利用蔗糖合酶提高水稻茎秆抗倒伏能力的方法与流程
本发明涉及转蔗糖合酶水稻的分子生物学技术,尤其涉及一种利用蔗糖合酶提高水稻抗倒伏能力的方法。
背景技术:
蔗糖合酶(sucrose synthase,EC 2.4.1.13,SUS)是一种糖基转移酶,催化可逆的反应:一般认为,它在植物体内主要起到蔗糖分解的作用[1,2](Tsai et al.,1970;Su and Preiss,1978)。单子叶模式物种水稻的蔗糖合酶基因家族有6个同源基因,分别命名为OsSUS1-6。大量研究表明SUS参与纤维素和淀粉的合成。如棉花中SUS突变或被抑制,棉花胚珠表皮纤维起始基本被抑制,棉纤维细胞延伸和种子发育受到影响,最终产生少纤维的棉花种子[3,4](Ruan et al.,1998,2003)。在棉花中超表达SUS基因,使棉纤维长度增加,纤维产量增加[5,6](Xu et al.,2012;Jiang et al.,2012)。在木本植物杨树中,超表达棉花SUS基因同样增加纤维素含量和次生细胞壁厚度[7](Coleman et al.,2009)。玉米SUS突变导致胚乳中淀粉含量显著降低,籽粒皱缩、干瘪[8](Chourey and Nelson,1976)。在玉米中转入马铃薯的蔗糖合酶基因StSUS4,转基因玉米的种子中含量增加[9](Li et al.,2013)。对马铃薯SUS基因进行RNA抑制后,块茎中的的SUS活性降低,淀粉含量降低[10](Zrenner et al.,1995)。但迄今为止,尚无蔗糖合酶对提高植物抗倒伏能力的报道。
参考文献:
1.Tsai CY,Salamini F,Nelson OE.Enzymes of carbohydrate metabolism in the developing endosperm of maize.Plant Physiol,1970,46:299-306.
2.Su JC,Preiss J.Purification and properties of sucrose synthase from maize kernels.Plant Physiol,1978,61:389-393.
3.Ruan YL,Chourey PS.A fiberless seed mutation in cotton is associated with lack of fiber cell initiation in ovule epidermis and alterations in sucrose synthase expression and carbon partitioning in developing seeds.Plant Physiol,1998,118:399-406.
4.Ruan YL.Suppression of sucrose synthase gene expression represses cotton fiber cell initiation,elongation,and seed development.Plant Cell,2003,15:952-964.
5.Xu SM,Brill E,Llewellyn DJ,Furbank RT,Ruan YL.Overexpression of a potato sucrose synthase gene in cotton accelerates leaf expansion,reduces seed abortion,and enhances fiber production.Mol Plant,2012,5:430-441.
6.Jiang Y,Guo W,Zhu H,Ruan YL,Zhang T.Overexpression of GhSusA1 increases plant biomass and improves cotton fiber yield and quality.Plant Biotechnol J,2012,10:301-312.
7.Coleman HD,Yan J,Mansfield SD.Sucrose synthase affects carbon partitioning to increase cellulose production and altered cell wall ultrastructure.Proc Natl Acad Sci USA,2009,106:13118-13123.
8.Chourey PS,Nelson OE.The enzymatic deficiency conditioned by the shrunken-1 mutations in maize.Biochem Genet,1976,14:1041-1055.
9.Li J,Baroja-Fernández E,Bahaji A et al.Enhancing sucrose synthase activity results in increased levels of starch and ADPglucose in maize(Zea mays L.)seed endosperms.Plant Cell Physiol,2013,54:282-294.
10.Zrenner R,Salanoubat M,Willmitzer L,Sonnewald U.Evidence of the crucial role of sucrose synthase for sink strength using transgenic potato plants(Solanum tuberosum L.).Plant J,1995,7:97-107。
技术实现要素:
本发明的目的在于克服现有技术存在的缺点和不足,提供一种利用蔗糖合酶提高水稻茎秆抗倒伏能力的方法,即利用转基因技术对水稻进行遗传改良,提高茎秆抗倒伏能力,为提高植物机械支撑力做理论以及技术上的探索。
本发明的目的是这样实现的:
利用蔗糖合酶提高水稻茎秆抗倒伏能力的方法(简称方法)
本方法包括下列步骤:
①从水稻中克隆OsSUS3基因cDNA
水稻蔗糖合酶3(OsSUS3)基因编码区序列如SEQ ID NO:1,cDNA长度为2643bp;
②构建OsSUS3基因在次生细胞壁合成时表达的载体,并转化水稻中花11,获得转基因植株:转OsSUS3基因水稻种子Sus3(Oryza sativa Sus3),于2017年1月3日保藏在中国典型培养物保藏中心(地址:中国武汉武汉大学邮编:430072),保藏编号为CCTCC NO:P201702;
③将获得的转OsSUS3基因的水稻材料开花后30天进行抗倒伏能力的测定增加OsSUS3基因在水稻中的表达量,使转基因水稻抗倒伏能力与对照相比提高了42%。
本发明具有下列优点和积极效果:
1、OsSUS3只在茎秆叶片等次生细胞壁大量合成的器官中增强表达,在胚乳中表达不增加;
2、增加OsSUS3基因在水稻中的表达量,使转基因水稻茎秆抗倒伏能力(开花后30天测定)提高了42%。
附图说明
图1是转OsSUS3基因水稻茎秆的抗倒伏能力的比较图片;
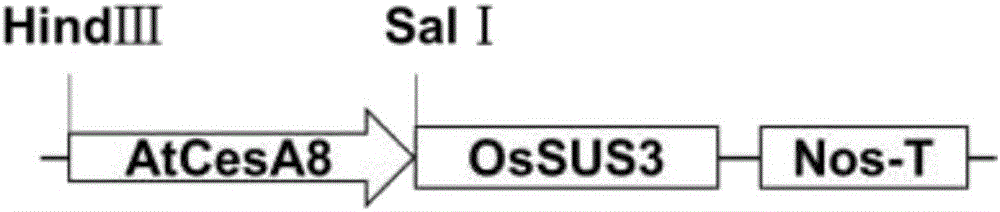
图2是植物表达载体的构建示意图。
转OsSUS3基因水稻种子Sus3(Oryza sativa Sus3),于2017年1月3日保藏在中国典型培养物保藏中心(地址:中国武汉武汉大学邮编:430072),保藏编号为CCTCC NO:P201702。
具体实施方式:
下面结合附图和实施例详细说明:
一、实施例1:AtCESA8启动子和OsSUS3基因cDNA的克隆
从拟南芥中克隆AtCESA8启动子全长,长度为949bp。从水稻中克隆OsSUS3基因cDNA,长度为2643bp,其中CDS为2451bp。PCR产物经过回收,酶切,连接载体。
——以拟南芥的基因组DNA为模板,合成引物扩增AtCESA8启动子。
F1:CCCAAGCTTCAGAGGAAACTCAGATGTGATGA;
R1:ACGCGTCGACCTTCGAATTCCCCTGTTTGGAGA;
——合成引物扩增OsSUS3cDNA
F2:ACGCGTCGACTTTCCTCCTCTTCTCCTTT;
R2:ACTGGCCCTGAAATCAAC
二、实施例2:表达载体的构建
1、拟南芥AtCESA8启动子超表达水稻的OsSUS3载体(pC1300T-AtCESA8-OsSUS3)的构建(见图2):
——将启动子和OsSUS3cDNA序列分别连接T载体后,酶切,转入表达载体pC1300T。
三、实施例3:植物表达载体转化农杆菌
1、农杆菌活化
将保存的农杆菌(EHA105)在固体LB培养基上画线(加抗生素:Kan,如果不加抗生素就有可能造成这些菌株的Ti质粒丢失,导致农杆菌缺乏侵染性),抗生素浓度为:50μg/mL,28℃培养1-2天,然后转移到新的加有抗生素的固体LB培养基上再培养2天;
2、农杆菌感受态细胞的制备
将100μL接种于1mLLB液体培养基中,150rpm 28℃振荡培养过夜;
吸取1mL菌液接种到100mL液体培养基中培养至OD600=0.5;
将菌液置冰上30min,使培养物冷却到0℃;
4℃,5000rpm离心30s,弃去上清液;
沉淀用60mL 0.1M CaCl2悬浮,冰浴30min;
5000rpm离心30s,弃去上清液;
每管用100μL含15%甘油的20mMCaCl2悬浮,用于转化;
制备好的感受态细胞可马上使用,也可按每管200μL分装于无菌离心管中,于4℃保存48小时内使用,长期贮存时必须在液氮中速冻后转-80℃保存,使用时从-80℃取出,置冰上融化后使用;
3、DNA直接转化农杆菌
200μL农杆菌感受态细胞中加入质粒DNA 0.1~1μg(5-10μL),之后冰浴30min;
放入液氮中7-8min,然后立即放入37℃水浴锅中水浴5min;
取出离心管,冰上放置2min,加入0.8mLLB,28℃,150rpm振荡培养3~4hr;
取出菌液于Kan抗生素的LB平板上涂板,在培养箱中28℃条件下倒置培养,2天左右菌落可见;
4、重组农杆菌鉴定
挑取单菌落,接种于含相应抗生素的LB液体培养基中,28℃振荡培养过夜;
小量提取质粒DNA,加GTE同时加5μL溶菌酶(50μg/mL-1,贮藏浓度为50mg/mL或10mg/mL);
质粒酶切或PCR鉴定。
四、实施例4:水稻成熟胚愈伤组织的诱导、分化、生根及移栽方法
1、愈伤诱导
将成熟的水稻种子去壳,先用无菌水冲洗3遍,然后用70%的乙醇处理1分钟,不时摇动;
无菌水冲洗3遍,每次30秒,不时摇动;
0.1%升汞溶液消毒12分钟(或2.5%NaClO消毒30min),不时摇动;
无菌水冲洗5遍以上,每次30秒,不时摇动;
将种子放在灭菌滤纸上吸干水分,然后放在诱导培养基上;
28℃,暗培养4周;
2、愈伤继代
挑选亮黄色、紧实且相对干燥的胚性愈伤(即散落到培养基上的愈伤,不要选从种子上发出来的愈伤),放于继代培养基上黑暗下培养2周,温度28℃;
3、预培养
挑选紧实且相对干燥的胚性愈伤,放于预培养基上黑暗下培养4-14天,温度28℃;
4、愈伤与农杆菌共培养
1)农杆菌培养
在带有对应抗性选择的LB培养基上预培养农杆菌两天,温度28℃;
将农杆菌转移至装有悬浮培养基的带塞试管或离心管里,重悬农杆菌,调节农杆菌的悬浮液至OD600为0.8-1.0;
2)农杆菌侵染
将预培养的愈伤转移至灭菌好的三角瓶内;
将愈伤在农杆菌悬浮液中浸泡30分钟;
转移愈伤至灭菌好的滤纸上吹干(30min-1h);
然后放置在已铺有一层滤纸的共培养基上培养3天,温度19-20℃。
5)选择培养
将共培养的愈伤转移至灭菌好的三角瓶内;
灭菌水洗涤愈伤至看不见农杆菌;
浸泡在含400ppm羧苄青霉素的灭菌水中30分钟;
转移愈伤至灭菌好的滤纸上吸干(1h以上);
转移愈伤至选择培养基上选择培养2次,每次2周。(第一次羧苄青霉素筛选浓度为400ppm,第二次为250ppm);
6)分化培养
将抗性愈伤转移至预分化培养基上黑暗处培养5-7天(可省略);
转移预分化培养的愈伤至分化培养基上,光照下培养,温度26℃;
7)生根培养
剪掉分化时产生的根(留1-2mm);
然后将其转移至生根培养基中光照下培养2周,温度26℃;
8)移栽
待苗高10-12cm,根系发生较好,打开瓶盖,洗掉根上的残留培养基(注意不要伤根),移栽花盆中。在最初的几天,蒙上保鲜膜,保持水分湿润。
五、实施例5:转基因阳性苗的鉴定
1、基因插入鉴定
检测引物:潮霉素通用引物和基因特异性引物;
2、表达鉴定
基因特异引物。
实施例6:水稻秸杆抗倒伏的测定
抗倒伏指数测定:用开花后30天水稻材料进行倒四节倒伏指数测定。倒四节倒伏指数计算公式如下:倒伏指数=(鲜重×株高)/折断力。鲜重:地上部分倒四节基部以上所有重量(包括茎秆、叶片、叶鞘、穗子);株高:倒四节基部到穗顶端长度;折断力:将待测样品倒四节水平放置在宽带度为5cm的装置中,采用折断力仪(DIK 7401;Daiki,Osaka,Japan)对倒四节进行折断力测定,结果见图1。
序列表
<110>华中农业大学
<120>利用蔗糖合酶提高水稻茎秆抗倒伏能力的方法
<140>
<141>
<160>5
<210>1
<211>2643
<212>OsSUS3基因的cDNA(包含CDS2451bp)
<400>
TTTCCTCCTCTTCTCCTTTAGTGCAAGGCTTGAGGATCCATCTAGAAGATAGCAATGGGGGAAACTACTGGAGAACGTGCCCTGACCCGTCTCCACAGCATGAGGGAGCGCATCGGCGATTCCCTCTCCGCGCACACCAATGAGCTTGTGGCTGTCTTCTCAAGGCTTGTGAACCAAGGAAAGGGAATGCTACAGCCCCACCAGATCATTGCTGAGTACAACGCCGCAATCCCTGAGGGCGAGCGTGAGAAGCTGAAGGACTCTGCCTTAGAGGATGTCCTGAGGGGAGCACAGGAGGCGATTGTCATCCCTCCATGGATTGCCCTTGCCATTCGCCCAAGGCCTGGTGTCTGGGAGTATCTGAGGATCAATGTAAGCCAGCTTGGTGTTGAGGAGCTGAGTGTCCCTGAATACTTGCAGTTCAAGGAGCAGCTTGTGGATGGAAGCACCCAGAACAACTTTGTGCTTGAGCTGGACTTTGAGCCATTCAATGCCTCCTTCCCTCGCCCATCGTTGTCGAAGTCTATTGGCAATGGGGTGCAGTTCTTGAACAGGCACCTGTCGTCAAAGCTGTTCCATGACAAAGAGAGCATGTACCCCCTGCTCAACTTTCTTCGTGCGCACAACTACAAAGGGATGACCATGATGTTGAACGACAGGATTCGCAGTCTCGATGCTCTCCAAGGTGCATTGAGGAAGGCAGAAAAACATCTTGCAGGCATTACAGCTGACACCCCATATTCAGAGTTCCATCACAGGTTCCAAGAGCTTGGTTTGGAGAAGGGTTGGGGTGACTGCGCTCAGCGAGTGCGTGAGACTATTCACCTTCTCTTGGACCTTCTTGAGGCCCCTGAGCCGTCCGCCTTGGAGAAGTTCCTTGGAACAATCCCAATGGTGTTCAATGTTGTTATCCTCTCCCCGCATGGTTACTTTGCACAGGCTAATGTCTTGGGGTACCCTGATACCGGTGGGCAGGTTGTCTACATTTTGGATCAAGTCCGTGCTATGGAGAATGAGATGCTGCTGAGGATCAAGCAACAAGGTCTAAACATCACACCAAGGATTCTCATTGTGACCAGGTTGCTACCTGATGCGCATGGCACCACATGTGGCCAGCGCCTTGAGAAGGTCCTAGGCACTGAGCACACTCATATCCTGCGTGTGCCATTCCGAACAGAAAATGGGACTGTTCGCAAATGGATCTCGCGTTTTGAAGTCTGGCCTTACCTGGAAACTTACACCGATGATGTGGCACACGAGATTTCTGGAGAGCTGCAGGCCACCCCTGACCTGATCATTGGGAACTACAGTGATGGCAACCTTGTTGCATGTTTGCTGGCACACAAGTTGGGTGTCACTCATTGTACAATCGCCCATGCACTTGAGAAAACCAAGTACCCCAACTCCGACCTTTACTGGAAGAAGTTTGAGGATCACTATCACTTCTCCTGCCAGTTCACAGCTGACCTGATTGCAATGAACCATGCTGACTTCATCATCACAAGTACCTTCCAGGAGATTGCTGGAAACAAGGAAACTGTGGGGCAGTATGAGTCTCACATGGCATTCACAATGCCTGGCCTTTATCGTGTTGTCCATGGTATCGATGTCTTTGACCCCAAGTTCAACATCGTCTCTCCTGGTGCTGACATGTCCATCTACTTCCCATTCACCGAATCACAGAAGAGGCTCACCTCTCTCCATTTAGAGATAGAGGAGCTACTCTTCAGTGATGTTGAAAACACTGAGCACAAGTTTGTTCTGAAGGACAAGAAGAAGCCAATCATCTTCTCGATGGCTAGGCTAGACCATGTCAAGAATTTGACTGGTCTGGTTGAGTTGTATGGTCGGAACCCTCGCCTGCAAGAGCTAGTAAACCTTGTGGTTGTCTGTGGTGACCATGGCAAGGAATCCAAGGACAAAGAAGAGCAGGCTGAGTTCAAGAAGATGTTTAATCTGATCGAGCAGTACAATTTGAATGGCCACATCCGCTGGATCTCCGCTCAGATGAACCGTGTCCGCAATGGTGAGCTCTACCGCTACATCTGCGACATGAGGGGAGCCTTTGTGCAGCCCGCTCTCTATGAGGCCTTTGGGCTAACTGTGATTGAGGCCATGACCTGTGGTCTTCCAACATTTGCAACTGCCTATGGTGGTCCAGCCGAGATCATCGTGCACGGCGTGTCTGGCTACCACATTGATCCTTACCAGAACGACAAGGCCTCGGCGCTGCTCGTGGAGTTCTTTGAGAAGTGTCAGGAAGACCCAAACCACTGGATCAAGATCTCGCAGGGTGGACTTCAGCGCATCGAGGAGAAGTACACATGGAAGCTCTACTCTGAGAGGCTGATGACTCTCTCCGGTGTCTACGGTTTCTGGAAGTATGTCACCAACCTCGACAGGCGTGAGACACGCCGCTACCTGGAGATGCTGTACGCCCTCAAGTACCGCAAGATGGCTACCACCGTTCCATTGGCCATTGAGGGAGAGGCCTCCACCAAATGATCTGGCCTTACCCGGTGAAAAGAATGGGCAATGGGTGCTCCATTGTTGCAGTGCTGATCCAGGGGTGAAGAAAAACAGAAATCGAGGAACGAATGCATCCATTTAGTTTCTAAGGGTTTAGTTGATTTCAGGGCCAGT;
<210>2
<211>AtCESA8启动子的正向引物
<212>32
<400>CCCAAGCTTCAGAGGAAACTCAGATGTGATGA;
<210>3
<211>AtCESA8启动子的反向引物
<212>33
<400>ACGCGTCGACCTTCGAATTCCCCTGTTTGGAGA;
<210>4
<211>OsSUS3cDNA的正向引物
<212>29
<400>ACGCGTCGACTTTCCTCCTCTTCTCCTTT;
<210>5
<211>OsSUS3cDNA的正向引物
<212>18
<400>ACTGGCCCTGAAATCAAC。
序列表
<110>华中农业大学
<120>利用蔗糖合酶提高水稻茎秆抗倒伏能力的方法
<140>
<141>
<160>5
<210>1
<211>2643
<212>OsSUS3基因的cDNA(包含CDS2451bp)
<400>
TTTCCTCCTCTTCTCCTTTAGTGCAAGGCTTGAGGATCCATCTAGAAGATAGCAATGGGGGAAACTACTGGAGAACGTGCCCTGACCCGTCTCCACAGCATGAGGGAGCGCATCGGCGATTCCCTCTCCGCGCACACCAATGAGCTTGTGGCTGTCTTCTCAAGGCTTGTGAACCAAGGAAAGGGAATGCTACAGCCCCACCAGATCATTGCTGAGTACAACGCCGCAATCCCTGAGGGCGAGCGTGAGAAGCTGAAGGACTCTGCCTTAGAGGATGTCCTGAGGGGAGCACAGGAGGCGATTGTCATCCCTCCATGGATTGCCCTTGCCATTCGCCCAAGGCCTGGTGTCTGGGAGTATCTGAGGATCAATGTAAGCCAGCTTGGTGTTGAGGAGCTGAGTGTCCCTGAATACTTGCAGTTCAAGGAGCAGCTTGTGGATGGAAGCACCCAGAACAACTTTGTGCTTGAGCTGGACTTTGAGCCATTCAATGCCTCCTTCCCTCGCCCATCGTTGTCGAAGTCTATTGGCAATGGGGTGCAGTTCTTGAACAGGCACCTGTCGTCAAAGCTGTTCCATGACAAAGAGAGCATGTACCCCCTGCTCAACTTTCTTCGTGCGCACAACTACAAAGGGATGACCATGATGTTGAACGACAGGATTCGCAGTCTCGATGCTCTCCAAGGTGCATTGAGGAAGGCAGAAAAACATCTTGCAGGCATTACAGCTGACACCCCATATTCAGAGTTCCATCACAGGTTCCAAGAGCTTGGTTTGGAGAAGGGTTGGGGTGACTGCGCTCAGCGAGTGCGTGAGACTATTCACCTTCTCTTGGACCTTCTTGAGGCCCCTGAGCCGTCCGCCTTGGAGAAGTTCCTTGGAACAATCCCAATGGTGTTCAATGTTGTTATCCTCTCCCCGCATGGTTACTTTGCACAGGCTAATGTCTTGGGGTACCCTGATACCGGTGGGCAGGTTGTCTACATTTTGGATCAAGTCCGTGCTATGGAGAATGAGATGCTGCTGAGGATCAAGCAACAAGGTCTAAACATCACACCAAGGATTCTCATTGTGACCAGGTTGCTACCTGATGCGCATGGCACCACATGTGGCCAGCGCCTTGAGAAGGTCCTAGGCACTGAGCACACTCATATCCTGCGTGTGCCATTCCGAACAGAAAATGGGACTGTTCGCAAATGGATCTCGCGTTTTGAAGTCTGGCCTTACCTGGAAACTTACACCGATGATGTGGCACACGAGATTTCTGGAGAGCTGCAGGCCACCCCTGACCTGATCATTGGGAACTACAGTGATGGCAACCTTGTTGCATGTTTGCTGGCACACAAGTTGGGTGTCACTCATTGTACAATCGCCCATGCACTTGAGAAAACCAAGTACCCCAACTCCGACCTTTACTGGAAGAAGTTTGAGGATCACTATCACTTCTCCTGCCAGTTCACAGCTGACCTGATTGCAATGAACCATGCTGACTTCATCATCACAAGTACCTTCCAGGAGATTGCTGGAAACAAGGAAACTGTGGGGCAGTATGAGTCTCACATGGCATTCACAATGCCTGGCCTTTATCGTGTTGTCCATGGTATCGATGTCTTTGACCCCAAGTTCAACATCGTCTCTCCTGGTGCTGACATGTCCATCTACTTCCCATTCACCGAATCACAGAAGAGGCTCACCTCTCTCCATTTAGAGATAGAGGAGCTACTCTTCAGTGATGTTGAAAACACTGAGCACAAGTTTGTTCTGAAGGACAAGAAGAAGCCAATCATCTTCTCGATGGCTAGGCTAGACCATGTCAAGAATTTGACTGGTCTGGTTGAGTTGTATGGTCGGAACCCTCGCCTGCAAGAGCTAGTAAACCTTGTGGTTGTCTGTGGTGACCATGGCAAGGAATCCAAGGACAAAGAAGAGCAGGCTGAGTTCAAGAAGATGTTTAATCTGATCGAGCAGTACAATTTGAATGGCCACATCCGCTGGATCTCCGCTCAGATGAACCGTGTCCGCAATGGTGAGCTCTACCGCTACATCTGCGACATGAGGGGAGCCTTTGTGCAGCCCGCTCTCTATGAGGCCTTTGGGCTAACTGTGATTGAGGCCATGACCTGTGGTCTTCCAACATTTGCAACTGCCTATGGTGGTCCAGCCGAGATCATCGTGCACGGCGTGTCTGGCTACCACATTGATCCTTACCAGAACGACAAGGCCTCGGCGCTGCTCGTGGAGTTCTTTGAGAAGTGTCAGGAAGACCCAAACCACTGGATCAAGATCTCGCAGGGTGGACTTCAGCGCATCGAGGAGAAGTACACATGGAAGCTCTACTCTGAGAGGCTGATGACTCTCTCCGGTGTCTACGGTTTCTGGAAGTATGTCACCAACCTCGACAGGCGTGAGACACGCCGCTACCTGGAGATGCTGTACGCCCTCAAGTACCGCAAGATGGCTACCACCGTTCCATTGGCCATTGAGGGAGAGGCCTCCACCAAATGATCTGGCCTTACCCGGTGAAAAGAATGGGCAATGGGTGCTCCATTGTTGCAGTGCTGATCCAGGGGTGAAGAAAAACAGAAATCGAGGAACGAATGCATCCATTTAGTTTCTAAGGGTTTAGTTGATTTCAGGGCCAGT;
<210>2
<211>AtCESA8启动子的正向引物
<212>32
<400> CCCAAGCTTCAGAGGAAACTCAGATGTGATGA;
<210>3
<211>AtCESA8启动子的反向引物
<212>33
<400>ACGCGTCGACCTTCGAATTCCCCTGTTTGGAGA;
<210>4
<211>OsSUS3cDNA的正向引物
<212>29
<400>ACGCGTCGACTTTCCTCCTCTTCTCCTTT;
<210>5
<211>OsSUS3cDNA的正向引物
<212>18
<400>ACTGGCCCTGAAATCAAC。
- 还没有人留言评论。精彩留言会获得点赞!