CYP71Z18基因及其编码蛋白与应用的制作方法
本发明属于农作物基因工程领域,具体地说,涉及一种CYP71Z18基因及其编码蛋白与应用。
背景技术:
天然抗菌物质植保素能响应病原菌侵染,提供抗病能力。水稻萜类植保素在早期研究中就被发现,这使水稻成为研究植保素形成的模式系统。玉米中首次发现的萜类植保素近年被报道,包括二萜类植保素kauralexins(Schmelz E A,Kaplan F,Huffaker A,et al.Identity,regulation,and activity of inducible diterpenoid phytoalexins in maize[J].Proceedings of the National Academy of Sciences of the United States of America,2011,108(13):5455-60.)和倍半萜类植保素zealexins(Huffaker A,Kaplan F,Vaughan M M,et al.Novel acidic sesquiterpenoids constitute a dominant class of pathogen-induced phytoalexins in maize[J].Plant Physiology,2011,156(4):2082-97.)。CYP71Z18参与zealexin生物合成,催化β–macrocarpene C15的甲基形成羧基(Mao H,Jiang L,Fei R,et al.Characterization of CYP71Z18 indicates a role in maize zealexin biosynthesis[J].Phytochemistry,2015,121:4-10.)。
二萜类化合物生物合成是在萜类化合物前体物质合成的基础上,通过异戊二烯基转移酶(Prenyl Transferases,PT)的作用产生20个碳原子的牻牛儿基牻牛儿基焦磷酸(Geranylgeranyl Diphosphate,GGPP),GGPP进一步在柯巴基焦磷酸合酶(Copalyl Diphosphate Synthase,CPS)和类贝壳杉烯合酶(Kaurene Synthase Like,KSL)的作用下形成二萜类化合物的基本骨架二萜烯,在萜烯的基础上经过羟基化和羧基化等形成二萜类化合物,其中细胞色素P450在修饰过程发挥着重要作用。
细胞色素P450(CYPs,P450)是一类亚铁血红素-硫醇盐蛋白的超家族,它以硫醇盐结合亚铁血红素为活性中心催化底物发生氧化反应,把一个氧原子加入到底物中,通常形成羟基化的产物。P450能够催化多种反应,除了羟基化还包括N-、O-和S-端的脱烷基化、环氧基化、脱氨基作用、脱硫、脱卤作用和过氧化反应等。P450催化时需要内质网膜上的细胞色素P450还原酶的存在,以提供氧化反应所需的电子。
水稻产生20多种二萜类植保素,几乎所有相关的萜类合酶都通过重组表达酶的生化方式被鉴定了功能(Xu M,Wilderman P R,Morrone D,et al.Functional characterization of the rice kaurene synthase-like gene family[J].Phytochemistry,2007,68(3):312-26.)。另一方面,P450催化特性各异,然而如何利用其多样的催化功能将水稻中多种二萜底物转化成新的产物还未见报道。
技术实现要素:
有鉴于此,本发明针对上述的问题,提供了一种CYP71Z18基因及其编码蛋白与应用,本发明通过大肠杆菌代谢工程,利用玉米倍半萜植保素生物合成中CYP71Z18编码蛋白对多种水稻二萜进行催化活性分析,发现CYP71Z18能够催化多种水稻二萜形成新的产物,其催化底物广泛。因此,可利用其催化特性在工业上生产多种新的二萜类化合物,以及改造水稻二萜类化合物的合成途径等。
为了解决上述技术问题,本发明公开了一种CYP71Z18基因,该基因的核苷酸序列如SEQ ID NO:1所示,该基因是与萜类化合物合成代谢有关的基因。该基因的DNA序列由1518个碱基组成。
本发明还公开了一种CYP71Z18基因编码的蛋白,具有序列表中SEQ ID NO.2的氨基酸残基序列以及具有SEQ IDNO.2的氨基酸残基序列相同活性的由SEQ IDNO.2衍生的蛋白质。该编码蛋白由505个氨基酸残基组成。
含有本发明基因CYP71Z18的原核表达载体、细胞系及宿主菌和使用该基因在调节和生产植物二萜类化合物及水稻育种中的应用也在本发明的保护范围之内。将SEQ ID No.1基因克隆到原核表达载体pCDFDuet的第二个多克隆位点(MCS2)限制性内切酶Nde I和BglⅡ位点间,pCDFDuet的MCS1含有拟南芥细胞色素P450还原酶基因AtCPR1(Mizutani M,Ohta D.Two isoforms of NADPH:cytochrome P450reductase in Arabidopsis thaliana.Gene structure,heterologous expression in insect cells,and differential regulation.[J].Plant Physiology,1998,116(1):357-67.),AtCPR1为CYP450提供所需的还原力,构建带有CYP71Z18基因的重组表达载体pCDF-AtCPR1-CYP71Z18。pACYCDuet的MCS1连有水稻同向柯巴基焦磷酸合酶(OsCPS4),MCS2连有大冷杉牻牛儿牻牛儿焦磷酸合酶(AgGGPS),载体简称pGGsC(Cyr A,Wilderman P R,Determan M,et al.A modular approach for facile biosynthesis of labdane-related diterpenes.[J].Journal of the American Chemical Society,2007,129(21):6684-5.)。pGGeC的MCS1连有拟南芥内部柯巴基焦磷酸合酶AtCPS,MCS2连有大冷杉牻牛儿牻牛儿焦磷酸合酶(AgGGPS)。pDEST14-OsKSL分别连有多种水稻二萜合酶基因(OsKSL4,OsKS7,OsKSL8,OsKSL10,OsKSL11)。将pGGsC/pGGeC、pDEST14-OsKSL与pCDF-AtCPR1-CYP71Z18共同转入大肠杆菌菌株C41(DE3)过表达宿主菌,采用异丙基-β-D-硫代吡喃半乳糖苷(IPTG)诱导表达,并在诱导表达时加入细胞色素P450反应所需辅因子。通过正己烷萃取菌液,GC-MS分析产物。
本发明还提供一种上述的CYP71Z18基因的编码蛋白在调节和生产植物二萜类化合物中的应用。
进一步地,该应用,包括以下步骤:
将pGGsC/pGGeC、pDEST14-OsKSL与pCDF-AtCPR1-CYP71Z18三种重组质粒各100-150ng同时加入40μL大肠杆菌C41(DE3)感受态细胞中,冰浴20min,42℃热激1min30s,冰上冷却2min后加入液体LB培养基200μL复苏2h,再涂布于同时含有氯霉素50mg/L、硫酸卡那霉素50mg/L和盐酸壮观霉素50mg/L的固体LB培养基平板上;37℃过夜培养,第二天挑选平均菌落,置于同时含有氯霉素50mg/L、硫酸卡那霉素50mg/L和盐酸壮观霉素50mg/L的5mL液体LB培养基中,37℃,过夜摇菌制备菌种;这些菌种为大肠杆菌C41(DE3)背景株系,且每个菌种分别含有三种质粒:
①pGGsC,pDEST14-OsKSL4,pCDF-AtCPR1-CYP71Z18;
②pGGsC,pDEST14-OsKSL8,pCDF-AtCPR1-CYP71Z18;
③pGGsC,pDEST14-OsKSL10,pCDF-AtCPR1-CYP71Z18;
④pGGsC,pDEST14-OsKSL11,pCDF-AtCPR1-CYP71Z18;
⑤pGGeC,pDEST14-OsKSL7,pCDF-AtCPR1-CYP71Z18;
⑥pGGeC,pDEST14-OsKSL10,pCDF-AtCPR1-CYP71Z18;
次日取1mL菌种置于50mL TB液体培养基,培养基中加入相应的抗生素,37℃,200rpm培养,菌液养至A600达到0.8-1.0,向其中加入1mM异丙基-β-D-硫代吡喃半乳糖苷诱导表达,并在诱导表达时加入细胞色素P450反应所需辅因子75mg/Lδ-氨基乙酰丙酸,1mM硫胺素,5mg/L核黄素;然后转至16℃,200rpm诱导培养72h;72h后用等体积的正己烷萃取萜类产物;所有的产物都经过叠氮甲烷甲基化后用于GC-MS检测羧酸类产物,制备得到羧酸类产物。
进一步地,液体LB培养基具体为:酵母提取物5g,胰蛋白胨10g,NaCl 10g,去离子水1L,用1mol/L的NaOH调PH值至7.0;高压灭菌,4℃保存;固体LB培养基在灭菌前加上15g/L的琼脂。
进一步地,TB液体培养基具体为:NaCl 5g,MgSO4.7H2O 2g,酵母提取物24g,水解酪蛋白12g,甘油20mL,去离子水900mL;10×TB缓冲液:KH2PO4 2.31g,K2HPO4 12.54g,去离子水100mL;高压灭菌,4℃保存,使用时加入10×TB缓冲液100mL。
本发明还公开了一种上述的CYP71Z18基因的编码蛋白在水稻育种中的应用。
与现有技术相比,本发明可以获得包括以下技术效果:
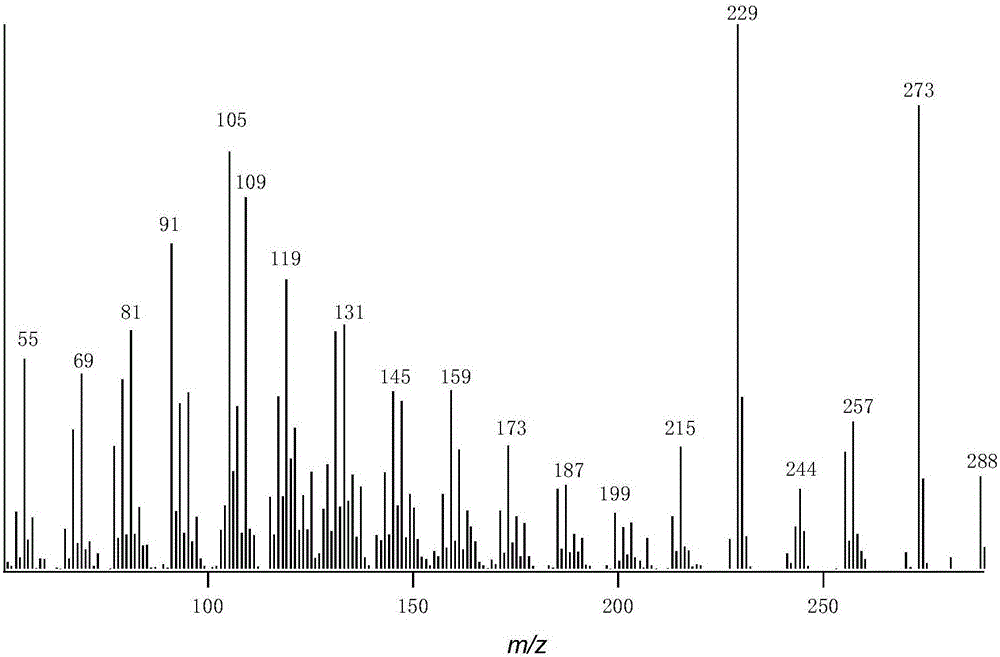
1)GC-MS分析结果表明:CYP71Z18蛋白酶催化syn-pimaradiene生成分子量为288(m/z)的新物质,通过核磁共振分析鉴定该产物为未被报道的新物质。CYP71Z18蛋白酶催化syn-stemarene生成甲基化后分子量为318(m/z)的新物质,其保留时间为19.778分钟;CYP71Z18蛋白酶催化syn-labdatriene生成分子量为304(m/z)和甲基化后分子量为316(m/z)的新物质,其保留时间分别为20.464分钟和19.527分钟;CYP71Z18蛋白酶催化syn-stemodene生成分子量为288(m/z)和甲基化后分子量为318(m/z)的新物质,其保留时间分别为20.361分钟和21.350分钟;CYP71Z18蛋白酶催化ent-cassadiene生成分子量为288(m/z)的新物质,其保留时间为21.935分钟;CYP71Z18蛋白酶催化ent-sandaracopimaradiene生成分子量为288(m/z)的新物质,其保留时间为19.414分钟。
2)研究结果表明,本发明涉及玉米倍半萜植保素生物合成中CYP71Z18能够催化多种水稻二萜形成新的产物,其催化底物十分广泛。
3)本发明有助于利用CYP71Z18催化特性在工业上生产多种新的二萜类化合物,以及改造水稻二萜类化合物的合成途径等具有重要理论及实践意义。
当然,实施本发明的任一产品并不一定需要同时达到以上所述的所有技术效果。
附图说明
此处所说明的附图用来提供对本发明的进一步理解,构成本发明的一部分,本发明的示意性实施例及其说明用于解释本发明,并不构成对本发明的不当限定。在附图中:
图1是构建的重组表达载体pCDF-AtCPR1-CYP71Z18;
图2是本发明CYP71Z18催化syn-pimaradiene反应产物GC-MS结构解析的质谱图,该产物分子量为288(m/z);
图3是本发明CYP71Z18催化syn-pimaradiene反应产物NMR结构解析;
图4是本发明CYP71Z18催化syn-stemarene反应产物GC-MS分析的总离子色谱图;
图5是本发明图4中色谱图中峰1对应的质谱图;
图6是本发明CYP71Z18催化syn-labdatriene反应产物GC-MS分析的总离子色谱图;
图7是本发明图6色谱图中峰2对应的质谱图;
图8是本发明图6色谱图中峰3对应的质谱图;
图9是本发明CYP71Z18催化syn-stemodene反应产物GC-MS分析的总离子色谱图;
图10是本发明图9色谱图中峰4对应的质谱图;
图11是本发明图9色谱图中峰5对应的质谱图;
图12是本发明CYP71Z18催化ent-cassadiene反应产物GC-MS分析的总离子色谱图;
图13是本发明图12色谱图中峰6对应的质谱图;
图14本发明CYP71Z18催化ent-sandaracopimaradiene反应产物GC-MS分析的总离子色谱图;
图15是本发明图14色谱图中峰7对应的质谱图。
具体实施方式
下列使用的所有化学药品,除正己烷和乙酸乙酯是色谱纯外,其余都是分析纯。GC-MS分析使用的是安捷伦6890-5973型号,HP-5GC柱,四倍质谱检测仪,采用电子轰击模式。样品进样1μL,以不分流模式,初始温度70℃,炉温保持70℃2min,以10℃/min的速度升至300℃,保持300℃2min。MS数据收集从保留时间10min开始直至程序结束,质荷比显示从50至600。
玉米B73株系的种子用20%次氯酸钠消毒20min,并置于水-琼脂培养基上发芽。7天后,无菌幼苗用于真菌侵染和RNA提取。禾谷镰刀菌在土豆葡萄糖琼脂培养基上生长,然后转移至产孢培养基(羧甲基纤维素钠7.5g/L,磷酸二氢钾0.5g/L,酵母提取物0.5g/L,硝酸铵0.5g/L,硫酸镁0.25g/L),5天后收集孢子。划伤叶片后将106mL-1孢子溶于0.1%吐温20,对叶片进行孢子接种。
实施例1:玉米中CYP71Z18基因的克隆及修饰
CYP71Z18在NCBI基因库中的编号是NM_001147894,根据其编码区核苷酸序列,设计引物从玉米中克隆全长序列;引物如下:
CYP71Z18F:5’-ATGGAGGACAAGGTGCTCATCGC-3’;其核苷酸序列如SEQ ID NO:3所示;
CYP71Z18R:5’-TTAGGCATTAACCGGAGCAATGTGG-3’;其核苷酸序列如SEQ ID NO:4所示。
玉米叶片被禾谷镰刀菌孢子接种24h后提取RNA,用反转录酶M-MLV(Takara)反转录得到cDNA。以cDNA为模板,CYP71Z18F和CYP71Z18R为引物,用高保真酶PrimeSTAR HS DNA polymerase(Takara)扩增CYP71Z18编码区序列。PCR程序:95℃5min预变性,35个循环:98℃10s,55℃5s,72℃1min30s,72℃5min,12℃∞。扩增完成后,将PCR产物进行1%的琼脂糖凝胶电泳检测并利用胶回收试剂盒(OMEGA)切胶回收目的片段。将胶回收得到的1518bp的片段克隆到pGM-T(Tiangen,Beijing)载体中,经测序确认为数据库中公布的CYP71Z18编码区序列。
为了顺利在大肠杆菌中表达CYP71Z18蛋白,参照前人研究方法(Swaminathan S,Morrone D,Wang Q,et al.CYP76M7 Is an ent-Cassadiene C11-Hydroxylase Defining a Second Multifunctional Diterpenoid Biosynthetic Gene Cluster in Rice[J].Plant Cell,2009,21(10):3315-25.),将CYP71Z18的N端前32个氨基酸替换成10个富赖氨酸序列MAKKTSSKGK。设计N端修饰引物:
CYP71Z18-d32 ES-F1:CCTCCAGGGCCATGGACGCT;其核苷酸序列如SEQ ID NO:5所示;
CYP71Z18-d32 ES-F2:AACATATGGCGAAAAAAACCAGCAGCAAAGGTAAACCTCCAGGGCCATGG;其核苷酸序列如SEQ ID NO:6所示;
CYP71Z18-ES-R:AAAGATCTTTAGGCATTAACCGGAGCAATGTGG;其核苷酸序列如SEQ ID NO:7所示;
其中,下划线所示序列为酶切识别位点。
经过两步PCR,第一步PCR以pGM-T/CYP71Z18为模板,CYP71Z18-d32 ES-F1和CYP71Z18-ES-R为引物,用高保真酶PrimeSTAR HS DNA polymerase扩增得到片段,经胶回收试剂盒回收长为1422bp的目的片段。第二步PCR以上步回收得到的1422bp目的片段为模板,CYP71Z18-d32 ES-F2和CYP71Z18-ES-R为引物,用高保真酶PrimeSTAR HS DNA polymerase扩增得到修饰后的片段长为1468bp,切胶回收片段。
修饰后的基因被亚克隆到pCDF-Duet(Novagen)载体的第二个多克隆位点(MCS)限制性内切酶Nde I和BglⅡ位点间,pCDF-Duet第一个多克隆位点的限制性内切酶Nco I和Sal I位点间含有来自拟南芥中的依赖NADPH的P450还原酶(AtCPR1)。将上步回收得到修饰后的片段和pCDF-AtCPR1分别用Nde I和BglⅡ(Takara)双酶切并酶切液回收,利用T4DNA Ligase(Takara)连接,最终构建的重组表达载体结构如图1所示。
实施例2:CYP71Z18基因的编码蛋白与多种水稻二萜合酶的原核重组表达
CYP71Z18基因的编码蛋白在大肠杆菌C41(DE3)(Lucigen)过表达株系中表达,将AgGGPS、OsCPS4/AtCPS、OsKSL、AtCPR1和CYP71Z18共同转入大肠杆菌,AgGGPS、OsCPS4/AtCPS用以提供前体GGPP和CPP,AtCPR1提供CYP71Z18所需的还原力。OsKSL提供多种二萜底物,其中OsKSL4催化同向柯巴基焦磷酸(syn-CPP)产生syn-pimaradiene;OsKSL8催化syn-CPP产生syn-stemarene;OsKSL10催化syn-CPP产生syn-labdatriene;OsKSL11催化syn-CPP产生syn-stemodene。OsKSL7催化内部柯巴基焦磷酸(ent-CPP)产生ent-cassadiene;OsKSL10催化ent-CPP产生ent-sandaracopimaradiene。
具体实施方式如下:将pGGsC/pGGeC、pDEST14-OsKSL与pCDF-AtCPR1-CYP71Z18三种重组质粒各100-150ng同时加入40μL大肠杆菌C41(DE3)感受态细胞中,冰浴20min,42℃热激1min30s,冰上冷却2min后加入液体LB培养基(酵母提取物5g,胰蛋白胨10g,NaCl 10g,去离子水1L,用1mol/L的NaOH调PH值至7.0。高压灭菌,4℃保存。固体LB培养基在灭菌前加上15g/L的琼脂即可。)200μL复苏2h,再涂布于同时含有三种质粒抗性(氯霉素50mg/L,硫酸卡那霉素50mg/L,盐酸壮观霉素50mg/L)的固体LB培养基平板上。37℃过夜培养,第二天挑选平均菌落,置于同时含有三种质粒抗性的5mL液体LB培养基中,37℃,过夜摇菌制备得到6中菌种。这些菌种为大肠杆菌C41(DE3)背景株系,且每个菌种分别含有三种质粒:
①pGGsC,pDEST14-OsKSL4,pCDF-AtCPR1-CYP71Z18;
②pGGsC,pDEST14-OsKSL8,pCDF-AtCPR1-CYP71Z18;
③pGGsC,pDEST14-OsKSL10,pCDF-AtCPR1-CYP71Z18;
④pGGsC,pDEST14-OsKSL11,pCDF-AtCPR1-CYP71Z18;
⑤pGGeC,pDEST14-OsKSL7,pCDF-AtCPR1-CYP71Z18;
⑥pGGeC,pDEST14-OsKSL10,pCDF-AtCPR1-CYP71Z18。
次日取1mL菌种置于50mL TB液体培养基(NaCl 5g,MgSO4.7H2O 2g,酵母提取物24g,水解酪蛋白12g,甘油20mL,去离子水900mL;10×TB缓冲液:KH2PO42.31g,K2HPO412.54g,去离子水100mL;高压灭菌,4℃保存,使用时加入10×TB缓冲液100mL。),培养基中加入相应的抗生素,37℃,200rpm培养,菌液养至A600达到0.8-1.0,向其中加入1mM异丙基-β-D-硫代吡喃半乳糖苷(IPTG)诱导表达,并在诱导表达时加入细胞色素P450反应所需辅因子75mg/Lδ-氨基乙酰丙酸,1mM硫胺素,5mg/L核黄素。然后转至16℃,200rpm诱导培养72h。72h后用等体积的正己烷萃取萜类产物。所有的产物都经过叠氮甲烷甲基化后用于GC-MS检测羧酸类产物。为了得到足够的CYP71Z18催化syn-pimaradiene的产物而进行核磁共振(NMR)分析,培养液的体积增加至6L,并用正己烷萃取产物,通过旋转蒸发仪浓缩。浓缩蒸干后的产物用5mL正己烷重悬,通过硅胶柱层析法分离。使用正己烷,二氯甲烷混合溶剂(体积比为10:1)为洗脱液。最终得到CYP71Z18催化syn-pimaradiene的主要产物约5mg,其纯度为95%,并用于核磁共振分析。该方法应用于工业发酵,即扩大发酵菌液的体积,则可产生大量的如上所述多种新的二萜类物质,实现工业大量合成二萜类新物质的愿景。
结果如图2~图15,在大肠杆菌代谢工程体系中,CYP71Z18催化syn-pimaradiene形成生成分子量为288(m/z)的新物质,通过核磁共振分析鉴定该产物为syn-pimaradiene的C15和C16位碳原子被环氧化的产物,是一种未被报道的新物质。CYP71Z18蛋白酶催化syn-stemarene生成甲基化后分子量为318(m/z)的新物质,其保留时间为19.778分钟;CYP71Z18蛋白酶催化syn-labdatriene生成分子量为304(m/z)和甲基化后分子量为316(m/z)的新物质,其保留时间分别为20.464分钟和19.527分钟;CYP71Z18蛋白酶催化syn-stemodene生成分子量为288(m/z)和甲基化后分子量为318(m/z)的新物质,其保留时间分别为20.361分钟和21.350分钟。CYP71Z18蛋白酶催化ent-cassadiene生成分子量为288(m/z)的新物质,其保留时间为21.935分钟;CYP71Z18蛋白酶催化ent-sandaracopimaradiene生成分子量为288(m/z)的新物质,其保留时间为19.414分钟。研究结果表明,本发明涉及玉米倍半萜植保素生物合成中CYP71Z18能够催化多种水稻二萜形成新的产物,其催化底物十分广泛。
鉴于此,本发明通过转基因技术将玉米中的CYP71Z18在水稻中过表达,则CYP71Z18能够对水稻体内多种参与水稻二萜类植保素合成相关的二萜烯进行氧化,从而产生新的二萜类物质,这可能改变水稻体内二萜植保素的生物合成途径,抑或产生新的二萜植保素,改变水稻的抗逆性。
上述说明示出并描述了发明的若干优选实施例,但如前所述,应当理解发明并非局限于本文所披露的形式,不应看作是对其他实施例的排除,而可用于各种其他组合、修改和环境,并能够在本文所述发明构想范围内,通过上述教导或相关领域的技术或知识进行改动。而本领域人员所进行的改动和变化不脱离发明的精神和范围,则都应在发明所附权利要求的保护范围内。
- 还没有人留言评论。精彩留言会获得点赞!
- 蝎神经毒素AaIT及其编码基因与应用的制造方法与工艺
- 腈水解酶突变体及其编码基因和应用的制造方法与工艺
- 植物抗旱相关蛋白AsTMK1及其编码基因和应用的制造方法与工艺
- 植物抗旱相关蛋白EtSnRK2.2及其编码基因和应用的制造方法与工艺
- OsSAPK7蛋白及其编码基因在提高水稻白叶枯病抗性中的应用的制造方法与工艺
- 一种抗除草剂抗虫融合基因、编码蛋白及应用的制造方法与工艺
- 毛果杨PtrZFP103基因及其编码蛋白和应用的制造方法与工艺
- 大豆bHLH转录因子基因GmFER及其编码蛋白和应用的制造方法与工艺
- 一种花狭口蛙的基因及其编码的多肽和该多肽的用途的制造方法与工艺
- 一种泽陆蛙基因及其编码的多肽和该多肽的用途的制造方法与工艺
- 降低种子重量的白桦AP2基因及其编码蛋白的制造方法与工艺
- 可编码ArtRD蛋白基因及其蛋白和应用的制造方法与工艺
- 蝎神经毒素AaIT及其编码基因与应用的制造方法与工艺
- 腈水解酶突变体及其编码基因和应用的制造方法与工艺
- 植物抗旱相关蛋白AsTMK1及其编码基因和应用的制造方法与工艺
- 植物抗旱相关蛋白EtSnRK2.2及其编码基因和应用的制造方法与工艺
- OsSAPK7蛋白及其编码基因在提高水稻白叶枯病抗性中的应用的制造方法与工艺
- 一种抗除草剂抗虫融合基因、编码蛋白及应用的制造方法与工艺
- 毛果杨PtrZFP103基因及其编码蛋白和应用的制造方法与工艺
- 大豆bHLH转录因子基因GmFER及其编码蛋白和应用的制造方法与工艺
- 腈水解酶突变体及其编码基因和应用的制造方法与工艺
- 植物抗旱相关蛋白AsTMK1及其编码基因和应用的制造方法与工艺
- 植物抗旱相关蛋白EtSnRK2.2及其编码基因和应用的制造方法与工艺
- OsSAPK7蛋白及其编码基因在提高水稻白叶枯病抗性中的应用的制造方法与工艺
- 一种抗除草剂抗虫融合基因、编码蛋白及应用的制造方法与工艺
- 毛果杨PtrZFP103基因及其编码蛋白和应用的制造方法与工艺
- 大豆bHLH转录因子基因GmFER及其编码蛋白和应用的制造方法与工艺
- 一种花狭口蛙的基因及其编码的多肽和该多肽的用途的制造方法与工艺
- 一种泽陆蛙基因及其编码的多肽和该多肽的用途的制造方法与工艺
- 一种水稻MIT1基因、其编码蛋白及应用的制造方法与工艺
- 植物抗旱相关蛋白EtSnRK2.2及其编码基因和应用的制造方法与工艺
- OsSAPK7蛋白及其编码基因在提高水稻白叶枯病抗性中的应用的制造方法与工艺
- 一种抗除草剂抗虫融合基因、编码蛋白及应用的制造方法与工艺
- 毛果杨PtrZFP103基因及其编码蛋白和应用的制造方法与工艺
- 大豆bHLH转录因子基因GmFER及其编码蛋白和应用的制造方法与工艺
- 一种水稻MIT1基因、其编码蛋白及应用的制造方法与工艺
- 蛋白StTrxF、编码基因及其在提高植物淀粉含量中的应用的制造方法与工艺
- 蛋白质及其编码基因在调控植物抗黄萎病性中的应用的制造方法与工艺
- 甘薯耐逆相关蛋白IbFBA2、编码基因、重组载体与应用的制造方法与工艺
- 植物抗旱相关蛋白ZmNAC111及其编码基因与应用的制造方法与工艺
- 植物抗旱相关蛋白EtSnRK2.2及其编码基因和应用的制造方法与工艺
- OsSAPK7蛋白及其编码基因在提高水稻白叶枯病抗性中的应用的制造方法与工艺
- 一种抗除草剂抗虫融合基因、编码蛋白及应用的制造方法与工艺
- 毛果杨PtrZFP103基因及其编码蛋白和应用的制造方法与工艺
- 大豆bHLH转录因子基因GmFER及其编码蛋白和应用的制造方法与工艺
- 一种水稻MIT1基因、其编码蛋白及应用的制造方法与工艺
- 蛋白StTrxF、编码基因及其在提高植物淀粉含量中的应用的制造方法与工艺
- 植物抗旱相关蛋白ZmNAC111及其编码基因与应用的制造方法与工艺
- 核盘菌凝集素SSL‑6蛋白及其编码基因与应用的制造方法与工艺
- 植物抗病毒相关蛋白GmSN1及其编码基因和应用的制造方法与工艺
- 植物抗旱相关蛋白EtSnRK2.2及其编码基因和应用的制造方法与工艺
- OsSAPK7蛋白及其编码基因在提高水稻白叶枯病抗性中的应用的制造方法与工艺
- 一种抗除草剂抗虫融合基因、编码蛋白及应用的制造方法与工艺
- 毛果杨PtrZFP103基因及其编码蛋白和应用的制造方法与工艺
- 大豆bHLH转录因子基因GmFER及其编码蛋白和应用的制造方法与工艺
- 一种水稻MIT1基因、其编码蛋白及应用的制造方法与工艺
- 蛋白StTrxF、编码基因及其在提高植物淀粉含量中的应用的制造方法与工艺
- 蛋白质及其编码基因在调控植物抗黄萎病性中的应用的制造方法与工艺
- 植物抗旱相关蛋白ZmNAC111及其编码基因与应用的制造方法与工艺
- 核盘菌凝集素SSL‑6蛋白及其编码基因与应用的制造方法与工艺