一种具有高转糖苷活性的β‑半乳糖苷酶组合突变体及其制备方法和应用与流程
本申请是申请号为201410514519.1,申请日为2014年9月29日,名称为“一种具有高转糖苷活性的β-半乳糖苷酶组合突变体及其制备方法和应用”的发明专利申请的分案申请。
本发明涉及基因工程和遗传工程领域,具体涉及一种具有高转糖苷活性的β-半乳糖苷酶组合突变体及其制备方法和应用。
背景技术:
低聚半乳糖(gos)是一类在人体胃肠道内不能被消化吸收而直接进入大肠被各种双歧杆菌良好利用的具有特殊生物学功能的低聚糖。它能够改善人体内微生态环境,有利于双歧杆菌和其它有益菌的增殖,提高人体免疫功能。同时,gos经代谢产生有机酸使肠内ph值降低,抑制肠内沙门氏菌和腐败菌的生长,减少有毒发酵产物及有害细菌酶的产生,调节胃肠功能,减轻了肝脏分解毒素的负担。由于低聚半乳糖有着比其他功能性低聚糖更优良的性质,从而使低聚半乳糖作为添加剂大规模应用更方便易行,能够适应更多的食品种类和更广的消费群体,有巨大的应用价值和市场潜力。
gos制备方法通常有五种:从天然原料中提取、天然多糖的酸水解、化学法合成、发酵法及酶法合成。由于gos在自然界中含量低,无色不带电荷故难以提取分离;天然多糖转化产品得率低,产物成分复杂,难以纯化;化学法毒性大易残留,对环境污染重;发酵法生产gos的研究很少,仍处于实验室规模,无法进行大量生产。目前低聚半乳糖的工业化生产是通过β-半乳糖苷酶完成的。β-半乳糖苷酶(β-d-galactosidegalatohydrolase,ec3.2.1.23)又称乳糖酶(lactase),具有水解和转糖苷的双重作用。早先对于β-半乳糖苷酶的研究主要集中在利用其水解功能生产低乳糖乳制品,来解除乳糖不耐受患者因食用乳制品而引起的腹泻、腹胀等多种不良反应。随着低聚半乳糖特殊的保健功能的确定,利用β-半乳糖苷酶的转糖苷作用生产低聚半乳糖成为研究热点。主要集中在以下三个方面:
1、筛选高转糖苷活性的β-半乳糖苷酶产生菌
酵母、芽孢杆菌、曲霉、青霉、双歧杆菌等多种微生物中都有具转糖苷酶活性的β-半乳糖苷酶。研究表明,不同来源的β-半乳糖苷酶因其酶学性质各异,合成低聚半乳糖时的反应条件大不相同。β-半乳糖苷酶根据最适ph可分为酸性和中性两种。通常霉菌来源的β-半乳糖苷酶为酸性酶,其作用最适ph在ph2.5~5.5,最适反应温度较高(50~60℃);酵母和细菌所产的β-半乳糖苷酶为中性酶,其最适ph介于ph6~7.5,最适反应温度较低(30~40℃)。不同来源的β-半乳糖苷酶作用的底物类型也不尽相同,由此生成的低聚半乳糖中寡糖的种类和比例也千差万别,这就使低聚半乳糖家族的新成员层出不穷。尽管如此,筛选到的天然β-半乳糖苷酶的转糖苷活性普遍较低,低聚半乳糖的最高产率通常只有5~30%,无法满足工业化生产的要求。
2、优化反应条件、改进生产工艺
有学者试图通过优化低聚半乳糖的生产条件和工艺来弥补其转糖苷活性低的缺陷,提高低聚半乳糖的得率,已取得了一定的效果。主要的方法有:增加起始乳糖的浓度、用有机溶剂控制水活度以及采用固定化技术。由于β-半乳糖苷酶的水解和转糖苷反应是可逆的。当底物浓度较低时,水解产物半乳糖浓度较低,其对水解反应的抑制作用较小,此时酶表现出水解活力较高,而转糖苷活力较低,因此产物中单糖含量较高。当乳糖浓度较高时,水解产物半乳糖浓度较高,当其达到一定值就对水解酶活产生抑制作用。半乳糖是转糖苷的底物,浓度高有利于半乳糖寡糖的合成,因此底物浓度较高时,产物中寡糖含量较高。使用有机溶剂有利于低聚糖的合成是因为有机溶剂能降低反应体系中水的活度以影响酶的活性位点和反应机制,引导水解酶催化逆向的转半乳糖苷反应,使反应平衡由水解偏移向低聚糖合成。利用固定化技术则可大大增加游离酶的ph和热稳定性并可反复利用,降低生产成本。mozaffar将β-半乳糖苷酶吸附于酚醛树脂并用戊二醛交联后,低聚糖的产率提高了20%;但也有研究发现,固定化酶作用于较高浓度的乳糖溶液时,产生的低聚糖比用游离酶还少。由此可见单纯依靠优化条件始终无法从根本上解决问题。
3、利用基因工程手段提高β-半乳糖苷酶的表达和性质改良
自然界中野生的β-半乳糖苷酶gos产量一般都维持在20~45%之间,产量较低,难以满足生产需求,因此通过分子改造筛选转糖苷性质优良的突变酶成为研究热点。hanseno.(2001)等发现来源于双歧杆菌中的β-半乳糖苷酶bif3,缺失掉c末端580个氨基酸后将酶蛋白变为一个高效的转糖苷酶,能够利用几乎90%乳糖生成gos,而水解成分占10%,而且在10%至40%的乳糖浓度下始终能够保持9:1比率的转糖苷活力和水解活力;placierg.于2009年对来源于嗜热脂肪土芽孢杆菌kve39的β-半乳糖苷酶进行定向进化,将转糖苷活力提高则水解活力降低作为筛选依据,成功筛选到三株突变体r109w,r109v,r109k,在18%(w/v)乳糖中低聚糖产量分别为23%,11.5%,21%,而野生酶只有2%;wuy.(2013)等对硫化叶菌来源的β-半乳糖苷酶进行分子改造,研究其最适gos生成条件,在各自最适条件下突变体f441ygos产量为61.7%,f359q为58.3%,而野生酶为50.9%。
继而,有学者提出一种迭代组合突变的方法(jij,fank.etal,2012),即将每一轮筛选结果里最高活力的突变体作为模板进行下一轮组合突变,直至生成的突变体活力均无法超越出发模板时,迭代组合结束。通过该方法从四轮组合突变共24株突变体中筛选到11株活力提高的突变体,最终活力最高突变体(c155yy184hv275ic281y)比原始野生酶提高1874%,但该组合突变的方法尚未有报道应用于β-半乳糖苷酶的分子改造中。
到目前为止,无论是天然酶的筛选分离、工艺优化还是通过基因工程手段提高β-半乳糖苷酶的表达和性质改良,都没有改变β-半乳糖苷酶转糖苷活性低和产量低的现状,从而也就造成低聚半乳糖合成产率低,反应时间长,生产成本太高,这严重制约了低聚半乳糖的廉价生产和推广应用。
因此,创制高转糖苷活性的新型β-半乳糖苷酶,并使其廉价生产是目前研究和生产中亟需解决的主要问题之一。
技术实现要素:
针对现有技术中存在的上述缺陷,本发明一方面提供了一种具有高转糖苷活性的β-半乳糖苷酶组合突变体,其是在亮白曲霉或米曲霉β-半乳糖苷酶的基础上,通过双位点的定点饱和突变而获得的,优选是在序列2或序列4所示的氨基酸序列的基础上,通过双位点的定点饱和突变所获得的,所述突变体在低聚糖生成量保持不变的情况下,转糖苷反应速率比野生型提高100%以上,优选提高150%以上,更加优选提高200%以上。
在本发明优选的实施方案中,突变位点分别为第219位和第785位的氨基酸。
在本发明进一步优选的实施方案中,双位点的定点饱和突变分别为用甘氨酸残基取代第219位的丝氨酸残基(s219g)和用缬氨酸残基取代第785位的谷氨酸残基(e785v)、用丙氨酸残基取代第219位的丝氨酸残基(s219a)和用缬氨酸残基取代第785位的谷氨酸残基(e785v)、用天冬酰胺残基取代第219位的丝氨酸残基(s219n)和用缬氨酸残基取代第785位的谷氨酸残基(e785v)、或用缬氨酸残基取代第219位的丝氨酸残基(s219v)和用缬氨酸残基取代第785位的谷氨酸残基(e785v)。
本发明的另一方面提供了编码上述组合突变体的dna分子。
本发明的再一方面提供了含有上述dna分子的重组表达载体,优选为重组酵母表达载体。
本发明的再一方面提供了表达上述dna分子的宿主细胞,优选糖酵母属、克鲁维氏酵母属、裂殖糖酵母属和甲基营养型酵母菌株,甲基营养型酵母菌株更加优选为毕赤氏酵母属菌株。
本发明的再一方面提供了一种制备具有高转糖苷活性的β-半乳糖苷酶的方法,包括以下步骤:
1、用上述的重组表达载体转化宿主细胞,获得重组菌株;
2、培养上述重组菌株,诱导重组β-半乳糖苷酶的表达;
3、回收并纯化所表达的高转糖苷活性的β-半乳糖苷酶。
本发明的最后一方面提供了本发明所述的组合突变体、dna分子、重组表达载体以及宿主细胞在制备β-半乳糖苷酶中的应用。
本发明采用双位点的定点饱和突变技术对去除自身信号肽的亮白曲霉的β-半乳糖苷酶基因lacbˊ和去除自身信号肽的米曲霉的β-半乳糖苷酶基因lacoˊ进行了定点饱和突变,获得了具有高转糖苷活性的β-半乳糖苷酶组合突变体,从而使其转糖苷反应速率比野生型提高了100%以上,甚至提高了200%以上,使制备高效率且高转糖苷活性的β-半乳糖苷酶成为现实,为β-半乳糖苷酶的应用奠定了良好的基础。
附图说明
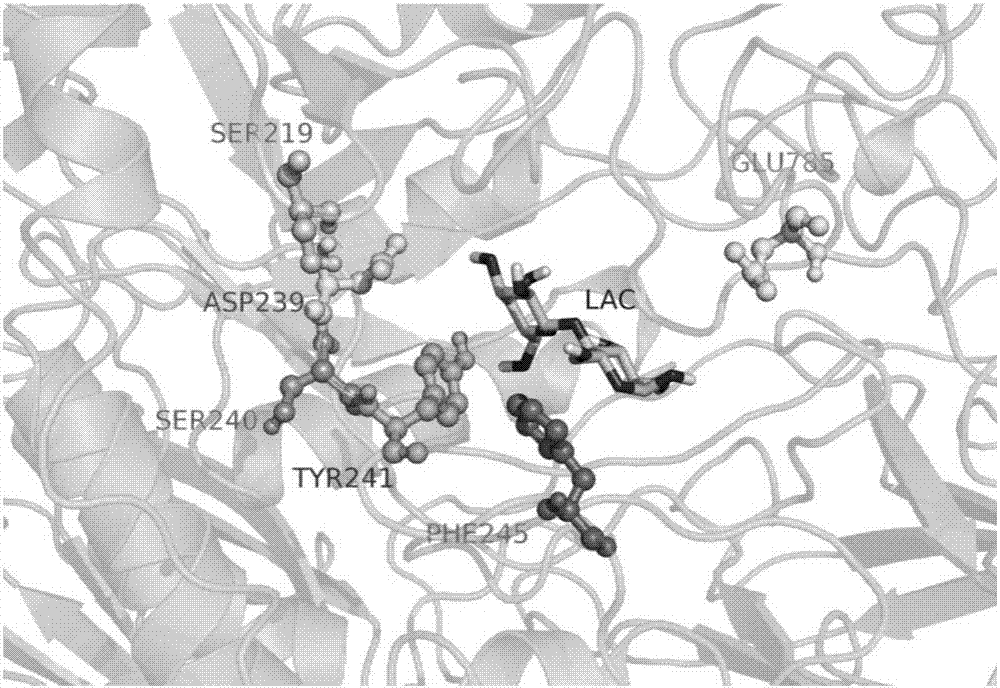
图1:亮白曲霉和米曲霉的β-半乳糖苷酶与底物的分子对接过程。
图2:亮白曲霉和米曲霉的β-半乳糖苷酶中突变位点与乳糖分子空间位置关系。
图3:含突变体β-半乳糖苷酶基因的重组表达载体的构建过程。
图4:s219位点典型突变体低聚半乳糖生成量。
图5:f245位点突变体库低聚半乳糖生成量趋势。
图6:f245位点典型突变体低聚半乳糖生成量。
图7:e785位点突变体库低聚半乳糖生成量趋势。
图8:s219/e785双位点突变体低聚半乳糖生成量。
具体实施方式
下面通过实施例对本发明作进一步的详细说明,旨在用于说明本发明而非限定本发明。应当指出,对于本领域技术人员而言,在不脱离本发明原理的前提下,还可以对本发明进行若干改进和修饰,这些改进和修饰也同样落入本发明的保护范围之内。
实施例1:β-半乳糖苷酶三级结构及突变位点的预测
去除自身信号肽的β-半乳糖苷酶基因lacbˊ,为本实验室从亮白曲霉(aspergilluscandidus)中克隆得到的,去除自身信号肽序列的基因由2958个核苷酸组成,具体序列如序列1所示,该基因编码的蛋白质由986个氨基酸组成,具体序列如序列2所示。
去除自身信号肽的β-半乳糖苷酶基因lacoˊ,为本实验室从米曲霉(aspergillusoryze)中克隆得到的,其也由2958个核苷酸组成,具体序列如序列3所示,该基因编码的蛋白质也由986个氨基酸组成,具体序列如序列4所示。其氨基酸序列与lacbˊ基因编码的蛋白质仅有三个氨基酸的差异:分别在第231位:lacbˊ(gly),lacoˊ(ser);第401位:lacbˊ(met),lacoˊ(ile);第970位:lacbˊ(asp),lacoˊ(asn)。
以亮白曲霉和米曲霉的β-半乳糖苷酶为研究材料。以获得晶体结构的青霉属β-半乳糖苷酶(pdb登录号:1tg7),米曲霉β-半乳糖苷酶(pdb登录号:4iug)及里氏木霉β-半乳糖苷酶(pdb登录号:3og2)的蛋白质晶体结构为同源模型,对β-半乳糖苷酶的3d结构及与底物结合的区域进行预测。预测的结构与文献报道的预测结果高度相似(参见thecrystalstructureofacidicβ-galactosidasefromaspergillusoryzae,mirkom.maksimainen,internationaljournalofbiologicalmacromolecules,2013,109-115)。酶蛋白由5个结构域组成:结构域1(1~394氨基酸)靠近n端,是该酶的活性中心所在,活性中心是tim桶型结构;结构域2(395~573氨基酸)由16个反向平行的β-折叠片和1个α-螺旋组成,包含了一个类似免疫球蛋白的亚结构域;结构域3(574~661氨基酸)由8个反向平行的β-折叠片组成一个“希腊钥匙”的β-夹层和1个α-螺旋组成;结构域4(662~857氨基酸)和结构域5(858~1005氨基酸)主要由“春卷”型的拓扑结构组成。对活性中心重点分析发现,位于tim桶活性中心第4和第7条β-折叠片上的glu160、glu258可能是参与催化反应必不可少的氨基酸,而asn140和tyr96可能用来固定乳糖分子。
根据得到的β-半乳糖苷酶的三维结构,通过discoverystudio软件进行酶与底物的分子对接模拟(参见附图1),根据对接结果解析,可获知与底物存在相互作用的氨基酸(参见附图2)。使用计算生物学软件逐个评估这些氨基酸的进化熵,最终筛选确定了六个进化熵变化较大的氨基酸位点s219,d239,s240,y241,f245和e785(参见表1)用于定点饱和突变。
表1β-半乳糖苷酶典型氨基酸的进化熵
实施例2:毕赤酵母单点饱和突变体库的构建
1、材料与方法
(1)菌种与载体
野生型基因来源于去除自身信号肽的亮白曲霉的β-半乳糖苷酶基因lacbˊ,以及来源于米曲霉的β-半乳糖苷酶基因lacoˊ,为本实验室前期克隆得到,具体序列如序列1和序列3所示,连于ppic9表达载体上,并在毕赤酵母gs115中表达;escherichiacolitrans1-t1感受态细胞购于transgen公司;ppic9表达载体,毕赤酵母gs115购于invitrogen公司。
(2)培养基及相关溶液的配制
毕赤酵母转化、培养及筛选常规培养基及试剂参照invitrogen公司的说明书。
ptm微量盐:0.6%cuso4,0.008%nai2,0.3%mnso4,0.02%na2moo4,0.002%h3bo3,0.05%cocl2,2%zncl2,6.5%feso4,0.5%硫酸(v/v)。
酵母发酵基础盐培养基(fbsm):0.5%kh2po4,5%nh4h2po4,1.485%mgso4,1.82%k2so4,0.093%caso4,0.15%koh,0.00011%biotin,0.44%ptm微量盐,2%葡萄糖。
酵母发酵基础盐诱导培养基(fbim):0.5%kh2po4,5%nh4h2po4,1.485%mgso4,1.82%k2so4,0.093%caso4,0.15%koh,0.00011%biotin,0.44%ptm微量盐,0.5%甲醇。
na2hpo4-柠檬酸缓冲液(0.1mol/lph5.2):0.2mol/l磷酸氢二钠536ml,0.1mol/l柠檬酸464ml,混匀后调ph至5.2。
(3)寡核苷酸引物
基因突变中所采用的具体引物序列如表2所示。
表2基因突变中所用引物表
2、重叠pcr扩增突变位点
采用重叠pcr(overlappcr)的方法对基因的单个位点进行饱和突变。即两个片段分别经pcr扩增,然后通过重叠延伸进行融合。设计一对兼并引物,使其在靶位点附近有一定程度的重叠(参见附图3中的b、c引物),分别与基因5′和3′端引物(参见附图3中的a、d引物)组合,扩增含突变靶位点的上游片段和下游片段,由于这些引物是互补的,因而产生的pcr产物链将相互重叠,将上下游片段在靶位点处交错,互为模板重叠延伸成全长基因。取1μlppic9-lacb′质粒为模板,分别用引物对a和c以及b和d,使用transstartfastpfudna聚合酶进行扩增将pcr产物用琼脂糖凝胶电泳进行检测,回收大小正确的片段(方法参照天根生化试剂(北京)公司,琼脂糖凝胶dna回收试剂盒)。
3、体外同源重组构建表达载体
将带有同源臂的两段pcr片段,等摩尔量混合,加入同源重组酶进行体外重组。将混合物在25℃反应30min,之后冰上放置5min。可立即转化或者-20℃保存。将同源重组产物分别取10ul利用化学法转入100ul大肠杆菌trans1-t1感受态细胞中,并涂布于含有amp的lb平板上,37℃倒置培养过夜。
lb平板上长出3~5倍于理论突变子数量(突变密码子为mnn,变体库的理论值为4×4×2=32,单点饱和突变挑选32个克隆子即可覆盖全部突变)就可以覆盖所有突变位点。随机从每个突变体库的lb平板上挑取6~8个单克隆进行dna序列测定,测序工作委托北京迈奥德恩生物科技有限公司完成。将每个突变体库按突变位点不同,依次命名为s219库、d239库、s240库、y241库、f245库和e785库。
4、β-半乳糖苷酶突变体在毕赤酵母中的表达与高转糖苷活性菌株的筛选方法
(1)重组质粒在毕赤酵母中的表达
从各个突变体库的大肠杆菌中提取混合质粒(约200-230μg),用足量bglii酶进行完全酶切消化后,异丙醇沉淀,70%乙醇洗涤后溶于去离子水中转化毕赤酵母。将转化酵母涂布到含有x-gal的mm平板上,在mm平板上变蓝的菌株为有β-半乳糖苷酶活性的阳性克隆,将其对应的md平板上的菌落挑到48孔培养板中,不同突变体在48孔板中先经fbsm培养基培养使其快速生长48h后,再经fbim培养基诱导培养48h后取上清测定阳性突变株的β-半乳糖苷酶活性。
(2)β-半乳糖苷酶以onpg底物的活性测定方法
准确称取onpg底物0.1g,溶于40mlna2hpo4-柠檬酸缓冲液(ph5.2,0.1mol/l),即为浓度为0.25%(w/v)的onpg溶液。待测粗酶液使用ph5.2,0.1mol/l的na2hpo4-柠檬酸稀释至合适倍数,取吸800μl的底物溶液加入试管中于60℃水浴锅中预热2min,加入200μl稀释后酶液混匀,反应15min后依次加入1ml10%tca终止,2ml1mol/lna2co3显色,测定420nm处的光吸收值(od420)。以加入na2hpo4-柠檬酸缓冲液(浓度为0.1mol/l,ph5.2)作为空白对照,利用标准曲线,计算反应生成的onp的量,进而计算出β-半乳糖苷酶的酶活力。酶活力单位定义:一个单位(1u)的β-半乳糖苷酶活性指在60℃,ph5.2条件下,每分钟催化底物邻硝基苯酚-β-d-吡喃半乳糖苷(onpg)生成1μmol邻硝基酚(onp)所需的酶量。
根据β-半乳糖苷酶标准曲线的结果,酶活力计算公式为:
酶活性(u/ml)=5*n*(0.9472x+0.0046)/15
x:420nm处光吸收值;n:酶液的稀释倍数;15:反应15min;5:将200μl稀释酶液中的酶活折算为1ml的酶活性。
(3)突变株转糖苷活力测定基本反应体系及反应条件
将各突变体的粗酶液经ph5.2,0.1mol/l的na2hpo4-柠檬酸缓冲液稀释至等蛋白浓度,即每60μl酶液中含有5μg蛋白(浓度约0.08mg/ml)。吸取60μl稀释后酶液置于ep管中,加入440μl30%(w/v)的乳糖底物,底物与酶液混合尽量快速,保证每个样品起始反应时间间隔最小。将各反应物置于50℃,200rpm恒温摇床中反应不同时间。反应结束后12000rpm离心10min,置于100℃水浴中煮沸10min终止反应。
反应产物使用超纯水稀释16倍后12000rpm离心10min,取700μl进行hplc检测。
高效液相色谱仪(hplc)检测条件:
hplc定量检测前绘制葡萄糖、半乳糖、乳糖的标准曲线。葡萄糖、半乳糖、乳糖的检测区间均为0–25.6mg/ml,检测条件为:waterse2695separationsmoule,流动相:50mmedta钙钠盐;柱温:85℃;流速,时间:0.5ml/min,12min/sample。
低聚糖gos产量(mg/ml)=起始乳糖量(mg/ml)-剩余乳糖量(mg/ml)-葡萄糖量(mg/ml)-半乳糖量(mg/ml)(jorgensenfetal,2001);
gos转化率=低聚糖量(mg/ml)/起始乳糖量(mg/ml);
消耗乳糖转化为gos的得率=低聚糖量(mg/ml)/[起始乳糖量(mg/ml)-剩余乳糖量(mg/ml)]。
实施例3:s219饱和突变体库的筛选及低聚糖的合成
选取s219突变体库的200个毕赤酵母阳性克隆测定其转糖苷活性(低聚糖生产量),并测定核苷酸序列。测序表明,这些突变体分别突变为8种不同的氨基酸均可提高突变酶的转糖苷活性(参见表3),尤其以带有较小侧链的氨基酸如gly、ala、val及带负电荷的极性氨基酸glu较突出,其中突变为gly最为明显(参见附图4),低聚糖生成量可提高26.6%。突变为小侧链且带负电的glu后低聚糖生成量提高25.7%,当s219突变为ala、val后低聚糖生成量分别提高15.0%和15.5%,突变为asp、arg、leu低聚糖生成量分别提高10.4%,7.9%和8.2%,突变为大侧链芳香族氨基酸phe其转糖苷也有大幅提升,提高16.4%。而突变为pro和trp其转糖苷能力明显下降,分别下降16.7%和28.7%。由此可见,s219突变为其他氨基酸后转糖苷能力呈现有高有低的态势,表明该位点是一个与β-半乳糖苷酶转糖苷能力相关的重要位点。该位点在空间上位于β-半乳糖苷酶活性中心-tim桶内,通过电荷和极性作用与乳糖底物产生一定作用且是该活性中心内的非保守氨基酸。
表3s219不同氨基酸突变体低聚糖生成情况
注:wt代表野生酶。
实施例4:f245饱和突变体库的筛选及低聚糖的合成
从f245库中筛选了200个阳性突变体进行转糖苷活性的测定。结合转糖苷活性测定结果和测序结果表明,所有突变体在低聚糖生成量上呈现有高有低的态势(参见附图5),部分突变体低聚糖生成量比野生酶下降很多,部分突变体低聚糖生成量比野生酶高出30%左右。由此可见,f245位点也是一个与β-半乳糖苷酶低聚糖生成相关的重要位点。
具体的,f245位点突变为arg后低聚糖生成量提高最多,约35%,其次是lys和gly,分别提高约30%和24.7%。突变为其他氨基酸如ser、glu、ala、thr和met等后,低聚糖产量也都有大幅提升(参见附图6)。
实施例5:e785饱和突变体库的筛选及低聚糖的合成
与野生型相比,e785饱和突变体库中,仅有少量突变体的低聚糖合成量提高,且提高幅度不大于20%(参见附图7)。e785突变体库低聚糖生成量提高的突变体中,glu突变为val后提高15%,其余大部分突变体低聚糖产量低于野生型或与野生酶差异不大。
实施例6组合突变体库的构建和筛选
1、材料与方法
(1)菌种与载体
起始突变体为亮白曲霉的β-半乳糖苷酶基因lacbˊ的突变体e785v,连接于ppic9表达载体上,并在毕赤酵母gs115中表达;escherichiacolitrans1-t1感受态细胞购于transgen公司;ppic9表达载体和毕赤酵母gs115购于invitrogen公司。
(2)培养基及相关溶液的配制
毕赤酵母转化、培养及筛选常规培养基及试剂参照invitrogen公司的说明书。
ptm微量盐:0.6%cuso4,0.008%nai2,0.3%mnso4,0.02%na2moo4,0.002%h3bo3,0.05%cocl2,2%zncl2,6.5%feso4,0.5%硫酸(v/v)。
酵母发酵基础盐培养基(fbsm):0.5%kh2po4,5%nh4h2po4,1.485%mgso4,1.82%k2so4,0.093%caso4,0.15%koh,0.00011%biotin,0.44%ptm微量盐,2%葡萄糖。
酵母发酵基础盐诱导培养基(fbim):0.5%kh2po4,5%nh4h2po4,1.485%mgso4,1.82%k2so4,0.093%caso4,0.15%koh,0.00011%biotin,0.44%ptm微量盐,0.5%甲醇。
na2hpo4-柠檬酸缓冲液(0.1mol/lph5.2):0.2mol/l磷酸氢二钠536ml,0.1mol/l柠檬酸464ml,混匀后调ph至5.2。
(3)突变用寡核苷酸引物
组合突变中所采用的具体引物序列如表4所示。
表4组合突变中所用引物表
注:加粗字体为突变碱基。
2、组合突变体库的构建
基于上述对不同位点饱和突变体库的分析,选择s219以及e785两个位点作为组合突变的位点,并根据对每个位点正突变体的测序结果确定每个位点的目的突变氨基酸。。确定突变位点以及目的突变氨基酸后设计引物,组合突变体库的构建采用分次突变的方法,即先构建其中一个位点的突变质粒,在此基础上再对另一位点进行突变,直至完成目的突变。对于s219/e785双点组合突变而言,采用先突变e785单点,再利用e785单点突变质粒进行s219位点的突变。
组合突变重组质粒的构建则建立在上述单点突变的基础上,即以单点突变的混合质粒为模板继续通过上述的pcr扩增和重组技术获得双点突变体。
将组合突变重组质粒转化到毕赤酵母gs115中,在毕赤酵母gs115中进行表达,并测定阳性突变株的β-半乳糖苷酶活性。
3、组合突变体库的筛选
从s219/e785双点突变体库中随机挑取500个突变体,测定其β-半乳糖苷酶活性。如图8所示,s219/e785双点突变体库中大部分突变体的低聚糖生成量都高于野生酶。更重要的是,部分组合突变体的gos生成速率较野生型大大加快,达到最高gos产量所需的时间也大大缩短,不同突变体转糖苷的性质分别列于表5中。
表5不同突变体的转糖苷性质
由表5的数据可知,筛选出的高转糖苷突变酶在相同乳糖底物中低聚糖的生成速率明显加快,也即意味着转糖苷反应速率加快,以s219g/e785v突变体最为明显。s219g/e785v在反应2h时低聚糖的产量就极高,大部分突变体在5–6h左右就达到了低聚糖合成最大量,即水解和转糖苷反应速度相平衡阶段,只有e785v突变体在36h左右才达到最大低聚糖生成量,而野生酶在48h左右才达到转糖苷和水解的平衡阶段,反应周期很长。大部分突变体在加快反应速率的同时增加了gos得率,最高的如s219v/e785v转化率由23.3%到27.1%提高了16%。以反应2h低聚糖的产量来计算每个突变体低聚糖的生成速率可见,除e785v突变体比野生酶提高了22%左右,其余突变体提高量都在100%以上,其中反应速率最快的s219g/e785v突变体提高量在211%左右,是野生酶的3倍多。
序列表
<110>中国农业科学院生物技术研究所
<120>一种具有高转糖苷活性的β-半乳糖苷酶组合突变体及其制备方法和应用
<160>24
<170>patentinversion3.3
<210>1
<211>2958
<212>dna
<213>人工序列
<400>1
tccatcaagcatcgtctcaatggcttcacgatcctggaacatccggatccggcgaaaaga60
gacttgctgcaagacattgttacatgggatgacaaatctctgttcatcaatggagagagg120
attatgttattcagcggagaagtgcatcctttcagattgccagtaccttcgctttggctt180
gatatcttccacaagatcagagctcttggtttcaactgtgtatctttctatattgattgg240
gctcttctggagggaaagcctggcgactacagagcagaaggcatctttgctctggaaccc300
ttcttcgatgcagccaaggaagcaggcatttatctgatcgcccgccccggttcgtacatc360
aatgccgaggtctcaggcggtggcttccctggatggttgcagagggtcaatggcactctt420
cgctcgtctgatgagccattccttaaagctactgataactatatcgccaatgccgctgct480
gccgtggcgaaggctcaaatcacgaatggagggccagtaattctctaccagcccgaaaac540
gaatacagcggtggctgctgcggtgtcaaataccccgatgcagactacatgcagtatgtt600
atggatcaggcccggaaggctgacattgttgtacctttcatcagcaacgatgcctcacct660
tctgggcacaatgctcctggaagtggaacgggcgctgttgatatttatggtcacgatagc720
tatccccttggctttgattgcgcaaacccatccgtatggcccgagggtaaactgcccgac780
aacttccgcacgctccatcttgagcagagcccatcaactccgtattcacttcttgagttc840
caagcgggtgctttcgacccatggggtggacccggctttgaaaaatgctatgccctcgtt900
aaccacgaattctcgagagttttctataggaacgacttgagtttcggagtttctaccttt960
aacttatacatgactttcggcggaacaaactggggtaacctcggacatcccggtggatat1020
acatcctacgactacggctcgcctataactgaaacgcgaaacgttacacgggagaagtac1080
agcgacataaagctccttgccaactttgtcaaagcatcgccatcctatctcaccgctact1140
cccagaaacctgactactggtgtttacacagacacatctgacctggctgtcaccccgtta1200
atgggtgatagtccaggctcattcttcgtggtcagacatacggactattccagccaagag1260
tcaacctcgtacaaacttaagcttcctaccagtgctggtaacctgactattccccagctg1320
gagggcactctaagtctcaacggacgtgactcaaaaattcatgttgttgattataatgtg1380
tctggaacgaacattatctattcgacagctgaagtcttcacctggaagaagtttgacggt1440
aacaaggtcctggtgttatacggcggaccgaaggaacaccatgaattggccattgcctcc1500
aagtcaaatgtgaccatcatcgaaggttcggactctggaattgtctcaacgaggaagggc1560
agctctgttatcattggctgggatgtctcttctactcgtcgcatcgttcaagtcggtgac1620
ttgagagtgttcctgcttgatagaaactctgcttacaactactgggtccccgaactcccc1680
acagaaggtacttctcccgggttcagcacttcgaagacgaccgcctcctccattattgtg1740
aaggccggctacctcctccgaggggctcacctggatggtgctgatcttcatcttactgct1800
gatttcaatgccaccaccccgattgaagtgatcggtgctccaacaggcgccaagaatctg1860
ttcgtgaatggtgaaaaggctagccacacagtcgacaaaaacggcatctggagtagtgag1920
gtcaagtacgcggctccagagatcaagctccccggtttgaaggatttggactggaagtat1980
ctggacacgcttcccgaaattaagtcttcctatgatgactcggcctgggtttcggcagac2040
cttccaaagacaaagaacactcaccgtcctcttgacacaccaacatcgctatactcctct2100
gactatggcttccacactggctacctgatctacaggggtcacttcgttgccaacggcaag2160
gaaagcgaattttttattcgcacacaaggcggtagcgcattcggaagttccgtatggctg2220
aacgagacgtatctgggctcttggactggtgccgattatgcgatggacggtaactctacc2280
tacaagctatctcagctggagtcgggcaagaattacgtcatcactgtggttattgataac2340
ctgggtctcgacgagaattggacggtcggcgaggaaaccatgaagaatcctcgtggtatt2400
cttagctacaagctgagcggacaagacgccagcgcaatcacctggaagctcactggtaac2460
ctcggaggagaagactaccaggataaggttagaggacctctcaacgaaggtggactgtac2520
gcagagcgccagggcttccatcagcctcagcctccaagcgaatcctgggagtcgggcagt2580
ccccttgaaggcctgtcgaagccgggtatcggattctacactgcccagttcgaccttgac2640
ctcccgaagggctgggatgtgccgctgtacttcaactttggcaacaacacccaggcggct2700
cgggcccagctctacgtcaacggttaccagtatggcaagttcactggaaacgttgggcca2760
cagaccagcttccctgttcccgaagggatcctgaactaccgcggaaccaactatgtggca2820
ctgagtctttgggcattggagtcggacggtgctaagctgggtagcttcgaactgtcctac2880
accaccccagtgctgaccggatacggggatgttgagtcacctgagcagcccaagtatgag2940
cagcggaagggagcatac2958
<210>2
<211>986
<212>prt
<213>人工序列
<400>2
serilelyshisargleuasnglyphethrileleugluhisproasp
151015
proalalysargaspleuleuglnaspilevalthrtrpaspasplys
202530
serleupheileasnglygluargilemetleupheserglygluval
354045
hispropheargleuprovalproserleutrpleuaspilephehis
505560
lysileargalaleuglypheasncysvalserphetyrileasptrp
65707580
alaleuleugluglylysproglyasptyrargalagluglyilephe
859095
alaleugluprophepheaspalaalalysglualaglyiletyrleu
100105110
ilealaargproglysertyrileasnalagluvalserglyglygly
115120125
pheproglytrpleuglnargvalasnglythrleuargserserasp
130135140
glupropheleulysalathraspasntyrilealaasnalaalaala
145150155160
alavalalalysalaglnilethrasnglyglyprovalileleutyr
165170175
glnprogluasnglutyrserglyglycyscysglyvallystyrpro
180185190
aspalaasptyrmetglntyrvalmetaspglnalaarglysalaasp
195200205
ilevalvalpropheileserasnaspalaserproserglyhisasn
210215220
alaproglyserglythrglyalavalaspiletyrglyhisaspser
225230235240
tyrproleuglypheaspcysalaasnproservaltrpproglugly
245250255
lysleuproaspasnpheargthrleuhisleugluglnserproser
260265270
thrprotyrserleuleuglupheglnalaglyalapheaspprotrp
275280285
glyglyproglypheglulyscystyralaleuvalasnhisgluphe
290295300
serargvalphetyrargasnaspleuserpheglyvalserthrphe
305310315320
asnleutyrmetthrpheglyglythrasntrpglyasnleuglyhis
325330335
proglyglytyrthrsertyrasptyrglyserproilethrgluthr
340345350
argasnvalthrargglulystyrseraspilelysleuleualaasn
355360365
phevallysalaserprosertyrleuthralathrproargasnleu
370375380
thrthrglyvaltyrthraspthrseraspleualavalthrproleu
385390395400
metglyaspserproglyserphephevalvalarghisthrasptyr
405410415
serserglngluserthrsertyrlysleulysleuprothrserala
420425430
glyasnleuthrileproglnleugluglythrleuserleuasngly
435440445
argaspserlysilehisvalvalasptyrasnvalserglythrasn
450455460
ileiletyrserthralagluvalphethrtrplyslyspheaspgly
465470475480
asnlysvalleuvalleutyrglyglyprolysgluhishisgluleu
485490495
alailealaserlysserasnvalthrileilegluglyseraspser
500505510
glyilevalserthrarglysglyserservalileileglytrpasp
515520525
valserserthrargargilevalglnvalglyaspleuargvalphe
530535540
leuleuaspargasnseralatyrasntyrtrpvalprogluleupro
545550555560
thrgluglythrserproglypheserthrserlysthrthralaser
565570575
serileilevallysalaglytyrleuleuargglyalahisleuasp
580585590
glyalaaspleuhisleuthralaasppheasnalathrthrproile
595600605
gluvalileglyalaprothrglyalalysasnleuphevalasngly
610615620
glulysalaserhisthrvalasplysasnglyiletrpserserglu
625630635640
vallystyralaalaprogluilelysleuproglyleulysaspleu
645650655
asptrplystyrleuaspthrleuprogluilelyssersertyrasp
660665670
aspseralatrpvalseralaaspleuprolysthrlysasnthrhis
675680685
argproleuaspthrprothrserleutyrserserasptyrglyphe
690695700
histhrglytyrleuiletyrargglyhisphevalalaasnglylys
705710715720
glusergluphepheileargthrglnglyglyseralapheglyser
725730735
servaltrpleuasngluthrtyrleuglysertrpthrglyalaasp
740745750
tyralametaspglyasnserthrtyrlysleuserglnleugluser
755760765
glylysasntyrvalilethrvalvalileaspasnleuglyleuasp
770775780
gluasntrpthrvalglyglugluthrmetlysasnproargglyile
785790795800
leusertyrlysleuserglyglnaspalaseralailethrtrplys
805810815
leuthrglyasnleuglyglygluasptyrglnasplysvalarggly
820825830
proleuasngluglyglyleutyralagluargglnglyphehisgln
835840845
proglnproproserglusertrpgluserglyserproleuglugly
850855860
leuserlysproglyileglyphetyrthralaglnpheaspleuasp
865870875880
leuprolysglytrpaspvalproleutyrpheasnpheglyasnasn
885890895
thrglnalaalaargalaglnleutyrvalasnglytyrglntyrgly
900905910
lysphethrglyasnvalglyproglnthrserpheprovalproglu
915920925
glyileleuasntyrargglythrasntyrvalalaleuserleutrp
930935940
alaleugluseraspglyalalysleuglyserphegluleusertyr
945950955960
thrthrprovalleuthrglytyrglyaspvalgluserproglugln
965970975
prolystyrgluglnarglysglyalatyr
980985
<210>3
<211>2958
<212>dna
<213>人工序列
<400>3
tccatcaagcatcgtctcaatggcttcacgatcctggaacatccggatccggcgaaaaga60
gacttgctgcaagacattgttacatgggatgacaaatctctgttcatcaatggagagagg120
attatgttattcagcggagaagtgcatcctttcagattgccagtaccttcgctttggctt180
gatatcttccacaagatcagagctcttggtttcaactgtgtatctttctatattgattgg240
gctcttctggagggaaagcctggcgactacagagcagaaggcatctttgctctggaaccc300
ttctttgatgcagccaaggaagcaggcatttatctgatcgcccgccccggttcgtacatc360
aatgccgaggtctcaggcggtggcttccctggatggttgcagagggtcaatggcactctt420
cgctcgtctgatgagccattccttaaagctactgataactatatcgccaatgccgctgct480
gccgtggcgaaggctcaaatcacgaatggagggccagtaattctctaccagcccgaaaac540
gaatacagcggtggctgctgcggtgtcaaataccccgatgcagactacatgcagtatgtt600
atggatcaggcccggaaggctgacattgttgtacctttcatcagcaacgatgcctcacct660
tctgggcacaatgctcctggaagtggaacgagcgctgttgatatttatggtcacgatagc720
tatcccctcggctttgattgcgcaaacccatccgtatggcccgagggtaaactgcccgac780
aacttccgcacgctccatcttgagcagagcccatcaactccgtattcacttcttgagttc840
caagcgggtgctttcgacccatggggtggacccggctttgaaaaatgctatgccctcgtt900
aaccacgaattctcgagagttttctataggaacgacttgagtttcggagtttctaccttt960
aacttatacatgactttcggcggaacaaactggggtaacctcggacatcccggtggatat1020
acatcctacgactacggatcgcctataactgaaacgcgaaacgttacgcgggagaagtac1080
agcgacataaagctccttgccaactttgtcaaagcatcgccatcctatctcaccgctact1140
cccagaaacctgactactggtgtttacacagacacatctgacctggctgtcaccccgtta1200
attggtgatagtccaggctcattcttcgtggtcagacatacggactattccagccaagag1260
tcaacctcgtacaaacttaagcttcctaccagtgctggtaacctgactattccccagctg1320
gagggcactctaagtctcaacggacgtgactcaaaaattcatgttgttgattataatgtg1380
tctggaacgaacattatctattcgacagctgaagtcttcacctggaagaagtttgacggt1440
aacaaggtcctggtgttatacggcggaccgaaggaacaccatgaattggccattgcctcc1500
aagtcaaatgtgaccatcatcgaaggttcggactctggaattgtctcaacgaggaagggc1560
agctctgttatcattggctgggatgtctcttctactcgtcgcatcgttcaagtcggtgac1620
ttgagagtgttcctgcttgataggaactctgcttacaactactgggtccccgaactcccc1680
acagaaggtacttctcccgggttcagcacttcgaagacgaccgcctcctccattattgtg1740
aaggctggctacctcctccgaggcgctcaccttgatggtgctgatcttcatcttactgct1800
gatttcaatgccaccaccccgattgaagtgatcggtgctccaacaggcgctaagaatctg1860
ttcgtgaatggtgaaaaggctagccacacagtcgacaagaacggcatctggagcagtgag1920
gtcaagtacgcggctccagagatcaagctccccggtttgaaggatttggactggaagtat1980
ctggacacgcttcccgaaattaagtcttcctatgatgactcggcctgggtttcggcagac2040
cttccaaagacaaagaacactcaccgtcctcttgacacaccaacatcgctatactcctct2100
gactatggcttccacactggctacctgatctacaggggtcacttcgttgccaacggcaag2160
gaaagcgaattttttattcgcacacaaggcggtagcgcattcggaagttccgtatggctg2220
aacgagacgtatctgggctcttggactggtgccgattatgcgatggacggtaactctacc2280
tacaagctatctcagctggagtcgggcaagaattacgtcatcactgtggttattgataac2340
ctgggtctcgacgagaattggacggtcggcgaggaaaccatgaagaatcctcgtggtatt2400
cttagctacaagctgagcggacaagacgccagcgcaatcacctggaagctcactggtaac2460
ctcggaggagaagactaccaggataaggttagaggacctctcaacgaaggtggactgtac2520
gcagagcgccagggcttccatcagcctcagcctccaagcgaatcctgggagtcgggcagt2580
ccccttgaaggcctgtcgaagccgggtatcggattctacactgcccagttcgaccttgac2640
ctcccgaagggctgggatgtgccgctgtacttcaactttggcaacaacacccaggcggct2700
cgggcccagctctacgtcaacggttaccagtatggcaagttcactggaaacgttgggcca2760
cagaccagcttccctgttcccgaaggtatcctgaactaccgcggaaccaactatgtggca2820
ctgagtctttgggcattggagtcggacggtgctaagctgggtagcttcgaactgtcctac2880
accaccccagtgctgaccggatacgggaatgttgagtcacctgagcagcccaagtatgag2940
cagcggaagggagcatac2958
<210>4
<211>986
<212>prt
<213>人工序列
<400>4
serilelyshisargleuasnglyphethrileleugluhisproasp
151015
proalalysargaspleuleuglnaspilevalthrtrpaspasplys
202530
serleupheileasnglygluargilemetleupheserglygluval
354045
hispropheargleuprovalproserleutrpleuaspilephehis
505560
lysileargalaleuglypheasncysvalserphetyrileasptrp
65707580
alaleuleugluglylysproglyasptyrargalagluglyilephe
859095
alaleugluprophepheaspalaalalysglualaglyiletyrleu
100105110
ilealaargproglysertyrileasnalagluvalserglyglygly
115120125
pheproglytrpleuglnargvalasnglythrleuargserserasp
130135140
glupropheleulysalathraspasntyrilealaasnalaalaala
145150155160
alavalalalysalaglnilethrasnglyglyprovalileleutyr
165170175
glnprogluasnglutyrserglyglycyscysglyvallystyrpro
180185190
aspalaasptyrmetglntyrvalmetaspglnalaarglysalaasp
195200205
ilevalvalpropheileserasnaspalaserproserglyhisasn
210215220
alaproglyserglythrseralavalaspiletyrglyhisaspser
225230235240
tyrproleuglypheaspcysalaasnproservaltrpproglugly
245250255
lysleuproaspasnpheargthrleuhisleugluglnserproser
260265270
thrprotyrserleuleuglupheglnalaglyalapheaspprotrp
275280285
glyglyproglypheglulyscystyralaleuvalasnhisgluphe
290295300
serargvalphetyrargasnaspleuserpheglyvalserthrphe
305310315320
asnleutyrmetthrpheglyglythrasntrpglyasnleuglyhis
325330335
proglyglytyrthrsertyrasptyrglyserproilethrgluthr
340345350
argasnvalthrargglulystyrseraspilelysleuleualaasn
355360365
phevallysalaserprosertyrleuthralathrproargasnleu
370375380
thrthrglyvaltyrthraspthrseraspleualavalthrproleu
385390395400
ileglyaspserproglyserphephevalvalarghisthrasptyr
405410415
serserglngluserthrsertyrlysleulysleuprothrserala
420425430
glyasnleuthrileproglnleugluglythrleuserleuasngly
435440445
argaspserlysilehisvalvalasptyrasnvalserglythrasn
450455460
ileiletyrserthralagluvalphethrtrplyslyspheaspgly
465470475480
asnlysvalleuvalleutyrglyglyprolysgluhishisgluleu
485490495
alailealaserlysserasnvalthrileilegluglyseraspser
500505510
glyilevalserthrarglysglyserservalileileglytrpasp
515520525
valserserthrargargilevalglnvalglyaspleuargvalphe
530535540
leuleuaspargasnseralatyrasntyrtrpvalprogluleupro
545550555560
thrgluglythrserproglypheserthrserlysthrthralaser
565570575
serileilevallysalaglytyrleuleuargglyalahisleuasp
580585590
glyalaaspleuhisleuthralaasppheasnalathrthrproile
595600605
gluvalileglyalaprothrglyalalysasnleuphevalasngly
610615620
glulysalaserhisthrvalasplysasnglyiletrpserserglu
625630635640
vallystyralaalaprogluilelysleuproglyleulysaspleu
645650655
asptrplystyrleuaspthrleuprogluilelyssersertyrasp
660665670
aspseralatrpvalseralaaspleuprolysthrlysasnthrhis
675680685
argproleuaspthrprothrserleutyrserserasptyrglyphe
690695700
histhrglytyrleuiletyrargglyhisphevalalaasnglylys
705710715720
glusergluphepheileargthrglnglyglyseralapheglyser
725730735
servaltrpleuasngluthrtyrleuglysertrpthrglyalaasp
740745750
tyralametaspglyasnserthrtyrlysleuserglnleugluser
755760765
glylysasntyrvalilethrvalvalileaspasnleuglyleuasp
770775780
gluasntrpthrvalglyglugluthrmetlysasnproargglyile
785790795800
leusertyrlysleuserglyglnaspalaseralailethrtrplys
805810815
leuthrglyasnleuglyglygluasptyrglnasplysvalarggly
820825830
proleuasngluglyglyleutyralagluargglnglyphehisgln
835840845
proglnproproserglusertrpgluserglyserproleuglugly
850855860
leuserlysproglyileglyphetyrthralaglnpheaspleuasp
865870875880
leuprolysglytrpaspvalproleutyrpheasnpheglyasnasn
885890895
thrglnalaalaargalaglnleutyrvalasnglytyrglntyrgly
900905910
lysphethrglyasnvalglyproglnthrserpheprovalproglu
915920925
glyileleuasntyrargglythrasntyrvalalaleuserleutrp
930935940
alaleugluseraspglyalalysleuglyserphegluleusertyr
945950955960
thrthrprovalleuthrglytyrglyasnvalgluserproglugln
965970975
prolystyrgluglnarglysglyalatyr
980985
<210>5
<211>32
<212>dna
<213>人工序列
<400>5
cgcgaggcagagatcttgagataaatttcacg32
<210>6
<211>33
<212>dna
<213>人工序列
<400>6
acgtgaaatttatctcaagatctctgcctcgcg33
<210>7
<211>40
<212>dna
<213>人工序列
<220>
<221>misc_feature
<222>(29)..(30)
<223>nisa,c,g,ort
<400>7
acttccaggagcattgtgcccagaaggmnnggcatcgttg40
<210>8
<211>31
<212>dna
<213>人工序列
<400>8
ttctgggcacaatgctcctggaagtggaacg31
<210>9
<211>36
<212>dna
<213>人工序列
<220>
<221>misc_feature
<222>(29)..(30)
<223>nisa,c,g,ort
<400>9
tgcgcaatcaaagccaaggggatagctmnngtgacc36
<210>10
<211>27
<212>dna
<213>人工序列
<400>10
tccccttggctttgattgcgcaaaccc27
<210>11
<211>36
<212>dna
<213>人工序列
<220>
<221>misc_feature
<222>(26)..(27)
<223>nisa,c,g,ort
<400>11
tgcgcaatcaaagccaaggggatamnnatcgtgacc36
<210>12
<211>32
<212>dna
<213>人工序列
<220>
<221>misc_feature
<222>(25)..(26)
<223>nisa,c,g,ort
<400>12
tttgcgcaatcaaagccaaggggmnngctatc32
<210>13
<211>30
<212>dna
<213>人工序列
<400>13
tggctttgattgcgcaaacccatccgtatg30
<210>14
<211>31
<212>dna
<213>人工序列
<220>
<221>misc_feature
<222>(24)..(25)
<223>nisa,c,g,ort
<400>14
atacggatgggtttgcgcaatcmnngccaag31
<210>15
<211>24
<212>dna
<213>人工序列
<400>15
tgcgcaaacccatccgtatggccc24
<210>16
<211>29
<212>dna
<213>人工序列
<400>16
tttcctcgccgaccgtccaattaacgtcg29
<210>17
<211>27
<212>dna
<213>人工序列
<400>17
tggacggtcggcgaggaaaccatgaag27
<210>18
<211>32
<212>dna
<213>人工序列
<400>18
cgcgaggcagagatcttgagataaatttcacg32
<210>19
<211>33
<212>dna
<213>人工序列
<400>19
acgtgaaatttatctcaagatctctgcctcgcg33
<210>20
<211>40
<212>dna
<213>人工序列
<400>20
acttccaggagcattgtgcccagaaggaakggcatcgttg40
<210>21
<211>40
<212>dna
<213>人工序列
<220>
<221>misc_feature
<222>(29)
<223>nisa,c,g,ort
<400>21
acttccaggagcattgtgcccagaaggmncggcatcgttg40
<210>22
<211>31
<212>dna
<213>人工序列
<400>22
ttctgggcacaatgctcctggaagtggaacg31
<210>23
<211>29
<212>dna
<213>人工序列
<400>23
tttcctcgccgaccgtccaattaacgtcg29
<210>24
<211>27
<212>dna
<213>人工序列
<400>24
tggacggtcggcgaggaaaccatgaag27
- 还没有人留言评论。精彩留言会获得点赞!