一种编码糖转运蛋白的氨基酸序列、突变体及应用的制作方法
本公开涉及酿酒酵母糖转运蛋白技术领域,具体涉及一种编码糖转运蛋白的氨基酸序列、核苷酸突变体、包含该序列的重组质粒及受体细胞在提高酿酒酵母糖转运能力方面的应用。
背景技术:
公开该背景技术部分的信息仅仅旨在增加对本公开的总体背景的理解,而不必然被视为承认或以任何形式暗示该信息构成已经成为本领域一般技术人员所公知的现有技术。
酿酒酵母(saccharomycescerevisiae)是广泛用于食品和工业生产的微生物,广泛应用于酿酒工业及燃料乙醇制造业,其具有鲁棒性强、代谢旺盛以及食品级安全等特性。酿酒酵母的生长和发酵与己糖转运蛋白家族密切相关,发酵过程中的酿酒酵母可以在有氧和无氧的条件下发酵已糖获取能量,酵母细胞的发酵效率与已糖转蛋白的活性息息相关。
木质纤维素资源种类繁多,年产量高,提高木糖组分的综合利用可以显著提高资源利用率,降低食品生产成本。自然的酿酒酵母不能利用木糖,仅能利用木酮糖。通过基因工程手段,在酿酒酵母中同时表达木糖还原酶和木糖醇脱氢酶,或者单独表达木糖异构酶可以赋予酿酒酵母将木糖转化为木酮糖的能力,进而通过相关代谢途径涉及的酶的优化表达,可以获得能够将木糖转化为各种目标产品的细胞工厂。
木糖向酿酒酵母细胞内的运输是木糖被利用的第一步。高水平表达糖转运蛋白可以促进酿酒酵母对木糖的吸收。酿酒酵母没有专门的木糖转运蛋白,其对木糖的吸收依赖于己糖转运蛋白。发明人认为,虽然有报道酿酒酵母本源的gal2蛋白是一个作用广泛、具有多种己糖及戊糖转运能力的糖转运蛋白(youngetal.2011,appliedandenvironmentalmicrobiology77(10):3311-3319),但是这些非特异性的转运蛋白对木糖的运输受到葡萄糖强烈的竞争性抑制,从而影响了菌株在有葡萄糖环境下的木糖利用效率(subtilandboles2012,biotechnologyforbiofuels5:14),对于相关微生物发酵食品及饲料的生产带来了限制。
技术实现要素:
针对上述技术问题,发明人认为,为了进一步提高酿酒酵母的生产效率,一方面可以通过基因改造的方式提高己糖的转运效率;另一方面,提升自然界中存在多种天然可利用木糖的微生物,从这些微生物中寻找优良的木糖转运蛋白,通过基因改造并导入酿酒酵母,可以消除底物对木糖转运的竞争性抑制,提高酿酒酵母对木糖的转运效率,降低生产成本,通过对糖转运蛋白的改造有望实现对酿酒酵母培养基的控制,获得具有碳源倾向性的酿酒酵母,为食品生产及乙醇燃料的制备提供更多的便利。
为了实现上述技术效果,本公开提供以下技术方案:
本公开第一方面,提供一种糖转运蛋白的氨基酸序列,所述氨基酸序列为以下序列之一:
(1)genebank中蛋白protein_id="xp_006966515.1"的氨基酸序列,该氨基酸序列命名为xltr1;
(2)所述(1)中氨基酸序列xltr1的突变体,所述突变体为:xltr1f300a、xltr1n326a、xltr1d39a、xltr1r132a、xltr1q288a、xltr1q291a、xltr1q292a、xltr1f298a、xltr1i299a、xltr1y301a、xltr1y302a、xltr1w407a、xltr1n434a、xltr1q292s、xltr1q292y、xltr1q292w、xltr1q292f、xltr1f300s、xltr1f300y、xltr1f300w、xltr1f300k、xltr1n326s、xltr1n326f、xltr1n326y、xltr1n326w;
(3)与(1)或(2)具有80%及以上相似性的氨基酸序列。
序列(1)即seqidno:1所示的氨基酸序列,其中优选的突变位点为xltr1f300a、xltr1n326f、xltr1q288a、xltr1q291a、xltr1q292a、xltr1f300a、xltr1n326a、xltr1n434a、xltr1q292s、xltr1q292f、xltr1f300s、xltr1f300y、xltr1f300w、xltr1f300k、xltr1n326s。
xltr1蛋白作为里氏木霉的胞外代谢物,具有糖转运蛋白活性,本公开的研究进一步发现,该蛋白可以作为酿酒酵母的糖转运蛋白,不仅能够有效提高酿酒酵母对葡萄糖、甘露糖及果糖的利用率,还能够增加显著增加酿酒酵母对木糖的利用率,是酿酒酵母本源蛋白gal2p的8.25倍,可以有效的提高酿酒酵母对木糖的利用率。
上述突变体xltr1f300a表示protein_id="xp_006966515.1"所指示的氨基酸序列其外显子第300位由苯丙氨酸f突变为丙氨酸a;
同理,xltr1n326a表示protein_id="xp_006966515.1"所指示的氨基酸序列其外显子第326位由天门冬酰胺n突变为丙氨酸a;其他突变体表示的含义可以以此类推。
上述突变体相比酿酒酵母本源蛋白均具有一定的技术效果,例如xltr1q288a、xltr1q291a、xltr1q292a、xltr1f300a、xltr1n326a、xltr1q292s、xltr1q292f、xltr1f300s、xltr1f300y、xltr1f300w、xltr1f300k、xltr1n326s等对碳源具有良好的利用效果;其中,xltr1f300a虽然转运葡萄糖的能力只有gal2p的67.5%,但是它的木糖转运能力几乎完全消失。另一方面,xltr1n326f葡萄糖转运能力完全消失,而木糖转运能力远远高于gal2p。将上述突变体应用于酿酒酵母,可以实现对底物中碳源的控制,实现更多的生产可能。
进一步优选的突变体为xltr1f300a、xltr1n326f、xltr1f300s、xltr1n434a。
xltr1f300s突变株在混合己糖(葡萄糖,甘露糖,果糖)利用条件下,对葡萄糖的利用速率加快,而对甘露糖及果糖的利用速度降低,可应用于混合糖的分离,去除混合糖中的葡萄糖成分,扩展了该菌株的应用。
xltr1n326f突变株在混合糖(葡萄糖,木糖)利用条件下,葡萄糖转运能力完全消失,而木糖转运能力远远高于gal2p,是它的2.37倍。
xltr1n434a为本公开首次发现的木糖利用关键位点,此前并未公开该突变后的氨基酸序列能够应用于糖蛋白转运。
本公开研究表明,xltr1f300a没有木糖转运能力,但保留了己糖的转运能力。该特性同样可以应用于混合糖的分离,此外,该突变结果可以说明此位点对于己糖的转运十分关键,为进一步开发已糖相关转运蛋白具有宝贵的借鉴作用。
本公开第二方面,提供一种编码糖转运蛋白的核苷酸序列,所述核苷酸序列为以下序列之一:
(1)genebank中geneid:18487109;
(2)由于密码子简并性能够翻译得到第一方面(1)部分所述序列的核苷酸序列;
(3)由于密码子简并性能够翻译得到第一方面(2)部分所述突变体的核苷酸序列。
(4)与第二方面(1)或(2)或(3)部分所述核苷酸序列具有80%及以上相似性的核苷酸序列。
本公开第三方面,提供一种重组质粒,所述重组质粒具有第二方面所述核苷酸序列的完整编码阅读框序列。
优选的,所述重组质粒的载体为pjfe3。
本公开第四方面,提供一种表达糖转运蛋白的受体细胞,所述受体细胞内包含第一方面所述糖转运蛋白的氨基酸序列和/或第二方面所述编码糖转运蛋白的核苷酸序列和/或第三方面所述的重组质粒。
优选的,所述受体细胞为细菌或真菌;进一步优选的,为酿酒酵母。
本公开第五方面,提供一种提高酿酒酵母糖吸收和利用能力的方法,该方法包括以下步骤:将第三方面所述的重组质粒导入酿酒酵母细胞中。
本公开第六方面,提供第一方面所述糖转运蛋白的氨基酸序列、第二方面所述糖转运蛋白的核苷酸序列、第三方面所述重组质粒或第四方面所述受体细胞在提高酿酒酵母糖利用方面的应用。
优选的,所述应用为利用酿酒酵母生产木糖醇、乙醇、多元醇、有机酸、烃类或萜类化合物中的应用。
与现有技术相比,本公开的有益效果:
1.本公开提供了xltr1蛋白作为酿酒酵母糖转运蛋白的应用,针对自然酿酒酵母细胞不能利用木糖的现状,本公开提供了该蛋白作为酿酒酵母的糖转运蛋白可以有效的提高酿酒酵母对碳源的利用率及对木糖的运输能力。通过基因改造的方式将核苷酸序列导入酿酒酵母细胞中,使细胞可以木糖作为碳源进行发酵,且同一培养条件下,导入本公开序列得酿酒酵母对麦芽糖的利用率显著升高,远远超过对葡萄糖的利用率。将本申请中的序列导入酿酒酵母,可以有效消除培养基中其他碳源的竞争性抑制,提高细胞对木糖的利用率,有望带来良好的经济效益。
2.本公开还提供了xltr1蛋白的突变体,作为酿酒酵母的糖转运蛋白,同样具有良好的运输活性,除了提高酿酒酵母对碳源的利用率,还有望实现控制酿酒酵母对单一碳源的摄取,实现更多的生产方式。
附图说明
构成本公开的一部分的说明书附图用来提供对本公开的进一步理解,本公开的示意性实施例及其说明用于解释本公开,并不构成对本公开的不当限定。
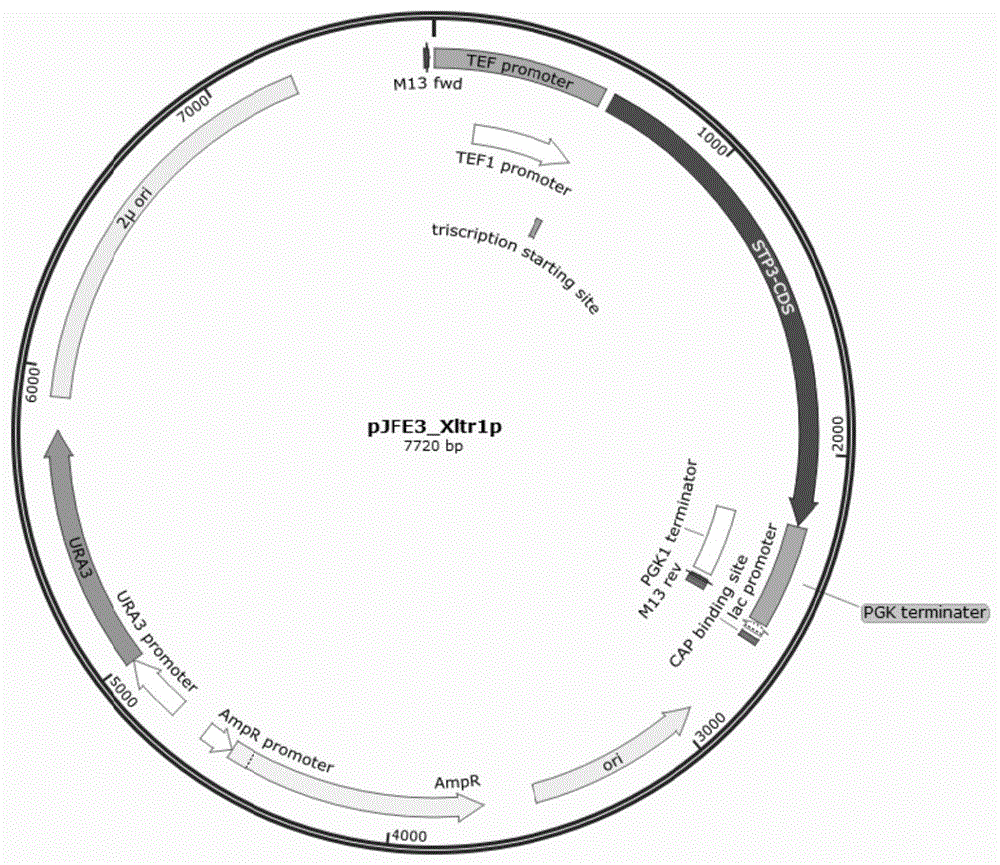
图1为实施例2中重组质粒pjfe3-xltr1物理图谱;
图2为实施例4中表达xltr1不同突变蛋白菌株在培养基生长状况图。
具体实施方式
应该指出,以下详细说明都是例示性的,旨在对本公开提供进一步的说明。除非另有指明,本文使用的所有技术和科学术语具有与本公开所属技术领域的普通技术人员通常理解的相同含义。
需要注意的是,这里所使用的术语仅是为了描述具体实施方式,而非意图限制根据本公开的示例性实施方式。如在这里所使用的,除非上下文另外明确指出,否则单数形式也意图包括复数形式,此外,还应当理解的是,当在本说明书中使用术语“包含”和/或“包括”时,其指明存在特征、步骤、操作、器件、组件和/或它们的组合。
正如背景技术所介绍的,酿酒酵母中糖转运蛋白的转运能力与酵母细胞发酵活力密切相关,提供具有良好转运能力的己糖转运蛋白有利于提高酿酒酵母的发酵效率;另一方面,通过基因改造使酿酒酵母获得木糖发酵能力,有利于提高资源利用率,提高生产效益。为了实现上述技术目的,本公开提供了xltr1蛋白作为酿酒酵母糖转运蛋白的应用及其他同样具有糖转运活性的突变体。
为了使得本领域技术人员能够更加清楚地了解本公开的技术方案,以下将结合具体的实施例与对比例详细说明本公开的技术方案。
以下实施例中:
(1)酿酒酵母:酿酒酵母菌株eby.vw4000(matαleu2-3,112ura3-52trp1-289his3-δ1mal2-8csuc2hxt17δhxt13δ::loxphxt15δ::loxphxt16δ::loxphxt14δ::loxphxt12δ::loxphxt9δ::loxphxt11δ::loxphxt10δ::loxphxt8δ::loxphxt514::loxphxt2δ::loxphxt367δ::loxpgal2pδstl1δ::loxpagt1δ::loxpydl247wδ::loxpyjr160cδ::loxp)(wieczorkeetal.1999,febsletters464(3):123-128)是一株缺失了所有內源己糖转运蛋白的菌株(hxtnull),其因缺乏向胞内运输己糖的能力不能在葡萄糖上生长,而只能在麦芽糖这种具有自身特异性运输蛋白的碳源底物上生长。在该菌株中表达糖转运蛋白可以使菌株重新在葡萄糖及其他己糖为单独碳源的培养基上生长,其生长能力可以反映出所表达转运蛋白的葡萄糖或者其他己糖运输能力。将表达糖转运蛋白的eby.vw4000在木糖中孵育一定时间,进而测定细胞内木糖和木糖醇的总积累量,可以反映所表达转运蛋白对木糖的运输能力。当在eby.vw4000中引入木糖利用途径之后,菌株在木糖为唯一碳源上的生长能力可以反映转运蛋白对木糖的运输能力。
(2)xltr1为seqidno:1所示的氨基酸序列。
(3)利用生物信息学手段,通过同源建模和分子对接,预测xltr1蛋白的蛋白结构,以及通过对保守氨基酸残基的分析,并根据xltr1蛋白与葡萄糖、果糖、甘露糖和木糖分子的对接结果,优选氨基酸位点进行突变。
下面结合实施例对本发明作进一步的说明。
实施例1转运蛋白编码基因的克隆
提取酿酒酵母实验室菌株基因组作为模板,以gal2p-f及gal2p-r为引物,进行gal2cds区的扩增,退火温度同样选择为56℃,使用clontech公司的in-fusionhdcloning高效克隆试剂盒将gal2片段与pjfe3质粒连接。提取trichodermareeseiqm6a(atcc13631)的rna进行反转录得到cdna作为模板,用toyobo公司的kod高保真酶进行pcr扩增,通过infusion的方法将xltr1片段与pjfe3质粒连接。再使用点突变试剂盒kod-plus-mutagenisis引入表1的定点突变。pcr扩增dna片段xltr1,其pcr扩增时选择的退火温度为56℃。引物列表如图1。
表1实验所用引物列表
备注:
1.“f”表示正向引物,“r”表示反向引物。
2.f300x-r/n326x-r等代表xltr1的突变前后的氨基酸残基,f300x代表f300a/s/w/y;n326x代表n326a/s/f。
实施例2构建pjfe3_xltr1p表达质粒
如图1所示,该重组质粒为酿酒酵母2μ质粒,gal2p和xltr1基因在酿酒酵母组成型启动子tef1启动子控制下表达;将gal2p和xltr1基因的dna片段采用gb-dir的方法连在pjfe3质粒上。通过对片段的测序筛选到正确的单克隆转化子,其中大小分别为7817bp,7720bp的质粒为正确的重组质粒,该重组质粒命名为pjfe3-gal2p和pjfe3-xltr1p,在该质粒上,gal2p和xltr1p基因在酿酒酵母组成型启动子tef1启动子控制下表达。
实施例3在酿酒酵母中表达转运蛋白
用空载体质粒pjfe3转化酿酒酵母菌株eby.vw4000,在添加20g/l麦芽糖的sc-ura营养缺陷型培养基平板上筛选转化子,未被转化的细胞不能在该平板上生长。获得的转化子命名为bsw4pp。
将重组质粒pjfe3-gal2p和pjfe3-xltr1p转化菌株eby.vw4000,在添加20g/l麦芽糖的sc-ura营养缺陷型培养基平板上筛选转化子,未被转化的细胞不能在该平板上生长。获得的转化子命名为fjtr3_xltr1。
将重组质粒pjfe3-xltr1的不同位点突变转化菌株eby.vw4000,在添加20g/l麦芽糖的sc-ura营养缺陷型培养基平板上筛选转化子,未被转化的细胞不能在该平板上生长。突变后的转化子分别命名为fjtr3_xltr1f300a和fjtr3_xltr1n326f等(见表2和表3)。
其中,酿酒酵母转化使用电转化的方法(costagliolietal.1994,currentgenetics27(1):26-30)。
其中,sc-ura营养缺陷培养基中含有1.7g/lynb(酵母基础氮源,购自上海生工生物工程股份有限公司),5g/l硫酸铵,0.77g/lsc-ura(购自mpbiomedicals);固体培养基另外添加20g/l琼脂粉,ph调为6.0~7.0;灭菌条件:115℃,30min;使用时添加终浓度为20g/l的麦芽糖作为碳源。
实施例4在不同的碳源上酵母的生长变化
如图2所示,转化菌株bsw4pp和fjtr3_xltr1在添加20g/l甘露糖或者葡萄糖或者木糖或者果糖为唯一碳源的培养基平板上培养。结果显示,对照空载菌株bsw4pp在木糖培养基上不能生长,在甘露糖和果糖培养基上仅仅能微弱生长,而菌株fjtr3_xltr1在木糖,甘露糖和果糖培养基上生长得很好(图2)。
实施例5木糖积累量法比较运输蛋白的木糖运输能力
在添加20g/l麦芽糖的sc-ura营养缺陷型培养基中于30℃下培养菌株fjtr3_xltr1及其突变菌株,12小时后转接培养物到新鲜的添加20g/l麦芽糖的sc-ura营养缺陷型培养基中,转接量为10%。于30℃下继续培养10小时后,8000转/分离心3分钟收集菌体。然后将细胞重悬于添加20g/l木糖的sc-ura营养缺陷型培养基中。调节菌株fjtr3_xltr1及其突变菌株的菌悬液od600一致,优选od600均等于20,至于30℃孵育。在孵育30min,60min和120min时,分别取各菌株10ml菌悬液,8000转/分离心3分钟,弃上清。用10ml温度为4℃的无菌水洗菌泥两次。洗的方法是用10ml温度为4℃的无菌水重悬细胞,然后8000转/分离心3分钟,弃上清。洗过的细胞重悬于3ml去离子水中,于37℃培养箱中过夜放置后离心,用hplc测定上清液中的木糖和木糖醇含量,根据其中木糖和木糖醇总含量的摩尔数,计算菌体细胞吸收的木糖总量(duetal.2010,molbiosyst6(11):2150-2156.)。用分光光度计biophotometerplus(购自eppendorf,德国)测定菌悬液的od600,并根据公式:细胞干重dcw(mg/ml)=0.2154*od600+0.1358,将细胞量换算为菌体干重。糖的转运能力用单位菌体细胞吸收的木糖总量来表示,其计量单位为mg/gdcw。结果见表2。
实施例6以生长速率比较的方法比较转运蛋白转运葡萄糖的能力
在添加20g/l麦芽糖的sc-ura营养缺陷型培养基中于30℃下培养菌株fjtr3_gal2p、fjtr3_xltr1及其突变菌株,12小时后转接培养物到新鲜的添加20g/l麦芽糖的sc-ura营养缺陷型培养基中,转接量为10%。于30℃下继续培养12小时后8000转离心3分钟收集菌体。然后用无菌双蒸水洗涤2次后重悬于无菌双蒸水中,转接10%菌体量到新鲜的添加20g/l葡萄糖的sc-ura营养缺陷型培养基中。调节菌悬液初始od600为1,30℃200rpm培养5天。结果见表2。
表2表达xltr1及其突变蛋白的菌株葡萄糖和转运木糖能力的比较
从表2可见,和酿酒酵母的本源蛋白gal2p相比,xltr1表达的酵母,木糖转运能力更强,是gal2p的8.25倍,而xltr1的突变体xltr1f300a虽然转运葡萄糖的能力只有gal2p的67.5%,但是它的木糖转运能力几乎完全消失。另一方面,xltr1的突变体xltr1n326f,它的葡萄糖转运能力完全消失,而木糖转运能力远远高于gal2p,是它的2.37倍。
实施例7
本实施例中,发明人针对以下突变位点编码的转运蛋白进行筛选,参照实施例1-3的方法构建重组质粒,并导入酿酒酵母菌株eby.vw4000中,考察以葡萄糖、甘露糖及果糖为碳源时的糖利用效率
1.获取方式:
利用生物信息学手段,通过同源建模和分子对接,根据xltr1蛋白与葡萄糖、果糖和甘露糖分子的对接结果,以及保守氨基酸序列的分析,优选氨基酸位点进行突变。
2.活性验证具体步骤:
a活化菌株:首先平板活化,后挑取单菌落在sc-ura添加麦芽糖的液体培养基中进行过夜培养,第二天转接至新鲜培养基,初始od600调整至0.1-0.2,培养16-18h,使od600在2.0左右;
b将菌体进行离心,并使用无菌水反复洗涤3次,至无残留培养基;
c转接至含有不同碳源(葡萄糖、甘露糖、果糖)培养基中进行;
d在不同时间点进行取样:0h,6h,12h,24h,36h,48h,72h;
e使用高效液相色谱对样品中剩余糖含量进行检测,结果如下表3所示:
表3表达xltr1及其突变蛋白的菌株在混合糖(葡萄糖、甘露糖、果糖)中糖利用速率的比较
注释:t30,t50:分别代表糖消耗30%和50%所消耗的时间,可以表征糖的消耗速率;nq代表无法计算时间,糖消耗30%和50%所消耗的时间约为无穷大。
突变体xltr1q291a、xltr1f300s、xltr1f300y、xltr1n326a、xltr1n326s在混合糖利用中,对葡萄糖的利用速率发生了明显的加快,突变体xltr1n326a、xltr1n326s对甘露糖的利用速率变慢,突变体xltr1q291a、xltr1n326s对甘露糖的利用速率则明显变快,突变体xltr1q288s、xltr1q291a、xltr1f300y、xltr1n326a、xltr1n326s对果糖的利用速率明显变快,突变体xltr1q292a、xltr1q292s、xltr1f300s、xltr1f300w对果糖的利用速率变慢。因此,可以通过将上述突变体导入酿酒酵母的方式实现对碳源的选择性利用及混合糖的分离。
以上所述仅为本公开的优选实施例而已,并不用于限制本公开,对于本领域的技术人员来说,本公开可以有各种更改和变化。凡在本公开的精神和原则之内,所作的任何修改、等同替换、改进等,均应包含在本公开的保护范围之内。
sequencelisting
<110>荣成市慧海创达生物科技有限公司
<120>一种编码糖转运蛋白的氨基酸序列、突变体及应用
<130>2010
<160>22
<170>patentinversion3.3
<210>1
<211>543
<212>prt
<213>人工序列
<400>1
metargpheserglulysleuglyphelysargproaspaspgluala
151015
glyalasertrpvalalaileleumetglyphephevalalaphegly
202530
glyvalleutyrglytyraspthrglythrileserglyilemetala
354045
metprotyrphelysaspleupheserthrglytyrargasnproasn
505560
glygluleuaspilethralathrglngluseralailevalserile
65707580
leuseralaglythrphepheglyalaleualaserproleuleuala
859095
asppheleuglyargargproalaleumetileserthrtrpvalphe
100105110
asnleuglyvalvalleuglnthrilealathralaileprometphe
115120125
leualaglyargphephealaglypheglyvalglyleuileserala
130135140
leuileproleutyrglnsergluthralaprolystrpilearggly
145150155160
alailevalglyalatyrglnleualailethrileglyleuleuleu
165170175
alaalavalvalasnasnalathralalysarghisaspserglyser
180185190
tyrargileproilealavalglnphealatrpserleuvalleuphe
195200205
valglymetilepheleuprogluthrproargpheleuvalargser
210215220
glylysleuglulysalaargalaalaleuserargileargargleu
225230235240
serprogluhisglualaleualaalagluleuglyglnileglnala
245250255
asnleuglualagluserservalarglysalathrtyralaaspcys
260265270
pheargargprometleulysargglnphethrglymetalaleugln
275280285
alaleuglnglnleuthrglyileasnpheilephetyrtyrglythr
290295300
argtyrpheglnasnserglyvalserserglyphethrileglymet
305310315320
ilethralaglyileasnvalalaserthrileproglyleuleuala
325330335
ileaspargtrpglyargargproleuleuleuleuglyalavalgly
340345350
metcysvalserglnleuilevalalavalvalglythrvalserthr
355360365
glyglnargproasnglygluilephevallysserleualaglygln
370375380
glnalaalavalalaphevalcysilepheilealaphephealaser
385390395400
thrtrpglyproleualatrpvalvalthrglygluiletyrproleu
405410415
alathrargalalysalaleusermetthrthralathrasntrpleu
420425430
pheasntrpalailealatyrserthrprotyrleuvalasntyrgly
435440445
proglytyralaasnleuglnserlysilephephevaltrpphegly
450455460
alacyspheleucysilealaleuvaltrpphepheiletyrgluthr
465470475480
lysglyleuserleuglugluvalaspgluleutyralagluvallys
485490495
valalaarglysserthrthrtrplysprothrproargleugluala
500505510
alaglyserthrthrserglugluserlysaspgluserglyprolys
515520525
glualaserprohisvalmetaspglnglyvalgluleuglnval
530535540
<210>2
<211>1632
<212>dna
<213>人工序列
<400>2
atgcgtttctccgagaagctcggcttcaagcgccccgacgatgaggccggcgcttcatgg60
gtcgccatcctcatgggcttctttgtcgccttcggcggtgtcctctacggatatgacact120
ggcaccatttccggaatcatggccatgccatacttcaaggatctcttcagcacgggatac180
cgcaaccccaacggcgagctcgacatcaccgccactcaggagagcgccatcgtctccatc240
ctgtcggctggtactttcttcggcgctcttgcgtcccctcttctggccgacttcctgggc300
cgccgccctgccttgatgatctcgacttgggtcttcaacctgggcgtcgtcctccaaaca360
atcgcaacggcaatccccatgttcctggctggtcgcttctttgccggcttcggcgtcggt420
ctcatctccgccttgatccccctgtaccagtccgaaacagcacccaaatggatccgcggc480
gccatcgtcggcgcctaccaactggccatcacaatcggcctcctcctcgccgccgtcgtc540
aacaacgccacggccaagcgccacgactcgggcagctaccgcatccccatcgccgtccag600
ttcgcctggtccctcgtcctcttcgtcggcatgatcttcctgcccgagacgccgcgcttc660
ctcgtccgctccggcaagctcgagaaagccagggccgccctctcgcgcatccgccgcctg720
tccccggagcacgaggctctcgccgccgagctcggccagatccaggccaatctcgaggcc780
gagagcagcgtccgcaaggccacgtacgcagactgcttccgccgccccatgctcaagcgc840
cagttcaccggcatggccctgcaggcgctgcagcagctgacgggcatcaacttcatcttc900
tactacgggacgcggtatttccagaactcgggcgtctccagcggcttcaccattggcatg960
atcacggcgggcatcaacgtcgcctccaccatccccggtctcttggccattgaccgctgg1020
ggccgccgccctctgctcctcctcggcgccgtcggcatgtgcgtctcccagctcatcgtc1080
gccgtggtcggcacagtttccaccggccagcgccccaacggagaaatctttgtaaagtcc1140
ctggccggccagcaggcggccgtcgccttcgtctgcatcttcatcgccttcttcgccagc1200
acctggggccccctcgcgtgggtcgtcaccggcgaaatctacccgctggccacgcgggca1260
aaggccctgagcatgaccacggccacaaactggctcttcaactgggccattgcctactcc1320
accccttacctcgtcaactacggcccgggctacgccaacctgcagtccaagatcttcttc1380
gtctggttcggcgcctgcttcctctgcatcgcccttgtctggttcttcatctacgagacc1440
aagggccttagcctggaggaggtcgacgagctctatgccgaggtcaaggtcgcccgcaag1500
tctaccacctggaagcccaccccgcgcctggaggctgctggaagtaccaccagcgaggag1560
tcaaaggatgagtctggtcccaaggaggcgagtcctcatgttatggaccaaggagttgaa1620
cttcaagtgtaa1632
<210>3
<211>25
<212>dna
<213>人工序列
<400>3
atggcagttgaggagaacaatatgc25
<210>4
<211>23
<212>dna
<213>人工序列
<400>4
cttattctagcatggccttgtac23
<210>5
<211>19
<212>dna
<213>人工序列
<400>5
ttcgtcgcctccaccatcc19
<210>6
<211>19
<212>dna
<213>人工序列
<400>6
agagtcgcctccaccatcc19
<210>7
<211>19
<212>dna
<213>人工序列
<400>7
gcggcgctgcagcagctga19
<210>8
<211>19
<212>dna
<213>人工序列
<400>8
cagggccatgccggtgaac19
<210>9
<211>21
<212>dna
<213>人工序列
<400>9
gcgcagctgacgggcatcaac21
<210>10
<211>19
<212>dna
<213>人工序列
<400>10
cagcgcctgcagggccatg19
<210>11
<211>21
<212>dna
<213>人工序列
<400>11
caggcgctgacgggcatcaac21
<210>12
<211>19
<212>dna
<213>人工序列
<400>12
cagcgcctgcagggccatg19
<210>13
<211>20
<212>dna
<213>人工序列
<400>13
gcctactacgggacgcggta20
<210>14
<211>20
<212>dna
<213>人工序列
<400>14
tggtactacgggacgcggta20
<210>15
<211>20
<212>dna
<213>人工序列
<400>15
tactactacgggacgcggta20
<210>16
<211>21
<212>dna
<213>人工序列
<400>16
gatgaagttgatgcccgtcag21
<210>17
<211>19
<212>dna
<213>人工序列
<400>17
gccgtcgcctccaccatcc19
<210>18
<211>19
<212>dna
<213>人工序列
<400>18
gatgcccgccgtgatcatg19
<210>19
<211>19
<212>dna
<213>人工序列
<400>19
ttcgtcgcctccaccatcc19
<210>20
<211>19
<212>dna
<213>人工序列
<400>20
tgggtcgcctccaccatcc19
<210>21
<211>19
<212>dna
<213>人工序列
<400>21
tacgtcgcctccaccatcc19
<210>22
<211>19
<212>dna
<213>人工序列
<400>22
gatgcccgccgtgatcatg19
- 还没有人留言评论。精彩留言会获得点赞!