一种热稳定性提高的脂肪氧合酶突变体及其构建方法与流程
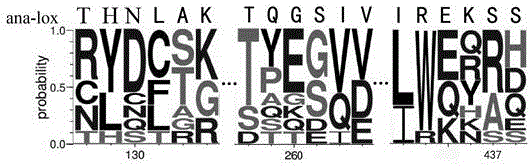
本发明涉及生物技术,具体涉及一种热稳定性提高的脂肪氧合酶突变体及其构建方法。
背景技术:
:LOX(Lipoxygenase,LOX)是非血红素、非硫铁过氧化酶,它能催化具有顺,顺-1,4-戊二烯结构的多元不饱和脂肪酸及其酯类发生空间及位置特异性的双加氧反应,生成相应的氢过氧化物。在植物中其天然底物主要是亚油酸和亚麻酸,在动物体内其天然底物主要是花生四烯酸。LOX作为无毒、无害、天然的添加剂在食品、医药、化工等领域有着重要的应用价值。在面粉的加工过程中它能够将面粉中的多不饱和脂肪酸催化氧化成氢过氧化物,这些氢过氧化物能够起到提高面粉白度,增强面筋蛋白强度的作用。另外,在污水处理与环境保护方面,可以用于染料的降解。然而脂肪氧合酶热稳定性差的缺陷限制了其在工业上的应用。因此,通过分子改造来提高脂肪氧合酶热稳定性以期获得酶学性质更适用工业应用的突变酶是目前的研究热点。技术实现要素:本发明目的是为了提供一种酶活与热稳定性同时提高的脂肪氧合酶突变体及其构建方法,提高了脂肪氧合酶突变体的热稳定性,扩展了应用范围。本发明的技术方案如下:一种热稳定性提高的脂肪氧合酶突变体,该脂肪氧合酶突变体具有如SEQIDNO.2、SEQIDNO.3、SEQIDNO.4、SEQIDNO.5或SEQIDNO.6中任一种所示的氨基酸序列。一种以上所述的热稳定性提高的脂肪氧合酶突变体中PCR反应的简并引物,其特征在于,该简并引物具体如下:N130-f:TTACTCACNNKCTGGCAAAATATGACATCAAG;N130-r:TTTGCCAGMNNGTGAGTAAGCTCATGTG;S437-f:GGAAAAATCANNKATATTGGAACCAGGACTTC;S437-r:TCCAATATMNNTGATTTTTCCCGAATGAGCG;G260-f:GCTAACGCAGNNKTCTATTGTTGATGTAA;G260-r:AACAATAGAMNNCTGCGTTAGCATG。一种以上所述的热稳定性提高的脂肪氧合酶突变体的构建方法,包括以下步骤:(1)鱼腥藻脂肪氧合酶突变位点确定:将鱼腥藻脂肪氧合酶自N端起第130位天冬酰胺、260位甘氨酸和437位丝氨酸作为饱和突变位点;(2)鱼腥藻脂肪氧合酶突变体库的建立及突变体筛选:以重组质粒为模板,在第130位天冬酰胺、260位甘氨酸和437位丝氨酸相对应的核苷酸处利用NNK代替原有密码子设计简并引物,该简并引物如以上所述,进行全质粒PCR反应,构建3个定点饱和突变库,DpnI消化模板,纯化后产物直接转化大肠杆菌,将转化后的大肠杆菌涂布于含有100μg/ml氨苄抗性LB平板上,37℃下培养16-24小时,得到3个饱和突变库;(3)取3个饱和突变库的菌落接种于含有氨苄LB培养基的96孔板中,于37℃过夜培养,得到3个突变体库的母版,取母版接种于含有氨苄LB培养基的96孔板中,37℃下培养3h,再加入IPTG低温诱导16h;然后裂解菌体,得到粗酶液,将粗酶液在50℃条件下孵育5min,冷却,测定残余活力,得到阳性突变体;;(4)将步骤(3)得到的阳性突变体接种于含有氨苄青霉素的LB液体培养基中,37℃,180rpm培养至OD600为0.6-0.8时,加IPTG低温诱导,离心收集菌体,用磷酸盐缓冲液重悬菌体,超声波破碎菌体,将上清液过Ni-NTA亲和柱纯化,透析去除咪唑即得到纯的脂肪氧合酶突变体。进一步地,所述的热稳定性提高的脂肪氧合酶突变体的构建方法,步骤(2)中大肠杆菌为大肠杆菌E.coliBL21(DE3)。进一步地,所述的热稳定性提高的脂肪氧合酶突变体的构建方法,其特征在于,步骤(2)中重组质粒为重组质粒pET32a-Ana-LOX。进一步地,所述的热稳定性提高的脂肪氧合酶突变体的构建方法,步骤(2)中全质粒PCR反应的反应条件为:96℃5min;然后98℃10s,55℃5s,72℃90s,共25个循环;最后72℃5min。进一步地,所述的热稳定性提高的脂肪氧合酶突变体的构建方法,步骤(3)中裂解菌体采用冻融破碎的方法进行,具体为-70℃冷冻2小时,室温融化1小时,反复冻融。以上所述的热稳定性提高的脂肪氧合酶突变体在食品、制药、造纸或污水处理中的应用。与野生型鱼腥藻脂肪氧合酶相比,本发明的脂肪氧合酶突变体的热稳定性获得了提高,以各自的比活力及半衰期t1/2来表示酶活与热稳定性的提高,本发明的脂肪氧合酶突变体热稳定性提高如表1所示。表1本发明首先利用已知的结构与生物信息学信息,找出可能的突变位点,在对这些位点进行饱和突变,构建突变文库;然后在96孔板上进行诱导表达;最后对文库进行筛选得到性质改良的突变体。再以此突变株作为模板,组合其余位点的突变,重复上述过程。本发明运用半理性设计的方法,对鱼腥藻脂肪氧合酶基因进行多轮定点饱和突变,获取脂肪氧合酶突变体,这些突变体包含氨基酸突变G260A、S437T、N130D及组合突变体N130D/S437Y、N130D/G260Q。以各自的比活力与半衰期t1/2来表示,脂肪氧合酶突变体的活力与热稳定性得到了提高,具有较高的实际应用价值和广阔的市场前景。附图说明图1为本发明实施例1中脂肪氧合酶家族蛋白的系统发育树图;图2为本发明实施例1中多重序列比对选择突变位点图;图3为本发明实施例1中脂肪氧合酶突变体热失活动力学图;图4为本发明实施例1中脂肪氧合酶突变体最适反应温度图。具体实施方式实施例11.饱和突变位点的确定:对脂肪氧合酶家族进行系统发育分析,选择与鱼腥藻脂肪氧合酶进化关系紧密的同一家族的氨基酸序列进行多重序列比对,结合结构信息确定下列位点为饱和突变位点:自N端起第130位天冬酰胺、260位甘氨酸、437位丝氨酸。其中鱼腥藻脂肪氧合酶氨基酸序列如SEQIDNO.1所示。突变位点选择过程具体为在pfam数据库中选择脂肪氧合酶家族序列,将其通道blastp与Ana-LOX进行比对,筛选出相似度30%-90%的序列,并构建系统发育树,最终选择与Ana-LOX亲缘关系最近的序列(图1)。将这些序列进行多重序列比对,比对结果与Ana-LOX进行比较,筛选出潜在的突变位点(图2)。Ana-LOX序列提交phyre2网站构建三维模型,将潜在的突变位点放在模型中结合作用力分析,最终确定自N端起第130位天冬酰胺、260位甘氨酸、437位丝氨酸为突变位点。其中图1为脂肪氧合酶家族蛋白的系统发育树图,通过构建Ana-LOX与其蛋白质家族的系统发育树筛选出8条序列与其进化关系紧密,选择这些序列进行多重序列比对。图2为多重序列比对选择突变位点图,可以发现Ana-LOX氨基酸序列与其多重序列比对存在大量的位点不一致,例如130号位Ana-LOX为天冬酰胺而该处频率最高的氨基酸为天冬氨酸,可以将其视为潜在的突变位点,可以做为进一步分析的候选。2.鱼腥藻脂肪氧合酶突变体库的建立及突变体筛选用下述方法筛选具有较好热稳定性的鱼腥藻脂肪氧合酶的突变体,具体过程包括以下步骤:1)根据突变位点构建饱和突变库并导入宿主细胞以重组质粒pET32a-Ana-LOX为模板,根据以上确定的突变位点,分别在第130位天冬酰胺、260位甘氨酸、437位丝氨酸相对应的核苷酸处利用NNK代替原有密码子,设计简并引物:N130-fTTACTCACNNKCTGGCAAAATATGACATCAAG(SEQIDNO.7)N130-rTTTGCCAGMNNGTGAGTAAGCTCATGTG(SEQIDNO.8)S437-fGGAAAAATCANNKATATTGGAACCAGGACTTC(SEQIDNO.9)S437-rTCCAATATMNNTGATTTTTCCCGAATGAGCG(SEQIDNO.10)G260-fGCTAACGCAGNNKTCTATTGTTGATGTAA(SEQIDNO.11)G260-rAACAATAGAMNNCTGCGTTAGCATG(SEQIDNO.12)以上简并引物中,N代表ATCG中的任一种;M代表A和C中的任一种;K代表G和T中的任一种。然后进行全质粒PCR反应,使用TaKaRa公司的PrimeSTARDNA聚合酶,PCR反应条件为:96℃5min;然后98℃10s,55℃5s,72℃90s,共25个循环;最后72℃5min。构建3个定点饱和突变库,DpnI消化模板,纯化后产物直接转化大肠杆菌E.coliBL21(DE3),将转化后的大肠杆菌涂布于含有100μg/ml氨苄抗性LB平板上,37℃下培养16-24小时,得到3个饱和突变库;2)从饱和突变体库中筛选热稳定性提高的突变体取饱和突变库中的菌落接种于含有100μg/ml氨苄LB培养基的96孔板中,于37℃过夜培养,得到3个突变体库的母版;从培养好的母版中取20μl接种至含有100μg/ml氨苄LB培养基的96孔板中,37℃下培养3h,在加入终浓度为100μg/ml的IPTG,16℃诱导16h;利用冻融破碎的方法裂解菌体,得到粗酶液;将所得到的粗酶液在50℃下孵育5min,在冰上冷却,测定每孔的残余活力;以野生型为对照,将残余活力高于野生型的作为阳性突变体。将筛选到的阳性突变体工程菌接种于含有100μg/mL氨苄青霉素的LB液体培养基中,37℃,180rpm培养至OD600为0.6-0.8时,加IPTG至100μg/mL,低温16℃诱导16h,离心收集菌体,用磷酸盐缓冲液重悬菌体,超声波破碎菌体,并将上清液过Ni-NTA亲和柱纯化,透析去除咪唑即可得到纯的脂肪氧合酶突变体;纯酶液测定50℃下的半衰期,确定每个突变体库中热稳定性最高的突变体,进行测序,至此完成了第一轮筛选,得到三个热稳定性与酶活提高的突变体,分别为N130D,G260A和S437T,分别对应SEQIDNO.2,3和4。以N130D的基因作为模板,进行突变体组合:针对260位甘氨酸、437位丝氨酸位点设计饱和突变库,并参考上述步骤进行新一轮的筛选;经过组合突变,获得了两株热稳定性与酶活力显著提高的突变株,经测序确定两株组合突变体分别为:N130D/G260Q,N130D/S437Y,分别对应SEQIDNO.5和6。为了测定脂肪氧合酶突变体的比活力与半衰期,对酶进行分离纯化。将超声破碎得到的粗酶液通过Ni2+离子亲和柱进行纯化,透析过夜即可得到纯的脂肪氧合酶突变体纯酶。脂肪氧合酶在50℃下半衰期的测定方法为:50℃下孵育酶液,在不同处理时间取样,测定残余活力百分比。以残余活力百分比的ln值对时间t(min)作图,直线的斜率为失活常数kd,由t1/2=ln2/kd得到脂肪氧合酶在该温度下的半衰期。图3为脂肪氧合酶突变体热失活动力学图在单点突变中N130D突变对酶热稳定性的改良最为显著。在N130D的基础上,组合突变体N130D/S437Y、N130D/G260Q的热稳定性也有了进一步提升。图4为脂肪氧合酶突变体最适反应温度图,突变体对温度的适应范围相比野生型变得更宽,在常温下25℃野生型活力低于50%,而突变体均超过60%,可以推测突变体活化能降低。野生型对稳定较为敏感,当温度超过最适反应温度35℃,酶活急剧下降,而突变体S437T与N130D/S437Y的最适反应温度可以达到45℃。组合突变体N130D/S437Y、N130D/G260Q即使达到55℃的高温依然可以保持70%以上的活力,而此时野生型已经失活。具有Ser437位置的突变体S437T与N130D/S437Y最适反应温度均提高了10℃,表明Ser437位置时影响最适反应温度的关键氨基酸。脂肪氧合酶目前在多个领域都有着广泛的应用,如在食品领域可使面粉增白,在造纸、染料、涂料、洗涤剂、污水处理、聚氯乙烯增塑剂的工业化生产,还可作为药物合成的中间体,所以脂肪氧合酶也被用于其他工业催化用途,如利用脂肪氧合酶对脂肪酸或脂肪酸酯进行选择性催化,形成特定的氢过氧化脂肪酸及其酯的衍生物,再将之环氧化,生成环氧化油,则可减小石油用量;氢过氧化油脂经还原得到羟基油脂,经过氧化物酶裂解得到不饱和酸及醛,再进一步反应可以得到二元酸,这些物质在食品、化工、生物技术等领域都有重要用途。因此,本发明提供的脂肪氧合酶突变体能够在以上领域进行更好的应用。序列表<110>南京农业大学<120>一种热稳定性提高的脂肪氧合酶突变体及其构建方法<130>1<160>12<170>PatentInversion3.3<210>1<211>455<212>PRT<213>鱼腥藻脂肪氧合酶野生型氨基酸序列<400>1GlyValSerGlyAlaLeuValHisTyrPheGlySerIleValArgAla151015GluArgThrGlnTyrLeuTyrGlySerLysAspAspLeuProGlyLys202530ProValTyrPheProLeuProValThrGluIleProSerLysArgPhe354045LeuPheLeuLeuGluLysTyrAsnPheLeuThrAspAsnSerTyrPro505560SerAspGlyGluHisAspLysIleGluAlaLeuValSerAlaMetPro65707580ThrThrAlaLeuAspLeuAlaValGlyThrThrAspProThrAspIle859095ProAspSerTyrPheLeuGluArgArgLeuAsnGlyTyrAsnProGly100105110AlaIleArgGluSerSerGlyGlnGluGlyTrpThrHisGluLeuThr115120125HisAsnLeuAlaLysTyrAspIleLysProGlyLeuHisPheProAsp130135140PheValGlnCysArgLeuPheValAspLysGlnAsnGlyValLysLeu145150155160HisSerIleLysIleAspAspHisGluIleThrProCysGlnGluGln165170175TrpGlnTyrAlaLysArgThrTyrLeuGlnAlaGluPheLeuSerGln180185190GluLeuLysLeuHisLeuAlaArgCysHisPheAsnIleGluGlnTyr195200205ValMetAlaIleLysArgArgLeuAlaProThrHisProValArgAla210215220PheIleAsnProHisLeuGluGlyLeuIlePheIleAsnSerSerAla225230235240ValProLysIleIleGlySerThrGlyPheIleProIleAlaSerMet245250255LeuThrGlnGlySerIleValAspValMetLysAsnGluLeuSerLys260265270LeuSerTyrMetTrpAsnProIleAlaAspLeuProArgAspIlePro275280285GlyAspLeuPheThrProAlaAlaThrAlaTyrTrpGluLeuLeuAsn290295300AsnTyrValGluGlnGlyLeuLeuGlnProPheGluAspGluLeuArg305310315320ThrGluValAsnAlaIleGlnValAspGluLeuPheAlaGluLeuLys325330335GluArgSerLeuTyrSerGlyAspGlnProProLysTyrAspSerSer340345350GluLeuLysSerLeuLeuMetTyrIleIleTyrHisSerSerPheLeu355360365HisSerTrpAlaAsnPheLysGlnTyrAspAspAlaGlyAsnProAsn370375380HisValSerMetGlyAspTyrSerGlnTyrAspGlnGlnThrGlnAsp385390395400LysIleArgPheSerGlnArgSerLeuThrTrpValLeuSerSerIle405410415ArgTyrAsnSerValAlaValTyrGlySerAspLeuLeuLysGlnLeu420425430IleArgGluLysSerSerIleLeuGluProGlyLeuProLeuGluAsp435440445LeuMetMetSerIleAsnIle450455<210>2<211>455<212>PRT<213>鱼腥藻脂肪氧合酶N130D突变体氨基酸序列<400>2GlyValSerGlyAlaLeuValHisTyrPheGlySerIleValArgAla151015GluArgThrGlnTyrLeuTyrGlySerLysAspAspLeuProGlyLys202530ProValTyrPheProLeuProValThrGluIleProSerLysArgPhe354045LeuPheLeuLeuGluLysTyrAsnPheLeuThrAspAsnSerTyrPro505560SerAspGlyGluHisAspLysIleGluAlaLeuValSerAlaMetPro65707580ThrThrAlaLeuAspLeuAlaValGlyThrThrAspProThrAspIle859095ProAspSerTyrPheLeuGluArgArgLeuAsnGlyTyrAsnProGly100105110AlaIleArgGluSerSerGlyGlnGluGlyTrpThrHisGluLeuThr115120125HisAspLeuAlaLysTyrAspIleLysProGlyLeuHisPheProAsp130135140PheValGlnCysArgLeuPheValAspLysGlnAsnGlyValLysLeu145150155160HisSerIleLysIleAspAspHisGluIleThrProCysGlnGluGln165170175TrpGlnTyrAlaLysArgThrTyrLeuGlnAlaGluPheLeuSerGln180185190GluLeuLysLeuHisLeuAlaArgCysHisPheAsnIleGluGlnTyr195200205ValMetAlaIleLysArgArgLeuAlaProThrHisProValArgAla210215220PheIleAsnProHisLeuGluGlyLeuIlePheIleAsnSerSerAla225230235240ValProLysIleIleGlySerThrGlyPheIleProIleAlaSerMet245250255LeuThrGlnGlySerIleValAspValMetLysAsnGluLeuSerLys260265270 LeuSerTyrMetTrpAsnProIleAlaAspLeuProArgAspIlePro275280285GlyAspLeuPheThrProAlaAlaThrAlaTyrTrpGluLeuLeuAsn290295300AsnTyrValGluGlnGlyLeuLeuGlnProPheGluAspGluLeuArg305310315320ThrGluValAsnAlaIleGlnValAspGluLeuPheAlaGluLeuLys325330335GluArgSerLeuTyrSerGlyAspGlnProProLysTyrAspSerSer340345350GluLeuLysSerLeuLeuMetTyrIleIleTyrHisSerSerPheLeu355360365HisSerTrpAlaAsnPheLysGlnTyrAspAspAlaGlyAsnProAsn370375380HisValSerMetGlyAspTyrSerGlnTyrAspGlnGlnThrGlnAsp385390395400LysIleArgPheSerGlnArgSerLeuThrTrpValLeuSerSerIle405410415ArgTyrAsnSerValAlaValTyrGlySerAspLeuLeuLysGlnLeu420425430IleArgGluLysSerSerIleLeuGluProGlyLeuProLeuGluAsp435440445LeuMetMetSerIleAsnIle450455<210>3<211>455<212>PRT<213>鱼腥藻脂肪氧合酶G260A突变体氨基酸序列<400>3GlyValSerGlyAlaLeuValHisTyrPheGlySerIleValArgAla151015GluArgThrGlnTyrLeuTyrGlySerLysAspAspLeuProGlyLys202530ProValTyrPheProLeuProValThrGluIleProSerLysArgPhe354045LeuPheLeuLeuGluLysTyrAsnPheLeuThrAspAsnSerTyrPro505560SerAspGlyGluHisAspLysIleGluAlaLeuValSerAlaMetPro65707580ThrThrAlaLeuAspLeuAlaValGlyThrThrAspProThrAspIle859095ProAspSerTyrPheLeuGluArgArgLeuAsnGlyTyrAsnProGly100105110AlaIleArgGluSerSerGlyGlnGluGlyTrpThrHisGluLeuThr115120125HisAsnLeuAlaLysTyrAspIleLysProGlyLeuHisPheProAsp130135140PheValGlnCysArgLeuPheValAspLysGlnAsnGlyValLysLeu145150155160HisSerIleLysIleAspAspHisGluIleThrProCysGlnGluGln165170175TrpGlnTyrAlaLysArgThrTyrLeuGlnAlaGluPheLeuSerGln180185190GluLeuLysLeuHisLeuAlaArgCysHisPheAsnIleGluGlnTyr195200205ValMetAlaIleLysArgArgLeuAlaProThrHisProValArgAla210215220PheIleAsnProHisLeuGluGlyLeuIlePheIleAsnSerSerAla225230235240ValProLysIleIleGlySerThrGlyPheIleProIleAlaSerMet245250255LeuThrGlnAlaSerIleValAspValMetLysAsnGluLeuSerLys260265270LeuSerTyrMetTrpAsnProIleAlaAspLeuProArgAspIlePro275280285GlyAspLeuPheThrProAlaAlaThrAlaTyrTrpGluLeuLeuAsn290295300AsnTyrValGluGlnGlyLeuLeuGlnProPheGluAspGluLeuArg305310315320ThrGluValAsnAlaIleGlnValAspGluLeuPheAlaGluLeuLys325330335GluArgSerLeuTyrSerGlyAspGlnProProLysTyrAspSerSer340345350GluLeuLysSerLeuLeuMetTyrIleIleTyrHisSerSerPheLeu355360365HisSerTrpAlaAsnPheLysGlnTyrAspAspAlaGlyAsnProAsn370375380HisValSerMetGlyAspTyrSerGlnTyrAspGlnGlnThrGlnAsp385390395400LysIleArgPheSerGlnArgSerLeuThrTrpValLeuSerSerIle405410415ArgTyrAsnSerValAlaValTyrGlySerAspLeuLeuLysGlnLeu420425430IleArgGluLysSerSerIleLeuGluProGlyLeuProLeuGluAsp435440445LeuMetMetSerIleAsnIle450455<210>4<211>455<212>PRT<213>鱼腥藻脂肪氧合酶S437T突变体氨基酸序列<400>4GlyValSerGlyAlaLeuValHisTyrPheGlySerIleValArgAla151015GluArgThrGlnTyrLeuTyrGlySerLysAspAspLeuProGlyLys202530ProValTyrPheProLeuProValThrGluIleProSerLysArgPhe354045LeuPheLeuLeuGluLysTyrAsnPheLeuThrAspAsnSerTyrPro505560SerAspGlyGluHisAspLysIleGluAlaLeuValSerAlaMetPro65707580ThrThrAlaLeuAspLeuAlaValGlyThrThrAspProThrAspIle859095ProAspSerTyrPheLeuGluArgArgLeuAsnGlyTyrAsnProGly100105110AlaIleArgGluSerSerGlyGlnGluGlyTrpThrHisGluLeuThr115120125HisAsnLeuAlaLysTyrAspIleLysProGlyLeuHisPheProAsp130135140PheValGlnCysArgLeuPheValAspLysGlnAsnGlyValLysLeu145150155160HisSerIleLysIleAspAspHisGluIleThrProCysGlnGluGln165170175TrpGlnTyrAlaLysArgThrTyrLeuGlnAlaGluPheLeuSerGln180185190GluLeuLysLeuHisLeuAlaArgCysHisPheAsnIleGluGlnTyr195200205ValMetAlaIleLysArgArgLeuAlaProThrHisProValArgAla210215220PheIleAsnProHisLeuGluGlyLeuIlePheIleAsnSerSerAla225230235240ValProLysIleIleGlySerThrGlyPheIleProIleAlaSerMet245250255LeuThrGlnGlySerIleValAspValMetLysAsnGluLeuSerLys260265270LeuSerTyrMetTrpAsnProIleAlaAspLeuProArgAspIlePro275280285GlyAspLeuPheThrProAlaAlaThrAlaTyrTrpGluLeuLeuAsn290295300AsnTyrValGluGlnGlyLeuLeuGlnProPheGluAspGluLeuArg305310315320ThrGluValAsnAlaIleGlnValAspGluLeuPheAlaGluLeuLys325330335GluArgSerLeuTyrSerGlyAspGlnProProLysTyrAspSerSer340345350GluLeuLysSerLeuLeuMetTyrIleIleTyrHisSerSerPheLeu355360365HisSerTrpAlaAsnPheLysGlnTyrAspAspAlaGlyAsnProAsn370375380HisValSerMetGlyAspTyrSerGlnTyrAspGlnGlnThrGlnAsp385390395400LysIleArgPheSerGlnArgSerLeuThrTrpValLeuSerSerIle405410415ArgTyrAsnSerValAlaValTyrGlySerAspLeuLeuLysGlnLeu420425430IleArgGluLysThrSerIleLeuGluProGlyLeuProLeuGluAsp435440445LeuMetMetSerIleAsnIle450455<210>5<211>455<212>PRT<213>鱼腥藻脂肪氧合酶N130D/G260Q突变体氨基酸序列<400>5GlyValSerGlyAlaLeuValHisTyrPheGlySerIleValArgAla151015GluArgThrGlnTyrLeuTyrGlySerLysAspAspLeuProGlyLys202530ProValTyrPheProLeuProValThrGluIleProSerLysArgPhe354045LeuPheLeuLeuGluLysTyrAsnPheLeuThrAspAsnSerTyrPro505560SerAspGlyGluHisAspLysIleGluAlaLeuValSerAlaMetPro65707580ThrThrAlaLeuAspLeuAlaValGlyThrThrAspProThrAspIle859095ProAspSerTyrPheLeuGluArgArgLeuAsnGlyTyrAsnProGly100105110AlaIleArgGluSerSerGlyGlnGluGlyTrpThrHisGluLeuThr115120125HisAspLeuAlaLysTyrAspIleLysProGlyLeuHisPheProAsp130135140PheValGlnCysArgLeuPheValAspLysGlnAsnGlyValLysLeu145150155160HisSerIleLysIleAspAspHisGluIleThrProCysGlnGluGln165170175TrpGlnTyrAlaLysArgThrTyrLeuGlnAlaGluPheLeuSerGln180185190GluLeuLysLeuHisLeuAlaArgCysHisPheAsnIleGluGlnTyr195200205ValMetAlaIleLysArgArgLeuAlaProThrHisProValArgAla210215220PheIleAsnProHisLeuGluGlyLeuIlePheIleAsnSerSerAla225230235240ValProLysIleIleGlySerThrGlyPheIleProIleAlaSerMet245250255LeuThrGlnGlnSerIleValAspValMetLysAsnGluLeuSerLys260265270LeuSerTyrMetTrpAsnProIleAlaAspLeuProArgAspIlePro275280285GlyAspLeuPheThrProAlaAlaThrAlaTyrTrpGluLeuLeuAsn290295300AsnTyrValGluGlnGlyLeuLeuGlnProPheGluAspGluLeuArg305310315320ThrGluValAsnAlaIleGlnValAspGluLeuPheAlaGluLeuLys325330335GluArgSerLeuTyrSerGlyAspGlnProProLysTyrAspSerSer340345350GluLeuLysSerLeuLeuMetTyrIleIleTyrHisSerSerPheLeu355360365 HisSerTrpAlaAsnPheLysGlnTyrAspAspAlaGlyAsnProAsn370375380HisValSerMetGlyAspTyrSerGlnTyrAspGlnGlnThrGlnAsp385390395400LysIleArgPheSerGlnArgSerLeuThrTrpValLeuSerSerIle405410415ArgTyrAsnSerValAlaValTyrGlySerAspLeuLeuLysGlnLeu420425430IleArgGluLysSerSerIleLeuGluProGlyLeuProLeuGluAsp435440445LeuMetMetSerIleAsnIle450455<210>6<211>455<212>PRT<213>鱼腥藻脂肪氧合酶N130D/S437Y突变体氨基酸序列<400>6GlyValSerGlyAlaLeuValHisTyrPheGlySerIleValArgAla151015GluArgThrGlnTyrLeuTyrGlySerLysAspAspLeuProGlyLys202530ProValTyrPheProLeuProValThrGluIleProSerLysArgPhe354045LeuPheLeuLeuGluLysTyrAsnPheLeuThrAspAsnSerTyrPro505560SerAspGlyGluHisAspLysIleGluAlaLeuValSerAlaMetPro65707580ThrThrAlaLeuAspLeuAlaValGlyThrThrAspProThrAspIle859095ProAspSerTyrPheLeuGluArgArgLeuAsnGlyTyrAsnProGly100105110AlaIleArgGluSerSerGlyGlnGluGlyTrpThrHisGluLeuThr115120125HisAspLeuAlaLysTyrAspIleLysProGlyLeuHisPheProAsp130135140PheValGlnCysArgLeuPheValAspLysGlnAsnGlyValLysLeu145150155160HisSerIleLysIleAspAspHisGluIleThrProCysGlnGluGln165170175TrpGlnTyrAlaLysArgThrTyrLeuGlnAlaGluPheLeuSerGln180185190GluLeuLysLeuHisLeuAlaArgCysHisPheAsnIleGluGlnTyr195200205ValMetAlaIleLysArgArgLeuAlaProThrHisProValArgAla210215220PheIleAsnProHisLeuGluGlyLeuIlePheIleAsnSerSerAla225230235240ValProLysIleIleGlySerThrGlyPheIleProIleAlaSerMet245250255LeuThrGlnGlySerIleValAspValMetLysAsnGluLeuSerLys260265270LeuSerTyrMetTrpAsnProIleAlaAspLeuProArgAspIlePro275280285GlyAspLeuPheThrProAlaAlaThrAlaTyrTrpGluLeuLeuAsn290295300AsnTyrValGluGlnGlyLeuLeuGlnProPheGluAspGluLeuArg305310315320ThrGluValAsnAlaIleGlnValAspGluLeuPheAlaGluLeuLys325330335GluArgSerLeuTyrSerGlyAspGlnProProLysTyrAspSerSer340345350GluLeuLysSerLeuLeuMetTyrIleIleTyrHisSerSerPheLeu355360365HisSerTrpAlaAsnPheLysGlnTyrAspAspAlaGlyAsnProAsn370375380HisValSerMetGlyAspTyrSerGlnTyrAspGlnGlnThrGlnAsp385390395400LysIleArgPheSerGlnArgSerLeuThrTrpValLeuSerSerIle405410415ArgTyrAsnSerValAlaValTyrGlySerAspLeuLeuLysGlnLeu420425430IleArgGluLysTyrSerIleLeuGluProGlyLeuProLeuGluAsp435440445LeuMetMetSerIleAsnIle450455<210>7<211>32<212>DNA<213>人工序列N130-f<220><221>misc_feature<222>(9)..(10)<223>nisa,c,g,ort<400>7ttactcacnnkctggcaaaatatgacatcaag32<210>8<211>28<212>DNA<213>人工序列N130-r<220><221>misc_feature<222>(10)..(11)<223>nisa,c,g,ort<400>8tttgccagmnngtgagtaagctcatgtg28<210>9<211>32<212>DNA<213>人工序列S437-f<220><221>misc_feature<222>(11)..(12)<223>nisa,c,g,ort<400>9ggaaaaatcannkatattggaaccaggacttc32<210>10<211>31<212>DNA<213>人工序列S437-r<220><221>misc_feature<222>(10)..(11)<223>nisa,c,g,ort<400>10tccaatatmnntgatttttcccgaatgagcg31<210>11<211>29<212>DNA<213>人工序列G260-f<220><221>misc_feature<222>(11)..(12)<223>nisa,c,g,ort<400>11gctaacgcagnnktctattgttgatgtaa29<210>12<211>25<212>DNA<213>人工序列G260-r<220><221>misc_feature<222>(11)..(12)<223>nisa,c,g,ort<400>12aacaatagamnnctgcgttagcatg25当前第1页1 2 3
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1