec的分析前与各种fzdcrd-fc蛋白质一起温育的5fam-fz7-21的代表性荧光痕迹。图4b提供在通过fsec的分析前与各种fzdcrd-fc蛋白质一起温育的5fam-fz7-21s的代表性荧光痕迹。图4c提供图4a和4b的荧光强度的量化(曲线下面积,auc)。图5a提供在通过fsec的分析前与各种人sfrp蛋白质一起温育的5fam-fz7-21的代表性荧光痕迹。图5b提供在通过fsec的分析前与各种sfrp蛋白质一起温育的5fam-fz7-21s的代表性荧光痕迹。图5c提供图5a和5b的荧光强度的量化(曲线下面积,auc)。图6a提供纯化的重组hfzdcrd的大小排阻层析概况。图6b显示来自图6a的合并的hfzd7crd的sds-page。图6c提供为了评估fz7-21是否诱导hfzd7crd多聚化而实施的实验的结果。图6d提供图6c的放大视图(1.5ml至2.0ml范围)。图7提供显示具有5fam-fz7-21或5fam-fz7-21s结合活性的人fzd富半胱氨酸域(crd)之间的进化保守性的分支图。图8a显示脱辅基hfzd7crd二聚体的晶体结构的飘带展示。图8b显示桥接脱辅基hfzd7crd二聚体界面的脂质结合腔的表面展示。图8c显示与fz7-21结合的hfzd7crd的晶体结构。图8d显示定位到与fz7-21结合的hfzd7crd的结构上的疏水腔的表面展示。图8e显示与fz7-21结合的hfzd7crd的晶体结构的顶视图表面展示。图8f显示与fz7-21结合的hfzd7crd的晶体结构的侧视图表面展示。图9a显示hfzd4的脂质结合沟的表面展示。图9b显示mfzd8的脂质结合沟的表面展示。图9c显示fzdcrd的叠加。图10a提供经由接头(seqidno:136)与fz7-21融合的hfzd7crd的示意图。图10b显示纯化的hfzd7crd-fz7-21融合构建物的大小排阻层析概况。图11a提供fz7-21二聚体中的分子内相互作用的图。图11b提供fz7-21二聚体中的分子内相互作用的图,其中各个相互作用以线描绘。图11c提供fz7-21二聚体结构上的作为梯度呈现的溶剂可及表面的描绘。图12a提供脱辅基hfzd7crd的结构的飘带展示,突显二聚体界面处的16°角。图12b提供与fz7-21结合的hfzd7crd的结构的飘带展示,突显二聚体界面处的90°角。图13突显与fz7-21结合的hfzd7crd的晶体结构内的选定fz7-21/hfzd7crd相互作用。图14提供hfzd7(seqidno:137),hfzd2(seqidno:138),hfzd1(seqidno:139),hfzd5(seqidno:140),mfzd8(seqidno:141),hfzd4(seqidno:142),hfzd9(seqidno:143),hfzd10(seqidno:144),hfzd6(seqidno:145)和hfzd3(seqidno:146)之crd的氨基酸序列的比对。图15显示为了评估dfz7-21,dfz7-21-l6a,dfz7-21-w9a,dfz7-21-m13a,dfz7-21-y14a,和dfz7-21δ2对hek293-tb细胞中wnt刺激的β-联蛋白信号传导的影响而实施的实验的结果。图16a提供fz7-21的nosey连通图。图16b显示dfz7-21的代表性nmr溶液结构,其基于dfz7-21的20种最低能量nmr结构的叠加(氨基酸侧链以线显示)。图16c显示dfz7-21的2dnoesy图。图16d显示fz7-21s的2dnoesy图。图16e显示fz7-21,fz7-21s和dfz7-21肽的1dnmr谱。图17a显示dmso处理对代表性小鼠肠类器官的形态的影响。图17b显示fz7-21s处理对代表性小鼠肠类器官的形态的影响。图17c显示抗lrp6阻断性抗体处理对代表性小鼠肠类器官的形态的影响。图17d显示200μm二聚化fz7-21处理对代表性小鼠肠类器官的形态的影响。图17e显示100μm二聚化fz7-21处理对代表性小鼠肠类器官的形态的影响。图17f显示10μm二聚化fz7-21处理对代表性小鼠肠类器官的形态的影响。图17g显示1μm二聚化fz7-21处理对代表性小鼠肠类器官的形态的影响。图18提供在肽处理后类器官干细胞(sc)潜能的量化。图19a提供为了评估dfz7-21或fz7-21s处理对小鼠肠类器官中的lrg5表达的影响而实施的实验的结果。图19b提供为了评估dfz7-21或fz7-21s处理对小鼠肠类器官中的ascl2表达的影响而实施的实验的结果。图19c提供为了评估dfz7-21或fz7-21s处理对小鼠肠类器官中的轴蛋白2表达的影响而实施的实验的结果。图20a提供为了评估dfz7-21或fz7-21s处理对自小鼠收集的肠上皮中的lrg5表达的影响而实施的实验的结果。图20b提供为了评估dfz7-21或fz7-21s处理对自小鼠收集的肠上皮中的ascl2表达的影响而实施的实验的结果。图20c提供为了评估dfz7-21或fz7-21s处理对自小鼠收集的肠上皮中的轴蛋白2表达的影响而实施的实验的结果。图21a显示为了确定肽dfz7-21δ2.m13adp对用50ng/ml重组wnt3a刺激的hek293-tb细胞中的β-联蛋白信号传导的影响而实施的实验的结果。图21b显示为了确定肽dfz7-21δ2.m13tbh对用50ng/ml重组wnt3a刺激的hek293-tb细胞中的β-联蛋白信号传导的影响而实施的实验的结果。图21c显示为了确定肽dfz7-21δ2.m13k(c8)对用50ng/ml重组wnt3a刺激的hek293-tb细胞中的β-联蛋白信号传导的影响而实施的实验的结果。图21d显示为了确定肽dfz7-21δ2.l6hof对用50ng/ml重组wnt3a刺激的hek293-tb细胞中的β-联蛋白信号传导的影响而实施的实验的结果。图21e显示为了确定肽dfz7-21δ2.m13c8对用50ng/ml重组wnt3a刺激的hek293-tb细胞中的β-联蛋白信号传导的影响而实施的实验的结果。图21f显示为了确定肽dfz7-21δ2.m13k(c10)对用50ng/ml重组wnt3a刺激的hek293-tb细胞中的β-联蛋白信号传导的影响而实施的实验的结果。图21g显示为了确定肽dfz7-21δ2.l6hol对用50ng/ml重组wnt3a刺激的hek293-tb细胞中的β-联蛋白信号传导的影响而实施的实验的结果。图21h显示为了确定肽dfz7-21δ2.l6kc(8)对用50ng/ml重组wnt3a刺激的hek293-tb细胞中的β-联蛋白信号传导的影响而实施的实验的结果。图21i显示为了确定肽dfz7-21δ2.m13k(c12)对用50ng/ml重组wnt3a刺激的hek293-tb细胞中的β-联蛋白信号传导的影响而实施的实验的结果。图21j显示为了确定肽dfz7-21δ2.m13k(c14)对用50ng/ml重组wnt3a刺激的hek293-tb细胞中的β-联蛋白信号传导的影响而实施的实验的结果。图21k显示为了确定肽dfz7-21δ2.m13k(c16)对用50ng/ml重组wnt3a刺激的hek293-tb细胞中的β-联蛋白信号传导的影响而实施的实验的结果。图22a显示为了评估在fz7-21以低于10μm的肽浓度存在下wnt5a对fzd1crd,fzd2crd,fzd4crd,和fzd7crd的结合而实施的实验的结果。图22b显示为了评估在dfz7-21以低于10μm的肽浓度存在下wnt5a对fzd1crd,fzd2crd,fzd4crd,和fzd7crd的结合而实施的实验的结果。图22c显示为了评估在fz7-21s以低于10μm的肽浓度存在下wnt5a对fzd1crd,fzd2crd,fzd4crd,和fzd7crd的结合而实施的实验的结果。图22d显示为了评估在fz7-21以低于10μm的肽浓度存在下wnt3a对fzd1crd,fzd2crd,fzd4crd,和fzd7crd的结合而实施的实验的结果。图22e显示为了评估在dfz7-21以低于10μm的肽浓度存在下wnt3a对fzd1crd,fzd2crd,fzd4crd,和fzd7crd的结合而实施的实验的结果。图22f显示为了评估在fz7-21s以低于10μm的肽浓度存在下wnt3a对fzd1crd,fzd2crd,fzd4crd,和fzd7crd的结合而实施的实验的结果。图23a显示为了评估在dfz7-21,fz7-21s,或dfz7-21δ2以低于10μm的肽浓度存在下wnt5a对fzd7crd的结合而实施的实验的结果。图23b显示为了评估在fz7-21s,dfz7-21,或m13adp以低于10μm的肽浓度存在下wnt5a对fzd7crd的结合而实施的实验的结果。图23c显示为了评估在fz7-21s,dfz7-21,或m13tbh以低于10μm的肽浓度存在下wnt5a对fzd7crd的结合而实施的实验的结果。图23d显示为了评估在fz7-21s,dfz7-21,或m13k(c8)以低于约0.5μm的肽浓度存在下wnt5a对fzd7crd的结合而实施的实验的结果。图23e显示为了评估在fz7-21s,dfz7-21,或l6hof以低于10μm的肽浓度存在下wnt5a对fzd7crd的结合而实施的实验的结果。图23f显示为了评估在fz7-21s,dfz7-21,或m13c8以低于10μm的肽浓度存在下wnt5a对fzd7crd的结合而实施的实验的结果。图23g显示为了评估在fz7-21s,dfz7-21,或m13k(c10)以低于10μm的肽浓度存在下wnt5a对fzd7crd的结合而实施的实验的结果。图23h显示为了评估在fz7-21s,dfz7-21,或l6hol以低于10μm的肽浓度存在下wnt5a对fzd7crd的结合而实施的实验的结果。图23i显示为了评估在fz7-21s,dfz7-21,或l6k(c8)以低于10μm的肽浓度存在下wnt5a对fzd7crd的结合而实施的实验的结果。图23j显示为了评估在fz7-21s,dfz7-21,m13k(c8),m13k(c10),m13k(c12),m13k(c14),或m13k(c16)以低于10μm的肽浓度存在下wnt5a对fzd7crd的结合而实施的实验的结果。图23k显示为了评估在fz7-21s,dfz7-21,dfz7-21δ2.m13adp,dfz7-21δ2.m13tbh,dfz7-21δ2.m13k(c8),dfz7-21δ2.l6hof,dfz7-21δ2.m13c8,dfz7-21δ2.m13k(c10),dfz7-21δ2(q519),dfz7-21δ2.l6hol,或dfz7-21δ2.l6kc(8)以低于10μm的肽浓度存在下wnt5a对fzd4crd的结合而实施的实验的结果。图23l显示为了评估在fz7-21s,dfz7-21,m13k(c8),m13k(c10),m13k(c12),m13k(c14),或m13k(c16)以低于10μm的肽浓度存在下wnt5a对fzd4crd的结合而实施的实验的结果。图24a显示通过uv吸收分析的分子量(mw)标准物,作为洗脱体积(ve)在空隙体积(vo)上的函数绘图。数值代表三项独立实验的均值±s.e.m.。图24b显示与5fam-fz7-21结合的fzdcrd-fc蛋白质的观察分子量(灰色圆形)较之预测fzdcrd-fc四聚体mw(黑色方形)。图24c显示所使用的不同fzdcrd-fc蛋白质(~2μg)的天然page(4-16%)。图25a显示脱辅基hfzd7crd晶体结构的飘带展示及图示fzd7内的crd放置的全长fzd7的示意图。图25b显示与fz7-21结合的hfzd7crd的结构的飘带展示。图25c提供脱辅基hfzd7crd中的疏水腔的放大侧视图。图25d提供来自图25c的顶视图。图25e提供与fz7-21结合的hfzd7crd的放大侧视图。图25f显示图25e的顶视图。图26显示dfz7-21的20种最低能量nmr结构的叠加。图27a显示用于测定hfzd7crd-gs的mw的相关分子量标准物。图27b显示图10b中的融合蛋白在还原性条件下的sds-page。图27c显示自图10b中的融合蛋白获得的晶体的明视野图像。图28a提供与它们的相应c24脂肪酸结合的hfzd7crd原体的叠加。图28b显示来自图28a的没有飘带展示的结构,展示接近c24脂肪酸的残基。c24脂肪酸碳自羧酸首基(c1)开始顺序编号至ω碳(c24)。水,结晶辅因子,和聚糖为清楚起见而隐藏。图28c提供脱辅基hfzd7crd(飘带展示)和与c24脂肪酸(球和棍展示)结合的hfzd7crd(飘带展示)的叠加。图29a提供与c24脂肪酸(球和棍展示)复合的hfzd7crd二聚体(飘带展示)的晶体结构。图29b提供定位到与c24脂肪酸(黑色;球和棍展示)复合的hfzd7crd(飘带展示)结构上的疏水腔的表面展示。图29c提供图29b的放大视图,展示接近疏水腔的残基(灰色,a链;白色,b链;c24脂肪酸,黑色;球和棍展示)。c24脂肪酸碳自羧酸首基(c1)开始顺序编号至ω碳(c24)。脂质结合腔的基部介于c9和c13之间。图29d显示脱辅基hfzd7crd(pdbid5t44;浅灰色,棍展示)和与c24脂肪酸(黑色,空间填充模型)结合的hfzd7crd(灰色,棍展示)的脂质结合腔的叠加。展示了氢键相互作用(黑色;短划线)。图30a显示与20(s)-羟基胆固醇(灰色;棍展示)结合的平滑crd(灰色;飘带展示;pdbid#5kzv)和与c24脂肪酸(白色;棍展示)结合的hfzd7crd(白色;飘带展示)的叠加(在108个原子对上r.m.s.)。放大插图突显紧密接近配体羟基基团的残基且展示了氢键相互作用(黑色;短划线)。图30b显示与20(s)-羟基胆固醇(灰色;棍展示)结合的平滑crd(灰色;飘带展示;pdbid#5kzv)和与c16:1脂肪酸(深灰色;棍展示)结合的hfzd5crd(褐色,a链;飘带展示)的叠加(在109个原子对上r.m.s.偏差为)。放大插图突显紧密接近配体羟基基团的残基且展示了氢键相互作用(黑色;短划线)。图31显示与c16:1n-7脂肪酸(黑色或深灰色;球和棍展示)结合的人(h)fzd5crd(a链,浅灰色;b链,中等灰色;飘带展示)的结构。a链内的每个c16:1n-7脂肪酸因对称配合平均而具有50%占据率。图32a显示与c16:1n-7脂肪酸(深灰色或黑色;球和棍展示)复合的hfzd5crd同二聚体(a链;飘带展示)的晶体结构。图32b提供定位到与每个具有50%占据率的两个c16:1n-7脂肪酸(黑色或灰色;球和棍展示)复合的hfzd5crd(飘带展示)上的hfzd5crda链同二聚体疏水腔(灰色)的表面展示。图32c显示图32b的没有飘带展示的放大视图。突显接近疏水腔的残基(深灰色,棍展示)。c16:1n-7脂肪酸碳自羧酸首基(c1)开始顺序编号至ω碳(c16)。脂质结合腔的基部介于c7和c10之间。图33a提供与每个具有50%占据率的两分子n-辛基-β-d-葡萄糖苷(bog)(灰色或黑色;球和棍展示)复合的人(h)fzd5crd二聚体的晶体结构。图33b显示定位到与n-辛基-β-d-葡萄糖苷(球和棍展示)结合的hfzd5crda链(白色;飘带展示)的结构上的hfzd5crda链之疏水腔的表面展示,突显距n-辛基-β-d-葡萄糖苷(展示)~内的残基。图33c显示图33b的没有展示碳主链的放大视图。突显hfzd5crd和n-辛基-β-d-葡萄糖苷的糖苷之间的氢键相互作用(短划线;黑色)。图33d显示n-辛基-β-d-葡萄糖苷的分子结构,突显分子构件。图34a显示c16:1n-7脂肪酸(飘带和棍展示)或n-辛基-β-d-葡萄糖苷(飘带和棍展示)结合的hfzd5crd的叠加(119个残基上r.m.s.偏差为)。a链内的每个c16:1n-7脂肪酸和n-辛基-β-d-葡萄糖苷因对称配合平均而具有50%占据率。图34b显示来自图34a的hfzd5crdb链的飘带展示,突显接近c16:1n-7脂肪酸的选定残基。图34c显示来自图34a的hfzd5crda链,突显接触c16:1n-7脂肪酸或n-辛基-β-d-葡萄糖苷的残基(vdw交叠>至-)。放大视图描绘显示fzd5crd和n-辛基-β-d-葡萄糖苷之间的氢键相互作用(黑色短划线)。图35a显示mfzd8crd二聚体(pdbid#1ijy;飘带展示)的晶体结构及同二聚体疏水腔的表面展示。指示了来自实施例和dann,c.e.etal.“insightsintowntbindingandsignalingfromthestructuresoftwofrizzledcysteine-richdomains.”nature412,86-90(2001)的二聚体界面。图35b显示a链和b链同二聚体的叠加及它们的疏水腔的表面展示。图35c显示图35b的放大视图,突显接近mfzd8crd之疏水腔的侧链。图35d显示bog结合的hfzd5crd(飘带展示),c16:1n-7脂肪酸结合的hfzd5crd(飘带展示),脱辅基fzd7crd(飘带展示),c24脂肪酸结合的hfzd7crd(飘带展示)和脱辅基mfzd8crd(飘带展示)的叠加。图35e显示wnt结合fzd受体之crd的一种模型,利用u形疏水腔实现顺式脂肪酸选择性。fzd5,7和8(可能fzd1和fzd2)受体(crd,褐色或蓝色椭圆形;7次跨膜域,黄色椭圆形)处于它们的无活性状态且可能形成二聚体构象,其中疏水腔形成连续且u形腔(红色)。一旦wnt结合,顺式-δ9-不饱和脂肪酸就占据脂质结合腔,利用“扭结”来横越二聚体界面,并募集辅因子来刺激下游wnt信号传导。图36a显示hfzd7crd,hfzd5crd和hfzd8crd的二聚体相互作用界面势能的表。图36b显示fzdcrd二聚体互补性得分(sc)的表。图36c显示hfzd7crd的环-环相互作用界面的二聚体互补性得分的可视化。依照二聚体互补性得分,相互作用界面的中心假设具有低互补性且外围处的螺旋具有高互补性。图36d显示mfzd8crd的环-环相互作用界面的二聚体互补性得分的可视化。依照二聚体互补性得分,相互作用界面的中心具有低互补性且外围处的螺旋具有高互补性。图36e显示hfzd5crd的环-环相互作用界面的二聚体互补性得分的可视化。依照二聚体互补性得分,相互作用界面的中心具有低互补性且外围处的螺旋具有高互补性。图36f显示hfzd5crd的fzd7样α螺旋二聚体相互作用界面的二聚体互补性得分的可视化。依照二聚体互补性得分,相互作用界面的中心具有低互补性且外围处的螺旋具有高互补性。图36g显示hfzd7crd的fzd7样α螺旋二聚体相互作用界面的二聚体互补性得分的可视化。依照二聚体互补性得分,相互作用界面的中心具有低互补性且外围处的螺旋具有高互补性。图36h显示hfzd7crd的fzd7样α螺旋二聚体相互作用界面的二聚体互补性得分的可视化。依照二聚体互补性得分,相互作用界面的中心具有低互补性且外围处的螺旋具有高互补性。图37a显示人fzdcrd家族成员的clustalomega序列比对。形成原体之间的晶体学fzd7样二聚体接触的残基标有下划线(vdw交叠>-0.4埃)。fzd家族成员之间fzd7样二聚体界面的保守性以序列比对上方的“+”表示。保守的半胱氨酸以蓝色突显。fzd7(seqidno:147),fzd2(seqidno:148),fzd1(seqidno:149),fzd5(seqidno:150),fzd8(seqidno:151),fzd4(seqidno:152),fzd9(seqidno:153),fzd10(seqidno:154),fzd6(seqidno:155),和fzd3(seqidno:156)。图37b显示使用脱辅基hfzd7crd(飘带展示)的结构作为替代品突显具有180°旋转的α螺旋fzd7二聚体界面。插图展示二聚体界面的放大视图且突显残基-残基氢键(黑色短划线)。图38a显示将与mfzd8crd(白色;飘带展示;pdbid#4f0a)复合的xwnt8(浅灰色;飘带展示)叠加到mfzd8crd(灰色;飘带展示;pdbid#1ijy)的α螺旋二聚体上。所示二聚体的疏水腔表面定位到前表面透明的fzdcrd结构上。c14脂肪酸(球和棍展示)在xwnt8s187处共价结合。水,结晶辅因子和糖基化为清楚起见而隐藏。图38b显示与复合了c24脂肪酸(黑色;球和棍展示)的hfzd7crd(灰色;飘带展示)复合的xwnt8(浅灰色;飘带展示)。所示二聚体的疏水腔表面定位到前表面透明的fzdcrd结构上。c14脂肪酸(浅灰色;球和棍展示)在xwnt8s187处共价结合。水,结晶辅因子和糖基化为清楚起见而隐藏。图38c显示与复合了c16:1n-7脂肪酸(灰色或黑色;球和棍展示)的hfzd5crda链同二聚体(灰色;飘带展示)复合的xwnt8(浅灰色;飘带展示)。所示二聚体的疏水腔表面定位到前表面透明的fzdcrd结构上。c14脂肪酸(浅灰色;球和棍展示)在xwnt8s187处共价结合。水,结晶辅因子和糖基化为清楚起见而隐藏。图39a提供与hfzd7crd(pdbid#5urv)复合的xwnt8a(pdbid#4f0a)的一种模型,其中延长的脂肪酰基模块(c16:n-7)在hfzd7crd的u形疏水腔中结合。图39b提供图39a的黑色框中的部分的放大视图。图39c提供在肽fz7-21存在下wnt与fzd7crd的相互作用的一种建议模型。图39d提供图39c的黑色框中的部分的放大视图。图40提供为了评估hfzd7crd上的肽结合区内的特定残基的突变是否与野生型hfzd7crd相比降低fz7-21的结合而实施的荧光大小排阻层析术(fsec)实验的结果。图41a提供为了确定dfz7-21处理是否降低自lgr5-gfp小鼠衍生的类器官中lgr5-gfp干细胞的数目而实施的实验的结果。图41b提供为了确定dfz7-21处理是否降低自lgr5-gfp小鼠衍生的类器官中lgr5-gfp干细胞的数目而实施的实验的结果。图41c提供图41a和41b中显示的结果的量化。图42a提供为了评估dmso,fz7-21或fz7-21s处理对lgr5-gfp类器官中的轴蛋白2mrna水平的影响而实施的实验的结果。图42b提供为了评估dmso,fz7-21或fz7-21s处理对lgr5-gfp类器官中的ascl2mrna水平的影响而实施的实验的结果。图42c提供为了评估dmso,fz7-21或fz7-21s处理对apcmin类器官中的轴蛋白2mrna水平的影响而实施的实验的结果。图42d提供为了评估dmso,fz7-21或fz7-21s处理对apcmin类器官中的ascl2mrna水平的影响而实施的实验的结果。图42e提供为了评估dmso,fz7-21或fz7-21s处理对apcmin类器官中的lgr5mrna水平的影响而实施的实验的结果。发明详述提供的是包含结合或特异性结合卷曲蛋白7(fzd7)受体之富半胱氨酸域(crd)的非天然发生肽的配体。在某些实施方案中,这些配体进一步结合选自由卷曲蛋白1(fzd1)和卷曲蛋白2(fzd2)组成的组的卷曲蛋白(fzd)受体之富半胱氨酸域(crd)。此类配体展现下面的特征中的一项或多项:以小于约100nm的ic50抑制wnt介导的β-联蛋白信号传导;以小于90nm的ec50值结合人fzd7crd和小鼠fzd7crd,和/或结合包含三个或更多个下述氨基酸的人fzd7crd之结合区:leu81,his84,gln85,tyr87,pro88,phe138,和phe140。还提供的是在治疗受试者中的癌症(诸如结肠癌,胰腺癌,非小细胞肺癌,特征在于rnf43中的突变的癌症,特征在于usp6过表达的癌症,或特征在于牵涉r-spondin(rspo)家族成员的基因融合的癌症)中使用包含结合或特异性结合fzd7之crd的非天然发生肽的配体的方法。还提供的是在治疗受试者中的癌症(诸如结肠癌,胰腺癌,非小细胞肺癌,特征在于rnf43中的突变的癌症,特征在于usp6过表达的癌症,或特征在于牵涉r-spondin(rspo)家族成员的基因融合的癌症)中使用包含进一步结合fzd1和/或fzd2之crd的非天然发生肽的配体的方法。还提供的是包含结合或特异性结合fzd7之crd的非天然发生肽的配体在制造用于治疗眼部疾病或病症的药物中的用途。还提供的是此类包含进一步结合fzd1和/或fzd2之crd的非天然发生肽的配体在制造用于治疗眼部疾病或病症的药物中的用途。除非另外指明,本公开文本的实践采用细胞生物学,毒理学,分子生物学,生物化学,细胞培养,免疫学,肿瘤学,重组dna领域和本领域技术内的相关领域中的标准方法和常规技术。此类技术在文献中有描述且由此是本领域技术人员可得的。参见例如alberts,b.etal.,“molecularbiologyofthecell,”5thedition,garlandscience,newyork,ny,2008;voet,d.etal.,“fundamentalsofbiochemistry:lifeatthemolecularlevel,”3rdedition,johnwiley&sons,hoboken,nj,2008;sambrook,j.etal.,“molecularcloning:alaboratorymanual,”3rdedition,coldspringharborlaboratorypress,2001;ausubel,f.etal.,“currentprotocolsinmolecularbiology,”johnwiley&sons,newyork,1987和定期更新;freshney,r.i.,“cultureofanimalcells:amanualofbasictechnique,”4thedition,johnwiley&sons,somerset,nj,2000;和丛书“methodsinenzymology,”academicpress,sandiego,ca。定义如本文中使用的,“非天然发生”意味着例如包含在自然界中没有找到的氨基酸序列的多肽或例如包含在自然界中没有找到的核苷酸序列的核酸。本文中提供的非天然发生肽可以通过遗传工程方法或通过化学合成方法来生成。如此,本文中描述的非天然发生肽可以是重组的,即由已经通过引入异源核酸或将天然核酸改变成对该细胞不是天然的形式而修饰的细胞,或核酸,或载体,或自如此修饰的细胞衍生的细胞生成的。或者,本文中描述的非天然发生肽可以经由化学肽合成来生成。如本文中使用的,“氨基酸改变”指例如肽序列中(诸如结合或特异性结合fzd7之富半胱氨酸域(crd)的非天然发生肽的序列中)至少一个氨基酸的添加,删除,或替代。如本文中使用的,“配体”指包含(诸如本质上由其组成或由其组成)至少一种本文中描述的结合或特异性结合例如卷曲蛋白7(fzd7)之富半胱氨酸域(crd)的非天然发生肽的分子。包含(诸如本质上由其组成或由其组成)至少一种本文中描述的非天然发生肽的配体对crd的结合可测量地不同于非特异性相互作用,而且可以通过例如测量蛋白质-配体结合的结合测定法或免疫测定法来检测。本发明的“结合”感兴趣受体的配体是如下的,其以足够亲和力结合受体,使得配体在靶向蛋白质或表达受体的细胞或组织中作为诊断和/或治疗剂是有用的。关于配体对靶分子或受体的结合,术语“特异性结合”特定多肽或特定多肽靶上的表位或“对其特异性的”意味着可测量地不同于非特异性相互作用的结合。特异性结合可以例如通过与对照分子的结合相比测定分子的结合来测量。例如,特异性结合可以通过与同靶物相似的对照分子(例如过量的未标记的靶物)的竞争来测定。在这种情况中,如果经标记的靶物对探针的结合受到过量的未标记的靶物的竞争性抑制,那么指示特异性结合。在一个特定的实施方案中,“特异性结合”指配体对它的规定靶fzd7crd域而非其它规定fzdcrd域的结合。例如,配体特异性结合fzd7crd但并不特异性结合fzd8crd。在某些实施方案中,配体对“非靶”fzd受体(诸如fzd3,fzd4,fzd5,fzd6,fzd8,fzd9,或fzd10)的结合的程度会小于配体对fzd1,fzd2,和/或fzd7的结合的约10%,如通过例如荧光激活细胞分选(facs)分析或放射免疫沉淀(ria)测定的。在某些实施方案中,本公开文本的配体以等于或低于100nm,任选低于10nm,任选低于1nm,任选低于0.5nm,任选低于0.1nm,任选低于0.01nm,或任选低于0.005nm的解离常数(kd)或ic50值特异性结合fzd1,fzd2,和/或fzd7;于约4℃,25℃,37℃,或45℃的温度测量。在某些实施方案中,配体对“非靶”fzd受体(诸如fzd1,fzd2,fzd3,fzd4,fzd5,fzd6,fzd8,fzd9,或fzd10)的结合的程度会小于配体对fzd7的结合的约10%,如通过例如荧光激活细胞分选(facs)分析或放射免疫沉淀(ria)测定的。在某些实施方案中,本公开文本的配体以等于或低于100nm,任选低于10nm,任选低于1nm,任选低于0.5nm,任选低于0.1nm,任选低于0.01nm,或任选低于0.005nm的解离常数(kd)或ic50值特异性结合fzd7;于约4℃,25℃,37℃,或45℃的温度测量。“分离的”配体是如下的,其已经自包含配体和污染物或杂质的组合物鉴定和分出和/或回收。污染物或杂质是会干扰包含(诸如本质上由其组成或由其组成)结合或特异性结合fzd7之富半胱氨酸域(crd)的非天然发生肽的配体的诊断或治疗用途的材料。污染物可包括例如宿主细胞酶,激素,和其它蛋白质性质或非蛋白质性质的溶质,即如果配体是重组生成的话,或例如盐,试剂,截短的或降解的序列,不完全去保护的序列,等,即如果配体是经由化学合成而生成的话。在优选的实施方案中,本文中提供的配体会纯化(1)至大于95%,以配体的重量计,如通过lowry方法测定的,和最优选超过99%,以重量计,(2)至足以通过使用转杯式测序仪获得至少15个残基的n端或内部氨基酸序列的程度,或(3)至同质,根据在还原性或非还原性条件下的sds-page,使用考马斯蓝或优选银染剂。分离的配体包括重组细胞内的原位配体。分离的包含(诸如本质上由其组成或由其组成)结合或特异性结合fzd7之富半胱氨酸域(crd)的非天然发生肽的配体会通过至少一个纯化步骤来制备。关于本文中鉴定的多肽序列的“百分比(%)氨基酸序列同一性”或“同源性”定义为作为序列同一性的一部分考虑任何保守替代,在比对序列之后,候选序列中与所比较的多肽中的氨基酸残基相同的氨基酸残基的百分比。为了确定百分比氨基酸序列同一性目的的比对可以以本领域技术内的多种方式来实现,例如使用公众可得的计算机软件,诸如blast,blast-2,align或megalign(dnastar)软件。本领域技术人员能确定用于测量比对的适宜参数,包括在所比较的序列的全长上实现最大比对所需要的任何算法。然而,为了本文中的目的,%氨基酸序列同一性值是使用序列比较计算机程序align-2生成的。align-2序列比较计算机程序由genentech,inc.编写且源代码已经与用户文档一起提交美国版权局(washingtond.c.,20559),在那里它以美国版权登记号txu510087注册。公众经由genentech,inc.(southsanfrancisco,california)可得到align-2程序。align-2程序应当汇编成供在unix操作系统,优选数码unixv4.0d上使用。所有序列比较参数由align-2程序设定且不变化。如本文中使用的,术语“结合区”指能够受到肽(诸如本文中提供的特异性结合fzd7之富半胱氨酸域(crd)的非天然发生肽)特异性结合的区域。结合区可包含对于结合区独特的空间构象中的约3-10个氨基酸。这些氨基酸可以是在蛋白质内线性的(即在氨基酸序列中连续的),或者它们可以位于蛋白质的不同部分(即在氨基酸序列中不连续的)。测定蛋白质内的或两个蛋白质的界面处的氨基酸的空间构象的方法是本领域知道的,而且包括例如x射线晶体学和二维核磁共振。为了治疗目的的“受试者”,“患者”,或“个体”指归类为哺乳动物的任何动物,包括人,驯养的和农场动物,和动物园,运动,或宠物动物,诸如犬,马,猫,牛,等。优选地,哺乳动物是人。如本文中公开的配体(或包含此类配体的组合物)的“有效量”是足以实现具体描述的目的的量。“有效量”可以根据描述的目的凭检验和通过已知方法来确定。术语“治疗有效量”指如本文中公开的配体或组合物有效“治疗”哺乳动物(诸如人患者)中的疾病或病症的量。在癌症的情况中,治疗有效量的本文中公开的配体能减少癌细胞的数目;降低肿瘤尺寸或重量;抑制(即一定程度地减缓和优选停止)癌细胞浸润入周围器官;抑制(即一定程度地减缓和优选停止)肿瘤转移;一定程度地抑制肿瘤生长;和/或一定程度地缓解一种或多种与癌症有关的症状。就如本文中公开的配体能阻止现有癌细胞生长和/或杀死现有癌细胞的程度而言,它可以是细胞抑制性的和/或细胞毒害性的。在一个实施方案中,治疗有效量是生长抑制量。在另一个实施方案中,治疗有效量是延长患者的存活的量。在另一个实施方案中,治疗有效量是改善患者的无进展存活的量。如本文中使用的,“处理/治疗”是用于获得有益的或想要的结果(包括临床结果)的办法。为了本发明的目的,有益的或想要的临床结果包括但不限于下面的一项或多项:缓解一种或多种源自疾病的症状,减轻疾病的程度,稳定疾病(例如预防或延迟疾病的恶化),预防或延迟疾病的扩散(例如转移),预防或延迟疾病的复发,延迟或减缓疾病的进展,改善疾病状态,提供疾病的消退(部分的或全部的),降低治疗疾病所需要的一种或多种其它药疗的剂量,延迟疾病的进展,提高或改善生命质量,提高重量增益,和/或延长存活。“处理/治疗”还涵盖降低癌症的病理学后果(诸如例如肿瘤体积)。本发明的方法涵盖处理/治疗的这些方面中的任一项或多项。“病症”是会受益于用包含(诸如本质上由其组成或由其组成)结合fzd7之富半胱氨酸域(crd)的非天然发生肽的配体或用包含(诸如本质上由其组成或由其组成)本文中描述的特异性结合fzd7之crd的非天然发生肽的配体治疗的任何状况。例如,罹患异常wnt表达或活性或需要针对异常wnt表达或活性的预防的哺乳动物。这包括慢性和急性病症或疾病,包括那些使哺乳动物倾向于所讨论的病症的病理学状况。本文中要治疗的病症的非限制性例子包括如本文中别处描述的癌症和转移性疾病。在某些实施方案中,可使用包含(诸如本质上由其组成或由其组成)结合fzd7之富半胱氨酸域的非天然发生肽的配体或包含(诸如本质上由其组成或由其组成)特异性结合fzd7之crd的非天然发生肽的配体来促进组织修复,伤口愈合,和骨生长。如本文中使用的,“制药学可接受的”或“药理学相容的”意味着材料不是生物学或其它方面不想要的,例如可以将材料掺入施用于患者的药物组合物,没有引起任何显著的不想要的生物学效应或与含有它的组合物的任何其它成分以有害方式相互作用。制药学可接受载剂或赋形剂优选达到毒理学和制造测试的期望标准和/或包括在由美国食品和药品管理局制备的无活性组分指导上。术语“检测”意图包括确定物质的存在或缺失或量化物质(诸如fzd7或其受体)的量。该术语如此指使用本文中提供的材料,组合物,和方法进行定性和定量测定。一般而言,用于检测的特定技术对于本发明的实践不是至关紧要的。例如,“检测”依照本发明可包括:观察fzd7的存在或缺失;fzd7水平的变化;和/或fzd7生物学功能/活性的变化。在某些实施方案中,“检测”可包括检测fzd7的水平(例如人fzd1,人fzd2,和/或人fzd7的多肽水平)。检测可包括量化当与对照相比时介于10%和90%之间的任何值,或介于30%和60%之间的任何值,或超过100%的变化(升高或降低)。检测可包括量化介于2倍至10倍之间的(含端点)或更多的(例如100倍)任何值的变化。当在本文中使用时,词语“标记物”指与包含(诸如本质上由其组成或由其组成)特异性结合卷曲蛋白7(fzd7)之富半胱氨酸域(crd)的非天然发生肽的配体直接或间接缀合的可检测化合物或组合物。标记物可以自身是通过自身可检测的(例如放射性同位素标记物或荧光标记物),或者在酶标记物的情况中,可以催化可检测的底物化合物或组合物的化学改变。在本文中提到“约”数值或参数指这个
技术领域:
中的技术人员容易知道的相应数值的通常误差范围。在本文中提到“约”数值或参数包括(且描述)指向该数值或参数本身的方面。例如,提到“约x”的描述包括“x”的描述。理解的是,本文中描述的发明的各方面和实施方案包括“包含”,“由……组成”,和“本质上由……组成”方面和实施方案。据此通过援引完整收录本文中引用的所有参考文献,包括专利申请和出版物。包含结合fzd7之富半胱氨酸域的非天然发生肽的配体提供的是一种包含(诸如本质上由其组成或由其组成)结合或特异性结合卷曲蛋白7(fzd7)之富半胱氨酸域(crd)的非天然发生肽的配体。在某些实施方案中,配体结合一种或多种选自由卷曲蛋白1(fzd1),卷曲蛋白2(fzd2),和卷曲蛋白7(fzd7)组成的组的卷曲蛋白(fzd)受体。在某些实施方案中,fzd1是人fzd1。在某些实施方案中,fzd1是小鼠fzd1。在某些实施方案中,fzd2是人fzd2。在某些实施方案中,fzd2是小鼠fzd2。在某些实施方案中,fzd7是人fzd7。在某些实施方案中,fzd7是小鼠fzd7。在具体的实施方案中,配体包含(诸如本质上由其组成或由其组成)特异性结合卷曲蛋白7(fzd7)(例如人fzd7或小鼠fzd7)之富半胱氨酸域(crd)的非天然发生肽。在某些实施方案中,配体包含(诸如本质上由其组成或由其组成)不结合卷曲蛋白3(fzd3),卷曲蛋白4(fzd4),卷曲蛋白5(fzd5),卷曲蛋白6(fzd6),卷曲蛋白8(fzd),卷曲蛋白9(fzd9)或卷曲蛋白10(fzd10)之crd的非天然发生肽。在某些实施方案中,配体包含(诸如本质上由其组成或由其组成)不结合卷曲蛋白1(fzd1),卷曲蛋白2(fzd2),卷曲蛋白3(fzd3),卷曲蛋白4(fzd4),卷曲蛋白5(fzd5),卷曲蛋白6(fzd6),卷曲蛋白8(fzd),卷曲蛋白9(fzd9)或卷曲蛋白10(fzd10)之crd的非天然发生肽。在某些实施方案中,配体包含(诸如本质上由其组成或由其组成)特异性结合或特异性结合fzd7之crd的非天然发生肽。在某些实施方案中,配体包含(诸如本质上由其组成或由其组成)结合或特异性结合hfzd7之crd的非天然发生肽。在某些实施方案中,配体包含(诸如本质上由其组成或由其组成)结合或特异性结合mfzd7之crd的非天然发生肽。在某些实施方案中,配体包含(诸如本质上由其组成或由其组成)结合或特异性结合fzd7之crd且另外结合1)fzd1,2)fzd2,或fzd1和fzd2中任一之crd的非天然发生肽。在某些实施方案中,配体包含(诸如本质上由其组成或由其组成)结合fzd1和fzd7之crd的非天然发生肽。在某些实施方案中,配体包含(诸如本质上由其组成或由其组成)结合fzd2和fzd7之crd的非天然发生肽。在某些实施方案中,配体包含竞争对受到包含氨基酸序列lpsddlefwchvmy(seqidno:13)的肽结合的fzd7crd之结合区的结合的非天然发生肽。在某些实施方案中,配体本质上由竞争对受到包含氨基酸序列lpsddlefwchvmy(seqidno:13)的肽结合的fzd7crd之结合区的结合的非天然发生肽组成(诸如由其组成)。在某些实施方案中,配体本质上由竞争对受到由氨基酸序列lpsddlefwchvmy(seqidno:13)组成的肽结合的fzd7crd之结合区的结合的非天然发生肽组成(诸如由其组成)。在某些实施方案中,配体包含特异性结合受到包含氨基酸序列lpsddlefwchvmy(seqidno:13)的肽结合的相同的fzd7crd之结合区的非天然发生肽。在某些实施方案中,配体本质上由特异性结合受到包含氨基酸序列lpsddlefwchvmy(seqidno:13)的肽结合的相同的fzd7crd之结合区的非天然发生肽组成(诸如由其组成)。在某些实施方案中,配体本质上由特异性结合受到由氨基酸序列lpsddlefwchvmy(seqidno:13)组成的肽结合的相同的fzd7crd之结合区的非天然发生肽组成(诸如由其组成)。在某些实施方案中,配体包含(诸如本质上由其组成或由其组成)特异性结合包含至少一个,至少两个,至少三个,至少四个,至少五个,至少六个,或至少七个选自由leu81,his84,gln85,tyr87,pro88,phe138,和phe140组成的组的氨基酸的fzd7之结合区的肽。在某些实施方案中,配体包含(诸如本质上由其组成或由其组成)特异性结合包含至少一个,至少两个,至少三个,至少四个,至少五个,至少六个,或至少七个选自由leu81,his84,gln85,tyr87,pro88,phe138,和phe140组成的组的氨基酸的fzd7之结合区的非天然发生肽。在某些实施方案中,配体包含(诸如本质上由其组成或由其组成)特异性结合距至少一个,至少两个,至少三个,至少四个,至少五个,至少六个,或至少七个选自由leu81,his84,gln85,tyr87,pro88,phe138,和phe140组成的组的氨基酸在内的fzd7之结合区的非天然发生肽。在某些实施方案中,配体是嵌合分子,其包含与一个或多个模块融合的本文中描述的结合或特异性结合fzd1,fzd2,和/或fzd7之crd的非天然发生肽。在某些实施方案中,配体是嵌合分子,其包含与一个或多个模块融合的本文中描述的特异性结合fzd7之crd的非天然发生肽。在某些实施方案中,模块融合至肽的n端。在某些实施方案中,模块融合至肽的c端。在某些实施方案中,第一模块融合至肽的n端,且第二模块融合至肽的n端。在某些实施方案中,一个或多个模块重组融合至肽。在某些实施方案中,一个或多个模块经由接头(诸如可切割接头)连接至肽。例示性的模块包括但不限于肽,多肽或其片段,蛋白质或其片段,融合蛋白。在某些实施方案中,配体包含与异源多肽或氨基酸序列重组融合的本文中描述的结合或特异性结合fzd7之crd的非天然发生肽。在某些实施方案中,配体包含与将配体靶向例如用于投递至各种组织或例如穿越血脑屏障的蛋白质转导域(使用例如人免疫缺陷病毒tat蛋白的蛋白质转导域(schwarzeetal.,1999,science285:1569-72))融合(例如在n端,c端,或n和c端二者)的本文中描述的结合或特异性结合fzd7之crd的非天然发生肽。在某些实施方案中,配体包含与细胞可渗透肽,诸如初级两亲性肽mpg(galflgflgaagstmgawsqpkkkrkvseqidno:104),pep-1(ketwwetwwtewsqpkkkrkvseqidno105),次级两亲性肽cady(ac-glwralwrllrslwrllwra-cyaseqidno:106)或八精氨酸(r(8)seqidno:107)融合的本文中描述的结合或特异性结合fzd7之crd的非天然发生肽。在某些实施方案中,配体包含与稳定非天然发生肽的构象(诸如α螺旋结构)的域或模块融合(例如在n端,c端,或n和c端二者)的本文中描述的结合或特异性结合fzd7之crd的非天然发生肽。在某些实施方案中,包含(诸如本质上由其组成或由其组成)本文中描述的结合或特异性结合fzd7之crd的非天然发生肽的配体可以作为单特异性的以单体形式或作为双或多特异性的(针对不同靶配体或相同靶配体的不同结合区)以多聚体形式使用。在某些实施方案中,包含(诸如本质上由其组成或由其组成)本文中描述的结合或特异性结合fzd7之crd的非天然发生肽的配体是与lrp6缀合的。附着可以是共价的或非共价的。在某些实施方案中,二聚体双特异性配体具有有针对第一靶或结合区的特异性的一个亚基和有针对第二靶配体或结合区的特异性的第二。包含本文中描述的结合或特异性结合fzd7之crd的非天然发生肽的配体可以以能提高结合的效价和由此亲合力的多种构造接合。在某些实施方案中,本文中提供的配体包含两个或更多个(诸如三个,四个,五个,六个,七个,八个,九个,十个,或超过十个)本文中提供的结合或特异性结合fzd7之crd的非天然发生肽。在某些实施方案中,可以工程化改造核酸以编码两个或更多个拷贝的单一的本文中提供的结合或特异性结合fzd7之crd的非天然发生肽,拷贝串联转录并翻译以生成相同亚基的共价连接的多聚体。在某些实施方案中,可以工程化改造核酸以编码两个或更多个不同的本文中提供的结合或特异性结合fzd7之crd的非天然发生肽,拷贝串联转录并翻译以生成不同亚基的共价连接的多聚体。在另一个实施方案中,配体包含与提供抗标签抗体能选择性结合的表位的标签多肽融合的本文中提供的结合或特异性结合fzd7之crd的非天然发生肽。表位标签一般放置在配体的氨基或羧基端处。此类带表位标签形式的配体的存在可以使用针对标签多肽的抗体来检测。还有,表位标签的提供使得包含本文中提供的结合或特异性结合fzd7之crd的非天然发生肽的配体能够容易地使用抗标签抗体或另一种类型的特异性结合表位标签的亲和基质通过亲和纯化来纯化。各种标签多肽和它们的相应抗体是本领域知道的。例子包括聚组氨酸(poly-his)或聚组氨酸-甘氨酸(poly-his-gly)标签;流感ha标签多肽和它的抗体12ca5(fieldetal.(1988)mol.cell.biol.8,2159-2165);c-myc标签及其8f9,3c7,6e10,g4,b7和9e10抗体(evanetal.(1985)mol.cell.biol.5,3610-3616);和单纯疱疹病毒糖蛋白d(gd)标签和它的抗体(paborskyetal.(1990)proteineng.,3,547-553)。其它标签多肽包括flag肽(hoppetal.(1988)biotechnology,6,1204-1210);kt3表位肽(martinetal.(1992)science,255,192-194);α-微管蛋白表位肽(skinneretal.(1991)j.biol.chem.266,15163-15166);和t7基因10蛋白质肽标签(lutz-freyermuthetal.(1990)proc.natl.acad.sci.usa87,6393-6397)。在某些实施方案中,配体包含与免疫球蛋白或免疫球蛋白的特定区域重组融合的本文中提供的结合或特异性结合fzd7之crd的非天然发生肽。对于二价形式的配体(例如“免疫粘附素”),此类融合物可以是与igg分子的fc区的。本文中提供的ig融合物包括包含大约或仅仅人的残基94-243,残基33-53或残基33-52来替换ig分子内的至少一个可变区的多肽。在一个特别优选的实施方案中,免疫球蛋白融合物包括igg1分子的铰链,ch2和ch3,或铰链,ch1,ch2和ch3区。对于免疫球蛋白融合物的生成,还参见1995年6月27日公告的美国专利no.5,428,130。在某些实施方案中,包含本文中提供的结合或特异性结合fzd7之crd的非天然发生肽的配体是例如在n或c端处与igg的恒定区(fc)融合的。在某些实施方案中,配体/fc融合分子活化免疫应答的补体成分。在某些实施方案中,配体/fc融合蛋白提高包含本文中提供的结合或特异性结合fzd7之crd的非天然发生肽的配体的治疗价值。在某些实施方案中,包含本文中提供的结合或特异性结合fzd7之crd的非天然发生肽的配体例如在n或c端处与补体蛋白质,诸如ciq融合(诸如重组融合)。多份出版物描述用于获得其半衰期改变的非天然发生蛋白质的方法,这通过将fcrn结合多肽引入分子(wo1997/43316,us5869046,us5747035,wo1996/32478,wo1991/14438)或通过将蛋白质与其fcrn结合亲和力保留但对其它fc受体的亲和力大大降低的抗体融合(wo1999/43713)或与抗体的fcrn结合域融合(wo2000/09560,us4703039)。延长生理学活性分子(例如包含本文中提供的结合或特异性结合fzd7之crd的非天然发生肽的配体)的半衰期的具体技术和方法还可以在us7083784中找到。在某些实施方案中,包含本文中提供的结合或特异性结合fzd7之crd的非天然发生肽的配体与包含氨基酸残基突变(如通过kabat中的eu索引编号的):m252y/s254t/t256e或h433k/n434f/y436h的来自igg的fc区融合。在某些实施方案中,配体包含与增加或延长体内或血清半衰期的分子融合的本文中提供的结合或特异性结合fzd7之crd的非天然发生肽。在某些实施方案中,配体包含与清蛋白,诸如人血清清蛋白(hsa),聚乙二醇(peg),多糖,免疫球蛋白分子(igg),补体,血红蛋白,结合肽,脂蛋白或增加它在血流中的半衰期和/或它的组织穿透的其它因子融合的本文中提供的结合或特异性结合fzd7之crd的非天然发生肽。可以经由基因改组,基序改组,外显子改组,和/或密码子改组(统称“dna改组”)的技术来生成另外的包含本文中提供的结合或特异性结合fzd7之crd的非天然发生肽的配体。可以采用dna改组来改变包含(诸如本质上由其组成或由其组成)本文中提供的结合或特异性结合fzd7之crd的非天然发生肽(例如具有较高亲和力和更低解离速率的非天然发生肽)的配体的活性。一般参见us5605793,us5811238,us5830721,us5834252,us5837458,pattenetal.(1997)curr.opinionbiotechnol.8,724-33;harayama(1998)trendsbiotechnol.16,76-82;hanssonetal.(1999)j.mol.biol.287,265-76;和lorenzoandblasco(1998)biotechniques24,308-313。在某些实施方案中,在重组前通过提交随机诱变(通过错误倾向pcr,随机核苷酸插入或其它方法)来改变包含(诸如本质上由其组成或由其组成)本文中提供的结合或特异性结合fzd7之crd的非天然发生肽的配体。在某些实施方案中,可以将编码包含(诸如本质上由其组成或由其组成)本文中提供的结合或特异性结合fzd7之crd的非天然发生肽的配体的多核苷酸的一个或多个部分与一种或多种异源分子的一种或多种构件,基序,部件,部分,域,片段,等重组。可以通过标准技术来生成包含(诸如本质上由其组成或由其组成)本文中提供的结合或特异性结合fzd7之crd的非天然发生肽的配体,例如通过自使用公众可得的基因序列构建的重组融合基因表达融合蛋白,或通过化学肽合成。在某些实施方案中,本文中提供的配体包含(诸如本质上由其组成或由其组成)在长度上介于5-45个氨基酸之间的非天然发生肽。在某些实施方案中,本文中提供的配体包含(诸如本质上由其组成或由其组成)在长度上介于7-30个氨基酸之间的非天然发生肽。在某些实施方案中,本文中提供的配体包含(诸如本质上由其组成或由其组成)在长度上介于10-20个氨基酸之间的非天然发生肽。在某些实施方案中,本文中提供的配体包含(诸如本质上由其组成或由其组成)在长度上介于11-14个氨基酸之间的非天然发生肽。在某些实施方案中,本文中提供的配体包含(诸如本质上由其组成或由其组成)在长度上小于45,40,35,30,25,20,15,10,9,8,7,6,或5个氨基酸的非天然发生肽。在某些实施方案中,配体包含包含(诸如本质上由其组成或由其组成)x1x2x3ddlx4x5wchvmy(seqidno:100)中所列氨基酸序列的非天然发生肽,其中x1-x3中每个是无氨基酸,任何氨基酸,或非天然氨基酸,且其中x4-x5是任何氨基酸或非天然氨基酸。在某些实施方案中,x1是l,x2是p,x3是s,x4是e,且x5是f。在某些实施方案中,非天然发生肽是环化的。在某些实施方案中,非天然发生肽是二聚化的。在某些实施方案中,配体包含包含(诸如本质上由其组成或由其组成)x1x2ddlx3x4wchvmy(seqidno:101)中所列氨基酸序列的非天然发生肽,其中x1-x2中每个是无氨基酸,任何氨基酸,或非天然氨基酸,且其中x3-x4是任何氨基酸或非天然氨基酸。在某些实施方案中,非天然发生肽是环化的。在某些实施方案中,非天然发生肽是二聚化的。在某些实施方案中,配体包含包含(诸如本质上由其组成或由其组成)x1ddlx2x3wchvmy(seqidno:102)中所列氨基酸序列的非天然发生肽,其中x1是无氨基酸,任何氨基酸,或非天然氨基酸,且其中x2-x3是任何氨基酸或非天然氨基酸。在某些实施方案中,x1是s,x2是e,且x3是f。在某些实施方案中,非天然发生肽是环化的。在某些实施方案中,非天然发生肽是二聚化的。在某些实施方案中,配体包含包含(诸如本质上由其组成或由其组成)ddlx1x2wchvmy(seqidno:103)中所列氨基酸序列的非天然发生肽,其中x1和x2中每个是任何氨基酸或非天然氨基酸。在某些实施方案中,非天然发生肽是环化的。在某些实施方案中,非天然发生肽是二聚化的。在某些实施方案中,配体本质上由包含(诸如本质上由其组成或由其组成)x1x2x3ddlx4x5wchvmy(seqidno:100)中所列氨基酸序列的非天然发生肽组成(诸如由其组成),其中x1-x3中每个是无氨基酸,任何氨基酸,或非天然氨基酸,且其中x4-x5是任何氨基酸或非天然氨基酸。在某些实施方案中,x1是l,x2是p,x3是s,x4是e,且x5是f。在某些实施方案中,非天然发生肽是环化的。在某些实施方案中,非天然发生肽是二聚化的。在某些实施方案中,配体本质上由包含(诸如本质上由其组成或由其组成)x1x2ddlx3x4wchvmy(seqidno:101)中所列氨基酸序列的非天然发生肽组成(诸如由其组成),其中x1-x2中每个是无氨基酸,任何氨基酸,或非天然氨基酸,且其中x3-x4是任何氨基酸或非天然氨基酸。在某些实施方案中,非天然发生肽是环化的。在某些实施方案中,非天然发生肽是二聚化的。在某些实施方案中,配体本质上由包含(诸如本质上由其组成或由其组成)x1ddlx2x3wchvmy(seqidno:102)中所列氨基酸序列的非天然发生肽组成(诸如由其组成),其中x1是无氨基酸,任何氨基酸,或非天然氨基酸,且其中x2-x3是任何氨基酸或非天然氨基酸。在某些实施方案中,x1是s,x2是e,且x3是f。在某些实施方案中,非天然发生肽是环化的。在某些实施方案中,非天然发生肽是二聚化的。在某些实施方案中,配体本质上由包含(诸如本质上由其组成或由其组成)ddlx1x2wchvmy(seqidno:103)中所列氨基酸序列的非天然发生肽组成(诸如由其组成),其中x1和x2中每个是任何氨基酸或非天然氨基酸。在某些实施方案中,非天然发生肽是环化的。在某些实施方案中,非天然发生肽是二聚化的。在某些实施方案中,配体本质上由包含(诸如本质上由其组成或由其组成)sddlefwchvxy(seqidno:114)中所列氨基酸序列的非天然发生肽组成(诸如由其组成),其中x是任何氨基酸,或非天然氨基酸。在某些实施方案中,x是2-氨基癸酸。在某些实施方案中,x是l-2-氨基十六烷酸。在某些实施方案中,x是赖氨酸的包含与赖氨酸厄普西隆氨基基团偶联的辛酸的衍生物。在某些实施方案中,x是赖氨酸的包含与赖氨酸厄普西隆氨基基团偶联的癸酸的衍生物。在某些实施方案中,x是赖氨酸的包含与赖氨酸厄普西隆氨基基团偶联的十二烷酸的衍生物。在某些实施方案中,x是赖氨酸的包含与赖氨酸厄普西隆氨基基团偶联的十四烷酸的衍生物。在某些实施方案中,x是赖氨酸的包含与赖氨酸厄普西隆氨基基团偶联的十六烷酸的衍生物。在某些实施方案中,x是2-氨基-6-羟基己酸。在某些实施方案中,x是氧代己酸叔丁氧基。在某些实施方案中,x是s-苄基-l-高半胱氨酸(即经由硫醚键与苄基基团偶联的高半胱氨酸)。在某些实施方案中,x是由cas#374899-60-2代表的非天然氨基酸。在某些实施方案中,x是2-氨基-3-癸氧基-丙酸。在某些实施方案中,x是l-高苯丙氨酸。在某些实施方案中,x是2-氨基苯基戊酸。在某些实施方案中,x是l-α-氨基己二酸δ-叔丁酯。在某些实施方案中,x是由cas#:159751-47-0代表的非天然氨基酸。在某些实施方案中,x是丁酰基赖氨酸。在某些实施方案中,x是戊基赖氨酸。在某些实施方案中,x是氨基-8-(苄基氧基)-8-氧代辛酸。在某些实施方案中,x是由cas#:182059-59-2代表的非天然氨基酸。在某些实施方案中,x是l-2-氨基庚酸。在某些实施方案中,x是l-3-苯乙烯基丙氨酸。在某些实施方案中,x是6-羟基-l-正亮氨酸。在某些实施方案中,非天然发生肽是环化的。在某些实施方案中,非天然发生肽是二聚化的。在某些实施方案中,配体本质上由包含(诸如本质上由其组成或由其组成)sddxefwchvmy(seqidno:115)中所列氨基酸序列的非天然发生肽组成(诸如由其组成),其中x是任何氨基酸,或非天然氨基酸。在某些实施方案中,x是赖氨酸的包含与赖氨酸厄普西隆氨基基团偶联的辛酸的衍生物。在某些实施方案中,x是赖氨酸的包含与赖氨酸厄普西隆氨基基团偶联的癸酸的衍生物。在某些实施方案中,x是赖氨酸的包含与赖氨酸厄普西隆氨基基团偶联的十二烷酸的衍生物。在某些实施方案中,x是高苯丙氨酸。在某些实施方案中,x是l-高亮氨酸。在某些实施方案中,x是由cas#180414-94-2代表的非天然氨基酸。在某些实施方案中,x是叔丁基丙氨酸。在某些实施方案中,x是环丁基丙氨酸。在某些实施方案中,x是环戊基l-丙氨酸。在某些实施方案中,x是3-苯乙烯基苯丙氨酸。在某些实施方案中,x是联苯基。在某些实施方案中,x是l-2-氨基庚酸。在某些实施方案中,x是l-2-氨基辛酸。在某些实施方案中,x是l-2-氨基癸酸。在某些实施方案中,x是3-喹啉基-l-丙氨酸。在某些实施方案中,x是4-喹啉基-l-丙氨酸。在某些实施方案中,x是三氟甲基-l-亮氨酸。在某些实施方案中,x是环己基-l-丙氨酸。在某些实施方案中,x是f(苯丙氨酸)。在某些实施方案中,非天然发生肽是环化的。在某些实施方案中,非天然发生肽是二聚化的。在某些实施方案中,配体本质上由包含(诸如本质上由其组成或由其组成)sddx1efwchvx2y(seqidno:116)中所列氨基酸序列的非天然发生肽组成(诸如由其组成),其中x1和x2中每个是任何氨基酸或非天然氨基酸。在某些实施方案中,x1是l-高苯丙氨酸,且x2是赖氨酸的包含与赖氨酸厄普西隆氨基基团偶联的十四烷酸的衍生物。在某些实施方案中,非天然发生肽是环化的。在某些实施方案中,非天然发生肽是二聚化的。在某些实施方案中,配体本质上由包含(诸如本质上由其组成或由其组成)sddlefwchxmy(seqidno:117)中所列氨基酸序列的非天然发生肽组成(诸如由其组成),其中x是任何氨基酸,或非天然氨基酸。在某些实施方案中,x是l。在某些实施方案中,x是i。在某些实施方案中,x是t。在某些实施方案中,x是3-氨基-l-丙氨酸。在某些实施方案中,x是由cas#:181954-34-7代表的非天然氨基酸。在某些实施方案中,x是β-羟基正缬氨酸。在某些实施方案中,x是叔丁基丙氨酸。在某些实施方案中,x是叔丁基-l-丙氨酸。在某些实施方案中,x是环丁基-l-甘氨酸。在某些实施方案中,x是环丙基-l-丙氨酸。在某些实施方案中,x是环戊基-l-丙氨酸。在某些实施方案中,非天然发生肽是环化的。在某些实施方案中,非天然发生肽是二聚化的。在某些实施方案中,配体本质上由包含(诸如本质上由其组成或由其组成)lpsddlefwchvmx(seqidno:118)中所列氨基酸序列的非天然发生肽组成(诸如由其组成),其中x是任何氨基酸,或非天然氨基酸。在某些实施方案中,x是4-(三氟甲基)-l-苯丙氨酸。在某些实施方案中,x是4-氯-l-苯丙氨酸。在某些实施方案中,x是4-甲基-l-苯丙氨酸。在某些实施方案中,x是3-(3-喹啉基)-l-丙氨酸。在某些实施方案中,x是3-(2-喹啉基)-l-丙氨酸。在某些实施方案中,x是3-(2-喹噁啉基)-l-丙氨酸。在某些实施方案中,x是3-[3,4-二(三氟甲基)苯基]-l-丙氨酸。在某些实施方案中,x是3,4-二氟-l-苯丙氨酸。在某些实施方案中,非天然发生肽是环化的。在某些实施方案中,非天然发生肽是二聚化的。在某些实施方案中,配体本质上由包含(诸如本质上由其组成或由其组成)sdxlefwchvmy(seqidno:119)中所列氨基酸序列的非天然发生肽组成(诸如由其组成),其中x是任何氨基酸,或非天然氨基酸。在某些实施方案中,x是e(谷氨酸)。在某些实施方案中,x是l-α-氨基己二酸。在某些实施方案中,x是由cas#250384-77-1代表的非天然氨基酸。在某些实施方案中,非天然发生肽是环化的。在某些实施方案中,非天然发生肽是二聚化的。在某些实施方案中,配体本质上由包含(诸如本质上由其组成或由其组成)lpsdxlefwchvmy(seqidno:120)中所列氨基酸序列的非天然发生肽组成(诸如由其组成),其中x是任何氨基酸,或非天然氨基酸。在某些实施方案中,x是q(谷氨酰胺)。在某些实施方案中,非天然发生肽是环化的。在某些实施方案中,非天然发生肽是二聚化的。在某些实施方案中,配体本质上由包含(诸如本质上由其组成或由其组成)sddlexwchvmy(seqidno:121)中所列氨基酸序列的非天然发生肽组成(诸如由其组成),其中x是任何氨基酸,或非天然氨基酸。在某些实施方案中,x是4-氯-l-苯丙氨酸。在某些实施方案中,非天然发生肽是环化的。在某些实施方案中,非天然发生肽是二聚化的。在某些实施方案中,配体本质上由包含(诸如本质上由其组成或由其组成)xddlefwchvmy(seqidno:122)中所列氨基酸序列的非天然发生肽组成(诸如由其组成),其中x是任何氨基酸,或非天然氨基酸。在某些实施方案中,x是t(苏氨酸)。在某些实施方案中,x是β-羟基正缬氨酸。在某些实施方案中,非天然发生肽是环化的。在某些实施方案中,非天然发生肽是二聚化的。在某些实施方案中,配体本质上由包含(诸如本质上由其组成或由其组成)lpsddlefwxhvmy(seqidno:123)中所列氨基酸序列的非天然发生肽组成(诸如由其组成),其中x是任何氨基酸,或非天然氨基酸。在某些实施方案中,x是a(丙氨酸)。在某些实施方案中,非天然发生肽是环化的。在某些实施方案中,非天然发生肽是二聚化的。在某些实施方案中,本文中提供的配体包含包含氨基酸序列lpsddlefwchvmy(seqidno:13)的非天然发生肽。在某些实施方案中,本文中提供的配体包含由氨基酸序列lpsddlefwchvmy(seqidno:13)组成的肽。在某些实施方案中,本文中提供的配体由包含氨基酸序列lpsddlefwchvmy(seqidno:13)的非天然发生肽组成。在某些实施方案中,本文中提供的配体由由氨基酸序列lpsddlefwchvmy(seqidno:13)组成的非天然发生肽组成。在某些实施方案中,肽是以使它的α螺旋结构成核或稳定的方式经过修饰的,如例如mahonetal.(2012)drugdiscoverytoday:tech.9:e57-e62;foroodetal.(1993)procnatlacadsciu.s.a.90:838-842;klein(2014)medchemlett.5:838-839;和其中引用的参考文献中描述的。在某些实施方案中,肽是订合(stapled)肽。在某些实施方案中,肽是环化的。在某些实施方案中,肽是二聚化的。在某些实施方案中,本文中提供的配体包含包含氨基酸序列sddlefwchvmy(seqidno:99)的非天然发生肽。在某些实施方案中,本文中提供的配体包含由氨基酸序列sddlefwchvmy(seqidno:99)组成的非天然发生肽。在某些实施方案中,本文中提供的配体是包含氨基酸序列sddlefwchvmy(seqidno:99)的非天然发生肽。在某些实施方案中,本文中提供的配体是由氨基酸序列sddlefwchvmy(seqidno:99)组成的非天然发生肽。在某些实施方案中,肽是以使它的α螺旋结构成核或稳定的方式经过修饰的,如例如mahonetal.(2012)drugdiscoverytoday:tech.9:e57-e62;foroodetal.(1993)procnatlacadsciu.s.a.90:838-842;klein(2014)medchemlett.5:838-839;和其中引用的参考文献中描述的。在某些实施方案中,肽是订合肽。在某些实施方案中,肽是环化的。在某些实施方案中,肽是二聚化的。在某些实施方案中,配体包含(诸如本质上由其组成或由其组成)非天然发生肽,它是lpsddlefwchvmy(seqidno:13)的变体。在某些实施方案中,此类肽变体包含seqidno:13中的至少1,至少2,至少3,至少4,或至少5处氨基酸替代。在某些实施方案中,seqidno:13中的位置1,2,3,7,和/或8处的氨基酸是替代的。在某些实施方案中,配体包含(诸如本质上由其组成或由其组成)非天然发生肽,它是sddlefwchvmy(seqidno:99)的变体。在某些实施方案中,此类变体包含seqidno:99中的至少1,至少2,或至少3处氨基酸替代。在某些实施方案中,seqidno:99中的位置1,5,和/或6处的氨基酸是替代的。在某些实施方案中,氨基酸替代是保守氨基酸替代。在某些实施方案中,氨基酸替代是用非天然氨基酸的替代。在某些实施方案中,氨基酸替代并不实质性降低非天然发生肽结合fzd7之crd的能力。例如,可以进行并不实质性降低fzd7crd结合亲和力的保守改变(例如如本文中描述的保守替代)。可以使用下文实施例中描述的方法来评估包含(诸如本质上由其组成或由其组成)seqidno:13或seqidno:99的变体的配体的结合亲和力。保守替代在下文表1中在标题“保守替代”下显示。更加实质性变化在表1中在标题“例示性替代”下提供且如下文提及氨基酸侧链类别进一步描述的。可以将氨基酸替代引入seqidno:13或seqidno:99的变体,并使用实施例中描述的方法对产物筛选期望活性,例如保留/改善对fzd7之crd的结合。表1:保守替代非保守替代会需要用这些类别之一的成员交换另一类别。seqidno:13或seqidno:99的一种例示性替代变体是亲和力成熟肽,可以例如使用基于噬菌体展示的亲和力成熟技术,诸如实施例中描述的那些方便地生成。简言之,改变(即添加,删除,或替代)本文中描述的肽中的一个或多个残基并在噬菌体上展示变体肽并筛选fzd7crd结合亲和力。在亲和力成熟的某些实施方案中,通过多种方法任一(例如错误倾向pcr或寡核苷酸定向诱变)将多样性导入为成熟选择的可变肽。然后创建次级文库。然后筛选文库以鉴定具有对fzd7crd的期望亲和力的任何肽变体。在某些实施方案中,引入多样性牵涉随机化本文中描述的肽中的一个或多个残基。在某些实施方案中,可以例如使用丙氨酸扫描诱变,丝氨酸扫描诱变,缬氨酸扫描诱变,天冬氨酸扫描诱变或建模来鉴定本文中描述的肽中牵涉对fzd7之crd的结合的残基。在一些实施方案中,配体包含(诸如本质上由其组成或由其组成)包含(诸如本质上由其组成或由其组成)lpsddaefwchvmy(seqidno:109),lpsddlefachvmy(seqidno:110),lpsddlefwchvay(seqidno:111),lpsddlefwchvma(seqidno:112),lpsdqlefwchvmy(seqidno:131),或lpsddlefwahvmy(seqidno:133)中所列氨基酸序列的非天然发生肽。在某些实施方案中,肽的c端羧基基团是酰胺化的。在某些实施方案中,肽的n端胺是乙酰化的。在某些实施方案中,肽的c端羧基基团是酰胺化的,且肽的n端胺是乙酰化的。在某些实施方案中,非天然发生肽是环化的。在某些实施方案中,非天然发生肽是二聚化的。在某些实施方案中,二聚体中的肽是通过例如肽中的c10之间的二硫键连接的。在某些实施方案中,二聚体中的肽是通过化学接头,诸如本文中别处描述的化学接头连接的。在某些实施方案中,二聚体中的肽是与间隔肽(例如居间肽)融合的。在一些实施方案中,配体包含(诸如由其组成或本质上由其组成)包含(诸如本质上由其组成或由其组成)sddlefwchvey(seqidno:125),sddlefwchlmy(seqidno:126),sddlefwchimy(seqidno:127),sddlefwchtmy(seqidno:128),sddfefwchvmy(seqidno:129),sdelefwchvmy(seqidno:130),或tddlefwchvmy(seqidno:132)中所列氨基酸序列的非天然发生肽。在某些实施方案中,肽的c端羧基基团是酰胺化的。在某些实施方案中,肽的n端胺是乙酰化的。在某些实施方案中,肽的c端羧基基团是酰胺化的,且肽的n端胺是乙酰化的。在某些实施方案中,非天然发生肽是环化的。在某些实施方案中,非天然发生肽是二聚化的。在某些实施方案中,二聚体中的肽是通过例如肽中的c8之间的二硫键连接的。在某些实施方案中,二聚体中的肽是通过化学接头,诸如本文中别处描述的化学接头连接的。在某些实施方案中,二聚体中的肽是与间隔肽(例如居间肽)融合的。另外/或者,在某些实施方案中,配体包含(诸如本质上由其组成或由其组成)肽,它是lpsddlefwchvmy(seqidno:13)的变体或sddlefwchvmy(seqidno:99)的变体,其中一个或多个氨基酸是用非天然氨基酸替代的。另外/或者,在某些实施方案中,配体包含(诸如本质上由其组成或由其组成)肽,它是lpsddlefwchvmy(seqidno:13)的变体或sddlefwchvmy(seqidno:99)的变体,其中添加(例如在n端,在c端,或在n和c端二者)一个或多个非天然氨基酸残基。在某些实施方案中,非天然氨基酸是本文中别处描述的非天然氨基酸。在某些实施方案中,配体包含(诸如本质上由其组成或由其组成)肽,它是lpsddlefwchvmy(seqidno:13)的变体或sddlefwchvmy(seqidno:99)的变体,其中肽的c端羧基基团是酰胺化的。在某些实施方案中,配体包含(诸如本质上由其组成或由其组成)肽,它是lpsddlefwchvmy(seqidno:13)的变体或sddlefwchvmy(seqidno:99)的变体,其中肽的n端胺是乙酰化的。在某些实施方案中,配体包含(诸如本质上由其组成或由其组成)肽,它是lpsddlefwchvmy(seqidno:13)的变体或sddlefwchvmy(seqidno:99)的变体,其中肽的c端羧基基团是酰胺化的且其中肽的n端胺是乙酰化的。在某些实施方案中,本文中提供的配体包含(诸如本质上由其组成或由其组成)二聚化的特异性结合fzd7之crd的非天然发生肽。在某些实施方案中,二聚体中的每个肽包含(诸如本质上由其组成或由其组成)seqidno:13中所列氨基酸。在某些实施方案中,二聚体中的肽是通过例如seqidno:13的c10之间的二硫键连接的。在某些实施方案中,二聚体中的每个肽包含(诸如本质上由其组成或由其组成)seqidno:99中所列氨基酸。在某些实施方案中,二聚体中的肽是通过例如seqidno:99的c8之间的二硫键连接的。在某些实施方案中,二聚体中的肽是通过化学接头,诸如本文中别处描述的化学接头连接的。在某些实施方案中,二聚体中的肽是与间隔肽(例如居间肽)融合的。在某些实施方案中,配体包含(诸如本质上由其组成或由其组成)两个非天然发生肽。在某些实施方案中,第一非天然发生肽包含(诸如本质上由其组成或由其组成)sddlefwchvmyx(seqidno:134)中所列氨基酸序列,其中x是l-高炔丙基甘氨酸,且第二非天然发生肽包含(诸如本质上由其组成或由其组成)sddlxfwchvmy(seqidno:135)中所列氨基酸序列,其中x是5-叠氮基-l-鸟氨酸或s-乙酰氨基甲基-l-半胱氨酸。在某些实施方案中,第一和第二非天然发生肽是通过经由点击化学使非天然氨基酸反应而共价连接的。在某些实施方案中,第一非天然发生肽包含(诸如本质上由其组成或由其组成)sddlefwchvmyx(seqidno:134)中所列氨基酸序列,其中x是l-二高炔丙基甘氨酸,且第二非天然发生肽包含(诸如本质上由其组成或由其组成)sddlxfwchvmy(seqidno:135)中所列氨基酸序列,其中x是叠氮基-高丙氨酸或s-乙酰氨基甲基-l-半胱氨酸。在某些实施方案中,第一和第二非天然发生肽是通过经由点击化学使非天然氨基酸反应而共价连接的。在某些实施方案中,配体包含(诸如本质上由其组成或由其组成)包含(诸如本质上由其组成或由其组成)sddlefwxhvmy(seqidno:124)中所列氨基酸序列的非天然发生肽,其中x是l-硒代半胱氨酸。在某些实施方案中,肽是环化的和/或经由l-硒代半胱氨酸二聚化的。在某些实施方案中,本文中提供的配体包含包含seqidno:1-12,14-31,和39-98任一中所列氨基酸序列的非天然发生肽(诸如线性肽)。在某些实施方案中,本文中提供的配体包含由seqidno:1-12,14-31,和39-98任一中所列氨基酸序列组成的非天然发生肽(诸如线性肽)。在某些实施方案中,本文中提供的配体是包含seqidno:1-12,14-31,和39-98任一中所列氨基酸序列的非天然发生肽(诸如线性肽)。在某些实施方案中,本文中提供的配体是由seqidno:1-12,14-31,和39-98任一中所列氨基酸序列组成的非天然发生肽(诸如线性肽)。在某些实施方案中,本文中提供的配体包含包含seqidno:32-38任一中所列氨基酸序列的非天然发生(诸如环状肽)。在某些实施方案中,本文中提供的配体包含由seqidno:32-38任一中所列氨基酸序列组成的非天然发生肽(诸如环状肽)。在某些实施方案中,本文中提供的配体是包含seqidno:32-38任一中所列氨基酸序列的非天然发生肽(诸如环状肽)。在某些实施方案中,本文中提供的配体是由seqidno:32-38任一中所列氨基酸序列组成的非天然发生肽(诸如环状肽)。seqidno:1-12和14-98的氨基酸序列在下文表2中提供:表2在某些实施方案中,本文中提供的配体包含(诸如本质上由其组成或由其组成)与脂质缀合的本文中描述的结合或特异性结合fzd7之crd的非天然发生肽。在某些实施方案中,脂质是包含介于12-20个碳原子之间的长链脂肪酸(即lcfa)。在某些实施方案中,脂质是包含6个或更少碳原子的短链脂肪酸(即scfa)。在某些实施方案中,脂质是饱和脂肪酸。在某些实施方案中,脂质是不饱和脂肪酸。在某些实施方案中,脂质包含芳香族尾。在某些实施方案中,脂质是辛酸。在某些实施方案中,脂质是癸酸。在某些实施方案中,脂质是十二烷酸。在某些实施方案中,脂质是十四烷酸。在某些实施方案中,脂质是十六烷酸。在某些实施方案中,脂质是氨基-6-羟基己酸。在某些实施方案中,脂质是氨基-6-羟基己酸。在某些实施方案中,脂质是氨基苯基戊酸。在某些实施方案中,脂质是l-α-氨基己二酸δ-叔丁酯。在某些实施方案中,脂质是氨基-8-(苄基氧基)-8-氧代辛酸。在某些实施方案中,脂质是2-氨基庚酸。在某些实施方案中,脂质是l-2-氨基癸酸。在某些实施方案中,脂质是2-氨基辛酸。将脂质与肽偶联的方法是本领域公知的且典型地需要在标准酰胺偶联条件下使脂质的羧酸基团与肽中的赖氨酸侧链的厄普西隆胺反应。另外的方法在gerauer,m.,koch,s.,brunsveld,l.andwaldmann,h.2008.lipidatedpeptidesynthesis.wileyencyclopediaofchemicalbiology.1-11中综述。在某些实施方案中,如本文中别处描述的,使用标准肽合成技术将脂质偶联的氨基酸掺入肽。在某些实施方案中,本文中提供的配体包含(诸如本质上由其组成或由其组成)本文中描述的非天然发生肽,其中肽的c端羧基基团是酰胺化的。在某些实施方案中,本文中提供的配体包含(诸如本质上由其组成或由其组成)本文中描述的非天然发生肽,其中肽的n端胺是乙酰化的。在某些实施方案中,本文中提供的配体包含(诸如本质上由其组成或由其组成)本文中描述的非天然发生肽,其中肽的c端羧基基团是酰胺化的且其中肽的n端胺是乙酰化的。在某些实施方案中,本文中提供的配体包含(诸如本质上由其组成或由其组成)与核酸分子,小分子,模拟剂,合成药物,无机分子,和有机分子缀合(诸如共价或非共价)的本文中描述的结合或特异性结合fzd7之crd的非天然发生肽。在某些实施方案中,本文中提供的配体包含(诸如本质上由其组成或由其组成)与异源蛋白质或多肽(或其片段,与至少10,至少20,至少30,至少40,至少50,至少60,至少70,至少80,至少90或至少100个氨基酸的多肽)缀合(诸如共价或非共价)的本文中描述的结合或特异性结合fzd7之crd的非天然发生肽。在某些实施方案中,本文中提供的配体包含(诸如本质上由其组成或由其组成)与细胞毒剂,诸如化疗剂,毒素(例如细菌,真菌,植物,或动物起源的酶活性毒素,或其片段),或放射性同位素(即放射缀合物)缀合的本文中描述的结合或特异性结合fzd7之crd的非天然发生肽。可使用的酶活性毒素和其片段包括白喉a链,白喉毒素的非结合活性片段,外毒素a链(来自铜绿假单胞菌(pseudomonasaeruginosa)),蓖麻毒蛋白(ricin)a链,相思豆毒蛋白(abrin)a链,蒴莲根毒蛋白(modeccin)a链,α-帚曲霉素(sarcin),油桐(aleuritesfordii)毒蛋白,香石竹(dianthin)毒蛋白,美洲商陆(phytolacaamericana)毒蛋白(papi,papii,和pap-s),苦瓜(momordicacharantia)抑制物,麻疯树毒蛋白(curcin),巴豆毒蛋白(crotin),肥皂草(saponariaofficinalis)抑制物,白树毒蛋白(gelonin),丝林霉素(mitogellin),局限曲菌素(restrictocin),酚霉素(phenomycin),依诺霉素(enomycin),和单端孢菌素(tricothecene)。其它毒素包括美登木素和美登木生物碱,加利车霉素和其它细胞毒剂。多种放射性核素可用于生成放射缀合的包含(诸如本质上由其组成或由其组成)本文中提供的特异性结合fzd1,fzd2,和/或fzd7之crd的非天然发生肽的配体。例子包括212bi,131i,131in,90y,和186re。使用多种双功能蛋白质偶联剂来生成包含(诸如本质上由其组成或由其组成)本文中提供的结合或特异性结合fzd7之crd的非天然发生肽的配体和例如细胞毒剂的缀合物,双功能蛋白质偶联剂诸如n-琥珀酰亚胺基-3-(2-吡啶基二硫代)丙酸酯(spdp),亚氨基硫烷(it),亚氨酸酯(诸如盐酸己二酰亚氨酸二甲酯),活性酯(诸如辛二酸二琥珀酰亚胺基酯),醛(诸如戊二醛),双叠氮化合物(诸如双(对叠氮苯甲酰基)己二胺),双重氮衍生物(诸如双(对重氮苯甲酰基)-乙二胺),二异氰酸酯(诸如甲苯2,6-二异氰酸酯),和双活性氟化合物(诸如1,5-二氟-2,4-二硝基苯)的双功能衍生物。例如,蓖麻毒蛋白免疫毒素可以如vitettaetal.,science,238:1098(1987)中所述来制备。碳-14标记的1-异硫氰酸基苄基-3-甲基二亚乙基三胺五乙酸(mx-dtpa)是用于将放射性核素与包含(诸如本质上由其组成或由其组成)本文中提供的结合或特异性结合fzd7之crd的非天然发生肽的配体缀合的一种例示性螯合剂。参见wo94/11026。在某些实施方案中,工程化改造包含(诸如本质上由其组成或由其组成)本文中提供的结合或特异性结合fzd7之crd的非天然发生肽的配体以提供用于缀合的反应性基团。在某些实施方案中,n端和/或c端也可用来提供用于缀合的反应性基团。在某些实施方案中,将n端与一种模块(诸如但不限于peg)缀合而将c端与另一种模块(诸如但不限于生物素)缀合,或反之亦然。在某些实施方案中,可以将包含(诸如本质上由其组成或由其组成)本文中提供的结合或特异性结合fzd7之crd的非天然发生肽的配体与诊断剂或可检测剂缀合。此类配体缀合物对于疾病的发生或进展的监测或预后作为临床测试规程的一部分,诸如测定特定疗法的功效可能是有用的。此类诊断和检测可以通过将包含(诸如本质上由其组成或由其组成)本文中提供的结合或特异性结合fzd7之crd的非天然发生肽的配体与可检测物质偶联来实现,可检测物质包括但不限于各种酶,诸如但不限于辣根过氧化物酶,碱性磷酸酶,β-半乳糖苷酶,或乙酰胆碱酯酶;辅基,诸如但不限于链霉亲合素/生物素和亲合素/生物素;荧光材料,诸如但不限于伞形酮,荧光素,异硫氰酸荧光素,罗丹明,二氯三嗪基胺荧光素,丹酰氯或藻红蛋白;发光材料,诸如但不限于鲁米诺;生物发光材料,诸如但不限于萤光素酶,萤光素,和水母发光蛋白;放射性材料,诸如但不限于碘(131i,125i,123i,121i),碳(14c),硫(35s),氚(3h),铟(115in,113in,112in,111in),和锝(99tc),铊(201ti),镓(68ga,67ga),钯(103pd),钼(99mo),氙(133xe),氟(18f),153sm,177lu,159gd,149pm,140la,175yb,166ho,90y,47sc,186re,188re,142pr,105rh,97ru,68ge,57co,65zn,85sr,32p,153gd,169yb,51cr,54mn,75se,113sn,和117tn;正电子发射金属,使用各种正电子发射断层照相术,非放射性顺磁性金属离子,和放射性标记的或与特定放射性同位素缀合的分子。还提供的是与治疗性模块缀合的包含(诸如本质上由其组成或由其组成)本文中提供的结合或特异性结合fzd7之crd的非天然发生肽的配体。在某些实施方案中,可以将包含(诸如本质上由其组成或由其组成)本文中提供的结合或特异性结合fzd7之crd的非天然发生肽的配体与治疗性模块,诸如细胞毒素,例如抑制细胞剂或杀细胞剂,治疗剂或放射性金属离子,例如α-发射体缀合。细胞毒素或细胞毒剂包括任何对细胞有害的药剂。在某些实施方案中,将包含(诸如本质上由其组成或由其组成)本文中提供的结合或特异性结合fzd7之crd的非天然发生肽的配体与治疗性模块,诸如放射性金属离子,诸如α-发射体,诸如213bi或对于将放射性金属离子,包括但不限于131in,131lu,131y,131ho,131sm与多肽缀合有用的大环螯合剂缀合。在某些实施方案中,大环螯合剂是能经由接头分子附着于包含(诸如本质上由其组成或由其组成)本文中提供的结合或特异性结合fzd7之crd的非天然发生肽的配体的1,4,7,10-四氮杂环十二烷-n,n',n",n"'-四乙酸(dota)。此类接头分子是本领域普遍知道的且描述于例如denardoetal.(1998)clincancerres.4,2483-90;petersonetal.(1999)bioconjug.chem.10,553-557;和zimmermanetal.(1999)nucl.med.biol.26,943-50。用于将治疗性模块与抗体缀合的技术是公知的且可以应用于包含(诸如本质上由其组成或由其组成)本文中提供的结合或特异性结合fzd7之crd的非天然发生肽的配体(参见例如amonetal.,"monoclonalantibodiesforimmunotargetingofdrugsincancertherapy,”inmonoclonalantibodiesandcancertherapy,reisfeldetal.(eds.),pp.243-56(alanr.liss,inc.1985);hellstrometal.,"antibodiesfordrugdelivery",incontrolleddrugdelivery(2nded.),robinsonetal.(eds.),pp.623-53(marceldekker,inc.1987);thorpe,"antibodycarriersofcytotoxicagentsincancertherapy:areview",inmonoclonalantibodies84:biologicalandclinicalapplications,pincheraetal.(eds.),pp.475-506(1985);"analysis,results,andfutureprospectiveofthetherapeuticuseofradiolabeledantibodyincancertherapy",inmonoclonalantibodiesforcancerdetectionandtherapy,baldwinetal.(eds.),pp.303-16(academicpress1985);和thorpeetal.,1982,immunol.rev.62:119-58)。应当选择与包含(诸如本质上由其组成或由其组成)本文中提供的结合或特异性结合fzd7之crd的非天然发生肽的配体缀合的治疗性模块或药物来实现对受试者中的特定病症的期望预防或治疗效果。临床医生或其他医务人员在决定将哪种治疗性模块或药物与配体缀合时应当考虑下述:疾病的性质,疾病的严重程度,和受试者的状况。在某些实施方案中,还可以将包含(诸如本质上由其组成或由其组成)本文中提供的结合或特异性结合fzd7之crd的非天然发生肽的配体附着于对于靶抗原的纯化或免疫测定法特别有用的固体支持物。此类固体支持物包括但不限于玻璃,纤维素,聚丙烯酰胺,尼龙,聚苯乙烯,聚氯乙烯或聚丙烯。还涵盖编码包含(诸如本质上由其组成或由其组成)本文中提供的结合或特异性结合fzd7之crd的非天然发生肽的配体的核酸分子,包含编码配体的核酸分子的表达载体,和包含核酸分子的细胞。本文中还提供的是生成包含(诸如本质上由其组成或由其组成)本文中提供的结合或特异性结合fzd7之crd的非天然发生肽的配体的方法,其通过培养此类细胞,表达配体,和自细胞培养物回收配体。在某些实施方案中,如本文中别处描述的,经由体外翻译生成包含(诸如本质上由其组成或由其组成)本文中提供的结合或特异性结合fzd7之crd的非天然发生肽的配体。在某些实施方案中,经由化学肽合成生成包含(诸如本质上由其组成或由其组成)本文中提供的结合或特异性结合fzd7之crd的非天然发生肽的配体,例如通过将化学合成的本文中描述的非天然发生肽与一种或多种模块嫁接,或通过化学合成整个配体。在某些实施方案中,在其中牵涉异常wnt信号传导的疾病或状况的治疗中作为治疗剂使用包含(诸如本质上由其组成或由其组成)本文中提供的结合或特异性结合fzd7之crd的非天然发生肽的配体。在某些实施方案中,提供的是一种杀伤癌细胞(例如结肠癌细胞,胰腺癌细胞,非小细胞肺癌细胞,包含rnf43突变的癌细胞,特征在于usp6过表达的癌症,或特征在于牵涉r-spondin(rspo)家族成员的基因融合的癌细胞)的方法,其包含使癌细胞与包含(诸如本质上由其组成或由其组成)本文中提供的结合或特异性结合fzd7之crd域的非天然发生肽的配体接触。在某些实施方案中,提供的是一种杀伤癌干细胞(例如结肠癌干细胞,胰腺癌干细胞,非小细胞肺癌干细胞,包含rnf43突变的癌干细胞,特征在于usp6过表达的癌干细胞,或特征在于牵涉r-spondin(rspo)家族成员的基因融合的癌干细胞)的方法,其包含使癌细胞与包含(诸如本质上由其组成或由其组成)本文中提供的结合或特异性结合fzd7之crd域的非天然发生肽的配体接触。在某些实施方案中,提供的是一种抑制细胞(诸如结肠癌细胞,胰腺癌细胞,非小细胞肺癌细胞,包含rnf43突变的癌细胞,特征在于usp6过表达的癌细胞,或特征在于牵涉r-spondin(rspo)家族成员的基因融合的癌细胞)中的wnt介导的β-联蛋白信号传导的方法,其包含使癌细胞与包含(诸如本质上由其组成或由其组成)本文中提供的结合或特异性结合fzd7之crd域的非天然发生肽的配体接触。功能特征在某些实施方案中,包含(诸如本质上由其组成或由其组成)本文中提供的结合或特异性结合fzd7之crd的非天然发生肽的配体具有不超过约1x10-7m,优选不超过约1x10-8和最优选不超过约1x10-9m的结合亲和力(kd)值但具有比它对fzd7之crd的结合亲和力要弱至少约50倍,或至少约500倍,或至少约1000倍的对fzd3,fzd4,fzd5,fzd6,fzd8,fzd9,和/或fzd10之crd的结合亲和力。在某些实施方案中,包含(诸如本质上由其组成或由其组成)本文中提供的结合或特异性结合fzd7之crd的非天然发生肽的配体具有不超过约1x10-7m,优选不超过约1x10-8和最优选不超过约1x10-9m的结合亲和力(kd)值但具有比它对fzd7之crd的结合亲和力要弱至少约50倍,或至少约500倍,或至少约1000倍的对fzd1,fzd2,fzd3,fzd4,fzd5,fzd6,fzd8,fzd9,和/或fzd10之crd的结合亲和力。在某些实施方案中,包含(诸如本质上由其组成或由其组成)本文中提供的非天然发生肽的配体对例如非靶fzd受体(例如fzd1,fzd2,fzd3,fzd4,fzd5,fzd6,fzd8,fzd9,和/或fzd10)之crd的结合的程度小于包含(诸如本质上由其组成或由其组成)本文中提供的非天然发生肽的配体对fzd7之crd的结合的约10%,如通过本领域已知方法,诸如elisa,荧光激活细胞分选(facs)分析,或放射免疫沉淀(ria)测定的。在某些实施方案中,包含(诸如本质上由其组成或由其组成)本文中提供的非天然发生肽的配体对在结构上与fzd蛋白质有关的蛋白质(诸如分泌型卷曲蛋白相关蛋白,例如sfrp1,sfrp2,sfrp3,sfrp4,和/或sfrp5)的结合的程度小于包含(诸如本质上由其组成或由其组成)本文中提供的非天然发生肽的配体对fzd7之crd的结合的约10%,如通过本领域已知方法,诸如elisa,荧光激活细胞分选(facs)分析,或放射免疫沉淀(ria)测定的。特异性结合可以例如通过与对照分子(一般是具有相似结构且没有结合活性的分子)的结合相比测定分子的结合来测量。例如,特异性结合可以通过与同靶物相似的对照分子(例如过量的未标记的靶物)的竞争来确定。在这种情况中,如果经标记的靶物对探针的结合受到过量的未标记的靶物的竞争性抑制,那么指示特异性结合。实施例中描述了评估包含(诸如本质上由其组成或由其组成)本文中提供的“特异性结合”fzd7之crd的非天然发生肽的配体的结合的其它方法。如本文中使用的术语“特异性结合”特定多肽或特定多肽靶上的结合区或“对其特异性的”可以例如由具有至少约10-4m,或者至少约10-5m,或者至少约10-6m,或者至少约10-7m,或者至少约10-8m,或者至少约10-9m,或者至少约10-10m,或者至少约10-11m,或者至少约10-12m,或更大的对靶物的kd的分子展现。在一个实施方案中,术语“特异性结合”指如下的结合,其中分子结合特定多肽或特定多肽上的结合区且并不实质性结合任何其它多肽或结合区。在某些实施方案中,包含(诸如本质上由其组成或由其组成)本文中提供的非天然发生肽的配体以介于约1pm至约500nm之间的kd结合fzd7之crd。在某些实施方案中,包含(诸如本质上由其组成或由其组成)本文中提供的非天然发生肽的配体以介于约1pm至约50pm之间,介于约50pm至约250pm之间,介于约250pm至约500pm之间,介于约500pm至750pm之间,介于约750pm至约1nm之间,介于约1nm至约25nm之间,介于约25nm至约50nm之间,介于50nm至约100nm之间,介于约100nm至约250nm之间,或介于约250nm至约500nm之间的kd结合fzd7之crd。在某些实施方案中,kd是经由表面等离振子共振测定的。在某些实施方案中,包含(诸如本质上由其组成或由其组成)本文中提供的特异性结合fzd7之crd的非天然发生肽的配体以小于约300nm,275nm,250nm,200nm,175nm,150nm,140nm,130nm,120nm,110nm,100nm,90nm,80nm,70nm,60nm,50nm,40nm,30nm,20nm,10nm,5nm,1nm,0.9nm,0.8nm,0.7nm,0.6nm,0.5nm,0.4nm,0.3nm,0.2nm,或0.1nm中任一的ic50值抑制wnt介导的β-联蛋白信号传导,包括介于这些值之间的任何范围。在某些实施方案中,包含(诸如本质上由其组成或由其组成)本文中提供的特异性结合fzd7之crd的非天然发生肽的配体以介于10nm和200nm之间的ic50值抑制wnt介导的β-联蛋白信号传导。在某些实施方案中,包含(诸如本质上由其组成或由其组成)本文中提供的特异性结合fzd7之crd的非天然发生肽的配体以介于20nm和200nm之间的ic50值抑制wnt介导的β-联蛋白信号传导。在某些实施方案中,包含(诸如本质上由其组成或由其组成)本文中提供的特异性结合fzd7之crd的非天然发生肽的配体以介于30nm和200nm之间的ic50值抑制wnt介导的β-联蛋白信号传导。在某些实施方案中,包含(诸如本质上由其组成或由其组成)本文中提供的特异性结合fzd7之crd的非天然发生肽的配体以介于40nm和200nm之间的ic50值抑制wnt介导的β-联蛋白信号传导。在某些实施方案中,包含(诸如本质上由其组成或由其组成)本文中提供的特异性结合fzd7之crd的非天然发生肽的配体以介于50nm和200nm之间的ic50值抑制wnt介导的β-联蛋白信号传导。在某些实施方案中,包含(诸如本质上由其组成或由其组成)本文中提供的特异性结合fzd7之crd的非天然发生肽的配体以介于50nm和180nm之间的ic50值抑制wnt介导的β-联蛋白信号传导。在某些实施方案中,包含(诸如本质上由其组成或由其组成)本文中提供的特异性结合fzd7之crd的非天然发生肽的配体以之介于50nm和160nm间的ic50值抑制wnt介导的β-联蛋白信号传导。在某些实施方案中,包含(诸如本质上由其组成或由其组成)本文中提供的特异性结合fzd7之crd的非天然发生肽的配体以介于50nm和140nm之间的ic50值抑制wnt介导的β-联蛋白信号传导。在某些实施方案中,包含(诸如本质上由其组成或由其组成)本文中提供的特异性结合fzd7之crd的非天然发生肽的配体以介于50nm和120nm之间的ic50值抑制wnt介导的β-联蛋白信号传导。在某些实施方案中,包含(诸如本质上由其组成或由其组成)非本文中提供的特异性结合fzd7之crd的天然发生肽的配体以介于50nm和100nm之间的ic50值抑制wnt介导的β-联蛋白信号传导。在某些实施方案中,包含(诸如本质上由其组成或由其组成)本文中提供的特异性结合fzd7之crd的非天然发生肽的配体以介于40nm和100nm之间的ic50值抑制wnt介导的β-联蛋白信号传导。在某些实施方案中,包含(诸如本质上由其组成或由其组成)本文中提供的特异性结合fzd7之crd的非天然发生肽的配体以介于30nm和100nm之间的ic50值抑制wnt介导的β-联蛋白信号传导。在某些实施方案中,包含(诸如本质上由其组成或由其组成)本文中提供的特异性结合fzd7之crd的非天然发生肽的配体以介于20nm和100nm之间的ic50值抑制wnt介导的β-联蛋白信号传导。在某些实施方案中,包含(诸如本质上由其组成或由其组成)本文中提供的特异性结合fzd7之crd的非天然发生肽的配体以介于10nm和100nm之间的ic50值抑制wnt介导的β-联蛋白信号传导。在某些实施方案中,ic50是在双重萤光素酶测定法中作为萤光素酶活性的测量测定的,如实施例中进一步详细描述的。在某些实施方案中,包含(诸如本质上由其组成或由其组成)本文中提供的特异性结合fzd7之crd的非天然发生肽的配体具有小于约300nm,275nm,250nm,200nm,175nm,150nm,100nm,90nm,80nm,70nm,60nm,50nm,40nm,30nm,20nm,10nm,5nm,1nm,0.9nm,0.8nm,0.7nm,0.6nm,0.5nm,0.4nm,0.3nm,0.2nm,或0.1nm中任一的ec50值,包括介于这些值之间的任何范围。在某些实施方案中,ec50是使用5fam标记的肽经由fsec测定的,如实施例中进一步详细描述的。在某些实施方案中,包含(诸如本质上由其组成或由其组成)本文中提供的特异性结合fzd7之crd的非天然发生肽的配体增强wnt对fzd7之crd的募集和结合。在某些实施方案中,当肽以小于约10μm,5μm,1μm,0.9μm,0.8μm,0.7μm,0.6μm,0.5μm,0.4μm,0.3μm,0.2μm,0.1μm,0.005μm,0.001μm,或0.0005μm的浓度存在时,包括介于这些值之间的任何范围,wnt对fzd7之crd的募集和结合增强。在某些实施方案中,wnt是wnt5a。在某些实施方案中,wnt是wnt3a。在某些实施方案中,包含(诸如本质上由其组成或由其组成)本文中提供的特异性结合fzd7之crd的非天然发生肽的配体并不增强wnt对fzd3,fzd4,fzd5,fzd6,fzd8,fzd9,和/或fzd10之crd或对fzd1,fzd2,fzd3,fzd4,fzd5,fzd6,fzd8,fzd9,和/或fzd10之crd的募集和结合。在某些实施方案中,wnt是wnt5a。在某些实施方案中,wnt是wnt3a。鉴定特异性结合fzd7之富半胱氨酸域的非天然发生肽的方法在某些实施方案中,本文中提供的是一种获得结合或特异性结合fzd7之crd的非天然发生肽的方法。在某些实施方案中,方法包含(a)在容许非天然发生肽:crd复合物形成的条件下使fzd7之crd与非天然发生肽(诸如线性或环状肽)的文库接触,(b)检测复合物的形成,和(c)自复合物获得特异性结合fzd7之crd的非天然发生肽。在某些实施方案中,方法进一步包含(d)测定特异性结合fzd7之crd的非天然发生肽的核酸序列。在某些实施方案中,提供的是一种包含非天然发生肽和fzd7之crd的复合物。在某些实施方案中,将结合或特异性结合fzd7之crd的非天然发生肽提交亲和力成熟。在这个过程中,将已经发现结合fzd7之crd的肽提交选择升高的对fzd7crd的亲和力的方案(参见wuetal.(1998)procnatlacadsciusa.95,6037-42)。在某些实施方案中,在自文库筛选鉴定之后将特异性结合fzd7之crd的非天然发生肽进一步随机化。例如,在某些实施方案中,获得特异性结合fzd7之crd的非天然发生肽的方法进一步包含(e)随机化自先前鉴定的肽:crd获得的非天然发生肽的至少1,至少2,至少3,至少4,至少5,至少6,至少7,至少8,至少9,至少10,至少11,至少12,至少13,至少14,至少15,至少16,或超过16个氨基酸以生成进一步随机化的非天然发生肽,(f)使fzd7之crd与进一步随机化的非天然发生肽接触,(g)检测进一步随机化的肽:crd复合物的形成,和(h)自复合物获得进一步随机化的特异性结合fzd7之crd的非天然发生肽。在某些实施方案中,方法进一步包含(i)测定特异性结合fzd7之crd的非天然发生肽的核酸序列。在某些实施方案中,使用方法来鉴定特异性结合fzd1,fzd2,或fzd7之crd的肽。在某些实施方案中,使用方法来鉴定特异性结合fzd1,fzd2,和fzd7之crd的肽。在某些实施方案中,使用方法来鉴定特异性结合fzd1或fzd7之crd的肽。在某些实施方案中,使用方法来鉴定特异性结合fzd1和fzd7之crd的肽。在某些实施方案中,使用方法来鉴定特异性结合fzd2或fzd7之crd的肽。在某些实施方案中,使用方法来鉴定特异性结合fzd2和fzd7之crd的肽。在某些实施方案中,使用方法来鉴定特异性结合fzd1或fzd2之crd的肽。在某些实施方案中,使用方法来鉴定特异性结合fzd1和fzd2之crd的肽。在某些实施方案中,使用方法来鉴定特异性结合fzd1之crd的肽。在某些实施方案中,使用方法来鉴定特异性结合fzd2之crd的肽。在某些实施方案中,使用方法来鉴定特异性结合fzd7之crd的肽。在一个具体的实施方案中,使用方法来鉴定结合或特异性结合fzd7之crd的肽。可以实施多轮随机化,筛选和选择直至获得具有足够的对一种或多种靶crd的亲和力的非天然发生肽。如此,在某些实施方案中,步骤(e)-(h)或步骤(e)-(i)重复一,二,三,四,五,六,七,八,九,十,或超过十次以鉴定特异性结合fzd7之crd的非天然发生肽。在某些实施方案中,已经经历至少二,三,四,五,六,七,八,九,十,或超过十轮随机化,筛选和选择的肽以至少像已经经历一轮随机化,筛选,和选择的肽的亲和力一样高的亲和力结合fzd7之crd。在某些实施方案中,已经经历至少二,三,四,五,六,七,八,九,十,或超过十轮随机化,筛选和选择的非天然发生肽以比已经经历一轮随机化,筛选,和选择的非天然发生肽的亲和力要高的亲和力结合fzd7之crd。可以通过本领域已知的用于新的或改良的结合或特异性结合fzd7之crd的肽的进化的任何技术来筛选本文中描述的非天然发生肽的文库。在某些实施方案中,在固体支持物(诸如柱树脂或微量滴定板孔)上固定化fzd7之crd,并使fzd7之crd与候选非天然发生肽的文库接触。选择技术可以是例如噬菌体展示(smith(1985)science228:1315-1317),mrna展示(wilsonetal.(2001)procnatlacadsciusa98:3750-3755),细菌展示(georgiouetal.(1997)natbiotechnol15:29-34),酵母展示(boderandwittrup(1997)nat.biotechnol15:553-557)或核糖体展示(hanesandpluckthun(1997)procnatlacadsciusa94:4937-4942和wo2008/068637)。在某些实施方案中,提供的是展示本文中描述的结合或特异性结合fzd7之crd的非天然发生肽的噬菌体颗粒。噬菌体展示是作为与外壳蛋白的融合蛋白在噬菌体颗粒的表面上展示多数非天然发生肽变体的一种技术(smith,g.p.(1985)science228:1315-7;scott,j.k.andsmith,g.p.(1990)science249:386;sergeeva,a.,etal.(2006)adv.drugdeliv.rev.58:1622-54)。噬菌体展示的效用在于能对选择性随机化的蛋白质变体(或随机克隆的cdna)的大型文库快速且有效分选那些以高亲和力结合靶配体的序列的事实。已经使用肽(cwirla,s.e.etal.(1990)proc.natl.acad.sci.usa,87:6378)或蛋白质(lowman,h.b.etal.(1991)biochemistry,30:10832;clackson,t.etal.(1991)nature,352:624;marks,j.d.etal.(1991)j.mol.biol.,222:581;kang,a.s.etal.(1991)proc.natl.acad.sci.usa,88:8363)文库在噬菌体上的展示来对数以百万计的多肽或寡肽筛选具有特异性结合特性者(smith,g.p.(1991)currentopin.biotechnol.,2:668;wuetal.(1998)procnatlacadsciusa.may95,6037-42)。经由与丝状噬菌体的基因iii或基因viii任一的融合,已经使用多价噬菌体展示方法来展示小随机肽和小蛋白质(wellsandlowman,curr.opin.struct.biol.,3:355-362(1992),和其中引用的参考文献)。在单价噬菌体展示中,将蛋白质或肽文库与基因iii或其部分融合,并在野生型基因iii蛋白质存在下以低水平表达,使得噬菌体颗粒不展示或展示一个拷贝的融合蛋白。亲合效应相对于多价噬菌体降低,使得分选基于内在配体亲和力,而且使用噬菌粒载体,这简化dna操作(lowmanandwells,methods:acompaniontomethodsinenzymology,3:205-0216(1991))。分选结合fzd7之crd的非天然发生肽的噬菌体文库需要构建和扩充大量的变体,使用靶配体的亲和纯化的规程,和评估结合富集的结果的手段(参见例如us5223409,us5403484,us5571689,和us5663143)。大多数噬菌体展示方法使用丝状噬菌体(诸如m13噬菌体)。还知道λ类噬菌体展示系统(参见wo1995/34683,us5627024),t4噬菌体展示系统(renetal.(1998)gene,215:439;zhuetal.(1998)cancerresearch,58:3209-3214;jiangetal.(1997)infection&immunity,65:4770-4777;renetal.(1997)gene,195:303-311;ren(1996)proteinsci.,5:1833;efimovetal.(1995)virusgenes,10:173)和t7噬菌体展示系统(smithandscott(1993)methodsinenzymology,217:228-257;us5766905)。现在已经开发了基本噬菌体展示概念的许多其它改良和变型。这些改良增强展示系统对肽文库筛选对选定靶配体的结合和以对功能性蛋白质筛选期望特性的潜力展示这些蛋白质的能力。已经开发了用于噬菌体展示反应的组合反应装置(wo1998/14277)且已经使用噬菌体展示文库来分析和控制双分子相互作用(wo1998/20169;wo1998/20159)和受约束螺旋肽的特性(wo1998/20036)。wo1997/35196描述了一种分离亲和配体的方法,其中使噬菌体展示文库与其中配体会结合靶配体的一种溶液和其中亲和配体不会结合靶配体的第二溶液接触以选择性分离结合配体。wo1997/46251描述一种用亲和纯化的抗体生物淘选随机噬菌体展示文库,然后分离结合噬菌体,继以使用微量板孔来分离高亲和力结合噬菌体的微量淘选过程的方法。可以将此类方法应用于本文中公开的结合fzd1,fzd2,和/或fzd7之crd的非天然发生肽。还已经报告了金黄色葡萄球菌(staphylococcusaureus)蛋白a作为亲和标签的用途(lietal.(1998)molbiotech.9:187)。wo1997/47314描述底物减除文库用于区分酶特异性的用途,其中使用组合文库(可以是噬菌体展示文库)。另外的选择特异性结合蛋白质的方法描述于us5498538,us5432018,和wo1998/15833。生成肽文库和筛选这些文库的方法还描述于us5723286,us5432018,us5580717,us5427908,us5498530,us5770434,us5734018,us5698426,us5763192,和us5723323。生成包含特异性结合fzd1,fzd2,和/或fzd7之crd的非天然发生肽的配体的方法在某些实施方案中,经由遗传工程生成包含(诸如本质上由其组成或由其组成)本文中提供的结合或特异性结合fzd7之crd的非天然发生肽的配体。先前已经描述了多种用于诱变的方法(连同用于筛选或选择的适宜方法)。此类诱变方法包括但不限于例如错误倾向pcr,环改组,或在重组前的寡核苷酸定向诱变,随机核苷酸插入或其它方法。关于这些方法的进一步详情描述于例如abou-nadleretal.(2010)bioengineeredbugs1,337-340;firthetal.(2005)bioinformatics21,3314-3315;cirinoetal.(2003)methodsmolbiol231,3-9;pirakitikulr(2010)proteinsci19,2336-2346;steffensetal.(2007)j.biomoltech18,147-149;和其它。因而,在某些实施方案中,提供的是经由遗传工程技术生成的包含(诸如本质上由其组成或由其组成)本文中提供的特异性结合fzd7之crd的非天然发生肽的配体。在某些实施方案中,经由体外翻译生成包含(诸如本质上由其组成或由其组成)本文中提供的结合或特异性结合fzd7之crd的非天然发生肽的配体。简言之,体外翻译需要将蛋白质编码序列克隆入含有启动子的载体,通过用rna聚合酶转录克隆的序列来生成mrna,和通过体外翻译此mrna(例如使用无细胞提取物)来合成蛋白质。可以仅仅通过改变克隆的蛋白质编码序列来生成期望的变体蛋白质。能在小麦胚芽提取物中或在家兔网织红细胞裂解物中有效翻译许多mrna。关于体外翻译的进一步详情描述于例如hopeetal.(1985)cell43,177-188;hopeetal.(1986)cell46,885-894;hopeetal.(1987)emboj.6,2781-2784;hopeetal.(1988)nature333,635-640;和meltonetal.(1984)nucl.acidsres.12,7057-7070。因而,提供的是编码包含(诸如本质上由其组成或由其组成)本文中提供的结合或特异性结合fzd7之crd的非天然发生肽的配体的核酸分子。还提供与编码此类配体的核酸分子可操作连接的表达载体。还提供包含编码此类配体的核酸的宿主细胞(包括例如原核宿主细胞,诸如大肠杆菌,真核宿主细胞,诸如酵母细胞,哺乳动物细胞,cho细胞,等)。在某些实施方案中,经由体外翻译生成包含(诸如本质上由其组成或由其组成)本文中提供的结合或特异性结合fzd7之crd的非天然发生肽的配体。简言之,体外翻译需要将克隆的蛋白质编码序列克隆入含有启动子的载体,通过用rna聚合酶转录克隆的序列来生成mrna,和通过体外翻译此mrna(例如使用无细胞提取物)来合成蛋白质。可以仅仅通过改变克隆的蛋白质编码序列来生成期望的突变体蛋白质。能在小麦胚芽提取物中或在家兔网织红细胞裂解物中有效翻译许多mrna。关于体外翻译的进一步详情描述于例如hopeetal.(1985)cell43,177-188;hopeetal.(1986)cell46,885-894;hopeetal.(1987)emboj.6,2781-2784;hopeetal.(1988)nature333,635-640;和meltonetal.(1984)nucl.acidsres.12,7057-7070。在某些实施方案中,经由化学合成生成包含(诸如本质上由其组成或由其组成)本文中提供的结合或特异性结合fzd7之crd的非天然发生肽的配体。在某些实施方案中,化学合成本文中提供的结合或特异性结合fzd7之crd的非天然发生肽并嫁接(诸如共价连接)至一个或多个模块,如本文中别处描述的。固相和液相肽合成的方法是本领域公知的且详细描述于例如methodsofmolecularbiology,35,peptidesynthesisprotocols(m.w.penningtonandb.m.dunneds),springer,1994;welschetal.(2010)curropinchembiol14,1-15;methodsofenzymology,289,solidphasepeptidesynthesis(g.b.fieldsed.),academicpress,1997;chemicalapproachestothesynthesisofpeptidesandproteins(p.lloyd-williams,f.albericio,ande.giralteds),crcpress,1997;fmocsolidphasepeptidesynthesis,apracticalapproach(w.c.chan,p.d.whiteeds),oxforduniversitypress,2000;solidphasesynthesis,apracticalguide(s.f.kates,falbericioeds),marceldekker,2000;p.seneci,solid-phasesynthesisandcombinatorialtechnologies,johnwiley&sons,2000;synthesisofpeptidesandpeptidomimetics(m.goodman,editor-in-chief,a.felix,l.moroder,c.tmioloeds),thieme,2002;n.l.benoiton,chemistryofpeptidesynthesis,crcpress,2005;methodsinmolecularbiology,298,peptidesynthesisandapplications(j.howled)humanapress,2005;和aminoacids,peptidesandproteinsinorganicchemistry,volume3,buildingblocks,catalystsandcouplingchemistry(a.b.hughs,ed.)wiley-vch,2011。共价修饰还涵盖包含(诸如本质上由其组成或由其组成)本文中提供的结合或特异性结合fzd7之crd的非天然发生肽的配体的共价修饰。一种类型的共价修饰包括使包含(诸如本质上由其组成或由其组成)本文中提供的结合或特异性结合fzd7之crd的非天然发生肽的配体的靶定氨基酸残基与能够与配体的选定侧链或n或c端残基反应的有机衍生剂反应。用双功能剂的衍生化例如对于将配体与供纯化fzd7的方法中使用的水不溶性支持物基质或表面交联是有用的,反之亦然。常用的交联剂包括例如1,1-双(重氮乙酰基)-2-苯乙烷,戊二醛,n-羟基琥珀酰亚胺酯(例如与4-叠氮水杨酸的酯),同双功能酰亚胺酯(包括二琥珀酰亚胺基酯,诸如3,3'-二硫代双(琥珀酰亚胺基-丙酸酯)),双功能马来酰亚胺(诸如双-n-马来酰亚胺基-1,8-辛烷)和诸如甲基-3-[(对-叠氮苯基)二硫代]丙酰亚胺酯等试剂。其它修饰包括谷氨酰胺酰和天冬酰胺酰残基分别脱酰胺成相应的谷氨酰和天冬氨酰残基,脯氨酸和赖氨酸的羟基化,丝氨酰或苏氨酰残基的羟基基团的磷酸化,赖氨酸,精氨酸,和组氨酸侧链的α-氨基基团的甲基化(t.e.creighton,proteins:structureandmolecularproperties,w.h.freeman&co.,sanfrancisco,pp.79-86(1983)),n端胺的乙酰化,和任何c端羧基基团的酰胺化。包含(诸如本质上由其组成或由其组成)本文中提供的结合或特异性结合fzd7之crd的非天然发生肽的配体的另一种类型的共价修饰包含以us4640835,us4496689,us4301144,us4670417,us4791192或us4179337中所列方式将配体与多种非蛋白质性质聚合物之一连接,例如聚乙二醇(peg),聚丙二醇,或聚氧化烯。术语“聚乙二醇”或“peg”意味着聚乙二醇化合物或其衍生物,有或没有偶联剂,偶联或活化模块(例如有硫醇,三氟甲磺酸酯,三氟乙磺酸酯,氮杂环丙烷,氧杂环丙烷,n-羟基琥珀酰亚胺或马来酰亚胺模块)。术语“peg”意图指示分子量介于500和150,000da之间的聚乙二醇,包括其类似物,其中例如末端or基团已经用甲氧基基团替换(称作mpeg)。在某些实施方案中,用聚乙二醇(peg)衍生化包含(诸如本质上由其组成或由其组成)本文中提供的结合或特异性结合fzd7之crd的非天然发生肽的配体。peg是具有两个末端羟基基团的氧化乙烯重复单元的线性水溶性聚合物。peg通过它们的分子量来分类,典型地范围为约500道尔顿至约40,000道尔顿。在一个当前优选的实施方案中,所采用的peg具有范围为5,000道尔顿至约20,000道尔顿的分子量。与包含(诸如本质上由其组成或由其组成)本文中提供的结合fzd7之crd的非天然发生肽的配体偶联的peg可以是分支的或不分支的。参见例如monfardini,c.etal.,bioconjugatechem6:62-69(1995)。peg可商购自nektarinc.,sigmachemicalco.和其它公司。此类peg包括但不限于单甲氧基聚乙二醇(mepeg-oh),单甲氧基聚乙二醇-琥珀酸酯(mepeg-s),单甲氧基聚乙二醇-琥珀酰亚胺基琥珀酸酯(mepeg-s-nhs),单甲氧基聚乙二醇-胺(mepeg-nh2),单甲氧基聚乙二醇-三氟乙磺酸酯(mepeg-tres),和单甲氧基聚乙二醇-咪唑基-羰基(mepeg-im)。在某些实施方案中,对所采用的亲水性聚合物(例如peg)在一个末端处通过非反应性基团(诸如甲氧基或乙氧基基团)加帽。其后,在另一末端通过与合适的活化剂的反应来活化聚合物,活化剂诸如氰尿酰卤(例如氰尿酰氯,溴或氟),二咪唑(diimadozle),酸酐试剂(例如二卤琥珀酸酐,诸如二溴琥珀酸酐),酰基叠氮化物,对重氮苄基醚,3-(对重氮苯氧基)-2-羟基丙基醚)等等。然后使活化的聚合物与包含(诸如本质上由其组成或由其组成)本文中提供的结合或特异性结合fzd7之crd的非天然发生肽的配体反应以生成用聚合物衍生化的配体。或者,可以为了与聚合物的反应活化包含(诸如本质上由其组成或由其组成)本文中提供的结合或特异性结合fzd7之crd的非天然发生肽的配体中的官能团,或可以使用已知的偶联方法在协同偶联反应中接合两个基团。会容易领会的是,可以使用本领域技术人员知道且使用的无数其它反应方案用peg衍生化包含(诸如本质上由其组成或由其组成)本文中提供的结合或特异性结合fzd7之crd的非天然发生肽的配体。脂质体还可以作为脂质体来配制包含(诸如本质上由其组成或由其组成)本文中提供的结合或特异性结合fzd7之crd的非天然发生肽的配体。可以通过本领域已知方法来制备此类脂质体,诸如描述于epsteinetal.,procnatlacadsciusa,82:3688(1985);hwangetal.,procnatlacadsciusa,77:4030(1980);和美国专利no.4,485,045和4,544,545。具有增强的循环时间的脂质体公开于例如美国专利no.5,013,556。可以用包含磷脂酰胆碱,胆固醇,和peg衍生化磷脂酰乙醇胺(peg-pe)的脂质组合物通过反相蒸发方法来生成特别有用的脂质体。穿过期望孔径的滤器挤出脂质体来生成具有期望直径的脂质体。脂质体内任选还含有第二治疗剂。参见gabizonetal.,j.nationalcancerinst.,81(19):1484(1989)。药物组合物和配制剂在某些实施方案中,本文中提供的是药物组合物,其包含包含(诸如本质上由其组成或由其组成)本文中提供的结合或特异性结合fzd7之crd的非天然发生肽的配体和制药学可接受赋形剂。在某些实施方案中,组合物还可以含有缓冲剂,载剂,稳定剂,防腐剂和/或填充剂,以使得组合物适合于眼部施用于患者以实现期望的效果或结果。包含(诸如本质上由其组成或由其组成)本文中提供的结合或特异性结合fzd7之crd的非天然发生肽的配体可以与合适的载剂或赋形剂一起配制,使得它们适合于施用。通过混合具有期望纯度的本文中公开的配体与任选的制药学可接受载剂,赋形剂或稳定剂(remington’spharmaceuticalsciences16thedition,osol,a.ed.(1980))以冻干配制剂或水溶液形式获得本文中公开的配体的合适的配制剂。可接受的载剂,赋形剂,或稳定剂在所采用的剂量和浓度对接受者是无毒的,而且包括缓冲剂,诸如磷酸盐,柠檬酸盐,和其它有机酸;抗氧化剂,包括抗坏血酸和甲硫氨酸;防腐剂(诸如氯化十八烷基二甲基苄基铵;氯化己烷双胺;苯扎氯铵,苄索氯铵;酚,丁醇或苯甲醇;对羟基苯甲酸烃基酯,诸如对羟基苯甲酸甲酯或丙酯;邻苯二酚;间苯二酚;环己醇;3-戊醇;和间甲酚);低分子量(少于约10个残基)多肽;蛋白质,诸如血清清蛋白,明胶,或免疫球蛋白;亲水性聚合物,诸如聚乙烯吡咯烷酮;氨基酸,诸如甘氨酸,谷氨酰胺,天冬酰胺,组氨酸,精氨酸,或赖氨酸;单糖,二糖,和其它碳水化合物,包括葡萄糖,甘露糖,或糊精;螯合剂,诸如edta;糖,诸如蔗糖,甘露醇,海藻糖或山梨醇;成盐反荷离子,诸如钠;金属复合物(例如zn-蛋白质复合物);和/或非离子表面活性剂,诸如tweentm,pluronicstm或聚乙二醇(peg)。可以应用于包含(诸如本质上由其组成或由其组成)本文中提供的特异性结合fzd1,fzd2,和/或fzd7之crd的非天然发生肽的配体,或包含(诸如本质上由其组成或由其组成)本文中提供的特异性结合fzd7之crd的非天然发生肽的配体的例示性抗体配制剂描述于wo98/56418,通过援引明确收入本文。适应皮下施用的冻干配制剂描述于wo97/04801。可以用合适的稀释剂将此类冻干配制剂重建成高蛋白质浓度且可以将重建的配制剂皮下施用于本文中要治疗的哺乳动物。本文中的配制剂还可以含有所治疗的特定适应症所必需的超过一种活性化合物,优选那些活性互补且彼此没有不利影响的。例如,可能想要进一步提供抗肿瘤剂,生长抑制剂,细胞毒剂,或化疗剂。此类分子合适地以对于预定目的有效的量组合存在。此类其它药剂的有效量取决于配制剂中的量,疾病或病症或治疗的类型,和上文讨论的其它因素。这些一般以与本文所述相同的剂量和施用路径使用或以过去采用的剂量的约1至99%使用。活性组分还可以包载在例如通过凝聚技术或通过界面聚合制备的微胶囊中(例如分别是羟甲基纤维素或明胶微胶囊和聚(甲基丙烯酸甲酯)微胶囊),在胶状药物投递系统中(例如脂质体,清蛋白微球体,微乳剂,纳米颗粒和纳米胶囊)或在粗滴乳状液中。此类技术披露于remington’spharmaceuticalsciences16thedition,osol,a.ed.(1980)。可以制备持续释放制剂。持续释放制剂的合适的例子包括含有拮抗剂的固体疏水性聚合物的半透性基质,基质为成形商品形式,例如膜,或微胶囊。持续释放基质的例子包括聚酯,水凝胶(例如聚(2-羟乙基-甲基丙烯酸酯),或聚(乙烯醇)),聚交酯(美国专利no.3,773,919),l-谷氨酸和l-谷氨酸γ-乙酯的共聚物,不可降解的乙烯-乙酸乙烯,可降解的乳酸-乙醇酸共聚物诸如luprondepottm(由乳酸-乙醇酸共聚物和醋酸亮丙瑞林构成的可注射微球体),和聚-d-(-)-3-羟基丁酸。可以使用lipofectin或脂质体将本文中提供的包含(诸如本质上由其组成或由其组成)本文中提供的结合或特异性结合fzd7之crd的非天然发生肽的配体投递入细胞。活性组分还可以包载在例如通过凝聚技术或通过界面聚合制备的微胶囊中(例如分别是羟甲基纤维素或明胶微胶囊和聚(甲基丙烯酸甲酯)微胶囊),在胶状药物投递系统中(例如脂质体,清蛋白微球体,微乳剂,纳米颗粒,和纳米胶囊)或在粗滴乳状液中。此类技术披露于remington’spharmaceuticalsciences,见上文。可以制备持续释放制剂。持续释放制剂的合适的例子包括含有包含(诸如本质上由其组成或由其组成)本文中提供的结合或特异性结合fzd7之crd的非天然发生肽的配体的固体疏水性聚合物的半透性基质,基质为成形商品形式,例如膜,或微胶囊。持续释放基质的例子包括聚酯,水凝胶(例如聚(2-羟乙基-甲基丙烯酸酯),或聚(乙烯醇)),聚交酯(美国专利no.3,773,919),l-谷氨酸和l-谷氨酸γ-乙酯的共聚物,不可降解的乙烯-乙酸乙烯,可降解的乳酸-乙醇酸共聚物诸如luprondepottm(由乳酸-乙醇酸共聚物和醋酸亮丙瑞林构成的可注射微球体),和聚-d-(-)-3-羟基丁酸。虽然诸如乙烯-乙酸乙烯和乳酸-乙醇酸的聚合物能够释放分子达100天以上,但是某些水凝胶释放蛋白质的时间段较短。当所封装的包含(诸如本质上由其组成或由其组成)本文中提供的结合或特异性结合crd的非天然发生肽的配体在体内长时间维持时,它们可能由于于37℃暴露于湿气而变性或聚集,导致生物学活性损失和免疫原性可能变化。可以根据所牵涉的机制来设计稳定化的理性策略。例如,如果发现聚集机制是经由硫-二硫化物互换的分子间s-s键形成,那么可以通过修饰巯基残基,自酸性溶液冻干,控制含水量,使用适宜的添加剂,和开发特定聚合物基质组合物来实现稳定化。要用于体内施用的配制剂必须是无菌的。这容易通过例如穿过无菌滤膜过滤来实现。使用包含结合或特异性结合fzd7之富半胱氨酸域的非天然发生肽的配体的方法多种多样的器官内的连续组织维持需要干细胞。wnt信号传导已经鉴定为调节数种器官中的干细胞(包括例如胃肠道,乳腺,皮肤,肾,和卵巢)。参见cleversetal.(2014)sciencedoi:10.1126/science.1248012。干细胞能够自主增殖和自我更新,如果它们位于它们在组织内的生态位环境中的话,而且因而早就拥有癌细胞的一些特征(phesseetal.(2009)br.j.cancer100,221-227)。因此,干细胞已经鉴定为数种不同癌症的起源的细胞,包括肠,胃,前列腺和肺(visvaderetal.(2011)nature469,314-322)。wnt信号传导已经显示调节数种器官中的干细胞且因此干细胞中失调的wnt信号传导能够诱导这些器官和组织中的肿瘤发生(barkeretal.(2009)nature457,608-611)。fzd7最近已经证明是将至关紧要的wnt信号发送至lgr5+肠干细胞的主要受体(flanaganetal.(2015)stemcellreports4,759-767)。常常在人结肠肿瘤活检中观察到e3家族连接酶znrf3和rnf43(用来负调节fzd受体周转)的功能丧失(lof)突变(tcga.comprehensivemolecularcharacterizationofhumancolonandrectalcancer.nature2012,487,330-337)。rnf43的灭活性突变也赋予胰腺导管腺癌中的wnt依赖性(jiangetal.(2013)procnatlacadsciusa110,12649-12654)。r-spondin(rspo)融合产物已经显示激活结肠癌中的wnt信号传导(seshagirietal.(2012)“recurrentr-spondinfusionsincoloncancer.”nature488,660-664)。另外,usp6癌基因通过使卷曲蛋白脱泛素化来促进wnt信号传导(madanetal.(2016)procnatlacadsciusa113,e2945-2954(2016)。此类肿瘤预测对wnt信号传导过敏。如此,在某些实施方案中,提供的是一种抑制干细胞增殖的方法,其包含使干细胞与包含(诸如本质上由其组成或由其组成)本文中提供的结合或特异性结合fzd7之crd域的非天然发生肽的配体接触。在某些实施方案中,提供的是一种抑制干细胞增殖的方法,其包含使干细胞与包含(诸如本质上由其组成或由其组成)本文中提供的特异性结合fzd7之crd域的非天然发生肽的配体接触。在某些实施方案中,干细胞是肠干细胞。在某些实施方案中,提供的是一种治疗受试者中的癌症(例如结肠癌,胰腺癌,非小细胞肺癌,特征在于rnf43中的突变的癌症,特征在于usp6过表达的癌症,或特征在于牵涉r-spondin(rspo)家族成员的基因融合的癌症)的方法,其包含施用有效量的包含(诸如本质上由其组成或由其组成)本文中提供的结合或特异性结合fzd7之crd的非天然发生肽的配体。在某些实施方案中,提供的是一种治疗受试者中的癌症(例如结肠癌,胰腺癌,非小细胞肺癌,特征在于rnf43中的突变的癌症,特征在于usp6过表达的癌症,或特征在于牵涉r-spondin(rspo)家族成员的基因融合的癌症)的方法,其包含施用有效量的包含(诸如本质上由其组成或由其组成)本文中提供的特异性结合fzd7之crd的非天然发生肽的配体。在某些实施方案中,提供的是包含(诸如本质上由其组成或由其组成)本文中提供的结合或特异性结合fzd7之crd的非天然发生肽的配体,供在制造用于治疗癌症(例如结肠癌,胰腺癌,非小细胞肺癌,特征在于rnf43中的突变的癌症,特征在于usp6过表达的癌症,或特征在于牵涉r-spondin(rspo)家族成员的基因融合的癌症)的药物中使用。在某些实施方案中,提供的是包含(诸如本质上由其组成或由其组成)本文中提供的特异性结合fzd7之crd的非天然发生肽的配体,供在制造用于治疗癌症(例如结肠癌,胰腺癌,非小细胞肺癌,特征在于rnf43中的突变的癌症,特征在于usp6过表达的癌症,或特征在于牵涉r-spondin(rspo)家族成员的基因融合的癌症)的药物中使用。在某些实施方案中,提供的是包含(诸如本质上由其组成或由其组成)本文中提供的结合或特异性结合fzd7之crd的非天然发生肽的配体,供治疗受试者中的癌症(例如结肠癌,胰腺癌,非小细胞肺癌,特征在于rnf43中的突变的癌症,特征在于usp6过表达的癌症,或特征在于牵涉r-spondin(rspo)家族成员的基因融合的癌症)中使用。在某些实施方案中,提供的是包含(诸如本质上由其组成或由其组成)本文中提供的特异性结合fzd7之crd的非天然发生肽的配体,供治疗受试者中的癌症(例如结肠癌,胰腺癌,非小细胞肺癌,特征在于rnf43中的突变的癌症,特征在于usp6过表达的癌症,或特征在于牵涉r-spondin(rspo)家族成员的基因融合的癌症)中使用。在某些实施方案中,提供的是包含包含(诸如本质上由其组成或由其组成)本文中提供的结合或特异性结合fzd7之crd的非天然发生肽的配体的组合物(诸如药物组合物),供治疗受试者中的癌症(例如结肠癌,胰腺癌,非小细胞肺癌,特征在于rnf43中的突变的癌症,特征在于usp6过表达的癌症,或特征在于牵涉r-spondin(rspo)家族成员的基因融合的癌症)中使用。在某些实施方案中,提供的是包含包含(诸如本质上由其组成或由其组成)本文中提供的特异性结合fzd7之crd的非天然发生肽的配体的组合物(诸如药物组合物),供治疗受试者中的癌症(例如结肠癌,胰腺癌,非小细胞肺癌,特征在于rnf43中的突变的癌症,特征在于usp6过表达的癌症,或特征在于牵涉r-spondin(rspo)家族成员的基因融合的癌症)中使用。在某些实施方案中,配体包含(诸如本质上由其组成或由其组成)包含seqidno:13或seqidno:99中所列氨基酸序列的非天然发生肽。在某些实施方案中,要治疗的受试者是哺乳动物(例如人,非人灵长动物,大鼠,小鼠,牛,马,猪,绵羊,山羊,犬,猫,等)。在某些实施方案中,受试者是人。在某些实施方案中,受试者是临床患者,临床试验志愿者,实验动物,等。在某些实施方案中,受试者怀疑具有癌症或处于具有癌症的风险(例如结肠癌,胰腺癌,非小细胞肺癌,特征在于rnf43中的突变的癌症,特征在于usp6过表达的癌症,或特征在于牵涉r-spondin(rspo)家族成员的基因融合的癌症)。施用和剂量给药包含(诸如本质上由其组成或由其组成)本文中提供的结合或特异性结合fzd7之crd的非天然发生肽的配体的施用可以通过任何合适路径,包括例如静脉内,肌肉内,或皮下。在某些实施方案中,与第二,第三,或第四药剂(包括例如抗肿瘤剂,生长抑制剂,细胞毒剂,或化疗剂)组合施用包含(诸如本质上由其组成或由其组成)本文中提供的特异性结合crd的结合或特异性结合fzd7之crd的非天然发生肽的配体以治疗牵涉例如异常wnt活性的疾病或病症。在某些实施方案中,包含(诸如本质上由其组成或由其组成)本文中提供的结合或特异性结合fzd7之crd的非天然发生肽的配体是与另外的药剂缀合的。此类药剂包括例如化疗剂。在某些实施方案中,包含(诸如本质上由其组成或由其组成)本文中提供的结合或特异性结合fzd7之crd的非天然发生肽的配体是与另外的药剂缀合的。可以经由任何路径将包含(诸如本质上由其组成或由其组成)本文中提供的结合或特异性结合fzd7之crd的非天然发生肽的配体施用于个体,包括但不限于静脉内(例如通过输注泵),腹膜内,眼内,动脉内,肺内,口服,吸入,囊内,肌肉内,气管内,皮下,眼内,鞘内,经皮,经胸膜,动脉内,表面,吸入(例如作为喷雾),粘膜(诸如经由鼻粘膜),皮下,经皮,胃肠,关节内,池内,室内,直肠(即经由栓剂),阴道(即经由阴道栓剂),颅内,尿道内,肝内,和肿瘤内。在一些实施方案中,包含(诸如本质上由其组成或由其组成)本文中提供的结合或特异性结合fzd7之crd的非天然发生肽的配体是系统施用的(例如通过静脉内注射)。在一些实施方案中,包含(诸如本质上由其组成或由其组成)本文中提供的结合或特异性结合fzd7之crd的非天然发生肽的配体是局部施用的(例如通过动脉内或眼内注射)。取决于要治疗的适应症和本领域熟练内科医师会熟悉的与剂量给药有关的因素,会以对于治疗该适应症有效同时最小化毒性和副作用的剂量施用包含(诸如本质上由其组成或由其组成)本文中提供的结合或特异性结合fzd7之crd的非天然发生肽的配体。一种典型剂量可以例如在0.001至1000μg的范围中;然而,在这种例示性范围以下或以上的剂量在本发明的范围内。日剂量可以是约0.1μg/kg至约100mg/kg总体重(例如约5μg/kg,约10μg/kg,约100μg/kg,约500μg/kg,约1mg/kg,约50mg/kg,或由任何两个上述数值定义的范围),优选约0.3μg/kg至约10mg/kg总体重(例如约0.5μg/kg,约1μg/kg,约50μg/kg,约150μg/kg,约300μg/kg,约750μg/kg,约1.5mg/kg,约5mg/kg,或由任何两个上述数值定义的范围),更优选约1μg/kg至1mg/kg总体重(例如约3μg/kg,约15μg/kg,约75μg/kg,约300μg/kg,约900μg/kg,或由任何两个上述数值定义的范围),和甚至更优选约0.5至10mg/kg体重每天(例如约2mg/kg,约4mg/kg,约7mg/kg,约9mg/kg,或由任何两个上述数值定义的范围)。如上所述,可以通过定期评估所治疗的患者来监测治疗或预防功效。对于数天或更久的重复施用,取决于状况,重复治疗直至疾病症状的期望遏制发生。然而,其它剂量方案可能是有用的且在本发明的范围内。可以通过单次推注施用组合物,通过多次推注施用组合物,或通过连续输注施用组合物来投递期望的剂量。可以一天一,二,三,或四次施用包含包含(诸如本质上由其组成或由其组成)本文中提供的结合或特异性结合fzd7之crd的非天然发生肽的配体的药物组合物。还可以以比每天要低的频率施用组合物,例如一周六次,一周五次,一周四次,一周三次,一周两次,一周一次,每两周一次,每三周一次,一月一次,每两月一次,每三月一次,或每六月一次。还可以在持续释放配制剂中,诸如在植入物中施用组合物,逐渐释放组合物供一段时间上的使用,且容许以较低的频率施用组合物,诸如一月一次,每2-6个月一次,每年一次,或甚至单次施用。可以通过注射来施用持续释放装置(诸如团粒,纳米颗粒,微粒,纳米球体,微球体,等等)。可以以单个日剂量施用包含(诸如本质上由其组成或由其组成)本文中提供的结合或特异性结合fzd7之crd的非天然发生肽的配体,或者可以以一天两,三,或四次的分剂量施用总的日剂量。还可以以比每天要低的频率施用组合物,例如一周六次,一周五次,一周四次,一周三次,一周两次,一周一次,每两周一次,每三周一次,一月一次,每两月一次,每三月一次,或每六月一次。还可以在持续释放配制剂中,诸如在植入物中施用组合物,逐渐释放组合物供一段时间上的使用,且容许以较低的频率施用组合物,诸如一月一次,每2-6个月一次,每年一次,或甚至单次施用。可以在各种位置中通过注射或手术植入来施用持续释放装置(诸如团粒,纳米颗粒,微粒,纳米球体,微球体,等等)。制品和试剂盒在某些实施方案中,提供的是一种制品,其含有本文中的包含(诸如本质上由其组成或由其组成)本文中提供的结合或特异性结合fzd7之crd的非天然发生肽的配体和/或包含此类配体的药物组合物,以及对于治疗癌症(诸如结肠癌,胰腺癌,非小细胞肺癌,特征在于rnf43中的突变的癌症,特征在于usp6过表达的癌症,或特征在于牵涉r-spondin(rspo)家族成员的基因融合的癌症)有用的材料。制品可以包含容器和在容器上或与容器关联的标签或包装插页。合适的容器包括例如瓶,管形瓶,注射器,等。容器可以自多种材料形成,诸如玻璃或塑料。在某些实施方案中,容器装有无菌单剂包装。标签或包装插页指示组合物用于治疗癌症(诸如结肠癌,胰腺癌,非小细胞肺癌,特征在于rnf43中的突变的癌症,特征在于usp6过表达的癌症,或特征在于牵涉r-spondin(rspo)家族成员的基因融合的癌症)。标签或包装插页会进一步包含关于将包含(诸如本质上由其组成或由其组成)本文中提供的结合或特异性结合fzd7之crd的非天然发生肽的配体施用于患者的用法说明书。还涵盖包含本文中描述的组合疗法的制品和试剂盒。包装插页指通常包括在治疗性产品的商业包装中的用法说明书,其含有关于涉及此类治疗性产品使用的适应症,用法,剂量,施用,禁忌症和/或警告的信息。在某些实施方案中,包装插页指示包含(诸如本质上由其组成或由其组成)本文中提供的结合或特异性结合fzd7之crd的非天然发生肽的配体(或包含此类配体的药物组合物)用于治疗癌症(诸如结肠癌,胰腺癌,非小细胞肺癌,特征在于rnf43中的突变的癌症,特征在于usp6过表达的癌症,或特征在于牵涉r-spondin(rspo)家族成员的基因融合的癌症)。另外,制品可以进一步包含第二容器,其包含制药学可接受缓冲剂,诸如抑菌性注射用水(bwfi),磷酸盐缓冲盐水,林格(ringer)氏溶液和右旋糖溶液。它可以进一步包括从商业和使用者立场看想要的其它材料,包括其它缓冲剂,稀释剂,滤器,针头,和注射器。还提供对于多种目的有用的试剂盒,例如用于分离或检测患者中的fzd7,任选与制品组合。为了分离和纯化fzd7,试剂盒可以含有与珠(例如sepharose珠)偶联的包含(诸如本质上由其组成或由其组成)本文中提供的结合或特异性结合fzd7之crd的非天然发生肽的配体。为了分离和纯化fzd7,试剂盒可以含有与珠(例如sepharose珠)偶联的包含(诸如本质上由其组成或由其组成)本文中提供的特异性结合fzd7之crd的非天然发生肽的配体。可以提供试剂盒,其含有包含(诸如本质上由其组成或由其组成)本文中提供的结合或特异性结合fzd7之crd的非天然发生肽的配体,用于在体外检测和量化fzd7,例如在elisa或印迹中。与制品一样,试剂盒包含容器和在容器上或与容器关联的标签或包装插页。例如,容器装有包含至少一种本文中描述的包含(诸如本质上由其组成或由其组成)本文中提供的结合或特异性结合fzd7之crd的非天然发生肽的配体的组合物。可以包括另外的容器,其含有例如稀释剂和缓冲剂,对照抗体,等。标签或包装插页可以提供组合物的描述以及关于预定体外或诊断用途的用法说明书。实施例实施例1:用于实施例2-5的材料和方法试剂,质粒,抗体和重组蛋白。小鼠wnt3a(目录号1324-wn/cf)蛋白质购自r&dsystems。fzdcrd-fc和sfrp蛋白质(r&dsystems)在pbs中溶解至10μm且包括:hfzd1crd-fc(目录号5988-fz),mfzd2crd-fc(目录号1307-fc),hfzd4crd-fc(目录号5847-fz),hfzd5crd-fc(目录号1617-fz),hfzd7crd-fc(目录号6178-fz),mfzd7crd-fc(目录号198-fz),hfzd8crd-fc(目录号6129-fz),mfzd9crd-fc(目录号2440-fz),hfzd10crd-fc(目录号3459-fz),hsfrp1(目录号5396-sf),hsfrp2(目录号6838-fr),hsfrp-3(目录号192-sf),hsfrp-4(目录号1827-sf),和hsfrp-5(目录号6266-sf)。生物素化的wnt3a(bio-wnt3a)和wnt5a(bio-wnt5a)由r&dsystems生成(当前不是目录项目)。简言之,wnt3a或wnt5a透析入含有0.5%chaps的pbs,ph7.4并依照制造商的推荐生物素化(pierce,目录号21338)。pcdna3.2wnt1和wnt3a构建物得自opensourcewnt计划(najdietal.(2012)differentiation84,203-213)。抗lrp6和抗豚草抗体如先前所述生成(tianetal.(2015)cellrep11,33-42)。通过标准fmoc化学合成肽。其它试剂包括:生物素化的过氧化物酶(目录号432040;invitrogen),山羊抗人iggfc-hrp(目录号a18817;thermofisherscientific),dmso(目录号d2650;sigma),和fugene-hd(目录号e2311;promega)。hfzd7crd-his作为分泌型蛋白质在表达endoh的粉纹夜蛾(trichoplusiani)细胞中表达,用几夫碱处理,然后通过标准ni-nta亲和层析术继以大小排阻层析术纯化,如别处所述(bourhisetal.(2010)j.biol.chem.285,9172-9179)。用150mmnacl,50mmtris-hcl,ph7.5清洗亲和珠。对应于单体的大小的主峰用多轮大小排阻层析术纯化,在相同缓冲液中以0.5ml/min使用superdex75柱。与fz7-21融合的hfzd7crd作为分泌型蛋白质在粉纹夜蛾细胞中表达并通过ni-nta亲和层析术纯化。用150mmnacl,50mmtris-hcl,ph7.5清洗珠。二聚体主峰用多轮大小排阻层析术纯化,在相同缓冲液中以7ml/min使用srt-1021.2x300mmsec300柱。为fzd3和fzd6蛋白质设计的多种构建物在粉纹夜蛾或sf9昆虫细胞中作为分泌型蛋白质测试且它们的表达通过还原或非还原条件中的sds-page监测。无一测试条件为下游分析提供足够质量的蛋白质。使用下面的uniprotid进行序列比对:fzd1,q9up38;fzd2,q14332;fzd3,q9npg1;fzd4,q9ulv1;fzd5,q13467;fzd6,o60353;fzd7,o75084;fzd8,q9h461;fzd9,o00144;fzd10,q9ulw2;hsfrp1,q8n474;hsfrp2,q96hf1;hsfrp3,q92765;hsfrp4,q6fhj7;和hsfrp5,q5t4f7。针对hfzd7crd-fc的噬菌体分选。遵循标准方案(tonikianetal.(2007)natprotoc2,1368-1386)线性肽文库的噬菌体集合(stanger,k.etal.allostericpeptidesbindacaspasezymogenandmediatecaspasetetramerization.natchembiol8,655-660(2012))循环经受多轮用hfzd7crd-fc的结合选择。在所有轮次的分选期间herceptin(10μm)包括在分选溶液中以封闭潜在的fc结合物。在四轮结合选择之后,使用板固定化fzd7crd-fc作为靶或脱靶在高通量点噬菌体elisa中分析噬菌体克隆个体进行特异性测试(hfzd8crd-fc和herceptin)。为了测量脱靶亲和力,针对bsa测定噬菌体颗粒副本以测量非特异性结合。具有噬菌体结合信号>0.5和信噪比>10的克隆认为是阳性克隆且提交dna序列分析。使用microsoftexcel2010进行一般数据处理,除非另有说明。肽合成。所有肽使用标准9-芴基甲氧基羰基方案合成,如别处所述(zhangetal.(2009)natchembiol5,217-219),n端作为乙酰胺保护而c端作为羧酰胺保护,并通过rp-hplc纯化。肽质量(>90%纯度)通过lc-ms检查,而肽身份通过ms确认。细胞培养和转染。稳定整合有萤火虫萤光素酶wnt报告物(topbrite,tb)(zhangetal.(2009)natchembiol5,217-219)和prl-sv40海肾(renilla)萤光素酶的hek293细胞(目录号e2231;promega)在补充有10%fbs,2mmglutamaxtm(目录号35050-061;gibco)和40μg/ml潮霉素(目录号30-240-cr;cellgro)的dmem:f12(50:50)中生长。在任何实验之前细胞在5%co2增湿温箱中于37℃温育24小时。对于萤光素酶报告物测定法,在透明底白色聚苯乙烯96孔板(目录号353377;falcon)的每个孔中接种50μlhek293-tb细胞(20,000个细胞/孔)。在24小时之后,将细胞用指定wnt构建物和fugenehd(目录号e2312;promega;在10μloptimem[目录号31985-070;gibco]中混合cdna和fugene)转染24小时,然后用指定肽处理6小时,并用50μldual-glo萤光素酶测定系统(目录号e2940;promega)加工。对于用肽处理,所有肽在dmso中稀释,并以dmso终浓度1%添加至细胞。对于用重组wnt蛋白质处理,所有wnt在pbs中稀释,与肽同步添加至细胞,然后如上所述加工。在perkinelmer多标记物读数仪上测量萤火虫和海肾发光。计算萤火虫发光对海肾发光的比,减去背景,并针对仅用wnt3a(50ng/ml)处理的对照细胞中的比标准化。细胞系得自genentechgcell实验室且测试支原体污染并通过snp分析认证。facs分析。稳定表达gd-fzd的细胞系在补充有10%fbs(目录号1500-100;seradigm),1xglutmax(目录号35050-061;gibco)和g418(200μg/ml;目录号a1720;sigma)的dmem:f12(50:50)培养基中生长至~80%汇合,用含0.05%胰蛋白酶/edta的pbs(目录号15400-054;gibco)释放,在冰上收集并缓冲液交换入冰冷的facs缓冲液(含0.5%bsa和0.05%naaz的pbs)。100,000个细胞/孔分配入冰冷的u底板(目录号3799;costar)且细胞通过以1200rpm离心3分钟而形成团粒。细胞用facs缓冲液中补充有小鼠抗gd抗体(genentech;gd:952;500ng/ml)的dmso(目录号d2650;sigma)或5fam-fz7-21(12.5μm)在黑暗中在冰上处理1小时。用5fam-fz7-21s处理的细胞展示显著的非特异性结合且排除在分析外。细胞在冰上用冰冷的facs清洗缓冲液(含0.5%bsa和0.1%naaz的pbs)清洗三次。然后细胞用alexa647缀合的驴抗小鼠igg(h+l)二抗(500ng/ml;目录号a-31571;invitrogen)在黑暗中在冰上处理1小时。细胞用冰冷的facs清洗缓冲液在冰上清洗三次,然后与sytox(目录号s34857;invitrogen)一起温育15分钟,之后在bdfortessa仪器上分析。数据用facsdiva(bd;8.0.1版)收集并使用flowjo(flowjo,llc;v10.1)分析。elisa。384孔maxisorp板(目录号464718;thermoscientific)与30μl中性亲合素(目录号31000,thermoscientific;2μg/ml,在pbs中)一起于4℃温育过夜。然后板用pbst清洗并用pbs/1%bsa于室温封闭1小时,然后用pbst清洗,之后进一步加工。结合测定法在96孔板(目录号83007-374;vwr)中进行,它们用pbs/1%bsa于4℃封闭过夜。hfzd4crd-fc或hfzd7crd-fc(63ng/ml)与bio-wnt3a(25ng/ml)或bio-wnt5a(50ng/ml)一起在指定肽在pbs中的4倍连续稀释液存在下于4℃温育过夜。所有测定法包括hfzd9crd-migg2a蛋白质作为阴性对照。hfzd7crd-his在150mmnacl,50mmtris-hcl,ph7.5中悬浮且当在测定法中使用时使用等体积的相同缓冲液对照。结合测定法混合物转移至中性亲合素包被的板并于室温温育~1小时。然后孔用pbst清洗,与抗higg1-hrp(在补充有1%bsa的pbs中1:10,000悬浮;目录号a18817,invitrogen)一起温育,并再次用pbst清洗。随添加tmb试剂(遵循制造商的推荐;kpl;50-76-00)达15分钟,之后添加等体积的1m磷酸达10分钟来发生信号。使用tekanm1000读板仪于450nm测量吸光度。收集值并使用prismgraphpad软件绘图。所有使用肽的测定法条件处于1%dmso终浓度。荧光大小排阻层析术。使用aktafplc系统(generalelectriccompany)及unicorn软件包(5.31版)运行荧光大小排阻层析术(fsec)实验。在磷酸盐缓冲液中以0.5ml/min使用sec3000或sec2000(phenomenex;torrence,ca)柱解析样品。以正常模式(激发/发射,550/494nm;增益=100;std=32)使用fp-2020plus荧光检测仪(jascoanalyticalinstruments,easton,md)监测荧光。所有样品以dmso终浓度1%制备并于4℃温育过夜,之后进行样品分离。fzdcrd-fc样品在过量5fam标记肽(1μm)存在下以~250nm保存。人分泌型卷曲蛋白相关蛋白在过量5fam标记肽(10μm)存在下以~250nm维持。大小排阻层析术偶联多角度光散射。hfzd7crd(15.8μm)与fz7-21,fz7-21s或dmso一起以指定的蛋白质对肽比温育2小时,之后通过大小排阻层析术(sec)分析并用多角度静态光散射(mals)检测。在具有1%dmso终浓度的150mmnacl,50mmtris-hcl,ph7.5缓冲液中以0.15ml/min通过superdexs2003.2/300柱(generalelectrichealthcarelifescience,pittsburgh,pa;目录号28990946)解析样品。在连接dawnheleos-ii多角度静态光散射(mals)检测仪和optilabt-rex差分折射率(dri)检测仪(wyatttechnologies,santabarbara,california)的1260infinityhplc(agilenttechnology,santaclara,ca)上实施运行。x射线晶体学。纯化hfzd7crd-his,缓冲液交换入150mmnacl,50mmtris-hcl,ph7.5,然后通过离心(目录号ufc900325;milliporeamiconultra15)浓缩至~25mg/ml。用25%peg2000mme(w/v)及mesph6.5(目录号134312;qiagen)稀释(1:1)蛋白质并于19℃通过蒸气扩散来生长。在10天内长出足够质量的晶体,在补充有30%peg2000mme的母液中低温保护,并在液氮中骤冻。在斯坦福同步辐射光源(ssrl)12-2处收集数据集并以分辨率通过分子置换来解析(nileetal.(2017)“unsaturatedfattyacylrecognitionbyfrizzledreceptorsmediatesdimerization.”proc.natl.acad.sci.usa114,4147-4152,lid-4110.1073/pnas.1618293114[doi])。在表达endoh的粉纹夜蛾细胞中作为分泌型蛋白质表达与fz7-21融合的hfzd7crd[gln33–gly168]并用几夫碱处理。作为分泌型蛋白质将hfzd7crd插入pacgp67-a载体(bdbiosciences-pharmingen)。使与fz7-21融合的hfzd7crd在ni-nta珠(qiagen;1018401)上通过,用300mmnacl,50mmtris-hcl,ph7.5清洗,然后用含有300mm咪唑的相同缓冲液洗脱。收集洗出液,浓缩并缓冲液交换入150mmnacl,50mmtris-hcl,ph7.5并在150mmnacl,50mmtris-hcl,ph8.0中通过凝胶过滤(superdex200;gehealthcarelifesciences)收集二聚体集合。通过离心将二聚体级分浓缩至~20mg/ml并将蛋白质用1-丙醇14%(v/v;目录号09158;fluka),9%peg5000mme(目录号hr-2-615;hamptonresearch),和0.1mmes,ph6.9(目录号hr2-243;hamptonresearch)稀释成1:1混合物并于4℃通过蒸气扩散来生长。在具有像素阵列检测器(pilatus,dectrisag,switzerland)的高级光子源(argonnenationallaboratory)光束线17id处收集x射线衍射数据。x射线波长设置于在低温(-180℃)下用单晶收集完整数据集。使用程序xds(kabsch,w.integration,scaling,space-groupassignmentandpost-refinement.actacrystallographicasectiond:biologicalcrystallography66,133-144(2010))对衍射数据积分并使用ccp4包(murshudov,g.n.,vagin,a.a.&dodson,e.j.refinementofmacromolecularstructuresbythemaximum-likelihoodmethod.actacrystallogrdbiolcrystallogr53,240-255(1997))的程序aimless(blessing,r.h.anempiricalcorrectionforabsorptionanisotropy.actacrystallogra51(pt1),33-38(1995))缩放。使用程序phaser(mccoy,a.j.etal.phasercrystallographicsoftware.journalofappliedcrystallography40,658-674(2007))通过分子置换(mr)方法对结构定相。使用hfzd7crd结构作为mr搜索模型(pdbid#5urv)(nile,a.h.,mukund,s.,stanger,k.,wang,w.&hannoush,r.n.unsaturatedfattyacylrecognitionbyfrizzledreceptorsmediatesdimerization.proc.natl.acad.sci.usa114,4147-4152,lid-4110.1073/pnas.1618293114[doi](2017))。随后将结构提交用制图学程序coot(emsley,p.,lohkamp,b.,scott,w.g.&cowtan,k.featuresanddevelopmentofcoot.actacrystallographicasectiond:biologicalcrystallography66,486-501(2010))的迭代模型构建和程序phenix(adams,p.d.etal.phenix:acomprehensivepython-basedsystemformacromolecularstructuresolution.actacrystallographicasectiond66,213-221(2010))中编码的最大似然最小二乘精修方案。最终结构精修至分辨率,有88.3%的残基落在ramachandran图中的有利区域中,11.1%在允许区域中,0.3%在一般允许区域中且0.2%在不允许区域中。在fzd7crd疏水腔内观察到随机电子密度;然而,无法做出自信的指派。更加详细的衍射数据加工和结构精修统计量在表10中可得。在高级光子源(argonnenationallaboratory)光束线处收集x射线衍射数据。与fz7-21复合的fzd7crd的结构存放于pdb数据库,代码5wbs。结构分析。为了计算和展示fzd分子的保守性,作为替代品使用人(h)fzd8的表面以呈现10种hfzdcrd之间的保守性(hfzd1至hfzd10;青色,低保守性;栗色,高保守性)。自uniprot获取hfzdcrd序列,用ugene46(v1.14.1)(okonechnikov,k.,golosova,o.,fursov,m.&team,t.u.uniprougene:aunifiedbioinformaticstoolkit.bioinformatics28,1166-1167(2012))聚集,并用clustalomega33比对。通过迭代比对的指导,修剪序列,使用hfzd7crd(y35至d167)作为指导,严重依赖于绝对保守的半胱氨酸残基作为crd边界的参考。将比对文件输出至ucsfchimera(v1.10.2)(pettersen,e.f.etal.ucsfchimera--avisualizationsystemforexploratoryresearchandanalysis.jcomputchem25,1605-1612(2004))并使用al2co算法(pei,j.&grishin,n.v.al2co:calculationofpositionalconservationinaproteinsequencealignment.bioinformatics17,700-712(2001))计算序列保守性,使用通过独立计数的频率估算,经由基于熵的办法和平均窗口1及缺口分数0.5测量保守性。自al2co计算的保守性值代表与均值的标准偏差且范围为颜色梯度上的+1.678(最保守;栗色)至-1.539(最不保守;青色)且在mfzd8crd(pdbid#4f0a)中作为分子表面展示梯度。为了计算dfz7-21之间的分子内相互作用,使用ucsfchimera47内的找到碰撞/接触效用来鉴定两个原子之间的交叠,定义为它们的vdw半径之和减去它们之间的距离并减去潜在成氢键对的容许值[交叠ij=rvdwi+rvdwj–dij–容许ij]。此计算鉴定所有直接相互作用,包括极性的和非极性的,有利的和不利的(包括碰撞),其中vdw交叠>-没有减去潜在成氢键对的交叠。在ucsfchimera包内使用msms包(sanner,m.f.,olson,a.j.&spehner,j.c.reducedsurface:anefficientwaytocomputemolecularsurfaces.biopolymers38,305-320(1996))计算分离的dfz7-21分子(a链和b链)的溶剂可及表面(sas)分子表面(探针半径)。hfzd7crd疏水腔展示。为了显现与fz7-21复合的hfzd7crd内的疏水腔,采用needleman-wunsch比对算法和blosum-62矩阵,使用ucsfchimera中的matchmaker效用将与mfzd8crd复合的两个xwnt8分子(pdbid#f40a)叠加到hfzd7crd结构上。在脂肪酰基模块~内鉴定连续疏水性表面。在脱辅基hfzd7crd与f40a的叠加中,自它们的ω-碳截短脂肪酸以消除疏水腔内的脂肪酸之间的碰撞,然后如上文所述实施疏水腔搜索以鉴定在脂肪酰基基团~内的疏水性表面。计算机分析。在moe(chemicalcomputinggroup;2017.05版)中构建在图39a和39b中呈现的与hfzd7crd(pdbid#5urv)(有延长的脂肪酰基模块(c16:n-7)在hfzd7crd的u形疏水腔中结合)复合的xwnt8a(pdbid#4f0a)的模型。使用mfzd8crd(pdbid#4f0a)作为指导将xwnt8a叠加到c24结合的hfzd7crd结构(pdbid#5urv)上。使用缺省设置应用protonate3d和结构制作效用以优化氢放置。修剪并延长在xwnt8aser187处共价连接的c14脂肪酸以跟随hfzd7crd疏水腔内的c24脂肪酸的轮廓。然后使用mmff94x力场(eps=r;截留[8,10])及刚性水和0.1rmskcal/mol/a2梯度将密切靠近的脂肪酸和残基能量最小化。如上文所述在moe(chemicalcomputinggroup;2017.05版)中构建在图39c和39d中找到的与同fz7-21结合的hfzd7crd(有延长的脂肪酰基模块(c16:n-7)在hfzd7crd的疏水腔中结合)复合的两个分子的xwnt8a(pdbid#4f0a)的模型。对于每种结构展示,使用相应的配体来定义疏水腔。在结合的配体~内鉴定连续疏水性表面并依照腔的疏水性显现。基于kyte-doolittle打分度量(kyte,j.&doolittle,r.f.asimplemethodfordisplayingthehydropathiccharacterofaprotein.journalofmolecularbiology157,105-132(1982))对依据氨基酸疏水性的分子表面定标。将坐标输出入ucsfchimera(1.11版)以生成图像。nmr光谱学。在配有冷冻探针的brukerultrashieldplus600mhz光谱仪上实施所有nmr实验。将肽在含10mm磷酸盐缓冲液(ph7.3)和10%(v/v)乙腈-d3(isotec,inc.;目录号t82-05013-ml)的h2o中以1mm重悬浮以测定溶液中的nmr结构。于27℃分别以70和350ms的混合时间记录肽的总相关光谱术(tocsy)和核overhauser效应光谱术(noesy)。使用topspin(brukerbiospin)和ccpn分析进行谱加工,显现和峰采集(vrankenetal.(2005)proteins59,687-696)。基于同核tocsy和noesy谱,指派肽的残基。基于自旋系统鉴定和顺序指派(wüthrich,k.nmrofproteinsandnucleicacids.(wileyinterscience,newyork,1986)获得质子的几乎完全共振指派。自noesy谱获得质子间距离约束。最初使用cyana3.97包(guntert&buchner(2015)journalofbiomolecularnmr62,453-471;guntertetal.(1997)jmolbiol273,283-298)计算dfz7-21的三维结构。因为只观察到一组共振,所以在具有重复序列和对称距离约束的cyana计算中作为对称二聚体处理dfz7-21。见表16。然后用cns(brunger,a.t.etal.crystallography&nmrsystem:anewsoftwaresuiteformacromolecularstructuredetermination.actacrystallogrdbiolcrystallogr54,905-921(1998);brunger,a.t.version1.2ofthecrystallographyandnmrsystem.natprotoc2,2728-2733(2007))精修结构。生成总共50种结构并选择20种具有最低能量的结构。有序残基(残基6-14)通过procheck的ramachandran分析显示63.1%残基在最有利区域中,27.2%残基在另外允许区域中且9.7%残基在宽大允许区域中,有0%残基在不允许区域中。感兴趣肽没有同位素标记,所以没有获得主链核的指派(ca,cb,ha,co)。在没有主链指派的情况下,结构计算中不包括二面角约束。结构计算中使用的化学位移指派和noe约束存放入bmrb,代码30311。结构存放于pdb数据库,代码5w96。类器官培养。如先前所述(satoetal.(2009)nature459,262-265(2009))自分离的隐窝(自整个长度的小肠收集)建立小鼠类器官并维持。所有小鼠衍生组织依照genentech(roche基团的一个成员)和机构动物护理和使用委员会的动物使用指南实施,符合加利福尼亚州法律和伦理实践。不选择小鼠性别。类器官传代至少每周两次并使用intesticult类器官生长培养基(目录号06005;stemcelltechnologies)生长。在dmso中悬浮肽并在培养基中溶解至1%dmso终浓度,然后添加至类器官以启动药物处理。添加抗体至10μg/ml浓度。使用andorneo相机(1x1分仓;200ms曝光)使用具有nikonplanfluor10xph1dll物镜的nikoneclipsetiscope对类器官成像并使用niselements(v4.5064-bit;nikon)获取。如先前所述(melo,f.d.s.e.etal.adistinctroleforlgr5+stemcellsinprimaryandmetastaticcoloncancer.nature543,676-680(2017))生成apcmin类器官。对于用chir99021(5μm;stemcelltechnologies)处理的类器官,分拆类器官并用dmso或chir99021(5μm)处理24小时,之后添加肽或dmso达24小时。对于在补充图22中找到的共焦图像,使用leicasp5激光扫描共焦显微镜用40x油浸物镜(hcxplapocsuv,1.25na)在蓝色(dapi)和绿色(gfp)通道中收集图像。所有图像如下以顺序模式在相同条件下获取:序列1(蓝色通道):uv激光(405nm):85%激发功率,pmt设置:425-470nm发射,主动增益:900,偏移:-15;序列2(绿色通道):氩激光(488nm),20%输出,55%激发功率,pmt设置:500-550nm发射,主动增益700,偏移:-5。以400hz单向扫描速度收集2048x2048像素分辨率(1.5倍变焦)的图像,具有1个airy单位针孔。收集10-20个光学切面(0.968微米厚)的堆叠并在图中使用最大强度投射的图像。rna提取和rt-pcr。使用rneasymicro试剂盒(目录号74004;qiagen)依照制造商的说明书自类器官样品分离rna。使用rneasymini试剂盒(目录号74104;qiagen)收集小鼠小肠rna。使用一步实时rt-pcrmaster混合物(目录号4392938;lifetechnologies)依照制造商的说明书在具有50ng总rna的10μl反应中实施qrt-pcr。使用来自lifetechnologies的taqman探针:actb(mm01205647_g1);lgr5(mm01251801_m1);轴蛋白2(mm00443610_m1);ascl2(mm01268891_g1);olfm4(mm01320260_m1);muc2(mm01276696_m1);actb(hs99999903_m1);actb(mm00607939_s1);gapdh(mm99999915_g1);和gapdh(hs03929097_g1)。使用7900ht快速实时pcr系统(abi)以下面的热循环条件运行rt-pcr反应:保持步骤于48℃30分钟,继以保持步骤于95℃10分钟,和40个循环的于95℃10秒钟和于60℃1分钟。将值针对肌动蛋白转录物水平标准化,然后针对对照标准化,如图例中描述的。rna测序。使用truseqrna样品制备试剂盒(illumina,ca)制备rna-seq文库。在illuminahiseq2500测序仪上对文库测序以获得每份样品平均3400万单端读数(50bp)。首先将rnaseq读数与核糖体rna序列比对以去除核糖体读数。使用gsnap(版本2013-10-10)(wu,t.d.&nacu,s.fastandsnp-tolerantdetectionofcomplexvariantsandsplicinginshortreads.bioinformatics26,873-881(2010))将剩余读数与小鼠参照基因组(ncbibuild38)比对,容许每50个碱基的序列有最多两处错配(参数:‘-m2-n10-b2-i1-n1-w200000-e1--pairmax-rna=200000--clip-overlap’)。转录物注释基于refseq数据库(ncbiannotationrelease104)。为了量化基因表达水平,计算定位至每种refseq基因的外显子的读数的数目。将读数计数依据库容量定标,将分位数标准化并使用“voom”r包(law,c.w.,chen,y.,shi,w.&smyth,g.k.voom:precisionweightsunlocklinearmodelanalysistoolsforrna-seqreadcounts.genomebiology15,r29(2014))计算精确性权重。随后,通过分别在6小时和24小时对比dfz7-21处理的样品与dmso或fz7-21s任一处理的样品,使用“limma”r包(ritchie,m.e.etal.limmapowersdifferentialexpressionanalysesforrna-sequencingandmicroarraystudies.nucleicacidsresearch43,e47-e47(2015))对标准化计数数据实施差异表达分析。另外,如较早所述(srinivasan,k.etal.untanglingthebrain'sneuroinflammatoryandneurodegenerativetranscriptionalresponses.natcommun7,11295(2016))以标准化读数每千碱基基因模型每百万总读数(nrpkm)的形式获得基因表达。收集的rnaseq数据经由ncbi的geneexpressionomnibus在登录号gse94159下可得。基因集分析。我们实施基因表达定量集合分析(qusage)(yaari,g.,bolen,c.r.,thakar,j.&kleinstein,s.h.quantitativesetanalysisforgeneexpression:amethodtoquantifygenesetdifferentialexpressionincludinggene-genecorrelations.nucleicacidsresearch41,e170-e170(2013))以鉴定与wnt抑制有关的相关生物学过程。为了该目的,在6小时或24小时时对比dfz7-21处理的样品与dmso样品。对于每项对比,如kimetal.single-celltranscriptprofilesrevealmultilineagepriminginearlyprogenitorsderivedfromlgr5+intestinalstemcells.cellreports16,2053-2060(2016)中定义的计算与肠生物学有关的选定集合的基因集合活性(即log2构成集合的基因个体的表达的均值差异)。动物研究。将c57bl/6雌性小鼠(8周龄)提交每小时一次腹膜内注射(~150-200μl注射体积,用于五次注射)直至达到指定肽剂量。在实验开始时腹膜内施用对照抗lrp6或抗豚草抗体(15mg/kg)并在6小时之后收获小肠。在90.9mm碳酸氢铵,18.2mm乙酸组氨酸,218.2mm蔗糖,和0.018%聚山梨酯-20和介于7.5和8之间的ph中悬浮dfz7-21肽至20mg/ml(用于160mg/kg注射)或10mg/ml(用于80mg/kg注射)。在20mm乙酸组氨酸,240mm蔗糖,0.02%聚山梨酯-20,ph5.5中重悬浮抗体。在6小时之后,处死小鼠,并收集小肠用于mrna提取。所有牵涉动物的研究均得到genentech的机构动物护理和使用委员会批准且遵守nrc实验动物护理和使用指南。实施例2:选择性结合fzd7crd的肽的鉴定fzd7在广泛的干细胞过程中发挥至关紧要的作用,因为它在多种组织特异性干细胞中上调(vincan&barker(2008)clinical&experimentalmetastasis25,657-663;phesseetal.(2016)cancers8)。在十种哺乳动物卷曲蛋白中,fzd1,2和7(它们属于fzd7亚类)在哺乳动物成体肠隐窝的基部处富集,已知那里存在多能干细胞(mariadasonetal.(20051)gastroenterology128,1081-1088;gregorieffetal.(2005)gastroenterology129,626-638)。最近显示fzd7在lgr5+肠干细胞(isc)中富集,而且是伽马照射之后干细胞介导的肠上皮再生所需要的,暗示它是负责介导肠干细胞中的wnt活性的至关紧要fzd受体(flanaganetal.(2015)stemcellreports4,759-767)。fzd7基因敲除实验确立fzd7对于将人胚胎干细胞维持于它们的未分化状态也是必要的(fernandezetal.(2014)proceedingsofthenationalacademyofsciences111,1409-1414,doi:10.1073/pnas.1323697111)。此外,最近的数据证明fzd7在结肠,胰腺和胃肿瘤的子集中上调(vincanetal.(2008)clinical&experimentalmetastasis25,657-663;phesse(2016)cancers8)。另外,fzd7已经牵涉黑素瘤的肿瘤启动和转移性生长以及它们对braf抑制剂的药物抗性(tiwary&xu(2016)plosone11,e0147638;anastasetal.(2014)thejournalofclinicalinvestigation124,2877-2890)。这些发现突显fzd7受体是干细胞的至关紧要的调节物和与干细胞功能障碍有关的疾病的有吸引力的药理学靶物。wnt信号传导在分泌型wnt糖蛋白与fzd受体相互作用后在细胞表面处启动。wnt蛋白质上存在的共价连接的顺式不饱和脂肪酰基基团(棕榈油酸酯)结合fzd受体的细胞外n端crd内的脂质结合沟(jandaetal.(2012)“structuralbasisofwntrecognitionbyfrizzled.”science337,59-64;nileandhannoush(2016)“fattyacylationofwntproteins.”naturechemicalbiology12,60-69),导致核β-联蛋白的稳定化和下游wnt信号传导的启动。在结肠直肠癌中,大多数wnt途径突变发生于β-联蛋白的水平(polakis,p.(2012)coldspringharbperspectbiol4)。然而,wnt途径的异常激活也能发生于受体的水平,正如已经在胆管癌中观察到的(boulteretal.(2015)j.clin.invest.125,1269-1285,lid-1210.1172/jci76452[doi]lid-76452[pii]),而且由途径中的上游突变驱动。例如,肿瘤抑制基因e3泛素连接酶znrf3和rnf43(它们是结肠直肠和子宫内膜癌中频繁突变的基因)(giannakisetal.(2014)natgenet46,1264-1266)中的灭活性突变导致细胞表面处卷曲蛋白受体的稳定化和更高水平(jiangetal.(2015)molecularcell58,522-533)。用全局wnt棕榈酰化和分泌的小分子抑制剂c59处理阻止小鼠中rnf43-/-(ring指蛋白43缺陷的)和znf3-/-(锌/ring指蛋白3缺陷的)肠瘤形成的生长,突显上游wnt信号传导在这些肿瘤中的作用(kooetal.(2015)proceedingsofthenationalacademyofsciences112,7548-7550)。值得一提的是在用c59处理后没有观察到对邻近肠隐窝的显著不利影响。虽然这些发现提示wnt/fzd相互作用的上游抑制可能在高度增殖的干细胞隔室诸如肠中得到耐受,但是在未来的研究中详细评估wnt途径抑制的一般性毒性副作用仍然是重要的。总之,上述发现突显fzd在癌症中的生物学作用且指示药理学靶向fzd受体可能是一种有吸引力的用于在上游在细胞表面处抑制wnt途径的办法。对fzd受体蛋白质家族的分子了解限于人(h)fzd4,5和7crd以及小鼠(m)fzd8crd的x射线晶体结构。虽然报告了第一种与mfzd8crd复合的爪蟾(xenopus)wnt8的x射线共晶体结构,但是wnt-fzd相互作用的特异性的结构基础(dijksterhuisetal.(2015)jbiolchem290,6789-6798)仍然了解较差。尽管有wnt-fzd相互作用的较好结构了解,然而开发在脂质结合沟处结合且破坏fzd依赖性信号传导的高亲和力小配体是有挑战性的。由于高度的序列相似性,选择性靶向特定fzd受体也是一项挑战。见图1;leeetal.(2015)jbiolchem290,30596-30606;gurneyetal.(2012)procnatlacadsciusa109,11717-11722。图1显示与小鼠(m)fzd8crd(表面展示)复合的爪蟾(x)wnt8(飘带展示;用箭指出脂肪酸)的晶体结构。xwnt8在两个不同位点处与mfzd8crd相互作用。位点一(“拇指”)主要是wnt脂肪酰基基团和mfzd8crd上的疏水沟之间的脂质–蛋白质界面,而位点二(“食指”)由蛋白质-蛋白质相互作用界面构成。mfzd8crd作为单体与xwnt8复合结晶。对所有人fzdcrd的clustalomega(sievers,f.etal.fast,scalablegenerationofhigh-qualityproteinmultiplesequencealignmentsusingclustalomega.mol.sys.biol.7,539(2011))序列比对关于它们的百分比保守性打分,作为颜色梯度应用在作为替代结构的mfzd8crd的表面上(深灰色,高保守性;浅灰色,低保守性)。糖基化基团为清楚起见而隐藏。为了调查fzd7在肠干细胞功能中的作用,实施实验来鉴定选择性结合fzd7crd的配体。经由噬菌体展示针对fzd7crd-fc筛选幼稚线性肽文库(长度为4-16个氨基酸)和环状肽文库。然后经由点噬菌体elisa针对fzd7crd-fc,fzd8crd-fc,和herceptin筛选发现结合fzd7crd-fc的肽以鉴定结合fzd7crd但不结合fzd8crd或herceptin的候选者。候选fzd7crd结合性线性肽的氨基酸序列在下文表3中显示,而候选fzd7crd结合性环状肽的氨基酸序列在表4中显示。表3a“n”指每种肽的发生。b对于“信:噪比”,“信(号)”是针对fzd7crd,herceptin,或fzd8crd检测的点噬菌体elisa信号。“噪(声)”是针对bsa的elisa信号。表4a标有下划线的半胱氨酸残基可形成二硫键。b“n”指每种肽的发生。c对于“信:噪比”,“信(号)”是针对fzd7crd,herceptin,或fzd8crd检测的点噬菌体elisa信号,而“噪(声)”是针对bsa的elisa信号。选择五种肽(fz7-06,fz7-07,fz7-17,fz7-20,和fz7-21)进行化学合成和进一步表征。在fz7-21上实施鸟枪丙氨酸扫描以鉴定肽中对结合fzd7crd至关紧要的氨基酸残基。简言之,生成六十种fz7-21衍生肽(见表5),每种含有至少一处随机丙氨酸和/或缬氨酸和/或天冬氨酸和/或丝氨酸替代,并经由点噬菌体elisa(即如上文描述的)针对fzd7crd-fc和herceptin筛选。如表5中显示的,大多数丙氨酸,天冬氨酸,丝氨酸或缬氨酸替代的fz7-21衍生肽展现fzd7crd结合活性。表5a“s/w”值是信号减噪声,其中“信号”指针对在384孔maxisorp板上固定化的fzd8crd-fc或herceptin检测的点噬菌体elisa信号,且其中“噪声”指针对相同板上的bsa的elisa信号。b“s/n”值是信噪比。c“n”指每种肽的发生。接下来,为肽fz7-21中的每个氨基酸位置确定“w/a”值。通过将在特定酸位置具有野生型(wt)残基的fz7-21衍生肽的数目除以在该位置具有替代的fz7-21衍生肽的数目来计算“w/a”。(如果在某个氨基酸位置处没有找到替代,那么a=1。)自60种肽变体收集数据。“w/a”值代表每个野生型fz7-21残基对于与fzd7crd的相互作用的肽结合依赖性。在fz7-21对fzd7crd的结合中发挥至关紧要作用的残基定义为具有>25的w/a且归类为“2类”残基。在fz7-21对fzd7crd的结合中发挥重要作用的残基定义为具有介于6和19之间的w/a值(即5<w/a<25)且归类为“1类”残基。不必要的残基定义为具有<5的w/a且归类为“0类”残基。如下文表6中显示的,fz7-21中的d5,l6,w9,c10,m13,和y14确定为对于对fzd7crd的结合是至关紧要的;fz7-21中的d4,h11,和v12确定为对于对fzd7crd的结合是重要的;而fz7-21中的l1,p2,s3,e7,和f8发现是不必要的。表6合成fz7-21肽的含有d5n替代的衍生物(即fz7-21n)以探查fz7-21中的氨基酸位置5处的带电荷的天冬氨酸残基对fzd7crd结合的贡献。在一系列基于细胞的测定法中表征肽的功能活性。在一组测定法中,在hek293-tb细胞中测试fz7-06,fz7-07,fz7-17,fz7-20,fz7-21,和fz7-21n对wnt信号传导的影响。简言之,在fz7-06,fz7-07,fz7-17,fz7-20,fz7-21,或fz7-21n的3倍连续稀释液中将hek293-tb细胞(即已经用tcf/lef响应性萤火虫萤光素酶报告物构建物稳定转染且组成性表达海肾萤光素酶构建物的细胞)用重组mwnt3a(50ng/ml)刺激6小时。平行实施dmso对照。作为针对dmso对照标准化的萤火虫萤光素酶对海肾萤光素酶的比测量β-联蛋白信号传导。结果在表7中显示。表7中的值是至少3份独立复制品的平均值±标准偏差。最有力的肽fz7-21(seqidno:13)以介于约90-100nm之间的ic50削弱hek293细胞中wnt3a介导的β-联蛋白信号传导(见表7)。fz7-21n削弱用外源wnt3a刺激的hek293细胞中wnt3a介导的β-联蛋白信号传导(ic50=100nm)。在一组互补实验中,fz7-06,fz7-07,fz7-17,fz7-20,fz7-21,和fz7-21n对小鼠l细胞中wnt介导的β-联蛋白蛋白质的稳定化的影响(如hannoush(2008)plosone3,e3498中描述的)。将l细胞用wnt3a(2.5nm)且在fz7-06,fz7-07,fz7-17,fz7-20,fz7-21,或fz7-21n的3倍连续稀释液中处理5小时。平行实施dmso对照。在处理后,将l细胞固定,透化,用荧光标记的抗β-联蛋白抗体探查,并经由细胞中western测定法(即如hannoush,r.n.(2008)plosone3,e3498中描述的)观察以评估β-联蛋白蛋白质水平。结果在表7中显示。表7中的值是至少3份独立复制品的平均值+标准偏差。fz7-21阻断小鼠l细胞中wnt3a介导的β-联蛋白的稳定化,指示这种肽抑制wnt信号传导。fz7-21n阻断小鼠l细胞中wnt3a介导的β-联蛋白的稳定化,指示这种肽抑制wnt信号传导。表7值代表均值±标准偏差。>100μm指示肽在所测试的肽的最高浓度(100μm)显示部分抑制a通过来自含有tcf的β-联蛋白区的topbrite报告物的读出测量b通过经由免疫荧光检测的β-联蛋白的量测量c以100μm肽测量d值是至少2份复制品的平均值;±标准偏差e值是至少3份复制品的平均值;±标准偏差fβ-联蛋白活性在11.11μm肽达到~50%抑制的平台g以11.11μm肽测量不受理论束缚,在hek293和小鼠l细胞之间在最大wnt途径抑制中观察到的差异(如分别通过topbrite萤光素酶报告物和β-联蛋白成像测定法测量的)很可能是由于两种细胞系之间各种fzd受体的细胞表面表达水平的差异(zhouetal.(2014)dev.cell31,248-256,lid-210.1016/j.devcel.2014.1008.1018[doi]lid-s1534-5807(1014)00548-00546[pii])。合成fz7-21s(即fz7-21的含有c10s替代的衍生物)以探查fz7-21中的氨基酸位置10处的半胱氨酸残基对fzd7crd结合的贡献。fz7-21s的氨基酸序列是lpsddlefwshvmy(seqidno:113)。在第一种测定法中,将hek293-tb细胞用重组mwnt3a(50ng/ml)在dmso对照,fz7-21s,或fz7-21的3倍连续稀释液中刺激6小时。如图2a中显示的,fz7-21以93.7nm±27.9nm的ic50抑制wnt3a刺激的β-联蛋白信号传导。没有发现fz7-21s抑制wnt3a信号传导。在后续的一项测定法中,用5ngpcdna3.2-wnt3a或25ngpcdna3.2-wnt3a转染hek293-tb细胞。24小时后,将转染细胞用dmso,fz7-21s,或fz7-21的3倍连续稀释液处理6小时。如图2b中显示的,fz7-21以用5ngpcdna3.2-wnt3a转染的细胞中329.8nm±157nm的ic50和用25ngpcdna3.2-wnt3a转染的细胞中419.3nm±168.6nm的ic50抑制wnt3a刺激的β-联蛋白信号传导。没有发现fz7-21s抑制wnt3a信号传导。在又一项测定法中,用5ngpcdna3.2-wnt1或25ngpcdna3.2-wnt1转染hek293-tb细胞。24小时后,将转染细胞用dmso,fz7-21s,或fz7-21的3倍连续稀释液处理6小时。如图2c中显示的,fz7-21以用5ngpcdna3.2-wnt3a转染的细胞中1.0864μm±0.5077μm的ic50和用25ngpcdna3.2-wnt3a转染的细胞中2.661μm±1.124μm的ic50抑制wnt1刺激的β-联蛋白信号传导。没有发现fz7-21s抑制wnt1信号传导。接下来,用5ngpcdna3.2-wnt1或25ngpcdna3.2-wnt3a转染hek293-tb细胞。24小时后,将转染细胞用fz7-21c(即含有位置10处的d-半胱氨酸立体异构体的fz7-21衍生肽)的3倍连续稀释液处理6小时。如图2d中显示的,fz7-21的位置10处d-半胱氨酸对l-半胱氨酸的替代将wnt1抑制的效力降低16倍,且将wnt3a抑制的效力降低31倍。总之,图2a-2d中显示的结果突显cys10残基对fzd7crd结合的功能贡献。在用6-bio(6-溴靛玉红-3'-肟)处理的hek293-tb细胞中进行一项上位性(epistasis)研究,6-bio是稳定β-联蛋白且不依赖受体活化而激活下游wnt信号传导的gsk-3α/β的一种抑制剂(meijeretal.(2003)chemistry&biology10,1255-1266)。将hek293-tb细胞在dmso,fz7-21s,或fz7-21的3倍连续稀释液存在下用10μm6-bio刺激6小时。fz7-21或fz7-21s均不抑制用6-bio处理的细胞中的wnt信号传导(见图2e)。此类结果指示fz7-21作用于gsk3α和β-联蛋白的上游,很可能在fzd受体的水平。在图2a-2e中,作为针对dmso对照标准化的萤火虫对海肾的比测量β-联蛋白信号传导。将计算值减去背景(未用wnt3a刺激的细胞),针对用dmso处理的样品标准化,并代表至少三项独立实验(具有技术上的一式三份)的均值±s.e.m。使用graphpadprism(v6.05)使用log(抑制剂)对标准化响应生成抑制曲线[可变斜率方程(y=100/(1+10^((logic50-x)*坡斜率))]。ic50代表均值±95%置信区间。所有样品维持1%dmso终浓度。使用holm-sidak方法确定统计学显著性,以α=5%,其中p*<1.17*10-6。显著性假设所有样品来自具有相同分散的群体。接下来,生成5-羧基荧光素标记型式的fz7-21肽,5fam-fz7-21,并测试对hfzd1crd-fc,mfzd2crd-fc,hfzd4crd-fc,hfzd5crd-fc,hfzd7crd-fc,mfzd7crd-fc,hfzd8crd-fc,mfzd9crd-fc,和hfzd10crd-fc的结合。平行实施使用5fam-fz7-21s的对照实验。将5fam-fz7-21或5fam-fz7-21s与每一种上述fzdcrd-fc一起在pbs中于4℃温育过夜并经由sec解析。5fam-fz7-21显示对hfzd1crd-fc,hfzd2crd-fc,hfzd7crd-fc,和mfzd7crd-fc的选择性的且优先的结合,亲和力在低nm范围中(39-116nm)。见表8和图3a-3i。在图3a-3i中,将5fam-fz7-21或5fam-fz7-21s(50nm,在pbs中)与递增浓度的fzdcrd-fc(2倍连续稀释液)一起温育,并使用monolithnt.115或nt.115pico仪器(nanotempertechnologies)测量荧光强度。ec50值代表至少三项独立实验的95%置信区间,hfzd1crd-fc+5fam-fz7-21s除外,它进行两次。用prismgraphpad使用log(激动剂)对响应计算ec50值[可变斜率(四参数)函数:y=底+(顶-底)/(1+10^((logec-x)*坡斜率))]。绘图的值代表均值±s.e.m。表8anc=无变化与fz7-21特异性结合fzd7亚类之crd的假说一致,观察到5fam-fz7-21很少结合至不结合fzd5,fzd8,fzd9和fzd10crd。对照肽5fam-fz7-21s不显示可检测的对fzdcrd家族的任何成员的结合。见图3a-3i和表8。还通过层析术观察fzd7受体亚类的选择性靶向。5fam-fz7-21与fzd1crd-fc,fzd2crd-fc,和fzd7crd-fc的复合物能通过荧光大小排阻层析术(fsec)来解析。与之对比,对5fam-fz7-21s没有观察到复合物。见图4a和4b,其分别提供在通过fsec的分析前与各种fzdcrd-fc蛋白质(250nm)一起于4℃温育过夜的1μm5fam-fz7-21和1μm5fam-fz7-21s的代表性荧光痕迹。垂直点线代表分子量标准物的洗脱体积。标示了fzd7crd对肽的摩尔化学计量。图4a和4b的荧光强度的量化(曲线下面积,auc)在图4c中提供。图4a-4c中的结果指示5fam-fz7-21优先结合fzd7类crd,而5fam-fz7-21s不结合任何fzdcrd-fc。在这些实验中,fzdcrd-fc融合构建物显示与包含通过两个fc片段保持在一起的两个fzdcrd二聚体的四聚体的形成一致的保留概况。在图24a中,将通过uv吸收分析的分子量(mw)标准物作为洗脱体积(ve)在空隙体积(vo)上的函数绘图。数值代表三项独立实验的均值±s.e.m.。图24b显示与5fam-fz7-21结合的fzdcrd-fc蛋白质的观察分子量(灰色圆形)对预测fzdcrd-fc四聚体mw(黑色方形)。测量值代表三项独立实验的均值±标准偏差(sd)。图24c显示所使用的不同fzdcrd-fc蛋白质(~2μg)的天然page(4-16%)。使用nativmark作为分子量标准物。将蛋白质样品用天然page样品缓冲液稀释,使用深蓝色阴极运行缓冲液分开,解析,并固定,依照制造商的推荐进行。对重组fzd3和6crd的结合由于表达这些蛋白质的挑战而无法评估。然而,通过流式细胞术分析,5fam-fz7-21显示对fzd7的优势结合,以及对在hek293细胞中稳定表达的fzd4,5或6受体的最小限度结合,与上文讨论的层析实验一致。实施进一步的fsec实验以测定5fam-fz7-21或5fam-fz7-21s是否结合人sfrp1,sfrp2,sfrp3,sfrp4,或sfrp5(即分泌型卷曲蛋白相关蛋白,在结构上与fzd蛋白质有关的可溶性蛋白质的一个家族)。没有发现5fam-fz7-21或5fam-fz7-21s结合测试的任何sfrp。见例如图5a-5c。垂直点线代表分子量标准物的洗脱体积。对于图4c和5c,数值代表三项独立实验的均值±s.e.m.。图4c和5c的曲线下面积(auc)使用unicorn(v5.31;generalelectricbio-sciences)积分并代表来自三项独立实验的均值±s.e.m.。在磷酸盐缓冲盐水(pbs)中以0.5ml/min使用sec3000柱(phenomenex;torrence,ca)解析样品并用fp-2020plus荧光检测仪(jascoanalyticalinstruments,easton,md)使用正常模式(激发/发射550/494nm;增益=100;std=32)监测荧光。所有溶液维持1%最终dmso。将线迹向上调整n+20个单位以能够在痕迹间区分。在样品运行前通过标准物(biorad目录号151-1901)运行来测定分子量。实施进一步的实验以检查肽对可溶性单体形式的hfzd7crd的结合。使用经endoh和几夫碱处理的重组hfzd7crd-his实施大小排阻层析术(sec),它是自杆状病毒生成,用ni-nta琼脂糖树脂捕获,浓缩,并使用gehealthcarehiload16/60superdex75pg以0.5ml/min用150mmnacl,50mmtris-hcl,ph7.5解析。如图6a中显示的,hfzd7crd作为两个主峰洗脱,~16kda处的单体和更高分子量多聚体。“vo”=空隙体积。图6b显示来自图6a的合并的单体的sds-page。在通过sds-page的分析前,用补充有1mmdtt的样品缓冲液处理样品并加热至98℃达10分钟。m,标志物(10μl,见蓝色plus2预染色蛋白质标准物)。为了进一步了解fz7-21的作用机制,将hfzd7crd-his(15.8μm)与(a)fz7-21(以1:1,1:5,或1:10的比率),(b)fz7-21s(以1:10的比率),或(c)dmso一起温育2小时,之后经由大小排阻层析术(sec)解析和经由多角度光散射(mals)和λ280处的吸光度检测。通过superdexs2003.2/300柱(generalelectrichealthcarelifescience,pittsburgh,pa;28990946)以0.15ml/min在含1%dmso终浓度的150mmnacl,50mmtris-hcl,ph7.5缓冲液中解析样品。将柱与1260infinityhplc(agilenttechnology,santaclara,ca)连接,后者与dawnheleos-ii多角度静态光散射(mals)检测仪和optilabt-rex差示折射率(dri)检测仪(wyatttechnologies,santabarbara,california)连接。sec-mals分析的结果在图6c中显示。图6d显示图6c的放大视图(1.5ml至2.0ml范围),测试另外的肽浓度。mals信号的量化和相对于dmso对照的分子量变化(δmw)在表9中显示。与fz7-21一起温育促进ve=1.6ml附近的多聚体的形成。见图6d。表9蛋白质处理mw(kda)不确定性(%)δmw(kda)bsadmso70.10.3n/ahfzd7crddmso20.91.40hfzd7crdfz7-21(1:1)21.91.61hfzd7crdfz7-21(1:5)27.316.4hfzd7crdfz7-21(1:10)321.411.1hfzd7crdfz7-21s(1:10)23.51.32.6此类结果证明肽fz7-21结合单体hfzd7cdr-his。见表9。出乎意料的是,fz7-21以浓度依赖性方式诱导hfzd7crd-his同二聚化或寡聚化。见表9。总之,此类发现证明肽fz7-21展示对fzd7受体亚类的结合选择性。此类结果还指示fz7-21诱导单体fzd7crd二聚化,而且fz7-21结合预形成的四聚体fzd7crd-fc。见图6a-6d。上文讨论的结果证明fz7-21选择性结合fzd7亚类的进化上保守的蛋白质,即fzd1,fzd2,和fzd7。见图7。鉴于fzd蛋白质之间的高度序列相似性,此类结合选择性是出乎意料的。在图7中,手工修剪蛋白质crd并用clustalomega比对。图7中的分支图是使用phylip邻接及jones-taylor-thorton距离矩阵和100次自举迭代和转换/颠换比2.0生成的。在图7中的分支图的右边是5fam-fz7-21或5fam-fz7-21s对每一种蛋白质的结合偏好的汇总。“*”指示测试小鼠蛋白质。“**”指示测试小鼠和人蛋白质二者。“sfrp”指“分泌型卷曲蛋白相关蛋白”。使用下面的登录号作为源序列:hfzd1(q9up38);hfzd2(q14332);hfzd3(q9npg1);hfzd4(q9ulv1);hfzd5(q13467);hfzd6(o60353);hfzd7(o75084);hfzd8(q9h461);hfzd9(o00144);hfzd10(q9ulw2)。实施例3:fzd7crd和fz7-21之间的相互作用的结构表征最近报告了处于它的脱辅基形式或与c24脂肪酸复合的hfzd7crd的x射线晶体结构(nileetal.(2017)proc.natl.acad.sci.usa114,4147-4152,lid-4110.1073/pnas.1618293114[doi])。在两种结构中,hfzd7crd包含具有α-螺旋二聚体界面和桥接二聚体界面的u形脂质结合腔的二聚体(nileetal.(2017)proc.natl.acad.sci.usa114,4147-4152,lid-4110.1073/pnas.1618293114[doi])。为了获得对肽-fzdcrd选择性的机制的了解,实施进一步的实验以在分子水平表征肽-蛋白质相互作用。致力于获得与fz7-21复合的hfzd7crd的共晶体结构的广泛努力(包括共结晶和浸泡技术)没有成功。使用一种构建物,其中fz7-21的n端与hfzd7crd的c端融合,侧翼有接头。fzd7crd(脱辅基形式)和与肽fz7-21结合的hfzd7crd的x射线晶体结构是分别以和分辨率测定的(见下文图8a-8f和表10)。脱辅基fzd7crd的结构揭示具有独特构造的二聚体。如图8a-8f中显示的,fzd7crd的脂质结合腔的结构令人惊讶地不同于报告的其它fzdcrd家族成员的脂质结合腔的结构。fzd7crd二聚体中存在两个脂质结合沟。每个fzd7crd单体的尾部在二聚体界面处彼此面对。见图8a-8b。这创建桥接二聚体界面的连续且弯曲(u形)的脂质结合腔。见图8b。接近疏水腔的残基在图8b中以球棍形式展示。图8c显示与fz7-21结合的hfzd7crd的晶体结构。图8d显示定位到与fz7-21(飘带展示)结合的hfzd7crd(飘带展示)的结构上的疏水腔的表面展示。来自疏水腔的残基以球棍展示显示。图8e显示与fz7-21结合的hfzd7crd的晶体结构的顶视图表面展示(飘带展示;显示了二硫化物)。图8f显示与fz7-21结合的hfzd7crd的晶体结构的侧视图表面展示(飘带展示;显示了二硫化物)。fz7-21的结合表位(定义在距离内)在圆圈内。脂质结合腔以箭指示。对于所有结构,聚糖模块为清楚起见而隐藏。表10:脱辅基hfzd7crd和fz7-21结合的hfzd7crd的x射线晶体学数据加工和结构精修统计量1括弧中的值是最高分辨率壳的。2r对称=σ|ihi-ih|/σihi,其中ihi是反射h的第i个对称相关观察的定标强度而ih是均值。3r结晶=σh|foh-fch|/σhfoh,其中foh和fch是反射h的观察和计算结构因子振幅。4r游离的值是为5%随机选择的,精修中不包括的反射计算的。5c-核心;a-另外容许的;g-一般容许的;d-不容许的。这种排列与脂质结合沟在其它fzdcrd(诸如hfzd4(见图9a)和mfzd8(见图9b)的那些)上的定位形成鲜明对比,它们位于远离二聚体界面的相对侧,而且由此彼此分开(参见例如jandaetal.(2012)science337,59-64;dannetal.(2001)nature412,86-90;和shenetal.(2015)cellres.25,1078-1081)。hfzd4(见图9a)和mfzd8(见图9b)的脂质结合腔依照氨基酸疏水性在梯度上着阴影(深灰色=亲水性的;浅灰色=疏水性的)。而且,hfzd4crd和mfzd8crd的脂质结合沟与hfzd7crd的不同还在于它们是溶剂暴露的且展示‘延伸’构象。图9c提供脱辅基hfzd7crd与mfzd8crd和hfzd4crd的叠加,突显不同的二聚体界面。致力于获得与fz7-21复合的hfzd7crd的共晶体结构的广泛努力(包括共结晶和浸泡技术)没有成功。设计一种称作hfzd7crd-gs的构建物,其中fz7-21的n端与hfzd7crd的c端融合,侧翼有接头(图10a和10b和表10)。在昆虫细胞中表达融合构建物,并通过大小排阻层析术(sec)纯化至接近同质。图10b显示纯化的hfzd7crd-gs的sec概况,具有42.7kda的测定分子量(mw),对应于二聚体。图27a显示用于测定hfzd7crd-gs的mw的相关mw标准物。图27b显示图10a中的融合蛋白在还原性条件下的sds-page,对应于单体。图27c显示自图10a中的融合蛋白获得的晶体的明视野图像。融合构建物的晶体结构揭示肽fz7-21作为二聚体接近脂质结合沟结合,在hfzd7crd的二聚体界面处与脂质结合腔衬里的残基进行接触(图8c-8f和表11和12)。肽二聚体包含两个以相对于彼此~45°角取向的反平行α螺旋且通过cys10处的一个二硫键和众多主链和侧链相互作用保持在一起,围绕二硫键形成溶剂保护的核心(图8c-8f和图11a-11c)。不对称单元包含四个结构上相似的crd二聚体对,每个通过fz7-21二聚体肽来桥接。接头区由于缺乏电子密度而不能解析。肽结合的hfzd7crd的构造显示独特的构象。脂质结合腔的几何学自在二聚体界面处显示~16°角的弯曲形式(即处于脱辅基结构)改变成在二聚体界面处显示90°的延伸形式(即处于肽结合的形式)。如此,fz7-21二聚体对hfzd7crd的脂质结合腔的结合将二聚体界面置换~75°(图8a-8d,图12a和12b和图25a-25f)。图25a显示脱辅基hfzd7crd晶体结构的飘带展示(彩虹着色;n端,蓝色;c端,红色),全长fzd7的示意图图示fzd7内的crd放置,而图25b显示与fz7-21结合的hfzd7crd的结构的飘带展示(彩虹着色)。在脱辅基和结合结构二者中,在图25a和25b中显示二聚体界面处的选定配对残基。图25c提供脱辅基hfzd7crd中的疏水腔的放大侧视图,而图25d提供来自图25c的顶视图。图25e提供与fz7-21结合的hfzd7crd的放大侧视图,其中突显疏水腔且为了清楚起见而隐藏蛋白质主链,有180°旋转,而图25f显示图25e的顶视图。图25c-25e中的疏水腔描绘为深灰色,疏水性的;白色,中性的;蓝色,浅灰色。总之,构成fzd7crdα螺旋二聚体界面的残基的数目自脱辅基结构中的13个(p55,e77,g80,l81,h84,q85,y87,p88,k91,v92,l134,f138(贯穿说明书或者鉴定为f130),和f140)每个单体减少至肽结合结构中的5个(p88,k91,v92,k137和f138(贯穿说明书或者鉴定为f130))每个单体。见图14。在这种结合构造中,肽α螺旋二聚体在延伸的脂质结合沟的顶部形成“盖”,而且来自每个螺旋的结合残基与两个hfzd7crd单体上的残基进行接触(图8c-8f和下文表11和12)。如图13中显示的,fz7-21(b链)上的leu6与hfzd7crd的phe138(贯穿说明书或者鉴定为phe130)和phe140(a链)进行关键的疏水性接触,而fz7-21(b链)上的asp5的主链羰基与hfzd7crd的his84(b链)的侧链形成氢键。在图13中,在与fz7-21(飘带和棍展示)结合的hfzd7crd的晶体结构内突显选定的fz7-21–hfzd7crd相互作用。点线代表氢键相互作用。而且,fz7-21上的trp9的吲哚氮与hfzd7crd的gln85的主链羰基形成氢键。见图13。总之,fz7-21上有16个残基(8个残基每个单体)牵涉与hfzd7crd的相互作用,主要在脂质结合沟的表面衬里的α螺旋二聚体界面处(见表12)。表11:距fz7-21在内的fzd7crd二聚体残基*phe138贯穿说明书或者鉴定为phe130表12:距hfzd7crd二聚体在内的fz7-21残基部分基于fzdcrd家族内的肽结合表位的序列保守性,这些相互作用提供对fzd7受体亚类的肽选择性的基本原理。见图14,其提供hfzd7,hfzd1,hfzd2,mfzd8,hfzd5,hfzd4,hfzd9,hfzd10,hfzd3,和hfzd6的crd的氨基酸序列的比对。在图14中,fz7-21结合表位,和处于它的脱辅基形式的或与fz7-21复合的(完整fzd7界面;a链对b链)hfzd7crd(a链对b链)的α螺旋二聚体界面中的残基通过序列比对上方的“+”来表示。保守的半胱氨酸以灰色突显而保守的表位残基标有下划线。另外,hfzd7crd内的二聚体界面和脂质结合沟几何学有助于肽结合足迹且因此可能影响肽选择性。有助于稳定二聚体间肽螺旋的主链和侧链相互作用在表13中显示。表13:fz7-21分子内相互作用的汇总脱辅基fzd7crd的晶体结构揭示出乎意料的关于这个fzd亚类内的脂质结合腔的独特几何学的分子照片,这可能对于为什么wnt蛋白质上的顺式不饱和脂肪酸可能优先进化成结合hfzdcrd的脂质结合腔提供一种解释。值得注意的是,肽结合的fzd7crd的构造显示独特的构象,其中脂质结合腔的几何学自弯曲的u形(在脱辅基结构中,在图12a中显示)改变成延伸形式,偶联二聚体界面实质性置换75°以形成~90°角。脱辅基形式和fz7-21结合形式的fzd7crd的二聚体界面处的氨基酸残基之间的距离在下文表14中提供。表14:残基间距离自晶体学模型观察到的相互作用得到上文描述的数项功能分析的支持。第一,鸟枪丙氨酸扫描鉴定出fz7-21上的数个残基,即asp5,leu6,trp9,cys10,met13和tyr14,它们对于与hfzd7crd-fc的相互作用是重要的(见上文表6),与晶体结构一致(见图13)。第二,来自晶体结构的观察结果提示二聚体形式的fz7-21会展现改善的活性。dfz7-21(即合成获得的二聚体形式的fz7经由cys10处的二硫键二聚化fz7-21)在抑制hek293细胞中的wnt3a信号传导方面展现与单体fz7-21相比的~40倍改善(图15),如通过用重组mwnt3a(50ng/ml)刺激的hek293-tb细胞中的topbrite报告物测定法测量的。(图15中的值代表指定肽相对于dfz7-21的ic50倍数变化。为了考虑表达水平,将萤火虫发光信号针对海肾萤光素酶发光标准化。将计算值减去背景,针对模拟处理样品标准化,并代表至少三项独立实验的均值,每项具有技术上的一式三份。)另外,dfz7-21内预测破坏它与hfzd7crd的相互作用的多个位置处的丙氨酸点突变(例如l6a)导致细胞效力降低~800倍(见图15和表15)。与之对比,n端处的截短并不干扰dfz7-21活性(例如缺乏2个n端残基的肽dfz7-21-δ2),与晶体结构和鸟枪丙氨酸扫描数据一致(见表6,表15,和图15)。作为阴性对照平行测试fz7-21s。表15dfz7-21是合成获得的二聚体形式的fz7,经由cys10处的二硫键二聚化fz7-21为了获得图15和表15中的数据,通过用重组mwnt3a(50ng/ml)刺激的hek293-tb细胞中的topbrite报告物测定法测量wnt信号传导。图15和表15中的值代表指定肽相对于dfz7-21的ic50倍数变化。为了考虑表达水平,将萤火虫发光信号针对海肾萤光素酶发光标准化。将计算值减去背景,针对模拟处理样品标准化,并代表至少三项独立实验的均值,每项具有技术上的一式三份。进一步地,在hfzd7crd上的肽结合区内的特定残基处引入的突变(h84a,y87a,f138a(贯穿说明书或者鉴定为f130a),f140a)与野生型hfzd7crd相比降低fz7-21的结合(见图40),与结构预测一致。而且,fz7-21促进溶液中hfzd7crd的单体至二聚体转换(数据未显示),与描绘肽-crd结合的2:2化学计量的结构模型一致。第五,与含有肽序列内cys10至ser突变的融合构建物(其显示具有不太明显的二聚体种类的单体和寡聚体的混合群体(数据未显示))相比,fzd7crd-fz7-21融合构建物在溶液中主要是二聚体(数据未显示)。预测对于肽-肽或肽-fzd7相互作用重要的fz7-21残基的另外的ala突变(即l6a;w9a;y14a;l6a和w9a;l6a和y14a;和l6a,w9a,和y14a)导致fzd7crd–fz7-21融合构建物的二聚化降低(数据未显示)。这些发现与上文sec-mals数据和晶体学观察结果一致,进一步支持fz7-21的活性形式是增强fzd7crd二聚体形成的二聚体的观念。dfz7-21的nmr溶液结构揭示肽二聚体是结构化的,展现分子内相互作用和α螺旋特征(见图16a-16e和表16),与来自晶体结构的观察结果相似。图16a提供dfz7-21的noesy连通图。图26显示dfz7-21的20种最低能量nmr结构的叠加(a链和b链,飘带展示;侧链,线展示)。图16b显示dfz7-21的一种代表性nmr溶液结构,其基于dfz7-21的20种最低能量nmr结构的叠加(氨基酸侧链作为线显示)。在nmr溶液结构中,两条肽链彼此以90°角取向且n端区域看来是无序的,指示与fzd7crd的相互作用可能诱导构象变化且有助于促进肽内的二级结构形成。图16c显示展示α螺旋特征的dfz7-21的2dnoesy图。与之对比,图16d中显示的fz7-21s的2dnoesy图指示fz7-21s具有有限的二级结构。图16e显示fz7-21,fz7-21s和dfz7-21肽的1dnmr谱。在图16e中,fz7-21在测试的缓冲液条件中显示相对于fz7-21s和dfz7-21的峰变宽,使得它的二级结构的指派变得困难。图16e中的箭指示来自二硫化物半胱氨酸的酰胺基团的质子峰。总之,所呈现的结构-活性相关性和nmr数据对结构模型提供进一步的生物化学支持。表16:fz7-21二聚体的nmr统计量如上文讨论的,比对hfzd7,hfzd1,hfzd2,mfzd8,hfzd5,hfzd4,hfzd9,hfzd10,hfzd3,和hfzd6crd的序列以获得fz7-21对fzd7类crd的选择性的进一步了解。见图14。比对是使用clustalomega生成的,继以使用二硫化物连接的半胱氨酸作为指导的手工比对(半胱氨酸以灰色突显)。hfzd7上变成与fz7-21二聚体直接接触的残基和其它fzdcrd上的相同残基以粗体和下划线文本描绘。因为fz7-21只结合fzd7类crd(即fzd7crd,fzd1crd,和fzd2crd),所以图14中显示的结合位点1(即lxxhqxyp)和结合位点2(即fgf)二者的组合很可能是特异性和选择性fzd7类结合所要求的。hfzd7crd二聚体的排列与其它相关卷曲蛋白家族成员,诸如hfzd5和mfzd8crd相似,但是与更加远缘相关fzdcrd家族成员,诸如hfzd4crd形成鲜明对比(jandaetal.(2012)science337,59-64;nileetal.(2017)proc.natl.acad.sci.usa114,4147-4152,lid-4110.1073/pnas.1618293114[doi];dannetal.(2001)nature412,86-90;shenetal.(2015)cellres.25,1078-1081),其中脂质结合沟看来彼此相当分开且位于远离二聚体界面的相对侧(图9b)。然而,最近报告的与c16:1n-7脂肪酸复合的hfzd4crd的结构揭示具有介于在hfzd7crd/c16:1n-7复合物中找到的crd和hfzd7crd/fz7-21复合物中的cdr之间的中间构型的二聚体crd。在肽结合区内的氨基酸序列中的差异(图14)以外,hfzd4crd中二聚体界面与hfzd7crd相比的不同构造可能部分解释肽对hfzd4crd的结合的缺乏。虽然fzd5,7和8crd共享α螺旋二聚体构造(nileetal.(2017)proc.natl.acad.sci.usa114,4147-4152,lid-4110.1073/pnas.1618293114[doi]),但是构成二聚体界面的氨基酸残基中有细微变化。特别地,fzd7crd中的tyr87和phe138(贯穿说明书或者鉴定为f130)在fzd5/8crd中分别变成trp和tyr。这两个残基构成hfzd7crd上的fz7-21肽结合区的部分,而且是肽表位内在fzd7和fzd8类crd之间不保守的唯一残基(在全部七个crd残基中)。虽然残基差异基于计算机预测并不呈现重大氨基酸变化,但是f138y可能造成与fz7-21在leu6处和与fzd7crd二聚体配偶在val92和lys91处的空间不相容性,潜在破坏hfzd7crd的‘延伸’脂质结合沟形式。(f138y贯穿说明书或者鉴定为f130y。)部分基于fzdcrd家族内的肽结合表位的序列保守性,这些相互作用可能提供对fzd7受体亚类的肽选择性的基本原理。另外,hfzd7crd内的二聚体界面和脂质结合沟几何学有助于肽结合足迹且因此可能影响肽选择性,更不要说crd蛋白质动力学。实施例4:fzd7功能的体内表征为了解决fz7-21是否通过破坏wnt与fzd7的相互作用来干扰fzd7介导的信号传导,开发了一种酶联免疫吸附测定法(elisa)来测量在fz7-21,dfz7-21或fz7-21s存在或缺失下生物素化的wnt(bio-wnt)对不同fzdcrd的结合。用fz7-21或dfz7-21处理将bio-wnt3a或bio-wnt5a对fzd7类蛋白质的结合增强~1.5-2倍;然而,它并不显示对相同wnt蛋白质对不与fz7-21相互作用的fzd4的结合的任何影响。而且,正如预期的,对照肽fz7-21s显示对不同wnt对fzdcrd的结合没有影响。不受理论束缚,由dfz7-21对hfzd7crd诱导的实质性构象变化(如通过改变的脂质结合腔的几何学例示的)可能推动在hfzd7crd二聚体上同时容纳两个wnt分子(图4e-h)。分子建模提示肽结合的hfzd7crd结构中的延伸的疏水腔能潜在容纳两个wnt脂肪酰基模块,它们在其它情况中在脱辅基hfzd7crd结构中会是空间上不相容的,为观察到的elisa信号的2倍升高提供貌似合理的解释。总之,实验和建模数据提示dfz7-21能增强wnt3a和wnt5a朝向fzd7类crd而非fzd4crd的募集。在fz7-21-fzd7crd复合物中观察到的剧烈构象变化可能使得受体不能进行正常信号传导,尽管有增强的wnt结合,如通过肽对wnt信号传导的抑制作用证明的。为了在药理学上解决fzd7在干细胞功能中的作用,评估fz7-21处理对自成体小鼠肠上皮建立的类器官培养物的影响(fatehullahetal.(2016)“organoidsasaninvitromodelofhumandevelopmentanddisease.”natcellbiol18,246-254;barker,n.etal.(2007)“identificationofstemcellsinsmallintestineandcolonbymarkergenelgr5.”nature449,1003-1007)。类器官培养系统忠实地再现肠上皮,包括隐窝-绒毛形态的存在。值得注意的是,隐窝区域经历连续出芽,这是功能性肠干细胞(isc)的存在的直接度量。因此,通过对每个类器官形成的芽的数目打分来定量评估类器官群体的干细胞功能(一种wnt依赖性过程)(grabingeretal.(2014)“exvivocultureofintestinalcryptorganoidsasamodelsystemforassessingcelldeathinductioninintestinalepithelialcellsandenteropathy.”celldeath&disease5,e1228)。肠小鼠类器官在matrigel中在生长因子(头蛋白,egf,和r-spondin)和(a)dmso,(b)200μmfz7-21s(即阴性对照),(c)抗lrp6阻断性抗体(即阳性对照),(d)200μm二聚化的dfz7-21,(e)100μm二聚化的dfz7-21,(f)10μm二聚化的dfz7-21,或(g)1μm二聚化的dfz7-21存在下生长48小时。48小时处理后的代表性小鼠肠类器官的形态在图17a-g中显示。接下来,量化肽处理后的类器官干细胞(sc)潜力。类器官sc潜力指示具有≥1个芽每个类器官的类器官的%。如上处理类器官并在处理后48小时收集。如图18中显示的,用dfz7-21处理以浓度依赖性方式剧烈减少出芽事件,而阴性对照肽fz7-21s不显示任何重大影响。(图18中的值代表来自至少三份生物学复制品的均值±s.e.m。在每种条件中打分的类器官的总数:n=554(dmso);n=547(dfz7-21,200μm);n=627(dfz7-21,100μm);n=648(dfz7-21,10μm);n=287(dfz7-21,1μm);n=528(fz7-21s,200μm);n=715(抗lrp6抗体,10μg/ml)。)与对isc潜力的破坏一致,充分表征的isc标志物(clevers,h.(2012)“theintestinalcrypt,aprototypestemcellcompartment.”cell154,274-284)诸如lgr5(图19a)和ascl2(图19b)在用dfz7-21处理24和48小时后显著下调。用fz7-21s处理没有影响。见图19a和19b。类似地,充分表征的wnt靶基因轴蛋白2在dfz7-21处理后显著降低,但是在fz7-21s处理后不然。见图19c。(图19a-19c中的值代表六份生物学复制品的均值±s.e.m,每份具有两个技术复制品。将肽处理样品针对dmso对照标准化。用参数非配对t检验实施统计学,假设两个群体具有相同的sd。)与之对比,muc2在用dfz7-21处理24和48小时后上调但是fz7-21s不然。dfz7-21对isc潜力和干细胞转录物二者的影响与对照抗lrp6抗体(即wnt信号传导的通用抑制剂)相似。在体内进一步调查dfz7-21的药理学作用。简言之,自用dfz7-21,fz7-21s,抗豚草抗体(阴性对照),或抗lrp6抗体(抑制wnt信号传导的阳性对照)处理(即经由腹膜内施用)6小时的c57bl/6小鼠收集肠上皮。在处理后,实施rt-pcr以量化转录物水平。各种肠干细胞(isc)和wnt转录物诸如lgr5(图20a),ascl2(图20b)和轴蛋白2(图20c)在dfz7-21处理后显著下调(图20a-20c中的值代表五只小鼠的均值±s.e.m.,具有技术上的一式两份。用参数未配对t检验实施统计学,假设两个群体具有相同的sd。在使用rout异常值消除方法(其中q=1%)之后去除一个值。将所有值针对抗豚草阴性对照标准化。)在用抗lrp6抗体处理的样品中也观察到这种效果。与之对比,用fz7-21s处理不影响lgr5,ascl2和轴蛋白2转录物水平。见图20a-20c。这些结果与图17a-17g,图18,和图19a-19c中的数据一致。通过对用dfz7-21处理6和24小时的肠类器官样品进行rna测序来进一步调查dfz7-21的药理学作用。无偏移分析揭示用dfz7-21处理后已知在lgr5+isc和保留isc中表达的标志物的时间依赖性方式的实质性下调。同时,dfz7-21处理的类器官中也有肠细胞标志物(促分化)的上调。在dfz7-21处理后下调最多的基因中有lgr5,olfm4和ascl2。在用模拟肽fz7-21s或dmso处理的样品中观察到基因表达的很少变化或无变化。最后,为了评估dfz7-21对干细胞的直接影响,如tian,h.etal.“areservestemcellpopulationinsmallintestinerenderslgr5-positivecellsdispensable.”nature478,255-259(2011)所述监测自lgr5-gfp小鼠衍生的类器官中gfp+干细胞的存在。与对照dmso相比,用dfz7-21处理减少lgr5-gfp干细胞的数目,如通过流式细胞术(见图41a-41c)和共焦显微术(数据未显示)测定的。在图41a和41b中,解离lgr5-gfp类器官,用sytox处理并通过流式细胞术来分析。图41a显示用dmso处理的表达gfp+的活细胞的代表性图,而图41b显示用dfz7-21处理的表达gfp+的活细胞的代表性图。图41a和41b的量化在图41c中显示。为了验证dfz7-21抑制的机制和它的特异性,测试肽处理对展现活性下游wnt信号传导的类器官的影响。简言之,将lgr5-gfp类器官用dmso或gsk3β抑制剂chir99021(5μm)预处理24小时,然后用dmso,dfz7-21(100μm),或fz7-21s(100μm)±chir99021处理。通过针对轴蛋白2(图42a)和ascl2(图42b)的rt-pcr量化转录物。在第二组实验中,培养apcmin类器官并用dmso,dfz7-21(100μm),fz7-21s(100μm)处理24小时,并通过针对轴蛋白2(图42c),ascl2(图42d),和lgr5(图42c)的rt-pcr量化转录物。(apcmin(多发性肠瘤形成)小鼠携带单一突变体apc等位基因且到4-6月龄时在小肠中形成50-100个良性腺瘤,总是与剩余野生型基因丧失有关。)观察到对用chir99021(一种gsk3β抑制剂)预处理或具有apc突变体背景的肠类器官中的干细胞(lgr5,ascl2)和wnt信号传导(轴蛋白2)标志物的很少影响或无影响。这些发现指示dfz7-21在受体水平作用于β-联蛋白和apc上游,与2d细胞培养物数据(图2e)一致。总之,实施例中讨论的代表性结果证明dfz7-21在脂质结合沟处对fzd7crd亚类的选择性靶向足以损害wnt信号传导和干细胞功能。在fz7-21-fzd7crd复合物中观察到的剧烈构象变化很可能使得受体不能进行正确的信号传导,如通过肽在体外和在体内的抑制作用证明的,由此提供肽的作用模式的机制。已经描述了一种有力且选择性的小肽拮抗剂(即fz7-21和它的衍生物)。这种肽因它的高效力,选择性和药理学作用模式而不同于先前报告的靶向fzd受体家族的多种亚类的基于抗体或小分子的拮抗剂。参见例如leeetal.(2015)“structure-baseddiscoveryofnovelsmallmoleculewntsignalinginhibitorsbytargetingthecysteine-richdomainoffrizzled.”jbiolchem290,30596-30606和gurneyetal.(2012)“wntpathwayinhibitionviathetargetingoffrizzledreceptorsresultsindecreasedgrowthandtumorigenicityofhumantumors.”procnatlacadsciu.s.a.109,11717-11722。与fz7-21复合的hfzd7crd的晶体结构揭示一种新颖的crd构象。而且,本文中提供的数据将脂质沟结合机制定义为同等型选择性fzd抑制的基础。相对于脱辅基fzd7crd,肽fz7-21改变fzd7crd和它的脂质结合腔的二聚体构型,提供具有开放的二聚体界面和‘延伸的’脂质结合沟的开放状态fzdcrd的第一个例子(见图12a和12b)。肽不与高亲和力wnt配体竞争fzd7crd结合,这可能是有益的,尤其在体内背景中。虽然在体外在fz7-21存在下fzdcrd上有另外的wnt配体募集,但是在fz7-21–fzd7crd复合物中观察到的剧烈的构象变化很可能使得受体不能进行正确的信号传导,如通过肽在细胞和肠类器官中的抑制作用证明的,由此为肽的作用模式提供貌似合理的解释。而且,不受理论束缚,鉴于这些蛋白质间的高度序列保守性和来自elisa的类似发现,可能的是肽会以与fzd7crd相似的方式结合fzd1和fzd2crd。上文讨论的数据指示fzd7crd亚类在脂质结合沟处的选择性靶向足以损害wnt和干细胞功能。fz7-21充当一种选择性药理学工具,用于进一步探查fzd7在干细胞和癌症生物学中的作用。此类发现提示fzd7crd脂质结合沟几何学在介导wnt信号以调节lgr5+肠干细胞中有至关紧要的作用。在肠隐窝的底部明显缺乏fzd7类受体的功能冗余,偶联显示fzd7受体亚类的选择性靶向的数据,提示在依赖于活性上游wnt信号传导的肿瘤中用小配体在药理学上靶向fzd7的一种可能路径(seshagiri(2012)“recurrentr-spondinfusionsincoloncancer.”nature488,660-664;jiangetal.(2013)“inactivatingmutationsofrnf43conferwntdependencyinpancreaticductaladenocarcinoma”.procnatlacadsciu.s.a.110,12649-12654;madanetal.(2016)“usp6oncogenepromoteswntsignalingbydeubiquitylatingfrizzleds.”procnatlacadsciu.s.a.113,e2945-2954)。实施例5:fz7-21肽衍生物fz7-21的m13和l6面对fzd7crd上的开放脂质结合沟。见图13,25a,和25b。实施进一步的实验以评估在这些位置将脂质与fz7-21缀合是否会增强肽对fzd7crd的亲和力,干扰wnt对fzd7crd的结合,或二者。生成fz7-21的含脂质衍生物(在表17中列出)并评估它们抑制wnt3a介导的β-联蛋白信号传导的能力,即如上文描述的。简言之,将已经用tcf/lef响应性萤火虫萤光素酶报告物构建物稳定转染且组成性表达海肾萤光素酶构建物的hek293-tb细胞在下文表17中列出的二聚化的肽的3倍连续稀释液中用重组mwnt3a(50ng/ml)刺激6小时。平行实施dmso对照。作为针对dmso对照标准化的萤火虫萤光素酶对海肾萤光素酶的比测量β-联蛋白信号传导。如表17和图21a-21k中显示的,在用外源wnt3a刺激的hek293细胞中dfz7-d21δ2.l6h和fz7-21δ2以与dfz7-21相当的ic50值损害wnt3a介导的β-联蛋白信号传导。(表17和图21a-21k中的值是至少3份独立复制品(除非另有说明)的平均值±标准偏差。表17**经由cys10处的二硫键二聚化的肽。*经由cys8处的二硫键二聚化的肽。§在一式两份实验中测试肽。在一组后续实验中,实施elisa测定法以评估低浓度的fz7-21,dfz7-21,和fz7-21s(例如会在与~100nm的细胞ic50相同的范围内的浓度)对wnt对fzdcrd的结合的影响。简言之,使用(a)fzd1crd-fc,(b)fzd2crd-fc,(c)fzd4crd-fc,或(d)fzd7crd-fc作为捕捉试剂和生物素化的wnt5a作为检测试剂实施测定法。以介于0μm-10μm之间的递增浓度添加肽(1)dfz7-21,(2)fz7-21,(3)fz7-21s或(4)dmso对照。使用链霉亲合素-hrp来检测生物素化的wnt5a,并作为428nm处的化学发光检测的函数来测量hrp活性。值得注意的是,wnt5a对fzd1crd,fzd2crd,和fzd7crd的结合在浓度低于约0.5μm的fz7-21存在下升高(图22a),指示fz7-21以浓度依赖性方式增强fzd1crd,fzd2crd,和fzd7crd上的wnt5a募集。在0.1μmfz7-21存在下看到最大结合。与之对比,fz7-21对wnt5a对fzd4(即与fzd1,fzd2,和fzd7分享比fzd1,fzd2,和fzd7彼此分享的序列相似性要少的序列相似性的一种fzd)上的crd的结合没有此类影响。wnt5a对fzd1crd,fzd2crd,和fzd7crd的结合也在浓度低于约0.5μm的dfz7-21存在下升高(图22b),在0.05μmdfz7-21存在下看到最大结合。dfz7-21对wnt5a对fzd4上的crd的结合没有此类影响。fz7-21s对wnt5a对测试的任何fzdcrd的结合没有影响(图22c)。使用wnt3a观察到类似的结果。见图22d-22f。wnt3a对fzd1crd,fzd2crd,和fzd7crd的结合在浓度低于约0.5μm的fz7-21存在下升高(图22d),在0.05μm-0.1μmdfz7-21存在下看到最大结合。wnt3a对fzd1crd,fzd2crd,和fzd7crd的结合在浓度低于约0.5μm的dfz7-21存在下升高(图22e),在0.01μm-0.05μmdfz7-21存在下看到最大结合。fz7-21或dfz7-21均对wnt3a对fzd4crd的结合没有任何影响。fz7-21s对wnt3a对测试的任何fzdcrd的结合没有影响(图22f)。图22a-22f中的值是针对dmso对照标准化的。所有测定法是以1%dmso终浓度实施的。鉴于令人惊讶的fz7-21和dfz7-21对wnt对fzd1crd,fzd2crd,和fzd7crd的结合的影响,重复上文描述的测定法以测试表17中列出的fz7-21衍生肽二聚体对wnt5a对fzd4crd和fzd7crd的结合的影响。测试的所有fz7-21衍生肽二聚体抑制fzd7crd上的wnt5a募集。见图23a-23j。fz7-21衍生肽二聚体对wnt5a对fzd4crd的结合没有影响。见图23k和23l。fz7-21s对wnt5a对测试的fzdcrd的结合没有影响。见图23a-23l。总之,申请人令人惊讶地显示fz7-21衍生肽二聚体(每一种因疏水性非天然氨基酸的存在而不同于dfz7-21)抑制fzd7crd上的wnt5a募集。此类结果提示表17中列出的fz7-21衍生肽二聚体对fzd7crd的结合将fzd7crd的脂质结合腔自脱辅基结构中的弯曲形式改变成延伸形式(见图12a和12b),其容许每种fz7-21衍生肽二聚体中存在的非天然氨基酸的疏水部分渗透延长的脂质结合沟,如此阻断wnt5a对fzd7crd的结合。实施例6:实施例7的材料和方法试剂和重组蛋白质在表达endoh的粉纹夜蛾细胞中作为分泌型蛋白质表达hfzd7crd-his[gln33–gly168]并用几夫碱处理。然后通过标准ni-nta亲和层析术继以大小排阻层析术来纯化它,如更早描述的20。在粉纹夜蛾细胞中作为分泌型蛋白质表达hfzd5crd-his[ala27–ala155],然后通过标准ni-nta亲和层析术继以大小排阻层析术纯化,如bourhis,e.etal.reconstitutionofafrizzled8.wnt3a.lrp6signalingcomplexrevealsmultiplewntanddkk1bindingsitesonlrp6.j.biol.chem.285,9172-9(2010)中描述的。x射线晶体学纯化c24结合的hfzd7crd-his,缓冲液交换入150mmnacl,50mmtris-hcl,ph7.5,然后通过离心(目录号ufc900325;milliporeamiconultra15)浓缩至25mg/ml。将蛋白质与含有2.2m硫酸铵,100mmbis-trisph6.5的储备溶液混合(1:1)并通过蒸气扩散于19℃温育。在stanfordsynchrotronradiationlightsource(ssrl)12-2收集衍射数据并以分辨率通过分子置换来解析。纯化hfzd5crd-his并缓冲液交换入150mmnacl,50mmtris-hcl,ph8.0,然后通过离心浓缩至8mg/ml。将蛋白质与含有0.1m柠檬酸三钠二水合物ph5.5,22%聚乙二醇3350的储备溶液1:1混合并通过蒸气扩散于19℃温育。在用于fzd5bog共晶体的结晶介质中包括0.1%正辛基-β-d-葡萄糖苷(bog)。对于fzd5crd与棕榈油酸的共结晶,在含有50%dmso的150mmnacl,50mmtris-hcl,ph8.0缓冲液中作为储液(10mg/ml)制备后者,然后将它与fzd5crd混合(1:1)。通过混合蛋白质-脂肪酸复合物与储备溶液(1:1)来设置共结晶,如上文描述的。储备溶液还含有0.1%棕榈油酸。在advancedlightsource(als)光束线5.0.1收集hfzd5crd:c16:1n-7和hfzd5crd:bog晶体的衍射数据并分别以和分辨率用分子置换解析。统计学在补充表1中可得。hfzd7crd疏水腔展示为了显现hfzd8crd内的疏水腔,采用needleman-wunsch比对算法和blosum-62矩阵,使用ucsfchimera中的matchmaker效用将与mfzd8crd复合的两个xwnt8分子(pdbid#f40a)叠加到每个hfzd8crd二聚体对上。对于与bog或c16:1n-7结合的hfzd5crd和与c24脂肪酸结合的hfzd7crd,使用它们的相应配体来定义疏水腔。在距结合的配体内鉴定连续疏水表面并依照腔的疏水性显现。计算机fzdcrd二聚体界面分析使用ucsfchimera可视化套件(1.11版)生成图。使用势能效用及amber10:eht力场和born溶剂化,在moe(chemicalcomputinggroup;2017.05版)中计算二聚体界面能量学。来自力场的能量值以kcal/mol报告且没有定标来反映实际的结合自由能。使用sc算法的执行来对connolly表面之间的互补性排序,其忽略距表面的边缘内的表面(worldwideweb-ccp4.ac.uk/newsletters/newsletter39/02_sc.html)。在计算前,使用protonate3d将晶体学模型质子化以优化氢放置(moe,2017.05版)。使用ucsfchimera套件(1.11版)内的matchmaker效用使用指定结构的每个对齐且叠加的残基的α碳计算均方根偏差(rmsd)。实施例7:fz7-21肽衍生物fzd受体在多种多样的过程中介导wnt信号传导,范围自骨生长至干细胞活性。然而,fzd受体的crd对wnt顺式不饱和脂肪酰基基团的识别的分子基础仍然是难以捉摸的,直至创造本文中报告的晶体结构。这个实施例显示与c16:1顺式δ9-不饱和脂肪酸结合的人fzd5crd的第一种晶体结构。出乎意料的是,还获得与c24脂肪酸结合的人fzd7crd的晶体结构。两种结构分享crd的一种保守的新颖的二聚体排列。脂质结合沟跨越两个单体且采取容纳脂肪酸的u形几何学。小鼠fzd8crd结构揭示它也与fzd5和fzd7crd分享相同的构造。这个实施例显示多种fzd受体对wnt顺式不饱和脂肪酰基基团的识别的一种共同机制,而且有助于开发特异性fzd受体抑制剂。这个实施例的实验的初始目标是测定脱辅基hfzd7crd的晶体结构。作为可溶性分泌型蛋白质在昆虫细胞中表达hfzd7crd(残基gln33–gly168)并纯化。通过分子置换解析hfzd7crd的x射线晶体结构并精修至的分辨率(见表18)。hfzd7crd采取同二聚体排列,构成晶体学不对称单元的两个原体(链a和b)之间有α-螺旋二聚体界面。出乎意料的是,观察到脂质结合腔中额外的电子密度,显示延长的形状,其类似游离脂肪酸分子(c24)的特征。此类脂肪酸大概起源于昆虫细胞表达宿主。观察到的电子密度指示脂肪酸的羧酸根基团存在于一个单体(a链)中,而烃链的甲基末端存在于另一个单体(b链)中(图29a,29b,29c,和29d)。hfzd7crd二聚体中的每个单体含有一个亲脂沟。两个脂质结合沟在二聚体界面处相遇,形成连续且弯曲(u形)的腔(图29b和29c)。尽管c24结合的hfzd7crd二聚体有几乎严格的非晶体学对称(ncs)2倍排列,但是crd二聚体中的亲脂沟的内部结构中有重要的差异,很可能是由c24脂肪酸的内在不对称性诱导的(在所有118个原子对间a链对b链r.m.s.偏差为见图28a和28b)。值得注意的是,脂质结合沟的内表面衬里的两个关键残基(phe138和phe140)在链a和b之间采取不同的侧链旋转异构体,由此呈现用于结合不对称脂肪酸的不对称通道(见图28b)。另外,脂肪酸看来主要埋藏在u形脂质结合腔中,c9-c13位于疏水腔的基部(图29c和28b)。表18在不同结晶条件下,还获得脱辅基hfzd7crd的晶体结构,其显示与在c24结合的hfzd7crd的结构中观察到的相似的二聚体构造和脂质结合沟几何学。有趣的是,hfzd7crd这次在任何源自表达宿主的内源脂肪酸缺失下结晶。尽管脱辅基和脂质结合的结构均较好叠加(在所有117个原子对间r.m.s.偏差为见图28c),但是有一些关键构象变化,它们在脂质结合的结构中是明显的。例如,疏水腔衬里的残基诸如tyr87和phe138(图29d)的侧链取向改变,提示脂质结合腔是柔性的且可能容纳链长可变的脂肪酸。这些发现与更早的证明wnt蛋白质掺入13-16碳链脂肪酰基基团的生物化学数据一致并给出解释(gao,x.&hannoush,r.n.single-cellimagingofwntpalmitoylationbytheacyltransferaseporcupine.natchembiol10,61-68(2014))。最后,值得注意的是c24的羧酸根基团通过由水分子介导的氢键锚定至tyr76(图29d)。鉴于fzd家族成员间tyr76的高度序列保守性,这种相互作用突显fzd受体对游离脂肪酸的识别的一种潜在分子机制。值得注意的是tyr76在含有细胞外fzd样crd的hedgehog途径受体smoothened(smo)中也是保守的(sharpe,h.j.,wang,w.,hannoush,r.n.&desauvage,f.j.regulationoftheoncoproteinsmoothenedbysmallmolecules.natchembiol11,246-55(2015)),并与20(s)-羟基胆固醇的羟基基团形成极性接触(huang,p.etal.cellularcholesteroldirectlyactivatessmoothenedinhedgehogsignaling.cell166,1176-1187.e14(2016))。值得注意的是,后一种分子在游离脂肪酸在fzd7crd结构中能结合的相同位置在smo的疏水沟中结合(图30a)。上述出乎意料的hfzd7crd:c24复合物结构中的观察结果,特别是二聚体界面周围的脂质结合沟的u形几何学和它对脂肪酸的不对称识别,推动进一步检查其它fzdcrd如何结合c16:1顺式δ9-不饱和脂肪酸(c16:1n-7),它是wnt蛋白质上存在的生理学相关脂质。没有报告fzd5crd,或与不饱和脂肪酸复合的任何其它fzdcrd的结构。fzd5crd展现在进化上接近fzd7和fzd8crd(图7)。因此,在昆虫细胞中作为可溶性分泌型蛋白质表达hfzd5crd(残基ala27–ala155)并将它纯化至接近同质。hfzd5crd与c16:1n-7脂肪酸复合共结晶。通过分子置换解析获得的复合物的x射线晶体结构并精修至的分辨率(见表18)。晶体学不对称单元由两个hfzd5crd单体组成(图31)。对hfzd5crd结构和晶体学对称配偶的检查揭示另外的蛋白质-蛋白质界面(a链同二聚体),其与在脱辅基hfzd7crd和与c24结合的hfzd7crd的结构中观察到的相似(图32a和32b和图31)。有两个脂质结合沟,每个源自一个单体,它们形成连续的u形腔,与在hfzd7crd二聚体结构中观察到的相似。精修结合的c16:1n-7脂肪酸,由于它在双折轴上的特殊位置而在结构中具有50%占据率,这等同于疏水腔内的100%占据率。脂肪酸能以任一方向在脂质结合沟内结合,羧酸根首基位于trp46和gln44残基附近(图32c)。值得注意的是,烃链内的“扭结的”顺式δ9-不饱和位点(c9-c10)位于u形脂质结合腔的基部,在残基ile51和try98(分别是hfzd7crd中的val92和phe138)附近(图32c)。如此,如本文中公开的与c16:1n-7复合的hfzd5crd的晶体结构揭示脂质结合腔的几何学的空前原子分辨率视图和它如何容纳游离不饱和脂肪酸,可能解释fzd受体crd对wnt蛋白质上的顺式不饱和脂肪酰基的优先结合。最后,实施实验以测定脱辅基hfzd5crd的晶体结构。然而,在这种情况中,hfzd5crd在正辛基-β-d-葡萄糖苷(bog)存在下结晶(分辨率),其存在于最佳结晶缓冲液中。有趣的是,bog配体在hfzd5crd的疏水腔中结合,具有50%占据率(图34a,34b,34c,和34d和表18)。总之,在螺旋-螺旋二聚体界面,u形脂质结合腔和配体在疏水腔内的位置方面,bog结合的hfzd5crd结构同与c16:1n-7复合的fzd5crd(在119个残基上r.m.s.偏差为0.134;图34a,34b,34c,和34d)和与c24复合的fzd7crd非常相似。基于脱辅基hfzd7,hfzd7crd:c24,hfzd5crd:c16:1n-7和hfzd5crd:bog之间的结构相似性,特别是二聚体构造和脂质结合沟构造,再次检查发表的mfzd8crd(进化上最保守的fzdcrd)的晶体学数据(pdbid#1ijy)(dann,c.e.etal.insightsintowntbindingandsignallingfromthestructuresoftwofrizzledcysteine-richdomains.nature412,86-90(2001))(图7)。对称配偶分析展示与fzd7和fzd5crd二者相同的二聚体构造(图35a,35b,35c,和35d)。dann等人基于它与分泌型卷曲蛋白相关蛋白3(sfrp3)的相似性和它的介导二聚体界面的环-环区域之间的有利互补性得分来指派不对称单元(dann,c.e.etal.insightsintowntbindingandsignallingfromthestructuresoftwofrizzledcysteine-richdomains.nature412,86-90(2001))。在发表时,wnt的顺式不饱和脂肪酰化状态和它的脂肪酸介导的对fzdcrd的结合均是未知的(janda,c.y.,waghray,d.,levin,a.m.,thomas,c.&garcia,k.c.structuralbasisofwntrecognitionbyfrizzled.science337,59-64(2012);takada,r.etal.monounsaturatedfattyacidmodificationofwntprotein:itsroleinwntsecretion.developmentalcell11,791-801(2006)),没有提供关于u形脂质结合腔的线索。基于能量和互补性得分(lawrence,m.c.&colman,p.m.shapecomplementarityatprotein/proteininterfaces.j.mol.biol.234,946-950(1993)),本文中公开的计算机分析支持螺旋-螺旋(fzd7样)二聚体界面作为mfzd8crd二聚体内的潜在生物学界面(见图36a,36b,36c,36d,36e,36f,36g,和36h)。在计算上,环-环和螺旋-螺旋二聚体界面在它们的互补性得分方面均是相似;然而,对于hfzd7和mfzd8crd二者,螺旋-螺旋二聚体界面与环-环界面相比在能量上更加有利(图36a,36b,36c,36d,36e,36f,36g,和36h)。基于下面的观察结果,数项证据支持具有u形脂质结合沟的螺旋-螺旋二聚体界面构造很可能是生物学单元的观念:(a)脂肪酸的结合模式在不同条件下独立结晶的两种不同fzdcrd结构(fzd5和7)之间是一致的,揭示烃脂肪酰基链同时跨越两个fzdcrd单体上的两个脂质结合沟,羧酸末端在一个单体上而烃链的甲基末端在另一个单体上;(b)螺旋-螺旋二聚体界面在fzd7和8家族成员间的保守性(图37a和37b);和(c)“扭结的”疏水腔在多个fzd家族成员间的结构相似性和保守性,如这项研究中显示的(图35d)。需要另外的研究来在体外和在体内二者探索观察到的二聚体几何学和它对脂质结合腔的调节的功能相关性。无论如何,本文中公开的结构研究为被单个脂肪酸占据的连续疏水脂质结合腔提供有力支持。基于这项研究中呈现的结构数据,提出wnt与fzdcrd相互作用的一种分子模型。wnt脂肪酰基基团占据fzdcrd的u形脂质结合腔,‘扭结的’顺式δ9-不饱和位点位于腔的基部(图35e和图38a,38b,和38c)。这项研究中报告的hfzd5crd与发表的与xwnt8结合的mfzd8crd结构(janda,c.y.,waghray,d.,levin,a.m.,thomas,c.&garcia,k.c.structuralbasisofwntrecognitionbyfrizzled.science337,59-64(2012))的叠加显示源自单个xwnt8的顺式不饱和脂肪酰基链能同时占据两个脂质结合腔,由此桥接hfzd5crd二聚体界面(图38a,38b,和38c)。fzd7和fzd8crd二聚体同样如此,由此指示wnt–fzdcrd相互作用的1:2化学计量。这与发表的与xwnt8结合的mfzd8crd结构(janda,c.y.,waghray,d.,levin,a.m.,thomas,c.&garcia,k.c.structuralbasisofwntrecognitionbyfrizzled.science337,59-64(2012))中描绘的1:1化学计量相当不同。在这后一种情况中,脂质结合沟处于溶剂暴露取向,这是不利的,而且没有完全埋藏16碳脂肪酰基链(janda,c.y.,waghray,d.,levin,a.m.,thomas,c.&garcia,k.c.structuralbasisofwntrecognitionbyfrizzled.science337,59-64(2012))(图38a,38b,和38c)。能想到的是研究中使用的含有fc片段的xwnt8–mfzd8crd融合构建物可能没有有利的fzdcrd二聚体形成,由此为观察到的差异提供一种解释。有趣的是还注意到作者未能毫无疑义地确定存在的脂质是否是棕榈油酸或饱和棕榈酸。确实,在xwnt8–mfzd8crd结构中建模的脂肪酰基链令人惊讶地是直的(图38a),而非根据单不饱和脂肪酸会预期的扭结的,尽管分辨率低;如果是这种情况,那么此类脂质的存在会排除fzdcrd二聚化。无论如何,这里提出的wnt-fzdcrd1:2化学计量提示wnt蛋白质,经由它的不饱和脂肪酰基基团,可能有助于促进fzdcrd二聚化,由此增强fzd受体在细胞表面聚簇并推动后续下游信号体组装(bienz,m.,signalosomeassemblybydomainsundergoingdynamichead-to-tailpolymerization.trendsbiochemsci39,487-95(2014))。总之,本文中报告的是与游离脂肪酸复合的两种fzdcrd(fzd5和fzd7)的第一种晶体结构,为不饱和脂肪酸可能如何与fzdcrd相互作用提供长期追寻的结构原理。这些研究揭示脂质结合沟是柔性的,容纳同时结合两个crd单体的单一脂肪酸。在再次检查更早发表的结构(dann,c.e.etal.insightsintowntbindingandsignallingfromthestructuresoftwofrizzledcysteine-richdomains.nature412,86-90(2001))后,发现mfzd8crd也分享与这里报告的hfzd7和hfzd5crd相似的结构特征,包括α-螺旋二聚体界面和连续u形脂质结合腔,导致再次审视该结构中的二聚体的解读。总之,这个实施例中提供的实验数据提示多种fzdcrd间识别顺式不饱和脂肪酸的一种共同机制,而且提供关于wnt能如何与卷曲蛋白受体相互作用并促进它经由顺式不饱和脂肪酰基基团的二聚化的分子照片。最后,本文中的发现可推动开发利用新发现的二聚体界面,和脂质结合沟构造及其柔性靶向特定fzdcrd受体亚类的药理学策略。成体肠上皮的再生由位于隐窝基部的表达含富亮氨酸重复的g蛋白偶联受体5(lgr5)的循环干细胞的集合介导。在10种哺乳动物卷曲蛋白(fzd)受体中,fzd7在lgr5+肠干细胞中富集且在它们的自我更新中发挥至关紧要作用。最近的研究提示fzd7可能是与干细胞功能障碍有关的疾病的一种潜在药理学靶物;然而,用于选择性fzd抑制的办法和结构基础仍然定义不足。通过经由位于fzd富半胱氨酸域(crd)内的脂质结合沟部分结合它们的脂肪酰基基团,fzd与wnt蛋白质相互作用。这里我们鉴定一种高度有力且选择性的肽,它结合fzd7crd并改变它的脂质结合沟的构造。用fzd7结合肽处理损害wnt信号传导且下调主要在肠类器官的干细胞隔室中表达的基因。依照本文中的数据,脂质沟结合机制充当同等型选择性fzd抑制的基础,而且暗示fzd7crd脂质结合沟几何学在肠干细胞功能中的作用。前述实施例仅仅出于例示目的而提供,并非意图以任何方式限制本发明的范围。在本文中显示和描述的那些以外对本发明的各种修饰会根据前述描述而对本领域技术人员变得显而易见且落在所附权利要求的范围内。序列表<110>基因泰克公司(genentech,inc.)豪夫迈·罗氏有限公司(f.hoffmann-larocheag)a·h·尼罗(nile,aaronhugh)张映楠(zhang,yingnan)l·周(zhou,lijuan)r·海诺时(hannoush,rami)<120>卷曲蛋白的选择性肽抑制剂<130>146392036940<140>未指派<141>随本文<160>156<170>fastseqforwindowsversion4.0<210>1<211>14<212>prt<213>人工序列<220><223>合成构建物<400>1tyrgluhisleuhisaspleumetaspleuileargprotrp1510<210>2<211>16<212>prt<213>人工序列<220><223>合成构建物<400>2thrtyrpheaspaspilecysasnleuileleuprotrpalaasnpro151015<210>3<211>16<212>prt<213>人工序列<220><223>合成构建物<400>3proglnaspleuleuasptrpcyshistyrmetilevalserserasp151015<210>4<211>14<212>prt<213>人工序列<220><223>合成构建物<400>4alacyssertyrvalileaspleutrpasnglncysleuthr1510<210>5<211>16<212>prt<213>人工序列<220><223>合成构建物<400>5procysservalilecysleuproasptrpserserleuleupheile151015<210>6<211>12<212>prt<213>人工序列<220><223>合成构建物<400>6aspthraspleuhisglntrpcysleutrpphethr1510<210>7<211>14<212>prt<213>人工序列<220><223>合成构建物<400>7phetrpmetleuleuglngluglyphealaphetrpphepro1510<210>8<211>14<212>prt<213>人工序列<220><223>合成构建物<400>8phegluleuleuleuaspleuglyaspleuileargleutrp1510<210>9<211>14<212>prt<213>人工序列<220><223>合成构建物<400>9alacyssertyrvalileaspleutrpasnleucysleuarg1510<210>10<211>14<212>prt<213>人工序列<220><223>合成构建物<400>10alasergluleuhisasptrpcysargmetmetpheprotrp1510<210>11<211>14<212>prt<213>人工序列<220><223>合成构建物<400>11ileserleuileglualametilealaleuaspargvalphe1510<210>12<211>14<212>prt<213>人工序列<220><223>合成构建物<400>12proproasnvalhisgluglycystrpsermetpheprotrp1510<210>13<211>14<212>prt<213>人工序列<220><223>合成构建物<400>13leuproseraspaspleugluphetrpcyshisvalmettyr1510<210>14<211>12<212>prt<213>人工序列<220><223>合成构建物<400>14aspthraspleuleuglntrpcysleutrpphethr1510<210>15<211>14<212>prt<213>人工序列<220><223>合成构建物<400>15phetrpmetglnleuglnaspglyphealailetrpphepro1510<210>16<211>16<212>prt<213>人工序列<220><223>合成构建物<400>16procysservalilecysleuproasptrpserserleuleupheile151015<210>17<211>14<212>prt<213>人工序列<220><223>合成构建物<400>17glyaspphetrpproglyserleuleutrpgluileleuval1510<210>18<211>14<212>prt<213>人工序列<220><223>合成构建物<400>18ileleuthrpheglutyrphetrpileleuglyleuileleu1510<210>19<211>10<212>prt<213>人工序列<220><223>合成构建物<400>19leuproleuphepheleusertyrvalleu1510<210>20<211>16<212>prt<213>人工序列<220><223>合成构建物<400>20pheleuproaspglnhisserhisleupheleuprotrpglyglupro151015<210>21<211>16<212>prt<213>人工序列<220><223>合成构建物<400>21sercysglnmettrpserasnleuargvalleupheleusertyrtrp151015<210>22<211>12<212>prt<213>人工序列<220><223>合成构建物<400>22valphevalprophesergluleuthrserleucys1510<210>23<211>14<212>prt<213>人工序列<220><223>合成构建物<400>23iletrpphelysglyargphevalglupheserserleuval1510<210>24<211>16<212>prt<213>人工序列<220><223>合成构建物<400>24asnalaphetrpargaspglncysleuglutrppheileilecysleu151015<210>25<211>16<212>prt<213>人工序列<220><223>合成构建物<400>25gluhisaspleuleuleuargalametasnserphevalleuilephe151015<210>26<211>10<212>prt<213>人工序列<220><223>合成构建物<400>26phecysgluasnprotyrileilecystrp1510<210>27<211>10<212>prt<213>人工序列<220><223>合成构建物<400>27asnproproproglucyspheleuserlys1510<210>28<211>16<212>prt<213>人工序列<220><223>合成构建物<400>28valphephetyrhisserleuphepheilelysleuileleuasppro151015<210>29<211>14<212>prt<213>人工序列<220><223>合成构建物<400>29gluargargvalcystyrprotrpphegluvalserglnpro1510<210>30<211>16<212>prt<213>人工序列<220><223>合成构建物<400>30leuserserglylyslysvalsersertyrtrppheasnphetrpphe151015<210>31<211>8<212>prt<213>人工序列<220><223>合成构建物<400>31phetrppheaspphetrpphegly15<210>32<211>16<212>prt<213>人工序列<220><223>合成构建物<400>32serserasppheserglycysleusertrpcysaspleuilephegly151015<210>33<211>16<212>prt<213>人工序列<220><223>合成构建物<400>33pheaspphecysservalmetproglnpheiletyrcysproglyasp151015<210>34<211>16<212>prt<213>人工序列<220><223>合成构建物<400>34hisleuseraspvalcyscysserasptrpcysaspleuvalphetrp151015<210>35<211>16<212>prt<213>人工序列<220><223>合成构建物<400>35thrseraspphesertrpcysleusertrpcysaspleuilephetrp151015<210>36<211>16<212>prt<213>人工序列<220><223>合成构建物<400>36pheaspphecysthrvalmetprohispheiletyrcysproglyasp151015<210>37<211>16<212>prt<213>人工序列<220><223>合成构建物<400>37pheaspphecysservalmetprohispheiletyrcysproglyasp151015<210>38<211>16<212>prt<213>人工序列<220><223>合成构建物<400>38hisleuseraspvalphecysserasptrpcysaspleuvalphetrp151015<210>39<211>14<212>prt<213>人工序列<220><223>合成构建物<400>39valalaalaaspaspleualaalatrpcyshisvalmettyr1510<210>40<211>14<212>prt<213>人工序列<220><223>合成构建物<400>40alaalaseraspaspleugluphetrpcyshisvalmettyr1510<210>41<211>14<212>prt<213>人工序列<220><223>合成构建物<400>41alaalaseraspaspleugluphetrpcyshisvalmettyr1510<210>42<211>14<212>prt<213>人工序列<220><223>合成构建物<400>42alaproseraspaspvalalaphetrpcyshisvalmettyr1510<210>43<211>14<212>prt<213>人工序列<220><223>合成构建物<400>43alaproalaaspaspvalgluphetrpcyshisvalmettyr1510<210>44<211>14<212>prt<213>人工序列<220><223>合成构建物<400>44alaproseraspaspleugluphetrpcyshisvalmettyr1510<210>45<211>14<212>prt<213>人工序列<220><223>合成构建物<400>45alaproalaaspaspleuglualatrpcyshisvalmettyr1510<210>46<211>14<212>prt<213>人工序列<220><223>合成构建物<400>46alaproseraspaspleugluphetrpcyshisalamettyr1510<210>47<211>14<212>prt<213>人工序列<220><223>合成构建物<400>47valalaseraspaspleuglualatrpcyshisvalmettyr1510<210>48<211>14<212>prt<213>人工序列<220><223>合成构建物<400>48alaalaalaaspaspleugluphetrpcyshisvalmettyr1510<210>49<211>14<212>prt<213>人工序列<220><223>合成构建物<400>49alaalaseraspaspleualaalatrpcyshisvalmettyr1510<210>50<211>14<212>prt<213>人工序列<220><223>合成构建物<400>50alaalaseraspaspleuglusertrpcyshisvalmettyr1510<210>51<211>14<212>prt<213>人工序列<220><223>合成构建物<400>51alaproalaaspaspleualaphetrpcyshisvalmettyr1510<210>52<211>14<212>prt<213>人工序列<220><223>合成构建物<400>52leuproalaaspaspleualavaltrpcysaspvalmettyr1510<210>53<211>14<212>prt<213>人工序列<220><223>合成构建物<400>53leuproseraspaspleuglusertrpcyshisvalmettyr1510<210>54<211>14<212>prt<213>人工序列<220><223>合成构建物<400>54alaalaalaaspaspleugluvaltrpcyshisvalmettyr1510<210>55<211>14<212>prt<213>人工序列<220><223>合成构建物<400>55valalaalaaspalaleugluphetrpcyshisvalmettyr1510<210>56<211>14<212>prt<213>人工序列<220><223>合成构建物<400>56alaproseraspaspleualaalatrpcyshisvalvaltyr1510<210>57<211>14<212>prt<213>人工序列<220><223>合成构建物<400>57alaalaalaaspaspleualaalatrpcysaspvalmettyr1510<210>58<211>14<212>prt<213>人工序列<220><223>合成构建物<400>58valalaseraspaspleugluphetrpcyshisvalmettyr1510<210>59<211>14<212>prt<213>人工序列<220><223>合成构建物<400>59alaalaalaaspaspleuglualatrpcysalavalmettyr1510<210>60<211>14<212>prt<213>人工序列<220><223>合成构建物<400>60leualaalaaspaspleuglusertrpcyshisvalmettyr1510<210>61<211>14<212>prt<213>人工序列<220><223>合成构建物<400>61alaproalaaspaspleualasertrpcyshisvalmettyr1510<210>62<211>14<212>prt<213>人工序列<220><223>合成构建物<400>62valalaseraspaspleualasertrpcyshisalamettyr1510<210>63<211>14<212>prt<213>人工序列<220><223>合成构建物<400>63valproalaaspaspleualasertrpcyshisvalmettyr1510<210>64<211>14<212>prt<213>人工序列<220><223>合成构建物<400>64alaproalaaspaspleugluphetrpcyshisvalvaltyr1510<210>65<211>14<212>prt<213>人工序列<220><223>合成构建物<400>65valproseraspaspleualaphetrpcyshisvalmettyr1510<210>66<211>14<212>prt<213>人工序列<220><223>合成构建物<400>66valproalaaspalaleualavaltrpcysaspvalmettyr1510<210>67<211>14<212>prt<213>人工序列<220><223>合成构建物<400>67valproalaaspaspleualaphetrpcyshisvalmettyr1510<210>68<211>14<212>prt<213>人工序列<220><223>合成构建物<400>68valproseraspaspleualasertrpcyshisvalmettyr1510<210>69<211>14<212>prt<213>人工序列<220><223>合成构建物<400>69valproseralaaspleuglusertrpcyshisvalmettyr1510<210>70<211>14<212>prt<213>人工序列<220><223>合成构建物<400>70alaalaseraspaspleuglualatrpcyshisvalmettyr1510<210>71<211>14<212>prt<213>人工序列<220><223>合成构建物<400>71alaproseraspaspleualasertrpcyshisvalmettyr1510<210>72<211>14<212>prt<213>人工序列<220><223>合成构建物<400>72alaproalaaspaspleugluphetrpcyshisvalmettyr1510<210>73<211>14<212>prt<213>人工序列<220><223>合成构建物<400>73alaproseraspaspleualaalatrpcyshisvalmettyr1510<210>74<211>14<212>prt<213>人工序列<220><223>合成构建物<400>74alaproalaaspaspleualaphetrpcyshisvalvaltyr1510<210>75<211>14<212>prt<213>人工序列<220><223>合成构建物<400>75alaproseraspaspleuglualatrpcysaspvalmettyr1510<210>76<211>14<212>prt<213>人工序列<220><223>合成构建物<400>76valproseraspaspleuglualatrpcysaspvalmettyr1510<210>77<211>14<212>prt<213>人工序列<220><223>合成构建物<400>77alaalaseraspaspleualaphetrpcyshisvalmettyr1510<210>78<211>14<212>prt<213>人工序列<220><223>合成构建物<400>78valproalaaspaspleualasertrpcysaspvalmettyr1510<210>79<211>14<212>prt<213>人工序列<220><223>合成构建物<400>79leuproseralaaspleuglusertrpcyshisvalmettyr1510<210>80<211>14<212>prt<213>人工序列<220><223>合成构建物<400>80alaalaalaaspaspleualaphetrpcyshisvalmettyr1510<210>81<211>14<212>prt<213>人工序列<220><223>合成构建物<400>81leualaseraspaspleugluphetrpcyshisvalmettyr1510<210>82<211>14<212>prt<213>人工序列<220><223>合成构建物<400>82leuproalaalaaspleualaalatrpcyshisvalmettyr1510<210>83<211>14<212>prt<213>人工序列<220><223>合成构建物<400>83valproseralaaspleugluthrtrpcyshisvalmettyr1510<210>84<211>14<212>prt<213>人工序列<220><223>合成构建物<400>84leuproalaaspaspleualaalatrpcyshisvalmettyr1510<210>85<211>14<212>prt<213>人工序列<220><223>合成构建物<400>85proproalaaspaspleualaphetrpcysaspvalmettyr1510<210>86<211>14<212>prt<213>人工序列<220><223>合成构建物<400>86valalaseraspaspleualasertrpcyshisvalvaltyr1510<210>87<211>14<212>prt<213>人工序列<220><223>合成构建物<400>87alaalaalaaspaspvalalasertrpcyshisvalmettyr1510<210>88<211>14<212>prt<213>人工序列<220><223>合成构建物<400>88valalaalaaspaspleualaphetrpcysaspvalmettyr1510<210>89<211>14<212>prt<213>人工序列<220><223>合成构建物<400>89alaproalaaspaspleugluphetrpcyshisalamettyr1510<210>90<211>14<212>prt<213>人工序列<220><223>合成构建物<400>90alaproalaaspaspleualaphetrpcysaspvalmettyr1510<210>91<211>14<212>prt<213>人工序列<220><223>合成构建物<400>91alaproseraspaspleualaphetrpcysaspvalmettyr1510<210>92<211>14<212>prt<213>人工序列<220><223>合成构建物<400>92leuproalaaspaspleualaphetrpcysaspvalmettyr1510<210>93<211>14<212>prt<213>人工序列<220><223>合成构建物<400>93alaalaalaaspaspleualaphetrpcysaspvalmettyr1510<210>94<211>14<212>prt<213>人工序列<220><223>合成构建物<400>94leuproalaaspaspleugluphetrpcyshisvalmettyr1510<210>95<211>14<212>prt<213>人工序列<220><223>合成构建物<400>95valproseraspaspleugluphetrpcysalavalmettyr1510<210>96<211>14<212>prt<213>人工序列<220><223>合成构建物<400>96alaproalaaspaspleuglusertrpcyshisvalmettyr1510<210>97<211>14<212>prt<213>人工序列<220><223>合成构建物<400>97alaproseraspaspleualaphetrpcyshisvalvaltyr1510<210>98<211>14<212>prt<213>人工序列<220><223>合成构建物<400>98alaalaalaaspaspleualaalatrpcyshisvalvaltyr1510<210>99<211>12<212>prt<213>人工序列<220><223>合成构建物<400>99seraspaspleugluphetrpcyshisvalmettyr1510<210>100<211>14<212>prt<213>人工序列<220><223>合成构建物<220><221>变体<222>1,2,3<223>xaa=任何氨基酸或非天然氨基酸且可以存在或缺失<220><221>变体<222>7,8<223>xaa=任何氨基酸或非天然氨基酸<400>100xaaxaaxaaaspaspleuxaaxaatrpcyshisvalmettyr1510<210>101<211>13<212>prt<213>人工序列<220><223>合成构建物<220><221>变体<222>1,2<223>xaa=任何氨基酸或非天然氨基酸且可以存在或缺失<220><221>变体<222>6,7<223>xaa=任何氨基酸或非天然氨基酸<400>101xaaxaaaspaspleuxaaxaatrpcyshisvalmettyr1510<210>102<211>12<212>prt<213>人工序列<220><223>合成构建物<220><221>变体<222>1<223>xaa=任何氨基酸或非天然氨基酸且可以存在或缺失<220><221>变体<222>5,6<223>xaa=任何氨基酸或非天然氨基酸<400>102xaaaspaspleuxaaxaatrpcyshisvalmettyr1510<210>103<211>11<212>prt<213>人工序列<220><223>合成构建物<220><221>变体<222>4,5<223>xaa=任何氨基酸或非天然氨基酸<400>103aspaspleuxaaxaatrpcyshisvalmettyr1510<210>104<211>27<212>prt<213>人工序列<220><223>合成构建物<400>104glyalaleupheleuglypheleuglyalaalaglyserthrmetgly151015alatrpserglnprolyslyslysarglysval2025<210>105<211>21<212>prt<213>人工序列<220><223>合成构建物<400>105lysgluthrtrptrpgluthrtrptrpthrglutrpserglnprolys151015lyslysarglysval20<210>106<211>20<212>prt<213>人工序列<220><223>合成构建物<220><221>乙酰化<222>1<220><221>变体<222>20<223>cya修饰<400>106glyleutrpargalaleutrpargleuleuargserleutrpargleu151015leutrpargala20<210>107<211>8<212>prt<213>人工序列<220><223>合成构建物<400>107argargargargargargargarg15<210>108<211>14<212>prt<213>人工序列<220><223>合成构建物<400>108leuproseraspasnleugluphetrpcyshisvalmettyr1510<210>109<211>14<212>prt<213>人工序列<220><223>合成构建物<400>109leuproseraspaspalagluphetrpcyshisvalmettyr1510<210>110<211>14<212>prt<213>人工序列<220><223>合成构建物<400>110leuproseraspaspleugluphealacyshisvalmettyr1510<210>111<211>14<212>prt<213>人工序列<220><223>合成构建物<400>111leuproseraspaspleugluphetrpcyshisvalalatyr1510<210>112<211>14<212>prt<213>人工序列<220><223>合成构建物<400>112leuproseraspaspleugluphetrpcyshisvalmetala1510<210>113<211>14<212>prt<213>人工序列<220><223>合成构建物<400>113leuproseraspaspleugluphetrpserhisvalmettyr1510<210>114<211>12<212>prt<213>人工序列<220><223>合成构建物<220><221>变体<222>11<223>xaa=任何氨基酸或非天然氨基酸<400>114seraspaspleugluphetrpcyshisvalxaatyr1510<210>115<211>12<212>prt<213>人工序列<220><223>合成构建物<220><221>变体<222>4<223>xaa=任何氨基酸或非天然氨基酸<400>115seraspaspxaagluphetrpcyshisvalmettyr1510<210>116<211>12<212>prt<213>人工序列<220><223>合成构建物<220><221>变体<222>4,11<223>xaa=任何氨基酸或非天然氨基酸<400>116seraspaspxaagluphetrpcyshisvalxaatyr1510<210>117<211>12<212>prt<213>人工序列<220><223>合成构建物<220><221>变体<222>10<223>xaa=任何氨基酸或非天然氨基酸<400>117seraspaspleugluphetrpcyshisxaamettyr1510<210>118<211>14<212>prt<213>人工序列<220><223>合成构建物<220><221>变体<222>14<223>xaa=任何氨基酸或非天然氨基酸<400>118leuproseraspaspleugluphetrpcyshisvalmetxaa1510<210>119<211>12<212>prt<213>人工序列<220><223>合成构建物<220><221>变体<222>3<223>xaa=任何氨基酸或非天然氨基酸<400>119seraspxaaleugluphetrpcyshisvalmettyr1510<210>120<211>14<212>prt<213>人工序列<220><223>合成构建物<220><221>变体<222>5<223>xaa=任何氨基酸或非天然氨基酸<400>120leuproseraspxaaleugluphetrpcyshisvalmettyr1510<210>121<211>12<212>prt<213>人工序列<220><223>合成构建物<220><221>变体<222>6<223>xaa=任何氨基酸或非天然氨基酸<400>121seraspaspleugluxaatrpcyshisvalmettyr1510<210>122<211>12<212>prt<213>人工序列<220><223>合成构建物<220><221>变体<222>1<223>xaa=任何氨基酸或非天然氨基酸<400>122xaaaspaspleugluphetrpcyshisvalmettyr1510<210>123<211>14<212>prt<213>人工序列<220><223>合成构建物<220><221>变体<222>10<223>xaa=任何氨基酸或非天然氨基酸<400>123leuproseraspaspleugluphetrpxaahisvalmettyr1510<210>124<211>12<212>prt<213>人工序列<220><223>合成构建物<220><221>变体<222>8<223>xaa=l-硒代半胱氨酸<400>124seraspaspleugluphetrpxaahisvalmettyr1510<210>125<211>12<212>prt<213>人工序列<220><223>合成构建物<400>125seraspaspleugluphetrpcyshisvalglutyr1510<210>126<211>12<212>prt<213>人工序列<220><223>合成构建物<400>126seraspaspleugluphetrpcyshisleumettyr1510<210>127<211>12<212>prt<213>人工序列<220><223>合成构建物<400>127seraspaspleugluphetrpcyshisilemettyr1510<210>128<211>12<212>prt<213>人工序列<220><223>合成构建物<400>128seraspaspleugluphetrpcyshisthrmettyr1510<210>129<211>12<212>prt<213>人工序列<220><223>合成构建物<400>129seraspaspphegluphetrpcyshisvalmettyr1510<210>130<211>12<212>prt<213>人工序列<220><223>合成构建物<400>130seraspgluleugluphetrpcyshisvalmettyr1510<210>131<211>14<212>prt<213>人工序列<220><223>合成构建物<400>131leuproseraspglnleugluphetrpcyshisvalmettyr1510<210>132<211>12<212>prt<213>人工序列<220><223>合成构建物<400>132thraspaspleugluphetrpcyshisvalmettyr1510<210>133<211>14<212>prt<213>人工序列<220><223>合成构建物<400>133leuproseraspaspleugluphetrpalahisvalmettyr1510<210>134<211>13<212>prt<213>人工序列<220><223>合成构建物<220><221>变体<222>13<223>xaa=l-高炔丙基甘氨酸或l-二高炔丙基甘氨酸<400>134seraspaspleugluphetrpcyshisvalmettyrxaa1510<210>135<211>12<212>prt<213>人工序列<220><223>合成构建物<220><221>变体<222>5<223>xaa=5-叠氮基-l-鸟氨酸或s-乙酰胺基甲基-l-半胱氨酸或叠氮基-高丙氨酸或s-乙酰胺基甲基-l-半胱氨酸<400>135seraspaspleuxaaphetrpcyshisvalmettyr1510<210>136<211>190<212>prt<213>人工序列<220><223>合成构建物<400>136alaglyserglnprotyrhisglyglulysglyileservalproasp151015hisglyphecysglnproileserileproleucysthraspileala202530tyrasnglnthrileleuproasnleuleuglyhisthrasnglnglu354045aspalaglyleugluvalhisglnphetyrproleuvallysvalgln505560cysserprogluleuargphepheleucyssermettyralaproval65707580cysthrvalleuaspglnalaileproprocysargserleucysglu859095argalaargglnglycysglualaleumetasnlyspheglyphegln100105110trpprogluargleuargcysgluasnpheprovalhisglyalagly115120125gluilecysvalglyglnasnthrseraspglyglyglyserglygly130135140serglyglyserglyglyserglyglyserglyglyserglyglyser145150155160glyglyserleuproseraspaspleugluphetrpcyshisvalmet165170175tyrglyserglyserglyasnserhishishishishishis180185190<210>137<211>65<212>prt<213>人工序列<220><223>合成构建物<400>137aspalaglyleugluvalhisglnphetyrproleuvallysvalgln151015cysserprogluleuargphepheleucyssermettyralaproval202530cysthrvalleuaspglnalaileproprocysargserleucysglu354045argalaargglnglycysglualaleumetasnlyspheglyphegln505560trp65<210>138<211>65<212>prt<213>人工序列<220><223>合成构建物<400>138aspalaglyleugluvalhisglnphetyrproleuvallysvalgln151015cysserprogluleuargphepheleucyssermettyralaproval202530cysthrvalleugluglnalaileproprocysargserilecysglu354045argalaargglnglycysglualaleumetasnlyspheglyphegln505560trp65<210>139<211>65<212>prt<213>人工序列<220><223>合成构建物<400>139aspalaglyleugluvalhisglnphetyrproleuvallysvalgln151015cysseralagluleulysphepheleucyssermettyralaproval202530cysthrvalleugluglnalaleuproprocysargserleucysglu354045argalaargglnglycysglualaleumetasnlyspheglyphegln505560trp65<210>140<211>66<212>prt<213>人工序列<220><223>合成构建物<400>140glualaglyleugluvalhisglnphetrpproleuvalgluilegln151015cysserproaspleuargphepheleucyssermettyrthrproile202530cysleuproasptyrhislysproleuproprocysargservalcys354045gluargalalysalaglycysserproleumetargglntyrglyphe505560alatrp65<210>141<211>66<212>prt<213>人工序列<220><223>合成构建物<400>141glualaglyleugluvalhisglnphetrpproleuvalgluilegln151015cysserproaspleulysphepheleucyssermettyrthrproile202530cysleugluasptyrlyslysproleuproprocysargservalcys354045gluargalalysalaglycysalaproleumetargglntyrglyphe505560alatrp65<210>142<211>66<212>prt<213>人工序列<220><223>合成构建物<400>142aspalagluleuglnleuthrthrphethrproleuileglntyrgly151015cysserserglnleuglnphepheleucysservaltyrvalpromet202530cysthrglulysileasnileproileglyprocysglyglymetcys354045leuservallysargargcysgluprovalleulysglupheglyphe505560alatrp65<210>143<211>66<212>prt<213>人工序列<220><223>合成构建物<400>143glualaalaalagluleualagluphealaproleuvalglntyrgly151015cyshisserhisleuargphepheleucysserleutyralapromet202530cysthraspglnvalserthrproileproalacysargprometcys354045gluglnalaargleuargcysalaproilemetgluglnpheasnphe505560glytrp65<210>144<211>66<212>prt<213>人工序列<220><223>合成构建物<400>144glualaalaileglnleuhisgluphealaproleuvalglutyrgly151015cyshisglyhisleuargphepheleucysserleutyralapromet202530cysthrgluglnvalserthrproileproalacysargvalmetcys354045gluglnalaargleulyscysserproilemetgluglnpheasnphe505560lystrp65<210>145<211>65<212>prt<213>人工序列<220><223>合成构建物<400>145ilealaalavalglumetgluhispheleuproleualaasnleuglu151015cysserproasnilegluthrpheleucyslysalaphevalprothr202530cysilegluglnilehisvalvalproprocysarglysleucysglu354045lysvaltyrseraspcyslyslysleuileaspthrpheglyilearg505560trp65<210>146<211>65<212>prt<213>人工序列<220><223>合成构建物<400>146thralaalaleualametgluprophehisprometvalasnleuasp151015cysserargasppheargpropheleucysalaleutyralaproile202530cysmetglutyrglyargvalthrleuprocysargargleucysgln354045argalatyrserglucysserlysleumetglumetpheglyvalpro505560trp65<210>147<211>109<212>prt<213>人(homosapiens)<400>147proasphisglyphecysglnproileserileproleucysthrasp151015ilealatyrasnglnthrileleuproasnleuleuglyhisthrasn202530glngluaspalaglyleugluvalhisglnphetyrproleuvallys354045valglncysserprogluleuargphepheleucyssermettyrala505560provalcysthrvalleuaspglnalaileproprocysargserleu65707580cysgluargalaargglnglycysglualaleumetasnlysphegly859095pheglntrpprogluargleuargcysgluasnphepro100105<210>148<211>109<212>prt<213>人(homosapiens)<400>148proasphisglyphecysglnproileserileproleucysthrasp151015ilealatyrasnglnthrilemetproasnleuleuglyhisthrasn202530glngluaspalaglyleugluvalhisglnphetyrproleuvallys354045valglncysserprogluleuargphepheleucyssermettyrala505560provalcysthrvalleugluglnalaileproprocysargserile65707580cysgluargalaargglnglycysglualaleumetasnlysphegly859095pheglntrpprogluargleuargcysgluhisphepro100105<210>149<211>109<212>prt<213>人(homosapiens)<400>149proasphisglytyrcysglnproileserileproleucysthrasp151015ilealatyrasnglnthrilemetproasnleuleuglyhisthrasn202530glngluaspalaglyleugluvalhisglnphetyrproleuvallys354045valglncysseralagluleulysphepheleucyssermettyrala505560provalcysthrvalleugluglnalaleuproprocysargserleu65707580cysgluargalaargglnglycysglualaleumetasnlysphegly859095pheglntrpproaspthrleulyscysglulysphepro100105<210>150<211>110<212>prt<213>人(homosapiens)<400>150serlysalaprovalcysglngluilethrvalprometcysarggly151015ileglytyrasnleuthrhismetproasnglnpheasnhisaspthr202530glnaspglualaglyleugluvalhisglnphetrpproleuvalglu354045ileglncysserproaspleuargphepheleucyssermettyrthr505560proilecysleuproasptyrhislysproleuproprocysargser65707580valcysgluargalalysalaglycysserproleumetargglntyr859095glyphealatrpprogluargmetsercysaspargleupro100105110<210>151<211>110<212>prt<213>人(homosapiens)<400>151alalysgluleualacysglngluilethrvalproleucyslysgly151015ileglytyrasntyrthrtyrmetproasnglnpheasnhisaspthr202530glnaspglualaglyleugluvalhisglnphetrpproleuvalglu354045ileglncysserproaspleulysphepheleucyssermettyrthr505560proilecysleugluasptyrlyslysproleuproprocysargser65707580valcysgluargalalysalaglycysalaproleumetargglntyr859095glyphealatrpproaspargmetargcysaspargleupro100105110<210>152<211>110<212>prt<213>人(homosapiens)<400>152gluglugluargargcysaspproileargilesermetcysglnasn151015leuglytyrasnvalthrlysmetproasnleuvalglyhisgluleu202530glnthraspalagluleuglnleuthrthrphethrproleuilegln354045tyrglycysserserglnleuglnphepheleucysservaltyrval505560prometcysthrglulysileasnileproileglyprocysglygly65707580metcysleuservallysargargcysgluprovalleulysgluphe859095glyphealatrpprogluserleuasncysserlysphepro100105110<210>153<211>110<212>prt<213>人(homosapiens)<400>153argglyalaalaprocysglnalavalgluileprometcysarggly151015ileglytyrasnleuthrargmetproasnleuleuglyhisthrser202530glnglyglualaalaalagluleualagluphealaproleuvalgln354045tyrglycyshisserhisleuargphepheleucysserleutyrala505560prometcysthraspglnvalserthrproileproalacysargpro65707580metcysgluglnalaargleuargcysalaproilemetgluglnphe859095asnpheglytrpproaspserleuaspcysalaargleupro100105110<210>154<211>110<212>prt<213>人(homosapiens)<400>154proglyaspglylyscysglnproilegluileprometcyslysasp151015ileglytyrasnmetthrargmetproasnleumetglyhisgluasn202530glnargglualaalaileglnleuhisgluphealaproleuvalglu354045tyrglycyshisglyhisleuargphepheleucysserleutyrala505560prometcysthrgluglnvalserthrproileproalacysargval65707580metcysgluglnalaargleulyscysserproilemetgluglnphe859095asnphelystrpproaspserleuaspcysarglysleupro100105110<210>155<211>109<212>prt<213>人(homosapiens)<400>155hisserleuphethrcysgluproilethrvalproargcysmetlys151015metalatyrasnmetthrphepheproasnleumetglyhistyrasp202530glnserilealaalavalglumetgluhispheleuproleualaasn354045leuglucysserproasnilegluthrpheleucyslysalapheval505560prothrcysilegluglnilehisvalvalproprocysarglysleu65707580cysglulysvaltyrseraspcyslyslysleuileaspthrphegly859095ileargtrpproglugluleuglucysaspargleugln100105<210>156<211>109<212>prt<213>人(homosapiens)<400>156hisserleuphesercysgluproilethrleuargmetcysglnasp151015leuprotyrasnthrthrphemetproasnleuleuasnhistyrasp202530glnglnthralaalaleualametgluprophehisprometvalasn354045leuaspcysserargasppheargpropheleucysalaleutyrala505560proilecysmetglutyrglyargvalthrleuprocysargargleu65707580cysglnargalatyrserglucysserlysleumetglumetphegly859095tyrprotrpprogluaspmetglucysserargphepro100105当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1
- SPNP‑I在制备Nav1.7通道工具试剂中的应用的制造方法与工艺
- SPNP‑I在制备Nav1.2通道工具试剂中的应用的制造方法与工艺
- SPNP‑IX在制备Nav1.3通道抑制剂中的应用的制造方法与工艺
- SPNP‑IX在制备Kv4.2通道工具试剂中的应用的制造方法与工艺
- SPNP‑IX在制备Kv4.3通道工具试剂中的应用的制造方法与工艺
- SPNP‑27在制备Nav1.5通道抑制剂中的应用的制造方法与工艺
- SPNP‑27在制备Nav1.3通道抑制剂中的应用的制造方法与工艺
- 一种多肽在制备Nav1.3钠通道工具试剂中的应用的制造方法与工艺
- SPKP‑XI在制备Kv4.2通道抑制剂中的应用的制造方法与工艺
- SGLT‑2糖尿病抑制剂及其中间体的制备方法与流程