一种生长素调控蛋白、编码基因及应用的制作方法
本发明涉及植物基因工程领域,更具体的说,涉及一种生长素调控蛋白oscole1、编码基因及应用。
背景技术:
生长素(iaa)作为最重要的植物激素之一,在植物的生长发育过程中发挥着重要作用。生长素转运过程作为生长素相关研究中最热门的组成部分之一,在生长素实现其生理效应的过程中发挥着决定性的作用。相比于在生长素极性运输机理及生理作用研究方面取得的大量成果,生长素在细胞内的转运过程鲜有研究涉及,使得人们对于这一重要的植物生理过程仍知之甚少。
生长素最明显的作用是促进生长,但对茎、芽、根生长的促进作用因浓度而异。三者的最适浓度是茎>芽>根,大约分别为每升10-5摩尔、10-8摩尔、10-10摩尔。植物体内吲哚乙酸的运转方向表现明显的极性,主要是由上而下。生长素活化氢离子泵,降低质膜外的ph值,还大大提高细胞壁的弹性和可塑性,从而使细胞壁变松,并提高吸水力。生长素还有促进愈伤组织形成和诱导生根的作用。
在植物体内吲哚乙酸可与其它物质结合而失去活性,可与天冬氨酸结合为吲哚乙酰天冬氨酸,与肌醇结合成吲哚乙酸肌醇,与葡萄糖结合成葡萄糖苷,与蛋白质结合成吲哚乙酸-蛋白质络合物等。结合态吲哚乙酸常可占植物体内吲哚乙酸的50-90%,可能是生长素在植物组织中的一种储藏形式,它们经水解可以产生游离吲哚乙酸。
生长素有多方面的生理效应,这与其浓度有关。低浓度时可以促进生长,高浓度时则会抑制生长,甚至使植物死亡。生长素可刺激形成层细胞分裂;促进木质部、韧皮部细胞分化,促进插条发根、调节愈伤组织的形态建成。 吲哚乙酸可造成顶端优势,延缓叶片衰老;生长素控制幼苗中胚轴伸长的可逆性红光抑制,当吲哚乙酸转移至枝条下侧即产生枝条的向地性,而吲哚乙酸转移至枝条的背光侧产生枝条的向光性;生长素可以促进开花,诱导单性果实的发育,延迟果实成熟等。
生长素极性运输过程的实现,离不开在植物体内存在的多种生长素转运体。这些转运体或通过极性定位分布于细胞膜的顶部或基部,或非极性地分布于细胞膜的两侧,通过介导生长素的单向输入或输出细胞,实现生长素向特定组织器官中的运输。就已有的生长素转运体的研究成果来看,共有3类生长素转运体参与介导了生长素的极性运输过程:pin-formed蛋白质家族转运体(pins),atp结合盒b家族蛋白质转运体(atp-bindingcassette(abc)-b),auxin1/like-aux1蛋白质家族转运体(auxs/laxs),且这3类转运体在双子叶植物拟南芥与单子叶植物水稻中均同源存在。植物体内除了内质网具有调控细胞内生长素动态平衡的作用外,近年来在拟南芥细胞中中,液泡也因被发现其含有大量的生长素代谢产物及具有膜定位的生长素转运体,而被认为同样参与了细胞内的生长素代谢调控过程。被鉴定的定位于液泡膜上的生长素转运体为wallarethin1(atwat1),该蛋白质在二级结构上与atpin5具有高度的相似性。生长素转运功能的鉴定表明:atwat1可以介导液泡内的iaa转运至细胞质中,但其在细胞内的转运过程可能需要相因的调控体进行调控。
此外,该蛋白质在决定植株基部自由iaa含量,细胞壁中次生壁厚度,植株高度及植物的维管束病菌抗性上具有重要作用。atwat1的发现,暗示在植物细胞内存在广泛的细胞器参与的细胞内生长素动态平衡调节过程,且该种生长素调控模式可能在进化上要早于存在于细胞与细胞之间的生长素极性运输过程。
技术实现要素:
本发明涉及到一个调控植物体内iaa含量的蛋白oscole1以及编码该蛋白的基因oscole1,该基因在水稻中的抑制表达导致植物体内的生长素含 量降低,植株变矮,叶夹角变小,开花提前,而过表达导致水稻生长素含量增加,生长期延长,叶夹角变大。因此该基因可用于水稻株型及花期的调控。
本发明所提供的蛋白,名称为oscole1,来源于水稻-日本晴,属于调控蛋白类。
本发明采用rt-pcr扩增的方法,获得了oscole1基因的cds序列。其cdna序列全长为822bp(序列1),编码产生长度为273氨基酸的蛋白质序列(序列2)。
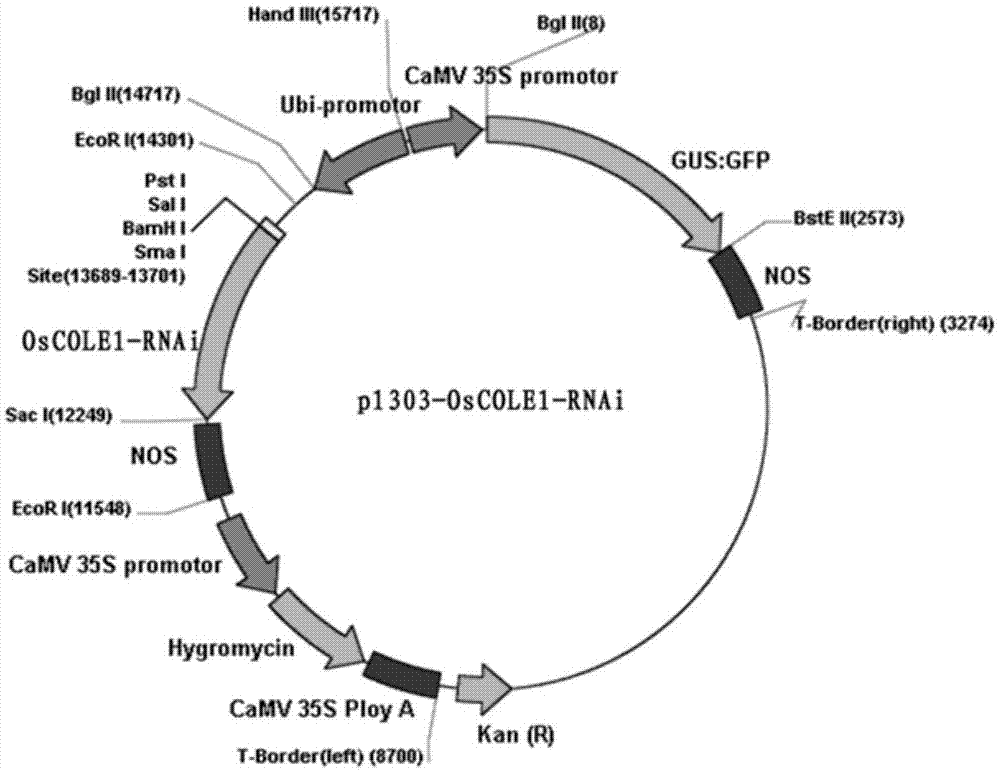
本发明把oscole1基因构建到植物表达载体上,启动子为ubiqutin,分别得到oscole1过表达(图1)及抑制表达rnai质粒(图2),将其转化到农杆菌eha105中,用农杆菌转化水稻并获得了转基因植株。
通过对转基因植株与未转基因的野生型植株在正常生长发育条件下的比较,结果表明,oscole1过表达导致水稻生长素含量增加,植株变大,生长期变长。oscole1抑制表达可降低胞内生长素含量,植株变矮,叶夹角变小,开花提前(图3)。进一步分析表明水稻的第一、第二节的长度受到明显影响(图4)。以上结果表明oscole1参与了植物株高、株型和开花的调控。
附图说明
为了更清楚地说明本发明实施例中的技术方案,下面将对实施例描述中所需要使用的附图作简单地介绍。附图中:
图1为oscole1过表达载体的构建图谱;
图2为oscole1rnai载体的构建图谱;
图3为oscole1转基因水稻的表型分析;
图4为oscole1转基因水稻第1、2、3、4节长度统计。
具体实施方式
下面将结合实施例,对本发明实施例中的技术方案进行清楚、完整地描述。
实施例1oscole1基因cdna的克隆
1.1rna的提取及dna的去除:
1)取约200mg日本晴幼苗为材料,液氮研磨,用trizol试剂(invitrogen)提取rna。
2)将rna溶于85μl水中,加入10×buffer10μl和5μlrq1rnafreednase(1u/μl),37℃,15min,以去除dna的污染。
3)加入100μl的酚氯仿抽提一次,取上清,用等体积的异丙醇沉淀rna,70%的乙醇洗一次,溶于50μl的水中。
4)定量rna的浓度为1μg/μl。
1.2逆转录:
按照promega公司的goscripttmreversetranscriptionsystem(promegaa5000)试剂盒进行,反应程序如下:
1)依次加入下列试剂,混匀后进行反应:
2)上述反应结束后,加入下列试剂继续进行反应:
3)最后加入m-mlv(200u/μl)1μl;42℃,50min;70℃,15min灭活,备用。
1.3pcr扩增:
1)pcr引物如下:
oscole1-oe-f:
5’-gtcgactctagaggatccaccatgggcgacgagaagtctccgc-3’;
oscole1-oe-r:
5’-atggtctttgtagtcggtaccgcttctgcttgctcctgttctg-3’。
2)扩增条件:
把扩增出dna片段从琼脂糖胶中回收,利用重组系统将pcr产物重组到载体pcambia1303上(bamhⅰ和sacⅰ线性化),图1为oscole1过表达载体的构建图谱,转化e.coli,酶切鉴定后测序;
测序结果表明:得到cdna为序列1所示的基因,将该基因命名为oscole1,全长822bp,具体序列如下:
atgggcgacgagaagtctccgctgagccagatgggcagccgcgaccgggaccgggagctcctcatccccgtctccggcggcggctcggcccccggagacggcgacggggacggcgacagggcggcctcctcctccgcgtccgccgctctgtcctcctccagccgcgaggcttttcataaggtggtccgaagttgggcttcaaagaaattcatgactggatgtgtaattctcttccccatagcaataacattctacatcacatggtggttcattcattttgttgacggattcttctcgccaatctatgctcaactgggaatcaacatgtttggtcttggctttatcacttctgttaccttcatatttgtggttggagtcttcatgtcatcttgggttggagcatctgtccttagccttggtgagtggattattaagcgcatgccacttgtccgtcatatctacaatgcatccaagcaaataagtgctgcaatatcaccagatcagaacaaacaggcattcaaggaagtggtcatcataaggcatcctcgtattggagaatatgcatttggtttcattacatcatcagtctctctccagagttatactggtcaagaggaactgtactgcgtgtatgtcccaaccaaccatctttatattggtgatatcttcatggtaaattcgaaggatgttataaggccaaatctttctgtccgtgaaggcatcgaaattgttgtatctggtggcatgtcgatgc cccaaattctatcaactctggacccacagacaattcttggagacagaacaggagcaagcagaagctga
其编码的蛋白的氨基酸序列如序列2所示,编码273个氨基酸(末端终止子未计入,即序列中***号未计入),其编码的蛋白的氨基酸序列如下:
1.4rnai载体的构建:
以该cdna为模板进行rt-pcr扩增,rnai载体构建的发卡rna设计为正向链的287到815bp,以及互补链的99到532bp。
1)pcr引物如下:
oscole1-ri-f1:
5’-gcaggtcgactctagaggatccattcttctcgccaatctatgc-3’;
oscole1-ri-r1:
5’-ggtaccagcgctatcgctgcttgctcctgttctgtct-3’;
oscole1-ri-f2:
5’-cgatagcgctggtaccacaacaatttcgatgccttca-3’;
oscole1-ri-r2:
5’-acgatcggggaaattcgagctcattcttctcgccaatctatgc-3’。
2)扩增条件是:
把扩增出的dna片段从琼脂糖胶中回收,利用重组系统将pcr产物重组到载体pcambia1303上(bamhⅰ和sacⅰ线性化),图2为oscole1rnai载体的构建图谱,转化e.coli,酶切鉴定后测序。
实施例2转基因水稻的获得
2.1水稻成熟胚愈伤组织的诱导培养
将成熟的水稻日本晴种子去除颖壳后,置于75%的乙醇中浸泡1-2min。去除乙醇后,加入0.1%的升汞溶液浸泡种子30min(期间定时摇晃)。去除升汞溶液,以无菌水冲洗灭菌种子3-4次。将种子置于无菌滤纸上吸干水分后,转移至成熟胚愈伤诱导培养基上,26℃条件下暗培养约2周。将成熟胚盾片长出的愈伤组织小心取下,转移到成熟胚继代培养基上。在相同条件下继代培养。愈伤组织每两周继代培养一次。挑选继代培养5-7d且色泽淡黄的愈伤组织用于共培养。
2.2农杆菌转化子侵染液的制备
将含有相应转化载体的eha105农杆菌在含有50mg/l利福平及卡那霉素的yeb固体培养基上划线,28℃培养3d。收集培养基上的农杆菌菌体,悬浮于共培养液体培养基中。调整菌体浓度至od600=0.5后,加入乙酰丁香酮溶液至终浓度为100mm。即得到用于转化水稻的农杆菌侵染液。
2.3水稻愈伤组织与农杆菌的共培养
选取生长状态好的愈伤组织,置于无菌三角瓶中。加入适量农杆菌侵染液刚好浸没愈伤组织。室温放置20min,期间不断晃动三角瓶。倒掉菌液后,用无菌滤纸吸去愈伤组织上的多余菌液,转移愈伤组织到铺有一层无菌滤纸 的固体共培养基上,26℃黑暗培养2-3d。
2.4抗性愈伤组织的筛选
将共培养后的愈伤组织转移到含有50μg/ml潮霉素的筛选培养基上,26℃暗培养两周。而后将愈伤组织转到新的筛选培养基上,继续在相同条件下筛选培养两周。
2.5抗性愈伤组织的选取及分化
从经两轮筛选后的愈伤组织中,挑取乳黄色致密的抗性愈伤组织。转移抗性愈伤组织至含有50μg/ml潮霉素的分化培养基上。26℃暗培养3d,再转移到26℃,每天15h光照的条件下培养。经过20d左右,抗性愈伤组织上出现绿点,30-40d后进一步分化出小苗。
2.6抗性愈伤组织生根、壮苗和移栽
当抗性愈伤组织分化的芽长至约2cm时,转移小苗至生根培养基上,培养两周左右后,选择叶鞘及根系均发育茁壮的转化苗,用于移栽及转化鉴定。
实施例3转基因水稻的鉴定
3.1dna的提取:
1)剪取100mg转基因材料叶片于液氮中速冻,转移到盛有液氮的研钵中研磨粉碎,将粉末转至1.5ml离心管中;
2)加入600μlsds溶液,颠倒混匀,65℃水浴中放置60min,每隔15min颠倒混匀一次);
3)加入等体积氯仿:异戊醇(v:v=24:1),轻轻颠倒混匀,静置5min后,12000rpm离心15min;
4)转移上清液到另一新离心管中,加入等体积的异丙醇,颠倒混匀,静置5min,12000rpm离心15min;
5)弃上清,以75%乙醇洗涤沉淀,干燥,加入30μl无菌水溶解dna(无菌水中加rnase以去除rna),4℃保存备用;
6)取1μl提取dna电泳检测提取浓度和质量。
3.2pcr鉴定:
1)以提取的转基因材料dna为模板,进行pcr鉴定,pcr引物如下:
oscole1fw:5’-actggtcaagaggaactgtac-3’;
nosrv:5’-gaccggcaacaggattcaatc-3’。
2)扩增条件如下:
共获得oscole1过表达阳性植株20株;rnai植株10个。
实施例4转基因水稻性状考察
4.1株高统计:
各类转基因材料选取灌浆期的两个独立转化事件,每个转化事件随机选取5株,所测定的株高为第一节间与根部连接处到最高稻穗顶部处的长度,选取典型材料进行拍照,oscole1转基因水稻的表型分析如图3。
4.2节间长度统计:
各类转基因材料选取灌浆期的一个独立转化事件,每个转化事件随机选取5株,每株选取株高最高的4个分蘖。图4为oscole1转基因水稻第1、2、3、4节长度统计,p1302ck为对照;oscole1-rnai-1为oscole1基因抑制表达植株;oscole1-oe-1为oscole1基因过表达植株。如图4所示,按从下至上分别为第1、2、3、4节间的顺序,对节间的长度进行测量,结果表明oscole1过表达植株第1、2节的长度明显大于对照,而oscole1抑制表达的植株第1、2节的长度明显短于对照。
实施例5转基因水稻iaa含量测定
选取灌浆期转基因材料中差异最大的茎秆第一、二节间进行iaa含量的测定,具体方法如下:
1)剪取的茎秆材料,使用液氮进行研磨粉碎。液氮环境下,准确称取200mg的样品,加入含有内标2h2-iaa(cdnisotopes)及抗氧化剂的甲醇中,-20℃过夜萃取;
2)萃取后的混合物在18000rpm,4℃的条件下离心15min。收集得到的上清液,利用氮气吹干,残余物以5%的氨溶液溶解;
3)得到的溶解粗产物,利用通过4ml甲醇,4ml水,以及4ml5%的氨溶液进行预处理的oasismaxspe柱进行纯化。当溶解粗产物被加至spe柱后,依次利用4ml5%的氨溶液,4ml水,4ml甲醇进行洗涤,得到的iaa被溶于含有5%甲酸的甲醇溶液中;
4)iaa溶液在氮气吹干后,最终使用水/甲醇(v:v=20:80)的溶液溶解,以用于uplc系统的分析。
测定结果表明,oscole1过表达植株中iaa的含量达到了4.4ng/g鲜重,显著高于转空载体的对照2.1ng/g;rnai植株则为2.1ng/g鲜重,明显低于。
显然,上面描述的实施例仅仅是本发明一部分实施例,而不是全部的实施例。基于本发明中的实施例,本领域普通技术人员在没有做出创造性劳动的前提下所获得的所有其他实施例,都属于本发明保护的范围。
- 还没有人留言评论。精彩留言会获得点赞!
- 蝎神经毒素AaIT及其编码基因与应用的制造方法与工艺
- 腈水解酶突变体及其编码基因和应用的制造方法与工艺
- 植物抗旱相关蛋白AsTMK1及其编码基因和应用的制造方法与工艺
- 植物抗旱相关蛋白EtSnRK2.2及其编码基因和应用的制造方法与工艺
- OsSAPK7蛋白及其编码基因在提高水稻白叶枯病抗性中的应用的制造方法与工艺
- 一种抗除草剂抗虫融合基因、编码蛋白及应用的制造方法与工艺
- 毛果杨PtrZFP103基因及其编码蛋白和应用的制造方法与工艺
- 大豆bHLH转录因子基因GmFER及其编码蛋白和应用的制造方法与工艺
- 一种花狭口蛙的基因及其编码的多肽和该多肽的用途的制造方法与工艺
- 一种泽陆蛙基因及其编码的多肽和该多肽的用途的制造方法与工艺
- 降低种子重量的白桦AP2基因及其编码蛋白的制造方法与工艺
- 可编码ArtRD蛋白基因及其蛋白和应用的制造方法与工艺
- 蝎神经毒素AaIT及其编码基因与应用的制造方法与工艺
- 腈水解酶突变体及其编码基因和应用的制造方法与工艺
- 植物抗旱相关蛋白AsTMK1及其编码基因和应用的制造方法与工艺
- 植物抗旱相关蛋白EtSnRK2.2及其编码基因和应用的制造方法与工艺
- OsSAPK7蛋白及其编码基因在提高水稻白叶枯病抗性中的应用的制造方法与工艺
- 一种抗除草剂抗虫融合基因、编码蛋白及应用的制造方法与工艺
- 毛果杨PtrZFP103基因及其编码蛋白和应用的制造方法与工艺
- 大豆bHLH转录因子基因GmFER及其编码蛋白和应用的制造方法与工艺
- 腈水解酶突变体及其编码基因和应用的制造方法与工艺
- 植物抗旱相关蛋白AsTMK1及其编码基因和应用的制造方法与工艺
- 植物抗旱相关蛋白EtSnRK2.2及其编码基因和应用的制造方法与工艺
- OsSAPK7蛋白及其编码基因在提高水稻白叶枯病抗性中的应用的制造方法与工艺
- 一种抗除草剂抗虫融合基因、编码蛋白及应用的制造方法与工艺
- 毛果杨PtrZFP103基因及其编码蛋白和应用的制造方法与工艺
- 大豆bHLH转录因子基因GmFER及其编码蛋白和应用的制造方法与工艺
- 一种花狭口蛙的基因及其编码的多肽和该多肽的用途的制造方法与工艺
- 一种泽陆蛙基因及其编码的多肽和该多肽的用途的制造方法与工艺
- 一种水稻MIT1基因、其编码蛋白及应用的制造方法与工艺
- 植物抗旱相关蛋白EtSnRK2.2及其编码基因和应用的制造方法与工艺
- OsSAPK7蛋白及其编码基因在提高水稻白叶枯病抗性中的应用的制造方法与工艺
- 一种抗除草剂抗虫融合基因、编码蛋白及应用的制造方法与工艺
- 毛果杨PtrZFP103基因及其编码蛋白和应用的制造方法与工艺
- 大豆bHLH转录因子基因GmFER及其编码蛋白和应用的制造方法与工艺
- 一种水稻MIT1基因、其编码蛋白及应用的制造方法与工艺
- 蛋白StTrxF、编码基因及其在提高植物淀粉含量中的应用的制造方法与工艺
- 蛋白质及其编码基因在调控植物抗黄萎病性中的应用的制造方法与工艺
- 甘薯耐逆相关蛋白IbFBA2、编码基因、重组载体与应用的制造方法与工艺
- 植物抗旱相关蛋白ZmNAC111及其编码基因与应用的制造方法与工艺
- 植物抗旱相关蛋白EtSnRK2.2及其编码基因和应用的制造方法与工艺
- OsSAPK7蛋白及其编码基因在提高水稻白叶枯病抗性中的应用的制造方法与工艺
- 一种抗除草剂抗虫融合基因、编码蛋白及应用的制造方法与工艺
- 毛果杨PtrZFP103基因及其编码蛋白和应用的制造方法与工艺
- 大豆bHLH转录因子基因GmFER及其编码蛋白和应用的制造方法与工艺
- 一种水稻MIT1基因、其编码蛋白及应用的制造方法与工艺
- 蛋白StTrxF、编码基因及其在提高植物淀粉含量中的应用的制造方法与工艺
- 植物抗旱相关蛋白ZmNAC111及其编码基因与应用的制造方法与工艺
- 核盘菌凝集素SSL‑6蛋白及其编码基因与应用的制造方法与工艺
- 植物抗病毒相关蛋白GmSN1及其编码基因和应用的制造方法与工艺