BK通道蛋白在制备成骨细胞增殖异常干预药物中的用途的制作方法
本发明属生物制药领域,涉及bk通道蛋白在制备成骨细胞干预药物中的用途,尤其是bk通道蛋白在制备成骨细胞增殖分化异常干预药物中的用途。
背景技术:
现有技术公开了bk通道蛋白大电导钙激活钾通道(bk),又名maxik/kca1.1通道,由kcnma1基因编码并在大多数细胞和组织中广泛表达;bk通道蛋白的独特之处在于它可以通过膜电压和局部细胞内钙来调节。关于bk通道蛋白在兴奋性细胞中的研究已有很多,而其在非兴奋性组织如骨细胞、肿瘤细胞和免疫细胞中的作用鲜有报道。bk通道存在于人骨肉瘤细胞(mg-63、saos-2和g292细胞)和成骨前体细胞(c1细胞)。有研究表明,非特异性钾抑制剂四乙胺(tea)在低浓度下会增加细胞数量而在高浓度下会减少细胞数量;更重要的是,bk基因敲除的小鼠呈现骨质疏松,长骨密度减少,小梁孔隙增大,这表明了在骨细胞中bk通道的重要作用。然而,成骨细胞中bk通道的作用尚不清楚。虽然zhang发现了骨髓间充质干细胞中由shrna敲除kcnma1基因会抑制细胞增殖与成骨,但是rna干扰介导的敲除是不完整的,只减弱基因功能。理想的基因分析方法是通过遗传密码(敲除)的改变来彻底消除基因功能。
zfn(锌指核酸酶)的出现,talen(转录激活因子样效应物核酸酶)和规律成簇间隔短回文重复(crispr)-casenzymes技术近年来在真核细胞和实验动物的目标基因组修饰中广泛应用,tale在植物病原菌xanthomonassp中被发现,talen由tale蛋白及foki核酸酶融合构建而成。tale蛋白有重复可变的双氨基酸残基(rvd),这决定了核苷酸结合的特异性(ni=a,hd=c,ng=t,nn=g或a);通过组装不同的tale模块,研究人员可以针对任何核苷酸序列和控制内源性基因的表达。此外,foki核酸酶结构域融合c-末端两对合成tales可以提供有效的dna序列特异性核酸内切酶活性。正常情况下,可识别目标区域左右两臂的两个talen形成一个功能性foki二聚体,并导致目标区域内的dna双链断裂(dsb),引起非同源末端连接修复或同源重组 修复,从而导致核苷酸的不匹配,插入,或缺失,和功能性基因敲除的引入。
基于现有技术有关成骨细胞在骨形成和骨相关疾病的潜在作用研究的基础,本申请的发明人拟提供在成骨细胞中的bk通道蛋白的的作用,尤其涉及bk通道蛋白在用于制备成骨细胞增殖分化异常干预药物中的用途。
与本发明有关的现有技术有:
1.bentzen,b.h.,olesen,s.p.,ronn,l.c.andgrunnet,m.(2014)bkchannelactivatorsandtheirtherapeuticperspectives.frontiersinphysiology5,389.
2.chen,j.,zhang,w.,lin,j.,wang,f.,wu,m.,chen,c.,zheng,y.,peng,x.,li,j.andyuan,z.(2014)anefficientantiviralstrategyfortargetinghepatitisbvirusgenomeusingtranscriptionactivator-likeeffectornucleases.moleculartherapy:thejournaloftheamericansocietyofgenetherapy22,303-311.
3.ding,q.,lee,y.k.,schaefer,e.a.,peters,d.t.,veres,a.,kim,k.,kuperwasser,n.,motola,d.l.,meissner,t.b.,hendriks,w.t.,etal.(2013)atalengenome-editingsystemforgeneratinghumanstemcell-baseddiseasemodels.cellstemcell12,238-251.
4.zhang,y.y.,yue,j.,che,h.,sun,h.y.,tse,h.f.andli,g.r.(2014)bkcaandheag1channelsregulatecellproliferationanddifferentiationinhumanbonemarrow-derivedmesenchymalstemcells.journalofcellularphysiology229,202-212.
5.zhaowang,j.l.,huanghuang,ganchengwang,mingjunjiang,shenyiyin,changhongsun,,hanshuozhang,fengfengzhuang,jianzhongjeffxi(2012)anintegratedchipforthehigh-throughputsynthesisoftranscriptionactivator-likeeffectors.angewcheminted51,,8505-8508.。
技术实现要素:
本发明的目的在于提供bk通道蛋白在制备成骨细胞干预药物中的用途,尤其是bk通道蛋白在制备成骨细胞增殖分化异常干预药物中的用途。
基于bk参与若干重要的生理功能,如神经兴奋组织、心脏和肌肉,而在骨组织中,成骨细胞bk通道报道鲜见。本发明建立了大电导钙激活钾通道(又称bk通道蛋白或bk通道)敲除和成骨细胞表型与降低成骨细胞的功能的联系模型,利用该模型模拟成骨细胞的病理状态以及进行成骨细胞在骨相关疾病中功能的研究;使用转录激活因子样 效应核酸酶(talen)技术对大鼠ros17/2.8成骨细胞bk通道进行敲除,并检测bk敲除细胞系额增殖矿化,观察成骨细胞bk通道的作用,实验结果显示,bk敲除克隆后增殖和矿化明显下降,成骨细胞分化标志基因的总体表达明显低于野生型成骨细胞。
本发明设计了在大鼠成骨细胞ros17/28上kcnma1(nm_031828)基因靶向的talens,并成功地建立了bk敲除细胞系;该敲除细胞系呈现了与野生型细胞相比较低的增殖和分化能力,结果表明,bk通道是调节成骨细胞增殖与骨相关疾病的治疗的重要靶点,所述的bk通道可用于制备成骨细胞干预药物,尤其可用于制备成骨细胞增殖分化异常干预药物。
本发明中,bk敲除细胞可作为成骨细胞功能研究的工具。
本发明中,进行了包括以下步骤的实验:
(1)细胞培养与试剂;
(2)talens的设计和构建;
(3)pcr扩增与dna序列分析序列;
(4)基因敲除克隆的建立;
(5)mrna表达的分析;
(6)免疫荧光染色;
(7)矿化;
(8)茜素红s染色;
(9)cck8测定;
(10)统计分析。
实验结果显示:
(1)bk敲除克隆在ros17/2.8细胞系的构建
为了在ros17/2.8细胞建立bk基因克隆,本申请实验中将talens转入ros17/2.8并加入嘌呤霉素进行药筛,剩余的细胞被稀释成单细胞培养,在96孔板生长成单克隆;kcnma1基因敲除克隆用免疫荧光显微镜和dna序列进行筛选;基因测序显示在靶点位置之间和之后有重叠峰出现,表明talen靶向成功;免疫荧光分析显示bk蛋白在所有克隆完全消失;
(2)bk基因敲除对ros17/28细胞增殖分化的影响
采用cck-8细胞增殖实验研究kcnma1基因敲除对ros17/2.8细胞增殖的影响, 结果显示,ko细胞群增殖率明显低于野生型细胞群(如图1所示);
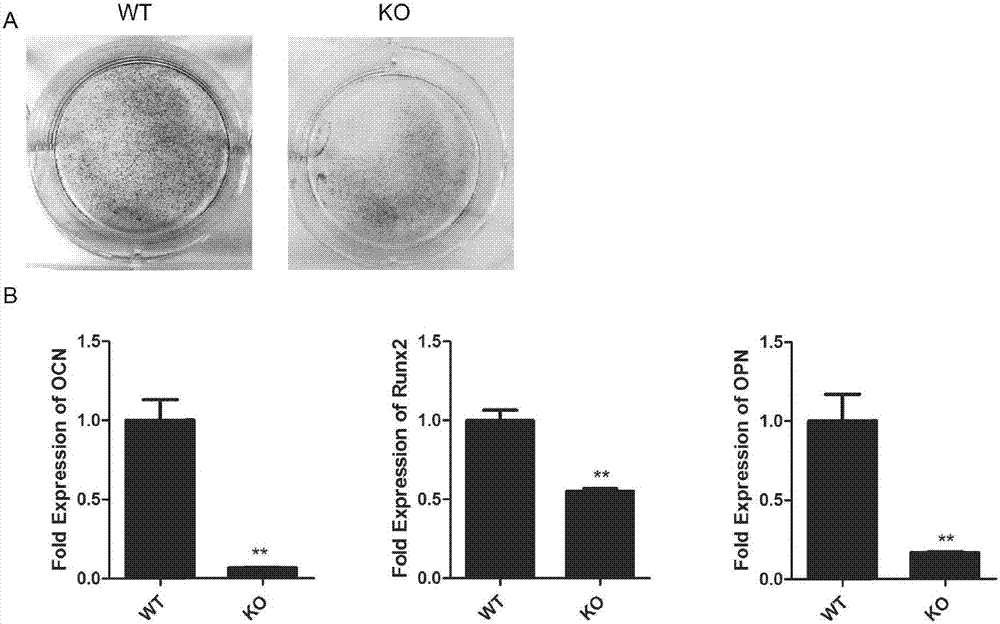
通过比较野生型和基因克隆矿化结节形成研究成骨细胞的分化,结果表明(如图2所示)ros17/2.8细胞bk敲除后的矿化能力显著下降;继续进行成骨转录因子和成骨细胞分化的标记的表达实验,包括runx2的表达骨钙素(ocn)、骨桥蛋白(opn),运用实时荧光定量rt-pcr方法分析在ros17/2.8细胞kcnma1基因敲除,结果显示,runx2的mrna的表达,ocn和opn与野生型相比显著减少,表明成骨细胞的分化能力下降。
实验结果表明,bk通道可作为调节成骨细胞增殖与骨相关疾病的治疗的重要靶点,所述的bk通道可用于制备成骨细胞干预药物,尤其可用于制备成骨细胞增殖分化异常干预药物。
附图说明
图1:ros17/2.8细胞系bk敲除克隆的建立,其中,
a)脱氧核糖核酸序列图,重叠峰显示在靶点位置之间和之后;
b)免疫荧光染色,野生型(wt)和kcnma1基因(ko)的bk敲除克隆,刻度尺=5μ。
图2:kcnma1基因对成骨细胞分化的影响,其中,
a)在kcnma1基因敲除细胞和野生型细胞矿化结节形成,细胞在培养10天之后,矿化茜素红s染色;
b)在kcnma1基因敲除细胞和野生型细胞的成骨细胞标志物mrna的表达,成骨转录因子mrna的表达(runx2)、成骨细胞标志物(骨钙素(ocn)、骨桥蛋白(opn))的实时rt-pcr分析,平均±标准差,n=3,p<0.05。
具体实施方式
实施例1
1、材料和方法
(1)细胞培养与试剂
ros17/2.8细胞(johnshopkins),细胞在ham’sf-12营养混合物培养基(sigmaadrich) 中培养,其中含有碳酸氢钠,辅以10%胎牛e血清(fbs、gibco)处于5%的二氧化碳和95%的空气的培养箱中,每两天传代培养,在密度达到90%时,细胞用胰蛋白酶-edta消化并接种于6孔培养板;
大鼠抗bk多克隆抗体(ap107)均alomone,e.z.n.a.
(2)talens的设计和构建
talen按由sidansai生物技术运用fasttaletm组装的试剂盒的详细说明构建(sidansai,中国),kcnma1靶向的talen左臂和右臂在网站(https://tale-nt.cac.cornell.edu/)设计,聚合酶链式反应(pcr)扩增进行聚集talen,选取各臂的12个克隆并用e.z.n.a.
(3)pcr扩增与dna序列分析序列
基因组dna提取试剂盒提取基因组dna,进行pcr扩增,
正向引物为5’-atatccacgcgaaccatctcagcct-3’,
反向引物5’-ggccgtagccaccaataaccctaca-3’,
pcr扩增在cfx96-pcr仪(bio-rad实验室hercules,ca)运行,95℃下,初始激活为30秒,42次循环变性(95°,5秒),退火(60℃,34秒),延伸(72℃,30秒);pcr产物由sangonbiotech(上海)分析;
(4)建立基因敲除克隆
为了构建bk敲除克隆,将talen质粒转染至ros17/2.8细胞上,然后从转染后的第二天起用4μg/ml嘌呤霉素给药3天,药筛后其余的克隆细胞恢复成长1周,形成单个的细胞克隆团;细胞克隆团通过免疫荧光染色和dna序列分析筛选建立kcnma1基因敲除的细胞系;
(5)免疫荧光染色
bk敲除克隆和野生型细胞均接种在细胞皿中,70%汇合程度时,细胞用4%多聚甲醛固定15分钟,pbs冲洗后,细胞yong0.1%的tritonx-100透析10分钟,3%牛血清白蛋白(bsa)室温p封闭1小时,bk蛋白抗体(1:200,abcam,英国)4℃彻夜培养,荧光二抗alexafluor647抗体(jacksonimmunoresearch,美国)室温条件下孵育1小时,使用leicatcssp5共聚焦系统聚焦显微镜(leica,wetzlar,德国)捕捉图像;
(6)矿化
ros17/2.8细胞接种于48孔板。密度达到90%左右后,将β-甘油磷酸钠(10mm)和l-抗坏血酸磷酸酯(50μg/ml)添加到培养基中作为矿化培养基,矿化培养基每三天更换一次,10天后,细胞用pbs洗两次准备茜素红染色;
(7)茜素红s染色μ
细胞用4%多聚甲醛处理,室温下固定10min,再用pbs轻柔地洗涤三次,每皿添加500μl0.1%茜素红s(yeasen,上海)室温染色15min;染料吸除后,细胞用蒸馏水500μl洗三次并振荡5min;
(8)cck8测定
bk基因敲除细胞接种于浓度为2000细胞/孔板的96孔板,在接种之后第2,3,4,5,7和9天,加入10μlcck8溶液,37℃培养2小时,用酶标仪(tecaninfinitem200pro,瑞士)波长450nm处测定吸光度;
bk敲除克隆在ros17/2.8细胞系的构建结果显示,为了在ros17/2.8细胞建立bk基因克隆,将talens转入ros17/2.8并加入嘌呤霉素进行药筛,剩余的细胞被稀释成单细胞培养,在96孔板生长成单克隆;kcnma1基因敲除克隆用免疫荧光显微镜和dna序列进行筛选,基因测序显示在靶点位置之间和之后有重叠峰出现,表明talen靶向成功;免疫荧光分析显示bk蛋白在所有克隆完全消失;
bk基因敲除对ros17/28细胞增殖分化的影响结果显示,采用cck-8细胞增殖实验证实kcnma1基因敲除对ros17/2.8细胞增殖有影响,ko细胞群增殖率明显低于野生型细胞群(如图1所示);通过比较野生型和基因克隆矿化结节形成研究成骨细胞的分化,结果如图2所示,ros17/2.8细胞bk敲除后的矿化能力显著下降;进一步研究成骨转录因子和成骨细胞分化的标记的表达,包括runx2的表达骨钙素(ocn)、骨桥蛋白(opn),运用实时荧光定量rt-pcr方法分析在ros17/2.8细胞kcnma1基因敲除,结果显示,runx2的mrna的表达,ocn和opn与野生型相比显著减少,表明成骨细胞的分化能力下降。
- 还没有人留言评论。精彩留言会获得点赞!