稻瘟病菌的突变体菌株K2e12及其用途的制作方法
本发明涉及一种稻瘟病菌的突变体K2e12,以及其在使植物不致病上的用途,属于微生物及微生物应用领域。
背景技术:
药物靶标是药物作用的对象,通常是具有重要生理或者病理功能,能够与药物相结合并产生药理作用的生物大分子。药物靶标的筛选和功能研究是发现特异、高效、低毒性药物的前提。在病害真菌中,药物靶标应该该具备以下特征:首先它们应该是菌体生长所必需的基因或基因产物并且具有高度保守和特异的序列,这样才能筛选出对菌体高度特异而对人体或其他宿主无害的药物;其次,药物靶标应该是病原真菌生长和生存必不可少的,至少在侵染宿主的过程中至关重要;最后药物靶标的生化功能要确认,必须有结合小分子化合物的化学特性。目前应用的抗真菌药物按其靶点大致可分为三类:作用于真菌细胞膜的抗真菌药物、作用于核酸合成的抗真菌药物和作用于细胞壁合成的抗真菌药物。第一类药物靶标的代表是真菌细胞膜合成中的关键酶羊毛甾醇14α-去甲基化酶,使用氮唑类抗真菌药物可以抑制该酶的活性,从而阻断真菌细胞膜合成,导致真菌破裂死亡。第二类药物的代表5-氟胞嘧啶(5-FC)进入真菌细胞后可以被转化5-氟脱氧鸟嘧啶单磷酸从而抑制胸苷酸合成酶活性进而影响DNA的合成。第三类靶标的代表为β-1,3-葡聚糖合成酶,作为真菌细胞壁特有多糖的合成酶是一类比较理想的靶点。
近年来,随着基因组学、蛋白质组学、生物信息学的快速发展,新的药物靶标筛选技术不断出现,如DNA芯片技术、蛋白质芯片技术、计算机辅助药物设计技术、组合化学技术、化学遗传学技术。这些技术大大提高了药物靶标筛选的效率,加快了新药研发的速度。从筛选出的潜在药物靶标到最终成为药物靶标还需要进一步的确认。目前药物靶标的确认主要采用基因敲除、RNAi和抗体捕获技术等技术,其中基因敲除具有其它方法难以比拟的直接性和有效性,仍是目前确认药物靶标最有力的手段。高通量基因敲除技术在植物病原真菌中的使用使得通过直接敲除基因筛选药物靶标成为可能。
蛋白激酶是一类催化蛋白质磷酸化反应的酶,它通过把ATP或GTP的γ-磷酸基团转移到底物蛋白的氨基酸残基上来活化底物。在真核细胞中,大约有30%蛋白质都有共价连接的磷酸基团,由此可见蛋白激酶在生命活动中扮演的重要角色。事实上蛋白激酶参与几乎所有的细胞过程,如细胞代谢、转录、细胞循环、细胞的骨架构建、凋亡、分化、信号传导、细胞间互作、细胞与细胞外基质相互作用等。蛋白激酶广泛存在于真核生物中,从低等的原生动物到高等的哺乳动物,从低等藻类到高等植物以及真菌中都有蛋白激酶。由于蛋白激酶的成员众多,Hanks和Hunter把它们归类为蛋白激酶超家族。
真核生物蛋白激酶的催化结构域由250-300个氨基酸残基组成,其氨基酸序列保守,又可分为12个功能亚域。Hanks和Hunter对人类370种蛋白激酶的氨基酸序列进行比对,以依赖cAMP蛋白激酶α催化亚基(PKA-Cα)作对照,发现亚域Ⅰ的Gly50和Gly52,亚域Ⅱ的Lys72,亚域Ⅲ的Glu91,亚域ⅥB的Asp171,亚域Ⅶ的Asp184和Gly186,亚域Ⅷ的Clu208,亚域Ⅸ的Asp220和Gly225,亚域Ⅺ的Ary280在95%的序列中保守。其中VIB、VIII和IX在整个蛋白激酶超家族中非常保守,而不同类型蛋白激酶的区别主要集中在Ⅵ、Ⅷ和Ⅸ亚域。蛋白激酶催化核心的功能包括:结合和定向ATP或GTP,该过程有二价阳离子(Mg2+或Mn2+)参与;结合和定向蛋白质或多肽;催化ATP或GTPγ-磷酸基团转移到受体蛋白氨基酸残基上。
蛋白激酶可以磷酸化底物蛋白的9种氨基酸残基,分别是丝氨酸(serine)、苏氨酸(threonine)、酪氨酸(tyrosine)、半胱氨酸(cysteine)、精氨酸(arginine)、赖氨酸(lysine)、天冬氨酸(aspartate)、谷氨酸(glutamate)和组氨酸(histidine)。根据其催化底物残基的不同可以将其分成5类,其中丝氨酸和苏氨酸蛋白激酶与酪氨酸蛋白激酶是最主要的两类。在人类激酶组学研究中,对2244个蛋白质上6600个磷酸化位点进行分析,发现磷酸化的丝氨酸(pSer)、苏氨酸(pThr)和酪氨酸(pTyr)位点分别占86.4%,11.8%,和1.8%也表明了这一点。
Hanks和Hunter在蛋白激酶的研究中做出了突出贡献,他们第一次在分子系统发生层面描述了人类蛋白激酶超家族,奠定了后人研究蛋白激酶分类的基础。Manning在Hanks和Hunter研究的基础上,将所有的激酶按组,家族,亚家族三种层次分类并将人类的蛋白激酶分类扩展到9个组,134个家族,201个亚家族,这为研究人类与模式生物相关激酶的功能和进化关系提供了便利。蛋白激酶的分类主要依据是催化结构域的序列相似程度,催化结构域以外的序列和结构的相似度以及生化功能也是分类的重要依据。按照Manning的分类,人类的518种蛋白激酶分成“传统”的蛋白激酶(ePK)和“非典型”的蛋白激酶(aPK)两大类。ePK含478种蛋白激酶,又被分成9个组:AGC,CAMK,CK1,CMGC,RGC,STE,TK,TKL,Other;aPK含40种蛋白激酶,被分为8个组:Alpha,PIKK,PDHK,RIO,A6,ABC1,BRD,Other。由于aPK在序列上与ePK并没有明确的相似性,但aPK具有蛋白激酶的功能,因此单独分为一类。
技术实现要素:
本发明要解决的技术问题是提供一种稻瘟病菌的突变体菌株K2e12及其用途。
为了解决上述技术问题,本发明提供一种稻瘟病菌的突变体菌株K2e12,为Magnaporthe oryzae K2e12(稻瘟病菌Magnaporthe oryzae K2e12);其保藏号为:CCTCC NO:M 2016107。
作为本发明的稻瘟病菌的突变体菌株K2e12的改进:该稻瘟病菌的突变体菌株K2e12是MGG_07012基因缺失菌株。
本发明还同时提供了上述稻瘟病菌的突变体菌株K2e12的用途:用于侵染植物,使植物不致病;所述病为稻瘟病。
作为本发明的稻瘟病菌的突变体菌株K2e12的用途的改进:所述植物为水稻或大麦;水稻例如为Oryzae sativa cv.CO-39;大麦例如为Hordeum vulgare cv.Golden promise。
本发明还同时提供了上述稻瘟病菌的突变体菌株K2e12的另一种用途:利用K2e12蛋白的表达或修饰作为靶标,筛选抗真菌的药物。
本发明的菌株Magnaporthe oryzae K2e12,其保藏信息具体如下:
保藏单位:中国典型培养物保藏中心;保藏地址:中国武汉武汉大学;保藏日期:2016年3月14日,保藏名称:Magnaporthe oryzae K2e12;保藏号:CCTCC NO:M 2016107。
本发明所涉及的稻瘟病菌突变体菌株K2e12,确切的说是MGG_07012基因缺失突变体,更确切的说与稻瘟病菌野生型菌株Guy11相比,K2e12在基因组DNA水平上的差异在于K2e12不含MGG_07012基因,即稻瘟病菌突变体菌株K2e12是MGG_07012基因缺失菌株。在蛋白水平上的差异在于K2e12不含MGG_07012蛋白,在表达水平上的差异在于MGG_07012基因在K2e12中不表达。
综上所述,菌株Magnaporthe oryzae K2e12来源于稻瘟病菌的突变体,属于真菌界,子囊菌门,该菌株的致病作用丧失。
附图说明
下面结合附图对本发明的具体实施方式作进一步详细说明。
图1为双重PCR验证突变体结果图;
图1中的A代表第一轮PCR结果:将验证目的基因的引物K2e12innerF/innerR和内参基因β-tubulin验证引物tubulinF+tubulinR加入到同一反应体系里进行PCR,经琼脂糖电泳后会出现如图1A所示的结果,K2e12突变体中仅有一条条带,而野生型Guy11中有两条条带;图1中的B代表第二轮PCR结果,电泳后条带如图1B所示,K2e12突变体中有一条同源置换的条带,而野生型Guy11中没有条带。两轮PCR的结果显示,稻瘟病菌MGG_07012基因在K2e12中已经缺失。
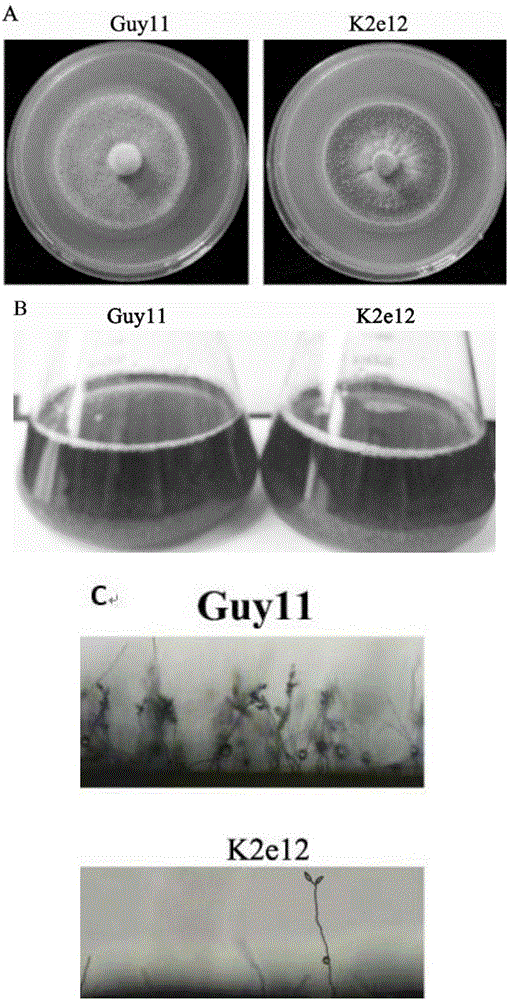
图2为K2e12与野生型菌株Guy11的对比图,
A为2种菌株在CM培养基上25℃暗培养7d后的菌落形态;
B为2种菌株的液体CM培养,K2e12与野生型Guy11无差别;
C为2种菌株在CM固体培养基上菌丝和孢子的形态比较;与野生型Guy11相比,K2e12气生菌丝稀疏,几乎不产生分生孢子。
图3为稻瘟病菌突变体菌株和野生型菌株在大麦离体叶片上的致病性测试试验,图3的下图为K2e12对大麦叶片致病性丧失,没有可见病斑;相比之下,图3的上图接种了野生型菌株的在接种处产生面积较大的扩散性黑色病斑,而且病斑会快速沿着叶脉扩散至周围的健康区域,病斑中央区域逐渐腐烂,并存在着大量的菌丝;因此得知:菌株K2e12对大麦的致病性丧失。
图4为稻瘟病菌突变体菌株和野生型菌株在水稻苗上的致病性测试试验,图4的下图为K2e12对水稻丧失致病性,没有产生任何病斑;相比之下,图4的上图为野生型菌株在水稻上产生典型的梭形病斑;因此得知:菌株K2e12对水稻的致病性丧失。
具体实施方式
下面结合具体实施例对本发明进行进一步描述,但本发明的保护范围并不仅限于此。
参照上述附图,对本发明的具体实施方式进行详细说明。
备注说明:下述所有的培养基使用前均需要按照常规方式进行高温高压的灭菌处理(121℃、0.24MPa、20分钟)。
实施例1、敲除载体pKO1B-K2e12的构建
使用ClonExpressTM MultiSOne Step Cloning Kit(诺维赞公司)进行敲除载体的构建:首先利用引物K2e12upF+upR和K2e12downF+downR分别扩增目的基因的上游和下游片段,长度约为1.5k;然后用引物BarF+BarR扩增草铵膦抗性基因(引物序列如下)。随后利用该试剂盒将PCR得到三个片段和经XbaI和HindIII双酶切得到的pKO1B线性载体重组成目地载体pKO1B-K2e12。
引物序列:
K2e12upF:5’-GGTACCCGGGGATCCTCTAGACCGTAAGTTCAGGGTCATCATC-3’
K2e12upR:5’-CTCCTTCAATATCATCTTCTAGATGCGGCGATGTCTCC-3’
K2e12downF:5’-TGCCCGTCACCGAGATTTAGTCGTCTGCGGAGTCTGTC-3’
K2e12downR:5’-ACGACGGCCAGTGCCAAGCTTATGAAGAGTTGGTGGGATGAC-3’
BarF:5’-AGAAGATGATATTGAAGGAG-3’
BarR:5’-CTAAATCTCGGTGACGGGCA-3’。
实施例2、稻瘟病菌突变体菌株的获得
农杆菌(Agrobacterium tumefaciens)菌株AGL1及质粒pCAMBIA1300等常规物均能通过常规方式获得。农杆菌一般在28℃黑暗培养,生长的基本培养基为LB液体培养基,在4%的甘油中-70℃下保存。
CM(Complete Medium)培养基:20×Nitrate salts 50ml,1000×Trace Elements 1ml,D-glucose(葡萄糖)10g,Peptone(蛋白胨)2g,Yeast extract(酵母提取物)1g,Casamino acid(酪蛋白氨基酸)1g,1000×Vitamin solution 1ml,用1mol/L NaOH调pH值至6.5,加蒸馏水至1L。
CM固体培养基为上述CM(Complete Medium)培养基的基础上加入琼脂粉12g。
20×Nitrate salts(1000ml)的配置:NaNO3 120g,KCl 10.4g,MgSO4.7H2O 10.4g,KH2PO4 30.4g,水定容至1L。
1000×Trace Elements(100ml)的配置:ZnSO4.7H2O 2.2g,H3BO3 1.1g,MnCl2.4H2O 0.5g,FeSO4.7H2O 0.5g,CoCl2.6H2O 0.17g,CuSO4.5H2O 0.16g,Na2MoO4.5H2O 0.15g,Na4EDTA5g,水定容至100ml。
1000×Vitamin solution(100ml)的配置:Biotin 0.01g,Pyridoxin 0.01g,Thiamine 0.01g,Riboflavin 0.01g,PABA(p-aminobenzonic acid)0.01g,Nicotinic acid 0.01g,水定容至100ml。
水琼脂培养基:蒸馏水加1.5%琼脂粉(即琼脂粉与蒸馏水的质量比为1.5:98.5),高压灭菌,用于单孢分离。
LB液体培养基(Luria-Bertani):Tryptone(胰蛋白胨)10g、Yeast extract(酵母提取物)5g、NaCl(氯化钠)10g,加去离子水至1000ml,用NaOH溶液调至pH7.5,高压灭菌。
LB固体培养基:Tryptone(胰蛋白胨)10g、Yeast extract(酵母提取物)5g、NaCl(氯化钠)10g、琼脂粉12g,加去离子水至1000ml,用NaOH溶液调至pH7.5,高压灭菌。
农杆菌诱导培养基AIM配方:0.8ml 1.25K-Phosphate-buffer pH 4.8(用KH2PO4和K2HPO4配制),20ml MN-buffer(30g/l MgSO4·7H2O,15g/l NaCl,1L H2O2),1ml 1%CaCl2·2H2O(w/v),10ml 0.01%FeSO4(w/v),5ml spore elements(100mg/l ZnSO4·7H2O,100mg/l CuSO4·5H2O,100mg/l H3BO3,100mg/l Na2MoO4·2H2O,1L H2O2)(过滤灭菌),2.5ml 20%NH4NO3(w/v),10ml 50%glycerol(v/v),40ml 1M MES pH 5.5(用NaOH溶液调pH值),20%glucose(w/v):液体培养基中加10ml,固体培养基加5ml,加水至1L,固体培养基加1.5%琼脂粉,(200mM AS-用二甲基亚砜DMSO配制)。
上述所有的培养基均需要进行高温高压的灭菌处理(121℃、0.24MPa、20分钟)。
在实验室条件下,按照农杆菌介导的转化方法即可获得本发明的稻瘟病菌突变体菌株。具体如下:
首先,农杆菌感受态制备:将构建好的敲除载体pKO1B-K2e12通过冻融法转化到农杆菌菌株AGL1。
将农杆菌菌株在LB平板上活化,然后转到5mL LB液体培养基中28℃振荡过夜培养;转移2mL此培养液到装有50mL LB液体培养基的三角瓶中28℃振荡培养至OD600 0.5~1.0;冰上放置停止生长,4℃3000g离心5min;沉淀用1mL预冷的20mM CaCl2溶液悬浮,分装到预冷却的Eppendorf离心管中,每管0.1mL;
加1μg的载体(敲除载体pKO1B-K2e12)质粒DNA到上述离心管中,液氮中冷冻;37℃水浴5min解冻;加入1mLLB液体培养基,28℃轻摇培养2-4h;将培养好的菌液涂于含有卡那霉素(含50μg/ml)的LB平板上,正面放置30min,待菌液完全被培养基吸收后,倒置培养皿,28℃培养2-4d。
其次,稻瘟病菌的T-DNA转化:从上述培养的LB平板(含50μg/ml卡那霉素)上挑选农杆菌单菌落接种于5ml LB液体培养基(含50μg/ml卡那霉素),200rpm/min,28℃过夜培养;第二天将200-400μl培养液转移到5ml含50μg/ml卡那霉素的诱导液体培养基(AIM)中,OD值约0.15,28℃培养5~6个小时使OD600达到0.5~0.6;用10ml灭菌蒸馏水从培养大约10天的CM平板上洗下稻瘟病菌野生型分生孢子,三层擦镜纸过滤后用血球计数板计数,并用无菌水稀释孢子浓度为106个/ml;取100μl培养好的农杆菌AGL-1(含载体)菌液和100μl稀释好的稻瘟病菌分生孢子悬浮液混合,混合液(200μl)均匀涂于AIM平板上的硝酸纤维素膜表面,AIM平板含200μM乙酰丁香酮(AS,acetosyringone)或不含AS作为对照,22℃共培养48小时;然后将硝酸纤维素膜转移到含草铵膦与抗生素的DCM选择平板上,选择性培养基中含300μg/ml草铵膦、400μg/ml头孢霉素(cefotaxime)、60μg/ml链霉素(streptomycin),将平皿置于28℃下培养到转化子出现。
敲除载体pKO1B-K2e12被转入农杆菌,随后进行农杆菌介导的转化。农杆菌介导的转化得到的转化子提取基因组后需进行两轮PCR验证:第一轮为双重PCR,将验证目的基因的引物K2e12innerF/innerR和内参基因β-tubulin验证引物tubulinF+tubulinR加入到同一反应体系里进行PCR,经琼脂糖电泳后会出现如图1A所示的结果,K2e12突变体中仅有一条条带,而野生型Guy11中有两条条带;然后进行第二轮PCR,该轮PCR的引物为K2e12longF+LbarR,电泳后条带如图1B所示,K2e12突变体中有一条同源置换的条带,而野生型Guy11中没有条带。两轮PCR的结果显示,稻瘟病菌MGG_07012基因在K2e12中已经缺失。
稻瘟病菌MGG_07012基因,编码548个氨基酸,含有蛋白激酶结构域,属于AGC组,改组包括依赖环核苷酸的家族(PKA和PKG)、蛋白激酶C家族、β-肾上腺素受体激酶(βARK)、核糖体S6家族,这些激酶的催化位点是底物碱性氨基酸(Lys和Arg)附近的丝氨酸和苏氨酸残基。PKA,PKG,S6的底物从N端到磷酸结合位点间有特定碱性氨基酸残基。PKC和RAC的底物磷酸化位点两边都有碱性氨基酸残基。而G蛋白偶联的受体激酶(βARK)却相反,它的底物催化位点附近是酸性环境。稻瘟病菌突变体K2e12,是在稻瘟病菌野生型菌株Guy11中,利用草铵膦抗性基因置换MGG_07012基因所得,因此在突变体K2e12中,缺失MGG_07012基因,没有该基因的表达,同时没有该基因的蛋白存在。突变体K2e12与稻瘟病菌野生型菌株Guy11有显著差异,突变体K2e12的菌丝稀薄,几乎不产生分生孢子,对水稻和大麦的致病性丧失。相比较之下,稻瘟病菌野生型菌株Guy11中,MGG_07012基因正常表达,菌株Guy11的气生菌丝浓密,能够产生大量的分生孢子,对水稻和大麦有很强致病能力。
本发明获得基因敲除突变体3个,将其中1个(K2e12)进行保藏,保藏信息具体如下:
保藏单位:中国典型培养物保藏中心;保藏地址:中国武汉武汉大学;保藏日期:2016年3月14日,保藏名称:Magnaporthe oryzae K2e12;保藏号:CCTCC NO:M 2016107。
引物序列:
K2e12innerF:5’-CACACTGCTAAATGCCGAGA-3’
K2e12innerR:5’-GCACAGTGTTGCTGTGGACT-3’
tubulinF:5’-AGCCCTACAACGCTACCC-3’
tubulinR:5’-CGAGCACAGACAGCGAGT-3’
K2e12longF:5’-GAACAAGTGGCGTACAGACG-3’
K2e12LbarR:5’-GTCCTCGTTCCTGTCTGCTAAT-3。
实施例3、稻瘟病菌突变体菌株K2e12的形态观察:
在CM培养基上观察该突变体菌株的菌落形态、颜色、生长速率和产孢量。
观察结果(图2)显示:突变体菌株K2e12在CM培养基上25℃光/暗培养7d后,与野生型菌株相比,气生菌丝稀疏,菌落深灰色(图2A)。突变体菌株K2e12在液体CM培养基中培养,与野生型菌株类似,菌丝蓬松,液体呈黄绿色(图2B)。突变体K2e12与野生型菌株Guy11产孢能力有显著差异,突变体K2e12几乎不产生分生孢子,而野生型菌株Guy11产生大量的分生孢子(图2C)。
备注说明:光/暗培养为12小时光培养(光线强度3000lux),12小时暗培养,以下同。
实施例4、稻瘟病菌突变体菌株K2e12对大麦叶片的侵染致病性丧失:
大麦(ZJ-8)光/暗育苗6d,剪取表面无损的第一片叶,用于接种试验。25℃,在CM固体培养基上光/暗培养稻瘟病菌野生型Guy11及突变体菌株K2e12,7天后用0.5cm口径的打孔器打取菌饼,将菌饼接种于离体的大麦叶片上,每片叶3个菌饼,3个重复,25℃,12/12h光/暗保湿培养,4d后观察并记录发病情况,试验重复3次。
观察结果(图3)显示:稻瘟病菌突变体菌株K2e12对大麦叶片致病性丧失,没有可见病斑;相比之下,野生型菌株在接种处产生面积较大的扩散性黑色病斑,而且病斑会快速沿着叶脉扩散至周围的健康区域,病斑中央区域逐渐腐烂,并存在着大量的菌丝。
实施例5、稻瘟病菌突变体菌株K2e12对水稻的致病性丧失:
水稻(CO-39)光/暗育苗14d;CM平板培养10天的稻瘟病菌,水洗孢子并用三层灭菌擦镜纸过滤,用0.2%明胶稀释孢子悬浮液至浓度1×105个/ml;用小型喷雾器将2-3ml孢子液均匀喷布于水稻苗上,每盆水稻用等量孢子液,每处理3个重复;22℃,黑暗保湿48h,然后将接种苗置于26℃光照条件下培养;5-7天后,观察并记录发病情况,实验重复3次。
观察结果(图4)显示:稻瘟病菌突变体菌株K2e12对水稻丧失致病性,没有产生任何病斑;相比之下,野生型菌株在水稻上产生典型的梭形病斑。
实施例6、利用MoK2e12蛋白的表达或修饰作为靶标,筛选抗真菌的药物:
把受MoK2e12基因调控的下游基因的启动子与荧光蛋白GFP构建成融合蛋白载体,通过实施例2的方法转入到稻瘟病菌中,然后把在下游基因的启动子调控下的GFP表达的稻瘟病菌转化子菌株在液体完全培养基中大量培养,收集菌丝后分成若干小份,每一份中加入待筛选的化合物或候选药物继续在完全培养基培养数小时至数天,取菌丝在荧光显微镜下观察菌丝的GFP的绿色荧光表达情况。如果MoK2e12蛋白被候选药物抑制而失去活性,则失去对下游基因的启动子表达的调控能力,GFP蛋白不表达从而菌丝失去绿色荧光,说明该侯选药物具有抑制MoK2e12蛋白的作用。获得的具有抑制MoK2e12蛋白功能的候选药物再利用实施例3的方法,在接种的孢子液中加入侯选药物,测定野生型稻瘟病菌在这种侯选药物存在的条件下对水稻的致病性有没有减弱,进一步确定化合物的抗真菌效果。当测定结果为野生型稻瘟病菌株对水稻的致病性减弱时,说明候选药物具有明显的抗真菌效果,可以作为抗真菌药物的来源;当测定结果为野生型稻瘟病菌株对水稻的致病性没有变化或增强时,说明侯选药物没有抗真菌效果,不能用作抗真菌药物。
综上所述,稻瘟病菌突变体K2e12由于满足缺失了致病基因,通过设计和筛选能够破坏该基因的表达、剪切及其编码蛋白的表达的化合物,或设计和筛选能对该蛋白的氨基酸序列进行修饰的化合物,从而开发出新的抗真菌药物,或该突变体可以作为生防菌株在防治稻瘟病菌上进行开发应用,因此,其基因有望作为药物靶标,其菌株有望作为生防菌剂。
最后,还需要注意的是,以上列举的仅是本发明的若干个具体实施例。显然,本发明不限于以上实施例,还可以有许多变形。本领域的普通技术人员能从本发明公开的内容直接导出或联想到的所有变形,均应认为是本发明的保护范围。
<110> 浙江大学
<120> 稻瘟病菌的突变体菌株K2e12及其用途
<160> 6
<210> 1
<211> 43
<212> DNA
<213>人工序列
<220>
<223> 引物K2e12upF
<400> 1
ggtacccggg gatcctctag accgtaagtt cagggtcatc atc 43
<210> 2
<211> 38
<212> DNA
<213> 人工序列
<220>
<223> 引物K2e12upR
<400> 2
ctccttcaat atcatcttct agatgcggcg atgtctcc 38
<210> 3
<211> 38
<212> DNA
<213> 人工序列
<220>
<223> 引物K2e12downF
<400> 3
tgcccgtcac cgagatttag tcgtctgcgg agtctgtc 38
<210> 4
<211> 42
<212> DNA
<213> 人工序列
<220>
<223> 引物K2e12downR
<400> 4
acgacggcca gtgccaagct tatgaagagt tggtgggatg ac 42
<210> 5
<211> 20
<212> DNA
<213> 人工序列
<220>
<223> 引物BarF
<400> 5
agaagatgat attgaaggag 20
<210> 6
<211> 20
<212> DN A
<213> 人工序列
<220>
<223> 引物BarR
<400> 6
ctaaatctcg gtgacgggca 20
- 还没有人留言评论。精彩留言会获得点赞!