一种用于中枢神经系统疾病治疗的基因序列构建体的制作方法
本发明涉及基因治疗领域,具体涉及到一种用于中枢神经系统疾病治疗的基因序列构建体,该基因序列构建体可用于对帕金森病、阿尔茨海默症及其他神经退行性疾病进行预防或者治疗。
背景技术
帕金森病(pd)是一种神经退行性疾病,以黑质区多巴胺能神经元丢失为特征,导致中脑多巴胺水平降低而致病。帕金森病影响全球约1%的55岁以上的人群,随着社会步入老年化,患者群体还会持续扩大。帕金森病的常见治疗方法是口服给药多巴胺的前体—左旋多巴,在一定程度上可以缓解症状。然而,随着病情的发展,左旋多巴治疗不尽如人意。
基因治疗对于帕金森病治疗具有独特优势,通过将多巴胺合成基因靶向递送到纹状体可使多巴胺在纹状体内合成并释放,从而使帕金森病的治疗更为有效。酪氨酸经过酪氨酸羟化酶(th)催化合成左旋多巴,随后芳香族氨基酸脱羧酶(aadc)将左旋多巴转化为多巴胺。th需要由四氢生物蝶呤作为辅酶,而四氢生物蝶呤由gtp-环化水解酶1(gch1)催化合成,因此表达th、aadc和gch1可以高效合成多巴胺(azzouzm等人,2002.jneurosci.22(23):10302-12.;jarrayab等人,2009.scitranslmed.1(2):2)。
2014年,基于马传染性贫血病毒(eiav)的慢病毒载体携带以两个内部核糖体进入位点(ires)连接三种关键的多巴胺合成酶在i/ii期临床试验显示了良好的效果(palfis等人,2014.lancet.383(9923):1138-46.)。然而,这个方法具有多个问题,比如ires表达三个基因经常导致表达不平衡。因此,stewarthj等人将接头肽(linker)取代ires元件以产生包含多巴胺合成所需的三种酶活性中的两种或更多种的融合蛋白(stewarthj等人,2016.humgenetherclindev.27(3):100-10.)。然而,天然状态下th、aadc和gch1三个蛋白是以独立蛋白形式表达,融合蛋白有可能会影响多巴胺合成效率,因此,寻找新的蛋白连接表达方式,选择合适的载体导入至靶细胞,并在其中高效表达,仍是帕金森病治疗中亟待解决的问题。
技术实现要素:
本发明的目的是针对现有治疗技术的不足,通过自加工(auto-processing)表达载体的构建,表达含有酪氨酸羟化酶(th)、gtp-环化水解酶i(gch1)、芳香族氨基酸多巴脱羧酶(aadc),以及神经系统生长因子等;蛋白间通过自加工单元(auto-processingunit,apu)连接;利用病毒载体将其靶向性导入目的细胞最终可产生高效表达的具有独立功能的酪氨酸羟化酶(th)、gtp-环化水解酶i(gch1)及芳香族氨基酸多巴脱羧酶(aadc)等,用于治疗帕金森病。
本发明包括如下内容:一种用于中枢神经系统疾病治疗的基因序列构建体,所述的构建体为通过自加工单元(apu)连接的与治疗中枢神经系统疾病相关的核苷酸序列;
优选地,所述的用于中枢神经系统疾病治疗的基因序列构建体中的与中枢神经系统疾病相关的核苷酸序列选自于酪氨酸羟化酶(th)、gtp-环化水解酶i(gch1)、芳香族氨基酸多巴脱羧酶(aadc)、神经系统生长因子核苷酸序列中的两种或者两种以上;
所述的核苷酸序列之间至少有两个核苷酸序列之间通过自加工单元(apu)连接;所述的自加工单元(apu)包含n-端自加工结构域和/或c-端自加工结构域;
优选地,所述的神经系统生长因子包括神经生长因子(ngf)、脑衍生神经营养因子(bdnf)、神经生长因子-3(nt-3)、神经生长因子-4/5(nt-4/5)、神经生长因子-6(nt-6)、睫状神经营养因子(cntf)、胶质细胞系衍生神经营养因子(gdnf)和gdnf分子家族(gdnf的天然存在的类似物、神经秩蛋白、persephin和artemin);
优选地,所述的用于中枢神经系统疾病治疗的基因序列构建体包括酪氨酸羟化酶(th)、gtp-环化水解酶i(gch1)、芳香族氨基酸多巴脱羧酶(aadc);
所述的核苷酸序列之间至少有两个序列之间通过自加工单元(apu)连接;
优选地,所述的n-端自加工结构域包括intein、b型细菌体intein样结构域(bil)、furin序列,及其衍生物;
优选地,所述的c-端自加工结构域包括2a肽或类2a肽;
优选地,所述的用于中枢神经系统疾病治疗的基因序列构建体中的2a肽或类2a肽包括源于口蹄疫病毒的2a肽(f2a)、源于猪捷申病毒的2a肽(p2a)、源于昆虫病毒的2a肽(t2a),以及源于马鼻炎病毒的2a肽(e2a)。
自加工2a肽最早源于口蹄疫病毒(fmdv)。fmdv属于小rna病毒科口蹄疫病毒属。fmdv基因组编码的蛋白酶2a可在翻译过程中形成的高级结构对核糖体肽基转移酶中心造成空间位阻,导致无法形成正常的肽链连接,但同时核糖体却能继续翻译下游蛋白,从而形成一种类似蛋白水解酶的作用将前后两个蛋白顺式“切开”;与fmdv类似,微小核糖核酸病毒科中的心病毒、蒂勒氏鼠脑脊髓病毒、马鼻炎病毒、猪捷申病毒等也含有一2a肽段;此外,在昆虫病毒、c型轮状病毒及锥虫型重复序列中都发现具有2a肽功能类似的基因序列。2a肽或类2a肽自我加工序列与内部核糖体进入位点(ires)一样都常被应用于多基因表达,以达到独立表达两个及以上非融合外源蛋白的目的;与ires相比,2a肽或类2a肽在多基因表达载体构建上有明显的优势,比如,2a肽或类2a肽比较小,且2a元件所连接的上下游基因的表达平衡性好;本发明使用自加工肽p2a来连接th、aadc和gch1三个蛋白,以获得三个蛋白独立高效的表达。
一种病毒载体基因组,所述的病毒载体基因组包括上述任一所述的用于中枢神经系统疾病治疗的基因序列构建体;
所述的病毒载体包括慢病毒载体或腺相关病毒载体。
一种慢病毒载体系统,所述的慢病毒载体系统包括包含有上述的用于中枢神经系统疾病治疗的基因序列构建体的基因组,以及编码gag和pol蛋白的一个或者多个核苷酸序列以及其它必需的病毒包装组件核苷酸序列。
一种包括上述任一所述基因序列构建体的生物制剂,其在治疗和/或预防帕金森病、阿尔茨海默症及其他神经退行性疾病中的用途。
一种包括上面任一所述基因序列构建体的生物制剂,包括包含有上面的基因序列的构建体病毒载体系统,以及其它必需的病毒包装组件,进而产生病毒颗粒加上药学上可接受的载体或者稀释剂形成的生物制剂,以及用于制备用于体内产生多巴胺的药物,其在治疗和/或预防帕金森病、阿尔茨海默症及其他神经退行性疾病中的用途。
具体地,一种基因序列构建体,所述的构建体包括选自于酪氨酸羟化酶(th)、gtp-环化水解酶i(gch1)、芳香族氨基酸多巴脱羧酶(aadc)核苷酸序列中的两种或者两种以上;
所述的核苷酸序列之间至少有两个序列之间通过自加工单元(apu)连接。
现选取一种基因序列构建体包括酪氨酸羟化酶(th)、gtp-环化水解酶i(gch1)、芳香族氨基酸多巴脱羧酶(aadc)核苷酸序列,该核苷酸序列之间至少有两个序列之间通过自加工单元(apu)连接;该基因序列构建体包括如下构建方式:
th-apu-ch1-apu-aadc;th-apu-ch1-其他序列-aadc;th-其他序列-ch1-apu-aadc;
th-apu-aadc-apu-ch1;th-其他序列-aadc-apu-ch1;th-apu-aadc-其他序列-ch1;
ch1-apu-th-apu-aadc;ch1-apu-th-其他序列-aadc;ch1-其他序列-th-apu-aadc;
ch1-apu-aadc-apu-th;ch1-apu-aadc-其他序列-th;ch1-其他序列-aadc-apu-th;
aadc-apu-th-apu-ch1;aadc-apu-th-其他序列-ch1;aadc-其他序列-th-apu-ch1;
aadc-apu-ch1-apu-th;aadc-apu-ch1-其他序列-th;aadc-其他序列-ch1-apu-th;
apu:自加工单元;其他序列:包括接头肽编码序列(linker)、内部核糖体进入位点(ires)、启动子或内含肽;
该构建体中的自加工单元(apu)包括2a肽或类2a肽;
基因序列构建体中的2a肽或类2a肽包括源于口蹄疫病毒的2a肽(f2a)、源于猪捷申病毒的2a肽(p2a)、源于昆虫病毒的2a肽(t2a),以及源于马鼻炎病毒的2a肽(e2a)。
基因序列构建体中如果包括启动子的话,其启动子为组成型启动子或者组织特异性启动子;组成型启动子,包括cmv启动子、磷酸甘油酸激酶启动子或者胸苷激酶启动子。组织特异性启动子,包括突触素(synapsin)启动子、cd68启动子、gfap启动子或其他合成的启动子。
与现有技术相比,本发明具有如下优点:
1、本发明提供了一种新的连接多巴胺合成的基因的方法;本发明实验研究表明,该方法可提高目标蛋白表达的均衡性,多巴胺及其代谢产物合成水平升高,可提高抗帕金森病的治疗效果。
2、本发明首次确定了利用p2a的自加工肽连接各蛋白的表达,最终可产生表达稳定且均衡的具有独立功能的酪氨酸羟化酶(th)、gtp-环化水解酶i(gch1)、芳香族氨基酸多巴脱羧酶(aadc),以及神经系统生长因子,能够为帕金森病及其其他神经退行性疾病的治疗和/或预防提供新的策略。
附图说明
为了更清楚地说明本发明实施例或现有技术中的技术方案,下面将对实施例或现有技术描述中所需要使用的附图作简单地介绍,显而易见地,下面描述中的附图仅仅是本发明的一些实施例,对于本领域普通技术人员来讲,在不付出创造性劳动性的前提下,还可以根据这些附图获得其他的附图。
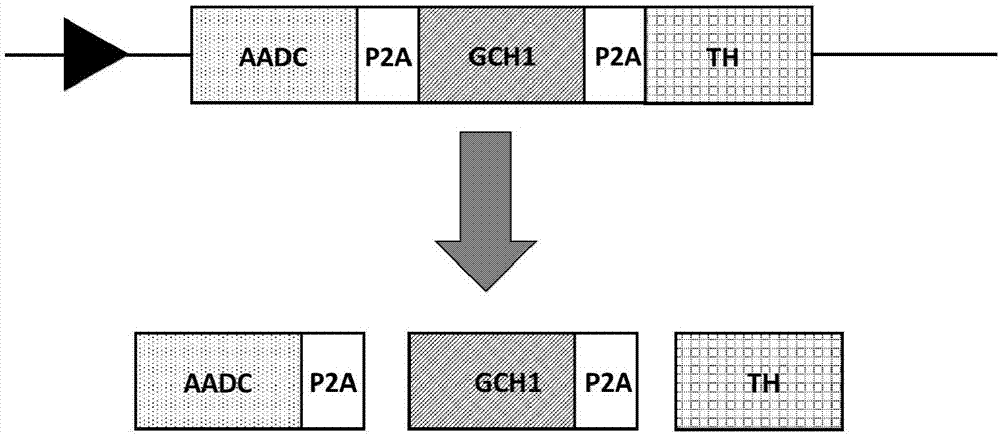
图1为本发明的用于帕金森病基因疗法的基因序列构建体示意图。
图2为本发明的构建体结构示意图。
图3转导293t细胞后,芳香族氨基酸多巴脱羧酶(aadc)、gtp-环化水解酶i(gch1),及酪氨酸羟化酶(th)蛋白的westernblot检测结果。
图4转导sh-sy5y细胞后,芳香族氨基酸多巴脱羧酶(aadc)、gtp-环化水解酶i(gch1),及酪氨酸羟化酶(th)蛋白的westernblot检测。
图5病毒转导sh-sy5y细胞后,hplc检测多巴胺da(图5a)及其代谢产物高香草酸hva(图5b)产量的结果。
其中图1中:包含表达芳香族氨基酸多巴脱羧酶(aadc)、gtp-环化水解酶i(gch1),及酪氨酸羟化酶(th)的编码序列,该蛋白间由自加工肽p2a连接;该基因转录翻译,经自加工肽加工后可产生独立的芳香族氨基酸多巴脱羧酶(aadc-p2a)、gtp-环化水解酶i(gch1-p2a),及酪氨酸羟化酶(th);p2a:猪捷申病毒2a自加工肽。
图2中:aadc:芳香族氨基酸多巴脱羧酶;gch1:gtp-环化水解酶i;th:酪氨酸羟化酶;p2a:猪捷申病毒的2a加工肽;synapsin、cmv、sv40及pgk皆为启动子。
图3中:blank:无病毒转导组;gfp:cmvpromoter-egfp病毒转导组;pd-1:synapsinpromoter-aadc-p2a-gch1-p2a-th病毒转导组;pd-2:cmvpromoter-aadc-p2a-gch1-p2a-th病毒转导组;p:cmvpromoter-aadc-sv40promoter-th-pgkpromoter-gch1病毒转导组;。endo-th:为细胞内源性的th;exo-th:为外源性过表达的仅催化结构域的th。
图4中:blank:无病毒转导组;gfp:cmvpromoter-egfp病毒转导组;pd-1:synapsinpromoter-aadc-p2a-gch1-p2a-th病毒转导组;pd-2:cmvpromoter-aadc-p2a-gch1-p2a-th病毒转导组;p:cmvpromoter-aadc-sv40promoter-th-pgkpromoter-gch1病毒转导组;endo-th:为细胞内源性的th;exo-th:为外源性过表达的仅催化结构域的th。
图5中:gfp:gfp病毒转导组;pd-2:cmvpromoter-aadc-p2a-gch1-p2a-th病毒转导组;p:cmvpromoter-aadc-sv40promoter-th-pgkpromoter-gch1病毒转导组。
具体实施方式
下面结合实施例对本发明做进一步的详细说明,以下实施例是对本发明的解释而本发明并不局限于以下实施例。
实施例:
一、如图2所示,各种构建体的构建:
kl0039载体,人工合成的cmvenhancer-synapsinpromoter-aadc-p2a-gch1-p2a-th及aadc-sv40promoter-th-pgkpromoter-gch1序列(其中th为截短形式的th);其中,cmvenhancer-synapsinpromoter-aadc-p2a-gch1-p2a-th连接至puc57载体(puc57-synapsin-agt);此处的kl0039载体为一种慢病毒转移载体,选自现有的慢病毒载体或者根据需要对其进行部分改造的慢病毒载体。
1、pd1载体构建
以kl0039载体为模板,用age-f+sal-r扩增wpre到cppt之间的序列,电泳回收并纯化pcr产物。所用引物序列为age-f:ctgagtgccattggatgacaatcaacctctggattaca;sal-r:gattactattaataactactcacgcatgctcttctcca。同时用agei和sali酶切puc57-synapsin-agt质粒,回收4.1kb大小的条带;回收纯化的pcr产物与酶切后的synapsin-agt片段经t4dna连接酶连接后,转化dh5α感受态细胞。挑取数个单克隆进行pcr鉴定,阳性克隆再经测序确认
2、pd2载体构建
以kl0039载体为模板,用引物snabi-f:
tcagtacgtattagtcatcgctat和spei-r:
cgatactagtgagctctgcttatataga扩增cmvpromoter序列,电泳回收纯化pcr产物(245bp)。snabi和spei同时双酶切cmvpromoter和puc57-synapsin-agt质粒,分别回收纯化cmvpromoter和puc57-agt片段,经t4dna连接酶连接后转化dh5α感受态细胞。挑取单克隆转化子进行pcr鉴定。阳性克隆再经测序确认。此阳性克隆命名为puc57-cmv-agt质粒。然后以kl0039载体为模板,用age-f+sal-r扩增wpre到cppt之间的序列,电泳回收并纯化pcr产物。用agei和sali同时双酶切pcr产物和puc57-cmv-agt质粒,电泳回收纯化pcr产物及cmv-agt片段。经t4dna连接酶连接后转化dh5α感受态细胞。挑取单克隆转化子进行pcr鉴定。阳性克隆再经测序确认。
3、p载体构建:序列aadc-sv40promoter-th-pgkpromoter-gch1置换pd2载体中agt序列。
4.gfp载体:克隆egfp序列,置换pd2载体中的agt序列。
二、评估不同构建体转导293t及sh-sy5y细胞后目标蛋白的表达差异
利用慢病毒四质粒体系瞬时转染293t细胞系,分别包装gfp(cmvpromoter-egfp)、pd1(synapsinpromoter-agt)、pd2(cmvpromoter-agt)的慢病毒及阳性对照病毒p(cmvpromoter-aadc-sv40promoter-th-pgkpromoter-gch1)。初始病毒经纯化后浓缩,稀释后转导293t细胞,利用rt-pcr(wpre/alb)测定其滴度,所有构建体的载体滴度是相似的,滴度范围从3.4e+09tu/ml至8.74e+09tu/ml。
为了研究目标蛋白表达水平,以各组病毒moi=10转导293t细胞及moi=20转导sh-sy5y细胞,分别转导72小时后收集细胞裂解物进行aadc、gch1和th的westernblot分析。结果表明,在293t细胞中,内源性蛋白除检测到较低的th外,aadc与gch1皆未检测到。所有蛋白分子量与预测相同。与无病毒转导组blank及gfp病毒转导对照组相比较,所有三种蛋白的最高水平表达来自pd2病毒载体,虽然p病毒载体的aadc水平表达最高,但是其他两个蛋白gch1和th基本检测不到。而对于pd1病毒载体,因为其启动子为神经元特异性启动子synapsin,与预期相同,在293t细胞中三个目标蛋白皆无表达(图3)。我们进一步检测了三种不同构建体在神经细胞sh-sy5y中目标蛋白的表达,内源性蛋白th与aadc皆有表达,而gch1未检测出,所有蛋白分子量与预测相同。与293t细胞结果一致,与无病毒转导组blank及gfp病毒转导对照组相比较,所有三种蛋白的最高水平表达仍来自pd2病毒载体,虽然p病毒载体的aadc水平表达最高,但是其他两个蛋白gch1和th仍基本检测不到。而对于pd1病毒载体,仅见少量的aadc及th的表达,gch1基本检测不到(图4)。
三、评估不同构建体转导sh-sy5y细胞后儿茶酚胺产量差异
神经元内的da主要由单胺氧化酶(mao)转化为二羟基苯乙酸(dopac),它既可在神经元内又可在神经元外形成,在细胞外儿茶酚胺氧化甲基转移酶(comt)的作用下,dopac转化为高香草酸(hva)。
因此,我们通过hplc分析从病毒转导后的sh-sy5y细胞上清中产生的儿茶酚胺,包含da及hva的检测。病毒转导sh-sy5y细胞,60小时后,收集细胞培养上清,4500rpm离心5分钟,移取上清液进行hplc检测。结果如图5所示,与gfp病毒载体转导的细胞相比较,虽然p病毒载体转导sh-sy5y后westernblot蛋白检测结果显示外源性的aadc大量表达,但因其另外两种蛋白th及gch1表达极低甚至检测不到,所以并未见da及其代谢产物hva的产量提升;而对于pd2病毒载体,其转导的细胞三种蛋白均表现出高效的表达,hplc检测的结果也显示da产量的增加(图5a),hva产量则是极大地提升,表现出约5倍的增加(图5b),这证实cmv启动子及p2a自加工载体比其他启动子及接头更具有优势。
此外,需要说明的是,本说明书中所描述的具体实施例,其各种物质或者载体所取名称等可以不同。凡依本发明专利构思所述的构造及原理所做的等效或简单变化,均包括于本发明专利的保护范围内。本发明所属技术领域的技术人员可以对所描述的具体实施例做各种各样的修改或补充或采用类似的方式替代,只要不偏离本发明的结构或者超越本权利要求书所定义的范围,均应属于本发明的保护范围。
- 还没有人留言评论。精彩留言会获得点赞!