一种热稳定性改良的木聚糖酶xyn-LVK及其基因的制作方法
本发明涉及基因工程技术领域,特别是涉及一种热稳定性改良的木聚糖酶xyn-LVK及其基因。
背景技术:
木聚糖酶在许多工业中被广泛应用,如饲料行业、造纸行业和食品行业。然而,在这些应用中,往往需要酶具有良好的耐热性,宽广的pH和较高的比活。所以,人们不断去寻找新的具有优良性能的可用木聚糖酶,并通过随机突变或者理性设计方法来提高它的性能。其中,来源于瘤胃真菌(Neocallimastix patriciarum)的木聚糖酶催化结构域xyn-CD,通过易错PCR将其随机突变得到的重组木聚糖酶xyn-CDBFV(其氨基酸序列如SEQ ID NO.5所示,基因序列如SEQ ID NO.6所示)拥有较高的酶活、较宽的pH和良好的耐热性。因此,重组木聚糖酶xyn-CDBFV在工业产品中具有广泛的应用前景。目前,Cheng, Y.S等详细解析了该蛋白的结构,并结合实验提出了该蛋白的耐热因素(Cheng,Y.S. et al.(2014)Structural Analysis of a Glycoside Hydrolase Family 11 Xylanase from Neocallimastix patriciarum: Insights into the Molecular Basis of a Thermophilic Enzyme. J. Biol. Chem. )。xyn-CDBFV具有11家族木聚糖酶典型的β-jelly-roll折叠结构,但是xyn-CDBFV具有延伸出来的N端结构域(N-terminal region,NTR),这在11家族木聚糖酶结构中是罕见的。NTR是依靠堆积相互作用、氢键和一个二硫键稳定的附着在xyn-CDBFV的催化中心,而不是做为一个灵活的片段自由地悬挂在外面。Cheng,Y.S等的研究表明:仅删除由11个氨基酸组成的NTR后,xyn-CDBFV突变株的酶活在55℃和75℃分别降为原来的61.5%和19.5%;仅删除由NTR和催化中心形成的二硫键后,xyn-CDBFV突变株的酶活在55℃和75℃分别降为原来的86.8%和23.3%。由此可见,NTR在该酶热稳定性中有着重要作用,尤其是NTR和催化中心形成的二硫键更是起着关键作用,在二硫键的作用下,NTR像个夹子一样夹住催化中心,使它在高温下能稳定。
中国专利CN102757947公开了一种热稳定性改良的木聚糖酶xyn-CDBFV-m及其基因,其中,对木聚糖酶xyn-CDBFV的改良之一是将33位的丙氨酸和58位的苯丙氨酸分别替换为半胱氨酸,并称其是在木聚糖酶xyn-CDBFV的N端引入二硫键。但随着Cheng,Y.S等解析了木聚糖酶xyn-CDBFV蛋白的晶体结构,发现其NTR仅由11个氨基酸组成,因此,对于中国专利CN102757947中公开的的N端引入二硫键提高木聚糖酶xyn-CDBFV的热稳定性的结果值得怀疑,因为33位的丙氨酸和58位的苯丙氨酸并非位于木聚糖酶xyn-CDBFV的N端结构域。
技术实现要素:
在了解了NTR和催化中心形成的二硫键的重要性后,本发明的目的是通过改造木聚糖酶xyn-CDBFV,使改造得到的木聚糖酶xyn-LVK在高温时的稳定性大大提高。
具体地,本发明通过Disulfide by Design软件对xyn-CDBFV上的二硫键进行了在线分析预测。经分析,存在26个可以形成二硫键的潜在位点。它们分别是:4位/172位;7位/217位;7位/218位;11位/45位;28位/65位;29位/42位;50位/60位;55位/58位;57位/204位;61位/198位;70位/74位;83位/172位;83位/218位;91位/208位;95位/117位;95位/118位;97位/115位;100位/200位;102位/195位;129位/145位;131位/143位;141位/164位;176位/179位;184位/189位;199位/219位;205位/208位。其中,NTR和催化中心之间除了已形成的4位/172位二硫键外,还有7位/217位和11位/45位可以设计形成二硫键。结合预测结果,理性的在NTR和催化中心之间设计引入7位/217位和11位/45位二硫键,这样在木聚糖酶xyn-LVK中,NTR和催化中心之间将有三个二硫键,它们使NTR这个“夹子”更稳固的夹住催化中心,使木聚糖酶xyn-LVK在高温下更加稳定。
本发明的一方面是提供一种热稳定性改良的木聚糖酶xyn-LVK,其氨基酸序列如SEQ ID NO.1所示。所述SEQ ID NO.1的序列如下:
QSFCSSCSHSCQSVKVTGNKVGTIGGVGYELWADSGNNSATFYScGSFSCTFQNAGDYLCRSGLSFDSTKTPSQIGRMKADFKLVKQNSSNVGYSYVGVYGWTRSPLVEYYIVDNWLSPFPPGDWVGNKKHGSFTIDGAQYTVYENTRTGPSIDGDTTFNQYFSIRQQARDCGTIDISAHFDQWEKLGMTMGKLHEAKVLGEAGNVNGGASGTADFcYAKVYIGD。
本发明中,将SEQ ID NO.1所示的木聚糖酶的氨基酸序列命名为木聚糖酶xyn-LVK。
本发明的木聚糖酶xyn-LVK和原有木聚糖酶xyn-CDBFV相比,有4个氨基酸发生了突变,突变位点分别是A7C,G11C,D45C,P217C。
本发明的另一方面还涉及一种热稳定性改良的木聚糖酶基因xyn-LVK,其编码上述木聚糖酶xyn-LVK,其核苷酸序列如SEQ ID NO.2所示。所述SEQ ID NO.2的序列如下:
CAAAGTTTCTGTAGTTCATGTTCTCACTCTTGTCAAAGTGTAAAGGTAACCGGCAACAAGGTTGGAACTATTGGTGGTGTTGGTTACGAATTATGGGCTGATAGTGGTAATAACAGTGCTACTTTCTATTCTTGTGGTTCCTTCTCATGTACTTTCCAAAATGCTGGGGATTACTTATGTCGTAGTGGTCTTTCTTTCGATAGTACTAAGACCCCATCTCAAATTGGTCGTATGAAGGCTGATTTCAAACTTGTCAAACAAAATAGTTCCAATGTTGGTTATTCCTATGTTGGTGTTTACGGTTGGACTAGAAGTCCACTTGTCGAATACTACATTGTCGATAATTGGCTTAGTCCATTCCCACCAGGTGATTGGGTTGGTAACAAGAAGCATGGTTCTTTCACTATTGATGGTGCTCAATACACTGTTTATGAAAACACTCGTACTGGTCCATCTATTGATGGTGATACCACCTTCAATCAATACTTTAGTATTCGTCAACAAGCTCGTGATTGTGGTACCATTGATATTTCTGCTCACTTTGATCAATGGGAAAAGCTTGGTATGACTATGGGTAAATTACATGAAGCCAAGGTTTTAGGTGAAGCCGGTAACGTTAACGGTGGTGCCAGTGGTACTGCTGATTTCTGTTACGCAAAGGTTTACATTGGTGATTAA。
本发明的另一方面还涉及包含木聚糖酶基因xyn-LVK的表达载体,将本发明的木聚糖酶基因xyn-LVK插入到质粒pPIC9K的信号肽序列的下游并受其调控,得到重组表达质粒pPIC9K-xyn-LVK。
本发明的另一方面还涉及一种重组菌株,所述重组菌株含有所述的木聚糖酶基因xyn-LVK或所述的表达载体或所述的重组表达质粒,所述重组菌株为重组大肠杆菌菌株或重组毕赤酵母菌株。
本发明的一个实施方案中,所述重组菌株为重组大肠杆菌DH5α-xyn-LVK。
本发明的另一个实施方案中,所述重组菌株为重组毕赤酵母菌株GS115-xyn-LVK。
本发明的另一方面还涉及一组引物对,所述引物对用于对木聚糖酶基因xyn-LVK的成熟蛋白的编码区进行扩增,所述引物对的两条引物分别为序列CCgGAATTCCAAAGTTTCTGTAGTTCA(SEQ ID NO:3)和ATAAGAATGCGGCCGCTTAATCACCAATGTAAACCTTTGCGTA(SEQ ID NO:4)。
本发明的另一方面还涉及一种产热稳定性改良的木聚糖酶xyn-LVK的毕赤酵母基因工程菌的制备方法,包括以下步骤:
(1)合成木聚糖酶xyn-LVK基因序列;
(2)将所述木聚糖酶xyn-LVK基因序列克隆至pPIC9K中,得到重组表达质粒;
(3)将所述重组表达质粒转化大肠杆菌感受态细胞DH5α,阳性转化子进行DNA测序,测序表明序列正确的转化子用于大量制备所述重组表达质粒;
(4)抽提所述重组表达质粒,用限制性内切酶BglII进行线性化的重组表达质粒电击转化酵母GS115感受态细胞,涂布于组氨酸缺陷型的 RDB 平板,培养2~3天;
(5)用灭过菌的牙签从长有转化子的 RDB 平板上挑取单菌落, 按照编号先点到 MM 平板上,再点到相应编号的MD平板上,将点有转化子的MM、MD 平板置于培养箱中培养1~2天,至菌落长出;
(6)按编号从 MD 平板上挑取转化子接种于装有BMGY培养基的摇瓶中,培养48h后将菌液离心,去上清,改用含有体积分数为0.5%的甲醇的BMMY培养基诱导培养48h后取菌液离心, 取上清液用于酶活性检测,从中筛选出高木聚糖酶活性的转化子,即得到产热稳定性改良的木聚糖酶xyn-LVK的毕赤酵母基因工程菌;
其中,所述木聚糖酶xyn-LVK的氨基酸序列如SEQ ID NO.1所示,所述木聚糖酶基因xyn-LVK的核苷酸序列如SEQ ID NO.2所示。
具体地,所述产热稳定性改良的木聚糖酶xyn-LVK的毕赤酵母基因工程菌的制备方法,包括以下步骤:
(1)合成木聚糖酶xyn-LVK基因序列;
(2)利用PCR技术扩增木聚糖酶xyn-LVK基因序列,然后利用限制性内切酶EcoRI,NotI将所述木聚糖酶xyn-LVK基因序列克隆至pPIC9K中,得到重组表达质粒;
(3)将所述重组表达质粒转化大肠杆菌感受态细胞DH5α,阳性转化子进行DNA测序,测序表明序列正确的转化子用于大量制备所述重组表达质粒;
(4)抽提所述重组表达质粒,将5微克的用限制性内切酶BglII进行线性化的重组表达质粒电击转化酵母GS115感受态细胞,涂布于组氨酸缺陷型的 RDB 平板,30℃培养2~3天;
(5)用灭过菌的牙签从长有转化子的 RDB 平板上挑取单菌落, 按照编号先点到 MM 平板上,再点到相应编号的MD平板上,将点有转化子的MM、MD 平板置于30℃培养箱中培养1~2天,至菌落长出;
(6)按编号从 MD 平板上挑取转化子接种于装有BMGY培养基的摇瓶中,30℃、250rpm摇床培养48h后将菌液3,000×g离心15min,去上清,改用含有体积分数为0.5%的甲醇的BMMY培养基在30℃、260rpm诱导培养48h后取菌液3,000×g 离心 5min,取上清液用于酶活性检测,从中筛选出高木聚糖酶活性的转化子,即得到产热稳定性改良的木聚糖酶xyn-LVK的毕赤酵母基因工程菌。
本发明的有益效果是:本发明通过理性设计,在木聚糖酶xyn-CDBFV的NTR和催化中心之间设计引入7位/217位和11位/45位二硫键,得到重组木聚糖酶xyn-LVK,这样在木聚糖酶xyn-LVK中,NTR和催化中心之间将有三个二硫键,它们使NTR这个“夹子”更稳固的夹住催化中心,使突变酶在高温下更加稳定,将木聚糖酶基因xyn-LVK在毕赤酵母中表达后,与木聚糖酶xyn-CDBFV相比,突变后的酶在高温时的稳定性得到了显著提高,在90℃处理4min 还能保留65%的酶活,处理8min 后还有42%的剩余酶活性。
附图说明
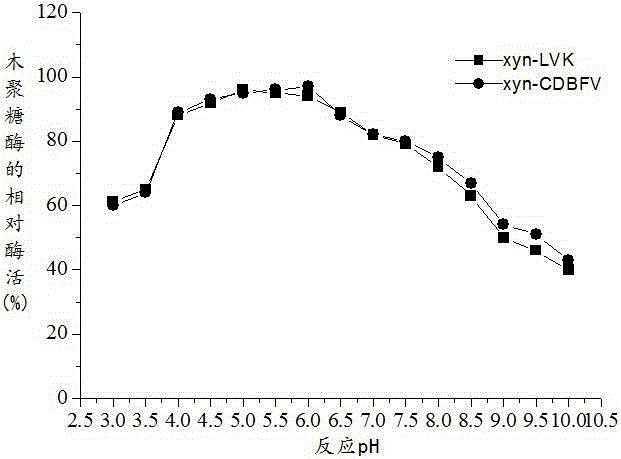
图1为木聚糖酶xyn-CDBFV和xyn-LVK的最适pH;
图2为木聚糖酶xyn-CDBFV和xyn-LVK的pH稳定性;
图3为木聚糖酶xyn-CDBFV和xyn-LVK的最适温度;
图4为木聚糖酶xyn-CDBFV和xyn-LVK的热稳定性。
具体实施方式
下面结合附图及具体实施例对本发明进行进一步描述:
本发明中,木聚糖酶的酶活检测方法参照《GB/T23874-2009饲料添加剂木聚糖酶活力的测定分光光度法》。
实验条件
1、 菌株与载体
毕赤酵母 GS115、载体 pPIC9K 购自 Invitrogen 公司;大肠杆菌DH5α 菌株已在《刘静华,包英华,陈燕飞;大肠杆菌DH5α 感受态细胞转化率的改进,韶关学院学报,2008,29(03) :87-90》文献中公开,大肠杆菌DH5α 菌株的感受态细胞可通过供应商天根生化科技( 北京) 有限公司购买,货号CB101-03。
2、酶类及其他生化试剂
内切酶、连接酶购自TaKaRa公司,其它都为国产试剂。
3、培养基
大肠杆菌培养基为LB(1%蛋白胨,0.5%酵母提取物,1%NaCl,pH7.0)。
酵母培养基为YPD(1%酵母提取物,2%蛋白胨,2%葡萄糖)。酵母筛选培养基为MD(2%葡萄糖,1.5%琼脂粉,0.00004%生物素,1.34%YNB)、MM(0.5%甲醇(体积分数),1.5%琼脂粉,0.00004%生物素,1.34%YNB)、RDB(1M山梨醇,2%葡萄糖,0.00004%生物素,1.34%YNB ,0.005%氨基酸)。酵母诱导培养基 BMGY(1%酵母提取物、2%蛋白胨、1.34%YNB、0.00004%生物素、1%甘油(体积分数))和BMMY(除以0.5%甲醇(体积分数)代替甘油,其余成份相与 BMGY 相同)。
实施例1:木聚糖酶xyn-LVK和木聚糖酶xyn-CDBFV 在毕赤酵母中的高效表达
(1)表达载体的构建及其在酵母的表达
以合成的木聚糖酶xyn-LVK基因为模板,设计合成了带有EcoRI和NotI限制性酶切位点的引物xyn-LVK-F 和 xyn-LVK-R,对xyn-LVK的成熟蛋白的编码区进行扩增,并利用EcoRI和NotI酶切PCR产物,连接进入表达载体 pPIC9K,使木聚糖酶xyn-LVK基因插入到上述表达载体的信号肽序列的下游,与信号肽形成正确的阅读框架,构建成酵母表达载体 pPIC9K-Xyn-LVK,转化大肠杆菌感受态细胞DH5α。阳性转化子进行DNA测序,测序表明序列正确的转化子用于大量制备重组质粒。约5微克的用限制性内切酶BglII进行线性化的重组表达质粒,电击转化酵母GS115感受态细胞,涂布于组氨酸缺陷型的 RDB 平板,30℃培养2-3天,挑取在RDB平板上生长的转化子进行进一步的表达实验。
酵母表达引物序列如下 :
xyn-LVK-F:CCGGAATTCCAAAGTTTCTGTAGTTCA(下划线为EcoRI酶切位点)
xyn-LVK-R:ATAAGAATGCGGCCGCTTAATCACCAATGTAAACCTTTGCGTA(下划线为NotI酶切位点)
(2)高木聚糖酶活性转化子的筛选
用灭过菌的牙签从长有转化子的 RDB 平板上挑取单菌落, 按照编号先点到 MM 平板上,再点到相应编号的MD平板上。将点有转化子的MM、MD 平板置于30℃培养箱中培养1~2天,至菌落长出。按编号从 MD 平板上挑取转化子接种于装有BMGY培养基的摇瓶中,30℃、250rpm摇床培养约48h;将摇瓶中培养48h的菌液3,000×g离心15min,去上清,改用含有体积分数为0.5%的甲醇的BMMY培养基,在30℃、260rpm诱导培养;诱导培养48h后,取菌液,3,000×g 离心 5min,取上清液用于酶活性检测,从中筛选出高木聚糖酶活性的转化子。
(3)高木聚糖酶活性转化子在发酵罐水平的小试表达
高细胞密度发酵用5L发酵罐进行实验,按照Invirogen公司的毕赤酵母发酵过程指导(Pichia Fermentation Process Guidelines)操作。将上述获得的高木聚糖酶活性转化子进行发酵水平高密度发酵试验。随着甲醇诱导时间的延长,发酵上清液中木聚糖酶酶活力显著增加,诱导180h后木聚糖酶活性可达72325 U/mL,菌体湿重达413g/L。
木聚糖酶 xyn-CDBFV在毕赤酵母中的表达方法同木聚糖酶xyn-LVK在毕赤酵母中的表达方法和表达引物一样,不同的是以合成的木聚糖酶xyn-CDBFV基因为模板。随着甲醇诱导时间的延长,发酵上清液中木聚糖酶酶活力显著增加,诱导180h后木聚糖酶活性可达68471 U/mL,菌体湿重达398g/L。
实施例2:木聚糖酶xyn-LVK和木聚糖酶xyn-CDBFV的纯化
木聚糖酶xyn-LVK纯化的具体过程是,将实施例1中发酵罐表达的上清液用超滤膜进行浓缩,然后在每100mL的浓缩液中缓慢加入质量分数为70%的硫酸铵溶液,并不断搅拌2.5h,然后在10000rpm离心15min,收集沉淀。沉淀用少量pH8.0的20mM Tris-HCl缓冲液溶解,并在大量该缓冲液中透析过夜,透析后的溶液过阴离子柱(HiTrap Q Sepharose XL 5mL)进行纯化。阴离子柱事先用pH8.0的20mM Tris-HCl缓冲液平衡,上样后用0~1.0mol/L线性梯度的NaCl溶液洗脱,收集有木聚糖酶活性的洗脱管,收集液再用超滤离心管处理,使其浓缩,经SDS-PAGE确认后用于酶学性质的分析。木聚糖酶xyn-CDBFV的纯化方法和木聚糖酶xyn-LVK的纯化方法基本一样。
实施例3:木聚糖酶xyn-LVK和木聚糖酶xyn-CDBFV的酶学性质分析
(1)木聚糖酶xyn-LVK和木聚糖酶xyn-CDBFV的最适pH和pH稳定性
将纯化的木聚糖酶xyn-LVK和木聚糖酶xyn-CDBFV稀释到所需浓度,将样品置于不同pH值下进行酶促反应以测定其最适pH,以最适pH值下的酶活作为100%对照,计算相对酶活,并作相对酶活曲线,结果如图1所示。由图1可知,木聚糖酶xyn-LVK和xyn-CDBFV的最适pH 曲线有所改变,xyn-LVK 最适pH 比xyn-CDBFV低,由原先的6.0降低到了5.0。
将纯化的木聚糖酶xyn-LVK和木聚糖酶xyn-CDBFV稀释到所需浓度,将样品置于不同pH值的缓冲液中,室温放置24h,然后检测其酶活,以未处理的原酶液作为100%对照,作相对酶活曲线,结果如图2所示。由图2可知,木聚糖酶xyn-LVK和xyn-CDBFV的pH 稳定性曲线基本没有改变。
(2)木聚糖酶xyn-LVK和木聚糖酶xyn-CDBFV的最适温度和热稳定性
将纯化的木聚糖酶xyn-LVK和木聚糖酶xyn-CDBFV稀释到所需浓度,将样品置于40℃、45℃、50℃、55℃、60℃、65℃、70℃、75℃、80℃、85℃、90℃、95℃、100℃温度下测定酶活,以确定该酶的最适温度,以最适温度下的酶活作为100%对照,计算相对酶活,并作相对酶活曲线,结果如图3所示。由图3可知, xyn-LVK 的最适温度有了明显的提高,从60℃提高到了75℃,并且在90℃时的相对酶活仍然能够保持在60%左右。
将纯化的木聚糖酶xyn-LVK和木聚糖酶xyn-CDBFV稀释到所需浓度,将样品置于90℃的烘箱中,分别在2 min、4 min、6 min、8 min、10 min、15min、20 min取出酶液,冷却至室温,然后检测其酶活,以未处理的原酶液作为100%对照,作相对酶活曲线,结果如图4所示。由图4可知,xyn-LVK 在90℃的热稳定性有了明显的提高,在90℃处理4min 还能保留65%的酶活,处理8min 后还有42%的剩余酶活性。而xyn-CDBFV 在90℃处理2min 酶活性就损失了85%,6min 酶活性完全损失。xyn-LVK 在90℃的稳定性实验结果表明,该木聚糖酶能够抵抗饲料高温制粒时短暂的高温处理。因此,本发明的优化改良的木聚糖酶xyn-LVK 可在饲料工业中显示出巨大的应用潜力。
尽管本发明的具体实施方式已经得到详细的描述,本领域技术人员将会理解,根据已经公开的所有教导,可以对那些细节进行各种修改和替换,这些改变均在本发明的保护范围之内。本发明的全部范围由所附权利要求及其任何等同物给出。
序列表
<110> 深圳市绿微康生物工程有限公司
<120> 一种热稳定性改良的木聚糖酶xyn-LVK及其基因
<130> LVK201501
<160> 6
<170> PatentIn version 3.3
<210> 1
<211> 225
<212> PRT
<213> 瘤胃真菌(Neocallimastix patriciarum)
<400> 1
Gln Ser Phe Cys Ser Ser Cys Ser His Ser Cys Gln Ser Val Lys Val
1 5 10 15
Thr Gly Asn Lys Val Gly Thr Ile Gly Gly Val Gly Tyr Glu Leu Trp
20 25 30
Ala Asp Ser Gly Asn Asn Ser Ala Thr Phe Tyr Ser Cys Gly Ser Phe
35 40 45
Ser Cys Thr Phe Gln Asn Ala Gly Asp Tyr Leu Cys Arg Ser Gly Leu
50 55 60
Ser Phe Asp Ser Thr Lys Thr Pro Ser Gln Ile Gly Arg Met Lys Ala
65 70 75 80
Asp Phe Lys Leu Val Lys Gln Asn Ser Ser Asn Val Gly Tyr Ser Tyr
85 90 95
Val Gly Val Tyr Gly Trp Thr Arg Ser Pro Leu Val Glu Tyr Tyr Ile
100 105 110
Val Asp Asn Trp Leu Ser Pro Phe Pro Pro Gly Asp Trp Val Gly Asn
115 120 125
Lys Lys His Gly Ser Phe Thr Ile Asp Gly Ala Gln Tyr Thr Val Tyr
130 135 140
Glu Asn Thr Arg Thr Gly Pro Ser Ile Asp Gly Asp Thr Thr Phe Asn
145 150 155 160
Gln Tyr Phe Ser Ile Arg Gln Gln Ala Arg Asp Cys Gly Thr Ile Asp
165 170 175
Ile Ser Ala His Phe Asp Gln Trp Glu Lys Leu Gly Met Thr Met Gly
180 185 190
Lys Leu His Glu Ala Lys Val Leu Gly Glu Ala Gly Asn Val Asn Gly
195 200 205
Gly Ala Ser Gly Thr Ala Asp Phe Cys Tyr Ala Lys Val Tyr Ile Gly
210 215 220
Asp
225
<210> 2
<211> 678
<212> DNA
<213> 瘤胃真菌(Neocallimastix patriciarum)
<400> 2
caaagtttct gtagttcatg ttctcactct tgtcaaagtg taaaggtaac cggcaacaag 60
gttggaacta ttggtggtgt tggttacgaa ttatgggctg atagtggtaa taacagtgct 120
actttctatt cttgtggttc cttctcatgt actttccaaa atgctgggga ttacttatgt 180
cgtagtggtc tttctttcga tagtactaag accccatctc aaattggtcg tatgaaggct 240
gatttcaaac ttgtcaaaca aaatagttcc aatgttggtt attcctatgt tggtgtttac 300
ggttggacta gaagtccact tgtcgaatac tacattgtcg ataattggct tagtccattc 360
ccaccaggtg attgggttgg taacaagaag catggttctt tcactattga tggtgctcaa 420
tacactgttt atgaaaacac tcgtactggt ccatctattg atggtgatac caccttcaat 480
caatacttta gtattcgtca acaagctcgt gattgtggta ccattgatat ttctgctcac 540
tttgatcaat gggaaaagct tggtatgact atgggtaaat tacatgaagc caaggtttta 600
ggtgaagccg gtaacgttaa cggtggtgcc agtggtactg ctgatttctg ttacgcaaag 660
gtttacattg gtgattaa 678
<210> 3
<211> 27
<212> DNA
<213> 人工序列
<400> 3
ccggaattcc aaagtttctg tagttca 27
<210> 4
<211> 43
<212> DNA
<213> 人工序列
<400> 4
ataagaatgc ggccgcttaa tcaccaatgt aaacctttgc gta 43
<210> 5
<211> 225
<212> PRT
<213> 瘤胃真菌(Neocallimastix patriciarum)
<400> 5
Gln Ser Phe Cys Ser Ser Ala Ser His Ser Gly Gln Ser Val Lys Val
1 5 10 15
Thr Gly Asn Lys Val Gly Thr Ile Gly Gly Val Gly Tyr Glu Leu Trp
20 25 30
Ala Asp Ser Gly Asn Asn Ser Ala Thr Phe Tyr Ser Asp Gly Ser Phe
35 40 45
Ser Cys Thr Phe Gln Asn Ala Gly Asp Tyr Leu Cys Arg Ser Gly Leu
50 55 60
Ser Phe Asp Ser Thr Lys Thr Pro Ser Gln Ile Gly Arg Met Lys Ala
65 70 75 80
Asp Phe Lys Leu Val Lys Gln Asn Ser Ser Asn Val Gly Tyr Ser Tyr
85 90 95
Val Gly Val Tyr Gly Trp Thr Arg Ser Pro Leu Val Glu Tyr Tyr Ile
100 105 110
Val Asp Asn Trp Leu Ser Pro Phe Pro Pro Gly Asp Trp Val Gly Asn
115 120 125
Lys Lys His Gly Ser Phe Thr Ile Asp Gly Ala Gln Tyr Thr Val Tyr
130 135 140
Glu Asn Thr Arg Thr Gly Pro Ser Ile Asp Gly Asp Thr Thr Phe Asn
145 150 155 160
Gln Tyr Phe Ser Ile Arg Gln Gln Ala Arg Asp Cys Gly Thr Ile Asp
165 170 175
Ile Ser Ala His Phe Asp Gln Trp Glu Lys Leu Gly Met Thr Met Gly
180 185 190
Lys Leu His Glu Ala Lys Val Leu Gly Glu Ala Gly Asn Val Asn Gly
195 200 205
Gly Ala Ser Gly Thr Ala Asp Phe Pro Tyr Ala Lys Val Tyr Ile Gly
210 215 220
Asp
225
<210> 6
<211> 678
<212> DNA
<213> 瘤胃真菌(Neocallimastix patriciarum)
<400> 6
caaagtttct gtagttcagc ttctcactct ggacaaagtg taaaggtaac cggcaacaag 60
gttggaacta ttggtggtgt tggttacgaa ttatgggctg atagtggtaa taacagtgct 120
actttctatt ctgatggttc cttctcatgt actttccaaa atgctgggga ttacttatgt 180
cgtagtggtc tttctttcga tagtactaag accccatctc aaattggtcg tatgaaggct 240
gatttcaaac ttgtcaaaca aaatagttcc aatgttggtt attcctatgt tggtgtttac 300
ggttggacta gaagtccact tgtcgaatac tacattgtcg ataattggct tagtccattc 360
ccaccaggtg attgggttgg taacaagaag catggttctt tcactattga tggtgctcaa 420
tacactgttt atgaaaacac tcgtactggt ccatctattg atggtgatac caccttcaat 480
caatacttta gtattcgtca acaagctcgt gattgtggta ccattgatat ttctgctcac 540
tttgatcaat gggaaaagct tggtatgact atgggtaaat tacatgaagc caaggtttta 600
ggtgaagccg gtaacgttaa cggtggtgcc agtggtactg ctgatttccc atacgcaaag 660
gtttacattg gtgattaa 678
- 还没有人留言评论。精彩留言会获得点赞!