马铃薯样磷脂酶7的应用的制作方法
本发明属于生物医学领域,具体涉及马铃薯样磷脂酶7蛋白在非酒精性脂肪肝病及高脂血症中的应用。
背景技术:
随着饮食结构的改变,营养过剩引起的肥胖和非酒精性脂肪肝病(nonalcoholicfattyliverdisease,nafld)已经成为一种全球流行性疾病。肥胖和nafld发病率的提高将直接导致慢性肝病的发生增多。
nafld是一种除酒精及其他确定的肝损因素如酒精、药物等所导致的肝脏脂质过度积累的病理综合症,是肝脏脂质堆积后发生的一系列病理过程。我国nafld的发病率逐年提升,脂肪肝病会进一步引起肝脏纤维化、肝硬化以及肝癌,严重威胁着人们的健康。肝脏中过多的甘油三酯(triacylglycerol,tag)通过极低密度脂蛋白(verylowdensitylipoproteins,vldl)分泌到血液中,肝脏vldl分泌紊乱导致nafld或高脂血症。血液中的vldl通过其表面的载脂蛋白apoe和apob被外周组织细胞表面的低密度脂蛋白受体(lowdensitylipoproteinreceptor,ldlr)识别,从而被外周组织吸收利用。肝脏vldl分泌紊乱导致nafld或高脂血症,但是其具体调控机制仍不清楚。前期研究发现肝脏中apoe的缺失抑制vldl分泌,从而导致脂肪肝的发生。
马铃薯样磷脂酶(patatin-likephospholipasedomaincontainingprotein,pnpla)家族蛋白是一类含有保守的磷脂酶结构域的蛋白家族,与脂代谢关系密切。如pnpla2是脂滴(lipiddroplets,ld)膜蛋白,是水解tag的限速酶,可以促进脂肪组织以及肝脏的脂动员;pnpla3氨基酸序列的第148位异亮氨酸残基突变为甲硫氨酸时可以引起严重的nafld。马铃薯样磷脂酶7(patatin-likephospholipasedomaincontainingprotein7,pnpla7,nm_146251.4)是一个在脂肪组织、肝脏和肌肉组织中高表达,体外具有溶血磷脂酶水解活性,但体内生理功能未知的pnpla家族蛋白。前期研究发现pnpla7蛋白是一个氨基末端具有一次跨膜结构的内质网蛋白,同时其羧基末端的磷脂酶结构域和ld相互作用。
技术实现要素:
解决的技术问题:本发明提供一种马铃薯样磷脂酶7的应用,作为治疗非酒精性脂肪肝病及其伴随的高脂血症的分子靶点,马铃薯样磷脂酶7可应用于制备治疗非酒精性脂肪肝病及其伴随的高脂血症药物。
技术方案:马铃薯样磷脂酶7在制备治疗非酒精性脂肪肝病预防及治疗药物中的应用。
降低马铃薯样磷脂酶7基因表达的腺病毒或慢病毒在制备治疗高脂血症药物中的应用。
所述腺病毒所针对的敲降序列如seqidno.1所示。
所述慢病毒所针对的敲降序列如seqidno.2所示。
马铃薯样磷脂酶7在制备筛选治疗与检测非酒精性脂肪肝以及高脂血症试剂盒中的应用。
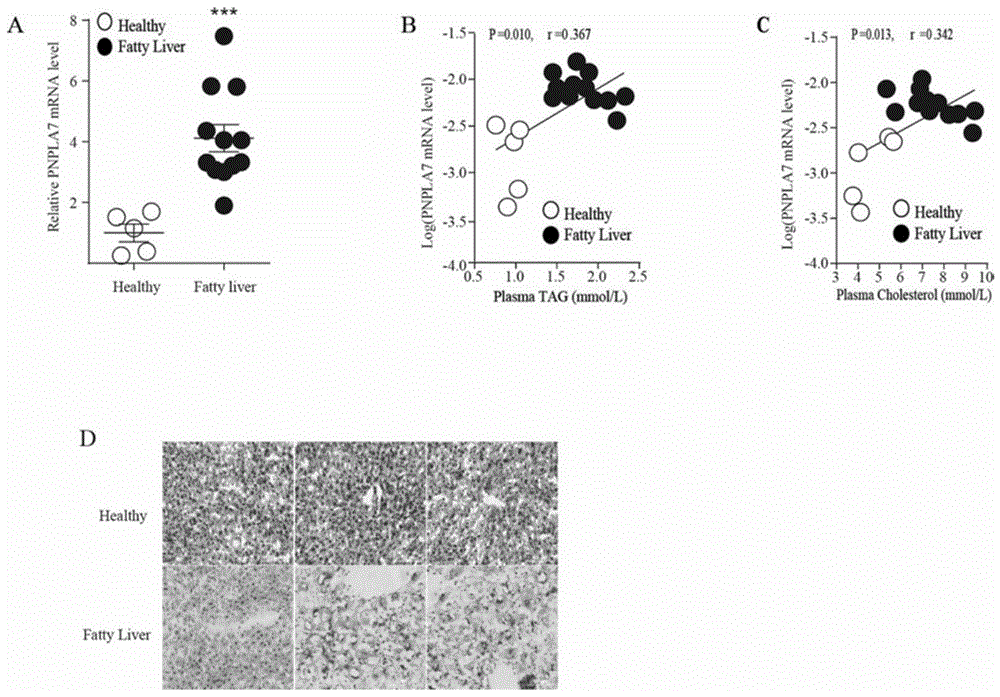
有益效果:本发明首先发现脂肪肝人体肝脏组织样本与正常人肝脏组织样本中pnpla7表达量相比较,显著升高(图1-a),且pnpla7表达量与机体血液中tag含量、tc含量呈正相关(图1-b和c)。如图1d所示,脂肪肝人体样本的肝脏组织进行油红染色后,显示其肝脏脂质含量显著升高。本发明成功在db/db小鼠以及c57bl/6j小鼠肝脏敲降pnpla7图2中a和b显示了腺病毒尾静脉注射后db/db小鼠肝脏pnpla7的敲降效率。我们通过尾静脉注射lpl的抑制剂wr1339后检测了pnpla7敲降对小鼠肝脏vldl分泌的影响。如图3-a所示,与对照组相比,不论是正常饮食还是高脂饮食条件下,肝脏中敲降pnpla7后都可以显著抑制vldl的分泌。如图3-b所示血液的fplc分析结果显示敲降pnpla后,正常饮食与高脂饮食组在vldl组分处的tag含量均显著降低,ldl和hdl处的tag含量无显著差异;vldl、ldl和hdl组分处的tc含量无明显变化,我们又将血清加入15%碘克沙醇中,超速离心取最上层vldl组分,并进行wb分析,结果显示vldl组分中的apob含量显著减少(图3-c);将这一组分进行负染,电镜观察pnpla7缺失后血清中vldl直径及数目。我们对vldl直径进行了统计发现,与对照组比较pnpla7敲降小鼠血清中的大直径vldl颗粒显著减少(图3-d)。如图4中a和b所示,成功分离肝脏内质网,高尔基体,并且发现高尔基体内tag含量显著减少,细胞质内tag含量显著增多。apoe敲除鼠原代肝细胞分泌的vldl-tag含量显著降低(图5-a),胞内tag含量显著升高(图5-b),且其apoe敲除效率达到99%。敲降pnpla7后的原代肝细胞分泌到胞外的tag含量显著降低,回补apoe蛋白后,分泌到胞外的tag含量恢复到正常水平(图5-c)。敲降pnpla7后的原代肝细胞胞内的tag含量显著升高,回补apoe蛋白后,胞内的tag含量恢复到正常水平(图5-d)。且敲降pnpla7与回补apoe在蛋白水平均成功(图5-e)。在原代肝细胞中pnpla7与apoe有显著相互作用(图6-a)。在huh7细胞中pnpla7过表达可以稳定apoe(图6-b),且不改变apoe的mrna水平(图6-c)。在原代肝细胞以及huh7细胞中pnpla7均能改变apoe的半衰期(图6-d)。apoe的蛋白降解途径为蛋白酶体途径(图6-e)。在293t细胞中过表达pnpla7可以降低apoe蛋白的泛素化水平(图6-f),在原代肝细胞敲降pnpla7可以增加apoe的泛素化水平(图6-g)。
附图说明
图1为pnpla7在脂肪肝病人中的表达,(a)人体脂肪肝病人中pnpla7mrna表达量;(b)人体脂肪肝病人的肝脏中pnpla7表达量与血液tag含量成正相关;(c)人体脂肪肝病人的肝脏中pnpla7表达量与血液tc含量成正相关;(d)脂肪肝病人与正常人体肝脏油红染色。
图2为利用高压尾静脉注射db/db小鼠以及c57bl/6j小鼠7天后,肝脏pnpla7的敲降效率(a)db/db小鼠肝脏蛋白,(b)c57bl/6j小鼠肝脏蛋白;
图3说明c57bl/6j小鼠肝脏vldl分泌减少;(a)小鼠尾静脉注射腺病毒敲降5天后,饥饿8小时,然后尾静脉注射wr-1339后按照时间点收取小鼠血液,检测血液中tag含量。(b)小鼠尾静脉注射腺病毒敲降7天后,饥饿16小时后取血液,进行fplc检测。(c)(b)图中的血液配成15%碘克沙醇,进行密度梯度离心,取最上层密度小于1.006的组分即为vldl组分,wb检测组分中apob含量;(d)电镜观察组分中vldl颗粒大小及数量,并进行统计。
图4为分离pnpla7敲降后的肝脏组织高尔基体、内质网以及细胞质,并检测其tag含量。(a)小鼠肝脏细胞器分离(具体实验步骤见材料与方法),并利用westernblot检测各组分的标记蛋白,其中gm130为高尔基体的marker;grp78为内质网的marker;apob为内质网合成并运输到高尔基体的组分,因此在内质网与高尔基体都有分布;(b)将(a)图中的高尔基体组分与内质网组分分别合并,离心后检测高尔基体与内质网以及细胞质中的tag含量。
图5说明pnpla7通过apoe调控肝脏vldl分泌。(a和b)分离8周c57bl/6j小鼠以及apoe-/-小鼠原代肝细胞,细胞计数后按照6孔板5×105个细胞/孔种板,细胞贴壁2h后换为无酚红的完全培养基继续培养24h。处理结束后收集培养液以及细胞,分别检测tag含量,并用蛋白浓度校正。(c和d)分离8周c57bl/6j小鼠原代肝细胞,细胞计数后按照6孔板5×105个细胞/孔种板,细胞贴壁2h后感染ad-shpnpla7腺病毒以及对照腺病毒6h后换为完全培养基培养6h;然后利用lipo2000转染flag-apoe质粒,转染6h后换为完全培养基培养6h;换为无酚红的完全培养基继续培养24h。处理结束后收集培养液以及细胞,分别检测tag含量,并用蛋白浓度校正。(e)收取原代肝细胞蛋白,利用westernblot的方法检测细胞内pnpla7以及apoe的蛋白表达量。
图6说明pnpla7可以稳定apoe蛋白。(a)原代肝细胞中过表达flag-pnpla7,并用flag标签抗体将其沉淀,检测apoe,apob蛋白是否被同时沉淀。(b和c)在huh7细胞中过表达或敲降pnpla7后,检测apoe蛋白表达量及mrna表达量。(d)在huh7细胞中敲降或过表达pnpla7后,chx处理后检测apoe蛋白表达并计算其半衰期。(e)293t细胞中利用不同的蛋白酶抑制剂以及蛋白酶体抑制剂、自噬体抑制剂处理细胞后,chx处理2h检测apoe蛋白表达。(f)原代肝细胞敲降pnpla7后,apoe抗体沉淀apoe蛋白,检测apoe蛋白的泛素化水平。(g)293t细胞中过表达pnpla7与apoe蛋白,利用myc抗体把apoe沉淀,检测apoe泛素化水平。
具体实施方式
seqidno.1为nm-146251.4(mousepnpla7)序列中的一部分靶标序列,敲降时,shrna结合到靶序列上将基因mrna全长降解。
seqidno.2为nm_152286.3(humanpnpla7)序列中的一部分靶标序列,敲降时,shrna结合到靶序列上将基因mrna全长降解。
慢病毒构建
将靶序列设计为shrna序列连接进入plko.1载体,在293t细胞中同时转染plko.1及其包装包膜蛋白载体(pspax2,pmd2.g),使其在293t细胞中包装出成熟慢病毒。收集72h与96h细胞培养液上清,利用0.45μm滤膜过滤,过滤后的上清加入感染试剂polybrene用来感染细胞。
腺病毒构建
将靶序列设计为shrna连接进入pshuttle-silence载体中,与ad-easy重组后在293a细胞中包装腺病毒。腺病毒纯化利用cscl密度梯度离心的方法。
实施例1
如图1-a所示,收集5例正常人体肝脏样本与12例脂肪肝病病人样本,提取rna,检测pnpla7基因表达水平。进一步通过分析17例样本的血清tag与tc水平与pnpla7表达关系,发现pnpla7与人体血液中tag与tc水平均成正相关。
实施例2
肝脏敲降pnpla7后,小鼠肝脏tag含量显著升高,血清tag显著减少
通过小鼠尾静脉注射敲降pnpla7腺病毒(敲降序列如seqidno.1所示),进行肝脏pnpla7敲降(图2-a和b)。病毒注射7天后收取小鼠肝脏及血液,利用folch法提取肝脏脂质,并利用tag检测试剂盒(wako)检测小鼠肝脏tag含量与小鼠血清tag含量,发现敲降pnpla7后小鼠肝脏tag含量显著增多,血清tag含量显著减少。
实施例3
肝脏敲降pnpla7后,vldl分泌速率降低
通过小鼠高压尾静脉注射敲降pnpla7腺病毒,进行肝脏pnpla7敲降。
小鼠尾静脉注射腺病毒5天后,禁食8h,尾静脉注射500mg/kg体重的tritonwr-1339,用于阻断小鼠血液中对vldl的清除和降解。取0、1、2、3h时间点的血液,室温静置1h,4000rpm,4℃离心30min,转移上清,再次离心13000rpm,4℃,10min,转移上清至新的离心管,检测血清中的tag含量,以代表血清中vldl的量。如图3-a所示,肝脏敲降pnpla7后,小鼠肝脏vldl分泌显著减少。
实施例4
血清快速蛋白液相色谱(fplc)
取等量3只小鼠血清混成300μl混合液,利用superose6column通过fplc方法将其按照密度大小分离成不同组分。用tag及tc试剂和检测每个组分中的tag及tc含量,fplc结果中第5-8个组分为vldl的分布位置;第8-12组分为ldl;第13-21组分为hdl。
如图3-b所示,小鼠肝脏敲降pnpla7后,小鼠肝脏分泌的vldl含量显著减少。
实施例5
血清密度梯度离心
新鲜血清取100μl加入2.9mlpbs进行稀释,稀释好的血清,取90μl加入30μl4×sds上样缓冲液作为input,取2.75ml加入1.25ml碘克沙醇配成15%浓度的碘克沙醇溶液,混合均匀后置入5mlbeckman塑封离心管中,用nvt100转子,350000g,4℃离心3h,离心结束后取最上层200μl即为vldl组分,可用于wb,电镜等的检测观察。
如图3-c和d所示,小鼠肝脏敲降pnpla7后,其血清中vldl含量显著减少。
实施例6
肝脏vldl合成分泌途径检测
肝脏内质网与高尔基体分离方法是根据已有的实验方法进行改动。8周大小鼠尾静脉注射腺病毒7天后,禁食16h取肝脏组织,在冰上将肝脏组织置于5ml匀浆液(10mmhepes,ph7.4,150mmsucrose,0.5mmdtt,1xedta-freeproteaseinhibitor)中并快速匀浆。将匀浆液1900g,4℃离心10min后,将上清置于beckman的nvt100转子离心管中,100000g,4℃,90min离心,此时得到的沉淀为高尔基体,最上层组分为脂滴,中间清液部分为细胞质,收集脂滴及细胞质组分。最下层沉淀用2.3ml8.58%的蔗糖溶液重悬,轻轻的加在提前铺好的蔗糖密度梯度溶液的顶端,其中提前铺好的蔗糖密度梯度层从下到上其浓度及体积为:56%(0.46ml),50%(0.92ml),45%(1.38ml),40%(2.3ml),35%(2.3ml),30%(1.38ml),20%(0.46ml)。然后用beckmansw41转子,39000rpm,4℃离心18h。离心结束后,轻轻取出离心管,不要晃动,将样品从顶端按照每1ml作为一个组分,依次取出,共12个组分,每个组分留样跑蛋白电泳,如图4所示分别用golgi的标记蛋白(gm130)和er的标记蛋白(gp78、cnx)检测golgi和er在各组分间的分布。再用bio-radimagej软件检测各蛋白条带的相对灰度值,以最高值作为100%,做分布图。将高尔基体组分合并,内质网组分合并,离心后收集高尔基体与内质网,并检测其tag含量。
实施例7
apoe参与pnpla7调控vldl通路
分离apoe-/-小鼠原代肝细胞,贴壁2h后,pbs洗3遍细胞,将培养液换为无酚红低糖培养基培养24h。检测培养基中tag含量代表细胞分泌的vldl-tag量;检测细胞中tag含量,代表胞内tag含量。分离wt鼠原代肝细胞,贴壁2h后感染ad-shcon与ad-shpnpla7腺病毒6h,换液培养6h后细胞转染apoe-flag质粒,继续培养12h后,细胞换液为无酚红dmem,培养24h,检测培养基中tag含量代表细胞分泌的vldl-tag量;检测细胞中tag含量,代表胞内积累的tag含量。如图5所示,pnpla7通过apoe调控肝脏vldl代谢。
实施例8
在huh7细胞中利用慢病毒敲降pnpla7(敲降序列如seqidno.2所示)后,如图6-b所示,细胞内apoe蛋白表达量显著下调,但是胞内的apoemrna表达量无显著差异。
进一步证实pnpla7通过apoe调控肝细胞vldl分泌的过程在人源细胞与鼠源细胞中高度一致。
序列表
<110>南京医科大学
<120>马铃薯样磷脂酶7的应用
<160>2
<170>siposequencelisting1.0
<210>1
<211>19
<212>dna
<213>人工序列(artificialsequence)
<400>1
ccaagaggattctgcgctt19
<210>2
<211>21
<212>dna
<213>人工序列(artificialsequence)
<400>2
gaaagtcaaggtgttgaacat21
- 还没有人留言评论。精彩留言会获得点赞!