高度稳定且抗蛋白酶的大肠杆菌天冬酰胺酶的制作方法
发明领域本发明可应用于医学领域,特别是用于治疗肿瘤,更具体地说,用于治疗急性淋巴细胞白血病(all)。
背景技术:
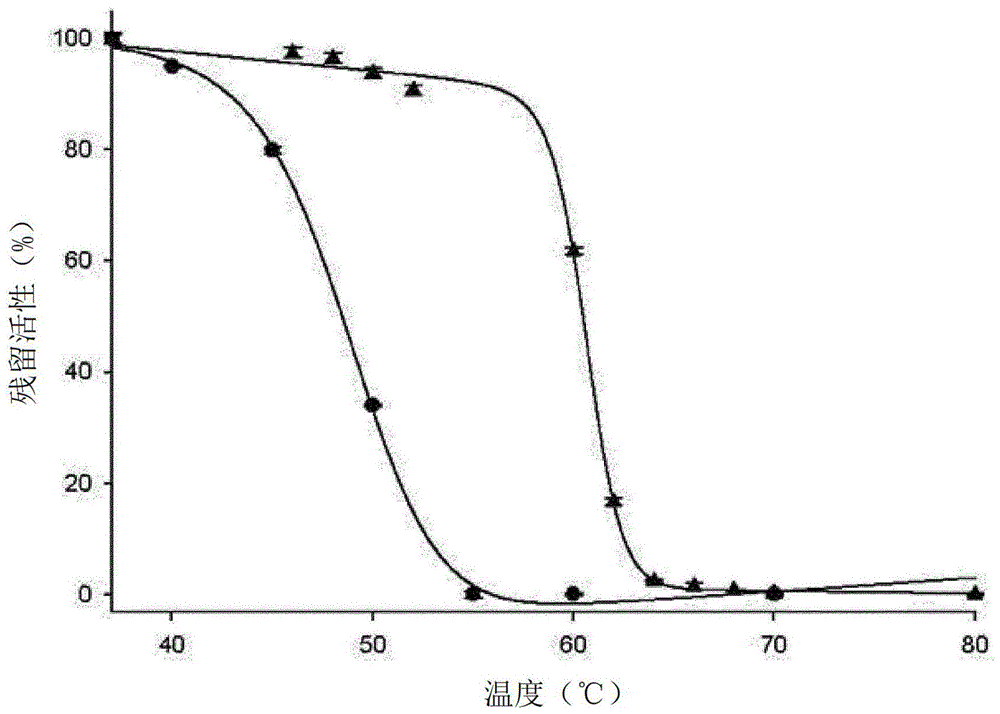
:细菌ii型l-天冬酰胺酶(asnase)是酰胺水解酶(ec3.5.1.1),其催化l-天冬酰胺(l-asn)水解成l-天冬氨酸(l-asp)和氨。由于效率降低,asnase还能够使l-谷氨酰胺(l-gln)脱氨基释放l-谷氨酸(l-glu)和氨。在结构上,asnase是大约130-140kda的同源四聚体。asnase单体具有两个完全不同的结构域,n-结构域(大约230个残基)和c-结构域(大约110个残基),通过有序环(orderedloop)连接。在四聚体折叠中,每个单体获得两组接触:一组较少延伸,代表疏远二聚体(distantdimer),另一组较多延伸,代表紧密二聚体(intimatedimer)。后一种二聚体是asnase的功能单元,两种紧密二聚体中的每一种在亚基界面处具有两个活性位点。每个活性位点由主要属于每个单体n-结构域的残基构成,并且在较低程度上由第二个同源单体的c-结构域构成。n结构域包含活性位点柔性环(残基3-27),这是一种特殊的结构元件,其作为盖子,在底物结合时封闭在活性位点上,带来两个必需的催化残基(thr和tyr)。asnase催化残基由两个高度保守的三联体(thr-asp-lys和thr-tyr-glu)描述,其中苏氨酸起主要作用[1]。l-天冬酰胺酶已被分离并在20世纪50年代早期被描述为抗增殖剂[2],从那时起,细菌已被用作酶药物的主要来源。asnase抗增殖机制依赖于从血液中迅速去除l-天冬酰胺,并且主要影响在l-asn从头生物合成途径中经常受损的肿瘤细胞。由于l-asn饥饿和没有从头生产,肿瘤细胞激活促凋亡和自噬途径[3]。直到几年前,asnase的二级l-谷氨酰胺酶活性被认为与药物毒性相关性较低,甚至有害,因为l-gln去除被认为是健康组织中asnase毒性的主要原因。如今,已经重新评估了l-gln去除在asnase抗肿瘤活性中的作用,并且已经在文献中报道了越来越多支持酶双重活性需求的证据[4-6]。l-天冬酰胺酶被用作儿科急性淋巴细胞白血病(all)治疗的一线药物已有40多年了。它们的巨大功效和特殊的饥饿活性(starvingactivity)使它们不仅对改善all疗法而且对建立新的和更有效的其他肿瘤治疗产生了极大的兴趣。不幸的是,基于asnase的疗法具有一些局限性,主要与以下方面相关:(i)代谢副作用,(ii)高免疫原性,(iii)成人患者的低效率,和(iv)体内高度不稳定性。在儿科患者中,抗asnase抗体的早期发展与治疗失败的高可能性相关,反过来又与患者的负面结果相关。此外,asnase免疫原性是其在成人患者中使用的主要限制,因为成人免疫系统对外源性分子的反应性高于儿童的免疫系统。产生抗asnase抗体的方法并不完全成功,因为它们可能引起过敏反应,常见的是轻微而不严重的,或沉默灭活药物。后一种现象是非常危险的,因为它导致药物的早期失活,没有任何症状或明显的表现,这表明缺乏药理学功效。目前,大肠杆菌(ecaii)和欧文氏菌(erwiniachrysanthemi)(eraii)ii型asnase可用于临床。天然或聚乙二醇化形式的ecaii用作首选药物;相反,eraii仅用于对大肠杆菌衍生的asnase的免疫反应。不幸的是,两种蛋白质的特征在于高的体内不稳定性、短半衰期和获得药理学活性浓度需要多次给药。此外,蛋白质不稳定性与抗体产生的可能性增加有关,反过来又与免疫反应有关。asnase临床应用考虑的另一个方面是分子对蛋白酶介导的降解的易感性。2009年,patel等[7]描述了两种能够通过水解灭活ecaii的溶酶体蛋白酶。两种蛋白酶是组织蛋白酶b(ctsb)和天冬酰胺内肽酶(aep),并且可以通过损害药物活性和药代动力学的白血病淋巴母细胞过表达。此外,eraii也导致对ctsb蛋白水解降解敏感。例如,在受费城阳性(ph+)和iamp21白血病影响的患者中报道了aep和ctsb的高表达,这是非常具有攻击性且较差反应性的白血病形式。在试图预防asnase免疫原性并同时改善分子稳定性的过程中,天然ecaii和eraii蛋白已通过peg化进行化学修饰。聚乙二醇化的ecaii(pegaspase)和最近聚乙二醇化的eraii(即pegcrisantaspase)常规用于临床。然而,pegaspase可能引起过敏反应,即使与原生ecaii相比频率降低,此外它还可引起高甘油三酯血症,导致严重的胰腺炎和代谢功能障碍。此外,pegaspase对天然酶的ctsb水解具有相同的易感性,阻止其用于治疗asnase灭活肿瘤。作为peg化天冬酰胺酶的替代物,最近已在临床中引入了加载到患者衍生的红细胞(graspa)中的天然形式的ecaii(ii/iii期临床试验)。新的药物递送系统似乎在降低asnase免疫原性和改善其半衰期方面非常成功,并且不仅在儿科患者中而且在成人患者中可以成为更安全的asnase递送的有效新平台。考虑到上述情况,认为需要进一步的天冬酰胺酶,其特征在于改善的热和蛋白水解抗性。技术实现要素:本发明基于以下发现:大肠杆菌ii型l-天冬酰胺酶的变体具有特别理想的性质,使其适用于改进的基于asnase的肿瘤疗法。发明目的根据第一个目的,本发明公开了一种大肠杆菌ii型天冬酰胺酶的变体,其特征在于具有对应于seq.id.n.6的序列。在另一个目的中,公开了制备本发明蛋白质的方法。根据第二个目的,本发明的天冬酰胺酶被公开为药物,特别是作为治疗肿瘤的药物。用作药物的用途适用于人类和动物。本发明的另一个目的是用于治疗肿瘤的方法,该方法包括给需要的患者施用适量的所公开的大肠杆菌ii型天冬酰胺酶的步骤。治疗方法适用于人类和动物。包含本发明蛋白质的药物制剂代表本发明的另一个目的。在另一个目的中,本发明公开了一种降低食品(特别是油炸食品)中丙烯酰胺含量的方法。附图说明图1显示了蛋白质稳定性测试的结果;图2显示了热变换分析的结果;图3报告了野生型和n24sasnase对sd1(图a)和reh细胞(图b)的影响结果;图4显示了sd1细胞裂解物(ponceaured染色)的ecaii野生型、n24s和elspar降解;图5报告了野生型和n24s突变体4周时的降解模式;图6说明了分别在野生型和突变蛋白中含有n24残基和s24残基的环的结构。具体实施方式根据第一个目的,公开了大肠杆菌ii型天冬酰胺酶(n24s突变体)的变体,其特征在于具有下述seq.id.n.6的氨基酸序列:特别是,该蛋白质由对应于下述seq.id.n.5的核酸编码:根据本发明的一个具体实施方案,所公开的蛋白质可以用合适的聚合物进行化学修饰。优选地,所述聚合物由聚乙二醇(peg)表示。出于本发明的目的,天冬酰胺酶peg化通过单甲氧基聚乙二醇(peg,分子量小于或等于5000da)与天然蛋白质的共价连接而获得[8]。通过将天然蛋白质与结合蛋白质表面上的特定残基的化学活化的peg混合来获得生物共轭。在另一个实施方案中,本发明的n24s蛋白可以包封在红细胞内。特别地,参考了众所周知的graspa技术。红细胞(rbc)中的包封是用于更安全地施用循环药物的众所周知的方法[9]。最近,临床上可获得包封在rbc中的商业形式的l-天冬酰胺酶(graspabyeritech)。包封技术包括源自患者血液的rbc的瞬时渗透压休克。红细胞渗透通过逐渐渗析进行,并且由于膨胀和毛孔打开而引起细胞膜的瞬时渗透。膨胀的rbc对l-天冬酰胺酶是可渗透的,其使用打开的孔进入细胞。随后,细胞恢复等渗性,l-天冬酰胺酶作为活性分子在细胞中持续存在[10]。将包封的rbc输注到患者体内,并且在溶血后,将l-天冬酰胺酶释放到患者血清中。用作l-天冬酰胺酶递送的生物反应器的rbc允许改善药物的稳定性以及由于患者循环系统中游离药物的短暂持久性而降低免疫原性。在另一个目的中,公开了制备本发明蛋白质的方法。特别地,本发明蛋白质的制备包括培养合适的宿主细胞的步骤,所述宿主细胞已经用包含对应于seq.id.n.5的编码序列的载体转化。在适当的培养基中,然后回收提取物并纯化蛋白质。根据本发明,可以在包括细菌、酵母或哺乳动物细胞的组中选择合适的宿主。在优选的实施方案中,宿主由大肠杆菌代表。在更优选的实施方案中,大肠杆菌细胞是菌株bl21(de3)的细胞。特别地,蛋白质的制备包括使用正义和反义引物,其特征在于具有以下序列:根据第二个目的,本发明的天冬酰胺酶被公开作为药物。更具体地,在下文中,本发明的天冬酰胺酶旨在是以下任何一种形式:-n24s蛋白,-聚乙二醇化的n24s蛋白,-包封在红细胞内的n24s蛋白,如上所述。特别地,天冬酰胺酶可用作治疗原发性和继发性实体瘤的药物。根据一个具体方面,实体瘤用于指选自下组的瘤形成:胃肠道瘤、肺肿瘤、乳腺肿瘤、前列腺肿瘤、卵巢肿瘤、肝肿瘤、肾肿瘤、甲状腺肿瘤、鼻咽肿瘤、肉瘤、淋巴瘤和多发性骨髓瘤。患者可由成人或儿童代表。在优选的实施方案中,该蛋白质可用于治疗急性淋巴细胞白血病(all)。作为更优选的实施方案,急性淋巴细胞白血病可以是费城阳性(ph+)或iamp21白血病。在一个具体实施方案中,急性淋巴细胞白血病的治疗通常在年龄小于18岁的儿童上进行。出于本发明的目的,用作本发明蛋白质的药物不仅适用于人类,也适用于动物。本发明的另一个目的是用于治疗肿瘤的方法,该方法包括给有需要的患者施用合适量的上述大肠杆菌ii型天冬酰胺酶的步骤。更具体地,该方法可包括给予上述本发明的天冬酰胺酶蛋白质中的任何一种:-n24s蛋白,-聚乙二醇化的n24s蛋白,-n24s蛋白包裹在红细胞内。特别地,所述肿瘤包括原发性和继发性实体瘤,并且可以影响成人和儿童。根据另一个具体实施方案,实体瘤用于指选自下组的瘤形成:胃肠道瘤、肺肿瘤、乳腺肿瘤、前列腺肿瘤、卵巢肿瘤、肝肿瘤、肾肿瘤、甲状腺肿瘤、鼻咽肿瘤、肉瘤、淋巴瘤和多发性骨髓瘤。出于本发明的目的,治疗肿瘤的方法不仅适用于人类,也适用于动物。作为本发明的优选方面,本发明的一种方法是用于治疗急性淋巴细胞白血病(all)的方法。作为替代,白血病可以是费城阳性(ph+)或iamp21白血病。在特定实施方案中,患者通常由年龄小于18岁的儿童代表。根据另一个实施方案,本发明的天冬酰胺酶可以与其他药物治疗组合,或者可以在已经进行的或传统治疗之后或作为其替代进行,用于治疗包括急性淋巴细胞白血病(all)的肿瘤。包含本发明蛋白质的药物制剂代表本发明的另一个目的。特别地,包含本发明蛋白质的制剂通过静脉内或肌肉内给药。因此,在本发明的目的范围内的合适的药物制剂包括有效量的本发明的蛋白质(n24s)和一种或多种药学上可接受的载体和/或赋形剂和/或防腐剂。药物制剂可根据不同的药理学方案给药。为了本发明的目的,药物制剂可以包括:-本发明的n24s蛋白质;-通过peg化修饰的本发明的n24s蛋白质;-已包封在红细胞内的本发明的n24s蛋白质。特别地,任选通过peg化学修饰的本发明的n24s蛋白质是上文公开的蛋白质。至于包封的蛋白质,参考众所周知的graspa技术。其中具有本发明的包封的n24s蛋白的红细胞的药物制剂代表了本发明的另一个目的。出于本发明的目的,除了上述形式的本发明蛋白质外,药物制剂还可包含一种或多种化学治疗药物。特别地,所述药物可选自:氨基戊二酰亚胺、安吖啶、阿那曲唑、天冬酰胺酶、苄乙酸、比卡鲁胺、博来霉素、硼替佐米、布舍瑞林、白消安、喜树碱、卡培他滨、卡铂、卡非佐米、卡莫司汀、苯丁酸氮芥、氯喹、顺铂、克拉屈滨氯膦酸盐、秋水仙碱、环磷酰胺、环丙孕酮、阿糖胞苷、达卡巴嗪、放线菌素、柔红霉素、去甲氧基嘧啶、二氯乙酸、二烯雌酚、己烯雌酚、多西紫杉醇、阿霉素、表阿霉素、雌二醇、雌莫司汀、依托泊苷、依维莫司、依西美坦、非格司亭、氟达拉滨、氟氢可的松、氟尿嘧啶、氟甲哌酮、氟他胺、吉西他滨、染料木黄酮、戈舍瑞林、羟基脲、伊达比星、异环磷酰胺、伊马替尼、干扰素、伊立替康、铁三嗪、来那度胺、来曲唑、甲酰四氢叶酸、亮丙瑞林、左旋咪唑、洛莫司汀、氯尼达明、氮芥、甲羟孕酮、甲地孕酮、美法仑、巯嘌呤、美司钠、二甲双胍、甲氨蝶呤、丝裂霉素、米托坦、米托蒽醌、尼罗替丁、诺氟唑、奥曲肽、奥沙利铂、紫杉醇、帕米膦酸、培美曲塞、喷司他丁、哌嗪、普利霉素、泊立度胺、卟吩、哌拉必利、雷替曲塞、利妥昔单抗、索拉非尼、链脲佐菌素、舒尼替尼、苏拉明、他莫昔芬、替莫唑胺、替西罗咪、替米罗甙、睾酮、沙利度胺、硫鸟嘌呤、塞替派、二氯二茂钛、拓扑替康、曲妥珠单抗、tretinoin、长春碱、长春新碱、长春地辛、长春瑞滨。作为本发明的另一个目的,公开了一种降低食品中丙烯酰胺含量的方法。在优选的实施方案中,所述食物由淀粉基食物代表。特别地,淀粉基食物包括例如马铃薯条,法式薯条,马铃薯片或炸丸子,含有选自黑麦、玉米(corn)、玉米(maize)、大麦、大米和燕麦的谷物的产品,小麦基产品。其他食品可以由咖啡、可可、坚果、蔬菜(例如芦笋)或水果(例如香蕉)代表。因此,减少食品加工中,特别是淀粉基食品加工过程中丙烯酰胺产量的方法包括在加工前将本发明的酶施用于食品的步骤。特别地,食物可以是之前列出的食物,而加工包括例如烘焙、油炸、煮沸或烘烤或在120℃或甚至更高温度下进行的任何其他加工。更详细地说,优选将酶应用于食品,例如,通过使酶与食物接触或通过在加工前将食物浸入含有酶的溶液中适当的时间,以允许本发明的酶发生作用并减少食物中天冬酰胺的含量。因此,本发明的蛋白质可用作食品添加剂。在以下实验部分中,本发明的蛋白质是指n24s突变体。另外,l-天冬酰胺酶的单位定义为在所述条件下将1微摩尔底物转化为产物所需酶的量。材料和方法1.突变基本原理设计了n24s突变体以获得改进的蛋白质。2.克隆来自大肠杆菌菌株k-12亚株mg1655的ansb基因的核苷酸序列获自ncbi核苷酸数据库(ncbi参考序列:nc_000913.2)。根据jennings和beacham[11]选择菌株。该序列含有用于分泌信号的63个核苷酸长的前导序列(leadersequence,ls),该分泌信号不包括在克隆的插入物中。通过使用合成形式的全长基因(pmaecansb,geneart)作为模板的pcr,获得在其5′端含有编码6xhis的18个密码子的ansb构建体。反应混合物包含:1x反应缓冲液(invitrogen),300μmdntp(sigma-aldrich),100ng模板(pmaecansb),300nm引物(sigma-aldrich,表1,其中带下划线的字母表示不存在于原始序列中的额外核苷酸),2.5uplatinumpfxdna聚合酶(invitrogen)。反应体积为50μl。获得的扩增产物用于克隆表达载体中的ansb基因。包含无ls的ansb基因和具有n-末端6xhis标签的载体用作定点诱变的模板。用于产生n24s突变体的引物报告于表2中,其中粗体字母表示突变密码子。带下划线的字母表示替换的核苷酸。pcr反应混合物含有:1x反应缓冲液(agilent),300μmdntp(sigma-aldrich),100ng模板,300nm引物(sigma-aldrich),2.5upfuultradna聚合酶(agilent)。反应体积为50μl。表1正义链引物caccatgcatcatcatcatcatcatttacccaatatcaccattttagc反义链引物caatctcgagttattattagtactgattgaagatctgctgg表23.蛋白质过表达和纯化为了获得重组蛋白过表达,使用大肠杆菌bl21(de3)δansa/ansb菌株。该菌株由douglasscottmerrell博士(universityoftheuniformedservicesuniversityofthehealthsciences,bethesda,md,usa)定制设计以缺少ansa和ansb基因,其分别编码大肠杆菌内源性i型和ii型l-天冬酰胺酶。根据studier[12],通过自诱导获得蛋白质过表达。将细胞提取物以2.0ml/min应用于在na缓冲液(50mm磷酸钠,300mmnacl,10mmimohph8.0)中平衡的histrap5ml柱。使用梯度梯度洗脱蛋白质(25、50、100、250、500mmimoh)以及收集5ml级分。检查级分的l-天冬酰胺酶活性,并使用12%丙烯酰胺/双丙烯酰胺凝胶通过sds-page分析。合并活性为阳性的级分,并使用在相同缓冲液中平衡的50ml脱盐柱(gehealthcare),相对于50mm磷酸钠(ph7.4)进行渗析。将洗脱的蛋白质样品应用于在qa缓冲液(50mm磷酸钠,ph7.4)中平衡的1mlhitrapqxl柱。收集流出级分,使用高盐浓度的洗脱缓冲液(50mm磷酸钠,1mnaclph7.4)洗脱结合的蛋白质,并收集10ml级分。检查流过物和结合的级分以获得l-天冬酰胺酶活性。使用12%丙烯酰胺/双丙烯酰胺凝胶通过sds-page分析等分试样。酶活性阳性和纯至均一的级分用于以下实验。使用50ml脱盐柱,相对于磷酸盐缓冲液(pbs,0.01mna/k-磷酸盐缓冲液,0.0027mkc1和0.137mnacl,ph7.4),对用于细胞毒性测定的蛋白质进行渗析。使用相同的柱,相对于ph8.0的0.03mtris-hcl,对用于结晶的蛋白质进行渗析。还通过sec在50mm磷酸钠(ph7.4)、100mmnacl中平衡的superose12hr10/300柱(gehealthcare)上分析样品。在sec之前,使用具有适当截止值的离心过滤器(amicon,merckmillipore)浓缩样品。4.l-天冬酰胺酶和l-谷氨酰胺酶活性测定为了确定l-天冬酰胺酶和l-谷氨酰胺酶活性,采用两种方法:使用谷氨酸脱氢酶(gadh)催化的偶联反应的分光光度法和使用nessler试剂的比色法。用于通过gdh偶联反应测量氨释放的标准反应混合物包含:50mm4-(2-羟乙基)哌嗪-1-乙磺酸(hepes)缓冲液ph7.5,1mmα-酮戊二酸(α-kg),0.24mmnadh,20u谷氨酸脱氢酶和10mml-asn或10mml-gln,终体积为0.5ml。通过添加酶溶液(5-300μg)开始反应,并使用单射线分光光度计在340nm监测nadh消耗。使用nessler试剂测定氨的标准方案进行调整,以便在96孔板中进行分析。简言之,将含有50mmhepesph7.5,0-10mml-asn或0-10mml-gln和5-30μg酶溶液的溶液在37℃温育15分钟以进行l-天冬酰胺酶活性测定或温育30分钟以进行l-谷氨酰胺酶活性测定。然后在冰上加入0.075m三氯乙酸(tca)保持15分钟来停止反应。离心(14000×g,15分钟)后,用nessler试剂对10μl反应物进行分析。根据制造商的说明,将nessler试剂稀释10倍,并将90μl工作溶液加入到10μl反应中,最终体积为100μl。在所有情况下,建立不含酶的阴性对照以确定由于从基质中自发释放氨而产生的背景噪音。为了确定氨的量,使用硫酸铵(10-0.005mm)作为氨源建立标准曲线。每个硫酸铵分子释放2分子氨。动力学参数km通过lineweaver-burk图使用用于sigmaplot(spssinc.)的enzymekineticmodule1.1测定。kcat或转换数是每秒催化事件的数量。kmis是反应速度为最大值的一半时的底物浓度。kcat/kmis是亚饱和底物浓度下催化效率的指标。5.细胞系用于实验的人急性淋巴细胞白血病(all)细胞列于表3中。molt-4、rs4;11、sd1和reh细胞的实验在内分泌和肥胖实验室-萨班(saban)研究所-儿童医院(洛杉矶,ca,美国)进行。人急性淋巴细胞白血病细胞(molt-4、sd1、reh、rs4;11)在rpmi1640完全培养基(10%胎牛血清、2mml-谷氨酰胺、50μg/ml庆大霉素)中生长。将细胞在37℃,5%co2存在下的潮湿培养箱中培养。表3:人all细胞系特征。6.细胞毒性评估为了比较重组野生型和n24secaii蛋白与商业elspar酶(medac)的细胞毒性,将1×105个细胞(即molt4,rs4;11,reh,sd1)接种于200μl中。完全培养基(rpmi1640,10%胎牛血清,2mml-谷氨酰胺,50μg/ml庆大霉素)。将冻干的asnase蛋白质重新悬浮于完全培养基中,并按照parmentier等[5]将其加入到孔中,每种细胞系的ic50值接近终浓度。每个实验一式三份进行,并在不同的天重复至少3次。盲观察者在72小时通过在孔中研磨细胞并通过在管中向10μl细胞样品中加入2x台盼蓝溶液来进行台盼蓝排除染料计数。仅计数活细胞。为了评估缺乏主要蛋白酶识别位点的n24s突变体的改善的稳定性,评估了其长期细胞毒性作用。选择sd1和reh细胞系作为研究的模型系统,因为sd1细胞过表达参与asnase失活的蛋白酶,而reh细胞则不然。将1×105个细胞接种在24孔板中的1.5ml完全培养基中。对于每种细胞系,用浓度低于ic50值的ecaii野生型和n24s处理细胞。特别地,0.5u/ml和1u/ml分别用于sd1和reh。通过台盼蓝活细胞排除染料在72小时和144小时测量细胞活力。数据表示为活细胞与初始平板(ip)的百分比。通过单因素方差分析和tukey检验比较数据。p值等于或小于0.05被认为是显著的。7.蛋白稳定性测定以t50值来测定蛋白质稳定性。为了确定每种蛋白质的t50,在50mmph7.4的磷酸钠中的酶使用热循环仪在不同温度下温育(37、40、42、44、46、48、50、52、54、56、58、60、70℃)10分钟。温育后,将酶溶液在冰上冷却,并如前所述通过奈氏比色法(nessleisation)测量酶活性。反应混合物含有10mml-asn。使用针对sigmaplot的temperaturemeltingplot(spssinc.)确定t50值。处理的酶残余活性表示为热处理样品相对于未处理样品的百分比。获得的t50值是蛋白质稳定性的良好指标:t50值越高,蛋白质稳定性越高。8.蛋白酶敏感性试验通过离心(3500rpm,5分钟,4℃)收集汇合的sd1细胞,并用pbs洗涤两次。使用1×106个细胞:0.1ml缓冲液比例将洗涤的细胞沉淀重悬于裂解缓冲液(50mm乙酸钠三钠缓冲液,ph4.5,5mmdtt)中。在-80℃下通过4个循环的冻融来裂解细胞。通过离心(8000rpm,10分钟,4℃)澄清细胞裂解物,并通过二辛可宁酸测定(bca,pierce)定量全蛋白质提取物。反应混合物的最终体积为50μl,并含有:100μg细胞裂解物和4μgasnase(即重组ecaii野生型和n24s,以及elspar),在ph4.5的50mm乙酸三钠、5mmdtt缓冲液中。在37℃温育24小时后,向样品中加入lds样品缓冲液(4x,thermofisherscientific)和样品还原剂(10x,thermofisherscientific),并在99℃下煮沸5分钟。使用预制梯度4-12%聚丙烯酰胺凝胶(凝胶,thermofisherscientific)通过电泳进行蛋白质分离。使用2装置(thermofisherscientific)将蛋白质转移到硝酸纤维素膜上。使用ponceaured染料对膜进行染色。9.热转换使用miniopticon实时系统(biorad)进行热变换测定。将在pbs缓冲液中的样品(0.5-1.5mg/ml)与20xsypro橙色染料(invitrogen)混合,在pcr管中终浓度为50μl。以1℃或0.5℃/min的速度将样品离心并从25℃加热至95℃。在517nm处监测荧光。根据以下等式对原始数据进行归一化:fu(t)=f(t)-fn/fd-fn[14],其中fu=给定温度(t)下的归一化荧光,f=给定温度下(t)的荧光,fn=荧光的最小值,fd:荧光的最大值。因此,值为1对应于完全展开的蛋白质。从获得的曲线中,外推每种蛋白质的t0.5。10.蛋白质印迹分析对于蛋白质印迹分析,将蛋白质从凝胶内转移至pvdf0.22μm膜(millipore)。将封闭的膜与1∶7000稀释的hrp缀合的抗his标签抗体(sigma)一起温育。11.结晶如上所述纯化大肠杆菌ii型l-天冬酰胺酶n24s突变体。相对于30mmtris-hcl(ph8.0),渗析纯化蛋白质级分并用于结晶。使用moleculardimensions(md1-03)的稀疏基质筛选进行结晶尝试。使用坐滴蒸气扩散法在21℃下生长晶体,并通过在2μl滴中混合等体积的母液和4mg/ml蛋白质溶液。原始结晶条件包含ph7.5的100mmhepes,10%w/vpeg8000,8%v/v乙二醇。为了改善晶体质量,优化条件以改变缓冲液ph和沉淀剂浓度。在低温安装在尼龙环上时,将待通过x射线分析的晶体浸泡在溶解在母液中的100μml-天冬氨酸(l-asp)中10分钟。12.结构解决方案x射线数据是在法国格勒诺布尔(grenoble)的欧洲同步加速器研究设施(europeansynchrotronresearchfacility,esrf)的光束线id-29处收集的[15]。数据与mosflm[16]整合,并与scala[17]合并。空间群用pointless确定[17]。通过用phenix-mr分子置换[18]并使用来自大肠杆菌野生型天冬酰胺酶(pdbid:3eca)[19]结构的l-天冬酰胺酶的紧密二聚体作为搜索模型来解析该结构。用phenix-refine[18]从没有配体的初始模型和属于活性位点柔性环的残基(残基8-35)开始进行结构精制。在每次精制运行中,手动添加残留物,其中存在透明电子密度。配体中加入了phenix-ligandfit,并使用l-天冬氨酸作为搜索模型[20]。结果1.定点突变使用标准t7引物对含有突变的n-末端6xhis标记的大肠杆菌ansb基因(即n-ansb_n24s)的载体进行测序用于插入(beckmancoulter)。测序后,将具有设计突变并且没有任何其他随机突变的构建体转化到大肠杆菌bl21(de3)δansa/ansb化学感受态细胞中用于蛋白质过表达。2.ecaii纯化在imac之后,n标记的ecaii野生型和n24s蛋白在对应于250mmimoh的级分中与一些污染物一起洗脱。将合并和渗析的级分(50mm磷酸钠ph7.4)上样到1mlhitrapqxl柱上。蛋白质在流过部分(flow-throughfraction)中洗脱,而所有其他污染物在结合部分(boundfraction)内洗脱。野生型和n24s突变体纯度均一,相对于l-天冬酰胺(l-asn)分别具有105±8.14和103.5±10.23u/mg的活性,相对于l-谷氨酰胺(l-gln)分别具有0.89±0.22和0.93±0.20u/mg的活性。使用gadh偶联方法并使用10mml-asn或l-gln作为底物测量比活性。使用在ph7.4的50mm磷酸钠中平衡的superose12hr10/300柱通过分析凝胶过滤评估蛋白质的适当四聚体折叠。蛋白质以13.41ml的体积洗脱为单个良好形状的峰,对应于约136kda(数据未显示)。3.ecaii野生型和n24s:动力学特征ecaii野生型和n24s变体的特征在于其相对于l-asn和l-gln的动力学特征。采用nesslerization方法测定酶michaelis常数(km)。动力学参数报告在表4中。由于用于确定ecaii动力学的nesslerization方法对于vmax外推是不可靠的,因此使用每种底物的酶的比活性计算kcat值。与l-asn和l-gln相比,野生型酶和n24s突变体具有双曲线反应。获得的野生型蛋白质的表观动力学参数类似于文献中公开的那些。该酶对l-asn比对l-gln(对于n24s为x111,对于野生型蛋白为x117)作为底物和类似的动力学参数具有更强的偏好。表4ecaii变体动力学参数4.ecaii野生型和n24s突变体的蛋白质稳定性为了评估ecaii野生型热稳定性,将纯度均一的蛋白质在ph7.4的50mm磷酸钠中在10个不同温度(范围为40-70℃)温育10分钟。温育后,将蛋白质在冰上冷却,并使用nessler方法在10mmasn存在下测量残余活性。残余活性计算为热处理样品相对于未处理蛋白质的活性百分比。解链温度,定义为温育10分钟后活性为未处理样品的一半的温度,测定为t50。ecaii的t50值等于49±1.3℃,n24s突变体的t50等于59±2.2℃(图1)。在+4℃下的ecaii野生型稳定性为1-1.5周,在第9周和第1年测试n24s突变体稳定性,并且在sds凝胶上未发现降解。蛋白质的时间依赖性降解导致酶活性丧失。通过热变换分析评估ecaii野生型和n24s蛋白的热诱导解折叠,使用一半蛋白质处于未折叠状态(t0.5)作为参考参数的温度。n24s突变体相对于野生型蛋白质具有高2.41℃的t0.5值(分别为t0.562.04±0.10℃vs.59.63±0.62℃,图2)。此外,野生型蛋白质具有更快的解折叠动力学,因为未折叠的部分在12℃(相当于7.5%/℃)从10%增加到100%,而n24s突变体具有较慢的解折叠动力学,展开率为5.3%/℃。野生型初始解折叠(即5%未折叠部分)在51℃开始,比n24s突变体(57℃)低6℃。可以通过不同方法评估酶稳定性。在这里,我们分析了ecaii野生型和n24s稳定性,它们都是生物活性保留(t50或残留活性)和解折叠速率(t0.5或热变换)的函数。前一种方法包括在不同温度下对酶进行热处理一段时间(即10分钟),然后立即在冰上冷却,最后进行酶活性评估。当用于评估酶在暴露于升高的温度时维持其功能性折叠的能力时,它是一个很好的参数,因此,较高的t50值是酶功能稳定性的良好指标。在这方面,ecaiin24s的t50值比野生型高10℃(分别为59℃vs.49℃),表明该变体能够在比野生型酶更极端的条件下维持其功能性折叠。n24s变体的这种特征是当作为药物给予患者时其保留活性的能力的非常有前景的象征。此外,在+4℃下储存时评估酶残留活性。n24s变体显示出比野生型蛋白质长8倍的稳定性,证实该变体改善了生物稳定性。还通过热变换评估野生型和n24s蛋白的解折叠率。t0.5δ值低于残留活性评估得到的值(δ,2.41℃vs.10℃)表明n24s改善的稳定性主要与活性位点稳定性而非四聚体稳定性相关。天冬酰胺酶活性位点在蛋白质稳定中起重要作用,特别是因为存在活性位点柔性环(残基3-27),其为含有催化的必需残基的一种移动的结构元件。该观点通过以下观察证实:蛋白质活性丧失与位于sdspage中的降解带的存在成比例,所述降解带位于比单体分子量低约3kda的位置。此外,较低的条带在蛋白质印迹中对抗his标签抗体不反应,表明包含6xhis标签和活性位点环的蛋白质n末端的丢失。进一步的确认来自结构研究。通过x射线分析获得的突变体n24s结构与野生型非常相似,如全局折叠(rms使用标准的pymol比对计算)以及界面溶剂化能量(野生型-291.3kcal/mol,n24s-290.3kcal/mol)和溶剂可及区域(野生型n24s)。在结合(δgint)和解离(δgdiss)能量中发现野生型和突变型四聚体装配之间的显著差异。特别是,n24sδgint的计算值比野生型低了10.1kcal/mol(分别为-50.8vs.-40.7kcal/mol),agdiss变化为4.8kcal/mol(野生型21.5kcal/mol,n24s26.3kcal/mol)。所有计算均使用embl-ebi开放式访问软件pisa完成。5.ecaii野生型和n24s突变体细胞毒性为了比较ecaii野生型和n24s蛋白与商业酶elspar(medac)的体外抗增殖作用,如材料和方法部分所述使用4种all模型细胞系(即molt-4、reh、rs4;11和sd1)进行细胞毒性测定。细胞活力表示为活细胞相对于未处理样品的百分比,并使用ecaii野生型和n24s和elspar浓度(u/ml)评估,该浓度接近parmentier等描述的ic50值[5]。表5中报告了每种细胞系的蛋白质浓度和72小时的活细胞百分比。重组野生型和n24secaii蛋白在除sd1外的所有细胞系上显示出与商业elspar蛋白相当的细胞毒性作用。实际上,相对于商业酶elspar,ecaii蛋白对sd1细胞在3.0u/ml的细胞毒性作用高近4.30倍。还在72和144小时评估了在sd1和reh细胞系上的长期细胞毒性。在72小时,野生型和n24s突变体对sd1和reh细胞具有相同的细胞毒作用。在144小时,ecaii变体n24s对sd1细胞的细胞毒性比野生型高47.5%,而野生型和突变体对reh细胞具有相同的细胞毒性(图2a和2b)。表572h的ecaii野生型、变体和elspar细胞毒性6.蛋白酶敏感性试验在sd1细胞裂解物存在下,将野生型和n24secaii和elspar在37℃o.n.下温育,以确定它们对肿瘤释放的蛋白酶的敏感性。温育后,通过sds-page分离样品中的蛋白质并印迹到硝酸纤维素膜上。在ponceaured染色后,在野生型和elspar样品中明显降解条带,并且在n24s样品中不存在(图4)。实验重复4次,都得到相同的结果。7.储存稳定性通过电泳分离和蛋白质印迹评估ecaii野生型和n24s蛋白的时间依赖性降解。降解后,可以在sds-page中观察到约30kda的清晰降解条带(图5a)。还通过天然page分析等份的完全降解的ecaii,以确定在降解时是否保持四聚体折叠。如图5b所示,完全降解的蛋白质(泳道2)作为单一物种在天然条件下运行,并且相对于新鲜纯化的蛋白质具有降低的分子量(泳道3)。蛋白质印迹分析(图5c)显示完全降解的蛋白质(泳道2)不与针对蛋白质n末端6xhis标签的抗his标签抗体反应。通过使用酪蛋白作为底物和folin-ciocalteu试剂作为比色指示剂的定性测试证实了蛋白酶的缺乏(数据未显示)。8.x射线结构测定在4天后,在21℃下,在ph7.5的100mmhepes,5%w/vpeg8000,4%v/v乙二醇中获得n24s突变体的单棱柱晶体。该条件具有高度可重复性,并且可有效地产生具有良好尺寸和良好衍射的晶体。本文中描述的晶体以的分辨率衍射。晶体不对称单元(asu)含有2个不同的亲密二聚体(即a1/c1和a2/c2);对应于l-asp产物的透明电子密度存在于asu的所有4个活性位点中。4种单体中的两种(即a1和c2)对属于活性位点柔性环(残基3-27)的残基呈现清晰的电子密度,而在单体a2和c1中缺少残基15-26。用丝氨酸取代24位的天冬酰胺导致局部h键重排。特别地,在突变蛋白质中,连接残基tyr25与附近单体的ala282的h键的丢失,有利于tyr25n和ser240oγ之间的新h-键(图6)。用丝氨酸取代天冬酰胺24可防止蛋白酶活性,其结果类似于对相同位置的其他突变体所描述的结果。此外,天冬酰胺侧链的去除可能防止在水存在下在天冬酰胺旁的肽键自发裂解,这是[21]所述的储存蛋白质中常见的不期望的反应。从以上描述中,本发明提供的优点对于本领域技术人员来说是显而易见的。特别地,与已知的野生型蛋白质相比,所公开的天冬酰胺酶已显示出保留功能参数。因此,蛋白质的保留的催化和抗增殖特性(其改善的长期细胞毒性)代表了其体内功效的坚实基础。然而,同时,本发明的蛋白质在培养和储存时具有大大改善的长期稳定性,这代表了在治疗中开发和引入药物的有希望的特征。鉴于收集的数据,可以得出结论,蛋白质的ic50值至少等于或甚至优于野生型。除上述之外,根据体外数据,预期所公开的蛋白质产生较低的肿瘤抗性。后一特征是至关重要的,因为蛋白酶抗性代表了药物特性的巨大改善,因为肿瘤相关蛋白酶可能在很大程度上通过降解asnase和揭示新的免疫原性表位来损害asnase的功效,从而增加了抗体产生和复发的风险。研究表明,蛋白水解活性的增加不仅可能是由于肿瘤的质量,也可能是由于肿瘤周围的细胞。已经在实体瘤中观察到增加的蛋白水解活性,其主要有助于癌发生和肿瘤转移。本发明还包括用peg化修饰蛋白质的可能性。所述化学修饰允许进一步改善n24s突变体特征,例如特别是稳定性,包括热稳定性,以及降低的免疫原性,其反过来导致药物的安全性和功效增强。作为另一种方法,本发明还包括使用graspa技术通过包封入红细胞来装载本发明蛋白质的可能性。众所周知,天冬酰胺在加工过程中(如煎炸或加热至120℃以上)会与淀粉类食品中所含的还原糖发生反应,因此产生丙烯酰胺作为美拉德反应的产物(参见,例如[22,23])。2002年在瑞典进行的研究表明,人们对膳食中丙烯酰胺暴露的担忧已经出现,这表明在各种食品的油炸或烘烤过程中形成了高含量的丙烯酰胺。粮农组织/世界卫生组织食品添加剂联合专家委员会(jecfa)于2005年审查了丙烯酰胺的安全性,并建议在世界各地正在进行的致癌性和长期神经毒性研究的结果可用时,重新评估丙烯酰胺,应继续采取适当措施降低食品中的丙烯酰胺浓度。因此,本发明还在食品工业中发现了其另外的有用的应用。特别地,与其他已知的酶相比,本发明的酶有利地减少了使丙烯酰胺失活所需的时间。参考文献1.maggi,m.和chiarellil.r.和valentinig.和scottic.,engineeringofhelicobacterpyloril-asparaginase:characterizationoftwofunctionallydistinctgroupsofmutants.plosone,2015.10(2):p.e0117025.2.kidd,j.g.,regressionoftransplantedlymphomasinducedinvivobymeansofnormalguineapigserum.ii.studiesonthenatureoftheactiveserumconstituent:histologicalmechanismoftheregression:testsforeffectsofguineapigserumonlymphomacellsinvitro.jexpmed,1953.98(6):p.583--606.3.covini,daniele和maggimaristella和tarditosaverio和bussolatiovidio和chiarellilaurentr.和pasquettomariav.和vecchialuca和valentinigiovanna和scotticlaudia,expandingtargetsforametabolictherapyofcancer:l-asparaginase.topicsinanti-cancerresearch,2015.3:p.418--445.4.chan,w.k.,lorenzi,p.l.,anishkin,a.,purwaha,p.,rogers,d.m.,sukharev,s.,rempe,s.b.,和weinstein,j.n.,theglutaminaseactivityofl-asparaginaseisnotrequiredforanticanceractivityagainstasns-negativecells.blood,2014.123(23):p.3596-606.5.parmentier,j.h.,maggi,m.,tarasco,e.,scotti,c.,avramis,v.i.,和mittelman,s.d.,glutaminaseactivitydeterminescytotoxicityofl-asparaginasesonmostleukemiacelllines.leukres,2015.39(7):p.757-62.6.avramis,v.i.,isglutaminedepletionneededinalldisease?blood,2014.123(23):p.3532-3.7.patel,n.,krishnan,s.,offman,m.n.,krol,m.,moss,c.x.,leighton,c.,vandelft,f.w.,holland,m.,liu,j.,alexander,s.,dempsey,c.,ariffin,h.,essink,m.,eden,t.o.,watts,c.,bates,p.a.,和saha,v.,adyadoflymphoblasticlysosomalcysteineproteasesdegradestheantileukemicdrugl-asparaginase.jclininvest,2009.119(7):p.1964-73.8.fu,ceciliah.和sakamotokathleenm.,peg-asparaginase.expertopiniononpharmacotherapy,2007.8(12):p.1977--84.9.kwon,y.m.,chung,h.s.,moon,c.,yockman,j.,park,y.j.,gitlin,s.d.,david,a.e.,和yang,v.c.,l-asparaginaseencapsulatedintacterythrocytesfortreatmentofacutelymphoblasticleukemia(all).jcontrolrelease,2009.139(3):p.182-9.10.godfrin,y.,thomas,x.,bertrand,y.,和duget,c.,l-asparaginaseloadedintoerythrocytes(graspa):principleandinterestsinacutelymphoblasticleukemia.blood,2007.110(11):p.4325.11.jennings,m.p.和beacham,i.r.,ahalysisoftheescherichiacoligeneencodingl-asparaginaseii,ansb,anditsregulationbycyclicampreceptorandfnrproteins.jbacteriol,1990.172(3):p.1491-8.12.studier,f.w.,proteinproductionbyauto-inductioninhighdensityshakingcultures.proteinexprpurif,2005.41(1):p.207-34.13.ehsanipour,e.a.,sheng,x.,behan,j.w.,wang,x.,butturini,a.,avramis,v.i.,和mittelman,s.d.,adipocytescauseleukemiacellresistancetol-asparaginaseviareleaseofglutamine.cancerres,2013.73(10):p.2998-3006.14.andreotti,g.,monticelli,m.,和cubellis,m.v.,lookingforproteinstabilizingdrugswiththermalshiftassay.drugtestanal,2015.7(9):p.831-4.15.id29:ahigh-intensityhighlyautomatedesrfbeamlineformacromolecularcrystallographyexperimentsexploitinganomalousscattering.j.synchrotronrad.j.synchrotronrad,2012.19(19):p.455--461.16.battye,t.geoffg.和kontogiannisluke和johnsonowen和powellharoldr.和leslieandrewg.w.,imosflm:anewgraphicalinterfacefordiffraction-imageprocessingwithmosflm.actacrystallographica.sectiond,biologicalcrystallography,2011.67(pt4):p.271--81.17.evans,philip,scalingandassessmentofdataquality.actacrystallographica.sectiond,biologicalcrystallography,2006.62(pt1):p.72--82.18.automatingcrystallographicstructuresolutionandrefinementofprotein-ligandcomplexes.actacrystallographicasectiondbiologicalcrystallography,2014.70(1):p.144--154.19.swain,a.l.,jaskólski,m.,housset,d.,rao,j.k.,和wlodawer,a.,crystalstructureofescherichiacolil-asparaginase,anenzymeusedincancertherapy.procnatlacadsciusa,1993.90(4):p.1474-8.20.klei,herberte.和moriartynigelw.和echolsnathaniel和terwilligerthomasc.和baldwinerict.和pokrossmatt和posyshana和adamspauld.和iucr和d.adamsp.和w.grosse-kunstlever.和l.w.hung和r.ioergert.和j.mccoya.和w.moriartyn.和j.readr.和c.sacchettinij.和k.sautern.和c.terwilligert.和j.badger和m.sauderj.和l.blundellt.和h.jhoti和c.abell和j.brunoi.和c.colej.和m.kessler和j.luo和s.motherwellw.d.和h.purkisl.和r.smithb.和r.taylor和i.cooperr.和e.harriss.和g.orpena.和e.carhartr.和h.smithd.和r.venkataraghavan和r.chancem.和p.cordellal.和p.foggia和c.sansone和m.vento和j.das和e.echols和w.moriartyn.和e.kleih.和v.afoninep.和g.bunkczi和j.headdj.和j.mccoya.和r.oeffner和j.readr.和c.terwilligert.和d.adamsp.和p.emsley和b.lohkamp和g.scottw.和k.cowtan和c.finzelb.和r.akavaram和a.ragipindi和r.vanvoorstj.和m.cahn和e.davism.和e.pokrossm.和s.sheriff和t.baldwine.和w.grosse-kunstlever.和v.afoninep.和k.sautern.和d.adamsp.和d.hawkinsp.c.和g.skillmana.和l.warreng.和a.ellingsonb.和t.stahlm.和k.henrick和e.hopcroftj.和m.karpr.和e.kleih.和k.kish和m.linp.f.和q.guo和j.friborg和e.roser.和y.zhang和v.goldfarb和r.langleyd.和m.wittekind和s.sheriff和j.kleywegtg.和a.kumar和h.j.chiu和l.axelrodh.和a.morse和m.a.elsliger和a.wilsoni.和a.deacon和a.lebedeva.和p.young和n.isupovm.和v.morozo.和a.vagina.和n.murshudovg.和c.liu和m.mooijw.t.和j.hartshornm.和j.ticklei.和j.sharffa.和l.verdonkm.和h.jhoti和w.moriartyn.和w.grosse-kunstlever.和d.adamsp.和j.oldfieldt.和j.oldfieldt.和l.quanm.和p.pintod.j.和a.rossik.和s.sheriff和s.alexanderr.和e.amparo和k.kish和m.knabbr.和m.luettgenj.和p.morin和a.smallwood和j.woernerf.和r.wexlerr.和j.rogersd.和t.tanimotot.和w.rosep.和b.beran和c.bi和f.bluhmw.和d.dimitropoulos和s.goodselld.和a.prlic和m.quesada和b.quinng.和d.westbrookj.和j.young和b.yukich和c.zardecki和m.bermanh.和e.bournep.和w.schttelkopfa.和f.vanaaltend.m.和c.terwilligert.和h.klei和d.adamsp.和w.moriartyn.和d.cohnj.和d.weininger和s.wlodek和g.skillmana.和a.nicholls,ligandplacementbasedonpriorstructures:theguidedligand-replacementmethod.actacrystallographicasectiondbiologicalcrystallography,2014.70(1):p.134--143.21.capasso,s.,mazzarella,l.,sorrentino,g.,balboni,g.,和kirby,a.j.,kineticsandmechanismofthecleavageofthepeptidebondnexttoasparagine.peptides,1996.17(6):p.1075-7.22.zyzak,d.v.,sanders,r.a.,stojanovic,m.,tallmadge,d.h.,eberhart,b.l.,ewald,d.k.,gruber,d.c.,morsch,t.r.,strothers,m.a.,rizzi,g.p.,和villagran,m.d.,acrylamideformationmechanisminheatedfoods.jagricfoodchem,2003.51(16):p.4782-7.23.mottram,d.s.,wedzicha,b.l.,和dodson,a.t.,acrylamideisformedinthemaillardreaction.nature,2002.419(6906):p.448-9。序列表<110>帕维亚大学<120>高度稳定且抗蛋白酶的大肠杆菌天冬酰胺酶<130>@e0105819<160>8<170>patentin版本3.5<210>1<211>48<212>dna<213>人工<220><223>引物<400>1caccatgcatcatcatcatcatcatttacccaatatcaccattttagc48<210>2<211>41<212>dna<213>人工<220><223>反义引物<400>2caatctcgagttattattagtactgattgaagatctgctgg41<210>3<211>23<212>dna<213>人工<220><223>正义引物构建体n24s<400>3aaccaaatctagctacacagtgg23<210>4<211>25<212>dna<213>人工<220><223>反义引物构建体n24s<400>4ccactgtgtagctagatttggttgc25<210>5<211>978<212>dna<213>人工<220><223>编码n24s的序列<400>5ttacccaatatcaccattttagcaaccggcgggaccattgccggtggtggtgactccgca60accaaatctagctacacagtgggtaaagttggcgtagaaaatctggttaatgcggtgccg120caactaaaagacattgcgaacgttaaaggcgagcaggtagtgaatatcggctcccaggac180atgaacgataatgtctggctgacactggcgaaaaaaattaacaccgactgcgataagacc240gacggcttcgtcattacccacggtaccgacacgatggaagaaactgcttacttcctcgac300ctgacggtgaaatgcgacaaaccggtggtgatggtcggcgcaatgcgtccgtccacgtct360atgagcgcagacggtccattcaacctgtataacgcggtagtgaccgcagctgataaagcc420tccgccaaccgtggcgtgctggtagtgatgaatgacaccgtgcttgatggccgtgacgtc480accaaaaccaacaccaccgacgtagcgaccttcaagtctgttaactacggtcctctgggt540tacattcacaacggtaagattgactaccagcgtaccccggcacgtaagcataccagcgac600acgccattcgatgtctctaagctgaatgaactgccgaaagtcggcattgtttataactac660gctaacgcatccgatcttccggctaaagcactggtagatgcgggctatgatggcatcgtt720agcgctggtgtgggtaacggcaacctgtataaatctgtgttcgacacgctggcgaccgcc780gcgaaaaccggtactgcagtcgtgcgttcttcccgcgtaccgacgggcgctaccactcag840gatgccgaagtggatgatgcgaaatacggcttcgtcgcctctggcacgctgaacccgcaa900aaagcgcgcgttctgctgcaactggctctgacgcaaaccaaagatccgcagcagatccag960cagatcttcaatcagtac978<210>6<211>326<212>prt<213>人工<220><223>n24s序列<400>6leuproasnilethrileleualathrglyglythrilealaglygly151015glyaspseralathrlyssersertyrthrvalglylysvalglyval202530gluasnleuvalasnalavalproglnleulysaspilealaasnval354045lysglygluglnvalvalasnileglyserglnaspmetasnaspasn505560valtrpleuthrleualalyslysileasnthraspcysasplysthr65707580aspglyphevalilethrhisglythraspthrmetglugluthrala859095tyrpheleuaspleuthrvallyscysasplysprovalvalmetval100105110glyalametargproserthrsermetseralaaspglypropheasn115120125leutyrasnalavalvalthralaalaasplysalaseralaasnarg130135140glyvalleuvalvalmetasnaspthrvalleuaspglyargaspval145150155160thrlysthrasnthrthraspvalalathrphelysservalasntyr165170175glyproleuglytyrilehisasnglylysileasptyrglnargthr180185190proalaarglyshisthrseraspthrpropheaspvalserlysleu195200205asngluleuprolysvalglyilevaltyrasntyralaasnalaser210215220aspleuproalalysalaleuvalaspalaglytyraspglyileval225230235240seralaglyvalglyasnglyasnleutyrlysservalpheaspthr245250255leualathralaalalysthrglythralavalvalargserserarg260265270valprothrglyalathrthrglnaspalagluvalaspaspalalys275280285tyrglyphevalalaserglythrleuasnproglnlysalaargval290295300leuleuglnleualaleuthrglnthrlysaspproglnglnilegln305310315320glnilepheasnglntyr325<210>7<211>978<212>dna<213>大肠杆菌<400>7ttacccaatatcaccattttagcaaccggcgggaccattgccggtggtggtgactccgca60accaaatctaactacacagtgggtaaagttggcgtagaaaatctggttaatgcggtgccg120caactaaaagacattgcgaacgttaaaggcgagcaggtagtgaatatcggctcccaggac180atgaacgataatgtctggctgacactggcgaaaaaaattaacaccgactgcgataagacc240gacggcttcgtcattacccacggtaccgacacgatggaagaaactgcttacttcctcgac300ctgacggtgaaatgcgacaaaccggtggtgatggtcggcgcaatgcgtccgtccacgtct360atgagcgcagacggtccattcaacctgtataacgcggtagtgaccgcagctgataaagcc420tccgccaaccgtggcgtgctggtagtgatgaatgacaccgtgcttgatggccgtgacgtc480accaaaaccaacaccaccgacgtagcgaccttcaagtctgttaactacggtcctctgggt540tacattcacaacggtaagattgactaccagcgtaccccggcacgtaagcataccagcgac600acgccattcgatgtctctaagctgaatgaactgccgaaagtcggcattgtttataactac660gctaacgcatccgatcttccggctaaagcactggtagatgcgggctatgatggcatcgtt720agcgctggtgtgggtaacggcaacctgtataaatctgtgttcgacacgctggcgaccgcc780gcgaaaaccggtactgcagtcgtgcgttcttcccgcgtaccgacgggcgctaccactcag840gatgccgaagtggatgatgcgaaatacggcttcgtcgcctctggcacgctgaacccgcaa900aaagcgcgcgttctgctgcaactggctctgacgcaaaccaaagatccgcagcagatccag960cagatcttcaatcagtac978<210>8<211>326<212>prt<213>大肠杆菌<400>8leuproasnilethrileleualathrglyglythrilealaglygly151015glyaspseralathrlysserasntyrthrvalglylysvalglyval202530gluasnleuvalasnalavalproglnleulysaspilealaasnval354045lysglygluglnvalvalasnileglyserglnaspmetasnaspasn505560valtrpleuthrleualalyslysileasnthraspcysasplysthr65707580aspglyphevalilethrhisglythraspthrmetglugluthrala859095tyrpheleuaspleuthrvallyscysasplysprovalvalmetval100105110glyalametargproserthrsermetseralaaspglypropheasn115120125leutyrasnalavalvalthralaalaasplysalaseralaasnarg130135140glyvalleuvalvalmetasnaspthrvalleuaspglyargaspval145150155160thrlysthrasnthrthraspvalalathrphelysservalasntyr165170175glyproleuglytyrilehisasnglylysileasptyrglnargthr180185190proalaarglyshisthrseraspthrpropheaspvalserlysleu195200205asngluleuprolysvalglyilevaltyrasntyralaasnalaser210215220aspleuproalalysalaleuvalaspalaglytyraspglyileval225230235240seralaglyvalglyasnglyasnleutyrlysservalpheaspthr245250255leualathralaalalysthrglythralavalvalargserserarg260265270valprothrglyalathrthrglnaspalagluvalaspaspalalys275280285tyrglyphevalalaserglythrleuasnproglnlysalaargval290295300leuleuglnleualaleuthrglnthrlysaspproglnglnilegln305310315320glnilepheasnglntyr325当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1