c‑met特异性人抗体及其制备方法与流程
本发明涉及与c-met特异性结合的抗体(后文称为c-met特异性抗体)或其片段,更具体地,涉及分离自人fv(scfv)噬菌体文库的对c-met具有高亲和力和特异性的全人抗体、编码所述抗体的多核苷酸、包含所述多核苷酸的重组载体、使用所述重组载体转化的细胞和通过培养所述细胞产生c-met特异性抗体的方法。
背景技术:
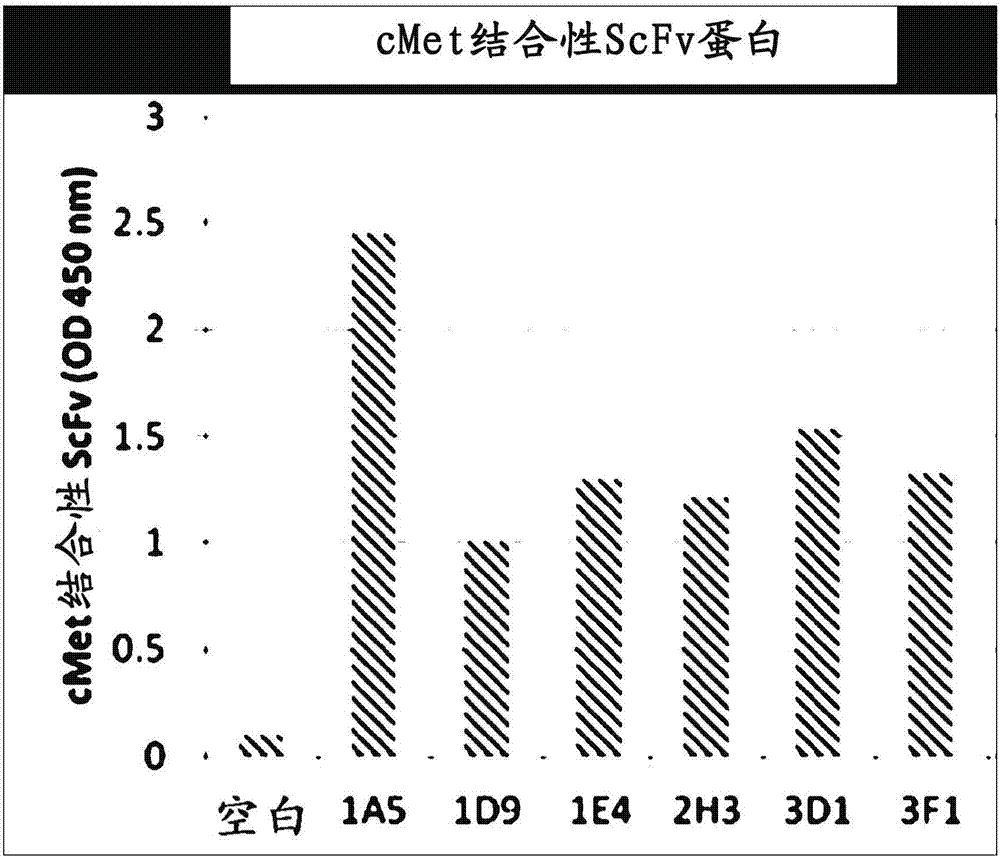
:发现了c-met(间充质上皮转化因子)是细胞表面上的受体,是受体酪氨酸激酶家族的癌基因(致癌基因,oncogene),其由α亚单元β亚单元构成,α亚单元仅由50kd胞外结构域构成,β亚单元由胞外、跨膜、酪氨酸激酶域以及磷酸化相关基序构成,总共145kd(deanetal.,nature,4;318(6044):385,1985;parketal.,pnas,84(18):6379,1987,maggioraetal.,j.cellphysiol.,173:183,1997)。已报导c-met或hgf的不当表达与多种类型的恶性肿瘤相关,且当其过表达时,预后较差(www.vai.org/met,ederetal.,clin.cancerres.,15:2207,2009)。c-met响应hgf且通过c-met的磷酸化刺激多种信号转导通路以促进转化,也就是说,促进癌细胞和血管细胞的分裂及细胞运动性,抑制细胞死亡、以及诱导血管生成以及向细胞外基质(ecm)的侵入和转移等(jeffersetal.,j.mol.med.,74:505,1996;amiconeetal.,emboj.,16:495,1997;matsumotoandnakamura,biochem.biophys.res.comm.,239:639,1997;corpsetal.,int.j.cancer,73:151,1997)。特别是,已报导了c-met和hgf在胶质瘤(koochekpouretal.,cancerres.,57:5391,1997)、乳腺癌(nagyetal.,surg.oncol.,5:15,1996;tucketal.,am.j.pathol.,148:225,1996)、胰腺癌(ebertetal.,cancerres.,54:5775,1994)、胸膜间皮瘤(tolpayetal.,j.cancerres.clin.oncol.,124:291,1998);(klomineketal.intl.j.cancer,76:240,1998)等多种癌症组织和细胞中同时过表达。然而,常常观察到无论hgf如何,通过c-met过表达发展成癌症的情况。例如,肝癌(肝细胞癌,suzukietal.,hepatology,20(5):1231,1996)、胃癌(taniguchietal.,cancer,82:2112-2122(1998))、肺癌(harveyetal.,j.pathol,180:.389,1996)、肾癌(natalietal.,intl.j.cancer,69:212,1996)、卵巢癌(nagyetal,j.surg.oncol,60:95,1995)、结肠直肠癌(hiscoxetal.,cancerinvest.,15:513,1997)等是很好的实例。这样c-met活化或过表达的情况下转化过程被促进,因此,已建立了多种抑制c-met活化的方法作为有前景的抗癌治疗策略。一个实例是设计为干扰atp(腺苷三磷酸)与c-met的相互作用的低分子量化合物。这些低分子量化合物包括k252a(fermentekbiotechnology)、su11274(sugen)、pha-665752(pfiza)等(morottietal.,oncogene,21(32):4885,2002;berthouetal.,oncogene,23(31):5387,2004;pfizer,christensenetal.,cancerres.,63(21):7345,2003),这些实例被设计为干扰c-met的磷酸化,使得信号传输的下游蛋白不被活化。然而,低分子量化合物的缺点是无法特异性地抑制c-met所致的磷酸化。此外,作为中和hgf/c-met信号传输的第二种方法,有抑制c-met和c-met配体hgf结合的方法。抑制c-met和hgf结合的方法包括使用失去的hgf片段(matsumoto&nakamura,cancersci.,94(4):321,2003)、可中和hgf的抗体(caoetal.,pnas.,98(13):7443,2001;kimetal.,clin.cancerres..,12:1292,2006;burgessetal.,cancerres.,66(3):1721,2006)或hgf前体(前-hgf,mazzoneetal.,j.clin.invest.,114(10):1418,2004)的方法,上述hgf前体不能活化c-met,但能比原始hgf更强的亲和力与c-met结合。此外,使用噬菌体展示方法和淘选技术来选出能够抑制c-met活性的肽序列,将相应的肽用于防止c-met的活化(kimetal.,biochembiophysrescommun.,354:115,2007)。选出的肽应用于分子成像,用于搜寻过表达c-met的体内组织或器官(caoetal.,clin.cancerres.,13(20);6049,2007)。虽然所选的肽受到仅抑制hgf依赖性c-met活化的限制,但现实中在体外或体内显示了抗癌效果,因而认为选定的肽可以通过如与现有化疗组合等有效地加以使用。此外,吉非替尼(gefitinib)和厄洛替尼(erlotinib)等表皮生长因子受体(egfr)激酶抑制剂已被用作有效的癌症治疗剂,但经常发生耐药,其原因据报导是c-met受体的强烈扩增(engelmanetal.,science,316(5827):1039,2007),因此,预期当c-met抑制剂合并处理时,抗癌效果会增大。此外,已知c-met由hgf活化,其自身有助于新生血管形成,并已知c-met通过与新生血管形成的主要受体vegfr-2的相互作用刺激新生血管形成。此处,可观察到当同时靶向c-met和vegfr-2加以抑制时,在人异种移植模型中肿瘤生长被协同地抑制(zhangetal.,idrugs,13:112,2010)。已知hgf和c-met间的失调节和c-met的过表达在转移性癌症的进展中起重要作用,因此,在抗癌剂中将c-met的抗体作为拮抗剂进行了大量开发。因此,本发明的发明人为开发能够与c-met以高亲和力特异性结合、由人衍生的序列构成、在体内施用时诱导免疫响应潜力低的c-met人抗体做出了努力,结果,通过使用噬菌体展示方法从单链fv(scfv)噬菌体文库分离了与c-met以高亲和力和特异性结合的完整人抗体,并完成了本发明。技术实现要素:本发明的目的是提供对c-met具有高亲和力和特异性的c-met特异性抗体的重链可变区。本发明的另一目的是提供对c-met具有高亲和力和特异性的c-met特异性抗体的轻链可变区。本发明的另一目的是提供包含c-met特异性抗体的重链可变区和c-met特异性抗体的轻链可变区的c-met特异性抗体或其片段。本发明的另一目的是提供编码所述c-met特异性抗体的重链可变区或所述c-met特异性抗体的轻链可变区的多核苷酸。本发明的另一目的是提供包含所述多核苷酸的重组载体和用所述重组载体转化的重组细胞。本发明的另一目的是提供使用所述重组细胞产生c-met特异性抗体的方法。为实现上述目的,本发明提供c-met特异性抗体的重链可变区,其包含seqidno:1的氨基酸序列所示的重链可变区cdr1;seqidno:2的氨基酸序列所示的重链可变区cdr2;和seqidno:3的氨基酸序列所示的重链可变区cdr3,且优选地,提供具有seqidno:7的氨基酸序列的c-met特异性抗体的重链可变区。此外,本发明提供c-met特异性抗体的轻链可变区,其包含seqidno:4的氨基酸序列所示的轻链可变区cdr1;seqidno:5的氨基酸序列所示的轻链可变区cdr2;和seqidno:6的氨基酸序列所示的轻链可变区cdr3,且优选地,提供具有seqidno:8的氨基酸序列的c-met特异性抗体的轻链可变区。此外,本发明提供c-met特异性抗体或其片段,其包含c-met特异性抗体的重链可变区和c-met特异性抗体的轻链可变区,所述重链可变区包含seqidno:1的氨基酸序列所示的重链可变区cdr1、seqidno:2的氨基酸序列所示的重链可变区cdr2、和seqidno:3的氨基酸序列所示的重链可变区cdr3;所述轻链可变区包含seqidno:4的氨基酸序列所示的轻链可变区cdr1、seqidno:5的氨基酸序列所示的轻链可变区cdr2、和seqidno:6的氨基酸序列所示的轻链可变区cdr3。根据本发明的c-met特异性抗体或其片段可包含seqidno:7所示的重链可变区和seqidno:8所示的轻链可变区。此外,本发明提供编码所述c-met特异性抗体的重链可变区或c-met特异性抗体的轻链可变区的多核苷酸。此外,本发明提供包含上述多核苷酸的重组载体,和使用所述重组载体转化的宿主细胞。此外,本发明提供产生c-met特异性抗体的方法,其包括通过培养上述重组细胞产生c-met特异性抗体;并回收所产生的c-met特异性抗体。附图说明图1显示了本发明中筛选的c-met特异性scfv与c-met结合的能力。图2显示了根据本发明的1e4-scfv与c-met结合的能力。图3显示了包含1e4-igg的重链恒定区和重链可变区的载体pigghd-1e4hvy的图谱。图4显示了包含1e4-igg的轻链恒定区和轻链可变区的载体piggld-1e4lgt的图谱。图5显示了纯化的1e4-igg的sds-page结果。图6显示了通过mdck-2细胞系确认根据本发明的c-met特异性抗体的分散能力(scattering)所得的结果。图7显示了通过蛋白免疫印迹确认根据本发明的c-met特异性抗体的磷酸化能力所得的结果。图8显示了根据本发明的c-met特异性抗体与小鼠c-met和与人c-met结合的能力。最佳模式只要未以其他方式定义,本说明书中使用的全部技术和科学术语具有与本发明所述领域的技术人员所一般理解相同的含义。一般而言,本说明书中使用的命名和下文将进行说明的实验方法是已知的,并且是本
技术领域:
通常使用的。根据本发明的抗体能够与c-met以高亲和力特异性结合,并包含来源于人的序列。在本发明的一个方面中,本发明提供c-met特异性抗体的重链可变区,其包含:seqidno:1的氨基酸序列所示的重链可变区cdr1;seqidno:2的氨基酸序列所示的重链可变区cdr2;和seqidno:3的氨基酸序列所示的重链可变区cdr3。本发明的另一个方面中,本发明提供c-met特异性抗体的轻链可变区,其包含:seqidno:4的氨基酸序列所示的轻链可变区cdr1;seqidno:5的氨基酸序列所示的轻链可变区cdr2;和seqidno:6的氨基酸序列所示的轻链可变区cdr3。本发明的另一个方面中,本发明提供c-met特异性抗体或其片段,其包含c-met特异性抗体的重链可变区和c-met特异性抗体的轻链可变区,所述重链可变区包含seqidno:1的氨基酸序列所示的重链可变区cdr1、seqidno:2的氨基酸序列所示的重链可变区cdr2、和seqidno:3的氨基酸序列所示的重链可变区cdr3;所述轻链可变区包含seqidno:4的氨基酸序列所示的轻链可变区cdr1、seqidno:5的氨基酸序列所示的轻链可变区cdr2、和seqidno:6的氨基酸序列所示的轻链可变区cdr3。具体而言,本发明通过使用噬菌体展示方法从单链fv(scfv)噬菌体文库获得了与c-met以高亲和力和特异性结合的完全人抗体。噬菌体文库和噬菌体展示可通过如美国专利号7,063,943、美国专利号6,172,197等公开的那样制备。通过上述的方法获得并提供的根据本发明的c-met特异性抗体具有重链可变区和轻链可变区,所述重链可变区包含分别具有seqidno:1、2和3的序列的重链可变区cdr1(hcdr1)、重链可变区cdr2(hcdr2)和重链可变区cdr3(hcdr3),所述轻链可变区包含分别具有seqidno:4、5和6的序列的轻链可变区cdr1(lcdr1)、轻链可变区cdr2(lcdr2)和轻链可变区cdr3(lcdr3)。在本发明示例性的实施方案中,根据本发明的c-met特异性抗体是1e4抗体,其包含具有seqidno:7的序列的重链可变区和具有seqidno:8的序列的轻链可变区。此外,本发明提供包含分别由seqidno:1、2和3所示的hcdr1、hcdr2和hcdr3的重链可变区和分别由seqidno:4、5和6所示的lcdr1、lcdr2和lcdr3的轻链可变区。优选地,本发明提供1e4抗体的由seqidno:7所示的重链可变区和1e4抗体的由seqidno:8所示的轻链可变区。【表1】根据本发明的c-met特异性抗体的重链和轻链的cdr和可变区序列如上文所述根据本发明的抗体片段可以为相同的目的使用。本发明的抗体片段包含具有与c-met结合的能力的单链抗体、双抗体、三抗体、四抗体、fab片段、f(ab')2片段、fd、scfv、域抗体、双特异性抗体、微抗体、scab、igd抗体、igm抗体、igg1抗体、igg2抗体、igg3抗体、igg4抗体、抗体恒定区的衍生物、基于蛋白支架的人工抗体等,但本发明不限于此。对于本领域的技术人员而言不言自明的是,只要保持与c-met的结合功能,根据本发明的抗体片段也具有与根据本发明的抗体相同的特征。根据本发明的抗体片段可通过切割完整的抗体或表达编码所述片段的dna而产生。所述抗体片段可通过如已知文件所述的方法产生(lamoyietal.,j.immunol.methods,56:235,1983;parham,j.immunol.,131:2895,1983)。在本发明的scfv中,重链可变区和轻链可变区序列可通过接头彼此连接,其中所述接头优选为氨基酸接头,且根据本发明的1e4抗体的scfv中接头的氨基酸序列可以是seqidno:9的序列,且整个scfv可以是seqidno:10的序列。在本发明示例性的实施方案中,在c-met特异性人抗体1e4的场合,确认了即使无hgf处理c-met亦被磷酸化,且通过使用犬肾上皮细胞mdck-2细胞确认细胞分散能力,结果确认了细胞分散能力与阳性对照组——hgf处理的细胞组在相同水平(图6)。此外,可变区中发生修饰的抗体只要保持本发明抗体的特征也包括在本发明的范围内。作为其实例,可包括可变区中氨基酸的保守取代。保守取代意为使用具有与原始氨基酸序列相似特征的其他氨基酸残基的取代,例如,赖氨酸、精氨酸和组氨酸的相似的特性在于具有碱性侧链;天冬氨酸和谷氨酸的相似特性在于具有酸性侧链。此外,甘氨酸、天冬酰胺、谷氨酰胺、丝氨酸、苏氨酸、酪氨酸、半胱氨酸和色氨酸的相似特点在于具有不带电的极性侧链,且丙氨酸、缬氨酸、亮氨酸、苏氨酸、异亮氨酸、脯氨酸、苯丙氨酸和甲硫氨酸的相似特点在于具有非极性侧链,且酪氨酸、苯丙氨酸、色氨酸和组氨酸的相似特点在于具有具有芳香族侧链。因此,对本领域的技术人员而言不言自明的是,即使即使发生上述具有相似特征的组内的氨基酸置换,也不会导致显著的特征改变,由于在可变区中保守取代而修饰的这些抗体也包括在本发明的范围内,只要其保持本发明抗体的特征。此外,根据本发明的c-met特异性抗体包括通过如直接突变、亲和力成熟、噬菌体展示、链改组等方法具有改进的亲和力和特异性的抗体。可通过突变cdr并筛选对应具有合意特征的抗原结合位点的cdr,来改进和修饰亲和力和特异性(yangetal.,j.mol.bio.,254:392,1995)。cdr可以多种方式突变。例如,一种方法是随机化开发的残基或残基组合,使得在相同抗原结合位点的群体中在特定位置处出现全部20种氨基酸,且可通过引起突变的易错pcr法在整个cdr残基的范围内引入突变(hawkinsetal.,j.mol.bio.,226:889,1992)。在大肠杆菌的突变株中复制包含重链可变区和轻链可变区基因的噬菌体展示载体(噬菌粒)(lowetal.,j.mol.biol.,250:359,1996)。这些诱变方法是本领域中通常使用的方法。此外,根据本发明的抗体或其片段可以与其他材料组合的偶联物的形式使用。因此,本发明提供抗体-药物偶联物,其中组合了根据本发明的c-met特异性抗体和治疗药物。可用于根据本发明的抗体-药物偶联物的治疗药物可不受限制使用,只要其满足本发明的目的。本发明提供药物组合物,其包含根据本发明的抗体或其片段。所述药物组合物可进一步包含药物上可接受的常规载剂、赋形剂等。另一方面,本发明提供编码c-met特异性抗体的重链可变区的多核苷酸、包含所述多核苷酸的重组载体、编码所述c-met特异性抗体轻链可变区的多核苷酸和包含所述多核苷酸的重组载体。编码根据本发明的抗体或其片段的可变区的多核苷酸序列优选具有eqidno:11的重链可变区和seqidno:12的轻链可变区,但本发明不限于此。此外,对本发明所述领域的技术人员(后文称为本领域的技术人员)不言自明的是,可能具有简并序列,即编码与seqidno:7所示的重链可变区和seqidno:8所示的轻链可变区的氨基酸序列相同的氨基酸序列但具有不同多核苷酸序列的序列,且本领域的技术人员显然也能够从seqidno:1-6所示的相应cdr序列推断编码本发明提供的每条重链和轻链的cdr序列的多核苷酸序列。【表2】编码根据本发明的c-met特异性抗体可变区的多核苷酸序列本发明提供包含所述多核苷酸序列的重组载体、包含所述载体的宿主细胞和通过使用重组载体或宿主细胞产生与本发明的c-met(c-met特异性抗体)特异性结合的抗体的方法。具体而言,本发明的c-met特异性抗体优选通过在基因重组方法中包括表达和纯化步骤而产生,且具体地,优选通过分别表达编码本发明的c-met特异性抗体的每一个可变区,或通过在一个宿主细胞中同时表达编码本发明的c-met特异性抗体的可变区而产生。此外,可让编码前导序列的多核苷酸位于本发明抗体的n-末端,用于产生根据本发明的抗体。此外,为了促进分离和纯化,并分析所产生的抗体,可附加标签。代表性的标签是his-标签,但本发明不限于此。本发明中的术语“重组载体”是能够在适当的宿主细胞中表达靶蛋白的表达载体,且表示包含彼此可操作连接的基本控制元件使得基因插入物得以表达的基因构建体。本发明中的术语“可操作连接”意为核酸表达控制序列和编码合意蛋白的核酸序列功能性地彼此连接,以实施一般功能。与重组载体的可操作连接可通过使用本领域已知的基因重组技术实施,位点特异性的dna裂解和连接可通过使用本领域一般知晓的酶容易地实施。本发明适当的表达载体可包括用于膜靶向或分泌的信号序列以及启动子、起始密码子、终止密码子和表达控制元件如多聚腺苷酸化信号和增强子。起始密码子和终止密码子一般视为编码免疫靶蛋白的核苷酸的部分,且当施用基因构建体时,应在注射的受试者中呈现功能,故应该与编码序列合框。一般启动子可以是构成型或诱导型的。原核细胞具有lac、tac、t3和t7启动子,但本发明不限于此。真核细胞具有猴病毒40(sv40)启动子、小鼠乳腺肿瘤病毒(mmtv)启动子、人免疫缺陷病毒(hiv)例如hiv长末端重复(ltr)启动子、莫洛尼病毒(moloneyvirus)、巨细胞病毒(moloneyvirus,cmv)、eb病毒(epstein-barrvirus,ebv)、劳斯肉瘤病毒(roussarcomavirus,rsv)启动子,且还具有β-肌动蛋白启动子、人类血红蛋白、人肌肉肌酸、人金属硫蛋白来源的启动子,但本发明不限于此。表达载体可包含用于选择包含载体的宿主细胞的选择标记。选择标记用于筛选所述载体转化的细胞,可以使用赋予药物耐性、营养缺陷型、对细胞毒性剂的耐性,或表面蛋白的表达等可选择表型的标记。由于只有表达选择标记的细胞才能够在使用选择剂处理的环境中存活,使得转化的细胞能够被选择。此外,当载体为可复制表达载体时,载体可包含复制起点,复制起点是起始复制的特定核酸序列。作为用于插入外源基因的重组表达载体,可使用多种载体如质粒、病毒、粘粒载体等。重组载体的种类没有特别限制,只要其在多种原核细胞和真核细胞的宿主细胞中表达合意的基因,并产生合意的蛋白即可,但优选保有强活性的启动子和强表达力,并能够以与天然状态相似的形式产生大量外源蛋白的载体。为表达根据本发明的二重靶抗体,可使用多种表达宿主/载体组合。适于真核宿主的表达载体的实例包括源自sv40、牛乳头瘤病毒(bovinepapillomavirus)、腺病毒(adenovirus)、腺伴随病毒(adeno-associatedvirus)、巨细胞病毒(cytomegalovirus)、杆状病毒(baculovirus)和逆转录病毒(retrovirus)的表达控制序列,但本发明不限于此。能够在细菌宿主中使用的表达载体的实例包括从大肠杆菌获得的细菌质粒,如pet、prset、pbluescript、pgex2t、puc载体、cole1、pcr1、pbr322、pmb9及其衍生物;具有更大宿主范围的质粒,如rp4;噬菌体dna,如λgt10和λgt11、nm989等非常多样的噬菌体λ衍生物,以及其他dna噬菌体如m13和丝状单链dna噬菌体。具体而言,对于在大肠杆菌中表达,可包括编码邻氨基苯甲酸合酶(trpe)和在羧基末端的多聚接头的dna序列,且其他表达载体系统基于β-半乳糖苷酶(pex);λpl麦芽糖结合蛋白(pmal)和谷胱甘肽s-转移酶(gene67:31,1988;peptideresearch3:167,1990)。当需要在酵母中实施表达时,适用于在酵母中使用的基因是在酵母质粒yrp7中存在的trpl基因(stinchcombetal.,nature,282:39,1979;kingsmanetal,gene,7:141,1979)。trpl基因提供在色氨酸中缺乏生长能力的酵母突变株,如atccno.44076或pep4-1(jones,genetics,85:12,1977)等的选择标记。因此,酵母宿主细胞基因组中trpl损坏的存在通过在缺乏色氨酸条件下的生长而提供了用于检测转化的有效环境。相似地,leu2-缺陷型酵母株(atccno.20,622或no.38,626)可被已知的包含leu2基因的质粒补全。在酵母细胞中有用的表达载体是2μ质粒及其衍生物,在昆虫细胞中有用的载体是pvl941。本发明的表达载体可包含与待表达的dna序列或其片段可操作地连接的一个或多个表达控制序列。表达控制序列插入载体以调节或控制克隆的dna序列的表达。有用的表达控制序列的实例可包括lac系统、trp系统、tac系统、trc系统、噬菌体λ的主要的操纵基因和启动子区、fd包被蛋白的控制区、酵母的糖酵解启动子如3-磷酸甘油酸激酶的启动子、酵母酸性磷酸酶的启动子如pho5的启动子和酵母α-交配因子和源自多瘤病毒、腺病毒、逆转录病毒和猴病毒的启动子如sv40的早期和晚期启动子,和其他已知控制原核细胞或真核细胞及其病毒的基因表达的序列,及其组合。另一方面,本发明涉及使用所述重组载体转化的重组细胞。在本发明中,所述重组细胞是使用包含编码c-met特异性抗体的重链可变区的多核苷酸的重组载体和包含编码c-met特异性抗体的轻链可变区的多核苷酸的重组载体转化的细胞。另一方面,本发明提供使用所述重组细胞或其片段产生c-met特异性抗体的方法,其中所述方法包括培养重组细胞并从所述培养的重组细胞分离c-met特异性抗体。优选地,根据本发明的c-met特异性抗体优选根据基因重组方法通过表达和纯化获得。具体地,编码抗体的重链可变区或完整重链区的基因序列和编码轻链可变区或完整轻链区的基因序列可在一个载体中、或分别在两个载体中表达,但本发明不限于此。具体地,产生c-met特异性抗体的方法可包括:通过将编码本发明的c-met特异性抗体的核苷酸序列插入载体产生重组载体;用该重组载体转化宿主细胞并培养;并从所述培养的转化体分离并纯化c-met特异性抗体。此外,c-met特异性抗体可通过在营养培养基中培养表达的重组载体的而转化体大量制造,培养基和培养条件可取决于宿主细胞使用适当地选择。温度、培养基的ph、培养时间等条件可适当控制,使其适于培养时的细胞生长和蛋白的大量生产。如上文所述重组产生的c-met特异性抗体可从培养基或细胞裂解物回收。在膜偶联类型的情况中,可通过使用适当的表面活性剂溶液(例如:tritone-x100)或酶裂解来分离膜。在表达c-met特异性抗体中使用的细胞可由多种物理或化学方式如冻融纯化、超声处理、机械破坏和细胞分解剂,可通过一般的生化分离技术分离和纯化(sambrooketal.,molecularcloning:alaboratorymanual,2nded.,coldspringharborlaboratorypress(1989);deuscher,m.,guidetoproteinpurificationmethodsenzymology,vol.182.academicpress.inc.,sandiego,ca(1990))。可用的方法有电泳、离心、凝胶过滤、沉淀、透析、色谱法(离子交换色谱、亲和色谱、免疫吸附色谱色谱、尺寸排阻色谱等)、等电聚焦和多种改变和复杂方法,但本发明不限于此。此后,本发明将参照下述实施例进行更详细的说明。然而,下述实施例仅为例示本发明,本领域的技术人员显然实现本发明的范围不应理解为受这些实施例限制。实施例1:生物淘选在生物淘选中使用的完全人非免疫单链抗体(scfv)噬菌体展示文库是在之前注册的专利中(韩国专利号10-0883430)建立的噬菌体展示文库。为回收其中展示单链的噬菌体,使噬菌体文库储液生长至对数期,并通过m13k07辅助噬菌体(gehealthcare,usa)拯救,且随后通过添加1mmiptg至包含34μg/ml的氯霉素(cm)和70μg/ml的卡那霉素的2xyt培养基,在30℃扩增过夜。使用人fc去除显示抗人fcscfv的噬菌体。为阻断其他非特异性结合,让噬菌体制备物在4%peg6000/0.5mnacl中沉淀,并在包含500μg/ml的人fc蛋白的2%脱脂牛奶/pbs中重悬,随后在37℃温育1小时。对于用5μg/ml的c-met-fc(r&dsystems,重组人hgfr/c-metfc嵌合(目录号358-mt/cf))包被的96孔nunc-immuno板(nunc,denmark),首先使用2%脱脂牛奶/pbs在室温封闭2小时,然后在室温接种3x1011pfu的噬菌体制剂1小时。使用pbst(包含0.1%tween20的pbs)洗涤管5次,再用pbs洗涤5次。结合型的噬菌体用100mm三乙胺溶液在室温历时10分钟洗脱。将洗脱的噬菌体与10ml的中对数期(mid-log)xl1-blue细胞共同在37℃静置30分钟,而后震荡培养30分钟。随后,将经感染的xl1-blue细胞在30℃于包含1%葡萄糖的2xyt/cm+板上培养过夜。在首次生物淘选后,通过将洗涤次数顺次增加至10次、15次和20次来实施第二、第三和第四次生物淘选。实施第4次淘选后,再次通过c-met结合测定确认所获得的噬菌体的c-met的结合能力。为实施c-met结合测定,将用100ng的c-met(r&dsystem)包被过夜的微孔板与2%脱脂牛奶/pbs在37℃反应2小时。使用pbs洗涤微孔板,并将每次淘选获得的噬菌体在室温反应1小时。使用pbs洗涤反应溶液,用hrp(辣根过氧化物酶)偶联的小鼠抗m13抗体(gehealthcare)在室温反应1小时。反应完成后的孔通过使用tmb溶液(bd)诱导显色并在450nm测量吸收(图1)。结果确认了1e4、1a5、1d9、2h3、3d1和3f1具有结合c-met的能力。实施例2:可溶性scfv的产生和纯化为了制造与实施例1中已确认了c-met结合能力的1e4-scfv、1a5-scfv、1d9-scfv、2h3-scfv、3d1-scfv和3f1-scfv相应的可溶性1e4-scfv、1a5-scfv、1d9-scfv、2h3-scfv、3d1-scfv和3f1-scfv,将通过生物淘选获得的pak-1e4、pak-1a5、pak-1d9、pak-2h3、pak-3d1和pak-3f1使用xbai和ecori切割,获得具有1e4-scfv、1a5-scfv、1d9-scfv、2h3-scfv、3d1-scfv和3f1-scfv序列的片段,并将这些片段插入使用相同限制酶获得的大肠杆菌表达载体(pet21b-6a6,novagen,u.s.a.),产生pet21b-1e4、pet21b-1a5、pet21b-1d9、pet21b-2h3、pet21b-3d1和pet21b-3f1。将产生的pet21b-1e4、pet21b-1a5、pet21b-1d9、pet21b-2h3、pet21b-3d1和pet21b-3f1转化入大肠杆菌bl21(de3)用于蛋白表达,利用转化体表达可溶性scfv蛋白,再离心细胞。随后,通过使用包含20%蔗糖,200μg/ml溶菌酶和蛋白酶抑制剂鸡尾酒(roche,switzerland)的50mmtris(ph8.0)溶液获得细胞的周质级分。获得的级分用ni-nta离心试剂盒(qiagen,germany)处理以纯化scfv蛋白。实施例3:使用scfv的c-met结合测定确认了实施例2中纯化的可溶性1e4-scfv、1a5-scfv、1d9-scfv、2h3-scfv、3d1-scfv和3f1-scfv是否具有结合c-met的能力。为此,用100ngc-met包被96孔nunc-immuno板(nunc,denmark),于4℃过夜,随后与2%脱脂牛奶/pbs在37℃反应2小时。反应完成后,使用pbs洗涤平板,将纯化的scfv添加到其中,室温反应1小时。反应完成后,使用pbs洗涤平板,添加hrp偶联的抗myc小鼠抗体(sigma,usa)在室温反应1小时。随后,使产物与tmb溶液反应,并在450nm测量吸收。结果,如图2所示,确认了纯化的1e4-scfv、1a5-scfv、1d9-scfv、2h3-scfv、3d1-scfv和3f1-scfv全部具有结合c-met的能力。实施例4:igg的表达和纯化为表达成为完整的igg形式,产生了包含完整的恒定区的重链表达载体和轻链表达载体。对于重链,将使用sfii处理过的具有人4-1bb重链骨架的pigghd载体(aprogen,korea)与使用sfii处理pakscfv的重链可变区获得的片段连接,产生包含完整重链恒定区和重链可变区的表达载体pigghd-1e4hvy、pigghd-1a5hvy、pigghd-1d9hvy、pigghd-2h3hvy、pigghd-3d1hvy和pigghd-3f1hvy(图3)。对于轻链,将使用bstxi处理的具有人4-1bb的轻链骨架的piggld载体(aprogen,korea)与使用bstxi处理pak-scfv的轻链可变区获得的片段连接,产生了包含完整轻链恒定区和轻链可变区的表达载体piggld-1e4lgt、piggld-1a5lgt、piggld-1d9lgt、piggld-2h3lgt、piggld-3d1lgt和piggld-3f1lgt。为了表达igg,将相同量的轻链表达载体和重链表达载体共转染入hek-293t细胞中。将转染的细胞在无血清培养基freestyle293中培养(37℃,5%co2),随后收集培养基。根据制造商所述的方案,使用蛋白a柱(gehealthcare,usa)通过亲和层析从上清液纯化1e4-igg、1a5-igg、1d9-igg、2h3-igg、3d1-igg和3f1-igg。将上清液注射入以20mm磷酸钠(ph7.0)和100mmnacl溶液平衡的蛋白a柱,并使用包含20mm磷酸钠(ph7.0)、1mmedta和500mmnacl的溶液洗涤,随后使用包含100mmnacl的0.1m甘氨酸-hcl(ph3.3)溶液洗脱。洗脱的蛋白使用1mtris中和。将洗脱的蛋白与5mm磷酸钠(ph6.0)以1:1的比率混合,并注射入用含有50mmnacl的5mm磷酸钠(ph6.0)平衡的预装填sp-琼脂糖(gehealthcare)柱。使用包含50mmnacl的磷酸钠(ph7.0)缓冲液洗脱与柱结合的蛋白,并注射入使用洗脱缓冲液平衡的预装填q-sepharose(gehealthcare),由此获得未结合的蛋白。获得的蛋白通过30kdvivaspin20(sartorius)浓缩并使用pbs透析。通过上述方法纯化的1e4-igg、1a5-igg、1d9-igg、2h3-igg、3d1-igg和3f1-igg蛋白中若干者的sds-page结果示于图5中。实施例5:c-met特异性抗体的分散能力(scattering)分析为评估6种c-met特异性抗体的分散能力,培养了犬肾上皮细胞mdck-2(madin-darby犬肾上皮细胞-2)。在24孔板中以2x104细胞/孔的浓度,使用添加有5%胎牛血清的dmem培养细胞。培养1天后,一旦细胞良好附着于平板生长,使用10μg/ml的抗体或30ng/ml或50ng/ml的hgf处理细胞。培养16小时后,通过使用水晶紫(sigma-aldrich,u.s.a)染色并拍摄图像来评估细胞的分散和生长(图6)。结果,如图6中所示,可以确认在6种抗体中,仅在1e4处理时细胞与阳性对照组hgf处理组的分散和生长相同。实施例6:通过蛋白免疫印迹的体内c-met磷酸化分析为再次确认如实施例5中所示的1e4作为激动剂的活性,通过蛋白免疫印迹测量了c-met特异性抗体1a5和1e4对c-met的磷酸化活性。将过表达c-met的直肠癌细胞系ht29细胞以2x105在6孔板中培养24小时,并在不包含胎牛血清的rpmi培养基条件下于无血清培养基中处理6小时,然后用10μg/ml的higg和c-met抗体预处理30分钟。随后在处理组中处理30ng/ml的人hgf15分钟。为了蛋白免疫印迹分析,用裂解缓冲液(1%(w/v)sds、10mmtris(ph7.4)、1mmna3vo4(原钒酸钠)、2mmegta、2mmedta、1mm苯甲基磺酰氟、1mm氟化钠和1x蛋白酶抑制剂结尾酒(sigma))处理获得裂解物,将裂解物煮沸,随后在4℃以10,000g离心5分钟以去除不可溶的沉淀。将上清与sds样品缓冲液混合并煮沸10分钟备用。sds-page和蛋白免疫印迹通过本领域公知的方法实施,且使用的样品如下提供:4-20%sds-聚丙烯酰胺凝胶(biorad,u.s.a.)、pvdf膜(millipore#ipvh00010,u.s.a.)、抗c-met抗体(santacruz,u.s.a.)和抗磷酸化-c-met抗体(cellsignalingtechnology,u.s.a.)作为一抗用于分析c-met磷酸化活性;抗p-erk抗体(cellsignalingtechnology,u.s.a.)和hrp-偶联的山羊抗小鼠igg抗体(santacruzbiotechnology,u.s.a.)和hrp-偶联的山羊抗兔igg(cellsignalingtechnology,u.s.a.)作为与一抗结合用于化学发光的二抗(图7)。结果,如图7中所示,可以确认1e4即使在单独处理时也具有磷酸化c-met的能力。实施例7:抗c-metigg(1e4-igg)结合能力的分析通过biacore(gehealthcare)测量抗体与c-met的结合能力。根据制造商的说明,通过在cm5芯片(gehealthcare,sweden)上固定小鼠c-met和人c-met(r&dsystem),并使不同量的抗体流过而获得传感图。以在每种浓度获得的传感图为基础测定速率常数kon和koff,并从速率常数的比率(koff/kon)计算kd(表3)。结果,1e4-igg不但具有与人c-met的结合能力,还具有与小鼠c-met的高结合能力(图8)。【表3】根据本发明的抗体的ka、kd和kd值ka(1/ms)kd(1/s)kd(m)小鼠c-met-fc2.58x1061.59x10-36.15x10-10人c-met-fc2.71x1068.07x10-42.98x10-10尽管已对本发明的特定实施方案进行了详述,对本领域的技术人员将显而易见的是,具体的说明仅为优选的示例性实施方案,不应理解为限制了本发明的范围。因此,本发明的实质范围将由随附的权利要求及其等同物所限定。工业实用性根据本发明的c-met特异性抗体能够特异性地与c-met以高亲和力结合并具有激动剂功能。当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1
- 用于治疗HBV感染和相关病症的三特异性结合分子的制造方法与工艺
- 对磺胺类药物具有特异性的固相微萃取纤维的制备方法与流程
- 一种检测猪初乳中抗PEDV特异性IgA抗体的夹心ELISA试剂盒的制造方法与工艺
- 用于施用CD19xCD3双特异性抗体的给药方案的制造方法与工艺
- 双特异性抗‑VEGF/抗‑ANG‑2抗体及其在治疗眼血管疾病中的应用的制造方法与工艺
- 特异性人EWS‑FLI1融合蛋白抗体的制备方法及在制备尤文肉瘤诊断抗体试剂的应用与制造工艺
- 双特异性四价抗体及其制造和使用方法与制造工艺
- 利用特异性挥发物筛选昆虫相关嗅觉蛋白的方法与制造工艺
- 使用与表面偶联的单特异性四聚抗体复合物组合物从样品中分离出靶实体的方法与制造工艺
- 抗-WT1/HLA双特异性抗体的制造方法与工艺
- 与CD56分子特异性结合的多肽及其应用的制造方法与工艺
- 用于治疗HBV感染和相关病症的三特异性结合分子的制造方法与工艺
- 对磺胺类药物具有特异性的固相微萃取纤维的制备方法与流程
- 避免信号干扰的肌原纤维蛋白丝氨酸磷酸化水平的测定方法与流程
- 一种检测猪初乳中抗PEDV特异性IgA抗体的夹心ELISA试剂盒的制造方法与工艺
- 用于施用CD19xCD3双特异性抗体的给药方案的制造方法与工艺
- 双特异性抗‑VEGF/抗‑ANG‑2抗体及其在治疗眼血管疾病中的应用的制造方法与工艺
- 特异性人EWS‑FLI1融合蛋白抗体的制备方法及在制备尤文肉瘤诊断抗体试剂的应用与制造工艺
- 活化人CD40的抗体和针对人PD‑L1的抗体的组合疗法的制造方法与工艺
- 双特异性四价抗体及其制造和使用方法与制造工艺