人源抗H7N9禽流感病毒中和性抗体1F7L及其应用的制作方法
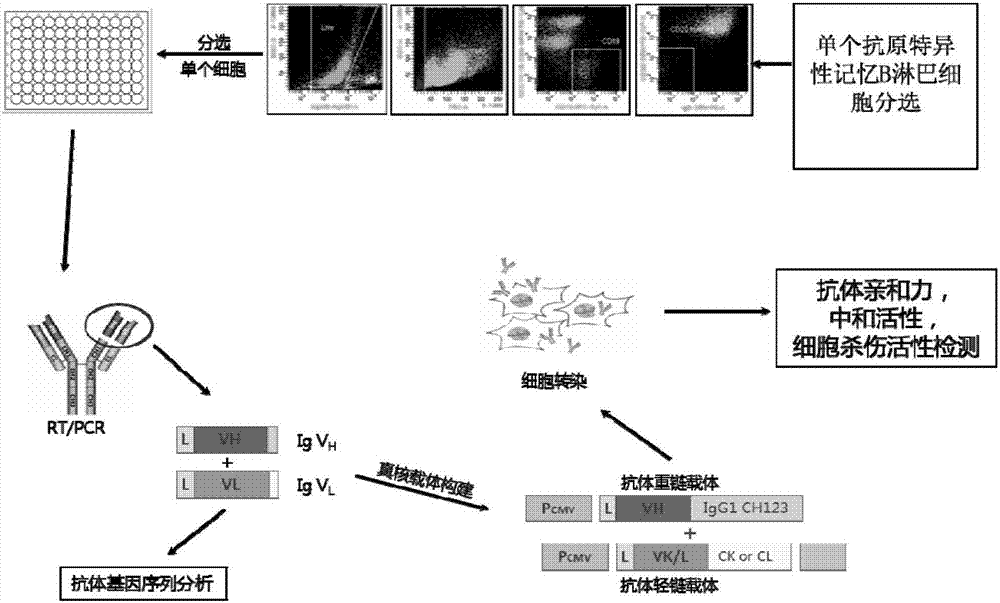
本发明涉及基因工程、单细胞分选技术及抗体文库展示技术领域,具体地说,涉及一种人源抗h7n9禽流感病毒中和性抗体1f7l及其应用。
背景技术:
自2013年2月以来,中国东南部陆续发现了由甲型h7n9禽流感病毒引起人感染病例,由上海、安徽逐渐蔓延到全国其他省市。截至2017年2月,全国共报道1079例h7n9流感感染,死亡病例较高。目前并无治疗该高致病型禽流感感染的特效药,而现有非特异性药物神经氨酸酶抑制剂-奥司他韦的疗效仅局限于感染早期。由于高致病性h7n9感染的前期症状与普通流感并无明显区别,大部分感染者错过最佳治疗时间而发展为急性呼吸窘迫综合征(ards)和多器官衰竭。高致病性禽流感病毒的抗原快速、灵敏诊断需求迫切。
单克隆抗体是由单一b细胞克隆产生的高度均一、仅针对某一特定抗原表位的免疫球蛋白。人源单克隆抗体由于副作用小,其应用于人类疾病治疗通常不需要进行人源化,避免了此过程中的亲和力丢失。人源单克隆抗体在感染性疾病的预防和控制过程中发挥了巨大作用,其能中和病毒,还能介导效应细胞对病毒感染细胞的杀伤,其保护效果在hiv、流感病毒、mers病毒,登革热病毒,汉坦病毒、麻疹病毒、rsv病毒、狂犬病毒等动物感染模型中得到充分的证实。
2008年以来,单个b细胞分选和抗体基因直接扩增技术成为人源抗体筛选的主要途径之一(tiller等,journalofimmunologicalmethods329,(2008),112–124)。2010年,wu等(science329,(2010)856-861)利用流式细胞分选技术分选hiv感染者外周血中抗原特异性的单个记忆性b细胞,进而利用逆转录pcr技术直接扩增单个细胞的抗体基因vh和vκ/vλ,然后将上述基因片段插入全长igg重组载体进行真核细胞转染和表达纯化,成功获得著名的hiv广谱中和单克隆抗体vrc01。由于该技术可以在最短的时间内获得单细胞来源的抗体基因和重组抗体蛋白,并且可以保证抗体重链和轻链的原始配对(抗体功能最优),单细胞分选和抗体基因直接扩增技术迅速成为抗体开发领域的重磅工具。迄今为止,该技术已在hiv,流感,mers,埃博拉,登革热等病毒的广谱中和单克隆抗体的筛选上成功运用,该技术获得的多个高效抗体先后进入药物临床研究。单个b细胞来源的基因工程抗体为抗原快速检测和抗体制药领域带来了新的希望和广阔前景。
以往的抗体开发主要以亲和力指标和中和能力指标对抗体进行衡量,然而最近的研究发现,不具有中和能力的抗体同样也可以实现对机体的保护,其抗体依赖细胞介导的细胞毒性杀伤(adcc)功能发挥了关键作用。著名的hiv临床试验rv144研究结果表明,疫苗的31%的保护效果与其所诱导抗体的adcc能力显著正相关,提示抗体adcc功能的重要性。目前已有的h7n9亚型特异性单克隆抗体具有一定的体外中和活性,并能保护动物免受感染或减轻症状,然而这些抗体并未报道或并不具有介导细胞依赖的毒性杀伤(adcc)能力,这将削弱以该类抗体为效应分子的药物的临床疗效。因此,既具有体外病毒中和活性又具有介导细胞毒性杀伤能力的全人源单克隆抗体将更有希望成为治疗性抗体药物。本发明的抗体具有体外病毒中和活性和介导病毒感染细胞杀伤的能力。
技术实现要素:
本发明的目的是提供一种人源抗h7n9禽流感病毒中和性抗体1f7l及其应用。
本发明的构思如下:全人源抗体的获得目前主要通过抗体文库展示筛选技术和单个b细胞分选技术。与单细胞分选法相比,抗体文库筛选获得的单克隆抗体原始轻重链配对的概率较低,而抗体的原始轻重链配对可能使抗体的功能实现最大化。单细胞分选法获得的抗体基因的轻重链,理论上均是来自于原始配对,因此,单细胞分选法获得高效功能性抗体的概率远高于抗体文库展示筛选法。
为了实现本发明目的,本发明人源抗h7n9禽流感病毒中和性抗体1f7l或其活性片段,所述中和性抗体1f7l或其活性片段的轻链及重链高变区cdr1、cdr2和cdr3的氨基酸序列如下表所示:
前述的中和性抗体1f7l,i)其轻链可变区的氨基酸序列如seqidno.2所示,或该序列经替换、缺失或添加一个或几个氨基酸形成的具有同等功能的氨基酸序列;以及
ii)其重链可变区的氨基酸序列如seqidno.5所示,或该序列经替换、缺失或添加一个或几个氨基酸形成的具有同等功能的氨基酸序列。
前述中和性抗体1f7l的轻链(vl)、重链(vh)全长氨基酸序列分别如seqidno.3和seqidno.6所示。
本发明还提供编码所述中和性抗体1f7l的基因。其中,编码轻链可变区和重链可变区的核苷酸序列分别如seqidno.1和seqidno.4所示。
本发明还提供表达盒、表达载体或克隆载体,其包括包含编码所述中和性抗体1f7l的基因序列的核酸。
本发明还提供含有所述中和性抗体1f7l的编码基因,或所述表达盒、载体的宿主细胞。
本发明还提供所述中和性抗体1f7l或其活性片段经改造得到的单链抗体scfv或fab抗体或全抗体免疫球蛋白igg。
本发明中所述h7n9流感病毒中和性抗体1f7l的活性片段是指能够与7型血凝素蛋白结合的人源h7n9中和抗体1f7l的fab片段。
本发明所述的人源抗h7n9禽流感病毒中和性抗体1f7l,可按如下方法制备:利用单个记忆性b细胞分选和抗体基因直接扩增法获得h7n9病毒血凝素蛋白特异性抗体的可变区片段,随后通过基因工程方法构建完整igg抗体的真核瞬间表达载体,并表达纯化出igg蛋白。
该抗体是由存在于抗体轻链和重链基因可变区中的高变区(cdrs)特异性基因序列决定的,并能够在真核细胞中获得有效表达的特异性结合h7n9流感病毒血凝素蛋白的功能性抗体。它能高效结合7型血凝素蛋白,高效结合h7n9病毒感染的mdck细胞,并具有抗h7n9流感病毒的中和活性功能和介导效应细胞对h7n9病毒感染细胞的杀伤功能。
h7n9流感病毒特异性抗体1f7l的轻链和重链基因来源于h7n9禽流感病毒感染患者外周血b细胞。其轻链和重链可变区相应的3个cdr区序列组合及其cdr区之间的框架区序列构成了该抗体可变区序列特征,抗体1f7l的轻链属于家族iglv1-47,重链属于家族ighv3-20。抗体蛋白功能由存在于抗体基因轻链和重链可变区的互补决定簇cdr1、cdr2和cdr3中特异性核苷酸序列及其互补序列所决定,6个相应的cdr区氨基酸序列构成了抗体的特异性抗原结合区域,决定了本发明抗体的抗原结合特征和抗h7n9禽流感病毒功能特征。
此外,考虑到密码子的简并性,例如可在其编码区,在不改变氨基酸序列的条件下,对编码1f7l可变区的基因序列进行改造,获得编码具有相同功能的抗体的基因。本领域技术人员可以根据表达抗体宿主的密码子偏爱性,人工合成改造基因,以提高抗体的表达效率。
进一步地,本发明所述h7n9流感病毒中和性抗体1f7l的轻链可变区和重链可变区可经过重组以形成较小分子量的fab抗体或更小分子量的单链抗体(scfv)。fab抗体和单链抗体同样具有识别h7n9流感病毒表面抗原的特性。小分子量的抗体具有穿透力强,易进入局部组织或细胞内发挥作用。
可将上述编码fab抗体的基因、scfv抗体的基因克隆到表达载体,进而转化宿主,通过诱导表达获得fab抗体和单链抗体(scfv)。
利用sds-page、elisa、体外病毒中和实验、抗体依赖细胞介导的细胞毒性杀伤(adcc)实验等方法对获得的igg抗体1f7l进行功能鉴定,结果表明表达纯化的人源igg抗体1f7l分子量大小符合预期(图2),其可以高效特异性结合7型血凝素蛋白,具有h7n9流感病毒中和活性,还具有介导效应细胞对h7n9病毒感染细胞的杀伤(adcc)活性。
本发明还提供所述中和性抗体1f7l或其活性片段在制备预防或治疗由h7n9禽流感病毒引起的疾病,特别是呼吸道疾病的药物中的应用。
本发明还提供所述中和性抗体1f7l或其活性片段在制备h7n9禽流感病毒抗原检测试剂或检测试剂盒中的应用。
本发明进一步提供含有所述中和性抗体1f7l或其活性片段的药物、检测试剂或检测试剂盒。
本发明采用多色荧光标记的流式细胞仪分选法获得7型血凝素蛋白抗原特异性记忆性b细胞,利用逆转录试剂盒获得单个b细胞的cdna,利用抗体特异性引物扩增抗体基因可变区片段,利用基因工程技术构建真核igg表达载体,利用细胞工程技术通过瞬间转染和抗体纯化技术获得纯化的目标igg抗体1f7l;利用上述获得的人源中和性h7n9禽流感病毒基因工程抗体可变区基因,可以以fab抗体、单链抗体基因及全长igg抗体的形式在原核细胞、真核细胞(包括酵母细胞)及任何重组蛋白表达系统中表达和生产该抗体,或以上述可变区基因为基础的改造后的含有该抗体基因的任何其他基因,获得具有中和h7n9禽流感病毒感染或杀伤病毒感染细胞的抗体产物,制成临床上用于预防和治疗由h7n9禽流感病毒引起的疾病,如急性呼吸道传染病的特异性抗体药物。
本发明提供的抗体1f7l可以高效结合h7n9流感病毒血凝素蛋白ha7,其针对ha7蛋白亲和力的ec50<0.0115μg/ml(0.077nm)(图3)。具体地,本发明提供的抗体1f7l在与7型血凝素蛋白亲和力检测的elisa实验中,当抗体浓度低至0.0115μg/ml((0.077nm)时,其od450值仍可以达到最大值的一半。本发明提供的抗体可以用于h7n9流感病毒抗原检测试剂的开发。
本发明提供的抗体1f7l可以特异性地识别h7n9流感病毒血凝素蛋白ha7,且其识别位点位于ha7的颈部区域即ha2段。通过elisa实验检测igg1f7l与h1n1,h5n1和h7n9流感病毒的血凝素蛋白的亲和力,结果发现igg1f7l特异性地识别h7n9的血凝素蛋白,对h1n1和h5n1流感病毒的血凝素蛋白不具有结合能力,说明其为ha7亚型特异性抗体;同时其能结合全长ha7血凝素蛋白,但是却不能结合ha7的ha1段,暗示其识别位点位于ha7的ha2段(图4)。本发明提供的抗体可以用于h7n9流感病毒抗原检测试剂的开发。
本发明提供的抗体1f7l可以有效地中和h7n9流感病毒,防止h7n9流感病毒感染mdck细胞。具体地,在体外中和实验中,1.85μg/ml的1f7l抗体浓度可以实现对200tcid50的h7n9病毒对mdck细胞感染的50%的抑制,也即ic50=1.85μg/ml(图5)。本发明提供的抗体可以用于h7n9流感病毒感染患者的治疗。
本发明提供的的抗体可以有效地介导效应细胞(如nk细胞)对h7n9感染细胞的杀伤。具体地,10μg/ml的1f7l抗体介导的针对h7n9感染的mdck细胞的杀伤百分数可达27.13%(图6)。本发明提供的抗体可以用于h7n9流感病毒感染患者的治疗。
附图说明
图1为本发明基于单细胞分选和抗体基因扩增的全人源单克隆抗体技术平台示意图。
图2为本发明igg1f7l的变性和非变性sds-page电泳检测抗体分子量和纯度。
图3为本发明实施例中igg1f7l针对ha7蛋白亲和力检测结果(elisa法)。其中,ha7蛋白以1μg/ml的浓度包被在elisa板上,igg1f7l从333.3nm开始4倍梯度稀释。当od450值为最高值一半时抗体1f7l的浓度即为其针对ha7的ec50值。igg9114l作为阴性对照同样进行梯度稀释检测其与ha7的亲和力,igg9114l为广谱中和抗体cr9114(参考文献:dreyfusc,laursenns,kwakst,zuijdgeestd,khayatr,ekiertdc,etal.highlyconservedprotectiveepitopesoninfluenzabviruses.science2012sep14;337(6100):1343-8)。
图4为本发明实施例中igg1f7l针对不同亚型及不同片段ha蛋白的亲和力检测结果(elisa法)。igg1f7l的孵育浓度为0.1μg/ml。h7为包被的influenzaah7n9(a/shanghai/2/2013)血凝素蛋白1μg/ml(acrobiosystems,货号ha9-v5227);h1为包被的influenzaah1n1(a/beijing/22808/2009)血凝素蛋白1μg/ml(北京义翘神州生物技术有限公司,货号40035-v08h-100),h5为包被的influenzaah5n1(a/commonmagpie/hongkong/2256/2006)血凝素蛋白1μg/ml(北京义翘神州生物技术有限公司,货号11700-v08h-100)。
图5为本发明实施例中igg1f7l针对h7n9禽流感病毒的体外中和活性检测结果。所用毒株为h7n9禽流感病毒深圳分离株sp17(a/shenzhen/sp17/2014(h7n9))。igg1f7l从50μg/ml开始3倍梯度稀释,与200tcid50的sp17病毒孵育后感染mdck细胞;抗体共设8个稀释梯度,每个梯度设6个复孔。感染72h后观察mdck细胞病理效应(cpe)并检测上清中病毒的血凝梯度以确定抗体的血凝抑制效果。
图6为本发明实施例中igg1f7l针对h7n9感染细胞的adcc活性检测结果。sp17病毒感染mdck细胞48h后,加入10μg/ml的抗体吸附细胞,去掉上清后加入健康人外周血淋巴细胞,培养4h后检测上清乳酸脱氢酶活性(od492)。每个抗体各设6个试验复孔,4个阴性对照复孔(未加抗体)和4个阳性对照复孔(加入裂解液),取平均值用于计算细胞杀伤活性:adcc%=100×(od492试验孔-od492阴性对照孔)/(od492阳性对照-od492阴性对照)。
具体实施方式
以下实施例用于说明本发明,但不用来限制本发明的范围。若未特别指明,实施例均按照常规实验条件,如sambrook等分子克隆实验手册(sambrookj&russelldw,molecularcloning:alaboratorymanual,2001),或按照制造厂商说明书建议的条件。
实施例1人源抗h7n9禽流感病毒中和性抗体1f7l及其制备方法
1、抗原的标记
从acrobiosystems(ha9-v5227)公司购买ha7蛋白,用pbs稀释成0.5-2mg/ml,然后利用thermal公司的生物素(ez-linktmsulfo-nhs-lc-biotin,21335)按照其试剂盒流程将ha7蛋白标记(分子数之比为蛋白:生物素=1:20-100),室温避光孵育0.5-2h,然后用10kd的离心半透柱(merckmillipore,ufc501096)以8000g离心4-6次,用无菌pbs补充,将多余的生物素分子去除干净,标记好的ha7蛋白分子将用于筛选ha7特异性的记忆性b细胞。
2、抗原特异性的记忆性b细胞分选及逆转录
分离h7n9感染患者恢复期外周血单个核细胞,用pbs洗涤一次,然后用含1%bsa的pbs(pbsa)重悬到106~108/ml,先加入生物素化的ha7蛋白使浓度达到10μg/ml,混匀后4℃孵育半小时,然后用pbs洗涤一次,再用同样体积的pbsa重悬pbmc,然后按照1:50的体积比加入购自biolegend公司的小鼠抗人cd19(apc-h7),小鼠抗人igg(apc),小鼠抗人igm(percp-cy5.5)和小鼠抗人cd27(fitc),同时按照1:200的体积比加入invitrogen公司的7aad(percp-cy5.5)和jacksonimmunolab的streptavidin-pe(016-110-084),混匀后4℃孵育半小时。pbs洗涤两遍,pbsa重悬后用于细胞分选。利用bdfacsariaiii分选患者pbmc中cd19+,igm-,igg+,cd27+,7aad-,ha7/pe+阳性的细胞,每孔1个细胞,用5μlresuspensionbuffer(superscripttmiiicellsdirectcdnasynthesissystem,invitrogen18080-300)收集细胞,-80℃保存或直接按照试剂盒的说明逆转录成cdna,-20℃保存。
3、抗体基因可变区的扩增
利用tiller于2008年发表在jimmunolmethods杂志上描述的引物和扩增方式扩增抗体基因重链可变区(vh)和轻链可变区(vk/vl)。vh,vk,vl分别进行两轮扩增,首先取3-5μlcdna作为模板,用抗体可变区基因先导序列特异性引物进行第一轮扩增,具体地,引物1-4和引物45、46扩增重链可变区(vh);引物17、18、19和引物51扩增kappa链可变区(vk),引物31-37和引物57扩增lambda链可变区(vl)。然后利用带有酶切位点的巢式引物,以5μl第一轮扩增的产物为模板进行第二轮扩增,具体地,引物5-16和引物48-50扩增重链可变区(vh);引物21-30和引物53-56扩增kappa链可变区(vk),引物38-43和引物58扩增lambda链可变区(vl)。pcr体系采用tiangen的superhifipcrmix(kt212)2×pcrmix。pcr程序为:95℃3min;95℃30sec,58-60℃30sec,72℃1min,50个循环;72℃10min;4℃10min。扩增产物利用1.5%的琼脂糖凝胶电泳(120v,40min),切取400bp左右的可变区片段。为了避免污染造成的假阳性,利用没有分选细胞的孔的cdna为模板作为第一轮pcr的阴性对照,利用没有加入第一轮pcr产物为模板的反应作为第二轮pcr的阴性对照。对于已经扩增出来的重链可变区和轻链可变区进行配对,只有同时扩增出重链可变区和轻链可变区的配对才进行下一步酶切和载体构建。
4、瞬时真核表达载体的构建
利用tiller于2008年发表在jimmunolmethods杂志上描述的抗体基因瞬间表达载体系统构建瞬间表达载体。该系统共含3个载体,分别用于表达igg1重链、kappa链和lambda链,分别命名为igh(accessionnumberdq407610)、igk(accessionnumberdq407610)和igl(accessionnumberfj517647)。按照该系统,只要将载体和带有酶切位点的可变区片段进行双酶切,连接,转化,即可得到能在真核细胞转染获得全长igg1的抗体。重链载体和片段采用agei和sali双酶切,kappa链载体和片段采用agei和bsiwi双酶切,lambda链载体和片段采用agei和xhoi双酶切。转化大肠杆菌后,挑取单克隆,利用5’absense引物(引物44)对载体插入序列进行测序,每个转化挑取3-5个细菌单克隆进行测序,以三个测序结果一致,插入序列和载体一起可以翻译成完整抗体片段,且插入序列不同于原始载体酶切前序列为载体构建成功。
抗体基因扩增所用的引物(5′-3′):
1l-vh1acaggtgcccactcccaggtgcag
2l-vh3aaggtgtccagtgtgargtgcag
3l-vh4/6cccagatgggtcctgtcccaggtgcag
4l-vh5caaggagtctgttccgaggtgcag
5ageivh1ctgcaaccggtgtacattcccaggtgcagctggtgcag
6ageivh1/5ctgcaaccggtgtacattccgaggtgcagctggtgcag
7ageivh3ctgcaaccggtgtacattctgaggtgcagctggtggag
8ageivh3–23ctgcaaccggtgtacattctgaggtgcagctgttggag
9ageivh4ctgcaaccggtgtacattcccaggtgcagctgcaggag
10ageivh4–34ctgcaaccggtgtacattcccaggtgcagctacagcagtg
11ageivh1–18ctgcaaccggtgtacattcccaggttcagctggtgcag
12ageivh1–24ctgcaaccggtgtacattcccaggtccagctggtacag
13ageivh3–33ctgcaaccggtgtacattctcaggtgcagctggtggag
14ageivh3–9ctgcaaccggtgtacattctgaagtgcagctggtggag
15ageivh4–39ctgcaaccggtgtacattcccagctgcagctgcaggag
16ageivh6–1ctgcaaccggtgtacattcccaggtacagctgcagcag
17lvκ1/2atgaggstcccygctcagctgctgg
18lvκ3ctcttcctcctgctactctggctcccag
19lvκ4atttctctgttgctctggatctctg
20panvκatgacccagwctccabycwccctg
21ageivκ1–5ctgcaaccggtgtacattctgacatccagatgacccagtc
22ageivκ1–9ttgtgctgcaaccggtgtacattcagacatccagttgacccagtct
23ageivκ1d–43ctgcaaccggtgtacattgtgccatccggatgacccagtc
24ageivκ2–24ctgcaaccggtgtacatggggatattgtgatgacccagac
25ageivκ2–28ctgcaaccggtgtacatggggatattgtgatgactcagtc
26ageivκ2–30ctgcaaccggtgtacatggggatgttgtgatgactcagtc
27agevκ3–11ttgtgctgcaaccggtgtacattcagaaattgtgttgacacagtc
28agevκ3–15ctgcaaccggtgtacattcagaaatagtgatgacgcagtc
29agevκ3–20ttgtgctgcaaccggtgtacattcagaaattgtgttgacgcagtct
30agevκ4–1ctgcaaccggtgtacattcggacatcgtgatgacccagtc
31lvλ1ggtcctgggcccagtctgtgctg
32lvλ2ggtcctgggcccagtctgccctg
33lvλ3gctctgtgacctcctatgagctg
34lvλ4/5ggtctctctcscagcytgtgctg
35lvλ6gttcttgggccaattttatgctg
36lvλ7ggtccaattcycaggctgtggtg
37lvλ8gagtggattctcagactgtggtg
38ageivλ1ctgctaccggttcctgggcccagtctgtgctgackcag
39ageivλ2ctgctaccggttcctgggcccagtctgccctgactcag
40ageivλ3ctgctaccggttctgtgacctcctatgagctgacwcag
41ageivλ4/5ctgctaccggttctctctcscagcytgtgctgactca
42ageivλ6ctgctaccggttcttgggccaattttatgctgactcag
43ageivλ7/8ctgctaccggttccaattcycagrctgtggtgacycag
44absensegcttcgttagaacgcggctac
45cγch1ggaaggtgtgcacgccgctggtc
46cμch1gggaattctcacaggagacga
47igg(internal)gttcggggaagtagtccttgac
48salljh1/2/4/5tgcgaagtcgacgctgaggagacggtgaccag
49salljh3tgcgaagtcgacgctgaagagacggtgaccattg
50salljh6tgcgaagtcgacgctgaggagacggtgaccgtg
51cκ543gtttctcgtagtctgctttgctca
52cκ494gtgctgtccttgctgtcctgct
53bsiwijκ1/4gccaccgtacgtttgatytccaccttggtc
54bsiwijκ2gccaccgtacgtttgatctccagcttggtc
55bsiwijκ3gccaccgtacgtttgatatccactttggtc
56bsiwijκ5gccaccgtacgtttaatctccagtcgtgtc
57cλcaccagtgtggccttgttggcttg
58xhoicλctcctcactcgagggygggaacagagtg
5、单克隆抗体的制备和纯化
将2ml含有构建成功载体的大肠杆菌接种到2yt培养基(trypton16g/l,yeastextract10g/l,nacl5g/l,ampicillin100μg/ml)200ml中,37℃220rpm培养16h。6000g离心15min收集菌体,按照thermal公司的质粒大量提取试剂盒(
为本发明基于单细胞分选和抗体基因扩增的全人源单克隆抗体技术平台示意图见图1。
实施例2单克隆抗体1f7l针对ha7蛋白的ec50测定
用elisa包被液稀释ha7蛋白至1μg/ml,然后每孔50μl包被elisa板(corning,3690),4℃过夜。pbst洗板,含5%脱脂牛奶的pbs封闭2h以上。将单克隆抗体从50μg/ml的起始浓度开始,用含5%脱脂牛奶的pbs进行4倍梯度稀释后,加入封闭好的elisa板。共设置10个梯度,两个重复,不加抗体的孔为阴性对照,加入梯度稀释的igg9114l(即cr9114,dreyfusc,laursenns,kwakst,zuijdgeestd,khayatr,ekiertdc,etal.highlyconservedprotectiveepitopesoninfluenzabviruses.science2012sep14;337(6100):1343-8)为抗体对照。羊抗人iggfc-hrp(1:10000,jacksonimmunolab,109-036-098)为二抗,100μltmb显色,100μl0.2m的硫酸终止,酶标仪读取od450值。取每个梯度的复孔的平均值,绘制od450-浓度曲线,按照graphpadprism的sigmoidaldoseresponse模型进行曲线拟合,并计算od450值为最高值一半(即1.6)时对应的抗体浓度,此即为抗体针对ha7的ec50值。
实施例3单克隆抗体1f7l针对h7n9(a/shenzhen/sp17/2014(h7n9))的中和活性测定
在96孔细胞培养板接种mdck细胞,培养至细胞密度约为70%-90%,pbs洗两遍后备用;将待检测的单克隆抗体在96孔微量滴定板上进行3倍的倍比稀释(100μg/ml起始),每个待检测抗体做4个孔的重复,8个梯度;根据病毒的tcid50滴度稀释病毒,使稀释后的病毒滴度为200tcid50/100μl;取60μl稀释后的病毒液与60μl稀释好的单克隆抗体样本混合,37℃培养箱中孵育2h使抗原抗体充分作用;然后取病毒血浆混合物100μl置洗好的96孔mdck细胞,37℃培养箱中感染1h,用加有tpck胰酶的mem培养基150μl置换病毒液,置37℃co2培养箱培养72h,观察cpe,红细胞凝集试验确认cpe结果。根据reed-muench法计算距离比,以能够抑制半数孔mdck细胞感染的浓度作为衡量单抗中和活性的ic50值。
实施例4单克隆抗体1f7l的抗体依赖细胞介导的细胞毒性杀伤(adcc)活性检测
在96孔细胞培养板接种mdck细胞,培养至细胞密度约为70%-90%,pbs洗两遍;用1000tcid50每孔的病毒量感染每孔mdck细胞,于含5%co2的37℃培养箱中感染1h,随后去掉上清,加170μl病毒生长液(mem培养基[thermal,11095-080)含1%青霉素-链霉素双抗(thermal,15140163)和0.1-0.5μg/mltpck胰酶(sigma,t1426)],置于37℃co2,浓度为5%的细胞培养箱中培养48h;pbs洗涤病毒感染的细胞两遍,每孔加入含10μg/ml待测抗体的mem培养基(thermal,11095-080),置于37℃,co2浓度为5%的细胞培养箱中培养30min,去掉上清,pbs洗涤一次,去掉上清,每孔加入200μl含5×105来自健康人pbmc的mem培养基,置于37℃,co2浓度为5%的细胞培养箱中培养4h。利用promega公司的非放射性细胞毒性试验检测试剂盒(g1780),按照操作流程检测培养上清中乳酸脱氢酶(ldh)活性。酶标仪读取od492值。未加抗体或加入非流感相关抗体的孔为阴性对照,加入试剂盒裂解液(未加pbmc)的孔为阳性对照。adcc%=100×(od492试验孔-od492阴性对照孔)/(od492阳性对照-od492阴性对照)。
本发明提供的抗体1f7l可以高效结合h7n9流感病毒血凝素蛋白ha7,其针对ha7蛋白亲和力的ec50<0.0115μg/ml(0.077nm)(图3)。具体地,本发明提供的抗体1f7l在与7型血凝素蛋白亲和力检测的elisa实验中,当抗体浓度低至0.0115μg/ml((0.077nm)时,其od450值仍可以达到最大值的一半。本发明提供的抗体可以用于h7n9流感病毒抗原检测试剂的开发。
本发明提供的抗体1f7l可以特异性地识别h7n9流感病毒血凝素蛋白ha7,且其识别位点位于ha7的颈部区域即ha2段。通过elisa实验检测igg1f7l与h1n1,h5n1和h7n9流感病毒的血凝素蛋白的亲和力,结果发现igg1f7l特异性地识别h7n9的血凝素蛋白,对h1n1和h5n1流感病毒的血凝素蛋白不具有结合能力,说明其为ha7亚型特异性抗体;同时其能结合全长ha7血凝素蛋白,但是却不能结合ha7的ha1段,暗示其识别位点位于ha7的ha2段(图4)。本发明提供的抗体可以用于h7n9流感病毒抗原检测试剂的开发。
本发明提供的抗体1f7l可以有效地中和h7n9流感病毒,防止h7n9流感病毒感染mdck细胞。具体地,在体外中和实验中,1.85μg/ml的1f7l抗体浓度可以实现对200tcid50的h7n9病毒对mdck细胞感染的50%的抑制,也即ic50=1.85μg/ml(图5)。本发明提供的抗体可以用于h7n9流感病毒感染患者的治疗。
本发明提供的的抗体可以有效地介导效应细胞(如nk细胞)对h7n9感染细胞的杀伤。具体地,10μg/ml的1f7l抗体介导的针对h7n9感染的mdck细胞的杀伤百分数可达27.13%(图6)。本发明提供的抗体可以用于h7n9流感病毒感染患者的治疗。
虽然,上文中已经用一般性说明及具体实施方案对本发明作了详尽的描述,但在本发明基础上,可以对之做一些修改或改进,这对本领域技术人员而言是显而易见的。因此,在不偏离本发明精神的基础上所做的这些修改或改进,均属于本发明要求保护的范围。
序列表
<110>深圳市第三人民医院
<120>人源抗h7n9禽流感病毒中和性抗体1f7l及其应用
<130>khp171113099.2
<160>6
<170>patentinversion3.3
<210>1
<211>330
<212>dna
<213>抗体1f7l轻链可变区dna序列
<400>1
agtctgtgctgactcagccaccctcagcgtctgggacccccgggcagagggtcaccatct60
cttgttctggaagcagctccaacatcggaagtaattatgtatactggtaccagcagctcc120
caggaacggcccccaaactcctcatctataggaataatcagcggccctcaggggtccctg180
accgattctctggctccaagtctggcacctcagcctccctggccatcagtgggctccggt240
ccgaggatgaggctgattattactgtgcagcatgggatgacagcctgagtggtgtggtat300
tcggcggagggacccagctgaccgtcctag330
<210>2
<211>110
<212>prt
<213>抗体1f7l轻链可变区
<400>2
glnservalleuthrglnproproseralaserglythrproglygln
151015
argvalthrilesercysserglyserserserasnileglyserasn
202530
tyrvaltyrtrptyrglnglnleuproglythralaprolysleuleu
354045
iletyrargasnasnglnargproserglyvalproaspargpheser
505560
glyserlysserglythrseralaserleualaileserglyleuarg
65707580
sergluaspglualaasptyrtyrcysalaalatrpaspaspserleu
859095
serglyvalvalpheglyglyglythrglnleuthrvalleu
100105110
<210>3
<211>216
<212>prt
<213>抗体1f7l轻链全长
<400>3
glnservalleuthrglnproproseralaserglythrproglygln
151015
argvalthrilesercysserglyserserserasnileglyserasn
202530
tyrvaltyrtrptyrglnglnleuproglythralaprolysleuleu
354045
iletyrargasnasnglnargproserglyvalproaspargpheser
505560
glyserlysserglythrseralaserleualaileserglyleuarg
65707580
sergluaspglualaasptyrtyrcysalaalatrpaspaspserleu
859095
serglyvalvalpheglyglyglythrglnleuthrvalleuglygln
100105110
prolysalaalaproservalthrleupheproprosersergluglu
115120125
leuglnalaasnlysalathrleuvalcysleuileseraspphetyr
130135140
proglyalavalthrvalalatrplysalaaspserserprovallys
145150155160
alaglyvalgluthrthrthrproserlysglnserasnasnlystyr
165170175
alaalasersertyrleuserleuthrprogluglntrplysserhis
180185190
argsertyrsercysglnvalthrhisgluglyserthrvalglulys
195200205
thrvalalaprothrglucysser
210215
<210>4
<211>385
<212>dna
<213>抗体1f7l重链可变区dna序列
<400>4
gaggtgcagctggtgcagtctgggggaggtgtggtacggcctggggggtccctgagactc60
tcctgtgcagcctctggattcagatttgatgattatggcatgagctgggtccgccaagct120
ccagggaagggactggagtgggtctctggtattaattggaatggtggtagcacagattat180
gcagactctgtgaagggccgattcaccatctccagagacaacgccaagaactccctgtat240
ctgcaaatgaacagtctgagagccgaggacacggccttgtattactgtgcgagagagggt300
ggcggggattttgtagtggtggtaggtggtaccacgtactactttgactactggggccag360
ggaaccctggtcaccgtctcctcag385
<210>5
<211>128
<212>prt
<213>抗体1f7l重链可变区
<400>5
gluvalglnleuvalglnserglyglyglyvalvalargproglygly
151015
serleuargleusercysalaalaserglypheargpheaspasptyr
202530
glymetsertrpvalargglnalaproglylysglyleuglutrpval
354045
serglyileasntrpasnglyglyserthrasptyralaaspserval
505560
lysglyargphethrileserargaspasnalalysasnserleutyr
65707580
leuglnmetasnserleuargalagluaspthralaleutyrtyrcys
859095
alaarggluglyglyglyaspphevalvalvalvalglyglythrthr
100105110
tyrtyrpheasptyrtrpglyglnglythrleuvalthrvalserser
115120125
<210>6
<211>458
<212>prt
<213>抗体1f7l重链全长
<400>6
gluvalglnleuvalglnserglyglyglyvalvalargproglygly
151015
serleuargleusercysalaalaserglypheargpheaspasptyr
202530
glymetsertrpvalargglnalaproglylysglyleuglutrpval
354045
serglyileasntrpasnglyglyserthrasptyralaaspserval
505560
lysglyargphethrileserargaspasnalalysasnserleutyr
65707580
leuglnmetasnserleuargalagluaspthralaleutyrtyrcys
859095
alaarggluglyglyglyaspphevalvalvalvalglyglythrthr
100105110
tyrtyrpheasptyrtrpglyglnglythrleuvalthrvalserser
115120125
alaserthrlysglyproservalpheproleualaproserserlys
130135140
serthrserglyglythralaalaleuglycysleuvallysasptyr
145150155160
pheprogluprovalthrvalsertrpasnserglyalaleuthrser
165170175
glyvalhisthrpheproalavalleuglnserserglyleutyrser
180185190
leuserservalvalthrvalproserserserleuglythrglnthr
195200205
tyrilecysasnvalasnhislysproserasnthrlysvalasplys
210215220
lysvalgluprolyssercysasplysthrhisthrcysproprocys
225230235240
proalaprogluleuleuglyglyproservalpheleuphepropro
245250255
lysprolysaspthrleumetileserargthrprogluvalthrcys
260265270
valvalvalaspvalserhisgluaspprogluvallyspheasntrp
275280285
tyrvalaspglyvalgluvalhisasnalalysthrlysproargglu
290295300
gluglntyrasnserthrtyrargvalvalservalleuthrvalleu
305310315320
hisglnasptrpleuasnglylysglutyrlyscyslysvalserasn
325330335
lysalaleuproalaproileglulysthrileserlysalalysgly
340345350
glnproarggluproglnvaltyrthrleuproproserargaspglu
355360365
leuthrlysasnglnvalserleuthrcysleuvallysglyphetyr
370375380
proseraspilealavalglutrpgluserasnglyglnprogluasn
385390395400
asntyrlysthrthrproprovalleuaspseraspglyserphephe
405410415
leutyrserlysleuthrvalasplysserargtrpglnglnglyasn
420425430
valphesercysservalmethisglualaleuhisasnhistyrthr
435440445
glnlysserleuserleuserproglylys
450455
- 还没有人留言评论。精彩留言会获得点赞!
- 人源抗H7N9禽流感病毒中和性抗体1F7L及其应用的制造方法与工艺
- 吲哚衍生物及其制备方法和其抗流感病毒作用与制造工艺
- 检测H5亚型禽流感病毒与鸡细小病毒的成套试剂与方法与制造工艺
- 检测禽流感病毒与鸡细小病毒的成套试剂与方法与制造工艺
- 一种流感病毒逆转录环介导等温扩增试剂盒及其应用的制造方法与工艺
- 用于快速检测流感病毒的狗肾细胞MDCK-NS1的离心小瓶及其应用方法与制造工艺
- 一种保护和稳定病毒RNA的采血管的制造方法与工艺
- H6亚型禽流感病毒荧光定量RT‑PCR检测方法与制造工艺
- 表达H7N9亚型禽流感病毒血凝素蛋白的重组火鸡疱疹病毒株rHOH及构建方法与制造工艺
- 一种提高禽流感疫苗毒株增殖滴度的方法与制造工艺