低饱和脂肪向日葵及相关方法与流程
本申请是2008年12月19日提交的申请号为200880127223.7(pct申请号为pct/us2008/087827)、发明名称为“低饱和脂肪向日葵及相关方法”的发明专利申请的分案申请。
优先权要求
本申请要求2007年12月20日提交的美国临时专利申请流水号61/015,591的申请日的权益。
本发明涉及新的且独特的向日葵以及相关方法,所述向日葵产生低饱和脂肪和任选地高亚油酸的种子。本发明进一步涉及具有草甘膦(glyphosate)抗性的非遗传修饰的、非诱变的向日葵及相关方法。
发明背景
栽培的向日葵(helianthusannuusl.)是植物油的全球主要来源。在美国,每年种植约四百万英亩向日葵,主要在达科他州(dakotas)和明尼苏达州(minnesota)。
在过去十年里美国的向日葵种植面积非常快速的扩张部分是由于向日葵育种和品种改良领域中的数项重要的发展。一个重大的发展是胞质雄性不育和育性恢复基因的发现,即一项容许产生杂种向日葵的发现。在上世纪70年代早期期间引入了如此产生的杂种。
fick,“breedingandgenetics,”于sunflowerscienceandtechnology279-338(j.f.carter编1978)提出了向日葵中胞质雄性不育(cms)和遗传育性恢复的描述。
向日葵油主要包含棕榈酸(16:0)、硬脂酸(18:0)、油酸(18:1)、亚油酸(18:2)和亚麻酸(18:3)。虽然植物中存在其它不常见的脂肪酸,但是棕榈酸、硬脂酸、油酸、亚油酸、和亚麻酸构成植物油的世界产量中存在的脂肪酸的约88%。(j.l.harwood,plantacyllipids:structure,distributionandanalysis,4lipids:structureandfunction,p.k.stumpf和e.e.conn编(1988)。)棕榈酸和硬脂酸是饱和脂肪酸,在某些研究中已经证明了它们促成血浆胆固醇水平升高,这种升高是冠心病(coronaryheartdisease)的一项因素。根据最近的研究,不饱和脂肪酸诸如油酸和亚油酸高的植物油可具有降低血浆胆固醇的能力。一般而言,饱和脂肪酸还具有比碳数目相同的不饱和脂肪酸高的熔点,这促成食品中的低温耐受性问题,而且能促成摄入过程中的蜡质(waxy)或油腻口感(feelinthemouth)。还知道的是,由具有小于约3%饱和脂肪酸的脂肪和油制成的食物产品通常每客(serving)会含有小于0.5克的饱和脂肪,并且因此可以在目前的标签规定下标记为含有“零饱和脂肪”。如此,就许多原因而言,生产具有低水平棕榈酸和硬脂酸且有高水平油酸或亚油酸的向日葵油是理想的。
任何新的、理想的植物种质的开发都有许多步骤。植物育种开始于分析和定义当前种质的问题和弱点、建立项目目的、和定义具体的育种目标。下一步是选择拥有满足项目目的的性状的种质。目的是在单一品种中组合来自亲本种质的期望性状的改良组合。这些重要的性状可以包括较高的种子产率(yield)、对疾病和昆虫的抗性、较好的茎和根、对干旱和热的耐受性、和较好的农艺学质量。
育种或选择方法的选择取决于植物繁殖的模式、所改良性状的遗传力、和商业上使用的栽培种类型(例如,f1杂种栽培种、纯系栽培种等)。对于高度可遗传的性状,选择在单个位置评估的卓越植物个体会是有效的,而对于具有低遗传力的性状,选择应当基于自重复评估相关植物的家族所获得的平均值。流行的选择方法通常包括谱系选择、改良的谱系选择、混合选择(massselection)、和轮回选择。
遗传的复杂性影响育种方法的选择。使用回交育种来将一种或少数几种对于高度可遗传的性状有利的基因转移入想要的栽培种中。此办法已经广泛地用于育种疾病抗性栽培种。使用多种轮回选择技术来改善受多个基因控制的数量遗传性状。轮回选择在自花传粉作物中的使用依赖于传粉容易、来自每次传粉的成功杂种的频率、和来自每次成功杂交的杂种后代的数目。
每个育种项目应当包括对育种方法效率的周期性客观评估。评估标准随着目的和目标而变化,但是应当包括基于与合适的标准品比较得到的每年来自选择的收益(gain)、高级育种系(advancedbreedingline)的总价值、和每单位投入(例如每年、花费的每一笔美元等)产生的成功栽培种的数目。
对有希望的高级育种系进行彻底地测试,并与代表商业目标区的环境中的合适标准品进行比较达三年或更多年。最好的品系是新商业栽培种的候选;那些在少数几个性状方面仍有缺陷的品系可以作为亲本使用以产生供进一步选择用的新种群。
这些过程(它们通向上市(marketing)和销售(distribution)的最终步骤)从进行第一次杂交的时间起通常花费8至12年。因此,新栽培种的开发是耗时的过程,其需要精确的预先计划、资源的有效使用、和最低限度的方向变化。
最困难的任务是鉴定遗传上卓越的个体,因为对于大多数性状,真实基因型的值被其它混杂的植物性状或环境因素掩盖。一种鉴定卓越植物的方法是相对于其它实验植物和广泛种植的标准栽培种观察其性能(performance)。如果单次观察没有结论,则重复观察可提供对其遗传价值的更好评估。
植物育种的目的是开发新的、独特的且卓越的向日葵栽培种和杂种。育种人员最初选择并杂交两种或更多种亲本品系,接着进行重复的自交和选择,产生许多新的遗传组合。理论上育种人员可以经由杂交、自交和突变产生几十亿不同的遗传组合。育种人员在细胞水平上没有直接对照(nodirectcontrol)。因此,两名育种人员绝不会开发出具有相同向日葵性状的相同品系,或者甚至非常类似的品系。
每年,植物育种人员选择种质来推进到下一世代。这种种质在独特的且不同的地理学、气候和土壤条件下种植,然后在生长期期间和结束时进行进一步选择。开发的栽培种是不可预测的。这种不可预测性是由于育种人员的选择,其发生在独特的环境中,在dna水平上没有对照(使用常规的育种方法),而且产生几百万不同的可能的遗传组合。具有本领域普通技术的育种人员不能预测他开发的最终所得品系,只是可能以非常粗略且大体的方式。同一育种人员不能通过使用完全相同的初始亲本和相同的选择技术来产生相同的栽培种两次。这种不可预测性导致花费大量的研究资金来开发卓越的新型向日葵栽培种。
新的向日葵栽培种的开发要求开发和选择向日葵品种、杂交这些品种、和选择卓越的杂种杂交。通过在选定的雄性能育亲本之间手工杂交或者通过使用雄性不育系统来产生杂种种子。针对某些单基因性状来选择这些杂种,诸如荚果颜色(podcolor)、花的颜色、短柔毛(pubescence)颜色、或除草剂抗性,它们指明种子确实是杂种。关于亲本品系,以及杂种表型的别的数据影响育种人员决定是否继续特定的杂种杂交。
使用谱系育种和轮回选择育种方法来从育种群体开发栽培种。育种项目将来自两种或更多种栽培种或多种运用广泛的(broad-based)来源的期望性状组合成育种集合(breedingpool),通过自交和选择期望的表型从所述育种集合开发栽培种。评估新的栽培种以确定哪些具有商业潜力。
谱系育种通常用于改良自花传粉的作物。将两种拥有有利的、互补性状的亲本杂交以产生f1。通过使一个或数个f1自交来产生f2群体。最好个体的选择可以在f2群体中开始;然后在f3中开始选择最好家族中的最好个体。对家族的重复测试可以在f4代中开始以改善选择具有低遗传力的性状的效率。在育种的高级阶段(即f6和f7),对最好的品系或表型上类似的品系的混合物测试作为新栽培种的推广(release)潜力。
可以使用混合和轮回选择来改善自花传粉或异花传粉作物的群体。通过使数种不同亲本互交来鉴定或创建杂合个体的遗传可变群体。基于个体优越性、杰出的子代、或卓越的组合能力来选择最好的植物。使选定的植物互交以产生新群体,在其中继续进一步的选择循环。
回交育种已经用于将简单遗传的、高度可遗传的性状的基因转移到作为回归亲本的期望的纯合栽培种或近交系中。要转移的性状来源称作供体亲本。预期所得的植物具有回归亲本(例如栽培种)的属性和自供体亲本转移的期望性状。初始杂交后,选择拥有供体亲本表型的个体,并与回归亲本重复杂交(回交)。预期所得的植物具有回归亲本(例如栽培种)的属性和自供体亲本转移的期望性状。
严格意义上的单种子遗传(single-seeddescent)方法指种植分离群,每株植物收获一粒种子样品,并使用这一粒种子样品来种植下一世代。当群体已经从f2推进至期望的育种水平时,衍生出品系的植物会各自上溯至不同的f2个体。群体中植物的数目每代都下降,这是由于一些种子不能萌发或者一些植物不能产生哪怕一粒种子。因此,当完成世代进度时,并不是所有最初在群体中取样的f2植物都会由子代代表。
在多种子方法中,向日葵育种人员通常从群体中的每株植物收获种子,并使它们脱粒在一起以形成整体(bulk)。使用整体的一部分来种植下一世代,而一部分进行保存。该方法已经被称为改良的单种子遗传。
已经使用多种子方法来在收获时节约劳动力。用机器取下种子比单种子方法中用手自每株取下一粒种子快得多。多种子方法还使得有可能每个近交世代种植相同数目的群体种子。收获足够的种子来补偿那些不萌发或不产生种子的植物。
关于通常用于不同性状和作物的其它育种方法的描述可以参见数本参考书之一(例如allard,1960;simmonds,1979;sneep等,1979;fehr,1987)。
合适的测试应当检测任何主要的不足,并建立高于当前栽培种的优越性或改进水平。除显示卓越的性能外,必须需要与工业标准相容或者产生新市场的新栽培种。新栽培种的引入可能由于专门的广告和行销、改变的种子和商业生产实践、和新产品利用而使种子生产者、种植者、加工者和消费者承受额外的成本。推广新栽培种前的测试应当考虑研究和开发成本以及最终栽培种的技术优越性。对于种子繁殖的栽培种,容易且经济地产生种子必须是可行的。
向日葵是一种重要且有价值的大田作物。如此,植物育种人员的持续目的是开发稳定的、高产率的农艺学上是可靠的(sound)向日葵栽培种。当前的目的是使所用土地上生产的谷粒量最大化,并为动物和人类供应食物。为了实现此目的,向日葵育种人员必须选择和开发具有产生卓越栽培种的性状的向日葵植物。
相关领域的上述例子及其相关限制意图是例示性的而非排他性的。在阅读说明书后,相关领域的其它限制对于本领域技术人员会是显而易见的。
发明概述
结合系统、工具和方法来描述以下实施方案,所述系统、工具和方法意欲为例示性和说明性的,而并不限制范围。在多个实施方案中,一个或多个上述问题已经得到减少或消除,而其它实施方案涉及其它改进。
根据本发明,提供了一种新的向日葵植物,其产生具有低饱和脂肪含量的种子。本发明部分涉及具有低饱和脂肪含量的向日葵种子、产生具有低饱和脂肪含量的种子的向日葵植物的植株或植株部分、及用于产生通过将产生具有低饱和脂肪含量的种子的向日葵植物与其自身或另一向日葵栽培种杂交来产生的向日葵植物的方法和通过诱变或转化产生具有低饱和脂肪含量的种子的向日葵植物来创建变体的方法。
本发明的各方面提供了新的向日葵植物,其产生具有低饱和脂肪含量和高亚油酸含量的种子。本发明部分涉及具有低饱和脂肪含量和高亚油酸含量的向日葵种子、产生具有低饱和脂肪含量和高亚油酸含量的种子的向日葵植物的植株或植株部分、及用于产生通过将产生具有低饱和脂肪含量和高亚油酸含量的种子的向日葵植物与其自身或另一向日葵栽培种杂交来产生的向日葵植物的方法和通过诱变或转化产生具有低饱和脂肪含量和高亚油酸含量的种子的向日葵植物来创建变体的方法。
具有低饱和脂肪含量的种子的例子包括但不限于如下的种子,其具有总计约2.8%或更少、约2.9%或更少、约3%或更少、约3.1%或更少、约3.2%或更少、或约3.3%或更少的棕榈酸(16:0)和硬脂酸(18:0)含量。
具有低饱和脂肪含量和高亚油酸(18:2)含量的种子的例子包括但不限于如下的种子,其具有总计约4.1%或更少、约5%或更少、约6%或更少、约7%或更少、约8%或更少、约9%或更少、约10%或更少、约11%或更少、或约12%或更少的棕榈酸(16:0)和硬脂酸(18:0)含量,而且具有约15%、约20%、约25%、约30%、约35%、约40%、约45%、约50%、约55%、约60%、约65%、约70%、或约74%或更多的亚油酸(18:2)。
如此,使用产生具有低饱和脂肪和任选地高亚油酸含量的种子的向日葵植物进行的任何此类方法是本发明的一部分(例如自交、回交、杂种生成、与群体杂交等)。使用产生具有低饱和脂肪和任选地高亚油酸含量的种子的向日葵植物作为亲本产生的所有植物在本发明的范围内。有利地,可以在与其它不同向日葵植物的杂交中使用所述向日葵植物以产生具有卓越特征的第一代(f1)向日葵杂种种子和植物。
在另一方面,本发明提供了单基因或多基因转变的向日葵植物,其产生具有低饱和脂肪和任选地高亚油酸含量的种子。优选地,转移的基因可以是显性或隐性等位基因。转移的基因可以赋予性状,诸如除草剂抗性、抗虫性(insectresistance)、对细菌、真菌或病毒疾病的抗性、雄性能育、雄性不育、提高的营养质量、和工业用途。基因可以是天然存在的向日葵基因或经由基因工程技术引入的转基因。
在另一方面,本发明提供了可再生细胞,其用于组织培养产生具有低饱和脂肪和任选地高亚油酸含量的种子的向日葵植物。组织培养能够再生具有上述产生具有低饱和脂肪和任选地高亚油酸含量之种子的向日葵植物的生理学和形态学特征的植物,而且能够再生与上述向日葵植物具有基本上相同基因型的植物。此类组织培养物中的可再生细胞可以是胚、原生质体、分生细胞、愈伤组织、花粉、叶、花药、根、根尖、花、种子、荚果或茎。更进一步,本发明提供了自本发明的组织培养物再生的向日葵植物。
在另一方面,本发明提供了一种将期望的性状引入产生具有低饱和脂肪和任选地高亚油酸含量的种子的向日葵植物中的方法,其中所述方法包括:将产生具有低饱和脂肪和任选地高亚油酸含量的种子的向日葵植物与包含期望性状的另一向日葵栽培种的植物杂交以产生f1子代植物,其中所述期望的性状选自下组:雄性不育、除草剂抗性、抗虫性、和对细菌疾病、真菌疾病或病毒疾病的抗性;选择一株或多株具有期望性状的子代植物以产生选定的子代植物;将所述选定的子代植物与产生具有低饱和脂肪和任选地高亚油酸含量的种子的向日葵植物杂交以产生回交子代植物;选择具有所述期望的性状和产生具有低饱和脂肪和任选地高亚油酸含量之种子的向日葵植物的生理学和形态学特征的回交子代植物,以产生选定的回交子代植物;并重复这些步骤来产生选定的第一或更高(firstorhigher)的回交子代植物,其包含所述期望的性状和产生具有低饱和脂肪和任选地高亚油酸含量之种子的向日葵植物的所有生理学和形态学特征。
除了上文所描述的例示性方面和实施方案外,通过研究以下描述,另外的方面和实施方案会变得显而易见。
具体地,本申请提供下列各项:
1.向日葵栽培种的种子,其中所述种子中的油的脂肪酸组成包含总计约3.3%或更少的棕榈酸(16:0)和硬脂酸(18:0)。
2.根据项1的向日葵栽培种的种子,其中所述种子的含油量包含共计大约或小于3%的棕榈酸(16:0)和硬脂酸(18:0)。
3.向日葵植物或其部分,其是通过种植项1的种子来产生的。
4.用于产生f1杂种向日葵种子的方法,其中所述方法包括将项3的植物与不同向日葵植物杂交,并收获所得的f1杂种向日葵种子。
5.通过项4的方法产生的杂种向日葵种子。
6.将期望的性状引入根据项3的向日葵植物中的方法,其中所述方法包括:(a)将根据项3的向日葵植物与包含期望性状的另一向日葵栽培种的植物杂交以产生f1子代植物,其中所述期望的性状选自下组:雄性不育、除草剂抗性、抗虫性、和对细菌疾病、真菌疾病或病毒疾病的抗性;(b)选择一株或多株具有所述期望性状的子代植物以产生选定的子代植物;(c)将所述选定的子代植物与根据项3的向日葵植物杂交以产生回交子代植物;(d)选择具有所述期望的性状并且能够产生具有如下含油量的种子的回交子代植物,即包含总计约3.3%或更少的棕榈酸(16:0)和硬脂酸(18:0)含量,以产生回交子代植物以产生选定的回交子代植物;并(e)重复步骤(c)和(d)三次或更多次以产生选定的第四或更高的回交子代植物,其包含所述期望的性状,并且能够产生具有如下含油量的种子,即包含总计约3.3%或更少的棕榈酸(16:0)和硬脂酸(18:0)含量。
7.项6的方法,其中根据项3的向日葵植物包含总计大约或小于2.8%的棕榈酸(16:0)和硬脂酸(18:0)含量。
8.在种子油方面稳定化的良种向日葵栽培种,所述种子油包含总计约3.3%或更少的棕榈酸(16:0)和硬脂酸(18:0)含量。
9.根据项8的在种子油方面稳定化的良种向日葵栽培种,其中所述种子油包含总计大约或小于3%的棕榈酸(16:0)和硬脂酸(18:0)含量。
10.项8的良种向日葵栽培种,其中所述期望的性状是除草剂抗性,而且赋予对选自下组的除草剂的抗性:咪唑啉酮、磺酰脲、草甘膦、草铵膦、l-草铵膦、三嗪、clearfield、麦草畏、2,4-d、和苯基氰。
11.至少一种良种向日葵植物的种子,所述种子在种子油方面是稳定化的,所述种子油包含总计约3.3%或更少的棕榈酸(16:0)和硬脂酸(18:0)含量。
12.根据项11的种子,其中所述种子油包含总计大约或小于3%的棕榈酸(16:0)和硬脂酸(18:0)含量。
13.自良种向日葵品种的向日葵种子提取的向日葵油的整装样品,所述向日葵油具有如下的脂肪酸谱,其包含总计约3.3%或更少的棕榈酸(16:0)和硬脂酸(18:0)含量,所述脂肪酸谱在所述良种向日葵品种的种子油中是稳定化的。
14.根据项19的自良种向日葵品种的向日葵种子提取的向日葵油的整装样品,其中所述向日葵油的脂肪酸谱包含总计大约或小于3%的棕榈酸(16:0)和硬脂酸(18:0)含量。
15.来自良种向日葵品种种子的油,所述油具有如下的脂肪酸谱,其包含总计约3.3%或更少的棕榈酸(16:0)和硬脂酸(18:0)含量。
16.根据项15的来自良种向日葵品种种子的油,其中未氢化油的脂肪酸谱包含总计大约或小于3%的棕榈酸(16:0)和硬脂酸(18:0)含量。
17.向日葵栽培种的种子中的油,所述油具有如下的脂肪酸含量,其包含总计3.3%或更少的棕榈酸(16:0)和硬脂酸(18:0)。
18.项17的油,其用于食品应用。
19.食物产品,其包含项17的油。
20.植物油组合物,其包含项17的油。
21.向日葵栽培种的种子,其中所述种子的含油量包含总计约12%或更少的棕榈酸(16:0)和硬脂酸(18:0)含量和约15%或更多的亚油酸(18:2)含量。
22.根据项21的向日葵栽培种的种子,其中总计棕榈酸(16:0)和硬脂酸(18:0)含量小于约11%,并且其中总亚油酸(18:2)含量大于约20%。
23.根据项21的向日葵栽培种的种子,其中总计棕榈酸(16:0)和硬脂酸(18:0)含量小于约4.1%,并且其中总亚油酸(18:2)含量大于约74%。
24.向日葵植物或其部分,其是通过种植项21的种子来产生的。
25.用于产生f1杂种向日葵种子的方法,其中所述方法包括将项23的植物与不同向日葵植物杂交,并收获所得的f1杂种向日葵种子。
26.通过项25的方法产生的杂种向日葵种子。
27.将期望的性状引入根据项24的向日葵植物中的方法,其中所述方法包括:(a)将根据项24的向日葵植物与包含期望性状的另一向日葵栽培种的植物杂交以产生f1子代植物,其中所述期望的性状选自下组:雄性不育、除草剂抗性、抗虫性、和对细菌疾病、真菌疾病或病毒疾病的抗性;(b)选择一株或多株具有所述期望性状的子代植物以产生选定的子代植物;(c)将所述选定的子代植物与根据项24的向日葵植物杂交以产生回交子代植物;(d)选择具有所述期望的性状并且能够产生具有如下含油量的种子的回交子代植物,即包含总计约3.3%或更少的棕榈酸(16:0)和硬脂酸(18:0)含量,以产生回交子代植物以产生选定的回交子代植物;并(e)重复步骤(c)和(d)三次或更多次以产生选定的第四或更高的回交子代植物,其包含所述期望的性状,并且能够产生具有如下含油量的种子,即包含总计约12%或更少的棕榈酸(16:0)和硬脂酸(18:0)含量和约15%或更多的亚油酸(18:2)含量。
28.在种子油方面稳定化的良种向日葵植物,其包含总计约12%或更少的棕榈酸(16:0)和硬脂酸(18:0)含量和约15%或更多的亚油酸(18:2)含量。
29.根据项28的在种子油方面稳定化的良种向日葵植物,其中所述种子油包含总计大约或小于11%的棕榈酸(16:0)和硬脂酸(18:0)含量和大约或大于20%的总亚油酸(18:2)含量。
30.至少一种良种向日葵植物的种子,所述种子在种子油方面是稳定化的,所述种子油包含总计约12%或更少的棕榈酸(16:0)和硬脂酸(18:0)含量和约15%或更多的亚油酸(18:2)含量。
31.根据项30的至少一种良种向日葵植物的种子,其中所述种子在种子油方面是稳定化的,所述种子油包含总计大约或小于11%的棕榈酸(16:0)和硬脂酸(18:0)含量和大约或大于20%的总亚油酸(18:2)含量。
32.自良种向日葵栽培种的向日葵种子提取的向日葵油的整装样品,所述向日葵油具有如下的脂肪酸谱,所述脂肪酸谱包含总计约12%或更少的棕榈酸(16:0)和硬脂酸(18:0)含量和约15%或更多的亚油酸(18:2)含量,所述脂肪酸谱在所述良种向日葵栽培种的种子油中是稳定化的。
33.根据项32的自良种向日葵栽培种的向日葵种子提取的向日葵油的整装样品,其中所述向日葵油的脂肪酸谱包含总计大约或小于11%的棕榈酸(16:0)和硬脂酸(18:0)含量和大约或大于20%的总亚油酸(18:2)含量。
34.自良种向日葵栽培种的种子提取的油,所述油具有如下的脂肪酸谱,其包含总计约12%或更少的棕榈酸(16:0)和硬脂酸(18:0)含量和约15%或更多的亚油酸(18:2)含量,所述脂肪酸谱在所述良种向日葵栽培种的种子油中是稳定化的。
35.根据项34的来自良种向日葵栽培种的种子的油,其中所述油的脂肪酸谱包含总计大约或小于11%的棕榈酸(16:0)和硬脂酸(18:0)含量和大约或大于20%的总亚油酸(18:2)含量。
36.向日葵栽培种的种子中的油,所述油具有如下的脂肪酸含量,其包含总计约12%或更少的棕榈酸(16:0)和硬脂酸(18:0)含量和约15%或更多的亚油酸(18:2)含量。
37.项36的油,其用于食品应用。
38.食物产品,其包含项36的油。
39.植物油组合物,其包含项36的油。
附图简述
图1显示了低硬脂酸qtl在lg17的ha1875-ha1865区间中的精细定位(小图a:具有新标志物(为蓝色)的lg17的图和小图b:低硬脂酸qtl相对于ha1875-ors565区间的精细定位);
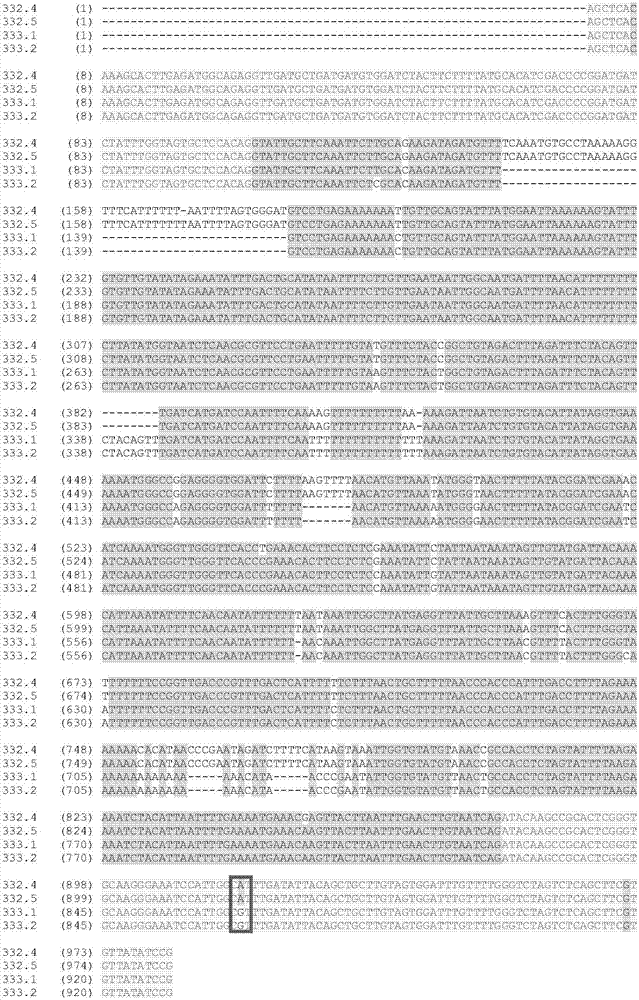
图2显示了来自两个亲本品系的kasii-2基因的序列比对,仅一个snp(以红色方框突出显示)位于外显子中,所有其它多态性都位于内含子中。显示了snp和indel(id号333.1(seqidno:38)和333.2(seqidno:39)代表来自ond163r扩增子的克隆,而332.4(seqidno:40)和332.5(seqidno:41)代表来自h280r[1]/687r-1-8-1扩增子的克隆);
图3显示了低棕榈酸qtl(小图a)和脂肪酸基因kasiii-2(小图b)在lg5上的共定位。
发明的实施方式
在以下描述和表格中,使用了许多术语。为了提供对说明书和权利要求书(包括此类术语要给予的范围)的清楚而一致的理解,提供了下列定义:
等位基因。等位基因指基因的一种或多种可选形式之任一种,所有该等位基因涉及一种性状或特征。在双倍体细胞或生物体中,给定基因的两个等位基因占据一对同源染色体上的相应基因座。
回交。回交指育种人员重复地将杂种子代往回与亲本之一(例如第一代杂种f1与f1杂种的亲本基因型之一)杂交的过程。
良种向日葵。已经在某些商业上重要的农艺学性状方面稳定化的向日葵栽培种,其相对于在同一生长位置、同时并且在相同条件下生长的验证品种的产率包含约100%或更大的稳定化产率。在一个实施方案中,“良种向日葵”意指在某些商业上重要的农艺学性状方面稳定化的向日葵栽培种,其相对于在同一生长位置、同时并且在相同条件下生长的验证品种的产率包含110%或更大的稳定化产率。在另一个实施方案中,“良种向日葵”意指在某些商业上重要的农艺学性状方面稳定化的向日葵栽培种,其相对于在同一生长位置、同时并且在相同条件下生长的验证品种的产率包含115%或更大的稳定化产率。
胚。胚指成熟种子内含有的小植株。
fame分析。脂肪酸甲酯分析是容许精确量化构成复合脂种类的脂肪酸的方法。
咪唑啉酮(imidazolinone)抗性(imi)。抗性和/或耐受性是由一种或多种改变乙酰乳酸合酶(als)(又称为乙酰羟酸合酶(ahas))的基因赋予的,所述基因容许所述酶对抗咪唑啉酮的作用。
诱变。诱变指用诱变剂(例如提高突变在靶标植物或植物部分中的频率的化学剂或物理因素)诱变植物或植物部分。作为非限制性例子,可以使用konzak的双重化学诱变技术(如记载于美国专利号6,696,294)来在内源植物基因中诱导突变等位基因。
含油量。含油量按照占完整干燥种子的百分比来测量,而且是不同品种的特征。它可以使用多种分析技术诸如nmr、nir、和soxhlet提取来测定。
总脂肪酸百分比。这通过如下测定,即从种子提取油样品,生成所述油样品中存在的脂肪酸的甲酯,并使用气相层析来分析各种脂肪酸在样品中的比例。脂肪酸组成也可以是品种的区别性特征。
单基因转变的(转变)。单基因转变的(转变)植物指通过称作回交的植物育种技术,或者经由基因工程开发的植物,其中除了经由回交技术或经由基因工程转移到品种中的单基因外,该品种的基本上所有期望的形态学和生理学特征也均得到复原。
稳定化的。从一个世代能通过繁殖传递到同一品种的近交植物的下一世代。
总饱和的(totsat)。油中的饱和脂肪(包括c12:0、c14:0、c16:0、c18:0、c20:0、c22:0和c24.0)占种子油的总百分比。
根据本发明的具体实施方案,提供了一种新的向日葵植物,其产生具有低饱和脂肪含量的种子。此实施方案涉及具有低饱和脂肪含量的向日葵种子、产生具有低饱和脂肪含量的种子的向日葵植物的植株或植株部分、及用于生产通过将产生具有低饱和脂肪含量的种子的向日葵植物与其自身或另一向日葵栽培种杂交而产生的向日葵植物的方法和通过诱变或转化产生具有低饱和脂肪含量的种子的向日葵植物来创建变体的方法。
本发明的其它方面提供了新的向日葵植物,其产生具有低饱和脂肪含量和高亚油酸含量的种子。此实施方案涉及具有低饱和脂肪含量和高亚油酸含量的向日葵种子、产生具有低饱和脂肪含量和高亚油酸含量的种子的向日葵植物的植株或植株部分、及用于生产通过将产生具有低饱和脂肪含量和高亚油酸含量的种子的向日葵植物与其自身或另一向日葵栽培种杂交而产生的向日葵植物的方法和通过诱变或转化产生具有低饱和脂肪含量和高亚油酸含量的种子的向日葵植物来创建变体的方法。
具有低饱和脂肪含量的种子的例子包括但不限于如下的种子,其具有总计约2.8%或更少、约2.9%或更少、约3%或更少、约3.1%或更少、约3.2%或更少、或约3.3%或更少的棕榈酸(16:0)和硬脂酸(18:0)含量。
具有低饱和脂肪含量和高亚油酸(18:2)含量的种子的例子包括但不限于如下的种子,其具有总计约6%或更少、约4.1%或更少、约5%或更少、约6%或更少、约7%或更少、约8%或更少、约9%或更少、约10%或更少、约11%或更少、或约12%或更少的棕榈酸(16:0)和硬脂酸(18:0)含量,而且具有约15%、约20%、约25%、约30%、约35%、约40%、约45%、约50%、约55%、约60%、约65%、约70%、或约74%或更多的亚油酸(18:2)。
如此,使用产生具有低饱和脂肪和任选地高亚油酸含量的种子的向日葵植物进行的任何此类方法是本发明的一部分(例如自交、回交、杂种生成、与群体杂交等)。使用产生具有低饱和脂肪和任选地高亚油酸含量的种子的向日葵植物作为亲本产生的所有植物在本发明的范围内。有利地,可以在与其它不同向日葵植物的杂交中使用所述向日葵植物以产生具有卓越特征的第一代(f1)向日葵杂种种子和植物。
在另一方面,本发明提供了单基因或多基因转变的向日葵植物,其产生具有低饱和脂肪和任选地高亚油酸含量的种子。优选地,转移的基因可以是显性或隐性等位基因。优选地,转移的基因会赋予性状,诸如除草剂抗性、抗虫性、细菌抗性、真菌抗性、病毒疾病抗性、雄性能育、雄性不育、提高的营养质量、和工业用途。基因可以是天然存在的向日葵基因或经由基因工程技术引入的转基因。
在另一方面,本发明提供了可再生细胞,其用于组织培养产生具有低饱和脂肪和任选地高亚油酸含量的种子的向日葵植物。优选地,组织培养会能够再生具有上述产生具有低饱和脂肪和任选地高亚油酸含量之种子向日葵植物的生理学和形态学特征的植物,而且能够再生与上述向日葵植物具有基本上相同基因型的植物。此类组织培养物中的可再生细胞可以是胚、原生质体、分生细胞、愈伤组织、花粉、叶、花药、根、根尖、花、种子、荚果或茎。更进一步,本发明的实施方案提供了自本发明的组织培养物再生的向日葵植物。
在另一方面,本发明提供了一种将期望的性状引入产生具有低饱和脂肪和任选地高亚油酸含量的种子的向日葵植物中的方法,其中所述方法包括:将产生具有低饱和脂肪和任选地高亚油酸含量的种子的向日葵植物与包含期望性状的另一向日葵栽培种的植物杂交以产生f1子代植物,其中所述期望的性状选自下组:雄性不育、除草剂抗性、抗虫性、和对细菌疾病、真菌疾病或病毒疾病的抗性;选择一株或多株具有期望性状的子代植物以产生选定的子代植物;将所述选定的子代植物与产生具有低饱和脂肪和任选地高亚油酸含量的种子的向日葵植物杂交以产生回交子代植物;选择具有所述期望的性状和产生具有低饱和脂肪和任选地高亚油酸含量之种子的向日葵植物的生理学和形态学特征的回交子代植物,以产生选定的回交子代植物;并重复这些步骤来产生选定的第一或更高的回交子代植物,其包含所述期望的性状和产生具有低饱和脂肪和任选地高亚油酸含量的种子的向日葵植物的所有生理学和形态学特征。
有用的方法包括但不限于使用直接基因转移方法诸如微粒介导的投递、dna注射、电穿孔等导入植物组织中的表达载体。可以使用微粒介质投递用生物射弹装置或使用土壤杆菌属(agrobacterium)介导的转化将表达载体导入植物组织中。用本发明的原生质获得的转化体植物意图在本发明的范围内。
随着使分离和表征编码特定蛋白质产物的基因成为可能的分子生物学技术的出现,植物生物学领域的科学家对以下方面形成了强烈的兴趣,即工程化改造植物的基因组以含有外来基因,或者另外的或经修饰型式的天然或内源基因(可能由不同启动子驱动),并进行表达以便以特定的方式改变植物的性状。此类外来的另外的和/或经修饰的基因在本文中统称为“转基因”。在过去15至20年里,已经开发了数种用于产生转基因植物的方法,并且在具体的实施方案中,本发明也涉及要求保护的品种或栽培种的转化型式。
植物转化牵涉构建会在植物细胞中发挥功能的表达载体。此类载体包含包括在调节元件(例如启动子)控制下的或者与调节元件(例如启动子)可操作连接的基因的dna。表达载体可以含有一种或多种此类可操作连接的基因/调控元件组合。载体可以是质粒形式,而且可以单独或者与其它质粒联合使用,以如下文所描述的那样使用转化方法提供经转化的向日葵植物,从而将转基因掺入向日葵植物的遗传物质中。
供向日葵转化用的表达载体:标志物基因
表达载体包括至少一种与调节元件(例如启动子)可操作连接的遗传标志,其容许通过负选择(即抑制不含该选择标志基因的细胞生长)或者通过正向选择(即筛选由该遗传标志编码的产物)回收含有所述标志的经转化细胞。许多通常用于植物转化的选择标志基因是转化领域中公知的,包括例如编码在代谢上使可以作为抗生素或除草剂的选择性化学剂解毒的酶的基因,或者编码对抑制剂不敏感的改变了的靶物的基因。几种正向选择方法也是本领域中已知的。
一种通常用于植物转化的选择标志基因是在植物调节信号控制下的新霉素磷酸转移酶ii(nptii)基因,其赋予对卡那霉素的抗性。参见例如fraley等,proc.natl.acad.sci.u.s.a.,80:4803(1983)。另一种常用的选择标志基因是潮霉素磷酸转移酶基因,其赋予对抗生素潮霉素的抗性。参见例如vandenelzen等,plantmol.biol.,5:299(1985)。
赋予对抗生素抗性的细菌起源的另外的选择标志基因包括庆大霉素乙酰转移酶、链霉素磷酸转移酶、氨基糖苷-3’-腺嘌呤转移酶(adenyltransferase)和博来霉素抗性决定子。参见hayford等,plantphysiol.86:1216(1988);jones等,mol.gen.genet.,210:86(1987);svab等,plantmol.biol.14:197(1990);hille等,plantmol.biol.7:171(1986)。其它选择标志基因赋予对除草剂诸如草甘膦、草铵膦(glufosinate)或溴苯腈(bromoxynil)的抗性。参见comai等,nature317:741-744(1985);gordon-kamm等,plantcell2:603-618(1990);和stalker等,science242:419-423(1988)。
其它供植物转化用的选择标志基因不是细菌起源的。这些基因包括例如小鼠二氢叶酸还原酶、植物5-烯醇丙酮酰莽草酸(enolpyruvylshikimate)-3-磷酸合酶和植物乙酰乳酸合酶。参见eichholtz等,somaticcellmol.genet.13:67(1987);shah等,science233:478(1986);charest等,plantcellrep.8:643(1990)。
另一类供植物转化用的标志基因需要筛选据推测经转化的植物细胞,而不对经转化的细胞直接遗传选择对毒性物质诸如抗生素的抗性。这些基因特别可用于量化或显现基因在特定组织中表达的空间样式,并且通常称为报告基因,因为它们可以与基因或基因调节序列融合以调查基因表达。通常用于筛选据推测经转化的细胞的基因包括β-葡糖醛酸糖苷酶(gus)、β-半乳糖苷酶、萤光素酶和氯霉素乙酰转移酶。参见r.a.jefferson,plantmol.biol.rep.5:387(1987);teeri等,emboj.8:343(1989);koncz等,proc.natl.acad.sciu.s.a.84:131(1987);deblock等,emboj.3:1681(1984)。
最近,已经可获得用于显现gus活性的体内方法,其不需要破坏植物组织。molecularprobespublication2908,imagene,t.m.green,第1页-第4页(1993);和naleway等,j.cellbiol.115:151a(1991)。然而,还没有证明这些用于显现gus活性的体内方法对于回收经转化的细胞是有用的,这是由于低灵敏度、高荧光背景和与使用萤光素酶基因作为选择标志有关的限制。
新近,已经利用编码绿色荧光蛋白(gfp)的基因作为原核和真核细胞中基因表达的标志物。参见chalfie等,science263:802(1994)。gfp和gfp突变体可以作为可筛选标志物使用。
供向日葵转化用的表达载体:启动子
表达载体中包含的基因必须由包含调节元件(例如启动子)的核苷酸序列驱动。数种类型的启动子现在是转化领域中公知的,可以单独使用或者与启动子联合使用的其它调节元件也是如此。
如本文中所使用的,“启动子”包括提及如下的dna区域,其在转录起始的上游,而且牵涉识别和结合rna聚合酶和其它蛋白质以启动转录。“植物启动子”是能够在植物细胞中启动转录的启动子。在发育控制下的启动子的例子包括优先在某些组织诸如叶、根、种子、纤维、木质部导管、管胞、或厚壁组织中启动转录的启动子。此类启动子称为“组织优选性的”。仅在某些组织中启动转录的启动子称为“组织特异性的”。“细胞类型”特异性启动子主要在一种或多种器官中的某些细胞类型(例如根或叶中的维管细胞)中驱动表达。“诱导型”启动子是在环境控制下的启动子。可招致诱导型启动子转录的环境条件的例子包括缺氧条件或光的存在。组织特异性的、组织优选性的、细胞类型特异性的、和诱导型启动子构成“非组成性”启动子类。“组成性”启动子是在大多数环境条件下有活性的启动子。
a.诱导型启动子
将诱导型启动子与向日葵中表达的基因可操作连接。任选地,将诱导型启动子与编码信号序列的核苷酸序列可操作连接,所述核苷酸序列与向日葵中表达的基因可操作连接。通过诱导型启动子,转录速率响应诱导剂而升高。
可以在本发明中使用任何诱导型启动子。参见ward等,plantmol.biol.22:361-366(1993)。例示性诱导型启动子包括但不限于:那些响应铜的acei系统的启动子(mett等,pnas90:4567-4571(1993));响应苯磺酰胺除草剂安全剂的玉米的in2基因(hershey等,mol.gen.genetics227:229-237(1991);和gatz等,mol.gen.genetics243:32-38(1994));和来自tn10的tet阻抑物(gatz等,mol.gen.genetics227:229-237(1991))。特别优选的诱导型启动子是响应植物正常不响应的诱导剂的启动子。例示性诱导型启动子是来自类固醇激素基因的诱导型启动子,其转录活性受糖皮质激素诱导。schena等,proc.natl.acad.sci.u.s.a.88:0421(1991)。
b.组成性启动子
将组成性启动子与向日葵中表达的基因可操作连接,或者将组成性启动子与编码信号序列的核苷酸序列可操作连接,所述编码信号序列的核苷酸序列与向日葵中表达的基因可操作连接。
可以在本发明中利用不同的组成性启动子。例示性的组成性启动子包括但不限于:来自植物病毒的启动子,诸如来自camv的35s启动子(odell等,nature313:810-812(1985));来自稻肌动蛋白基因的启动子(mcelroy等,plantcell2:163-171(1990));来自泛素的启动子(christensen等,plantmol.biol.12:619-632(1989),和christensen等,plantmol.biol.18:675-689(1992));来自pemu的启动子(last等,theor.appl.genet.81:581-588(1991));来自mas的启动子(velten等,emboj.3:2723-2730(1984));和来自玉米h3组蛋白的启动子(lepetit等,mol.gen.genetics231:276-285(1992),和atanassova等,plantjournal2(3):291-300(1992))。als启动子,即欧洲油菜(brassicanapus)als3结构基因5’的xba1/ncoi片段(或与该xba1/ncoi片段相似的核苷酸序列)代表一种特别有用的组成性启动子。参见pct申请wo96/30530。
c.组织特异性或组织优选性启动子
将组织特异性启动子与向日葵中表达的基因可操作连接。任选地,将组织特异性启动子与编码信号序列的核苷酸序列可操作连接,所述编码信号序列的核苷酸序列与向日葵中表达的基因可操作连接。用与组织特异性启动子可操作连接的感兴趣基因转化的植物可以专门或优先在特定组织中生成转基因的蛋白质产物。
可以在本发明中利用任何组织特异性或组织优选性启动子。例示性组织特异性或组织优选性启动子包括但不限于:根优选性启动子,诸如来自菜豆蛋白基因的启动子(murai等,science23:476-482(1983),和sengupta-gopalan等,proc.natl.acad.sci.u.s.a.82:3320-3324(1985));叶特异性和光诱导型启动子,诸如来自cab或核酮糖二磷酸羧化酶-加氧酶(rubisco)的启动子(simpson等,emboj.4(11):2723-2729(1985),和timko等,nature318:579-582(1985));花药特异性启动子,诸如来自lat52的启动子(twell等,mol.gen.genetics217:240-245(1989));花粉特异性启动子,诸如来自zm13的启动子(guerrero等,mol.gen.genetics244:161-168(1993))或小孢子优选性启动子,诸如来自apg的启动子(twell等,sex.plantreprod.6:217-224(1993))。
可以通过将编码信号序列的核苷酸序列可操作连接至编码感兴趣蛋白质的基因的5’和/或3’区域的手段来实现由转基因生成的蛋白质向亚细胞区室诸如叶绿体、液泡、过氧化物酶体、乙醛酸循环体、细胞壁或线粒体的转运或者分泌入质外体中。可以在蛋白质合成和加工过程中测定在结构基因的5’和/或3’端的靶向序列,其中编码的蛋白质最后被区室化(compartmentalized)。
信号序列的存在将多肽引导至胞内细胞器或亚细胞区室,或者分泌至质外体。许多信号序列是本领域中已知的。参见例如becker等,plantmol.biol.20:49(1992);p.s.close,master’sthesis,iowastateuniversity(1993);c.knox等,“structureandorganizationoftwodivergentalpha-amylasegenesfrombarley,”plantmol.biol.9:3-17(1987);lerner等,plantphysiol.91:124-129(1989);fontes等,plantcell3:483-496(1991);matsuoka等,proc.natlacad.sci.88:834(1991);gould等,j.cell.biol.108:1657(1989);creissen等,plantj.2:129(1991);kalderon等,ashortaminoacidsequenceabletospecifynuclearlocation,cell39:499-509(1984);steifel等,expressionofamaizecellwallhydroxyproline-richglycoproteingeneinearlyleafandrootvasculardifferentiation,plantcell2:785-793(1990)。
外来蛋白质基因和农艺学基因
通过根据本发明的转基因植物,可以以商业量生产外来蛋白质。如此,用于选择和繁殖经转化植物的技术(它们是本领域中充分了解的)产生多种转基因植物,它们以常规方式收获,然后可以从感兴趣的组织或总生物量提取外来蛋白质。可以通过由例如heney和orr,anal.biochem.114:92-6(1981)所讨论的已知方法来实现从植物生物量的蛋白质提取。
在本发明的各方面,为商业生产外来蛋白质提供的转基因植物是向日葵植物。在其它方面,感兴趣的生物量是种子。对于相对少量的显示较高表达水平的转基因植物,可以主要经由常规的rflp、pcr和ssr分析(其鉴定整合dna分子的近似染色体位置)来产生遗传图谱。关于这方面的例示性方法,参见glick和thompson,methodsinplantmolecularbiologyandbiotechnology,crcpress,bocaraton269:284(1993)。关于染色体位置的图谱信息对于主题转基因植物的所有权保护是有用的。如果进行未经授权的繁殖和与其它种质进行杂交,则可以将整合区域的图谱与可疑植物的类似图谱进行比较以确定后者是否与主题植物具有共同来源(parentage)。图谱比较会牵涉杂交、rflp、pcr、ssr和测序,它们都是常规技术。
同样地,可以在经转化的植物中表达农艺学基因。更具体地,可以对植物进行遗传工程化改造以表达农艺学感兴趣的各种表型。可以在这方面使用的例示性基因包括但不限于下文进行归类的基因。
1.赋予对害虫或疾病的抗性并编码下列各项的基因:
a)植物疾病抗性基因。植物防御常常由植物中的疾病抗性基因(r)产物与病原体中相应无毒性(avr)基因产物之间的特异性相互作用激活。可以用克隆的抗性基因转化植物品种以工程化改造对特定病原体株有抗性的植物。参见例如jones等,science266:789(1994)(克隆对黄枝孢霉(cladosporiumfulvum)有抗性的番茄cf-9基因);martin等,science262:1432(1993)(对丁香假单胞菌番茄致病变种(pseudomonassyringaepv.tomato)有抗性的番茄pto基因编码蛋白质激酶);mindrinos等,cell78:1089(1994)(对丁香假单胞菌(pseudomonassyringae)有抗性的拟南芥(arabidopsis)rsp2基因)。
b)赋予对害虫诸如大豆胞囊线虫有抗性的基因。参见例如pct申请wo96/30517;pct申请wo93/19181。
c)苏云金芽孢杆菌(bacillusthuringiensis)蛋白、其衍生物或模仿其的合成多肽。参见例如geiser等,gene48:109(1986),其披露了btδ-内毒素基因的克隆和核苷酸序列。此外,编码δ-内毒素基因的dna分子可以购自美国典型培养物保藏中心(americantypeculturecollection),manassas,va.,例如以atcc保藏号40098、670136、31995和31998购得。
d)凝集素。参见例如vandamme等,plantmolec.biol.24:25(1994)的公开内容,其披露了数种君子兰(cliviaminiata)甘露糖结合凝集素基因的核苷酸序列。
e)维生素结合蛋白诸如抗生物素蛋白。参见pct申请us93/06487。该申请教导了抗生物素蛋白和抗生物素蛋白同源物作为针对虫害的杀幼虫剂的用途。
f)酶抑制剂,例如蛋白酶或蛋白水解酶抑制剂或淀粉酶抑制剂。参见例如abe等,j.biol.chem.262:16793(1987)(稻半胱氨酸蛋白水解酶抑制剂的核苷酸序列);huub等,plantmolec.biol.21:985(1993)(编码烟草蛋白水解酶抑制剂i的cdna的核苷酸序列);sumitani等,biosci.biotech.biochem.57:1243(1993)(硝孢链霉菌(streptomycesnitrosporeus)α-淀粉酶抑制剂的核苷酸序列);和美国专利号5,494,813(hepher和atkinson,1996年2月27日公开)。
g)昆虫特异性激素或信息素,诸如蜕皮类固醇或保幼激素,其变体、基于其的模拟物、或其拮抗剂或激动剂。参见例如hammock等,nature344:458(1990)披露的克隆的保幼激素酯酶(即一种保幼激素灭活剂)的杆状病毒表达的内容。
h)昆虫特异性肽或神经肽,其在表达后破坏受到影响的害虫的生理学。例如参见regan,j.biol.chem.269:9(1994)(表达克隆产生编码昆虫利尿激素受体的dna)和pratt等,biochem.biophys.res.comm.163:1243(1989)(在太平洋折翅蠊(diplopterapuntata)中鉴定出咽侧体抑制素(allostatin))的公开内容。还可参见tomalski等的美国专利号5,266,317,其披露了编码昆虫特异性、麻痹性神经毒素的基因。
i)在自然界由蛇、黄蜂(wasp)等生成的昆虫特异性毒液。例如参见pang等,gene116:165(1992),是关于编码蝎昆虫毒性肽的基因在植物中的异源表达的公开内容。
j)酶,其负责超积累单萜、倍半萜、类固醇、异羟肟酸、类苯丙烷衍生物或另一具有杀昆虫活性的非蛋白质分子。
k)牵涉对生物学活性分子的修饰(包括翻译后修饰)的酶;例如,糖酵解酶、蛋白水解酶、脂肪分解酶、核酸酶、环化酶、转氨酶、酯酶、水解酶、磷酸酶、激酶、磷酸化酶、聚合酶、弹性蛋白酶、几丁质酶(chitinase)和葡聚糖酶(不管是天然的还是合成的)。参见以scott等名义的pct申请wo93/02197,其披露了callase基因的核苷酸序列。含有几丁质酶编码序列的dna分子可以例如从atcc以保藏号39637和67152获得。还可参见kramer等,insectbiochem.molec.biol.23:691(1993)(其教导了编码烟草天蛾几丁质酶的cdna的核苷酸序列)和kawalleck等,plantmolec.biol.21:673(1993)(其提供了欧芹ubi4-2多聚泛素基因的核苷酸序列)。
l)刺激信号转导的分子。例如参见botella等,plantmolec.biol.24:757(1994)(关于绿豆钙调蛋白cdna克隆的核苷酸序列)和griess等,plantphysiol.104:1467(1994)(其提供了玉米钙调蛋白cdna克隆的核苷酸序列)的公开内容。
m)疏水矩肽(hydrophobicmomentpeptide)。参见pct申请wo95/16776(抑制真菌植物病原体的鲎抗菌肽(tachyplesin)的肽衍生物的公开内容)和pct申请wo95/18855(教导了赋予疾病抗性的合成抗微生物肽)。
n)膜通透酶、通道形成剂或通道阻断剂。例如参见jaynes等,plantsci.89:43(1993)公开的杀菌肽-β溶胞肽类似物(cecropin-βlyticpeptideanalog)的异源表达以使转基因烟草植物对青枯假单胞菌(pseudomonassolanacearum)有抗性。
o)病毒侵入蛋白或自其衍生的复合毒素。例如,病毒外壳蛋白在经转化植物细胞中的积累赋予对由衍生出该外壳蛋白基因的病毒及相关病毒招致的病毒感染和/或疾病形成的抗性。参见beachy等,ann.rev.phytopathol.28:451(1990)。已经对经转化的植物赋予了针对苜蓿花叶病毒、黄瓜花叶病毒、烟草线条病毒、马铃薯x病毒、马铃薯y病毒、烟草蚀刻病毒、烟草脆裂病毒和烟草花叶病毒的外壳蛋白介导的抗性。同上。
p)昆虫特异性抗体或自其衍生的免疫毒素。如此,靶向昆虫肠中的重要代谢功能的抗体会使受影响的酶失活,这杀死昆虫。参见taylor等,摘要号497,seventhint’lsymposiumonmolecularplant-microbeinteractions(edinburgh,scotland)(1994)(经由生成单链抗体片段进行的转基因烟草中的酶促失活)。
q)病毒特异性抗体。参见例如tavladoraki等,nature366:469(1993),其显示了表达重组抗体基因的转基因植物受到保护免于病毒攻击。
r)在自然界由病原体或寄生物生成的发育阻滞蛋白(developmental-arrestiveprotein)。如此,真菌内α-l,4-d-多聚半乳糖醛酸酶通过溶解植物细胞壁同-α-1,4-d-半乳糖醛酸酶(galacturonase)来促进真菌定殖(colonization)和植物营养物释放。参见lamb等,bio/technology10:1436(1992)。编码豆内多聚半乳糖醛酸酶抑制蛋白的基因的克隆和表征由toubart等,plantj.2:367(1992)描述。
s)在自然界由植物生成的发育阻滞蛋白。例如logemann等,bio/technology10:305(1992)显示了表达大麦核糖体失活基因的转基因植物对真菌疾病具有升高的抗性。
2.赋予对除草剂的抗性的基因:
a)抑制生长点或分生组织的除草剂,诸如咪唑啉酮或磺酰脲。在这类中的例示性基因编码突变的als和ahas酶,如分别由例如lee等,emboj.7:1241(1988)和miki等,theor.appl.genet.80:449(1990)所描述的。
b)抑制光合作用的除草剂,诸如三嗪(psba和gs+基因)或苯基氰(腈水解酶基因)。przibila等,plantcell3:169(1991)描述了用编码突变的psba基因的质粒转化衣滴虫(chlamydomonas)。腈水解酶基因的核苷酸序列披露于stalker的美国专利号4,810,648,并且含有这些基因的dna分子可以以atcc保藏号53435、67441、和67442获得。编码谷胱甘肽s-转移酶的dna的克隆和表达由hayes等,biochem.j.285:173(1992)描述。
3.赋予或促成如下增值(value-added)性状的基因,诸如:
a)经修饰的脂肪酸代谢,例如通过用硬脂酰-acp去饱和酶的反义基因转化植物以提高植物的硬脂酸含量。参见knultzon等,proc.natl.acad.sci.u.s.a.89:2624(1992)。
b)降低的肌醇六磷酸盐含量——1)肌醇六磷酸酶编码基因的导入会增强肌醇六磷酸盐的分解,向经转化的植物添加更多游离的磷酸盐。例如,参见vanhartingsveldt等,gene127:87(1993),关于黑曲霉(aspergillusniger)肌醇六磷酸酶基因的核苷酸序列的公开内容。2)可以导入降低肌醇六磷酸盐含量的基因。例如在玉米中,这可以如下实现,即克隆与单一等位基因有关的dna,然后再次导入,所述单一等位基因负责特征为低水平肌醇六磷酸的玉米突变体。参见raboy等,maydica35:383(1990)。
c)例如通过用编码改变淀粉分支样式的酶的基因转化植物来实现的经修饰的碳水化合物组成。参见shiroza等,j.bacteol.170:810(1988)(链球菌(streptococcus)突变体果糖基转移酶基因的核苷酸序列),steinmetz等,mol.gen.genet.20:220(1985)(枯草芽孢杆菌(bacillussubtilis)果聚糖蔗糖酶(levansucrase)基因的核苷酸序列);pen等,bio/technology10:292(1992)(表达地衣芽胞杆菌(bacilluslichenifonnis)α-淀粉酶的转基因植物的生成);elliot等,plantmolec.biol.21:515(1993)(番茄转化酶基因的核苷酸序列);sogaard等,j.biol.chem.268:22480(1993)(大麦α-淀粉酶基因的定点诱变);和fisher等,plantphysiol.102:1045(1993)(玉米胚乳淀粉分支酶ii)。
用于向日葵转化的方法
已经开发了许多用于植物转化的方法,包括生物学和物理植物转化方案。参见例如miki等,“proceduresforintroducingforeigndnaintoplants”于methodsinplantmolecularbiologyandbiotechnology,b.r.glick和j.e.thompson编(crcpress,inc.,bocaraton,1993)第67页-第88页。另外,可获得用于植物细胞或组织转化及植物再生的表达载体和体外培养方法。参见例如gruber等,“vectorsforplanttransformation”于methodsinplantmolecularbiologyandbiotechnology,b.r.glick和j.e.thompson编(crcpress,inc.,bocaraton,1993)第89页-第119页。
a)土壤杆菌介导的转化——一种用于将表达载体导入植物中的方法是基于土壤杆菌属的天然转化系统。参见例如horsch等,science227:1229(1985).根癌土壤杆菌(a.tumefaciens)和发根土壤杆菌(a.rhizogenes)是在遗传上转化植物细胞的植物病原性土壤细菌。根癌土壤杆菌(a.tumefaciens)的ti质粒和发根土壤杆菌(a.rhizogenes)的ri质粒携带负责植物的遗传转化的基因。参见例如c.i.kado,crit.rev.plantsci.10:1(1991)。土壤杆菌属载体系统和土壤杆菌介导的基因转移方法的描述由gruber等,见上文,miki等,见上文,及moloney等,plantcellreports8:238(1989)提供。还可参见美国专利号5,563,055(townsend和thomas),1996年10月8日公开。
b)直接基因转移——已经开发了数种植物转化方法(统称为直接基因转移)作为土壤杆菌介导的转化的备选。植物转化普遍可适用的方法是微粒介导的转化,其中在测量为1至4μm的微粒的表面上携带dna。用生物射弹装置将表达载体导入植物组织中,所述生物射弹装置将微粒加速到300至600m/s的速度,其足以穿透植物细胞壁和细胞膜。sanford等,part.sci.technol.5:27(1987);j.c.sanford,trendsbiotech.6:299(1988);klein等,bio/technology6:559-563(1988);j.c.sanford,physiol.plant7:206(1990);klein等,biotechnology10:268(1992)。还可参见美国专利号5,015,580(christou等),1991年5月14日公开;美国专利号5,322,783(tomes等),1994年6月21日公开。
另一种用于将dna物理投递至植物的方法是对靶细胞的超声处理。zhang等,bio/technology9:996(1991)。或者,已经使用脂质体和原生质球融合来将表达载体导入植物中。deshayes等,emboj,4:2731(1985);christou等,procnatl.acad.sci.u.s.a.84:3962(1987)。还已经报道了使用cacl2沉淀、聚乙烯醇或多-l-鸟氨酸来将dna直接摄取到原生质体中。hain等,mol.gen.genet.199:161(1985),和draper等,plantcellphysiol.23:451(1982)。还已经描述了原生质体和全细胞和组织的电穿孔。donn等,于abstractsofviithinternationalcongressonplantcellandtissuecultureiaptc,a2-38,第53页(1990);d’halluin等,plantcell4:1495-1505(1992),和spencer等,plantmol.biol.24:51-61(1994)。
转化向日葵靶标组织后,上述选择性标志物基因的表达容许优先选择经转化的细胞、组织和/或植物,其中使用本领域中公知的再生和选择方法来实现。
前述转化方法通常会用于产生转基因品种。然后可以将转基因品种与另一(非转化的或经转化的)品种杂交,以产生新转基因品种。或者,可以使用植物育种领域中公知的传统回交技术将遗传性状移入另一栽培种中,所述遗传性状已经使用前述转化技术工程化改造入特定的向日葵栽培种中。例如,可以使用回交方法来将工程化改造的性状从公共的、非良种品种移入良种品种中,或者从在其基因组中含有外来基因的品种移入不含所述基因的一种或多种品种中。如本文中所使用的,“杂交”可以指简单的xxy杂交,或回交过程,这取决于上下文。
向日葵的组织培养
可以通过自花传粉或者通过组织培养和再生来进行向日葵植物的进一步生成,所述向日葵植物产生具有低饱和脂肪和任选地高亚油酸含量的种子。向日葵的各种组织的组织培养和自其再生植物是已知的。例如,通过组织培养繁殖向日葵栽培种记载于美国专利6,998,516。
可以通过组织培养和再生来进行品种的进一步繁殖。大豆的各种组织的组织培养和自其再生植物是公知且广泛公布的。例如,可以参照美国专利6,998,516。如此,本发明的另一方面是提供如下的细胞,其在生长和分化后产生具有含低饱和脂肪和任选地高亚油酸含量的种子的向日葵植物。
如本文中所使用的,术语“组织培养物”指包含相同或不同类型的分离的细胞的组合物,或者组织成植物部分的此类细胞的集合。组织培养物的例示性类型包括能产生组织培养物且在植物或植物部分中完整的原生质体、愈伤组织、植物块(plantclump)、和植物细胞,诸如胚、花粉、花、种子、荚果、叶、茎、根、根尖、花药等。用于制备和维持植物组织培养物的手段是本领域中公知的。举例而言,已经使用构成器官的组织培养物来产生再生植物。美国专利号5,959,185、5,973,234、5,977,445、和6,998,516描述了某些技术。
单基因转变的(转变)植物
当术语“向日葵植物”在本发明的语境中使用时,这还包括所述品种的任何单基因转变。如本文中所使用的,术语“单基因转变的植物”指通过称作回交的植物育种技术,或者经由基因工程开发的那些向日葵植物,其中除了经由回交技术转移到品种中的单基因外,该品种的基本上所有期望的形态学和生理学特征均得到复原。回交方法可以与本发明一起使用以改善品种或将某种特征引入品种中。如本文中所使用的,术语“回交”指杂种子代往回与回归亲本的重复杂交(即与回归亲本回交1、2、3、4、5、6、7、8或更多次)。亲本向日葵植物(其贡献期望特征的基因)称作“非回归”或“供体亲本”。此术语指在回交方案中使用非回归亲本一次,并且非回归亲本因此不再出现的事实。接受来自非回归亲本的一种或多种基因转移的亲本向日葵植物称为回归亲本,因为它在回交方案中被使用了数轮(poehlman和sleper,1994;fehr,1987)。在典型的回交方案中,将感兴趣的原始品种(回归亲本)与第二品种(非回归亲本)杂交,所述第二品种携带要转移的感兴趣的单基因。然后将自此杂交所得的子代再次与回归亲本杂交,并重复该过程,直至获得这样的向日葵植物,即其中除了来自非回归亲本的单转移基因外,回归亲本的基本上所有期望的形态学和生理学特征均得到复原。
选择合适的回归亲本是成功回交方法的重要步骤。回交方案的目的是改变或替换原始品种中的单一性状或特征。为了实现这点,对回归品种的单基因进行修饰或者用来自非回归亲本的期望基因替换,而保留原始品种的基本上所有剩余的期望的遗传构成,和因此保留基本上所有剩余的期望的生理学和形态学构成。特定非回归亲本的选择会取决于回交目的。主要目的之一是将一些商业上期望的、农艺学上重要的性状添加至植物。确切的回交方案会取决于所改变的特征或性状以确定合适的测试方案。虽然当所转移的特征是显性等位基因时回交方法得到简化,但是也可以转移隐性等位基因。在此例子中,引入子代测试来测定期望的特征是否已经得到成功转移可能是必要的。
已经鉴定出许多单基因性状,它们在新品种的开发中不会有规律地被选择,但是可以通过回交技术来改良。单基因性状可以是或者不是转基因的,这些性状的例子包括但不限于雄性不育、蜡质淀粉、除草剂抗性、对细菌、真菌、或病毒疾病的抗性、抗虫性、雄性能育、提高的营养质量、工业应用、产率稳定性和产率提高。这些基因一般经由细胞核遗传。这些单基因性状之中的数个记载于美国专利号5,959,185、5,973,234和5,977,445。
本发明还涉及用于产生向日葵植物的方法,其通过将第一亲本向日葵植物与第二亲本向日葵植物杂交来进行,其中所述第一或第二亲本向日葵植物是产生具有低饱和脂肪和任选地高亚油酸含量的种子的向日葵植物。此外,第一和第二亲本向日葵植物两者都可以来源于产生具有低饱和脂肪和任选地高亚油酸含量的种子的向日葵植物。如此,使用产生具有低饱和脂肪和任选地高亚油酸含量的种子的向日葵植物进行的任何此类方法是本发明的一部分(例如自交、回交、杂种生成、与群体杂交等)。使用产生具有低饱和脂肪和任选地高亚油酸含量的种子的向日葵植物作为亲本产生的所有植物在本发明的范围内,包括那些自从产生具有低饱和脂肪和任选地高亚油酸含量的种子的向日葵植物衍生的品种开发的。有利地,可以在与其它不同向日葵植物的杂交中使用所述向日葵品种以产生具有卓越特征的第一代(f1)向日葵杂种种子和植物。本发明的品种也可以用于转化,其中导入外源基因,并由本发明的品种进行表达。使用如下向日葵植物经由传统育种方法或者经由通过本领域技术人员已知的许多方案之任一种转化如下向日葵植物而创建的遗传变体意图在本发明的范围内,所述向日葵植物产生具有低饱和脂肪和任选地高亚油酸含量的种子。
实施例
本发明在以下实施例中进行进一步描述,所述实施例作为例示提供,而并不意图以任何方式限制本发明。
实施例1:产生具有低饱和脂肪含量的种子的向日葵
已经经由正常的育种技术开发出了具有低得异乎寻常的饱和类水平(saturatelevel)的向日葵种质。表1中提供了向日葵栽培种的种子含油量。
表1
实施例2:产生具有低饱和脂肪含量和高亚油酸含量的种子的向日葵
已经经由正常的育种技术开发出了具有低得异乎寻常的饱和类水平的向日葵种质。表2中提供了向日葵栽培种的种子含油量。
表2
实施例3:产生具有低饱和脂肪含量的种子的向日葵
已经经由正常的育种技术开发出了具有低得异乎寻常的饱和类水平的向日葵种质。表3中提供了向日葵栽培种的种子含油量。
表3
如表3中可看出的,数据表明如下的种子油,其具有高油酸(>80%)背景中低到2.33%的总饱和类,nusun(55-50%油酸)背景中没有饱和类(<3.5%)谱,高达95.30%的油酸水平;低到0.25%的硬脂酸水平,低到1.47%的棕榈酸水平,和亚油酸(<55%油酸)背景中的低饱和类(<7.0%)谱。
实施例4:产生具有低饱和脂肪、硬脂酸、和棕榈酸含量的种子的向日葵
已经经由正常的育种技术开发出了具有低得异乎寻常的饱和类水平的向日葵种质。表4中提供了向日葵栽培种的种子含油量。
如表4中可看出的,此组数据包括硬脂酸(0.23%)、棕榈酸(1.37%)、和总饱和油(2.28%)的低数值。
实施例5:低硬脂酸和低棕榈酸的标志物开发
如本文中所描述的,开发了标志物开发策略。首先,鉴定了来自靶标qtl区域的标志物(在dowagrosciences开发以及来自公共资源),并筛选相应作图群体的亲本品系之间的多态性。然后在作图群体中筛选了多态性标志物。对于单态(未提供信息的)标志物,设计了引物以扩增其相应的基因组基因座,并对扩增子测序以鉴定亲本品系之间的单核苷酸多态性(snp)(若有的话)。开发了taqmanmgb等位辨别测定法(allelicdiscriminationassay)用于鉴定出的snp,并在各自的群体上定位。第二,基于脂肪酸候选基因的序列,设计在内含子侧翼的引物来从亲本品系分离脂肪酸基因序列。将在序列水平的核苷酸多态性基于其多态性性质开发成标志物,然后在作图群体中筛选。采用joinmap3.0(vanooijen,2004a)来定位新开发的标志物,并使用mapqtl5(vanooijen,2004b)来精细定位qtl(tofinemapqtl)。
a)低硬脂酸的标志物开发
ssr标志物开发:对8种ssr标志物筛选onn687rxh757b/ls10760b-b-17-3-23-5作图群图的亲本品系onn687r和h757b/ls10760b-b-17-3-23-5之间的多态性,所述作图群体先前用于定位目标低硬脂酸qtl。(参见表5。)4种ssr标志物,即ha0442、crt22、ors565和ors732是多态性的。在abi3730测序仪上解析来自onn687r和h757b/ls10760b-b-17-3-23-5的ha0442和crt22扩增子,并且对于标志物ha0442分别为163bp和165bp,而对于crt22分别为290bp和261bp。在3%metaphor凝胶上解析来自onn687r和h757b/ls10760b-b-17-3-23-5的ors565和ors732扩增子。使用下列pcr引物和反应条件用ha0442、crt22、ors565和ors732对相应作图群体onn687rxh757b/ls10760b-b-17-3-23-5进行基因型分型。
ha0442正向引物:5’-hex-tggaactgtaaatggacccaag-3’(seqidno:1)
ha0442反向引物:5’-gcactgcaccatttatgagaag-3’(seqidno:2)
crt22正向引物:5’-hex-tcgagatgaaaccgaatgaagaaa-3’(seqidno:3)
crt22反向引物:5’-gtttcttgggactgatattgccaagtggg-3’(seqidno:4)
ors565正向引物:5’-tggtcaacggatttagagtcaa-3’(seqidno:5)
ors565反向引物:5’-tccagtttggtcttgatttgg-3’(seqidno:6)
ors732正向引物:5’-gcacggaactcctcaaatgt-3’(seqidno:7)
ors732反向引物:5’-gcacgggaaacaaagagtca-3’(seqidno:8)
pcr成分:
4nggdna
1xpcr缓冲液(qiagen,valencia,california)
0.25μm正向引物
0.25μm反向引物
1mmmgcl2
0.1mm每种dntp
0.4%pvp
0.04个单位的hotstartaqdna聚合酶(qiagen,valencia,california)
总体积:4.8μl
温度循环仪设置:
步骤1:94℃达12分钟
步骤2:94℃达30秒
步骤3:55℃达30秒
步骤4:72℃达30秒
步骤5:重复步骤2、3和4达35个循环
步骤6:72℃达30分钟
snp标志物开发:使用了8对引物来从onn687r和h757b/ls10760b-b-17-3-23-5两者扩增8个基因组基因座以开发snp标志物(表6)。基于来自限制性片段长度多态性(rflp)探针zvg76、zvg77和zvg78的序列(kolkman等2007),设计了三个引物对(zvg76snpf/r、zvg77snpf/r、和zvg78snpf/r)。ht57f/r、ht64f/r、ht131f/r、ht134f/r、和ht210f/r的引物序列来自lai等(2005)。发现snp在来自ht64f/r、ht210f/r、和zvg78snpf/r的扩增子中。将taqmanmgb等位辨别测定法开发用于ht64f/r扩增子中的一个snp基因座和zvg78snpf/r扩增子中的一个snp基因座(参见下文),并使用开发的taqman测定法用这两种snp标志物对onn687rxh757b/ls10760b-b-17-3-23-5作图群体进行基因型分型。
来自onn687r和h757b/ls10760b-b-17-3-23-5的ht64f/r扩增子中有4个snp基因座(以粗体标记)。将taqman测定法开发用于r-基因座。正向引物、反向引物、探针1和探针2的序列分别为5’-ccggctgcttctagaccttataag-3’(seqidno:9)、5’-tcgtcggtgggacacaca-3’(seqidno:10)、5’-6fam-actgttggatcggttc-3’(seqidno:11)、和5’-vic-cactgttggatcgatt-3’(seqidno-12)。
ttattctcggcttccggtgtgattttactctcatggttaagttttcaagagattgtcgcy(t/c)gctgaaaactttttatattgtttcggtatgatcttggagtttatagcctttgtaaggttaagaatgaaacacccggctgcttctagaccttataagatacccgtgggcactgttggatcgr(a/g)ttcttctgtgtgtcccaccgacgattttgatctgtgtcgtgttggctctttcttcactcaaggtcatgatcgttagy(t/c)gty(c/t)attgccatatttttcgggttcgcattgcaaccgtttttaaagtttgccgagaagaaaagatggcttaaattttcaactaaagccgatcttcccg(seqidno:13)
来自onn687r和h757b/ls10760b-b-17-3-23-5的zvg78snpf/r扩增子中也有4个snp基因座(以粗体标记)。将taqman测定法开发用于在5’末端的r-基因座。正向引物、反向引物、探针1和探针2的序列分别为5’-gtccatctttcctcaacgacttg-3’(seqidno:14)、5’-cctaaacgcctcgaaaaagct-3’(seqidno:15)、5’-6fam-ttaccatgtctataatgc-3’(seqidno:16)、和5’-vic-attaccatgtctgtaatgc-3’(seqidno:17)。
aactgagttctgtacgccagagatttgcccgaccatgaccgcaggtccaaagtaagtcttgctattgcacatttgcacgattaacggtttcttatatagaagatacatgattcttgaatttatgtaaataaaacttgacagatatgaataccgatgggctgatggtgtgcaaatcaagaagcctattgaagtttcggctccaaagtacgtagagttcttgatggattggattgagtcacaattggatgacgagtccatctttcctcaacgacttggtaattagttaattaccatgtctr(g/a)taatgcatcatttaataaagctttttcgaggcgtttaggaaactgaaatagtaattttcgattgy(t/c)cgtgcaggagcgccatttcccgccaattttagggacgttgtgaaaacgatatttaaacgcttgtttcgtgtatay(t/c)gcgcatatctaccacacr(g/a)cattttcagaagattgtgagtcttaaagaagaagcccatctaaacacttgtttcaagcatttcatattgtttacatgtgtaa(seqidno:18)
对这两种snp标志物使用下列pcr设置。
实时pcr成分:
25nggdna
1xtaqman通用pcr主混合物(mastermix)
22.5μm正向引物
22.5μm反向引物
5μm探针1
5μm探针2
总体积:25μl
bio-radicycler设置:
步骤1:95℃达15分钟
步骤2:94℃达30秒
步骤3:60℃达1分钟
步骤4:重复步骤2和3达65个循环
步骤5:4℃保持(forever)
indel标志物开发:设计引物来从两个亲本品系onn687r和h757b/ls10760b-b-17-3-23-5扩增了32个脂肪酸相关基因,并进行了测序。7种基因具有多态性,4种基因具有弱扩增,而所有其它基因是单态性的(表6)。用所有鉴定出的多态性筛选作图群体onn687rxh757b/ls10760b-b-17-3-23-5。
定位新标志物并精细定位低硬脂酸qtl:使用joinmap3.0(vanooijen,2004a)来定位所有新鉴定的多态性标志物。标志物crt22给出显著的分离异常,而且没有被定位。从候选基因方法开发的6种标志物定位到与靶染色体17不同的染色体(表6)。将7种标志物ha0442、ors565、ht64、zvg78、kasi-2、kasi-4、和ors732定位到染色体17。将脂肪酸基因kasi-2和kasi-4定位到染色体17,但不靠近靶标低硬脂酸qtl(图1)。通过新定位的标志物,用mapqtl5((vanooijen,2004b)在ha1875-ors565区间中精细定位低硬脂酸qtl,所述ha1875-ors565区间在lg17的上部端粒区中跨越27cm。精细定位的qtl具有23.2的高lod得分,而且解释了硬脂酸含量方面50.8%的变化。可以使用新定位的标志物来辅助在育种程序中选择低硬脂酸。
b)开发和定位棕榈酸qtl的indel标志物
在用脂肪酸基因kasiii-2的引物对得到的亲本品系h280r[1]/687r-1-8-1和ond163r的扩增子序列中观察到了snp和indels(表6,图2)。用此引物对筛选作图群体h280r[1]/687r-1-8-1xond163r,并在3%metaphor凝胶上解析扩增子。作图程序joinmap3.0(vanooijen,2004a)在连锁群5上的低棕榈酸qtl内部定位了这种indel标志物(图3)。
表6:调查的脂肪酸基因。
虽然本发明已经在某些实施方案中进行了描述,但是可以在本公开内容的精神和范围内进一步修饰本发明。因此,本申请意图覆盖本发明使用其一般原理的任何变型、用途、或改编。此外,本申请意图覆盖诸如在本发明所属领域中的已知或习惯实践内和落入所附权利要求书的限定内的对本公开内容的偏离。
序列表
<110>陶氏益农公司(dowagrosciencesllc)
gerdes,james
dohman,angela
radtke,james
<120>低饱和脂肪向日葵及相关方法
<130>29718493.1us
<150>61/015,591
<151>2007-12-20
<160>41
<170>patentinversion3.5
<210>1
<211>22
<212>dna
<213>人工序列
<220>
<223>化学合成的-ha0442正向引物
<400>1
tggaactgtaaatggacccaag22
<210>2
<211>22
<212>dna
<213>人工序列
<220>
<223>化学合成的-ha0442反向引物
<400>2
gcactgcaccatttatgagaag22
<210>3
<211>24
<212>dna
<213>人工序列
<220>
<223>化学合成的-crt22正向引物
<400>3
tcgagatgaaaccgaatgaagaaa24
<210>4
<211>29
<212>dna
<213>人工序列
<220>
<223>化学合成的-crt22反向引物
<400>4
gtttcttgggactgatattgccaagtggg29
<210>5
<211>22
<212>dna
<213>人工序列
<220>
<223>化学合成的-ors565正向引物
<400>5
tggtcaacggatttagagtcaa22
<210>6
<211>21
<212>dna
<213>人工序列
<220>
<223>化学合成的-ors565反向引物
<400>6
tccagtttggtcttgatttgg21
<210>7
<211>20
<212>dna
<213>人工序列
<220>
<223>化学合成的-ors732正向引物
<400>7
gcacggaactcctcaaatgt20
<210>8
<211>20
<212>dna
<213>人工序列
<220>
<223>化学合成的-ors732反向引物
<400>8
gcacgggaaacaaagagtca20
<210>9
<211>24
<212>dna
<213>人工序列
<220>
<223>化学合成的
<400>9
ccggctgcttctagaccttataag24
<210>10
<211>18
<212>dna
<213>人工序列
<220>
<223>化学合成的
<400>10
tcgtcggtgggacacaca18
<210>11
<211>16
<212>dna
<213>人工序列
<220>
<223>化学合成的
<400>11
actgttggatcggttc16
<210>12
<211>16
<212>dna
<213>人工序列
<220>
<223>化学合成的
<400>12
cactgttggatcgatt16
<210>13
<211>355
<212>dna
<213>人工序列
<220>
<223>化学合成的
<220>
<221>misc_feature
<222>(60)..(60)
<223>n可以是t或c
<220>
<221>misc_feature
<222>(181)..(181)
<223>n可以是a或g
<220>
<221>misc_feature
<222>(258)..(260)
<223>n可以是t或c
<220>
<221>misc_feature
<222>(261)..(261)
<223>n可以是t或c
<400>13
ttattctcggcttccggtgtgattttactctcatggttaagttttcaagagattgtcgcn60
gctgaaaactttttatattgtttcggtatgatcttggagtttatagcctttgtaaggtta120
agaatgaaacacccggctgcttctagaccttataagatacccgtgggcactgttggatcg180
nttcttctgtgtgtcccaccgacgattttgatctgtgtcgtgttggctctttcttcactc240
aaggtcatgatcgttagngtnattgccatatttttcgggttcgcattgcaaccgttttta300
aagtttgccgagaagaaaagatggcttaaattttcaactaaagccgatcttcccg355
<210>14
<211>23
<212>dna
<213>人工序列
<220>
<223>化学合成的
<400>14
gtccatctttcctcaacgacttg23
<210>15
<211>21
<212>dna
<213>人工序列
<220>
<223>化学合成的
<400>15
cctaaacgcctcgaaaaagct21
<210>16
<211>18
<212>dna
<213>人工序列
<220>
<223>化学合成的
<400>16
ttaccatgtctataatgc18
<210>17
<211>19
<212>dna
<213>人工序列
<220>
<223>化学合成的
<400>17
attaccatgtctgtaatgc19
<210>18
<211>539
<212>dna
<213>人工序列
<220>
<223>化学合成的
<220>
<221>misc_feature
<222>(299)..(299)
<223>n可以是g或a
<220>
<221>misc_feature
<222>(364)..(364)
<223>n可以是t或c
<220>
<221>misc_feature
<222>(439)..(439)
<223>n可以是t或c
<220>
<221>misc_feature
<222>(457)..(457)
<223>n可以是g或a
<400>18
aactgagttctgtacgccagagatttgcccgaccatgaccgcaggtccaaagtaagtctt60
gctattgcacatttgcacgattaacggtttcttatatagaagatacatgattcttgaatt120
tatgtaaataaaacttgacagatatgaataccgatgggctgatggtgtgcaaatcaagaa180
gcctattgaagtttcggctccaaagtacgtagagttcttgatggattggattgagtcaca240
attggatgacgagtccatctttcctcaacgacttggtaattagttaattaccatgtctnt300
aatgcatcatttaataaagctttttcgaggcgtttaggaaactgaaatagtaattttcga360
ttgncgtgcaggagcgccatttcccgccaattttagggacgttgtgaaaacgatatttaa420
acgcttgtttcgtgtatangcgcatatctaccacacncattttcagaagattgtgagtct480
taaagaagaagcccatctaaacacttgtttcaagcatttcatattgtttacatgtgtaa539
<210>19
<211>20
<212>dna
<213>人工序列
<220>
<223>化学合成的-ht57f
<400>19
gcgattattgttatggacgc20
<210>20
<211>20
<212>dna
<213>人工序列
<220>
<223>化学合成的-ht57r
<400>20
agcggaaactgttcttgttg20
<210>21
<211>18
<212>dna
<213>人工序列
<220>
<223>化学合成的-ht64f
<400>21
ttattctcggcttccggt18
<210>22
<211>18
<212>dna
<213>人工序列
<220>
<223>化学合成的-ht64r
<400>22
cgggaagatcggctttag18
<210>23
<211>20
<212>dna
<213>人工序列
<220>
<223>化学合成的-ht131f
<400>23
cgtaacatgcaagttgtgga20
<210>24
<211>20
<212>dna
<213>人工序列
<220>
<223>化学合成的-ht131r
<400>24
tgtactctaaacgggcaacc20
<210>25
<211>20
<212>dna
<213>人工序列
<220>
<223>化学合成的-ht134f
<400>25
agtcatgcttgaaggagctg20
<210>26
<211>20
<212>dna
<213>人工序列
<220>
<223>化学合成的-ht134r
<400>26
ctctgtcagcttgcaatgaa20
<210>27
<211>20
<212>dna
<213>人工序列
<220>
<223>化学合成的-ht210f
<400>27
ctaaaactgtcgcaagggaa20
<210>28
<211>20
<212>dna
<213>人工序列
<220>
<223>化学合成的-ht210r
<400>28
cctccatcaatggtaagcac20
<210>29
<211>21
<212>dna
<213>人工序列
<220>
<223>化学合成的-zvg76snpf
<400>29
tccaactcatgaacggactct21
<210>30
<211>20
<212>dna
<213>人工序列
<220>
<223>化学合成的-zvg77snpf
<400>30
ttggtgactcttgcagcatc20
<210>31
<211>20
<212>dna
<213>人工序列
<220>
<223>化学合成的-zvg77snpr
<400>31
aagtttaaaaccgcgtcgtg20
<210>32
<211>20
<212>dna
<213>人工序列
<220>
<223>化学合成的-zvg78snpf
<400>32
tatgagcctcttcggtctcg20
<210>33
<211>20
<212>dna
<213>人工序列
<220>
<223>化学合成的-zvg78snpr
<400>33
caccttattcagccccgata20
<210>34
<211>20
<212>dna
<213>人工序列
<220>
<223>化学合成的-ha0953f-hex
<400>34
caaaccaacaaccaccatca20
<210>35
<211>20
<212>dna
<213>人工序列
<220>
<223>化学合成的-ha0953r
<400>35
aaacgacaccgatgagaacc20
<210>36
<211>26
<212>dna
<213>人工序列
<220>
<223>化学合成的-ha1909f-fam
<400>36
ctgagtttcgtgtaccatttctattg26
<210>37
<211>22
<212>dna
<213>人工序列
<220>
<223>化学合成的-ha1909r
<400>37
acaccaatcagtgggtttcatc22
<210>38
<211>929
<212>dna
<213>人工序列
<220>
<223>化学合成的-
<400>38
agctcacaaagcacttgagatggcagaggttgatgctgatgatgtggatctacttctttt60
atgcacatcgaccccggatgatctatttggtagtgctccacaggtattgcttcaaattct120
tgcacaagatagatgtttgtcctgagaaaaaaactgttgcagtatttatggaattaaaaa180
agtatttgtgttgtatatagaaatatttgactgcatataattttcttgttgaataattgg240
caatgattttaacattttttttcttatatggtaatctcaacgcgttcctgaatttttgta300
agtttctactggctgtagactttagatttctacagttctacagtttgatcatgatccaat360
tttcaattttttttttttttttaaagattaatctgtgtacattataggtgaaaaaatggg420
ccagaggggtggatttttttaacatgttaaaaatggggaactttttatacggatcgaatc480
atcaaaatgggttgggttcacccgaaacacttcctctccaaatattgtattaataaatag540
ttgtatgattacaaacattaaatattttcaacaatattttttaacaaattggcttatgag600
gtttattgcttaacgttttactttgggtaattttttccggttgacccgtttgactcattt660
tctctttaactgctttttaacccacccatttgaccttttagaaaaaaaaaaaaaaaaaac720
ataacccgaatattggtgtatgttaactgccacctctagtattttaagaaaatctacatt780
aattttgaaaatgaaacaagttacttaatttgaacttgtaatcagatacaagccgcactc840
gggtgcaagggaaatccattggcgtttgatattacagctgcttgtagtggatttgttttg900
ggtctagtctcagcttcgtgttatatccg929
<210>39
<211>929
<212>dna
<213>人工序列
<220>
<223>化学合成的-
<400>39
agctcacaaagcacttgagatggcagaggttgatgctgatgatgtggatctacttctttt60
atgcacatcgaccccggatgatctatttggtagtgctccacaggtattgcttcaaattct120
cgcacaagatagatgtttgtcctgagaaaaaaactgttgcagtatttatggaattaaaaa180
agtatttgtgttgtatatagaaatatttgactgcatataattttcttgttgaataattgg240
caatgattttaacattttttttcttatatggtaatctcaacgcgttcctgaatttttgta300
agtttctactggctgtagactttagatttctacagttctacagtttgatcatgatccaat360
tttcaattttttttttttttttaaagattaatctgtgtacattataggtgaaaaaatggg420
ccagaggggtggatttttttaacatgttaaaaatggggaactttttatacggatcgaatc480
atcaaaatgggttgggttcacccgaaacacttcctctccaaatattgtattaataaatag540
ttgtatgattacaaacattaaatattttcaacaatattttttaacaaattggcttatgag600
gtttattgcttaacgttttactttgggcaattttttccggttgacccgtttgactcattt660
tctctttaactgctttttaacccacccatttgaccttttagaaaaaaaaaaaaaaaaaac720
ataacccgaatattggtgtatgttaactgccacctctagtattttaagaaaatctacatt780
aattttgaaaatgaaacaagttacttaatttgaacttgtaatcagatacaagccgcactc840
gggtgcaagggaaatccattggcgtttgatattacagctgcttgtagtggatttgttttg900
ggtctagtctcagcttcgtgttatatccg929
<210>40
<211>982
<212>dna
<213>人工序列
<220>
<223>化学合成的
<400>40
agctcacaaagcacttgagatggcagaggttgatgctgatgatgtggatctacttctttt60
atgcacatcgaccccggatgatctatttggtagtgctccacaggtattgcttcaaattct120
tgcagaagatagatgttttcaaatgtgcctaaaaaggtttcattttttaattttagtggg180
atgtcctgagaaaaaaattgttgcagtatttatggaattaaaaaagtatttgtgttgtat240
atagaaatatttgactgcatataattttcttgttgaataattggcaatgattttaacatt300
ttttttcttatatggtaatctcaacgcgttcctgaatttttgtatgtttctaccggctgt360
agactttagatttctacagtttgatcatgatccaattttcaaaagttttttttttaaaaa420
gattaatctgtgtacattataggtgaaaaaatgggccggaggggtggattcttttaagtt480
ttaacatgttaaatatgggtaactttttatacggatcgaaacatcaaaatgggttgggtt540
cacctgaaacacttcctctcgaaatattctattaataaatagttgtatgattacaaacat600
taaatattttcaacaatatttttttaataaattggcttatgaggtttattgcttaaagtt660
tcactttgggtatttttttccggttgacccgtttgactcattttttctttaactgctttt720
taacccacccatttgaccttttagaaaaaaaacacataacccgaatagatcttttcataa780
gtaaattggtgtatgtaaaccgccacctctagtattttaagaaaatctacattaattttg840
aaaatgaaacgagttacttaatttgaacttgtaatcagatacaagccgcactcgggtgca900
agggaaatccattggcatttgatattacagctgcttgtagtggatttgttttgggtctag960
tctcagcttcgtgttatatccg982
<210>41
<211>983
<212>dna
<213>人工序列
<220>
<223>化学合成的-
<400>41
agctcacaaagcacttgagatggcagaggttgatgctgatgatgtggatctacttctttt60
atgcacatcgaccccggatgatctatttggtagtgctccacaggtattgcttcaaattct120
tgcagaagatagatgttttcaaatgtgcctaaaaaggtttcatttttttaattttagtgg180
gatgtcctgagaaaaaaattgttgcagtatttatggaattaaaaaagtatttgtgttgta240
tatagaaatatttgactgcatataattttcttgttgaataattggcaatgattttaacat300
tttttttcttatatggtaatctcaacgcgttcctgaatttttgtatgtttctaccggctg360
tagactttagatttctacagtttgatcatgatccaattttcaaaagttttttttttaaaa420
agattaatctgtgtacattataggtgaaaaaatgggccggaggggtggattcttttaagt480
tttaacatgttaaatatgggtaactttttatacggatcgaaacatcaaaatgggttgggt540
tcacccgaaacacttcctctcgaaatattctattaataaatagttgtatgattacaaaca600
ttaaatattttcaacaatatttttttaataaattggcttatgaggtttattgcttaaagt660
ttcactttgggtatttttttccggttgacccgtttgactcattttttctttaactgcttt720
ttaacccacccatttgaccttttagaaaaaaaacacataacccgaatagatcttttcata780
agtaaattggtgtatgtaaaccgccacctctagtattttaagaaaatctacattaatttt840
gaaaatgaaacaagttacttaatttgaacttgtaatcagatacaagccgcactcgggtgc900
aagggaaatccattggcatttgatattacagctgcttgtagtggatttgttttgggtcta960
gtctcagcttcgtgttatatccg983
1
- 还没有人留言评论。精彩留言会获得点赞!