特异抑制3β-HSD基因表达的shRNA重组载体构建与应用的制作方法
本发明属于基因工程技术领域,尤其涉及一种特异抑制3β-hsd基因表达的shrna重组载体构建与应用。
背景技术
3β-羟类固醇脱氢酶(3β-hydroxysteroiddehydrogenase,3β-hsd)广泛分布于脑、心脏、肾脏、脂肪、皮肤及甾体激素生成组织中,主要参与糖皮质激素、盐皮质激素以及性激素合成,发挥限速酶作用。由于3β-hsd催化类固醇激素生成的起始步骤,若其表达异常,则可能影响所有类固醇激素的合成。
有研究表明,邻苯二甲酸二(2-乙基己基)酯(dehp)可引起3β-hsdmrna和蛋白表达水平升高,dehp能够通过影响类固醇激素合成过程中3β-hsd表达水平的变化,从而产生生殖毒性作用。
在转录水平调控基因表达方面,rna干扰(rnai)被广泛应用于特异抑制基因的表达及其功能研究。在现有rna干扰技术中,以质粒为载体将小干扰rna(sirna)导入细胞内,专一作用于靶向基因的细胞系。sirna与载体质粒组成的表达载体可以在短时间内沉默3β-hsd基因的表达,但其存在干扰效果明显、转染效率低、细胞毒性大且无法实现长效稳定干扰的缺点。现有技术中尚无转染效率高且可长效稳定表达、特异抑制3β-hsd基因表达的shrna表达载体。
技术实现要素:
基于此,有必要针对现有技术中rna干扰沉默3β-hsd基因的表达的效果不明显且效果不稳定的问题,提供一种抑制3β-hsd基因表达的干扰性shrna、重组载体及其构建方法和应用。
一种抑制3β-hsd基因表达的干扰性shrna,包括依次连接的靶序列、茎环结构、所述靶序列的互补序列以及终止位点,其中,
所述靶序列如seq.id.no.1所示,或者,
所述靶序列如seq.id.no.2所示,或者,
所述靶序列如seq.id.no.3所示。
在其中一个实施例中,所述干扰性shrna的正义链如seqidno.4所示,所述干扰性shrna的反义链如seqidno.5所示;或者,
所述干扰性shrna的正义链如seqidno.6所示,所述干扰性shrna的反义链如seqidno.7所示;或者,
所述干扰性shrna的正义链如seqidno.8所示,所述干扰性shrna的反义链如seqidno.9所示。
一种抑制3β-hsd基因表达的重组载体,包括慢病毒载体及插入在所述慢病毒载体中的上述干扰性shrna。
在其中一个实施例中,所述慢病毒载体为pcmv-vsv-g,所述pcmv-vsv-g包括bamhi酶切位点、ecori酶切位点及kpni酶切位点,所述干扰性shrna包括bamhi酶切位点粘性末端及ecori酶切位点粘性末端。
一种抑制3β-hsd基因表达的重组载体的构建方法,包括以下步骤:
提供上述抑制3β-hsd基因表达的干扰性shrna;及
将上述抑制3β-hsd基因表达的干扰性shrna插入慢病毒载体中,得到所述抑制3β-hsd基因表达的重组载体。
一种抑制3β-hsd基因表达的慢病毒,通过如下步骤制得:
将上述抑制3β-hsd基因表达的重组载体转染进入293ft细胞中;及
对转染后的所述293ft细胞扩增表达,获得所述抑制3β-hsd基因表达的慢病毒。
一种重组工程菌,所述重组工程菌内含有上述抑制3β-hsd基因表达的干扰性shrna、上述抑制3β-hsd基因表达的重组载体及上述抑制3β-hsd基因表达的慢病毒中的至少一种。
上述抑制3β-hsd基因表达的干扰性shrna、上述抑制3β-hsd基因表达的重组载体、上述抑制3β-hsd基因表达的慢病毒或上述重组工程菌在制备治疗3β-hsd基因异常表达相关疾病的药物中的应用。
上述抑制3β-hsd基因表达的干扰性shrna、上述抑制3β-hsd基因表达的重组载体、上述抑制3β-hsd基因表达的慢病毒或上述重组工程菌在制备缓解和治疗塑化剂中毒的药物中的应用。
一种药物组合物,包括上述抑制3β-hsd基因表达的干扰性shrna、上述抑制3β-hsd基因表达的重组载体、上述抑制3β-hsd基因表达的慢病毒或上述重组工程菌。
本研究在抑制3β-hsd基因表达的方面进行了大量的探索,预料不到地发现,上述干扰性shrna通过与3β-hsd基因中部分序列特异性地结合而持续、稳定、高效地抑制3β-hsd基因表达。经试验证实,含有上述干扰性shrna的重组载体能够转染到人源细胞中,转染效率高,且转染了该抑制3β-hsd基因表达的重组载体的乳腺癌细胞mcf-7中3β-hsd基因表达的抑制率为61.6%~91.9%,而对照组的乳腺癌细胞mcf-7中3β-hsd基因表达几乎无影响,说明抑制3β-hsd基因表达的干扰性shrna能够持续、稳定、高效且特异性地抑制人源细胞中3β-hsd基因的表达。
附图说明
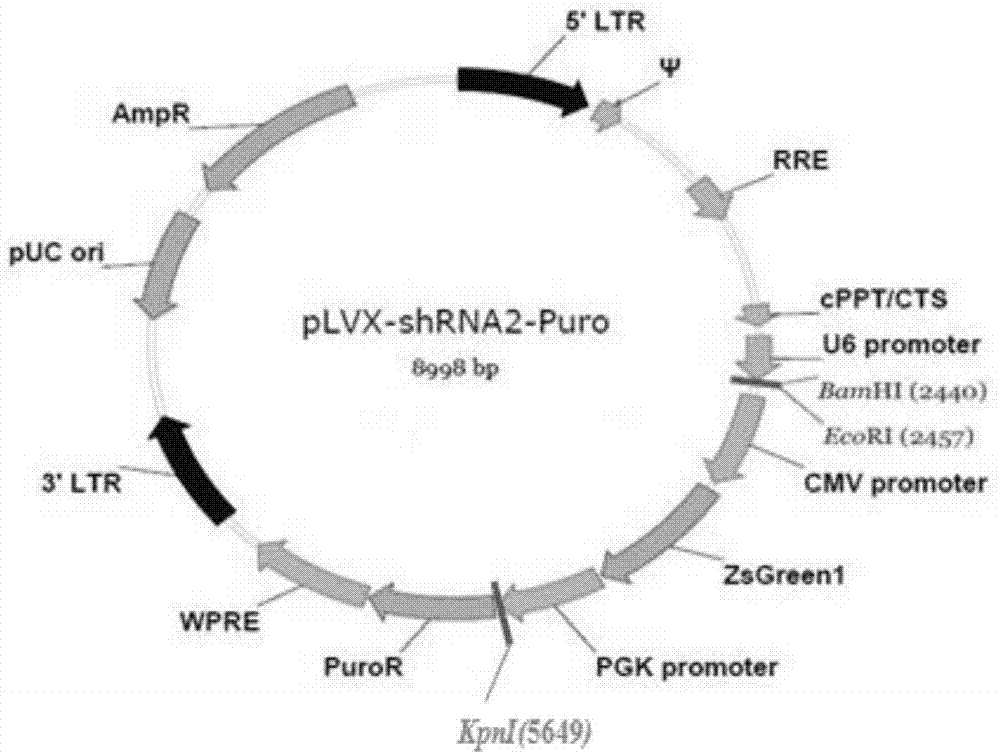
图1为plvx-shrna2p载体的图谱;
图2为实施例1中重组载体的酶切鉴定的电泳图;
图3为实施例1中plvx-shrna2pshrna1重组载体的部分测序结果图;
图4为实施例1中plvx-shrna2pshrna2重组载体的部分测序结果图;
图5为实施例1中plvx-shrna2pshrna3重组载体的部分测序结果图;
图6为实施例1中plvx-shrna2pshrnac重组载体的部分测序结果图;
图7为实施例4中荧光定量pcr检测四组沉默细胞中3β-hsd的mrna表达结果图;
图8为实施例5中westernblot检测plvx-shrna2pshrna2沉默细胞3β-hsd蛋白表达水平的定性结果图;
图9为实施例5中westernblot检测plvx-shrna2pshrna2沉默细胞3β-hsd蛋白表达水平的定量结果图;
图10为实施例6中传代培养后westernblot检测四组沉默细胞3β-hsd蛋白表达的定性结果图;
图11为实施例7中荧光定量pcr检测不同浓度的dehp条件下,mcf-7细胞、plvx-shrna2pshrna3沉默细胞凋亡基因bax的mrna表达水平结果图;
图12为实施例7中荧光定量pcr检测不同浓度的dehp条件下,mcf-7细胞、plvx-shrna2pshrna3沉默细胞凋亡基因caspase-3的mrna表达水平结果图;
图13为实施例7中荧光定量pcr检测不同浓度的dehp条件下,mcf-7细胞、plvx-shrna2pshrna3沉默细胞凋亡基因caspase-8的mrna表达水平结果图;
图14为实施例8中westernblot检测不同浓度的dehp条件下,mcf-7细胞、plvx-shrna2pshrna3沉默细胞凋亡基因bax的蛋白表达水平结果图;
图15为实施例8中westernblot检测不同浓度的dehp条件下,mcf-7细胞、plvx-shrna2pshrna3沉默细胞凋亡基因caspase-3的蛋白表达水平结果图;
图16为实施例8中westernblot检测不同浓度的dehp条件下,mcf-7细胞、plvx-shrna2pshrna3沉默细胞凋亡基因caspase-8的蛋白表达水平结果图。
具体实施方式
为了便于理解本发明,下面将参照相关附图对本发明进行更全面的描述。附图中给出了本发明的部分实施例。但是,本发明可以以许多不同的形式来实现,并不限于本文所描述的实施例。相反地,提供这些实施例的目的是使本发明公开内容更加透彻全面。
除非另有定义,本文所使用的所有的技术和科学术语与属于本发明的技术领域的技术人员通常理解的含义相同。本文中在本发明的说明书中所使用的术语只是为了描述具体的实施例的目的,不是旨在于限制本发明。
一种抑制3β-hsd基因表达的干扰性shrna,包括依次连接的靶序列、茎环结构、靶序列的互补序列以及终止位点。
在其中一个实施例中,靶序列如seq.id.no.1所示。
在其中一个实施例中,靶序列如seq.id.no.2所示。
在其中一个实施例中,靶序列如seq.id.no.3所示。
茎环结构用于使干扰性shrna呈茎环。
进一步地,茎环结构的碱基序列为5'-ttcaagaga-3'。
终止位点为rna聚合酶的转录终止位点。
在其中一个实施例中,终止位点为rnapolyiii聚合酶的转录终止位点。
在其中一个实施例中,终止位点的碱基序列为5'-ttttt-3'。
在其中一个实施例中,干扰性shrna还包括酶切位点粘性末端。使得干扰性shrna能够与载体中的酶切位点粘性末端连接。
进一步地,干扰性shrna从5'端到3'端包括依次连接的bamhi酶切位点粘性末端、靶序列、茎环结构、靶序列的互补序列、终止位点以及ecori酶切位点粘性末端。
当然,需要说明的是,干扰性shrna中的酶切位点粘性末端不限于为bamhi酶切位点粘性末端和ecori酶切位点粘性末端,还可以是其他酶切位点粘性末端,只要用于与干扰性shrna构成重组载体的载体具有相应的酶切位点,使得干扰性shrna能够插入其中即可。
在其中一个实施例中,干扰性shrna以双链的形式存在。双链形式的干扰性shrna能够插入病毒载体中,以转染宿主细胞,进而持续、稳定、高效且特异性地抑制宿主细胞中3β-hsd基因的表达。
进一步地,干扰性shrna的正义链如seqidno.4所示,干扰性shrna的反义链如seqidno.5所示。
进一步地,干扰性shrna的正义链如seqidno.6所示,干扰性shrna的反义链如seqidno.7所示。
进一步地,干扰性shrna的正义链如seqidno.8所示,干扰性shrna的反义链如seqidno.9所示。
本研究在抑制3β-hsd基因表达方面进行了大量的探索,预料不到地发现,上述干扰性shrna通过与3β-hsd基因中部分序列特异性地结合而持续、稳定、高效地抑制3β-hsd基因表达。经试验证实,含有上述干扰性shrna的重组载体能够转染到人源细胞中,转染效率高,且转染了该抑制3β-hsd基因表达的重组载体的乳腺癌细胞mcf-7中3β-hsd基因表达的抑制率为61.6%~79.4%,而对照组的乳腺癌细胞mcf-7中3β-hsd基因表达抑制效率约为1%,说明抑制3β-hsd基因表达的干扰性shrna能够持续、稳定、高效且特异性地抑制人源细胞中3β-hsd基因的表达。
一种抑制3β-hsd基因表达的重组载体,包括慢病毒载体及插入在慢病毒载体中的上述干扰性shrna。
具体地,慢病毒载体包括基本序列、抗性基因序列、多克隆位点序列以及启动子序列。
在其中一个实施例中,抗性基因序列包括氨苄青霉素抗性基因序列。
多克隆位点序列含有多个限制内切酶单一识别位点。能为外源dna提供多种可插入的位置或插入方案。
在其中一个实施例中,慢病毒载体为plvx-shrna2p。选用plvx-shrna2p是因为plvx-shrna2p载体带有gfp蛋白和puromycin抗性筛选标签,可双重筛选细胞株,快速、高效完成细胞株鉴定。另外,plvx-shrna2p含有zsgreen表达框,还可用于直接转染效率的检测和病毒滴度检测。
plvx-shrna2p包括bamhi酶切位点、ecori酶切位点及kpni酶切位点。plvx-shrna2p的bamhi酶切位点能够与干扰性shrna的bamhi酶切位点粘性末端连接,plvx-shrna2p的ecori酶切位点能够与干扰性shrna的ecori酶切位点粘性末端连接,从而形成抑制3β-hsd基因表达的重组载体。
plvx-shrna2p的kpni酶切位点能够与其bamhi酶切位点配合,以验证干扰性shrna是否成功插入plvx-shrna2p。
在其中一个实施方式中,干扰性shrna正向插入慢病毒载体的bamhi酶切位点和ecori酶切位点之间。
本研究在抑制3β-hsd基因表达的靶序列的选择、干扰性shrna的选择、载体的选择以及干扰性shrna插入位点等方面进行了方面进行了大量的探索,预料不到地发现,将上述干扰性shrna插入慢病毒表达载体中,成功构建能够特异性抑制3β-hsd基因的重组载体。将该重组载体转染到人源细胞中的转染效率高,表达载体的用量少,且该重组载体能够持续、稳定、高效地抑制3β-hsd基因表达。如,转染了该抑制3β-hsd基因表达的重组载体的乳腺癌细胞mcf-7中3β-hsd基因表达的抑制率为61.6%~79.4%,而对照组的乳腺癌细胞mcf-7中3β-hsd基因表基因表达抑制效率约为1%。该重组载体能够作为有力工具应用于制备治疗3β-hsd基因表达异常相关疾病的药物。
一实施方式的抑制3β-hsd基因表达的重组载体的构建方法,包括以下步骤:
s110、提供上上述抑制3β-hsd基因表达的干扰性shrna中的任意一种。
具体地,通过基因合成的方式获得干扰性shrna的正义链及反义链。然后将正义链和反义链混合,退火形成干扰性shrna。
s120、将干扰性shrna插入慢病毒载体中,得到抑制3β-hsd基因表达的重组载体。
具体地,将干扰性shrna插入到经过限制性内切酶bamhi和限制性内切酶ecori双酶切处理后的慢病毒载体中,得到抑制3β-hsd基因表达的重组载体。
进一步地,慢病毒载体先经过限制性内切酶bamhi与限制性内切酶ecori双酶切处理,打开缺口,形成慢病毒载体的酶切位点粘性末端,使得抑制3β-hsd基因表达的干扰性shrna与慢病毒载体的酶切位点粘性末端连接,从而获得抑制3β-hsd基因表达的重组载体。
具体地,用限制性内切酶bamhi与限制性内切酶ecori双酶切慢病毒载体后,再用dna连接酶将干扰性shrna连接到双酶切处理后的plvx-shrna2p的bamhi酶切位点与ecori酶切位点之间,获得抑制3β-hsd基因表达的重组载体。
进一步地,慢病毒载体为plvx-shrna2p。
进一步地,dna连接酶为t4dna连接酶。
在其中一个实施例中,得到抑制3β-hsd基因表达的重组载体后,还包括对抑制3β-hsd基因表达的重组载体进行扩增和鉴定。
具体地,将抑制3β-hsd基因表达的重组载体转化入大肠杆菌感受态细菌中,筛选阳性单克隆细菌,对阳性单克隆细菌进行鉴定。其中,大肠杆菌感受态细菌可以为jm107感受态细菌。
在本实施方式中,将抑制3β-hsd基因表达的重组载体转化入jm107感受态细菌中,涂布于具有抗性的培养基上培养,挑取阳性单克隆细菌进行限制性内切酶bamhi和限制性内切酶ecori双酶切鉴定。当然,需要说明的是,对阳性单克隆细菌进行鉴定的方式不限于限制性内切酶bamhi和限制性内切酶ecori双酶切鉴定,还可以对阳性单克隆细菌进行测序鉴定。
上述抑制3β-hsd基因表达的表达载体的构建方法操作简单,通过将含有靶序列的干扰性shrna插入慢病毒载体中,成功地构建能够特异性抑制3β-hsd基因的重组载体。实验结果表明该重组载体能够转染到人源细胞中,转染效率高,表达载体的用量少,且能够持续、稳定、高效且特异性地抑制人源细胞中3β-hsd基因的表达。
一实施方式的抑制3β-hsd基因表达的慢病毒,通过如下步骤制得:将上述抑制3β-hsd基因表达的重组载体转染进入293ft细胞中,对转染后的293ft细胞扩增表达,获得抑制3β-hsd基因表达的慢病毒。
具体地,将抑制3β-hsd基因表达的重组载体与包装载体混合并共转染到293ft细胞中,并培养转染后的293ft细胞,收集上清并过滤,即为抑制3β-hsd基因表达的慢病毒的上清液。
进一步地,包装载体包括包膜蛋白质粒pcmv-vsv-g及包装辅助质粒pcmv-dr8.91。
更进一步地,抑制3β-hsd基因表达的重组载体、pcmv-vsv-g和pcmv-dr8.91的质量比为1:0.5~2:0.5~2。
进一步地,培养转染后的293ft细胞的时间为24h~48h。
进一步地,使用lenti-xgostix试剂盒检测病毒滴度。
需要说明的是,包装载体也可以是其他载体,只要能够与抑制3β-hsd基因表达的重组载体配合促进其扩增及分泌即可。当然,包装载体还也可以省略,只要重组载体自身能够扩增及分泌即可。
上述抑制3β-hsd基因表达的慢病毒,能够转导到人源细胞中,且该慢病毒能够持续、稳定、高效地抑制该人源细胞中3β-hsd基因表达。
一实施方式的3β-hsd基因表达受抑制的细胞,通过如下步骤制备得到:用上述抑制3β-hsd基因表达的慢病毒感染的mcf-7细胞,并培养感染后的mcf-7细胞,得到3β-hsd基因表达受抑制的细胞。
具体地,培养mcf-7细胞至细胞汇合度至60%~90%,用上述抑制3β-hsd基因表达的慢病毒感染mcf-7细胞,并于36.5℃~38℃下培养24小时~48小时,得到3β-hsd基因表达受抑制的细胞。
一实施方式的重组工程菌,含有上述抑制3β-hsd基因表达的干扰性shrna、上述抑制3β-hsd基因表达的重组载体及上述抑制3β-hsd基因表达的慢病毒中的至少一种。
上述重组工程菌含有上述抑制3β-hsd基因表达的干扰性shrna、上述抑制3β-hsd基因表达的重组载体及上述抑制3β-hsd基因表达的慢病毒中的至少一种,能够持续、稳定、高效地抑制3β-hsd基因表达。
一实施方式的上述抑制3β-hsd基因表达的干扰性shrna、上述抑制3β-hsd基因表达的重组载体、上述抑制3β-hsd基因表达的慢病毒或上述在制备治疗3β-hsd基因异常表达相关疾病的药物中的应用。
一实施方式的上述抑制3β-hsd基因表达的干扰性shrna、上述抑制3β-hsd基因表达的重组载体、上述抑制3β-hsd基因表达的慢病毒或上述重组工程菌在制备缓解和治疗塑化剂中毒的药物中的应用。
在其中一个实施例中,塑化剂为邻苯二甲酸二(2-乙基)己酯(dehp)。
试验结果表明,用塑化剂分别处理未转染抑制3β-hsd基因表达的慢病毒的mcf-7与转染抑制3β-hsd基因表达的慢病毒后的mcf-7,转染后的mcf-7中凋亡基因的表达量明显低于未转染的mcf-7,说明上述抑制3β-hsd基因表达的干慢病毒能够降低增塑剂对细胞的损害作用,减缓细胞的凋亡,有望应用于缓解塑化剂毒性的药物中,为塑化剂中毒的治疗提供一种思路。
一实施方式的药物组合物,包括上述抑制3β-hsd基因表达的干扰性shrna、上述抑制3β-hsd基因表达的重组载体、上述抑制3β-hsd基因表达的慢病毒或上述重组工程菌。
可以理解,该药物组合物中还可以包括药学上可接受的辅助剂。
在其中一个实施例中,该药物组合物用于制备治疗3β-hsd基因异常表达相关疾病的药物。
在其中一个实施例中,该药物组合物用于制备缓解塑化剂的细胞毒性的药物。
在其中一个实施例中,该药物组合物用于制备缓解邻苯二甲酸二(2-乙基)己酯的细胞毒性的药物。
上述药物组合物能够用于治疗3β-hsd基因异常表达相关疾病或缓解塑化剂的细胞毒性。
以下为具体实施例部分
以下实施例中,如未特别说明,未注明具体条件的实验方法,通常按照常规条件,例如参见萨姆布鲁克、ef弗里奇、t曼尼阿蒂斯等(金冬雁,黎孟枫等译)所著的分子克隆实验指南[m](北京:科学出版社,1992)中所述的条件或者试剂盒生产厂家推荐的方法实现。实施例中所使用的试剂均为市售。
实施例中所用的材料:限制性内切酶bamhi和限制性内切酶ecori购于thermofish公司,t4dna连接酶购于neb公司,人乳腺癌mcf-7细胞购于上海生命科学院细胞资源中心、293ft细胞、lipofectaminetm2000均购于invitrogen公司,慢病毒表达载体plvx-shrna2p、包膜蛋白质粒pcmv-vsv-g和包装辅助质粒pcmv-dr8.91均购于biovector公司,rneasyminikit购于qiagen公司,lenti-xgostix试剂盒购于takara生物技术公司,rpmi-1640培养基和10%胎牛血清均购于gibco公司,lb培养基和氨苄青霉素均购于生工生物工程(上海)股份有限公司,dehp(纯度大于99%)购于sigma公司。dna胶回收试剂盒购于omega公司。rpmi-1640完全培养基即为含有10%胎牛血清,1%双抗的rpmi-1640培养基。
实施例1
构建抑制3β-hsd基因表达的表达载体
(1)制备干扰性shrna
制备四种干扰性shrna,四种干扰性rna分别为shrna1、shrna2、shrna3及shrnac,其中,shrnac为对照组,shrna1含有如seqidno.1所示的靶序列;shrna2如seqidno.2所示的靶序列;shrna3含有如seqidno.3所示的靶序列;shrnac含有如seqidno.10所示的靶序列。
具体地,合成干扰性shrna的正义链和反义链,将合成的正义链与反义链等量混合,退火形成干扰性shrna。
其中,退火程序为95℃加热30s、72℃加热120s、37℃加热120s、25℃保持120s、4℃保持20min。
其中,shrna1的正义链如seqidno.4所示,shrna1的反义链如seqidno.5所示,shrna2的正义链如seqidno.6所示,shrna2的反义链如seqidno.7所示,shrna3的正义链如seqidno.8所示,shrna3的反义链如seqidno.9所示,shrnac的正义链如seqidno.11所示,shrnac的反义链如seqidno.12所示。上述shrna的正义链和反义链均由上海生工生物工程技术服务有限公司合成。
(2)抑制3β-hsd基因表达的重组载体的构建
按照质粒提取试剂盒的使用说明提取plvx-shrna2p载体,plvx-shrna2p载体的谱图如图1所示。用限制性内切酶bamhi与限制性内切酶ecori双酶切处理plvx-shrna2p载体,电泳鉴定后,用dna胶回收试剂盒回收双酶切产物,得到双酶切后的plvx-shrna2p载体,再用t4dna连接酶分别将shrna1、shrna2、shrna3与shrnac连接到双酶切后的plvx-shrna2p载体中,得到连接产物。将连接产物转化到感受态大肠杆菌jm107中,并将转化后的大肠杆菌jm107涂布到含氨苄青霉素的lb培养基的平板上,于37℃培养14h。分别挑取含有上述重组载体的阳性单克隆的细菌进行分别进行限制性内切酶bamhi与限制性内切酶kpni双酶切处理、限制性内切酶ecori与限制性内切酶kpni双酶切处理,再对双酶切后的产物进行电泳鉴定,酶切鉴定结果如图2所示。
在图2中,m泳道对应于dnamaker;1泳道对应plvx-shrna2p与shrnac的连接产物经转化,然后bamh1、kpni双酶切的产物;2泳道对应于plvx-shrna2p与shrnac的连接产物经转化,然后ecori、kpni双酶切的产物;3泳道对应于plvx-shrna2p与shrna1的连接产物经转化,然后bamh1、kpni双酶切的产物;4泳道对应于plvx-shrna2p与shrna1的连接产物经转化,然后ecori、kpni双酶切的产物;5泳道对应于plvx-shrna2p与shrna2的连接产物经转化,然后bamh1、kpni双酶切的产物;6泳道对应于plvx-shrna2p与shrna2的连接产物经转化,然后ecori、kpni双酶切的产物;7泳道对应于plvx-shrna2p与shrna3的连接产物经转化,然后bamh1、kpni双酶切的产物;8泳道对应于plvx-shrna2p与shrna3的连接产物经转化,然后ecori、kpni双酶切的产物。
plvx-shrna2p具有bamh1酶切位点、kpni酶切位点及ecori酶切位点,但ecori酶切位点与干扰性shrna的ecori的粘性末端结合形成重组载体后,该ecori酶切位点不能再次被ecori酶切开,因此,重组载体在被ecori、kpni双酶切时,只有kpni一个酶切位点被切开,电泳时只出现一条分子量超过5000bp的条带。因此,由图2初步判定shrna1、shrna2、shrna3及shrnac均成功插入plvx-shrna2p。
挑取酶切鉴定后的阳性克隆的细菌进行培养,并将培养后的细菌菌液进行测序鉴定,测序由上海生工生物工程技术服务有限公司进行,测序鉴定结果如图3~6所示。
图3为shrna1与plvx-shrna2p构成的重组载体的部分测序图,shrna1的正义链位于168位~225位。图4为shrna2与plvx-shrna2p构成的重组载体的部分测序图,shrna2的正义链位于164位~221位。图5为shrna3与plvx-shrna2p构成的重组载体的部分测序图,shrna3的正义链位于168位~225位。图6为shrnac与plvx-shrna2p构成的重组载体的部分测序图,shrna3的正义链位于47位~105位。
因此,由图3~6可知,shrna1、shrna2、shrna3及shrnac均成功插入plvx-shrna2p。
将测序正确的菌液进行扩大培养,采用质粒试剂盒提取细菌中的重组载体,分别得到扩增的plvx-shrna2pshrna1重组载体、plvx-shrna2pshrna2重组载体、plvx-shrna2pshrna3重组载体、plvx-shrna2pshrnac重组载体。
实施例2
抑制3β-hsd基因表达的慢病毒的制备
培养293ft细胞,取生长状态良好的细胞接种到六孔板中,每孔106个细胞,获取实施例1提取的plvx-shrna2pshrna1重组载体、plvx-shrna2pshrna2重组载体、plvx-shrna2pshrna3重组载体、plvx-shrna2pshrnac重组载体各2μg,分别将2μg的各上述重组载体、1μg的pcmv-vsv-g和2μg的pcmv-dr8.91共转染到293ft细胞,置于37℃培养至48h和72h,收集上清培养基并用0.45μm滤膜过滤,分别得到含有plvx-shrna2pshrna1重组载体的病毒液、含有plvx-shrna2pshrna2重组载体的病毒液、含有plvx-shrna2pshrna3重组载体的病毒液、含有plvx-shrna2pshrnac重组载体的病毒液,使用lenti-xgostix试剂盒检测各病毒液的病毒滴度。其中,上述重组载体对应的病毒液的病毒滴度为5×107tu~5×108tu。
实施例3
慢病毒转导人乳腺癌mcf-7细胞
培养mcf-7细胞,取生长状态良好的细胞接种到六孔板中,每孔106个细胞,培养12h后细胞汇合度达50%。分别取实施例2获得的各病毒液,均用rpmi-1640完全培养基按10:1稀释后,再加入polybrene(聚凝胺)至终浓度为5μg/ml待用。去除六孔板中原有的培养基,加入含慢病毒的rpmi-1640完全培养基。转染24h后,去除含慢病毒的rpmi-1640完全培养基,加入正常的rpmi-1640完全培养基再培养24h,然后换用0.5μg/ml嘌呤霉素对细胞进行筛选。每隔3天换液一次,每次换液均增加嘌呤霉素的浓度,直至将嘌呤霉素的浓度增加至1.0μg/ml,筛选时间为10天,筛选得到抑制3β-hsd基因表达的mcf-7细胞株,即plvx-shrna2pshrna1沉默细胞、plvx-shrna2pshrna2沉默细胞、plvx-shrna2pshrna3沉默细胞及plvx-shrna2pshrnac沉默细胞。
根据转染前的mcf-7细胞的荧光图分别与plvx-shrna2pshrna1沉默细胞的荧光图、plvx-shrna2pshrna2沉默细胞的荧光图、plvx-shrna2pshrna3沉默细胞的荧光图及plvx-shrna2pshrnac沉默细胞的荧光图对比,计算得到含有plvx-shrna2pshrna1重组载体的病毒、含有plvx-shrna2pshrna2重组载体的病毒、含有plvx-shrna2pshrna3重组载体的病毒、含有plvx-shrna2pshrnac重组载体的病毒的转染率分别为98%、96%、98%、99%。
实施例4
荧光定量pcr检测3β-hsd的mrna表达水平
根据3β-hsd基因的mrna和β-actin基因mrna序列,利用引物设计软件primer5.0和oligo7.0设计pcr引物,引物序列如表1所示。
表1
分别接种正常的mcf-7细胞、plvx-shrna2pshrna1沉默细胞、plvx-shrna2pshrna2沉默细胞、plvx-shrna2pshrna3沉默细胞以及plvx-shrna2pshrnac沉默细胞至六孔板,细胞汇合度达到80%~90%时,用rneasyminikit提取各组细胞的总rna,利用primescriprtreagentkit将mrna逆转录为cdna,逆转录条件:37℃,15min;85℃,5s;4℃,∞。逆转录结束后,加入90μl的rnasefreedh2o稀释cdna,-20℃保存,以便后续检测使用。
取各组细胞的cdna1μl为模板,加入表1中的引物,以β-actin为内参,实时荧光定量pcr检测3β-hsd相对表达量,设置反应条件:95℃30s,1循环;55℃30s,40循环;95℃5s;60℃1min;95℃15s,利用sybrprimescriptrt-pcrkit检测各组细胞3β-hsd基因相对表达量,结果如图7所示。图7中每种沉默细胞3β-hsd基因的表达量与mcf-7细胞3β-hsd基因的表达量的比值,即相应沉默细胞3β-hsd基因的相对表达量。图7中:hsd-3β-sh-1对应于plvx-shrna2pshrna1沉默细胞中3β-hsd相对表达量,hsd-3β-sh-2对应于plvx-shrna2pshrna2沉默细胞中3β-hsd相对表达量,hsd-3β-sh-3对应于plvx-shrna2pshrna3沉默细胞中3β-hsd相对表达量,shrnac对应于plvx-shrna2pshrnac沉默细胞中3β-hsd相对表达量。
由图7可以看出,经含有3β-hsd基因干扰性shrna的病毒转导后的mcf-7细胞,其3β-hsd基因表达明显受到抑制,其中干扰片性shrna1、干扰片性shrna2、干扰片性shrna3的抑制效率分别为61.6%±0.05%、79.4%±0.06%、71.9%±0.08%。而作为对照的干扰片性shrnac对3β-hsd基因表达抑制效率0.049%±0.04%,从而证明了本研究中采用的plvx-shrna2p载体携带shrna1、shrna2或shrna3能特异抑制3β-hsd基因的表达,且抑制效果非常显著。
实施例5
westernblot检测3β-hsd蛋白表达水平
将mcf-7细胞、plvx-shrna2pshrna2沉默细胞接种到25cm2细胞培养瓶中,培养24h至细胞汇合度达70%。分别取上述细胞各1瓶,去除培养基,用冷pbs洗3次,加入200μl的细胞裂解液,并用细胞刮将裂解的细胞快速从瓶壁刮下,收集蛋白至500μl的ep管中,于4℃继续裂解30min,裂解结束后12000rpm、4℃离心20min,收集沉淀,向沉淀中加入5×sds-pagesampleloadingbuffer,100℃变性5min,进行12%sds-聚丙烯酰胺凝胶电泳,接着将蛋白电转至pvdf膜上,5%脱脂奶粉封闭1h。分别加入相应的3β-hsd抗体、β-actin抗体,4℃室温孵育过夜;tbst缓冲液洗膜3次,每10min一次;再加入二抗,室温孵育1h;tbst缓冲液洗膜3次,每10min一次。加入westernblot化学发光试剂后用进行成像分析及条带灰度定量分析,结果如图8及图9所示。
图8中沉默细胞及图9中3β-hsd沉默细胞均指plvx-shrna2pshrna2沉默细胞。
由图8及图9可以看出,携带3β-hsd基因干扰性shrna的病毒的转导mcf-7细胞中3β-hsd基因的蛋白质表达明显受抑制,其中干扰片段shrna2的抑制效果最高。而对照干扰性shrnac组细胞3β-hsd基因表达几乎没变化,以上结果和qpcr检测的结果相符合,也说明本研究中采用的plvx-shrna2p载体携带shrna能特异抑制3β-hsd基因的表达。
实施例6
传代培养
连续培养plvx-shrna2pshrna1沉默细胞、plvx-shrna2pshrna2沉默细胞、plvx-shrna2pshrna3沉默细胞以及plvx-shrna2pshrnac沉默细胞20代后,分别提取各组沉默型细胞,westernblotting分析3β-hsd基因表达量,结果如图10所示。图10中,1对应于plvx-shrna2pshrna1沉默细胞、2对应于plvx-shrna2pshrna2沉默细胞、3对应于plvx-shrna2pshrna3沉默细胞,4对应于plvx-shrna2pshrnac沉默细胞。
由图10可以看出,plvx-shrna2pshrna1沉默细胞、plvx-shrna2pshrna2沉默细胞及plvx-shrna2pshrna3沉默细胞都能有效的抑制3β-hsd基因的表达,而plvx-shrna2pshrnac沉默细胞对3β-hsd基因表达几乎没有影响。说明本发明构建的shrna慢病毒表达载体能够持续、高效的抑制3β-hsd的表达。
实施例7
抑制3β-hsd基因表达对凋亡基因的影响
(1)将mcf-7细胞、实施例3制备的plvx-shrna2pshrna2沉默细胞分别分接种到六孔板中,待细胞汇合度达70%时进行不同浓度的dehp处理。dehp终浓度分别为0mmol/l、0.05mmol/l、0.1mmol/l、0.2mmol/l、0.4mmol/l、0.8mmol/l,作用时间为24h。
(2)参见实施例4荧光定量pcr检测3β-hsd基因表达量的方法分别测定dehp处理后的各组细胞中bax、caspase-3和caspase-8基因表达水平的改变。具体地,用rneasyminikit提取各组细胞的总rna,利用primescriprtreagentkit将mrna逆转录为cdna,逆转录条件:37℃,15min;85℃,5s;4℃下保存。反转录结束后,加入90μl的rnasefreedh2o稀释cdna,-20℃保存,以便后面检测使用。
取各组细胞的cdna1μl为模板,加入表2中3β-hsd基因的引物,实时荧光定量pcr检测3β-hsd基因相对表达量,设置反应条件:95℃30s,1循环;55℃30s,40循环;95℃5s;60℃1min;95℃15s,利用sybrprimescriptrt-pcrkit检测各组细胞中bax基因、caspase-3基因和caspase-8基因相对表达量。其中,bax基因、caspase-3基因caspase-8基因及内参gapdh基因的pcr引物的序列如表2所示。
表2
在不同浓度的dehp条件下,各组细胞的凋亡基因中bax、caspase-3和caspase-8的表达水平测定结果请分别参见图11~13;图11~13中3β-hsd沉默细胞表示plvx-shrna2pshrna2沉默细胞。
从图11可以看出,dehp处理24h后,处理后的mcf-7细胞的bax基因表达水平高于未经处理的mcf-7细胞的bax基因表达水平,差异具有统计学意义(p<0.05或p<0.01)。说明dehp处理对细胞产生损害,激发细胞凋亡;随着dehp剂量的增加,被处理的mcf-7细胞中bax基因表达水平逐渐升高。在不同剂量dehp处理时,plvx-shrna2pshrna2沉默细胞的bax基因表达水平明显低于同样剂量下dehp处理的mcf-7细胞中bax基因表达水平,差异具有统计学意义(p<0.05或p<0.01)。
从图12可以看出,dehp处理24h后,处理后的mcf-7细胞的caspase-3基因表达水平高于未经dehp处理的mcf-7细胞的caspase-3基因表达水平,差异具有统计学意义(p<0.05或p<0.01)。说明dehp处理对细胞产生损害,激发细胞凋亡;随着dehp剂量的增加,被处理的mcf-7细胞中caspase-3基因表达水平逐渐升高。在不同剂量dehp处理时,plvx-shrna2pshrna2沉默细胞中caspase-3基因表达水平明显低于同样剂量下dehp处理的mcf-7细胞中caspase-3基因表达水平,差异具有统计学意义(p<0.05或p<0.01)。
从图13可以看出,dehp处理24h后,处理后的mcf-7细胞的caspase-8基因表达水平高于未经dehp处理的mcf-7细胞的caspase-8基因表达水平,差异具有统计学意义(p<0.05或p<0.01)。说明dehp处理对细胞产生损害,激发细胞凋亡;随着dehp剂量的增加,被处理的mcf-7细胞中caspase-8基因表达水平逐渐升高。在不同剂量dehp处理时,plvx-shrna2pshrna2沉默细胞中caspase-8基因表达水平明显低于同样剂量下dehp处理的mcf-7细胞中caspase-8基因表达水平,差异具有统计学意义(p<0.05或p<0.01)。
实施例8
抑制3β-hsd基因表达对凋亡基因的蛋白表达的影响
参见实施例5中westernblot检测3β-hsd蛋白表达水平的方法分别测定不同浓度dehp处理后的mcf-7细胞及plvx-shrna2pshrna2沉默细胞细胞中bax、caspase-3和caspase-8的蛋白表达水平的改变。
具体地,用冷pbs洗上述各组细胞,洗3次,加入200μl的细胞裂解液,并用细胞刮将裂解的细胞快速从瓶壁刮下,收集蛋白至500μl的ep管中,于4℃继续裂解30min,裂解结束后12000rpm、4℃离心20min,收集沉淀,向沉淀中加入5×sds-pagesampleloadingbuffer于100℃变性5min,进行12%sds-聚丙烯酰胺凝胶电泳,接着将蛋白电转至pvdf膜上,5%脱脂奶粉室温封闭1h。分别加入相应的凋亡基因抗体、gapdh抗体,4℃室温孵育过夜;tbst缓冲液洗膜3次,每10min一次;再加入二抗,室温孵育1h;tbst缓冲液洗膜3次,每10min一次。加入westernblot化学发光试剂后进行成像分析及条带灰度定量分析。
在不同浓度的dehp条件下,各组细胞的凋亡基因bax、caspase-3和caspase-8的蛋白表达水平测定结果请分别参见图14~16;图14~16中mcf-7细胞表示未经处理的正常mcf-7细胞;图14~16中3β-hsd表示plvx-shrna2pshrna2沉默细胞。
在图14中,a1部分为mcf-7细胞中bax蛋白表达的条带图,b1部分为plvx-shrna2pshrna2沉默细胞的bax蛋白表达条带图,c1部分为mcf-7细胞与plvx-shrna2pshrna2沉默细胞中bax蛋白表达含量对比图。
从图14可以看出,dehp处理24h后,处理后的mcf-7细胞的bax蛋白表达水平高于未经dehp处理的mcf-7细胞的bax蛋白表达水平,差异具有统计学意义(p<0.05或p<0.01)。说明dehp处理对细胞产生损害,激发细胞凋亡;随着dehp剂量的增加,处理后的mcf-7细胞中bax蛋白表达水平逐渐升高。在不同剂量dehp处理时,plvx-shrna2pshrna2沉默细胞中bax蛋白表达水平明显低于同样剂量下dehp处理的mcf-7细胞中bax蛋白表达水平,差异具有统计学意义(p<0.05或p<0.01)。
在图15中,a2部分为mcf-7细胞中caspase-3蛋白表达的条带图,b2部分为plvx-shrna2pshrna2沉默细胞的caspase-3蛋白表达条带图,c1部分为mcf-7细胞与plvx-shrna2pshrna2沉默细胞中caspase-3蛋白表达含量对比图。
从图15可以看出,dehp处理24h后,处理后的mcf-7细胞的caspase-3蛋白表达水平高于未经dehp处理的mcf-7细胞的caspase-3蛋白表达水平,差异具有统计学意义(p<0.05或p<0.01)。说明dehp处理对细胞产生损害,激发细胞凋亡;随着dehp剂量的增加,处理后的mcf-7细胞中caspase-3蛋白表达水平逐渐升高。在不同剂量dehp处理时,plvx-shrna2pshrna2沉默细胞中caspase-3蛋白表达水平明显低于同样剂量下dehp处理的mcf-7细胞中bax蛋白表达水平,差异具有统计学意义(p<0.05或p<0.01)。
在图16中,a3部分为mcf-7细胞中caspase-8蛋白表达的条带图,b3部分为plvx-shrna2pshrna2沉默细胞的caspase-8蛋白表达条带图,c3部分为mcf-7细胞与plvx-shrna2pshrna2沉默细胞中caspase-8蛋白表达含量对比图。
从图16可以看出,dehp处理24h后,处理后的mcf-7细胞的caspase-8蛋白表达水平高于未经dehp处理的mcf-7细胞的caspase-8蛋白表达水平,差异具有统计学意义(p<0.05或p<0.01)。说明dehp处理对细胞产生损害,激发细胞凋亡;随着dehp剂量的增加,处理后的mcf-7细胞中caspase-8蛋白表达水平逐渐升高。在不同剂量dehp处理时,plvx-shrna2pshrna2沉默细胞中caspase-8蛋白表达水平明显低于同样剂量下dehp处理的mcf-7细胞中caspase-8蛋白表达水平,差异具有统计学意义(p<0.05或p<0.01)。
以上所述实施例的各技术特征可以进行任意的组合,为使描述简洁,未对上述实施例中的各个技术特征所有可能的组合都进行描述,然而,只要这些技术特征的组合不存在矛盾,都应当认为是本说明书记载的范围。
以上所述实施例仅表达了本发明的几种实施方式,其描述较为具体和详细,但并不能因此而理解为对发明专利范围的限制。应当指出的是,对于本领域的普通技术人员来说,在不脱离本发明构思的前提下,还可以做出若干变形和改进,这些都属于本发明的保护范围。因此,本发明专利的保护范围应以所附权利要求为准。
序列表
<110>深圳市疾病预防控制中心
深圳市卫生检验中心
深圳市预防医学研究所
<120>特异抑制3β-hsd基因表达的shrna重组载体构建与应用
<160>24
<170>siposequencelisting1.0
<210>1
<211>19
<212>dna
<213>人工序列(artificialsequence)
<400>1
ggatgacagagatgtgcat19
<210>2
<211>19
<212>dna
<213>人工序列(artificialsequence)
<400>2
gtccaagactcagtgattt19
<210>3
<211>20
<212>dna
<213>人工序列(artificialsequence)
<400>3
gtggccaacttattgtattc20
<210>4
<211>58
<212>dna
<213>人工序列(artificialsequence)
<400>4
gatccggatgacagagatgtgcatttcaagagaatgcacatctctgtcatcctttttt58
<210>5
<211>58
<212>dna
<213>人工序列(artificialsequence)
<400>5
aattaaaaaaggatgacagagatgtgcattctcttgaaatgcacatctctgtcatccg58
<210>6
<211>58
<212>dna
<213>人工序列(artificialsequence)
<400>6
gatccgtccaagactcagtgatttttcaagagaaaatcactgagtcttggactttttt58
<210>7
<211>58
<212>dna
<213>人工序列(artificialsequence)
<400>7
aattaaaaaagtccaagactcagtgattttctcttgaaaaatcactgagtcttggacg58
<210>8
<211>59
<212>dna
<213>人工序列(artificialsequence)
<400>8
gatccgtggccaacttattgtattcttcaagagagaatacaataagttggccatttttt59
<210>9
<211>59
<212>dna
<213>人工序列(artificialsequence)
<400>9
gatccgtggccaacttattgtattcttcaagagagaatacaataagttggccatttttt59
<210>10
<211>19
<212>dna
<213>人工序列(artificialsequence)
<400>10
gctagttcgagattcgaat19
<210>11
<211>58
<212>dna
<213>人工序列(artificialsequence)
<400>11
gatccgctagttcgagattcgaatttcaagagaattcgaatctcgaactagctttttt58
<210>12
<211>58
<212>dna
<213>人工序列(artificialsequence)
<400>12
aattaaaaaagctagttcgagattcgaattctcttgaaattcgaatctcgaactagcg58
<210>13
<211>20
<212>dna
<213>人工序列(artificialsequence)
<400>13
cgggcatctatggtgaaggt20
<210>14
<211>20
<212>dna
<213>人工序列(artificialsequence)
<400>14
cacgtgcatccaagccacat20
<210>15
<211>20
<212>dna
<213>人工序列(artificialsequence)
<400>15
tcctgcaccaccaactgctt20
<210>16
<211>20
<212>dna
<213>人工序列(artificialsequence)
<400>16
gtcttctgggtggcagtgat20
<210>17
<211>20
<212>dna
<213>人工序列(artificialsequence)
<400>17
gccagcaaactggtgctcaa20
<210>18
<211>21
<212>dna
<213>人工序列(artificialsequence)
<400>18
atgtccagcccatgatggttc21
<210>19
<211>25
<212>dna
<213>人工序列(artificialsequence)
<400>19
gactctggaatatccctggacaaca25
<210>20
<211>22
<212>dna
<213>人工序列(artificialsequence)
<400>20
aggtttgctgcatcgacatctg22
<210>21
<211>22
<212>dna
<213>人工序列(artificialsequence)
<400>21
caaatgcaaactggatgatgac22
<210>22
<211>21
<212>dna
<213>人工序列(artificialsequence)
<400>22
agcaggctcttgttgatttgg21
<210>23
<211>20
<212>dna
<213>人工序列(artificialsequence)
<400>23
tcctgcaccaccaactgctt20
<210>24
<211>20
<212>dna
<213>人工序列(artificialsequence)
<400>24
gtcttctgggtggcagtgat20
- 还没有人留言评论。精彩留言会获得点赞!