用于产生具有提高的丁香苷生成的转基因植物的方法及使用所述方法产生的植物与流程
本发明涉及基于对芥子醇的显著酶特异性的具有优异的丁香苷合成活性的重组糖基转移酶UGT72E3/2基因,基于用参与类苯基丙烷生物合成途径的F5H、HCT和CHS基因和作为用于正调节参与木质素生物合成途径的基因的转录因子的Myb58基因的代谢工程,用于产生具有提高的丁香苷生成的转基因植物的方法,以及涉及通过该方法获得的植物。
背景技术:
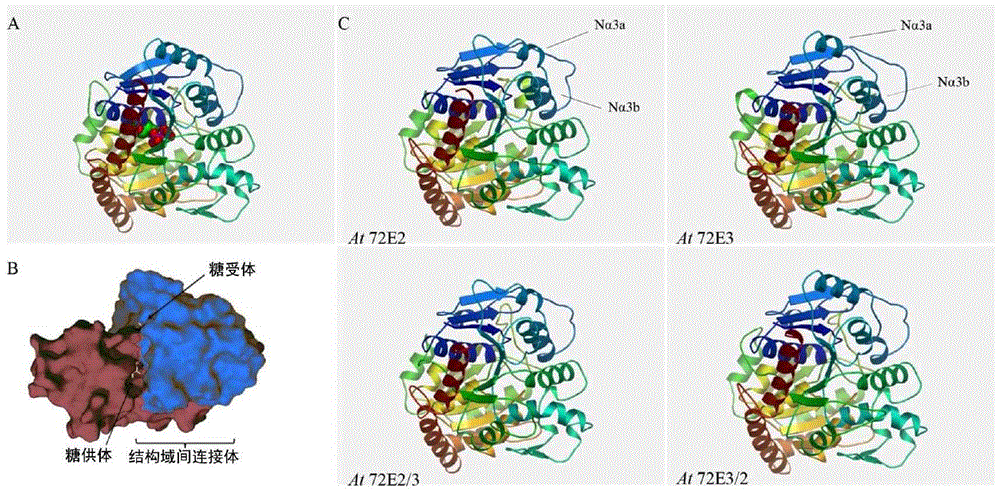
从通过类苯基丙烷合成途径产生的作为构成木质素的组分的芥子醇(s型木质素单体(monolignol))中,通过糖基转移酶(UDP-葡萄糖转移酶)产生作为s型木质素单体糖苷的丁香苷。在模式植物拟南芥(Arabidopsisthaliana)中有约100种糖基转移酶,已报道,其通过形成包括植物激素和次级代谢产物的多种化合物的糖苷,调节细胞活性和这些化合物在液泡中的储存。它们之中,报道过UGT72E2和UGT72E3作为糖基转移酶负责首先将木质素单体如松柏醇和芥子醇特异性地转化为木质素单体糖苷。特别是,作为生成丁香苷所需的芥子醇特异性糖基转移酶,UGT72E3对芥子醇具有很好的底物特异性,但其糖基转移酶活性很低,使其应用受到很大限制。因此,在丁香苷(其是药学上功能性次级代谢产物)大量生产和应用前,发展有利于在植物中有效生成丁香苷的新的糖基转移酶仍然是首要解决的问题。此外,为了通过类苯基丙烷合成途径在植物中有效生成丁香苷,还需要具有使作为以微量存在于植物细胞中的丁香苷前体的芥子醇的含量增加的代谢工程技术,以及对芥子醇具有底物特异性的强糖基转移酶活性。特别是,将刺五加苷B(丁香苷)归类为来自植物的代表性适应原,其 是来自刺五加(E.senticosus)的药用成分,针对应激具有良好的心理和身体的适应效果。适应原是表示增强活体响应各种应激的非特异性抗性而不引起副作用的植物次级代谢产物的术语。近期,据报道,分离出的纯态的丁香苷对成为现代城市中生活的人的严重问题的糖尿病和抑郁症表现出良好效果,并因此其应用较以往更加广泛。然而,由于刺五加的种植范围受到限制,且因此药用成分根据种植区域有巨大的变化,因此难以具有用于商业用途的丁香苷的生产所需的刺五加的稳定供应。从而需要基于使用生物工程技术调节植物代谢途径来发展用于稳定生产植物次级代谢产物如高价值产品丁香苷的技术。同时,韩国专利申请公开号2004-0004764公开了“包含具有肝脏保护活性的刺五加提取物或其丁醇可溶性部分和具有抗氧化和肝脏保护活性的丁香苷的丁醇馏分以及丁香脂素-双-O-β-D-吡喃葡糖苷衍生物”。此外,韩国专利申请公开号1998-0072707公开了“具有肝功能保护活性的丁香苷的药物组合物”。然而,之前从未记载本发明所公开的用于产生具有提高的丁香苷生成的转基因植物的方法以及由所述方法获得的植物。本发明的详细说明待解决的技术问题本发明是鉴于前述需求而设计,并用于产生具有提高的糖基转移活性同时保持高水平的底物特异性的新的重组糖基转移酶。通过UGT72E2和UGT72E3基因的结构域交换(domainswapping)产生重组基因UGT72E2/3和UGT72E3/2。用花浸渍法产生了超表达每个UGT72E家族的转基因拟南芥。此外,作为丁香苷合成效率的定量分析结果,证实了新产生的重组糖基转移酶UGT72E3/2表现出比野生型UGT72E2和UGT72E3显著提高的丁香苷合成。此外,为了通过调节植物代谢途径提高丁香苷的产生,产生了超表达F5H和HCT基因(其调节用于在类苯基丙烷合成途径中调节底物流的重要步骤)的每个拟南芥转基因植物、缺少CHS基因功能的转基因植物, 以及超表达作为木质素合成途径的正调节转录因子的Myb58的转基因植物。作为在将前述的转基因植物与超表达重组糖基转移酶UGT72E3/2的转基因植物杂交后,丁香苷合成效率的定量分析结果,已发现,来自同时超表达UGT72E3/2、F5H和Myb58蛋白的新产生的转基因植物的丁香苷生成量相比仅超表达UGT72E3/2蛋白的情况提高了10倍或以上,因此完成了本发明。解决问题的技术手段为了解决上述问题,本发明提供了重组糖基转移酶UGT72E3/2蛋白,其由氨基酸序列SEQIDNO:2构成。本发明还提供了编码UGT72E3/2蛋白的基因。本发明还提供了包含编码UGT72E3/2蛋白的基因的重组载体。本发明还提供了用重组载体转化的宿主细胞。本发明还提供了相比野生型提高植物中丁香苷合成的方法,包括用重组载体转化植物细胞,以超表达UGT72E3/2基因。本发明还提供了相比野生型具有提高的丁香苷生成的转基因植物,其中用包含编码重组糖基转移酶UGT72E3/2蛋白的基因的重组载体转化植物。本发明还提供了相比野生型具有提高的丁香苷生成的转基因植物,其中用包含编码重组糖基转移酶UGT72E3/2蛋白的基因的重组载体和包含编码F5H(阿魏酸5-羟化酶)蛋白的基因的重组载体转化植物。本发明还提供了相比野生型具有提高的丁香苷生成的转基因植物,其中用包含编码重组糖基转移酶UGT72E3/2蛋白的基因的重组载体、包含编码F5H(阿魏酸5-羟化酶)蛋白的基因的重组载体,以及包含编码Myb58或Myb63蛋白的基因的重组载体转化植物。本发明还提供了相比野生型在植物中具有提高的丁香苷生成的转基因植物,其中用包含编码重组糖基转移酶UGT72E3/2蛋白的基因的重组载体和敲除编码CHS(查尔酮合酶)蛋白的基因的重组载体转化植物。本发明还提供了用于产生相比野生型具有提高的丁香苷生成的转基 因植物的方法,其中用包含编码UGT72E3/2蛋白的基因的重组载体转化植物。本发明还提供了用于产生相比野生型具有提高的丁香苷生成的转基因植物的方法,包括使超表达UGT72E3/2蛋白的转基因植物和超表达F5H蛋白的转基因植物杂交,并选择同时超表达UGT72E3/2和F5H蛋白的转基因植物。本发明还提供了用于产生相比野生型具有提高的丁香苷生成的转基因植物的方法,包括使同时超表达UGT72E3/2蛋白和F5H蛋白的转基因植物和超表达Myb58或Myb63蛋白的转基因植物杂交,并选择同时超表达UGT72E3/2蛋白、F5H蛋白和Myb58或Myb63蛋白的转基因植物。本发明还提供了用于产生相比野生型具有提高的丁香苷生成的转基因植物的方法,包括使超表达UGT72E3/2蛋白的转基因植物和其中敲除编码CHS蛋白的基因的植物杂交,并选择超表达UGT72E3/2但抑制CHS蛋白表达的转基因植物。本发明还提供了通过上述每种方法产生的相比野生型具有提高的丁香苷生成的转基因植物及其种子。本发明又提供了用于提高植物中丁香苷合成的组合物,其包含作为有效成分的重组载体,所述重组载体包含由编码UGT72E3/2蛋白的核苷酸序列SEQIDNO:1组成的基因。本发明的有益效果根据本发明,提供了在各种植物中有效且大量生成丁香苷的新方法,丁香苷来自于植物且具有广泛的应用,如报道所言,对治疗成为现代城市中生活的人的严重问题的糖尿病和抑郁症表现出良好效果,通过该方法可在各种植物中有效且大规模生产药学上非常有用的丁香苷,基于作为丁香苷前体的芥子醇的充足生产和使用重组糖基转移酶UGT72E3/2利用代谢工程通过调节编码重组糖基转移酶UGT72E3/2蛋白的基因和参与植物中类苯基丙烷合成途径的F5H、CHS和Myb58基因提高糖基转移活性,所述丁香苷具有协同效应。因此,期望的是使关于作为食品或药品 具有高价值的农用生物材料的工业得到发展。附图说明图1示出拟南芥糖基转移酶UGT72E2和UGT72E3的一级和二级结构的对比。图2示出来自拟南芥或葡萄的糖基转移酶UGT72B1(A)和VvGT1(B)、来自拟南芥的糖基转移酶UGT72E2和UGT72E3以及本发明产生的重组糖基转移酶UGT72E2/3和UGT72E3/2(C)的三级结构的对比。图3示出(A)用于产生本发明的转化体的重组载体,(B)转入该转化体的基因的表达水平,以及(C)在转化的叶中针对紫外射线的反应性。图4示出了野生型拟南芥和转化体的叶中松柏苷和丁香苷的生成的定量HPLC分析,所述转化体表达4种糖基转移酶基因中的每一种,即UGT72E2、UGT72E3、UGT72E2/3和UGT72E3/2((C)中各层析谱的峰:1、松柏醇4-O-葡糖苷(松柏苷);2、芥子醇4-O-葡糖苷(丁香苷);3、松柏醇;以及4、芥子醇)。图5示出了野生型拟南芥和转化体的根中松柏苷和丁香苷的生成的定量HPLC分析,所述转化体表达4种糖基转移酶基因中的每一种,即UGT72E2、UGT72E3、UGT72E2/3和UGT72E3/2((C)中各层析谱的峰:1、松柏醇4-O-葡糖苷(1、松柏醇4-O-葡糖苷(松柏苷);2、芥子醇4-O-葡糖苷(丁香苷);3、松柏醇;以及4、芥子醇)。图6示出重组糖基转移酶UGT72E3/2的核苷酸序列和氨基酸序列。图7示出了从超表达糖基转移酶UGT72E2、UGT72E3、UGT72E2/3和UGT72E3/2中每个的拟南芥转化体或野生型的叶中制备的蛋白提取物中存在的糖基转移酶的活性的对比结果,其中通过测定向转基因的叶的蛋白提取物中加入松柏醇或芥子醇60分钟后产生的松柏苷和丁香苷,来间接测量各转化体的蛋白提取物中存在的糖基转移酶的活性。(A)松柏苷的生成量和(B)丁香苷的生成量。图8示出了用于合成丁香苷的类苯基丙烷合成途径以及用于本发明 的基因的调节位点。图9示出了基于RT-PCR测定超表达HCT、F5H和Myb58三者的拟南芥中各基因的表达量的结果。使用肌动蛋白2基因作为对照组。图10示出包括野生型在内的转化体的叶中生成的(A)松柏苷和(B)丁香苷的定量HPLC分析,用于确定糖基转移酶UGT72E3/2与类苯基丙烷合成途径的HCT、F5H和CHS基因之间的协同效应,所述转化体表达基因的各种组合。(C)中各层析谱的峰1:代表松柏醇4-O-葡糖苷(松柏苷);峰2代表芥子醇4-O-葡糖苷(丁香苷)。图11示出包括野生型在内的转化体的根中生成的(A)松柏苷和(B)丁香苷的定量HPLC分析,用于确定糖基转移酶UGT72E3/2与类苯基丙烷合成途径的HCT、F5H和CHS基因之间的协同效应,所述转化体表达基因的各种组合。(C)中各层析谱的峰1:代表松柏醇4-O-葡糖苷(松柏苷);峰2代表芥子醇4-O-葡糖苷(丁香苷)。图12为定量HPLC分析的图,其示出了转化体的叶中生成的(A)松柏苷和(B)丁香苷的剧增,其是基于下述的协同效应所致:糖基转移酶UGT72E3/2、类苯基丙烷合成途径中的F5H基因以及作为用于正调节参与木质素合成途径的基因的转录因子的Myb58基因的聚合(pyramiding)。(C)中各层析谱的峰1:代表松柏醇4-O-葡糖苷(松柏苷);峰2代表芥子醇4-O-葡糖苷(丁香苷)。图13示出了HPLC分析结果的图,其表明,基于糖基转移酶UGT72E3/2、类苯基丙烷合成途径中的F5H基因以及作为用于正调节参与木质素合成途径的基因的转录因子的Myb58基因的累积的协同效应,对转化体的根没有明显影响。(C)中各层析谱的峰1:代表松柏醇4-O-葡糖苷(松柏苷);峰2代表芥子醇4-O-葡糖苷(丁香苷)。图14示出了通过RT-PCT所测定的,在超表达UGT72E3/2、F5H和Myb58基因的转基因拟南芥、超表达UGT72E3/2和F5H基因的转基因拟南芥以及野生型拟南芥中参与类苯基丙烷合成途径的各种基因的表达量的图。使用肌动蛋白2基因作为对照组。实施本发明的最佳方式为了实现上述目的,本发明提供了重组糖基转移酶UGT72E3/2蛋白,其由氨基酸序列SEQIDNO:2组成。在本发明的重组糖基转移酶蛋白的范围内,包括了具有SEQIDNO:2所示的氨基酸序列的蛋白以及该蛋白的功能等同物。术语“功能等同物”是指由于氨基酸的添加、替换或缺失,与SEQIDNO:2所示的氨基酸序列具有至少70%、优选至少80%、更优选至少90%且甚至更优选至少95%序列同源性的蛋白,并且其表现出与SEQIDNO:2所示的蛋白实质上相同的生理活性。术语“实质上相同的生理活性”意指丁香苷合成的提高。本发明还包括重组糖基转移酶UGT72E3/2蛋白的“片段”、“衍生物”和“类似物”。本文使用的术语“片段”、“衍生物”和“类似物”指与本发明的重组糖基转移酶UGT72E3/2多肽具有实质上相同的生物学功能或活性的多肽。本发明的衍生物和类似物可以是以下的任意一种:(1)其中一个或多个保守型或非保守型氨基酸残基(优选保守型氨基酸残基)被替换(被替换的氨基酸残基可以被遗传密码编码或未编码)的多肽,(2)在一个或多个氨基酸上具有取代基的多肽,(3)衍生自与其他化合物(即能够提高多肽的半衰期的化合物;例如聚乙二醇)结合的成熟多肽的多肽,以及(4)衍生自与另外的氨基酸残基(例如前导序列、分泌序列、用于纯化多肽的序列、蛋白原序列或融合蛋白)结合的前述多肽的多肽。本文定义的片段、衍生物和类似物对于本领域技术人员而言是熟知的。编码SEQIDNO:2所示的成熟多肽的多核苷酸,包括仅编码成熟多肽的编码序列;编码成熟多肽的序列和各种附加的编码序列;以及编码成熟多肽的序列(以及任意附加的编码序列)和非编码序列。术语“编码多肽的多核苷酸”意指编码多肽的多核苷酸,或者另外包含编码和/或非编码序列的多核苷酸。本发明还涉及编码与本发明所述的序列相同的氨基酸序列的多核苷酸变体或含有其片段、类似物或衍生物的多肽。多核苷酸变体可以是天然存在的等位基因变体或非天然存在的变体。核苷酸变体包括取代变体、 缺失变体和插入变体。如在相关领域中所熟知的,等位基因变体是多核苷酸的替代物,并且其可以含有多核苷酸的一个或多个取代、缺失或插入。然而,变体编码的多核苷酸中没有产生实质的功能性变化。本发明还提供了编码重组糖基转移酶UGT72E3/2蛋白的基因。从源自拟南芥的UGT72E3和UGT72E2基因通过使用结构域交换法制备编码本发明的重组糖基转移酶UGT72E3/2蛋白的基因。在本发明中,为了具有提高的丁香苷生成率,由UGT72E2和UGT72E3基因通过使用结构域交换法制备重组基因UGT72E2/3和UGT72E3/2,以产生如UGT72E2一样具有高糖基转移酶活性同时如UGT72E3一样保持对芥子醇的底物特异性的新的重组糖基转移酶。首先确定来自拟南芥的约100种糖基转移酶的糖基转移酶UGT72E进化支的酶特性,已报道该进化支具有将糖转移到芥子醇(即丁香苷的前体)或结构相似的松柏醇的能力。UGT72E进化支还具有类似于普通糖基转移酶的结构特性,并且已报道氨基末端结构域具有底物识别位点,羧基末端结构域具有用于将被UDP激活的糖转移至底物的酶促激活区域,以及羧基末端上的PSPG(植物次级产物糖基转移酶)对于来自植物的糖基转移酶的活性特别重要。UGT72E进化支包括具有相似核苷酸的糖基转移酶UGT72E1、UGT72E2和UGT72E3。在本发明中,UGT72E2和UGT72E3的每个均分为包括编号1至编号344的氨基酸的氨基片段和包括从编号345至编号481的氨基酸的羧基片段。氨基片段包括用于决定底物识别特异性的区域,羧基末端包括PSPG基序,其对于糖基转移酶的活性很重要。为了在植物中有效生成丁香苷,底物特异性比糖基转移酶的活性更重要。因此,将大的氨基片段制备成包括全长的3/4,同时将羧基片段制备成包括PSPG基序的小片段,而不是具有精确的对切。结果,通过将UGT72E3的包括氨基末端上的编号1至编号344的氨基酸的氨基片段连接于UGT72E2的包括编号345至羧基末端上的编号481的氨基酸的羧基片段,产生了本发明的重组UGT72E3/2基因。本发明的基因可以是编码重组糖基转移酶UGT72E3/2蛋白的DNA或RNA。cDNA、基因组DNA和人工合成的DNA包括在DNA中。DNA可以是单链或双链的。DNA可以是编码序列或非编码序列。优选的是,本发明的基因可含有核苷酸序列SEQIDNO:1。此外,该核苷酸序列的同源物也在本发明的范围内。具体而言,上述基因可包含与核苷酸序列SEQIDNO:1具有优选至少70%、更优选至少80%,还更优选至少90%,且最优选至少95%同源性的核苷酸序列。某个多核苷酸的“序列同源性%”是通过比较最佳排列的两条序列的可比较区而确定的。在这点上,可比较区的一部分多核苷酸与参照序列(无任何添加或缺失)相比,相对于所述两条序列的最佳比对,可含添加或缺失(即缺口)。本发明还涉及与和上述核苷酸序列SEQIDNO:1具有至少50%、优选至少70%、更优选至少80%序列同源性的序列杂交的多核苷酸。本发明特别涉及在严格条件下与本发明的多核苷酸杂交的多核苷酸。如本文所述,“严格条件”意指:(1)在低离子强度和高温下的杂交和洗涤,如0.2xSSC,0.1%SDS,60℃;(2)存在变性剂如50%(v/v)甲酰胺、0.1%牛血清/0.1%Ficoll以及42℃下的杂交;或(3)仅在同源性为至少80%、优选至少90%、且更优选至少95%的两条序列之间发生的杂交。此外,由可杂交的核苷酸编码的多肽的生物学功能和活性与SEQIDNO:2所示的成熟多肽的生物学功能和活性相同。本发明还提供了包含编码前述重组糖基转移酶UGT72E3/2蛋白的基因的重组载体。术语“重组体”是指能够复制异源核苷酸,或能够表达所述核苷酸或肽、异源肽或由异源核苷酸编码的蛋白的细胞。重组细胞可以以有义或反义的形式表达在天然状态的细胞中未发现的基因或基因片段。此外,重组细胞可表达天然状态下存在的基因,只要通过人工手段将所述基因修饰或重转入细胞即可。本发明的重组植物表达载体可以用作允许外来基因在植物中瞬时表达的瞬时表达载体,并且还用作允许外来基因在植物中永久表达的植物 表达载体。可用于本发明的二元载体可以是包含T-DNA的RB(右边界)和LB(左边界)的任意二元载体,在与根瘤农杆菌(A.tumefaciens)的Ti质粒一起存在时,其可以转化植物。优选的是使用相关领域技术人员常用的pBI101(目录编号:6018-1,Clontech,USA)、pBIN19(基因库保藏号U09365)、pBI121、pCAMBIA等。本文使用的术语“载体”是指递送至细胞的DNA片段和核苷酸分子。载体能够复制DNA并能在宿主细胞中独立复制。术语“递送系统”和“载体”通常可交替使用。术语“表达载体”是指包含所需编码序列和在特定宿主生物体中表达可操作连接的编码序列所必需的其他适当核苷酸序列的重组DNA分子。可用于真核细胞中的启动子、增强子、终止信号和多聚腺苷酸化信号是本领域熟知的。植物表达载体的优选实例是Ti-质粒载体,当所述Ti-质粒载体存在于合适的宿主中(如根瘤农杆菌)时该载体可以将自身的一部分(即所谓的T区)转入植物细胞。其他类型的Ti-质粒载体(参见EP0116718B1)目前用于将杂交基因转移到可以产生新植物的原生质体,这是通过适当地将植物细胞或杂交DNA插入至植物基因组而实现的。Ti-质粒载体的特别优选形式是EP0120516B1和US专利4940838中公开的所谓二元载体。可用于将本发明的基因导入宿主植物的其他载体可选自双链植物病毒(例如CaMV)、单链植物病毒以及源自双粒病毒等的病毒载体(例如非完整植物病毒载体)。特别是,在植物宿主不能被适当的转化时,使用所述载体是有利的。表达载体可含有至少一个选择标记。所述选择标记是具有能通过普通化学方法选择靶基因的特性的核苷酸序列。实例包括可用于区分转化细胞和非转化细胞的所有基因。具体实例包括:除草剂(如草甘膦和膦丝菌素(phosphinotricine))抗性基因;抗生素(如卡那霉素、氨苄青霉素、G418、博莱霉素、潮霉素和氯霉素)抗性基因,但不限于此。对于本发明一个实施方案的植物表达载体而言,启动子可以是CaMV 35S、肌动蛋白、泛素、pEMU、MAS和组蛋白启动子中的任意一个,但不限于此。术语“启动子”是指结合RNA聚合酶以启动其转录的DNA分子且其对应于结构基因上游的DNA区域。术语“植物启动子”是指可启动植物细胞内转录的启动子。术语“组成型启动子”是指在大部分环境条件和生长状态或细胞分化状态下都有活性的启动子。因为可采用多种机制在不同阶段选择转化体,所以本发明优选组成型启动子。因此,本发明并不限制选择组成型启动子的可能性。在本发明的上述重组载体中,可以使用任意种类的典型终止子。实例包括胭脂碱合酶(NOS)、水稻α-淀粉酶RAmylA终止子、菜豆碱终止子以及根瘤农杆菌Optopine基因的终止子等,但不限于此。本发明还提供了用前述重组载体转化的宿主细胞。当用本发明的载体转化真核细胞时,可以用作宿主细胞的有酵母(酿酒酵母(Saccharomycecerevisiae))、昆虫细胞、人的细胞(例如CHO(中国地鼠卵巢细胞)的细胞系、W138、BHK、COS-7、293、HepG2、3T3、RIN和MDCK细胞系)、植物细胞等。宿主细胞优选为植物细胞。对于向宿主细胞递送本发明的载体的方法,可以通过微注射法、磷酸钙沉淀法、电穿孔法、脂质体介导的转染法、DEAE-葡聚糖处理法或基因轰击法将载体导入宿主细胞。本发明还提供了相比野生型提高植物中丁香苷合成的方法,包括用超表达UGT72E3/2基因的重组载体转化植物细胞。根据本发明的一个实施方案的方法,UGT72E3/2基因可以由核苷酸序列SEQIDNO:1组成,但不限于此。本发明还提供了相比野生型具有提高的丁香苷生成的转基因植物,其中用包含编码由氨基酸序列SEQIDNO:2组成的重组糖基转移酶UGT72E3/2蛋白的基因的重组载体转化植物。本发明还提供了相比野生型具有提高的丁香苷生成的转基因植物,其中用包含编码由氨基酸序列SEQIDNO:2组成的重组糖基转移酶UGT72E3/2蛋白的基因的重组载体和包含编码由氨基酸序列SEQIDNO: 4组成的F5H(阿魏酸5-羟化酶)蛋白的基因的重组载体转化植物。本发明还提供了相比野生型具有提高的丁香苷生成的转基因植物,其中用包含编码由氨基酸序列SEQIDNO:2组成的重组糖基转移酶UGT72E3/2蛋白的基因的重组载体、包含编码由氨基酸序列SEQIDNO:4组成的F5H(阿魏酸5-羟化酶)蛋白的基因的重组载体,以及包含编码由氨基酸序列SEQIDNO:6组成的Myb58蛋白的基因的重组载体转化植物。本发明还提供了相比野生型具有提高的丁香苷生成的转基因植物,其中用包含编码由氨基酸序列SEQIDNO:2组成的重组糖基转移酶UGT72E3/2蛋白的基因的重组载体、包含编码由氨基酸序列SEQIDNO:4组成的F5H(阿魏酸5-羟化酶)蛋白的基因的重组载体,以及包含编码由氨基酸序列SEQIDNO:8组成的Myb63蛋白的基因的重组载体转化植物。本发明还提供了相比野生型具有提高的丁香苷生成的转基因植物,其中用包含编码由氨基酸序列SEQIDNO:2组成的重组糖基转移酶UGT72E3/2蛋白的基因的重组载体、敲除编码由氨基酸序列SEQIDNO:10组成的CHS(查尔酮合酶)蛋白的基因的重组载体转化植物。在本发明中,为了向超表达重组糖基转移酶UGT72E3/2的转化体高效供应作为丁香苷的底物的芥子醇,利用了丁香苷合成途径中的每个步骤和工作酶。具体而言,为了提高向类苯基丙烷合成途径引入香豆酰辅酶A,超表达HCT(羟基肉桂酰辅酶A:莽草酸/奎尼酸羟基肉桂酰转移酶)基因,并降低转化为松柏醇的松柏醛的量,并且为了促进向芥子醇的转化,超表达F5H(阿魏酸5-羟化酶)基因。此外,为了降低从丁香苷合成途径到类黄酮途径损失的香豆酰辅酶A的量,使用了缺少CHS(查尔酮合酶)基因功能的突变体。为敲除CHS基因,可以使用沉默载体,但不限于此。本文所述的术语“敲除”是指从核苷酸序列中修饰或除去特定基因,以阻止所述特定基因的表达,并且其一般是指下调或完全抑制基因表达 的现象。本发明还提供了产生相比野生型具有提高的丁香苷合成的转基因植物的方法,包括:(a)用包含编码由氨基酸序列SEQIDNO:2组成的重组糖基转移酶UGT72E3/2蛋白的基因的重组载体转化植物,以及(b)由步骤(a)的转基因植物细胞再生植株。本发明还提供了产生相比野生型具有提高的丁香苷生成的转基因植物的方法,包括:(a)通过用包含编码由氨基酸序列SEQIDNO:2组成的重组糖基转移酶UGT72E3/2蛋白的基因的重组载体转化植物,产生超表达UGT72E3/2蛋白的转基因植物;(b)通过用包含编码由氨基酸序列SEQIDNO:4组成的F5H(阿魏酸5-羟化酶)蛋白的基因的重组载体转化植物,产生超表达F5H蛋白的转基因植物;(c)使步骤(a)的超表达UGT72E3/2蛋白的转基因植物和步骤(b)的超表达F5H蛋白的转基因植物杂交,并选择同时超表达UGT72E3/2蛋白和F5H蛋白的转基因植物。本发明还提供了产生相比野生型具有提高的丁香苷生成的方法,包括:(a)通过用包含编码由氨基酸序列SEQIDNO:2组成的重组糖基转移酶UGT72E3/2蛋白的基因的重组载体转化植物,产生超表达UGT72E3/2蛋白的转基因植物;(b)通过用包含编码由氨基酸序列SEQIDNO:4组成的F5H(阿魏酸5-羟化酶)蛋白的基因的重组载体转化植物,产生超表达F5H蛋白的转基因植物;(c)使步骤(a)的超表达UGT72E3/2蛋白的转基因植物和步骤(b)的超表达F5H蛋白的转基因植物杂交,并选择同时超表达UGT72E3/2蛋白和F5H蛋白的转基因植物;(d)通过用包含编码由氨基酸序列SEQIDNO:6组成的Myb58蛋白的基因的重组载体转化植物,产生超表达Myb58蛋白的转基因植物;(e)使步骤(c)的同时超表达UGT72E3/2蛋白和F5H蛋白的转基因植物和步骤(d)的超表达Myb58蛋白的转基因植物杂交,并选择同时超表达UGT72E3/2蛋白、F5H蛋白和Myb58蛋白的转基因植物。本发明还提供了产生相比野生型具有提高的丁香苷生成的转基因植物的方法,包括:(a)通过用包含编码由氨基酸序列SEQIDNO:2组成的重组糖基转移酶UGT72E3/2蛋白的基因的重组载体转化植物,产生超表达UGT72E3/2蛋白的转基因植物;(b)通过用包含编码由氨基酸序列SEQIDNO:4组成的F5H(阿魏酸5-羟化酶)蛋白的基因的重组载体转化植物,产生超表达F5H蛋白的转基因植物;(c)使步骤(a)的超表达UGT72E3/2蛋白的转基因植物和步骤(b)的超表达F5H蛋白的转基因植物杂交,并选择同时超表达UGT72E3/2蛋白和F5H蛋白的转基因植物;(d)通过用包含编码由氨基酸序列SEQIDNO:8组成的Myb63蛋白的基因的重组载体转化植物,产生超表达Myb63蛋白的转基因植物;(e)使步骤(c)的同时超表达UGT72E3/2蛋白和F5H蛋白的转基因植物和步骤(d)的超表达Myb63蛋白的转基因植物杂交,并选择同时超表达UGT72E3/2蛋白、F5H蛋白和Myb63蛋白的转基因植物。本发明还提供了产生相比野生型具有提高的丁香苷生成的转基因植物的方法,包括:(a)通过用包含编码由氨基酸序列SEQIDNO:2组成的重组糖基转移酶UGT72E3/2蛋白的基因的重组载体转化植物,产生超表达UGT72E3/2蛋白的转基因植物;(b)产生其中敲除编码由氨基酸序列SEQIDNO:10组成的CHS(查尔酮合酶)蛋白的基因的植物;以及(c)使步骤(a)的超表达UGT72E3/2蛋白的转基因植物和步骤(b)的敲除CHS蛋白编码基因的植物杂交,并选择超表达UGT72E3/2蛋白和抑制CHS蛋白表达的转基因植物。根据本发明的一个实施方案的方法,植物优选在叶或根处或最优选在叶中呈现提高的丁香苷合成,但并不限于此。本发明的方法包括用本发明的重组载体转化植物细胞的步骤,并且可以由根瘤农杆菌(Agrobacteriumtumefiaciens)介导所述转化。此外,本发明的方法包括由转基因植物细胞再生转基因植株的步骤。对于由转基因植物细胞再生转基因植株的方法,可以使用相关领域熟知的方法。需要将转基因植物细胞再生为整个植株。对于各种物种的通过愈合组织或原生质体的培养再生为成熟植株的技术是相关领域中熟知的(HandbookofPlantCellCulture(植物细胞培养手册),Vol.1-5,1983-1989Momillan,N.Y.)。本发明中所用的Myb58基因是用于正调节参与类苯基丙烷生物合成途径的基因的转录因子,预期已知具有相似功能的Myb63也预期在丁香苷合成的过程中具有协同效应。本发明还提供了通过上述每种方法产生的相比野生型具有提高的丁香苷生成的转基因植物及其种子。对于本发明的一个实施方案的植物而言,所述植物可优选为双子叶植物,如拟南芥(Arabidopsisthaliana)、烟草、茄子、辣椒、番茄、牛蒡、茼蒿、生菜、气球花、菠菜、甜菜、山药、芹菜、胡萝卜、水芹菜、水香菜、中国白菜、甘蓝、萝卜(Raphanussativusfor.raphnistroidesMAK)、西瓜、东方甜瓜(orientalmelon)、黄瓜、西葫芦、丝瓜、草莓、大豆、绿豆、芸豆或甜豌豆。最优选地,其可以是拟南芥,但并不限于此。本发明还提供了用于提高植物中丁香苷合成的组合物,其包含作为有效成分的重组载体,所述重组载体包含由SEQIDNO:1组成的编码UGT72E3/2蛋白的基因。所述组合物包含作为有效成分的重组载体,所述重组载体包含由SEQIDNO:1组成的编码UGT72E3/2蛋白的基因,并 且根据用所述重组载体转化植物,可以提高植物中丁香苷的合成。下面,通过实施例更详细地说明本发明。然而,很显然以下给出的实施例仅用于举例说明本发明,本发明并决不限于以下实施例。本发明所使用的基因和核苷酸序列的信息本发明所使用的基因和核苷酸序列的信息如下表1所述。[表1]基因名称核苷酸序列编号氨基酸序列编号UGT72E3/212F5H34Myb5856Myb6378CHS910HCT1112实施例1.拟南芥糖基转移酶UGT72E2和UGT72E3的一级结构和二级结构为了研发用于于植物中产生具有各种药物应用的丁香苷的新的糖基转移酶,通过使用SWISS-MODEL工作空间(图1),对具有非常优异的糖基转移酶效率但对丁香苷的前体松柏醇的底物特异性不佳的UGT72E2与对芥子醇具有非常优异的底物特异性但糖基转移酶效率低的UGT72E3蛋白之间进行一级结构与二级结构的对比。作为产生具有两种酶的所有优势的新的重组糖基转移酶的结构域交换的预备步骤,标记对糖基转移酶活性很重要的含有PSPG基序的羧基末端与用于决定底物特异性的氨基末端的一级结构和二级结构的差异以及发生结构域交换的区域。实施例2.拟南芥糖基转移酶UGT72E2和UGT72E3和新产生的重组糖基转移酶UGT72E2/3和UGT72E3/2的三级结构从分别来自拟南芥和葡萄的糖基转移酶UGT72B1和VvGT1(其三级结构已研究透彻)可见,糖基转移酶一般具有用于决定底物特异性的区域(即糖受体)和用于决定糖基转移酶活性的区域(即糖供体),两者在氨基末端结构域和羧基末端结构域之间的边界的深且窄的缺口(gap)处彼此相邻(图2A和2B)。通过使用SWISS-MODEL工作空间,获得UGT72E2、 UGT72E3、UGT72E2/3和UGT72E3/2的预期三级结构(图2C)。认为决定底物特异性的区域和决定糖基转移酶活性的区域附近的结构变化,与糖基转移酶UGT72E2、UGT72E3、UGT72E2/3和UGT72E3/2之间的底物特异性和糖基转移酶活性的差异有关。实施例3.用于转化体生产的重组载体、转移到转化体的基因的表达水平以及转化体响应UV射线的表型本发明的发明人探讨了,在拟南芥的100种糖基转移酶中,糖基转移酶UGT72E进化支的酶特征,已报道其具有将糖转为作为丁香苷的前体的芥子醇或结构相似的松柏醇的能力。UGT72E进化支也具有与常见糖基转移酶相似的结构特征。氨基末端结构域具有用于底物识别的区域,羧基末端具有用于将被UDP激活的糖转化为底物的酶激活区域。特别是,据报道,羧基末端的PSPG(植物次级产物糖基转移酶)基序对于来自植物的糖基转移酶的活性很重要。UGT72E进化支包括具有相似核苷酸序列的糖基转移酶UGT72E1、UGT72E2和UGT72E3。在本发明中,UGT72E2和UGT72E3的每个均分为包括编号1至编号344的氨基酸的氨基片段和包括从编号345至编号481的氨基酸的羧基片段。氨基片段包括用于决定底物识别特异性的区域,羧基末端包括对于糖基转移酶的活性很重要的PSPG基序。为了在植物中有效生成丁香苷,底物特异性比糖基转移酶的活性更重要。因此,,将大的氨基片段制备成包括全长的3/4,同时将羧基片段制备成包括PSPG基序的小片段,而不是具有精确的对切。通过将UGT72E2的包括氨基末端上的编号1至编号344的氨基酸的氨基片段与UGT72E3的包括编号345至羧基末端上的编号481的氨基酸的羧基片段连接,制备本发明所使用的重组UGT72E2/3基因。通过将UGT72E3的包括氨基末端上的编号1至编号344的氨基酸的氨基片段与UGT72E2的包括编号345至羧基末端上的编号481的氨基酸的羧基片段连接,制备重组UGT72E3/2基因。在本发明中,构建二元载体,使得从拟南芥分离出的UGT72E2和 UGT72E3与本发明新产生的重组的UGT72E2/3和UGT72E3/2基因的编码区受CaMV35S启动子和超级启动子控制。将所得载体并入根瘤农杆菌EHA105中,然后使用植物内方法(inplantamethod)用细菌转化拟南芥(图3)。由于转化体的表现型受引入的转基因表达水平的显著影响,因此基于RT-PCR,测定了在具有潮霉素抗性的拟南芥转化体之中,由来自拟南芥的转移酶基因UGT72E2、UGT72E3、UGT72E2/3和UGT72E3/2的稳定引入所产生的表达量。为了特异性扩增所引入的基因,使用UGT72E基因特异性正向引物(5’GGTTGGAGCTCGACGTTGGAAAGCGTC3’;SEQIDNO:13)和对载体的3’UTR区有特异性的反向引物(5’TTAAAGCAGGGCATGCCTGC3’;SEQIDNO:14)的组合。为修正相对RNA量,使用总在恒定水平表达的来自拟南芥的肌动蛋白1基因,作为参照基因。在已测定的每个基因的15种转化体中,最终鉴定出具有UGT72E2、UGT72E3、UGT72E2/3和UGT72E3/2的相似表达的转化体系(图3B)。根据叶中糖基转移酶的活化的芥子醇向丁香苷的转化,降低了叶中吸收UV射线的芥子酯(sinapylester)的生成。如此,当具有提高的糖基转移酶活性的转化体的叶暴露于UV射线时,大部分UV射线被叶绿素吸收,从而呈现很强的红色。当芥子酯的生成正常发生在叶中时,被叶绿素吸收的UV射线的量降低,呈现很弱的红色(图3C)。实施例4.转化体的叶中松柏苷和丁香苷生成的定量HPLC分析被子植物的叶中由类苯基丙烷合成途径的活化产生的木质素单体主要作为松柏醇存在,其通过糖基转移酶转化为松柏苷。由于酶的作用,极小部分的松柏醇被转化为作为丁香苷前体的芥子醇,并且由于酶活性低,被糖基转移酶引起的向丁香苷的转化率非常低。大部分芥子醇转化为芥子酯,其具有通过吸收UV射线保护叶的活性。因此,对松柏醇具有高底物特异性的UGT72E2基因的表达提高了松柏苷的生成,对芥子醇具有高底物特异性的UGT72E3的超表达相对提高了丁香苷的生成(图4A 和4B)。重组基因UGT72E3/2的超表达相比UGT72E3基因的超表达表现出植物中丁香苷的生成增加48.7%的效果(图4B和4C)。实施例5.转化体的根中松柏苷和丁香苷生成的定量HPLC分析与不断暴露于光的叶不同,植物的根一旦暴露于光便显示出参与基于信号转导机制的类苯基丙烷合成途径的各种基因的增强表达。结果,提高了包括大量松柏醇和芥子醇的木质素单体的合成。尽管松柏苷和丁香苷的生成(其在相同条件下栽培的野生型植株的叶中很难检测到)明显增加,但丁香苷的生成率仍很低,即仅是松柏苷的25%。此外,尽管在超表达具有高糖基转移酶活性且对松柏醇具有强底物特异性的UGT72E2的转化体的根中,松柏苷的生成增加了两倍以上,但丁香苷生成却没有大的变化(图5B和5C)。在超表达具有弱糖基转移酶活性且对芥子醇具有强底物特异性的UGT72E3基因的转化体的根中,松柏苷和丁香苷的生成与野生型几乎在相同水平。然而,重组基因UGT72E3/2的超表达相比UGT72E3基因的超表达呈现出根中生成的丁香苷增加11.7%的效果(图5B和5C)。实施例6.糖基转移酶UGT72E3/2的核苷酸序列和氨基酸序列图6示出了新的重组糖基转移酶UGT72E3/2基因的核苷酸序列和由该核苷酸序列编码的蛋白的氨基酸序列,其相比常规已知的糖基转移酶UGT72E3显示出糖基转移酶活性提高了48.7%以上,同时保持对芥子醇的底物特异性。实施例7.从野生型和超表达糖基转移酶UGT72E2、UGT72E3、UGT72E2/3和UGT72E3/2中的每个的转基因拟南芥的叶中制备的蛋白提取物中存在的糖基转移酶的活性的对比被子植物的叶中由类苯基丙烷合成途径的激活而生成的大部分木质素单体作为松柏醇存在,并且其转化为松柏苷,同时仅有极小部分通过F5H(阿魏酸5-羟化酶)、COMT(咖啡酸3-O-甲基转移酶)和CAD的连续酶作用转化为作为丁香苷前体的芥子醇。因此,作为底物的芥子醇的供应是决定丁香苷生成和糖基转移酶活性的重要因素。为了比较糖基转移酶的活性,将1mM松柏醇或芥子醇和5mMUDP-葡萄糖作为底物加入到从野生型和超表达糖基转移酶UGT72E2、UGT72E3、UGT72E2/3和UGT72E3/2中的每个的转基因拟南芥的叶中制备的蛋白提取物中。然后,使反应于22℃下发生60分钟。之后,将甲醇(2x体积)加入反应溶液中,以结束反应,并通过使用HPLC,通过HPLC对反应前和反应后生成的松柏苷和丁香苷进行定量。如前面在体内条件下所证明的,重组糖基转移酶UGT72E3/2对芥子醇具有高底物特异性和高糖基转移酶活性。特别是,根据底物的添加,UGT72E3/2比其他糖基转移酶呈现更好的丁香苷生成速度(图7)。以上结果证明,也能够基于代谢工程手段通过使用向超表达UGT72E3/2的转化体中增加芥子醇供应的方法,在植物中以高效率生成丁香苷。实施例8.类苯基丙烷合成途径和用于丁香苷合成的基因调节位点为了使用代谢工程手段向超表达重组糖基转移酶UGT72E3/2有效供应作为丁香苷底物的芥子醇,利用了类苯基丙烷合成途径的每个步骤和工作酶。具体而言,为了降低从丁香苷合成途径到类黄酮途径损失的香豆酰辅酶A的量,使用了缺少CHS(查尔酮合酶)基因的突变体。此外,为了提高向类苯基丙烷合成途径引入香豆酰辅酶A,超表达HCT(羟基肉桂酰辅酶A:莽草酸/奎尼酸羟基肉桂酰转移酶)基因,并降低松柏醛转化为松柏醇的量,并且为了促进向芥子醇的转化,采用了超表达F5H(阿魏酸5-羟化酶)基因的策略(图8)。实施例9.超表达作为调控类苯基丙烷合成途径的基因的HCT、F5H和Myb58的转基因拟南芥的生产以及各基因的表达量的测定构建二元载体,使得拟南芥的HCT、F5H和Myb58的编码区受超级启动子调控。在向根瘤农杆菌EHA105引入载体后,通过植物内方法使用前述细菌进行拟南芥的转化。那时,为了使通过使用潮霉素抗性选择标记利用杂交育种和选择方式产生的超表达UGT72E3/2的转化体中具有HCT、F5H和Myb58的聚合累积(pyramidalaccumulation),通过使用卡那 霉素(HCT和F5H)或除草剂(Myb58)抗性选择标记制备各基因的拟南芥转化体。由于转化体的表型受引入的转化基因表达水平的显著影响,因此在已选择的转化体中,基于RT-PCR,测定了由稳定引入HCT、F5H和Myb58基因所产生的表达量。为了特定扩增引入的基因,使用HCT基因特异性正向引物HCT-F(5'-CTGGTTACTTTGGGAATGTGATATTCAC-3';SEQIDNO:15)、F5H基因特异性正向引物F5H-F(5'-CAGACGAGTTGAAGAATCCGACATCGAG-3';SEQIDNO:16)和Myb58基因特异性正向引物Myb58-F(5'-CAGACGAGTTGAAGAATCCGACATCGAG-3';SEQIDNO:17)的组合物。此外,使用了载体的对3’UTR区具有特异性的UTR-R(5’TTAAAGCAGGGCATGCCTGC3’;SEQIDNO:14)作为反向引物。为了校准RNA的相对量,使用总在恒定水平表达的拟南芥的肌动蛋白2基因作为参照基因。对每个基因检测了10个转化体,结果,最终获得了具有优良的HCT、F5H和Myb58表达的转化体系(图9)。同时,仅有一种CHS基因存在于拟南芥中,且当缺少该基因时,种皮颜色转变为黄色。基于这种表型差异,分离出纯合转化体。实施例10.转化体的叶和根中松柏苷和丁香苷生成的定量HPLC分析通过类苯基丙烷合成途径,被子植物产生以下三种木质素单体:即利用p-香豆醇产生的H木质素单体、利用松柏醇产生的G木质素单体和利用芥子醇产生的S木质素单体。然而,大部分木质素单体以G木质素单体的形式存在,并因此松柏醇的浓度在植物细胞中相对最高。糖基转移酶UGT72E2的超表达使高浓度的松柏醇转化为松柏苷。由于F5H和COMT(咖啡酸3-O-甲基转移酶)的酶作用,使部分松柏醇转化为芥子醇,因此植物细胞中作为丁香苷前体的芥子醇的浓度非常低。因此,为了在植物细胞中有效生成丁香苷,除了具有高效糖基转移酶UGT72E3/2之外,还需要用于增加芥子醇的代谢工程方法。为了检测糖基转移酶UGT72E3/2与HCT、F5H和CHS基因之间的协同效应,进行转化体的叶和根中生成的松柏苷和丁香苷的定量HPLC分析。具体是,产生了超表达调节类苯基丙烷合成途径的重要步骤的HCT或F5H的转化体,并使其与超表达高效糖基转移酶UGT72E3/2的转化体杂交。之后,在F2代分离超表达HCT和UGT72E3/2或F5H与UGT72E3/2的转化体,并从下一代获得纯合株系。此外,为了防止作为类苯基丙烷合成途径到类黄酮合成途径的重要前体的p-香豆酰辅酶A的损失,使缺少负责p-香豆酰辅酶A向查尔酮转化的CHS(查尔酮合酶)的突变体与超表达UGT72E3/2的转化体进行杂交。在从F2代中分离出敲除CHS基因且超表达UGT72E3/2基因的株系后,从下一代确认纯合株系。作为通过使用HPLC测定转化体的叶和根中丁香苷合成效率的结果,发现相比仅超表达UGT72E3/2的转化体,在超表达HCT,或超表达F5H但缺少CHS基因功能的植物的叶中丁香苷合成分别提高了17.3%、71.3%和64.6%(图10)。然而,从植物的根来讲,相比仅超表达UGT72E3/2基因的转化体,仅具有UGT72E3/2和F5H的超表达的转化体表现出略微提高的丁香苷合成(图11)。实施例11.基于利用UGT72E3/2、F5H和Myb58基因的聚合的协同效应,转化体的叶与根中丁香苷生成的定量HPLC分析对芥子醇具有高特异性的新的糖基转移酶基因UGT72E3/2(其已通过酶工程方法研发)和参与类苯基丙烷途径中松柏醇向芥子醇的转化的F5H基因的累积的超表达,表现出明显提高丁香苷生成率的效果。同时,当暴露于光时,植物的根表现出参与基于光信号转导机制的类苯基丙烷合成途径的各种基因的提高的表达。结果,提高了包括大量松柏醇和芥子醇的木质素单体的合成,因此野生型植物根中丁香苷的合成量比叶高至少36倍。同时,超表达UGT72E3/2和F5H的转化体的叶中丁香苷的生成量相比野生型的叶提高了16倍或以上。然而,相比转化体的根中生成的丁香苷的量,其仍然低了至少4倍。与根不同,提高叶中生成的丁 香苷的优势在于,可在不破坏植物的情况下进行大量生产。因此,如暴露于光的根一样,增加参与类苯基丙烷合成的各种基因的方法对于叶也是需要的。为了解决前述问题,在本发明中通过定量HPLC分析测定了转化体的丁香苷生成量,丁香苷的生成基于使用糖基转移酶UGT72E3/2基因、类苯基丙烷合成途径的F5H基因和作为用于正调节与木质素合成途径有关的基因的转录因子的Myb58基因的聚合的协同效应。为了确认使用Myb58基因的协同效应,产生了超表达拟南芥中作为用于正调节木质素合成途径的特定转录因子的Myb58基因的转化体。由于Myb58基因不能提高F5H基因的表达,在与超表达UGT72E3/2基因和F5H基因的转化体杂交后,在F2代选择超表达Myb58、UGT72E3/2和F5H三者的转化体株系,并在下一代获得纯合株系。作为利用HPLC探讨这些转化体的叶和根中丁香苷合成效率的结果,已发现,在超表达UGT72E3/2、F5H和Myb58基因三者的转化体的叶中,丁香苷的生成量相比超表达UGT72E3/2和F5H基因的转化体提高了8倍,或者相比仅超表达UGT72E3/2基因的转化体提高了10倍,因此表现出优异的效果(图12)。基于与丁香苷生成有关的三种基因即UGT72E3/2、F5H和Myb58之间的协同效应,在叶中有效生成丁香苷的结果与在基因水平进行的分析结果相匹配。通过超表达Myb58并单独地超表达不受Myb58调控的UGT72E3/2和F5H基因来增强与类苯基丙烷合成途径有关的各种基因的表达,使得转化体的叶中丁香苷的生成量明显提高(图14)。此外,结果表明,转化体的叶中的丁香苷生成量相比根中的生成量提高了2倍或以上。因此,已确认,作为UGT72E3/2、F5H和Myb58基因超表达引起的协同效应,在根中丁香苷的生成最理想地降低大约2倍左右,同时在叶中明显提高。因此,用上述方法获得能够大量生成丁香苷的转基因植物。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1