核酸构建体的制作方法
发明领域
本发明涉及用于调节从单个载体共表达的多肽的相对表达的构建体和方法。特别地,本发明涉及调节从单个载体与第二多肽共表达的跨膜蛋白的表达。
发明背景
通常希望从同一载体表达不同的蛋白质,因为同一细胞的多重转导是困难的,昂贵的和不可预测的。因此已经开发了不同的方法以允许从单个载体共表达两种蛋白(参见图1)。
最初的尝试在同一盒中使用两种不同的启动子。这产生两个单独的转录物,其每个编码单独的蛋白质。因为一些原因,这是困难的方法。一个关键问题是“启动子干扰”,其中一个启动子占据优势并引起第二个启动子的沉默。此外,不同的启动子在不同的细胞环境中差异起作用,这使得难以实现每个转基因的相对表达的一致的“调整(tuning)”。
另一种方法是使用内部核糖体进入序列(ires)。这里,生成单个转录物。该转录物中的ires序列置于两个转基因的开放阅读框之间,并模拟mrna帽结构。因此,核糖体在5’帽或者ires起始翻译,导致两个单独的蛋白的表达。这种方法的关键限制是不能控制相对表达。3'转录物通常表达得少于5'转录物,但表达比例难以预测和调整。
在口蹄疫病毒(fmdv)2a肽在允许fmdv(和相关病毒)从单个开放阅读框(orf)表达多个蛋白质中的作用表征之后,提供了进一步的方法(donnellyetal;j.gen.virol.;82,1027–1041(2001))。在orf的翻译后,2a肽(和同源物)立即以非常高的效率切割,使得能够从单个orf表达多个肽。使用2a肽在同一orf中的不同肽之间切割的问题是表达被限制为1:1的比例。
因此,需要与上述缺点不相关的用于从单个载体表达多于一个蛋白质的替代方法。
发明概述
本发明基于确定当两个含有信号肽的蛋白作为在翻译后切割的多聚蛋白共表达时,可以改变信号肽的相对“强度”,从而控制两个蛋白质的相对表达。这不需要限于转基因对,而可用于允许控制最初翻译为多聚蛋白的多个蛋白的相对表达。
如本文所用,“多聚蛋白”是指下述的多肽序列,其从单个核酸构建体翻译成单个实体,但包含随后分离并且作为离散实体(例如分离的蛋白质)发挥功能的多肽序列。
因此,本发明的第一个方面提供核酸构建体,其包括以下结构:
a-x-b
其中
a是编码包含第一信号肽的第一多肽的核酸序列;
b是编码包含第二信号肽的第二多肽的核酸序列,并且
x是编码切割位点的核酸序列,
其中所述第一信号肽或所述第二信号肽包含一个或多个突变,使得其具有较少的疏水性氨基酸。
突变的信号肽可以比其衍生自的“野生型”信号肽具有更少的疏水性氨基酸。
突变的信号肽可以比另一多肽的信号肽具有更少的疏水性氨基酸。
疏水性氨基酸可以选自下组:丙氨酸(a);缬氨酸(v);异亮氨酸(i);亮氨酸(l);甲硫氨酸(m);苯丙氨酸(p);酪氨酸(y);色氨酸(w)。
第一信号肽和第二信号肽可衍生于相同的序列,但是与另一个信号肽相比,一个信号肽包含一个或多个氨基酸缺失或取代以去除或替换一个或多个疏水性氨基酸。
如下文更详细解释的,信号序列具有三分型(tripartite)结构,由疏水性核心区(h-区)和侧翼的n-和c-区组成。第一信号肽和第二信号肽可以具有相同的n-和c-区,但在h-区中可以不同:一个信号肽的h-区比另一信号肽的h-区具有更多的疏水性氨基酸。
从信号肽缺失或替换的疏水性氨基酸可以是选自下组:丙氨酸(a);缬氨酸(v);异亮氨酸(i);亮氨酸(l);甲硫氨酸(m);苯丙氨酸(p);酪氨酸(y);色氨酸(w)。
从信号肽缺失或替换的疏水性氨基酸可以是选自下组:缬氨酸(v);异亮氨酸(i);亮氨酸(l);和色氨酸(w)。
一个多肽的信号肽可以比另一个信号肽多包含直至5个疏水性氨基酸。可以对改变的信号肽替换或去除直至(upto)10%,直至20%,直至30%,直至40%或直至50%的其疏水性氨基酸。
第一和第二多肽可以是含有信号肽的多肽,例如分泌蛋白,跨膜蛋白或细胞器蛋白。本发明使得能够控制蛋白的相对表达,即在相关区室中蛋白的相对量。对于分泌蛋白,本发明使得能够控制由细胞产生的蛋白的相对量。对于跨膜蛋白,本发明使得能够控制两个(或更多个)蛋白的相对细胞表面表达。对于细胞器蛋白,本发明使得能够控制讨论中的细胞器之中或膜上的蛋白的相对表达。
第一和第二多肽可以都是跨膜蛋白,例如t细胞受体或嵌合抗原受体(car)。
在第一和第二蛋白是跨膜蛋白的情况下,第一和第二信号肽之间的差异可以使得当核酸构建体在细胞中表达时,在细胞表面存在第一和第二多肽的差异相对表达。
x可以是编码自切割肽,弗林蛋白酶切割位点或烟草蚀刻病毒切割位点的核酸序列。
x可以是编码来自口蹄病毒或心脏病毒的2a自切割肽或2a样肽。
在第二个方面,本发明提供载体,其包含根据前述权利要求任一项的核酸构建体。
载体可以是例如逆转录病毒载体或慢病毒载体。
在第三个方面,提供细胞,其包含根据本发明的第一个方面的核酸构建体或根据本发明的第二个方面的载体。
在这第三个方面,还提供细胞,其包含具有不同信号肽的两种嵌合抗原受体。
该两个信号肽可以具有不同数量的疏水性氨基酸。
在这一方面,具有含有较多数量的疏水性氨基酸的信号肽的嵌合抗原受体比具有含有较少数量的疏水性氨基酸的信号肽的嵌合抗原受体可以更高地在细胞表面表达。
还提供细胞,其包含具有不同信号肽的两种嵌合抗原受体,其中一种嵌合抗原受体的信号肽包含一个或多个突变,使得其具有较少的疏水性氨基酸。
具有突变的信号肽的嵌合抗原受体比具有未突变的信号肽的嵌合抗原受体以更低水平在细胞表面表达。
细胞可以是细胞溶解性免疫细胞,例如t细胞或自然杀伤(nk)细胞。
在第四个方面,提供用于制备根据本发明的第四个方面的细胞的方法,其包括将根据本发明的第一个方面的核酸构建体或根据本发明的第二个方面的载体引入细胞中的步骤。
在第五个方面,提供用于调节从单个核酸构建体与含有信号肽的第二蛋白一起表达的含有信号肽的第一蛋白的相对细胞表面表达的方法,其包括突变编码一个蛋白的信号肽的核酸序列的步骤,以便与另一个蛋白的信号肽相比去除或替换一个或多个疏水性氨基酸。
与具有未修饰的信号肽的跨膜蛋白相比,从跨膜蛋白的信号肽去除或替换疏水性氨基酸减少在细胞表面上表达的跨膜蛋白的量。因此,可以调节衍生自包括第二多肽的多聚蛋白的跨膜蛋白的相对表达水平。当跨膜蛋白仅在细胞表面具有活性(或主要在细胞表面上具有活性)时,降低蛋白的相对细胞表面表达也降低其相对活性。
可以延伸本发明以调节由切割位点分开的作为联接(concatenated)多肽表达的三种或更多种蛋白质的相对表达,并且通过不同活性的信号肽决定相对表面表达。
附图简述
图1:用于从同一载体表达不同蛋白的方法
(a)同一盒内的两个不同的启动子导致两个不同的转录物,其各自产生单独的蛋白。(b)使用内部核糖体进入序列(ires)导致单个转录物,所述单个转录物翻译成两个单独的蛋白。(c)fmdv2a肽的使用导致单个转录物和单个多聚蛋白,其快速切割成两个单独的蛋白。
图2:说明信号序列在蛋白质靶向中的功能的示意图
图3:编码两种car的核酸构建体示意图
图4:验证取代的信号序列的功能。
pct/gb2014/053452描述了编码两种嵌合抗原受体(car)(一种针对cd19,一种针对cd33)的载体系统。在该研究中用于car的信号肽是来自人cd8a信号序列的信号肽。为了本研究的目的,将这用来自鼠igkappa链v-iii区域的信号肽取代,所述信号肽具有以下序列:
图5:测试鼠igkappa链v-iii中一个氨基酸缺失的影响。创建了突变体1kappa链,其具有h-区中的以下缺失(以灰色示出)
图6:测试鼠igkappa链v-iii中两个氨基酸缺失的影响。创建了突变体2kappa链,其具有h-区中的以下缺失(以灰色示出)
图7:测试鼠igkappa链v-iii中三个氨基酸缺失的影响。创建了突变体2kappa链,其具有h-区中的以下缺失(以灰色示出)
图8:测试鼠igkappa链v-iii中五个氨基酸缺失的影响。创建了突变体2kappa链,其具有h-区中的以下缺失(以灰色示出)
发明详述
本发明提供核酸构建体,其包含以下结构:
a-x-b
其中
a是编码包含第一信号肽的第一多肽的核酸序列;
b是编码包含第二信号肽的第二多肽的核酸序列,并且
x是编码切割位点的核酸序列,
其中所述第一信号肽或所述第二信号肽包含一个或多个突变,使得其具有较少的疏水性氨基酸。
核酸构建体
多肽
由本发明的核酸构建体制备的多肽是含有信号肽的多肽,例如分泌蛋白,跨膜蛋白或细胞器蛋白。
信号肽
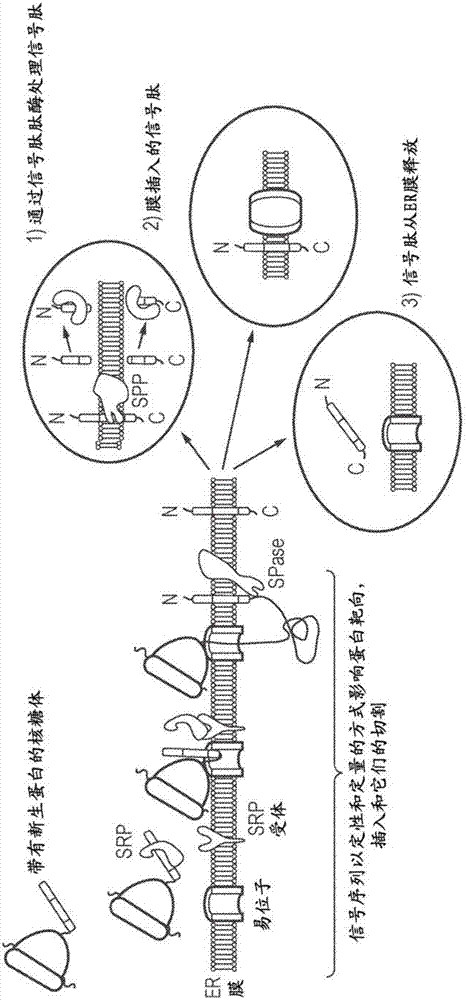
由本发明的核酸构建体编码的多肽a和b(和任选的其它,c,d等)各自可以包含信号肽,使得当该多肽在细胞内部表达时,新生蛋白被引导至内质网(er)(参见图2a)。
术语“信号肽”与“信号序列”同义。
信号肽是短肽,通常5-30个氨基酸长,存在于大多数注定向着分泌途径的新合成蛋白的n末端。这些蛋白包括存在于某些细胞器(例如,内质网,高尔基体或内体)内部的那些蛋白,从细胞分泌的那些蛋白,以及跨膜蛋白。
信号肽通常含有核心序列,其是具有形成单个α-螺旋的倾向的疏水性氨基酸的长段。信号肽可以从氨基酸的短带正电荷段开始,该段有助于在移位期间执行多肽的适当拓扑学。在信号肽的末端通常有被信号肽酶识别和切割的一段氨基酸。信号肽酶可以在移位期间或完成之后切割,以产生游离信号肽和成熟蛋白。然后游离的信号肽被特异性蛋白酶消化。
信号肽通常位于分子的氨基末端,尽管一些羧基末端信号肽是已知的。
如上文所述,信号序列具有三分型结构,其由疏水性核心区(h-区)和侧翼的n-和c-区组成。后者含有信号肽酶(spase)共有切割位点。通常,信号序列以共翻译方式(co-translationally)被切割,所得到的切割的信号序列称为信号肽。
在来自鼠igkappa链v-iii区的信号肽中,所述信号肽具有以下序列:
在本发明的核酸构建体中,两个(或更多个)多肽的信号序列在它们的h-区中不同。一个多肽(其具有较高的相对表达)在h-区中具有比另一个多肽(其具有较低的相对表达)更多数量的疏水性氨基酸。具有较低相对表达的多肽的信号肽相比于具有较低相对表达的多肽的信号肽在h-区中可以包含疏水性氨基酸的一个或多个氨基酸突变,例如取代或缺失。
第一信号肽和第二信号肽可以具有基本上相同的n-和c-区,但在h-区中不同,如上文解释。“基本上相同”表示第一和第二信号肽之间的n-和c-区可以是相同的,或者在不影响信号肽的功能的情况下,可以在n-或c链中相差一个,两个或三个氨基酸。
核心中的疏水性氨基酸可以是例如:丙氨酸(a);缬氨酸(v);异亮氨酸(i);亮氨酸(l);甲硫氨酸(m);苯丙氨酸(p);酪氨酸(y);或色氨酸(w)。
经突变以改变信号肽效率的疏水性氨基酸可以是来自上述列表的任一个,特别是:缬氨酸(v);异亮氨酸(i);亮氨酸(l);和色氨酸(w)。
在h-区中的残基中,一个信号肽(例如,改变的信号肽)可以比另一信号肽(例如,未改变的信号肽)包含少至少10%,20%,30%,40%或50%的疏水性氨基酸。
在h-区包含5-15个氨基酸的情况下,一个信号肽可以比另一个信号肽多包含1,2,3,4或5个疏水性氨基酸。
改变的信号肽可以包含疏水性氨基酸的1,2,3,4或5个氨基酸缺失或取代。可以使用非疏水性氨基酸,例如亲水性或中性氨基酸替换疏水性氨基酸。
可以使用软件技术(参见例如http://www.predisi.de/)检测或预测信号序列。
已知大量的信号序列,并可以在数据库中获得。例如http://www.signalpeptide.de在其数据库中列出了2109个经确认的哺乳动物信号肽。
表1提供了信号序列的列表,其纯粹是为了说明的目的。疏水性核心以粗体突出显示。这包括为了本发明的目的可以被取代或去除的氨基酸的实例。
表1
突变的信号肽包含一个或多个突变,使得其具有比其衍生自的野生型信号肽更少的疏水性氨基酸。术语“野生型”是指存在于其衍生自的天然蛋白中的信号肽的序列。例如,实施例中描述的信号肽是来自鼠igkappa链v-iii区的信号肽,其具有野生型序列:metdtlilwvllllvpgstg。
术语“野生型”还包括衍生自天然存在的蛋白的信号肽,其在n-或c-区中包含一个或多个氨基酸突变。例如,通常使用n末端的保守氨基酸取代修饰天然信号肽以引入限制性位点。对于本发明的目的,此类修饰的信号肽序列(其在h-区中不包含任突变)认为是“野生型”。
本发明还涉及不能参照野生型序列来定义的合成的信号肽序列。在该实施方案中,一种多肽的信号肽包含比另一多肽的信号序列更少的疏水性氨基酸。两个信号序列可以衍生自相同的合成信号肽序列,但核心区域中疏水性氨基酸的数目不同。
跨膜蛋白
本发明使得能够调节跨膜表面蛋白的相对表达。跨膜表面蛋白是在细胞表面表达的蛋白。当在细胞表面表达时,跨膜蛋白的至少一个结构域是胞质外的(即在细胞的外部)。
跨膜蛋白可以是单次跨膜蛋白,即其可以包含单个跨膜结构域,或者其可以包含多个跨膜结构域。
跨膜蛋白可以通过拓扑学分类,即参照n-和c-末端结构域的位置。i型,ii型和iii型跨膜蛋白是单通分子,而iv型跨膜蛋白是多通分子。i型跨膜蛋白以停止转移锚定序列(stop-transferanchorsequence)锚定到脂质膜上,并且在合成过程中其n末端结构域靶向至er腔(以及细胞外空间,当成熟形式位于质膜上时)。ii型和iii型以信号-锚定序列锚定,其中ii型以其c末端结构域靶向至er腔,而iii型使其n末端结构域靶向至er腔。iv型细分为iv-a,其中其n末端结构域靶向胞质溶胶,和iv-b,其中n端结构域靶向腔。
由本发明的核酸构建体制备的跨膜蛋白可以是i-iv型中的任一种。
跨膜结构域可以是在膜中热力学稳定的任何蛋白结构。这通常是包含几个疏水残基的α螺旋。任何跨膜蛋白的跨膜结构域可用于提供跨膜部分。蛋白的跨膜结构域的存在和跨度可以由本领域技术人员使用tmhmm算法(http://www.cbs.dtu.dk/services/tmhmm-2.0/)确定。此外,鉴于蛋白的跨膜结构域是相对简单的结构,即预测形成足够长度以跨越膜的疏水性α螺旋的多肽序列,也可以使用人工设计的tm结构域(us7052906b1描述了合成的跨膜组分)。
跨膜结构域可以衍生自cd28,其给出良好的稳定性。
i型跨膜蛋白的结构和加工是本领域公知的。此类蛋白质通常包含细胞外结构域,跨膜结构域和细胞内结构域,并且是具有穿过细胞膜的单个α-螺旋的单通分子。
i型跨膜蛋白通常具有信号肽,其被内质网(er)快速识别,因此翻译中的蛋白被快速再导向至er。疏水螺旋将蛋白锁定然后锚定在er的膜中。
如上文所述,i型跨膜蛋白通过停止转移锚定序列锚定到脂质膜上。停止转移序列停止多肽的进一步移位并充当跨膜锚。
如本文所使用的,术语i型跨膜蛋白涵盖包含i型跨膜结构域和停止转移锚定序列的任何蛋白,并且在不存在外源性细胞内保留信号的情况下靶向在细胞表面上表达。
适用于本发明的各种1型跨膜蛋白是本领域已知的。此类蛋白包括但不限于抑制性受体,刺激受体,细胞因子受体和g蛋白。
跨膜蛋白可以是t细胞受体α或β链。
跨膜蛋白可以是嵌合抗原受体(car)。
car是将抗原结合结构域移植到t细胞的效应物功能的蛋白。它们通常的形式是i型跨膜结构域蛋白的形式,具有抗原识别氨基末端,间隔区,跨膜结构域,全部连接到传输t细胞存活和激活信号的复合胞内域。
抗原结合结构域可以衍生自抗体或抗体模拟物,或者其可以是特异性结合抗原的另一实体,例如配体。
这些分子的最常见形式是衍生自识别靶抗原的单克隆抗体的单链可变片段(scfv)通过间隔区和跨膜结构域融合至信号传导胞内域的融合物。此类分子响应于scfv对其靶标的识别导致t细胞的活化。当t细胞表达此类car时,它们识别并杀死表达靶抗原的靶细胞。已经开发了几种针对肿瘤相关抗原的car,并且使用此类表达car的t细胞的过继转移方法目前正在临床试验中用于治疗各种癌症。
信号转导胞内域也可能存在于单独的分子上。与本发明有关的术语“car”还涵盖包含与跨膜结构域连接的抗原结合结构域的分子。此类car可以能够与单独的细胞内信号传导结构域相互作用以便于刺激t细胞活化。
在本发明中,核酸序列a或b可以是编码包含信号肽的跨膜蛋白的核酸序列。
大多数感兴趣的跨膜蛋白仅当在细胞表面时有活性,或者主要是有活性的。因此,使用低效的信号肽减少蛋白质在细胞表面的相对表达,并从而降低蛋白质的相对活性。
多聚蛋白
本发明的核酸构建体可以编码多聚蛋白,其包含第一和第二多肽。
第一和第二多肽可以是嵌合抗原受体(car)。
实施例中描述的核酸构建体编码以下多聚蛋白,其以组分被列出的顺序包含多个组分:
衍生于小鼠igkappa链v-iii区的信号肽
metdtlilwvllllvpgstg(见下文)
scfvacd19:
diqmtqttsslsaslgdrvtiscrasqdiskylnwyqqkpdgtvklliyhtsrlhsgvpsrfsgsgsgtdysltisnleqediatyfcqqgntlpytfgggtkleitkaggggsggggsggggsggggsevklqesgpglvapsqslsvtctvsgvslpdygvswirqpprkglewlgviwgsettyynsalksrltiikdnsksqvflkmnslqtddtaiyycakhyyyggsyamdywgqgtsvtvs
接头:
sd
人cd8astk:
ptttpaprpptpaptiasqplslrpeacrpaaggavhtrgldfacdi
人cd28tm:
fwvlvvvggvlacysllvtvafiifwv
人cd3zeta细胞内结构域:
rrvkfsrsadapayqqgqnqlynelnlgrreeydvldkrrgrdpemggkprrknpqeglynelqkdkmaeayseigmkgerrrgkghdglyqglstatkdtydalhmqalppr
2a肽:
raegrgslltcgdveenpgp
衍生于小鼠igkappa的信号肽:
mavptqvlgllllwltda
scfvacd33:
rcdiqmtqspsslsasvgdrvtitcrasediyfnlvwyqqkpgkapklliydtnrladgvpsrfsgsgsgtqytltisslqpedfatyycqhyknypltfgqgtkleikrsggggsggggsggggsggggsrsevqlvesggglvqpggslrlscaasgftlsnygmhwirqapgkglewvssislnggstyyrdsvkgrftisrdnakstlylqmnslraedtavyycaaqdaytggyfdywgqgtlvtvssm
接头:
dpa
衍生于人igg1的铰链和fc,具有突变以防止fcrg结合(hch2ch3pvaa):
epkspdkthtcppcpappvagpsvflfppkpkdtlmiartpevtcvvvdvshedpevkfnwyvdgvevhnaktkpreeqynstyrvvsvltvlhqdwlngkeykckvsnkalpapiektiskakgqprepqvytlppsrdeltknqvsltclvkgfypsdiavewesngqpennykttppvldsdgsfflyskltvdksrwqqgnvfscsvmhealhnhytqkslslspgk
接头:
kdpk
人cd148tm:
avfgcifgalvivtvggfifw
人cd148细胞内结构域:
rkkrkdaknnevsfsqikpkksklirvenfeayfkkqqadsncgfaeeyedlklvgisqpkyaaelaenrgknrynnvlpydisrvklsvqthstddyinanympgyhskkdfiatqgplpntlkdfwrmvweknvyaiimltkcveqgrtkceeywpskqaqdygditvamtseivlpewtirdftvkniqtseshplrqfhftswpdhgvpdttdllinfrylvrdymkqsppespilvhcsagvgrtgtfiaidrliyqienentvdvygivydlrmhrplmvqtedqyvflnqcvldivrsqkdskvdliyqnttamtiyenlapvttfgktngyia
如实施例中所述,改变来自鼠igkappa链v-iii区的信号序列,其具有序列:metdtlilwvllllvpgstg(疏水性残基加粗突出显示)。从疏水核心序贯缺失疏水性氨基酸,以便降低抗cd19car的表达水平。
切割位点
本发明第一个方面的核酸构建体包含位于编码第一和第二多肽的核酸序列之间的编码切割位点的序列,使得第一和第二多肽可以作为单独的实体表达。
切割位点可以是能够使包含第一和第二多肽的多肽分开的任何序列。
术语“切割”在本文中为了方便使用,但是切割位点可以通过除经典切割以外的机制将第一和第二多肽分离成单个实体。例如,对于口蹄疫病毒(fmdv)2a自切割肽(见下文),已经提出了各种模型来解释“切割”活性:宿主细胞蛋白酶的蛋白水解,自体蛋白水解或翻译效应(donnellyetal(2001)j.gen.virol.82:1027-1041)。此类“切割”的确切机制对于本发明的目的而言并不重要,只要切割位点当位于编码第一和第二多肽的核酸序列之间时,导致第一和第二多肽表达为单独的实体。
切割位点可以是弗林蛋白酶切割位点。
弗林蛋白酶是属于枯草杆菌蛋白酶样前蛋白转化酶家族的酶。该家族的成员是将潜伏的前体蛋白加工成其生物学活性产物的前蛋白转化酶。弗林蛋白酶是钙依赖性丝氨酸内切蛋白酶,其可以在它们配对的碱性氨基酸加工位点有效切割前体蛋白。弗林蛋白酶底物的实例包括甲状旁腺激素原,转化生长因子β1前体,白蛋白原,β-分泌酶原,膜1型基质金属蛋白酶,神经生长因子原的β亚基和willebrand因子。弗林蛋白酶正好在碱性氨基酸靶序列(典型地为arg-x-(arg/lys)-arg')的下游切割蛋白,并且在高尔基体中富集。
切割位点可以是烟草蚀刻病毒(tev)切割位点。
tev蛋白酶是高度序列特异性的半胱氨酸蛋白酶,其是糜蛋白酶样蛋白酶。它非常特异于其靶切割位点,因此经常用于体外和体内融合蛋白的受控切割。共有tev切割位点为enlyfq\s(其中“\”表示被切割的肽键)。哺乳动物细胞,如人类细胞,不表达tev蛋白酶。因此,在本核酸构建体包含tev切割位点并且在哺乳动物细胞中表达的实施方案中,必须在哺乳动物细胞中也表达外源tev蛋白酶。
切割位点可以编码自切割肽。
“自切割肽”是指下述肽,该肽发挥功能使得当包含第一和第二多肽和自切割肽的多肽产生时,其立即被“切割”或分离成独特的且离散的第一和第二多肽,而不需要任何外部切割活性。
自切割肽可以是来自口蹄病毒或心脏病毒的2a自切割肽。口蹄病毒和心脏病毒的主要2a/2b切割由在其自身c末端“切割”的2a介导。在口蹄病毒例如口蹄疫病毒(fmdv)和马鼻炎a型病毒中,2a区域是约18个氨基酸的短区段,其与蛋白2b的n末端残基(保守的脯氨酸残基)一起代表了能够在其自身c末端介导“切割”的自主元件。
较长的心脏病毒蛋白的c末端19个氨基酸与2b的n末端脯氨酸一起以大致等同于口蹄病毒fmdv2a序列的效率介导“切割”。心脏病毒包括脑心肌炎病毒(emcv)和theiler’s鼠脑炎病毒(tmev)。
emcv和fmdv2a的突变分析揭示了基序dxexnpgp密切参与“切割”活性(donellyetal(2001),如上文)。
本发明的切割位点可以包含以下氨基酸序列:
dx1ex2npgp,其中x1和x2是任意氨基酸。x1可以选自下列组:i,v,m和s。x2可以选自下列组:t,m,s,l,e,q和f。
例如,切割位点可以包含表2所示的氨基酸序列之一。
表2
基于2a序列的切割位点的长度可以是例如15-22个氨基酸。该序列可以包含2a蛋白的c末端,随后是脯氨酸残基(其对应于2b的n末端脯氨酸)。
突变研究还显示了,除天然存在的2a序列之外,一些变体也是活性的。切割位点可以对应于来自天然存在的2a多肽的变体序列,具有一个,两个或三个氨基酸取代,其保留了诱导多聚蛋白序列“切割”成两个或多个单独的蛋白的能力。
切割序列可以选自以下切割序列,其都已经显示在一定程度上是有活性的(donellyetal(2001),如上文):
llnfdllklagdvesnpgp(seqidno.11)
llnfdllklagdvqsnpgp(seqidno.12)
llnfdllklagdveinpgp(seqidno.13)
llnfdllklagdvefnpgp(seqidno.14)
llnfdllklagdveshpgp(seqidno.15)
llnfdllklagdvesepgp(seqidno.16)
llnfdllklagdvesqpgp(seqidno.17)
llnfdllklagdvesnpgg(seqidno.18)
基于dxexnpgp基序的序列,已经在除口蹄病毒或心脏病毒外的小核糖核酸病毒,“小rna病毒样”昆虫病毒,c型轮状病毒,锥虫属物种中的重复序列和细菌序列中发现了“2a样”序列(donellyetal(2001),如上文)。切割位点可以包含这些2a样序列之一,例如:
yhadyykqrlihdvemnpgp(seqidno.19)
hyagyfadllihdietnpgp(seqidno.20)
qctnyallklagdvesnpgp(seqidno.21)
atnfsllkqagdveenpgp(seqidno.22)
aarqmllllsgdvetnpgp(seqidno.23)
raegrgslltcgdveenpgp(seqidno.24)
traeiedeliragiesnpgp(seqidno.25)
traeiedeliradiesnpgp(seqidno.26)
akfqidkilisgdvelnpgp(seqidno.27)
ssiirtkmlvsgdveenpgp(seqidno.28)
cdaqrqklllsgdieqnpgp(seqidno.29)
ypidfggflvkadsefnpgp(seqidno.30)
切割位点可以包含如seqidno.24所示的2a样序列(raegrgslltcgdveenpgp)。
已经表明,包括5至39个氨基酸的n末端“延伸”可以增加活性(donellyetal(2001),如上文)。特别地,切割序列可以包含以下序列之一或其变体,所述变体具有例如直至5个氨基酸变化,其保留了切割位点活性:
vtellyrmkraetycprplaihptearhkqkivapvkqtlnfdllklagdvesnpgp(seqidno.31)
llaihptearhkqkivapvkqtlnfdllklagdvesnpgp(seqidno.32)
earhkqkivapvkqtlnfdllklagdvesnpgp(seqidno.33)
apvkqtlnfdllklagdvesnpgp(seqidno.34)
可调节性
可以使用本发明的方法,通过使用具有不同量或比例的疏水性氨基酸的信号肽精确调节一个或多个蛋白的相对表达。
当考虑多个蛋白的表达(每一个具有其自身的相对表达)时,使用不同信号肽的可调节性是特别有用的。例如,考虑具有以下结构的核酸构建体:
a-x-b-y-c
其中
a,b和c是编码多肽的核酸序列;和
x和y是编码切割位点的核酸序列。
核酸构建体将编码三个蛋白a,b和c,其任意或全部可以是跨膜蛋白,如果需要a,b和c表达为使得相对水平为a>b>c,则核酸序列a可以具有含有最多疏水性氨基酸的信号肽,核酸序列b可以具有含有中等量的疏水性氨基酸的信号肽,而核酸序列c可以具有含有最少疏水性氨基酸的信号肽。
载体
本发明还提供了包含根据本发明第一个方面的核酸构建体的载体。
可以使用此类载体将核酸构建体引入宿主细胞中,使得其表达第一和第二多肽。
载体可以是例如质粒或病毒载体,例如逆转录病毒载体或慢病毒载体,或基于转座子的载体或合成的mrna。
载体可以能够转染或转导哺乳动物细胞,例如t细胞。
细胞
本发明还提供了包含本发明的核酸构建体或载体的细胞,其表达由核酸序列编码的第一和第二多肽。
细胞可以是能够在细胞表面表达跨膜蛋白的任何真核细胞,例如免疫细胞。
细胞可以是细胞溶解性免疫细胞,例如t细胞或自然杀伤细胞。
方法
在进一步的方面,本发明提供了制备根据本发明的细胞的方法,其包括将本发明的核酸构建体或载体引入细胞的步骤。
可以通过转导或转染引入核酸构建体。
细胞可以是从受试者分离的细胞,例如从受试者分离的t细胞或nk细胞。
本发明还提供了一种用于调节从单个核酸构建体与第二蛋白一起表达的第一蛋白的相对细胞表面表达的方法,其包括突变编码一个蛋白的信号肽的核酸序列的步骤,以便与另一个蛋白的信号肽相比去除或替换一个或多个疏水性氨基酸
可以通过本领域已知的技术改变信号肽,例如定点诱变和重组技术。
现在将通过实施例进一步描述本发明,这些实施例旨在帮助本领域普通技术人员实施本发明,而不意图以任何方式限制本发明的范围。
实施例
实施例1-使用鼠igkappa链v-iii信号序列
pct/gb2014/053452描述了编码两种嵌合抗原受体(car)(一种针对cd19,而一种针对cd33)的载体系统。在该研究中用于car的信号肽是来自人cd8a信号序列的信号肽。为了本研究的目的,将这用来自鼠igkappa链v-iii区的信号肽取代,所述信号肽具有以下序列:metdtlilwvllllvpgstg(疏水性残基以粗体突出显示)。为了建立鼠igkappa链v-iii信号序列与来自人cd8a的信号序列一样好地发挥功能,进行了比较研究,进行了比较研究。对于两种信号序列,都观察到抗cd33car和抗cd19car的功能性表达。将该取代的信号序列及其所有随后的突变瞬时转染入293t细胞。转染三天后,用与兔fc链融合的可溶性嵌合cd19和与小鼠fc链融合的可溶性嵌合cd33两者染色293t细胞。然后用抗兔fc-fitc和抗小鼠fc-apc染色所有细胞。流式细胞术图显示了取代的信号序列作为与非转染(nt)和具有cd8信号序列的构建体的比较(图4)。发现鼠igkappa链v-iii信号序列与来自人cd8a的信号序列一样好地发挥功能。
实施例2-通过缺失信号肽中的疏水性残基改变相对表达
以逐步方式缺失疏水性残基,并观察对抗cd33car和抗cd19car的相对表达的影响。研究了一个,两个,三个和四个氨基酸缺失的影响,结果分别如图5至图8所示。
与抗cd33car相比,所有突变体构建体显示抗cd19car的相对表达降低。从1至3的氨基酸缺失的数量越大,抗cd19car表达的相对降低越大,但随后达到平稳(四个缺失产生与三个缺失相似的表达降低)。
因此,可以使用编码两个多肽的核酸构建体中的信号序列的修饰来控制两个多肽的相对表达。
上述说明书中提到的所有出版物通过引用并入本文。在不脱离本发明的范围和精神的情况下,本发明所述的方法和系统的各种修改和变化对于本领域技术人员将是显而易见的。虽然已经结合具体的优选实施方案描述了本发明,但是应当理解,所要求保护的本发明不应被过度限制于这些具体实施方案。实际上,对于分子生物学,细胞生物学或相关领域的技术人员显而易见的用于实施本发明的所述模式的各种修改旨在在所附权利要求书的范围内。
序列表
<110>ucl商务股份有限公司
<120>核酸构建体
<130>p107058pct
<150>gb1507108.7
<151>2015-04-27
<160>74
<170>patentinversion3.5
<210>1
<211>8
<212>prt
<213>人工序列
<220>
<223>切割位点,自切割肽
<400>1
aspilegluthrasnproglypro
15
<210>2
<211>8
<212>prt
<213>人工序列
<220>
<223>切割位点,自切割肽
<400>2
aspvalgluthrasnproglypro
15
<210>3
<211>8
<212>prt
<213>人工序列
<220>
<223>切割位点,自切割肽
<400>3
aspvalglumetasnproglypro
15
<210>4
<211>8
<212>prt
<213>人工序列
<220>
<223>切割位点,自切割肽
<400>4
aspvalgluserasnproglypro
15
<210>5
<211>8
<212>prt
<213>人工序列
<220>
<223>切割位点,自切割肽
<400>5
aspmetgluserasnproglypro
15
<210>6
<211>8
<212>prt
<213>人工序列
<220>
<223>切割位点,自切割肽
<400>6
aspvalgluleuasnproglypro
15
<210>7
<211>8
<212>prt
<213>人工序列
<220>
<223>切割位点,自切割肽
<400>7
aspvalglugluasnproglypro
15
<210>8
<211>8
<212>prt
<213>人工序列
<220>
<223>切割位点,自切割肽
<400>8
aspilegluleuasnproglypro
15
<210>9
<211>8
<212>prt
<213>人工序列
<220>
<223>切割位点,自切割肽
<400>9
aspilegluglnasnproglypro
15
<210>10
<211>8
<212>prt
<213>人工序列
<220>
<223>切割位点,自切割肽
<400>10
aspserglupheasnproglypro
15
<210>11
<211>19
<212>prt
<213>人工序列
<220>
<223>切割序列,自切割肽
<400>11
leuleuasnpheaspleuleulysleualaglyaspvalgluserasn
151015
proglypro
<210>12
<211>19
<212>prt
<213>人工序列
<220>
<223>切割序列,自切割肽
<400>12
leuleuasnpheaspleuleulysleualaglyaspvalglnserasn
151015
proglypro
<210>13
<211>19
<212>prt
<213>人工序列
<220>
<223>切割序列,自切割肽
<400>13
leuleuasnpheaspleuleulysleualaglyaspvalgluileasn
151015
proglypro
<210>14
<211>19
<212>prt
<213>人工序列
<220>
<223>切割序列,自切割肽
<400>14
leuleuasnpheaspleuleulysleualaglyaspvalglupheasn
151015
proglypro
<210>15
<211>19
<212>prt
<213>人工序列
<220>
<223>切割序列,自切割肽
<400>15
leuleuasnpheaspleuleulysleualaglyaspvalgluserhis
151015
proglypro
<210>16
<211>19
<212>prt
<213>人工序列
<220>
<223>切割序列,自切割肽
<400>16
leuleuasnpheaspleuleulysleualaglyaspvalgluserglu
151015
proglypro
<210>17
<211>19
<212>prt
<213>人工序列
<220>
<223>切割序列,自切割肽
<400>17
leuleuasnpheaspleuleulysleualaglyaspvalglusergln
151015
proglypro
<210>18
<211>19
<212>prt
<213>人工序列
<220>
<223>切割序列,自切割肽
<400>18
leuleuasnpheaspleuleulysleualaglyaspvalgluserasn
151015
proglygly
<210>19
<211>20
<212>prt
<213>人工序列
<220>
<223>2a样自切割肽
<400>19
tyrhisalaasptyrtyrlysglnargleuilehisaspvalglumet
151015
asnproglypro
20
<210>20
<211>20
<212>prt
<213>人工序列
<220>
<223>2a样自切割肽
<400>20
histyralaglytyrphealaaspleuleuilehisaspilegluthr
151015
asnproglypro
20
<210>21
<211>20
<212>prt
<213>人工序列
<220>
<223>2a样自切割肽
<400>21
glncysthrasntyralaleuleulysleualaglyaspvalgluser
151015
asnproglypro
20
<210>22
<211>19
<212>prt
<213>人工序列
<220>
<223>2a样自切割肽
<400>22
alathrasnpheserleuleulysglnalaglyaspvalglugluasn
151015
proglypro
<210>23
<211>19
<212>prt
<213>人工序列
<220>
<223>2a样自切割肽
<400>23
alaalaargglnmetleuleuleuleuserglyaspvalgluthrasn
151015
proglypro
<210>24
<211>20
<212>prt
<213>人工序列
<220>
<223>2a样自切割肽
<400>24
argalagluglyargglyserleuleuthrcysglyaspvalgluglu
151015
asnproglypro
20
<210>25
<211>20
<212>prt
<213>人工序列
<220>
<223>2a样自切割肽
<400>25
thrargalagluilegluaspgluleuileargalaglyilegluser
151015
asnproglypro
20
<210>26
<211>20
<212>prt
<213>人工序列
<220>
<223>2a样自切割肽
<400>26
thrargalagluilegluaspgluleuileargalaaspilegluser
151015
asnproglypro
20
<210>27
<211>20
<212>prt
<213>人工序列
<220>
<223>2a样自切割肽
<400>27
alalyspheglnileasplysileleuileserglyaspvalgluleu
151015
asnproglypro
20
<210>28
<211>20
<212>prt
<213>人工序列
<220>
<223>2a样自切割肽
<400>28
serserileileargthrlysmetleuvalserglyaspvalgluglu
151015
asnproglypro
20
<210>29
<211>20
<212>prt
<213>人工序列
<220>
<223>2a样自切割肽
<400>29
cysaspalaglnargglnlysleuleuleuserglyaspileglugln
151015
asnproglypro
20
<210>30
<211>20
<212>prt
<213>人工序列
<220>
<223>2a样自切割肽
<400>30
tyrproileasppheglyglypheleuvallysalaaspsergluphe
151015
asnproglypro
20
<210>31
<211>57
<212>prt
<213>人工序列
<220>
<223>自切割肽
<400>31
valthrgluleuleutyrargmetlysargalagluthrtyrcyspro
151015
argproleualailehisprothrglualaarghislysglnlysile
202530
valalaprovallysglnthrleuasnpheaspleuleulysleuala
354045
glyaspvalgluserasnproglypro
5055
<210>32
<211>40
<212>prt
<213>人工序列
<220>
<223>自切割肽
<400>32
leuleualailehisprothrglualaarghislysglnlysileval
151015
alaprovallysglnthrleuasnpheaspleuleulysleualagly
202530
aspvalgluserasnproglypro
3540
<210>33
<211>33
<212>prt
<213>人工序列
<220>
<223>自切割肽
<400>33
glualaarghislysglnlysilevalalaprovallysglnthrleu
151015
asnpheaspleuleulysleualaglyaspvalgluserasnprogly
202530
pro
<210>34
<211>24
<212>prt
<213>人工序列
<220>
<223>自切割肽
<400>34
alaprovallysglnthrleuasnpheaspleuleulysleualagly
151015
aspvalgluserasnproglypro
20
<210>35
<211>20
<212>prt
<213>小鼠
<400>35
metgluthraspthrleuileleutrpvalleuleuleuleuvalpro
151015
glyserthrgly
20
<210>36
<211>25
<212>prt
<213>人工序列
<220>
<223>信号肽,t细胞表面糖蛋白cd4
<400>36
metasnargglyvalprophearghisleuleuleuvalleuglnleu
151015
alaleuleuproalaalathrglngly
2025
<210>37
<211>23
<212>prt
<213>人工序列
<220>
<223>信号肽,白细胞共同抗原
<400>37
mettyrleutrpleulysleuleualapheglyphealapheleuasp
151015
thrgluvalphevalthrgly
20
<210>38
<211>21
<212>prt
<213>人工序列
<220>
<223>信号肽,t细胞表面糖蛋白cd8alpha链
<400>38
metalaleuprovalthralaleuleuleuproleualaleuleuleu
151015
hisalaalaargpro
20
<210>39
<211>21
<212>prt
<213>人工序列
<220>
<223>信号肽,t细胞表面糖蛋白cd8beta链
<400>39
metargproargleutrpleuleuleualaalaglnleuthrvalleu
151015
hisglyasnserval
20
<210>40
<211>24
<212>prt
<213>人工序列
<220>
<223>信号肽,t细胞表面抗原cd2
<400>40
metserpheprocyslysphevalalaserpheleuleuilepheasn
151015
valserserlysglyalavalser
20
<210>41
<211>24
<212>prt
<213>人工序列
<220>
<223>信号肽,t细胞表面糖蛋白cd5
<400>41
metprometglyserleuglnproleualathrleutyrleuleugly
151015
metleuvalalasercysleugly
20
<210>42
<211>25
<212>prt
<213>人工序列
<220>
<223>信号肽,t细胞抗原cd7
<400>42
metalaglyproproargleuleuleuleuproleuleuleualaleu
151015
alaargglyleuproglyalaleuala
2025
<210>43
<211>24
<212>prt
<213>人工序列
<220>
<223>信号肽,5,6-二羟基吲哚-2-羧酸氧化酶
<400>43
metseralaprolysleuleuserleuglycysilephepheproleu
151015
leuleupheglnglnalaargala
20
<210>44
<211>19
<212>prt
<213>人工序列
<220>
<223>信号肽,alpha-乳白蛋白
<400>44
metargphephevalproleupheleuvalglyileleupheproala
151015
ileleuala
<210>45
<211>23
<212>prt
<213>人工序列
<220>
<223>信号肽,beta-半乳糖苷酶
<400>45
metproglypheleuvalargileleuproleuleuleuvalleuleu
151015
leuleuglyprothrarggly
20
<210>46
<211>24
<212>prt
<213>人工序列
<220>
<223>信号肽,campath-1抗原
<400>46
metlysargpheleupheleuleuleuthrileserleuleuvalmet
151015
valglnileglnthrglyleuser
20
<210>47
<211>21
<212>prt
<213>人工序列
<220>
<223>信号肽,cd109抗原
<400>47
metglnglyproproleuleuthralaalahisleuleucysvalcys
151015
thralaalaleuala
20
<210>48
<211>22
<212>prt
<213>人工序列
<220>
<223>信号肽,补体c3
<400>48
metglyprothrserglyproserleuleuleuleuleuleuthrhis
151015
leuproleualaleugly
20
<210>49
<211>18
<212>prt
<213>人工序列
<220>
<223>信号肽,颗粒酶b
<400>49
metglnproileleuleuleuleualapheleuleuleuproargala
151015
aspala
<210>50
<211>20
<212>prt
<213>人工序列
<220>
<223>信号肽,igkappa链v-iii区vh
<400>50
metglualaproalaglnleuleupheleuleuleuleutrpleupro
151015
aspthrthrarg
20
<210>51
<211>20
<212>prt
<213>人工序列
<220>
<223>信号肽,igkappa链v-iv区
<400>51
metvalleuglnthrglnvalpheileserleuleuleutrpileser
151015
glyalatyrgly
20
<210>52
<211>19
<212>prt
<213>人工序列
<220>
<223>信号肽,iglambda链v-vi区eb4
<400>52
metalatrpalaproleuleuleuthrleuleualahiscysthrasp
151015
cystrpala
<210>53
<211>22
<212>prt
<213>人工序列
<220>
<223>信号肽,细胞因子受体共同gamma链
<400>53
metleulysproserleuprophethrserleuleupheleuglnleu
151015
proleuleuglyvalgly
20
<210>54
<211>20
<212>prt
<213>人工序列
<220>
<223>信号肽,杀菌/渗透性增加蛋白样1
<400>54
metalatrpalaserargleuglyleuleuleualaleuleuleupro
151015
valvalglyala
20
<210>55
<211>23
<212>prt
<213>人工序列
<220>
<223>信号肽,igg受体fcrn大亚基p51
<400>55
metglyvalproargproglnprotrpalaleuglyleuleuleuphe
151015
leuleuproglyserleugly
20
<210>56
<211>248
<212>prt
<213>人工序列
<220>
<223>scfvacd19,嵌合抗原受体(car)序列
<400>56
aspileglnmetthrglnthrthrserserleuseralaserleugly
151015
aspargvalthrilesercysargalaserglnaspileserlystyr
202530
leuasntrptyrglnglnlysproaspglythrvallysleuleuile
354045
tyrhisthrserargleuhisserglyvalproserargphesergly
505560
serglyserglythrasptyrserleuthrileserasnleuglugln
65707580
gluaspilealathrtyrphecysglnglnglyasnthrleuprotyr
859095
thrpheglyglyglythrlysleugluilethrlysalaglyglygly
100105110
glyserglyglyglyglyserglyglyglyglyserglyglyglygly
115120125
sergluvallysleuglngluserglyproglyleuvalalaproser
130135140
glnserleuservalthrcysthrvalserglyvalserleuproasp
145150155160
tyrglyvalsertrpileargglnproproarglysglyleuglutrp
165170175
leuglyvaliletrpglysergluthrthrtyrtyrasnseralaleu
180185190
lysserargleuthrileilelysaspasnserlysserglnvalphe
195200205
leulysmetasnserleuglnthraspaspthralailetyrtyrcys
210215220
alalyshistyrtyrtyrglyglysertyralametasptyrtrpgly
225230235240
glnglythrservalthrvalser
245
<210>57
<211>47
<212>prt
<213>人工序列
<220>
<223>人cd8astk
<400>57
prothrthrthrproalaproargproprothrproalaprothrile
151015
alaserglnproleuserleuargproglualacysargproalaala
202530
glyglyalavalhisthrargglyleuaspphealacysaspile
354045
<210>58
<211>27
<212>prt
<213>人工序列
<220>
<223>人cd28tm
<400>58
phetrpvalleuvalvalvalglyglyvalleualacystyrserleu
151015
leuvalthrvalalapheileilephetrpval
2025
<210>59
<211>113
<212>prt
<213>人工序列
<220>
<223>人cd3zeta细胞内结构域
<400>59
argargvallyspheserargseralaaspalaproalatyrglngln
151015
glyglnasnglnleutyrasngluleuasnleuglyargarggluglu
202530
tyraspvalleuasplysargargglyargaspproglumetglygly
354045
lysproargarglysasnproglngluglyleutyrasngluleugln
505560
lysasplysmetalaglualatyrsergluileglymetlysglyglu
65707580
argargargglylysglyhisaspglyleutyrglnglyleuserthr
859095
alathrlysaspthrtyraspalaleuhismetglnalaleupropro
100105110
arg
<210>60
<211>20
<212>prt
<213>人工序列
<220>
<223>2a自切割肽
<400>60
argalagluglyargglyserleuleuthrcysglyaspvalgluglu
151015
asnproglypro
20
<210>61
<211>18
<212>prt
<213>人工序列
<220>
<223>衍生自小鼠igkappa的信号肽
<400>61
metalavalprothrglnvalleuglyleuleuleuleutrpleuthr
151015
aspala
<210>62
<211>254
<212>prt
<213>人工序列
<220>
<223>scfvacd33,嵌合抗原受体(car)序列
<400>62
argcysaspileglnmetthrglnserproserserleuseralaser
151015
valglyaspargvalthrilethrcysargalasergluaspiletyr
202530
pheasnleuvaltrptyrglnglnlysproglylysalaprolysleu
354045
leuiletyraspthrasnargleualaaspglyvalproserargphe
505560
serglyserglyserglythrglntyrthrleuthrileserserleu
65707580
glnprogluaspphealathrtyrtyrcysglnhistyrlysasntyr
859095
proleuthrpheglyglnglythrlysleugluilelysargsergly
100105110
glyglyglyserglyglyglyglyserglyglyglyglyserglygly
115120125
glyglyserargsergluvalglnleuvalgluserglyglyglyleu
130135140
valglnproglyglyserleuargleusercysalaalaserglyphe
145150155160
thrleuserasntyrglymethistrpileargglnalaproglylys
165170175
glyleuglutrpvalserserileserleuasnglyglyserthrtyr
180185190
tyrargaspservallysglyargphethrileserargaspasnala
195200205
lysserthrleutyrleuglnmetasnserleuargalagluaspthr
210215220
alavaltyrtyrcysalaalaglnaspalatyrthrglyglytyrphe
225230235240
asptyrtrpglyglnglythrleuvalthrvalsersermet
245250
<210>63
<211>231
<212>prt
<213>人工序列
<220>
<223>衍生于人igg1的铰链和fc,具有突变以防止fcrg结合(hch2ch3pvaa)
<400>63
gluprolysserproasplysthrhisthrcysproprocysproala
151015
proprovalalaglyproservalpheleupheproprolysprolys
202530
aspthrleumetilealaargthrprogluvalthrcysvalvalval
354045
aspvalserhisgluaspprogluvallyspheasntrptyrvalasp
505560
glyvalgluvalhisasnalalysthrlysproargglugluglntyr
65707580
asnserthrtyrargvalvalservalleuthrvalleuhisglnasp
859095
trpleuasnglylysglutyrlyscyslysvalserasnlysalaleu
100105110
proalaproileglulysthrileserlysalalysglyglnproarg
115120125
gluproglnvaltyrthrleuproproserargaspgluleuthrlys
130135140
asnglnvalserleuthrcysleuvallysglyphetyrproserasp
145150155160
ilealavalglutrpgluserasnglyglnprogluasnasntyrlys
165170175
thrthrproprovalleuaspseraspglyserphepheleutyrser
180185190
lysleuthrvalasplysserargtrpglnglnglyasnvalpheser
195200205
cysservalmethisglualaleuhisasnhistyrthrglnlysser
210215220
leuserleuserproglylys
225230
<210>64
<211>4
<212>prt
<213>人工序列
<220>
<223>接头
<400>64
lysaspprolys
1
<210>65
<211>21
<212>prt
<213>人工序列
<220>
<223>人cd148tm
<400>65
alavalpheglycysilepheglyalaleuvalilevalthrvalgly
151015
glypheilephetrp
20
<210>66
<211>341
<212>prt
<213>人工序列
<220>
<223>人cd148细胞内结构域
<400>66
arglyslysarglysaspalalysasnasngluvalserphesergln
151015
ilelysprolyslysserlysleuileargvalgluasnphegluala
202530
tyrphelyslysglnglnalaaspserasncysglyphealagluglu
354045
tyrgluaspleulysleuvalglyileserglnprolystyralaala
505560
gluleualagluasnargglylysasnargtyrasnasnvalleupro
65707580
tyraspileserargvallysleuservalglnthrhisserthrasp
859095
asptyrileasnalaasntyrmetproglytyrhisserlyslysasp
100105110
pheilealathrglnglyproleuproasnthrleulysaspphetrp
115120125
argmetvaltrpglulysasnvaltyralaileilemetleuthrlys
130135140
cysvalgluglnglyargthrlyscysgluglutyrtrpproserlys
145150155160
glnalaglnasptyrglyaspilethrvalalametthrsergluile
165170175
valleuproglutrpthrileargaspphethrvallysasnilegln
180185190
thrsergluserhisproleuargglnphehisphethrsertrppro
195200205
asphisglyvalproaspthrthraspleuleuileasnpheargtyr
210215220
leuvalargasptyrmetlysglnserproprogluserproileleu
225230235240
valhiscysseralaglyvalglyargthrglythrpheilealaile
245250255
aspargleuiletyrglnilegluasngluasnthrvalaspvaltyr
260265270
glyilevaltyraspleuargmethisargproleumetvalglnthr
275280285
gluaspglntyrvalpheleuasnglncysvalleuaspilevalarg
290295300
serglnlysaspserlysvalaspleuiletyrglnasnthrthrala
305310315320
metthriletyrgluasnleualaprovalthrthrpheglylysthr
325330335
asnglytyrileala
340
<210>67
<211>4
<212>prt
<213>人工序列
<220>
<223>弗林蛋白酶切割位点
<220>
<221>misc_feature
<222>(2)..(2)
<223>xaa可以是任意天然存在的氨基酸
<400>67
argxaaargarg
1
<210>68
<211>4
<212>prt
<213>人工序列
<220>
<223>弗林蛋白酶切割位点
<220>
<221>misc_feature
<222>(2)..(2)
<223>xaa可以是任意天然存在的氨基酸
<400>68
argxaalysarg
1
<210>69
<211>7
<212>prt
<213>人工序列
<220>
<223>烟草蚀刻病毒(tev)切割位点
<400>69
gluasnleutyrpheglnser
15
<210>70
<211>8
<212>prt
<213>人工序列
<220>
<223>切割位点基序
<220>
<221>misc_feature
<222>(2)..(2)
<223>xaa可以是任意天然存在的氨基酸
<220>
<221>misc_feature
<222>(4)..(4)
<223>xaa可以是任意天然存在的氨基酸
<400>70
aspxaagluxaaasnproglypro
15
<210>71
<211>8
<212>prt
<213>人工序列
<220>
<223>切割位点基序
<220>
<221>misc_feature
<222>(2)..(2)
<223>xaa可以是ile,val,met或ser
<220>
<221>misc_feature
<222>(4)..(4)
<223>xaa可以是thr,met,ser,leu,glu,gln或phe
<400>71
aspxaagluxaaasnproglypro
15
<210>72
<211>4
<212>prt
<213>小鼠
<400>72
metgluthrasp
1
<210>73
<211>11
<212>prt
<213>小鼠
<400>73
thrleuileleutrpvalleuleuleuleuval
1510
<210>74
<211>5
<212>prt
<213>小鼠
<400>74
proglyserthrgly
15
- 还没有人留言评论。精彩留言会获得点赞!