一种L‑阿拉伯糖异构酶及其应用的制作方法
本发明属于生物工程技术领域,具体涉及一种L-阿拉伯糖异构酶及其应用。
背景技术:
D-塔格糖是一种新型功能性甜味剂,可广泛应用于食品、保健品、糖尿病药物和化学合成等行业。D-塔格糖最早发现于乔木的树胶中,也微量的存在于日常饮品如牛奶和可可中。由于D-塔格糖在人体健康方面展现出良好的应用前景,近年来D-塔格糖产业化进程迅猛,市场发展态势良好。
D-塔格糖的商业化生产长期以来依赖化学催化法,但化学催化法存在一些固有缺陷,比如剧烈的碱性环境会导致副产物形成(如山梨糖、甘露糖等)、酸和碱的使用增加了工艺下游的污水处理难度。近年来,研究人员一直在开发条件更为温和的生物法来生产D-塔格糖,以降低工艺成本和满足环境保护上的需求。
目前,生物法生产D-塔格糖最为有效的途径是依赖L-阿拉伯糖异构酶(L-AI)作为催化剂,将D-半乳糖转化为D-塔格糖。该方法所需的反应条件温和,副产物少,符合绿色生产的理念。由于L-阿拉伯糖与D-半乳糖在构象上非常相似,L-AI既能催化L-阿拉伯糖向L-核酮糖的转化,也能将D-半乳糖异构化为D-塔格糖,但其对L-阿拉伯糖的催化效率远高于对D-半乳糖催化效率,这限制了其在D-塔格糖工业化生产上的应用。此外,以往研究认为反应温度越高(95℃以内),对D-塔格糖的生产越有利,因此,研究者更多的关注于来自嗜热菌和极端嗜热菌的L-AI,这些L-AI的最适温度往往在65℃以上,但高温会导致褐变反应发生,从而影响D-塔格糖的品质。
利用以计算机同源建模为指导的基因定点突变技术可以实现对目标酶性质的快速改变。本发明中利用基因定点突变技术对来源于凝结芽孢杆菌(Bacillus coagulans)的L-AI进行改造,不仅增强了其催化D-半乳糖的活性,也将其最适催化温度由60℃降低为50℃。将该突变酶外源表达于大肠杆菌中,以全细胞催化体系进行D-半乳糖的异构化,大大提高了D-塔格糖产率和转化率,从而有效降低生产成本。
技术实现要素:
本发明要解决的技术问题是提供定点突变L-阿拉伯糖异构酶F279I。
本发明还要解决的技术问题是提供一株表达定点突变L-阿拉伯糖异构酶F279I的大肠杆菌。
本发明还要解决的技术问题是提供上述大肠杆菌的构建方法。
本发明最后要解决的技术问题是提供上述大肠杆菌在催化合成D-塔格糖中的应用。
为解决以上技术问题,本发明采用的技术方案如下:
一种L-阿拉伯糖异构酶,它是通过将凝结芽孢杆菌(Bacillus coagulans NL01)的L-阿拉伯糖异构酶第279位苯丙氨酸突变为异亮氨酸而得到;
其中,所述的L-阿拉伯糖异构酶基因araA,其核苷酸序列的GenBank登记号为KX356659。
一株表达定点突变L-阿拉伯糖异构酶的大肠杆菌,它是导入了上述L-阿拉伯糖异构酶的大肠杆菌。
上述的表达定点突变L-阿拉伯糖异构酶的大肠杆菌的构建方法,包括如下步骤:
(1)将定点突变后的L-阿拉伯糖异构酶基因araAF279I克隆到大肠杆菌表达载体pETDuet-1的多克隆位点处,得到重组质粒pETDuet-araAF279I;
(2)将该质粒转化大肠杆菌Escherichia coli BL21,利用氨苄青霉素平板筛选得到阳性转化子,即得到重组大肠杆菌E.coli BL21(pETDuet-araAF279I)。
以上所述的表达定点突变L-阿拉伯糖异构酶的大肠杆菌在催化制备D-塔格糖中的应用在本发明的保护范围之内。
其中,所述的应用包括如下步骤:
(1)细胞的准备:将重组大肠杆菌按体积比为0.1~3%的量转接到发酵培养基中,当细胞OD600达到0.4~0.8后,加入终浓度为0.1~1mmol/L的IPTG,在16~30℃继续培养1~12h,培养结束后,收集重组大肠杆菌;
(2)催化方法:将步骤(1)中得到的重组大肠杆菌加入含有D-半乳糖的反应体系中,催化制备D-塔格糖。
其中,步骤(1)中,所述的发酵培养基为蛋白胨10g/L,酵母粉5g/L,氯化钠10g/L,氨苄青霉素100mg/L,溶剂为水。
其中,步骤(2)中,反应的温度为在20~80℃,优选为50~70℃。
其中,步骤(2)中,反应的时间为5min~100h,优选为1~48h。
其中,重组大肠杆菌的干菌添加量为0.5~10g/L,优选为2.4~6g/L。
其中,步骤(2)中,所述的反应体系,其D-半乳糖的初始浓度为1~500g/L,优选为20~250g/L。
其中,步骤(2)中,所述的反应体系,其初始pH值为4~10,优选为6~8。
有益效果:
本发明具有如下显著的特征和效果:
本发明构建了一个有工业应用价值的重组大肠杆菌,其表达的F279I对D-半乳糖催化效率高。全细胞催化法反应体系最适反应温度温和,不会带来褐变反应,比野生型L-AI和其他常用的来自嗜热菌和极端嗜热菌的L-AI更有利于D-塔格糖的工业化生产。同时全细胞催化法反应体系成分简单,有较大的工业化生产、应用潜力和经济价值。
附图说明
图1催化反应温度对E.coli BL21(pETDuet-araAF279I)和E.coli BL21(pETDuet-araA)生产D-塔格糖的影响。(■),E.coli BL21(pETDuet-araAF279I);(□),E.coli BL21(pETDuet-araA)。
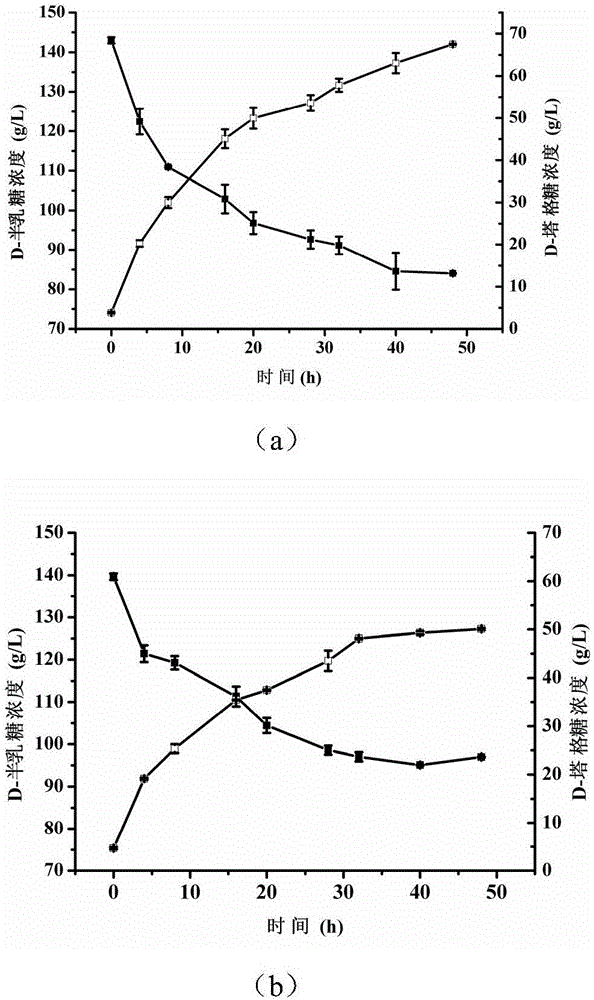
图2在D-半乳糖的初始浓度为150g/L时,E.coli BL21(pETDuet-araAF279I)(图a)和E.coli BL21(pETDuet-araA)(图b)全细胞催化制备D-塔格糖的时间曲线图。(■),D-半乳糖浓度;(□),D-塔格糖浓度。
图3在D-半乳糖的初始浓度为250g/L时,E.coli BL21(pETDuet-araAF279I)(图a)和E.coli BL21(pETDuet-araA)(图b)全细胞催化制备D-塔格糖的时间曲线图。(■),D-半乳糖浓度;(□),D-塔格糖浓度。
具体实施方式
根据下述实施例,可以更好地理解本发明。然而,本领域的技术人员容易理解,实施例所描述的内容仅用于说明本发明,而不应当也不会限制权利要求书中所详细描述的本发明。
以下实施例中D-半乳糖和D-塔格糖产量的检测方法如下:将反应混合物离心除去菌体,稀释上清液并用0.22微米滤膜过滤,采用Agilent 1200液相色谱仪,配备Sugar-pak 1(6.5×300mm,Waters)分离柱,柱温80℃,流动相为去离子水,流速为0.4mL/min,进样量10μL,示差折光检测器,控温35℃。
实施例1:大肠杆菌基因工程菌株E.coli BL21(pETDuet-araAF279I)的获得。
(1)L-阿拉伯糖异构酶基因araA的获得与重组质粒pETDuet-araA的构建。
设计引物,引入能插入pETDuet-1质粒多克隆位点处的的BamH I和EcoR I酶切位点,序列如下:
araA.f:5’-CGCGGATCCGATGTTGAAAATAAAAGA-3’
araA.r:5’-CCGGAATTCTCAAATTTTTACAGTTT-3’
以凝结芽孢杆菌基因组DNA为模板,采用上述引物进行PCR扩增。PCR扩增总体系为100μL,含超纯水64μL,5×FastPfu PCR Buffer 20μL,dNTPs 0.25mmol/L,引物各0.2~0.4μmol/L,模板50~200ng,TransGen公司的FastPfu DNA Polymerase 5U。PCR扩增条件为:95℃预变性2分钟,按如下参数30个循环(95℃变性20秒,60℃退火20秒,72℃延伸45秒),循环结束后72℃延伸5分钟。PCR反应产物用1%的琼脂糖凝胶电泳分析结果。将PCR反应获得的产物回收后用BamH I和EcoR I双酶切处理,同时,用BamH I和EcoR I双酶切处理pETDuet-1质粒,酶切产物经琼脂糖凝胶电泳分离纯化后采用NEB公司的T4DNA连接酶22℃连接1小时后转化大肠杆菌Mach1-T1感受态细胞,经过氨苄青霉素抗性筛选,提取阳性克隆质粒并酶切图谱分析,验证得到载有araA基因的重组质粒pETDuet-araA,随后将重组质粒pETDuet-araA转化导入大肠杆菌BL21(DE3),经氨苄青霉素筛选获得的阳性克隆菌株命名为E.coli BL21(pETDuet-araA)。
(2)运用同源建模和分子对接的方法得到凝结芽孢杆菌NL01的L-AI与D-半乳糖的对接模型。因为离底物距离越近的氨基酸残基,越有可能影响底物的结合与催化。分析发现,距离底物D-半乳糖分子较近的氨基酸残基有9个,分别为M185、R186、F279、E306、E331、H348、M349、I370和H447。但是E306、E331、H348和H447为关键催化氨基酸残基,这些位点的突变会导致L-AI完全失活,因此不能随意修改。故而将剩余的5个氨基酸残基选定为改造位点。
(3)选择M185、R186、F279、M349和I370位点进行定点饱和突变研究,即将质粒pETDuet-araA上编码M185、R186、F279、M349和I370的碱基分别替换为NNN(可随机其他种类的氨基酸),以筛选对D-半乳糖催化能力最强的突变。具体操作参考北京全式金生物技术有限公司的Fast Mutagensis System试剂盒。将饱和突变后得到的大量转化子分别进行诱导表达,收集表达后的菌体与D-半乳糖混合后发生催化反应生成D-塔格糖,用半胱氨酸咔唑法使生成的D-塔格糖显色,根据颜色深浅初步筛选对D-半乳糖酶活较高的转化子。
半胱氨酸咔唑法显色反应步骤如下:
适当稀释反应液,加入200μL 1.5%盐酸-半胱氨酸溶液,6mL 75%硫酸溶液。摇匀后,立即加入200μL咔唑酒精溶液,摇匀于50℃保温10min,冰浴冷却10min,在560nm下测定吸光值(A560)。根据吸光值大小判断颜色深浅。
显色结果显示,F279N转化子中,有1株显色特别深,通过质粒测序发现其279位的Phe(F)突变为了Ile(I)。该转化子即为E.coli BL21(pETDuet-araAF279I)。以L-阿拉伯糖为底物时其酶活为3.02U/mg,低于原始L-AI;以D-半乳糖为底物时其酶活为0.63U/mg,高于原始L-AI。
实施例2:原始L-AI和F279I的表达纯化及催化能力分析。
将实施例1得到的重组菌株E.coli BL21(pETDuet-araA)和E.coli BL21(pETDuet-araAF279I)分别接种于50mL含100μg/mL氨苄青霉素的LB液体培养基,37℃摇床培养12小时,转接于1L含100μg/mL氨苄青霉素的LB液体培养基,37℃摇床培养,当培养液OD600nm达到0.4~0.8后加入终浓度为0.5~1mmol/L的IPTG诱导表达,20℃继续培养8小时。诱导培养结束后,于12,000转/分钟离心5分钟,获得细胞沉淀,生理盐水洗涤两次后,重悬于100mL pH 7.4、50mmol/L的磷酸盐缓冲液中。在冰水混合物中利用超声波破碎细胞壁,破碎时间为20分钟,破壁后的溶液以16,000转/分钟离心60分钟,分别收集上清,经镍离子亲和层析进行纯化,得到L-AI和F279I的纯酶制品。在酶反应体系中加入12.5mM到700mM的L-阿拉伯糖或D-半乳糖(S),反应20min后用半胱氨酸咔唑硫酸法测定L-核酮糖或D-塔格糖的生成量,计算产物生成速率(v),利用双倒数曲线计算得到Km、Vmax和kcat,结果如表1。在此基础上计算得到kcat/Km,用以表征L-AI突变前后对L-阿拉伯糖和D-半乳糖的催化效率。结果显示,F279I对L-阿拉伯糖的催化效率由8.7min-1mM-1降低至5.2min-1mM-1,而对D-半乳糖的催化效率由1.0min-1mM-1提高为1.3min-1mM-1。
表1 L-AI和F279I的动力学常数
实施例3:E.coli BL21(pETDuet-araAF279I)和E.coli BL21(pETDuet-araA)在不同温度下生产D-塔格糖。
在10mL反应体系中,分别加入20g/L底物D-半乳糖、干重为4.8g/L的重组大肠杆菌湿菌体,混匀后在不同温度下振荡反应,反应体系的pH为7.5,15小时后将反应混合物离心除去菌体,取上清液检测生成的D-塔格糖产量。结果显示(图1),反应体系温度在40~70℃时,E.coli BL21(pETDuet-araAF279I)的转化率明显高于E.coli BL21(pETDuet-araA);此外,E.coli BL21(pETDuet-araAF279I)的转化率在50~70℃内差异不大,为了防止褐变反应发生,可以选取50℃进行催化反应;而对于E.coli BL21(pETDuet-araA),其最适催化温度为60℃。
实施例4:E.coli BL21(pETDuet-araAF279I)和E.coli BL21(pETDuet-araA)在初始D-半乳糖浓度为150g/L条件下生产D-塔格糖。
在10mL反应体系中,分别加入150g/L的底物D-半乳糖、干重为4.8g/L的重组大肠杆菌湿菌体,反应体系的pH为7.5,对于E.coli BL21(pETDuet-araAF279I),反应温度控制在50℃,对于E.coli BL21(pETDuet-araA),反应温度控制在60℃,在反应过程中间取样检测生成的D-塔格糖产量。结果显示,D-塔格糖产量随时间延长而升高,但含有E.coli BL21(pETDuet-araAF279I)的催化体系所产D-塔格糖产量始终高于对照菌株,48小时后,D-塔格糖最高产量为67.5g/L,产率为4.5g/(L h)(图2)。
实施例5:E.coli BL21(pETDuet-araAF279I)和E.coli BL21(pETDuet-araA)在初始D-半乳糖浓度为250g/L条件下生产D-塔格糖。
在10mL反应体系中,分别加入250g/L的底物D-半乳糖、干重为4.8g/L的重组大肠杆菌湿菌体,反应体系的pH为7.5,对于E.coli BL21(pETDuet-araAF279I),反应温度控制在50℃,对于E.coli BL21(pETDuet-araA),反应温度控制在60℃,在反应过程中间取样检测生成的D-塔格糖产量。结果显示,D-塔格糖产量随时间延长而升高,但含有E.coli BL21(pETDuet-araAF279I)的催化体系所产D-塔格糖产量始终高于对照菌株,48小时后,D-塔格糖最高产量为88.4g/L,产率为5.9g/(L h)(图3)。
SEQUENCE LISTING
<110> 南京林业大学
<120> 一种L-阿拉伯糖异构酶及其应用
<130> SG20161011001
<160> 2
<170> PatentIn version 3.5
<210> 1
<211> 27
<212> DNA
<213> Artificial Sequence
<220>
<223> araA.f
<400> 1
cgcggatccg atgttgaaaa taaaaga 27
<210> 2
<211> 26
<212> DNA
<213> Artificial Sequence
<220>
<223> araA.r
<400> 2
ccggaattct caaattttta cagttt 26
- 还没有人留言评论。精彩留言会获得点赞!
- 一种查尔酮基荧光传感器及其制备方法与应用与流程
- 一种含咔唑基的查尔酮与乙酰乙酸乙酯Micheal加成产物及其制备方法与流程
- 一种查尔酮类似物及其制备方法和用途与流程
- 一种制备4-溴-1,8-二氮萘的方法与制造工艺
- 一种利用磁性固相萃取吸附材料富集检测喜树碱的方法与制造工艺
- 拟南芥At‐UGE2基因及其过表达突变株和缺失突变株在调节植物性状中的应用的制造方法与工艺
- 一种马来酸顺反异构酶的热稳定性改造及其应用的制造方法与工艺
- 一种改进的基于加权图的超密集异构网络干扰协调方法与制造工艺
- 一种自旋标记骆驼宁碱A化合物、制备方法及其用途与制造工艺
- 一种顺‑4‑(2‑甲基‑3‑对叔戊基苯基丙酰基)‑2,6‑二甲基吗啉合成方法与制造工艺