基于KASP技术用于水稻产量基因分型的引物组合及其应用的制作方法
本发明涉及分子生物学及作物育种
技术领域:
,具体地说,涉及一种基于kasp技术用于水稻产量基因分型的引物组合及其应用。
背景技术:
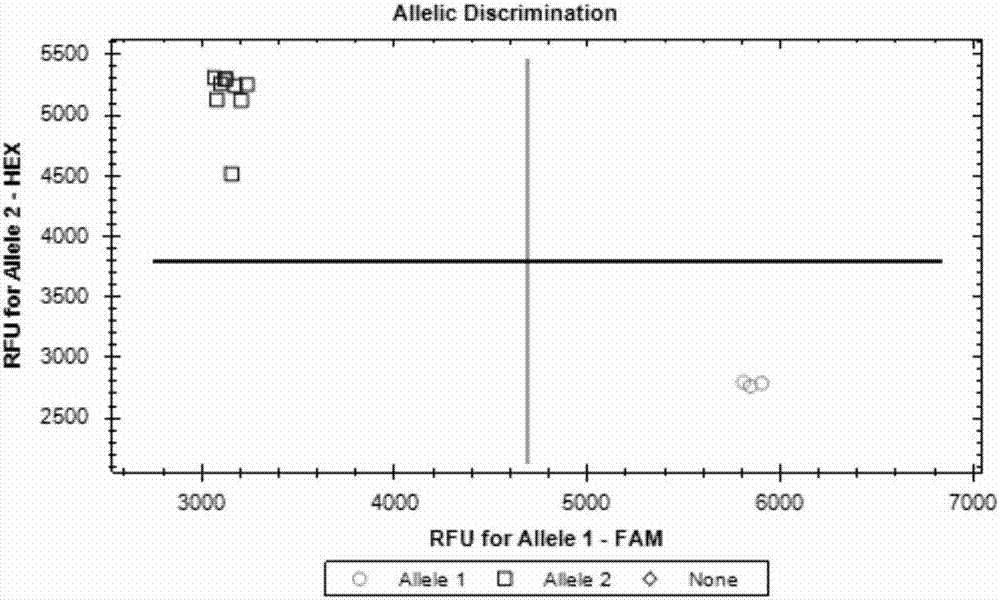
:近年来,育种家通过系谱法育成大量的高产水稻品种。但是传统杂交和回交选择往往需要大规模的群体,开展大量的表型考察工作,并且许多表型考察难度大,这样极大地增加了育种的工作量和成本。随着水稻重要基因的克隆,开发基因功能位点的分子标记,检测育种材料中目标性状的基因组合类型,根据特定的育种改良目标,导入相应优良基因,培育新品种,已经成为可能。另外,育种过程中许多性状如产量性状,易受环境影响,不容易准确调查,而利用控制该性状的重要基因等位变异组合进行鉴别,可以代替性状表型考察,且不受环境影响,增加了选择的准确性,提高了选择效率,加速育种进程。水稻产量是由数量性状位点控制的。目前,通过遗传分离群体已克隆了多个主效产量基因,如gn1a、nal1、ghd7、hd1、ghd7.1、ghd8。这些基因序列均存在广泛的自然变异。如gn1a编码区第1605位的碱基变异导致氨基酸变化,依据该单核苷酸多态性(singlenucleotidepolymorphism,snp)可将核心种质材料gn1a分作两种单倍型,二者存在极显著的产量差异。ghd7.1编码区域一个8bp缺失,造成氨基酸翻译移码,导致基因功能丧失。因此,利用导致基因功能变异的碱基位点开发标记,进行育种材料的有利等位变异鉴定,在水稻高产品种培育中具有重要作用。然而,传统的连锁标记主要是基于与基因连锁的分子标记进行基因型鉴定,不能用来准确鉴定材料目标性状有利基因单倍型组合以及进行定向的改良育种,并且操作复杂、通量小、劳动力消耗量大。另外,由于不同基因增加水稻产量的作用途径不同,如gn1a主要改良穗型,增加每穗颖花数,nal1主要增加叶片光合作用面积,改良株型,而hd1、ghd7、ghd7.1、ghd8能够同时改良多个性状(每穗颖花数,开花期等),因此,可以通过同时改良多个产量构成因子,进行高产育种。因而,开发成套相关基因的功能标记,可以利用分子标记辅助选择(marker-assistedselection,mas)技术针对不同育种材料作某个性状的定向改良,也可以进行多性状的聚合改良。kasp基因分型技术是一项独特的竞争型等位基因特异性pcr,可对各种基因组核酸样品进行snps和indels(insertiondeletion)高精度双等位基因分型。kasp技术操作简单,分析稳定、准确,并且成本较低。水稻基因组大小约400mb,经探索发现20-50ngdna为最适宜的模板用量,利用上述6个基因功能位点开发的分子标记,对育种材料的产量基因位点进行有利等位基因的鉴别以及组合筛选,进行特定有利基因的转移和聚合,可提高产量水平。从而可以避免传统育种中的盲目性,极大地减小群体规模,节省成本,同时增加选择的准确性和效率。技术实现要素:本发明的目的是提供一套基于kasp技术用于水稻产量基因分型的引物组合及其应用。为了实现本发明目的,本发明提供一套基于kasp技术开发的用于水稻产量基因分型的引物组合,其中所述水稻产量基因包括gn1a、nal1、ghd7、hd1、ghd7.1和ghd8,它们的功能性分子标记分别对应于表1中m1~m6,信息如下:表1编号分子标记类型物理位置等位基因m1snploc_os01g10110基因编码区第1605位[t/g]m2snploc_os04g52479基因编码区第698位[a/g]m3indelsloc_os06g1637基因编码区第1053-1056位[aaag/-]m4snploc_os07g15770基因编码区第455位[a/g]m5indelsloc_os07g49460基因编码区第1516-1523位[aacgttgt/-]m6snploc_os08g07740基因编码区第566位[a/c]基于kasp技术的用于检测上述功能性分子标记的引物组合,所述引物组合的信息见表2:表2本发明还提供含有所述引物组合的用于水稻产量基因分型的检测试剂盒。本发明还提供所述引物组合或试剂盒在改良水稻品种中的应用。本发明还提供所述引物组合或试剂盒在鉴定高产水稻品种中的应用。本发明还提供所述引物组合或试剂盒在水稻分子标记辅助育种中的应用。本发明进一步提供所述引物组合或试剂盒在水稻产量基因等位功能变异检测中的应用。所述应用包括以下步骤:1)提取待测水稻样品的dna;2)取模板dna2.5μl,引物混合液0.07μl,2×kaspmastermix2.5μl,进行pcr扩增;引物混合液中引物fam-f、引物hex-f终浓度均为12μm,引物r终浓度为25μm;3)采用荧光检测仪分析pcr扩增产物单倍型。pcr反应条件为:94℃预变性15分钟;第一步扩增反应:94℃变性20秒,61℃-55℃梯度退火并延伸60秒,10个循环,每个循环退火及延伸的温度降低0.6℃;第二步扩增反应,94℃变性20秒,55℃退火并延伸60秒,32个循环。本发明具有以下优点:(一)利用本发明提供的成套kasp功能标记及其引物组合能够快速检测水稻种质中产量基因的功能等位基因的组合方式,检测结果准确可靠,操作简单,成本低。(二)本发明方法能够实现水稻育种材料的重要产量功能基因的检测,为水稻高产新品种选育、改良提供科学指导。附图说明图1-图6分别为本发明实施例2中6个基因功能位点标记gn1a、nal1、hd1、ghd7、ghd7.1和ghd8在12份材料间的基因型簇图。图7为本发明实施例3中日本晴/珍汕97育种改良路线图。图8为本发明实施例4中明恢63/珍汕97育种改良路线图。具体实施方式以下实施例用于说明本发明,但不用来限制本发明的范围。若未特别指明,实施例均按照常规实验条件,如sambrook等分子克隆实验手册(sambrookj&russelldw,molecularcloning:alaboratorymanual,2001),或按照制造厂商说明书建议的条件。实施例1水稻重要产量基因功能标记的开发1、根据文献ashikarim,sakakibarah,lins,etal.cytokininoxidaseregulatesricegrainproduction[j].science,2005,309(5735):741-745.;wangj,xuh,lin,etal.artificialselectionofgn1aplaysanimportantroleinimprovingriceyieldsacrossdifferentecologicalregions[j].rice,2015,8(1):37。gn1a对应于msu基因编号:loc_os01g10110,该基因编码区第1605位snp:t/g变异直接导致基因功能差异。2、根据文献fujitad,trijatmikokr,tagleag,etal.nal1allelefromaricelandracegreatlyincreasesyieldinmodernindicacultivars[j].proceedingsofthenationalacademyofsciences,2013,110(51):20431-20436。nal1对应于msu基因编号:loc_os04g52479,针对该基因编码区第698位snp:a/g变异直接导致基因功能差异。3、根据文献takahashiy,teshimakm,yokois,etal.variationsinhd1proteins,hd3apromoters,andehd1expressionlevelscontributetodiversityoffloweringtimeincultivatedrice[j].proceedingsofthenationalacademyofsciences,2009,106(11):4555-4560。hd1对应于msu基因编号:loc_os06g1637,针对该基因编码区第1053-1056位indels:aaag/-变异直接导致基因功能差异。4、根据文献xuew,xingy,wengx,etal.naturalvariationinghd7isanimportantregulatorofheadingdateandyieldpotentialinrice[j].naturegenetics,2008,40(6):761-767。ghd7对应于msu基因编号:loc_os07g15770,针对该基因编码区第455位snp:a/g变异直接导致基因功能差异。5、根据文献yanw,liuh,zhoux,etal.naturalvariationinghd7.1playsanimportantroleingrainyieldandadaptationinrice[j].cellresearch,2013,23(7):969。ghd7.1对应于msu基因编号:loc_os07g49460,针对该基因编码区第1516-1523位indels:aacgttgt/-变异直接导致基因功能差异。6、根据文献yanw,wangp,chenhx,etal.amajorqtl,ghd8,playspleiotropicrolesinregulatinggrainproductivity,plantheight,andheadingdateinrice[j].molecularplant,2011,4(2):319-330。ghd8对应于msu基因编号:loc_os08g07740,针对该基因编码区第566位snp:a/c变异直接导致基因功能差异。提取1-6中snp或indels位点两侧100bp,利用primer5.0设计引物,委托生工生物工程(上海)股份有限公司合成,采用高纯度page方式纯化。引物信息见表3。表3gn1a对应于msu基因编号:loc_os01g10110,针对水稻种质中该基因编码区第1605位snp:t/g,特异的kasp引物:引物1、引物2和引物3;引物1依次为fam标签序列和gn1a有功能基因dna序列,引物2依次为hex标签序列和gn1a无功能基因dna序列,引物3为共用序列,序列如表3中引物3所示单链dna。nal1对应于msu基因编号:loc_os04g52479,针对水稻种质中该基因编码区第698位snp:a/g,特异的kasp引物:引物4、引物5和引物6;引物4依次为fam标签序列和nal1有功能基因dna序列,引物5依次为hex标签序列和nal1无功能基因dna序列,引物6为共用序列,序列如表3中引物6所示单链dna。hd1对应于msu基因编号:loc_os06g1637,针对水稻种质中该基因编码区第1053-1056位indels:aaag/-,特异的kasp引物:引物7、引物8和引物9;引物7依次为fam标签序列和hd1有功能基因dna序列,引物8依次为hex标签序列和hd1无功能基因dna序列,引物9为共用序列,序列如表3中引物9所示单链dna。ghd7对应于msu基因编号:loc_os07g15770,针对水稻种质中该基因编码区第455位snp:a/g,特异的kasp引物:引物10、引物11和引物12;引物10依次为fam标签序列和ghd7有功能基因dna序列,引物11依次为hex标签序列和ghd7无功能基因dna序列,引物12为共用序列,序列如表3中引物12所示单链dna。ghd7.1对应于msu基因编号:loc_os07g49460,针对水稻种质中该基因编码区第1516-1523位indels:aacgttgt/-,特异的kasp引物:引物13、引物14和引物15;引物13依次为fam标签序列和ghd7.1有功能基因dna序列,引物14依次为hex标签序列和ghd7.1无功能基因dna序列,引物15为共用序列,序列如表3中引物15所示单链dna。ghd8对应于msu基因编号:loc_os08g07740,针对水稻种质中该基因编码区第566位snp:a/c,特异的kasp引物:引物16、引物17和引物18;引物16依次为fam标签序列和ghd8有功能基因dna序列,引物17依次为hex标签序列和ghd8无功能基因dna序列,引物18为共用序列,序列如表3中引物18所示单链dna。所述标签fam的核苷酸序列为gaaggtgaccaagttcatgct;所述标签hex的核苷酸序列为gaaggtcggagtcaacggatt。实施例2水稻重要产量基因功能标记的多态性筛选1、针对6个水稻重要产量基因的功能位点分子标记,对包括非洲栽培稻、籼稻(包括生产上广泛推广品种黄华占、93-11等)、粳稻、普通野生稻在内的12份水稻种质材料进行kasp反应,测试标记分型情况。pcr扩增反应使用的体系为5μl:模板dna2.5μl,引物混合液0.07μl,2×kaspmastermix2.5μl,引物fam-f、引物hex-f终浓度均为12μm,引物r终浓度为25μm。pcr扩增反应的条件为:94℃预变性15分钟;第一步扩增反应:94℃变性20秒,61℃-55℃梯度退火并延伸60秒,10个循环,每个循环退火及延伸的温度降低0.6℃;第二步扩增反应,94℃变性20秒,55℃退火并延伸60秒,32个循环。pcr反应结束后,利用bio-radcfx荧光定量pcr仪对kasp反应产物进行荧光数据读取,荧光扫描的结果导出成列表格式,即得到表4中基因分型结果。结果显示,6个产量基因标记在12份材料存在多态性,6个基因功能位点标记在12份材料间基因型簇图分别见图1-图6。针对生产上广泛推广的水稻品种黄华占进行6个产量基因有利等位变异分析,发现黄华占拥有5个产量基因有利等位变异,可以定向增加nal1有利等位变异,提高颖花数,改良株型。而珍汕97中产量基因有利等位基因较少,可以添加日本晴来源的nal1、ghd7.1、ghd8和明恢63来源的hd1进行产量改良。表4测试种质材料分型结果水稻种质gn1a_kaspnal1_kasphd1_kaspghd7_kaspghd7.1_kaspghd8_kasp黄华占allele2allele2allele1allele2allele1allele2广西红米allele2allele2allele1allele2allele1allele12-33allele2allele2allele1allele2allele1allele1cypressallele2allele1allele2allele1allele1allele2kasalathallele2allele2allele2allele2allele1allele1中花11allele1allele1allele2allele1allele1allele293-11allele2allele2allele1allele2allele2allele1日本晴allele1allele1allele2allele1allele1allele2acc9allele2allele2allele2allele2allele1allele1acc10allele2allele2allele2allele2allele1allele1明恢63allele2allele2allele1allele2allele1allele1珍汕97allele2allele2allele2noneallele2allele1有利等位型allele2allele1allele1allele2allele1allele2注:表4中gn1a_kasp对应的allele1、2分别为t、g,nal1_kasp对应的allele1、2分别为a、g,hd1_kasp对应的allele1、2分别为-和aaag,ghd7_kasp对应的allele1、2分别为a、g,ghd7.1_kasp对应的allele1、2分别为aacgttgt和-,ghd8_kasp对应的allele1、2分别为a、c。none表示基因缺失。实施例3kasp标记在改良水稻种质珍汕97中的应用1、根据表4水稻种质产量基因分型结果,珍汕97(zs97b)仅在产量基因gn1a含有有利变异,为此,选择gn1a、nal1、ghd7.1与ghd8产量基因存在差异的种质材料日本晴与zs97b进行杂交,定向选择改良zs97b。2、利用gn1a-kasp选择日本晴与zs97b真杂种f1,与zs97b回交收获bc1f1,分别选择nal1、ghd7.1与ghd8基因杂合单株种植,与zs97b回交收获bc2f1。3、分别选择nal1、ghd7.1与ghd8基因杂合单株种植,与zs97b回交收获bc3f1。4、分别选择nal1、ghd7.1与ghd8基因杂合单株种植,与zs97b回交收获bc4f1。5、将nal1、ghd7.1与ghd8基因杂合单株种植,分别收获200粒自交种,将bc4f2种植于大田中,进行表型值测定,采取完全随机区组种植,育种改良路线图见图7。6、同时抽提bc4f2苗期叶片dna,进行基因分型,nal1-kasp、ghd7.1-kasp与ghd8-kasp可以将3个bc4f2群体分为3种基因型,即目标基因纯合zs97b,纯合日本晴,杂合型。分别选取nal1、ghd7.1与ghd8纯合基因型,考察产量性状见表5~表7。日本晴来源的nal1、ghd7.1与ghd8等位基因都能够增加zs97b每穗颖花数,nal1与ghd7.1显著提高单株产量。表5nal1功能等位变异产量性状近等基因系表型nil_nal1含有nal1优势单倍型基因,nil_nal1含有劣势单倍型基因。**表示在0.01水平上差异显著,下同。表6ghd7.1功能等位变异产量性状近等基因系表型nil_ghd7.1含有ghd7.1优势单倍型基因,nil_ghd7.1含有劣势单倍型基因。表7ghd8功能等位变异产量性状近等基因系表型nil_ghd8含有ghd8优势单倍型基因,nil_ghd8含有劣势单倍型基因。实施例4kasp标记在改良水稻种质zs97b中的应用根据12份水稻种质产量基因分型结果,选择含有hd1有利等位基因型材料明恢63(mh63)参照图8进行zs97b改良。利用mh63与zs97b进行杂交,利用标记hd1_kasp鉴定真杂种f1,f1与zs97b回交收获bc1f1,选择hd1基因杂合单株种植,与zs97b继续回交收获bc2f1,选择hd1基因杂合单株种植,与zs97b回交收获bc3f1,选择hd1基因杂合单株种植,与zs97b回交收获bc4f1,将hd1基因杂合单株种植,收获200粒自交种,bc4f2种植于大田中,进行表型值测定。同时抽提bc4f2苗期叶片抽提dna,进行基因分型,标记hd1-kasp可以将bc4f2群体分为3种基因型,基因型1:纯合zs97b;基因型2:纯合minghui63;基因型3:杂合型。选取hd1纯合基因型,考察产量性状见表8,mh63来源的hd1等位变异能够增加穗长、一次枝梗数、颖花数、千粒重,改良株高和抽穗期。表8hd1功能等位变异产量性状近等基因系表型虽然,上文中已经用一般性说明及具体实施方案对本发明作了详尽的描述,但在本发明基础上,可以对之做一些修改或改进,这对本领域技术人员而言是显而易见的。因此,在不偏离本发明精神的基础上所做的这些修改或改进,均属于本发明要求保护的范围。序列表<110>华中农业大学<120>基于kasp技术用于水稻产量基因分型的引物组合及其应用<130>khp171112519.8<160>18<170>patentinversion3.3<210>1<211>43<212>dna<213>人工序列<400>1gaaggtgaccaagttcatgctgaagcggcacttcggtgccaat43<210>2<211>43<212>dna<213>人工序列<400>2gaaggtcggagtcaacggattgaagcggcacttcggtgccaag43<210>3<211>22<212>dna<213>人工序列<400>3cgctgcacgaatcttggccaga22<210>4<211>45<212>dna<213>人工序列<400>4gaaggtgaccaagttcatgctgcaggttggtttcctcactaacca45<210>5<211>45<212>dna<213>人工序列<400>5gaaggtcggagtcaacggattgcaggttggtttcctcactaaccg45<210>6<211>24<212>dna<213>人工序列<400>6gtagtccaagtcaaccgcgacatg24<210>7<211>46<212>dna<213>人工序列<400>7gaaggtgaccaagttcatgctgacaatacgttatgaaacaaggcgt46<210>8<211>51<212>dna<213>人工序列<400>8gaaggtcggagtcaacggattgagaagacaatacgttatgaaacaagaaag51<210>9<211>14<212>dna<213>人工序列<400>9ggtcgtgcctctgc14<210>10<211>36<212>dna<213>人工序列<400>10gaaggtgaccaagttcatgctcgtctccctcggcga36<210>11<211>36<212>dna<213>人工序列<400>11gaaggtcggagtcaacggattcgtctccctcggcgg36<210>12<211>15<212>dna<213>人工序列<400>12gtccaggtcgggccg15<210>13<211>50<212>dna<213>人工序列<400>13gaaggtgaccaagttcatgctgacatgggttccactacaaagaacgttgt50<210>14<211>48<212>dna<213>人工序列<400>14gaaggtcggagtcaacggattcatgggttccactacaaaggacaaagc48<210>15<211>32<212>dna<213>人工序列<400>15agggtgacattactctctccttatttgtacta32<210>16<211>37<212>dna<213>人工序列<400>16gaaggtgaccaagttcatgctccgccggcgagcgggg37<210>17<211>37<212>dna<213>人工序列<400>17gaaggtcggagtcaacggattccgccggcgagcgggt37<210>18<211>20<212>dna<213>人工序列<400>18ggcgtcgtcatcgacgggca20当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1