植物抗逆性相关蛋白TabZIP14及其编码基因与应用的制作方法
本发明属于植物基因工程领域,涉及一种植物抗逆性相关蛋白TabZIP14及其编码基因与应用。
背景技术:
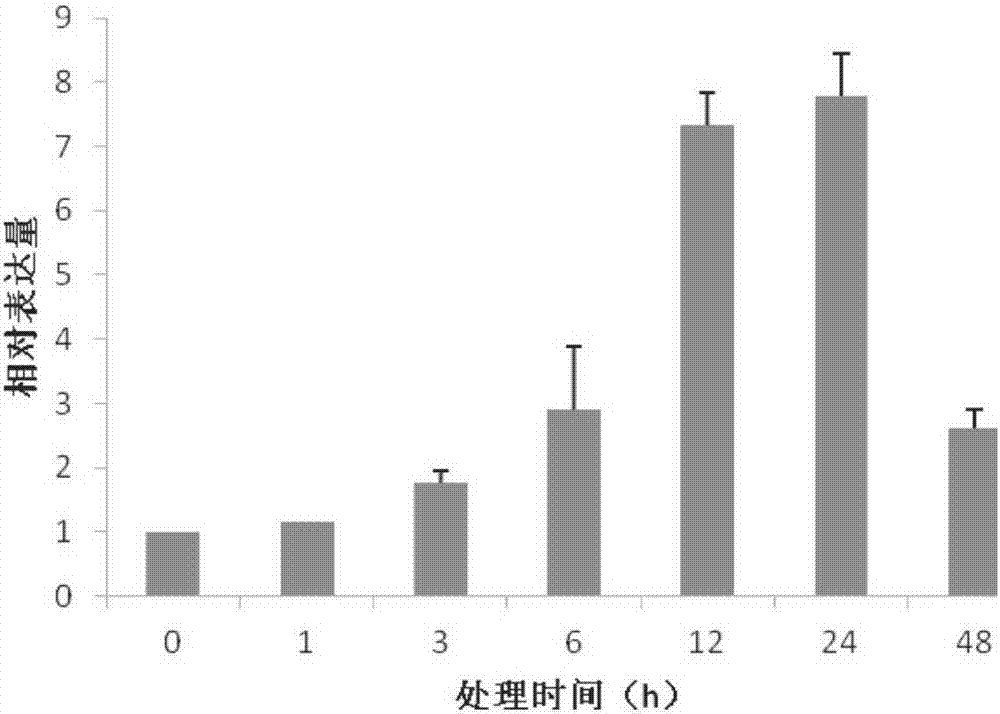
:非生物胁迫是影响作物生长发育和产量的主要因素之一,开发和利用抗逆品种在盐碱地和缺水地区显得尤为重要。最近几年,在提高植物抗逆性的基因方面取得一些进展。转录因子也是响应植物非生物胁迫逆境的关键调节因子,而且过表达一些转录因子基因可以提高植物的抗逆性。bZIP就是存在于植物中的一种转录因子。在逆境胁迫下植物体内会产生一系列应答反应,伴随着许多生理生化及发育上的变化。明确植物对逆境的反应机制,将为抗逆基因工程研究和应用提供科学论据。目前,植物抗逆性研究已逐渐深入到细胞、分子水平,并与遗传学和遗传工程研究相结合,探索用生物技术来改进植物生长特性,其目的是提高植物对逆境的适应能力。因此,利用现代分子生物技术,挖掘抗逆关键基因,通过基因工程改良植物特别是农作物的抗逆性,对于保障国家粮食安全具有重要意义。技术实现要素:本发明的目的是提供一种植物抗逆性相关蛋白TabZIP14及其编码基因与应用。本发明所提供的TabZIP14蛋白来源于小麦(TriticumaestivumL.)品种中国春,具体可为如下(a)或(b):(a)由序列表中序列1所示的氨基酸序列组成的蛋白质;(b)将序列1的氨基酸序列经过一个或几个氨基酸残基的取代和/或缺失和/或添加,且与植物抗逆性相关的由序列1衍生的蛋白质。为了便于上述(a)中所示蛋白质的纯化,可在由序列表中序列1的氨基酸残基序列组成的蛋白质的氨基末端或羧基末端连接上如下表所示的标签。表:标签的序列标签残基序列Poly-Arg5-6(通常为5个)RRRRRPoly-His2-10(通常为6个)HHHHHHFLAG8DYKDDDDKStrep-tagII8WSHPQFEKc-myc10EQKLISEEDL上述(b)中的蛋白质可人工合成,也可先合成其编码基因,再进行生物表达得到。上述(b)中的蛋白质的编码基因可通过将序列表中序列2所示的DNA序列中缺失一个或几个氨基酸残基的密码子,和/或进行一个或几个碱基对的错义突变。编码所述蛋白质的核酸分子也属于本发明的保护范围。所述核酸分子可以是DNA,如cDNA、基因组DNA或重组DNA;所述核酸分子也可以是RNA,如mRNA、hnRNA或tRNA等。在本发明的一个实施例中,所述核酸分子具体为编码所述蛋白质的基因,所述基因具体可为如下1)-4)中任一的DNA分子:1)序列表中序列2所示的DNA分子;2)序列表中序列2的第181-1383位所示的DNA分子;3)在严格条件下与1)或2)限定的DNA分子杂交且编码抗逆性相关蛋白的DNA分子;4)与1)-3)中任一限定的DNA序列具有90%以上同一性,且编码抗逆性相关蛋白的DNA分子。其中,序列2由1576个核苷酸组成,第181-1383位为ORF,编码序列表中序列1所示的蛋白质。含有上述核酸分子的重组载体、表达盒、转基因细胞系或重组微生物也属于本发明的保护范围。所述重组载体可为重组表达载体,也可为重组克隆载体。所述重组表达载体可用现有的植物表达载体构建。所述植物表达载体包括双元农杆菌载体和可用于植物微弹轰击的载体等,如pGreen0029、pCAMBIA3301、pCAMBIA1300、pBI121、pBin19、pCAMBIA2301、pCAMBIA1301-UbiN或其它衍生植物表达载体。所述植物表达载体还可包含外源基因的3’端非翻译区域,即包含聚腺苷酸信号和任何其它参与mRNA加工或基因表达的DNA片段。所述聚腺苷酸信号可引导聚腺苷酸加入到mRNA前体的3’端。使用所述基因构建重组表达载体时,在其转录起始核苷酸前可加上任何一种增强型、组成型、组织特异型或诱导型启动子,例如花椰菜花叶病毒(CAMV)35S启动子、泛素基因Ubiquitin启动子(pUbi)、胁迫诱导型启动子rd29A等,它们可单独使用或与其它的植物启动子结合使用;此外,使用本发明的基因构建重组表达载体时,还可使用增强子,包括翻译增强子或转录增强子,这些增强子区域可以是ATG起始密码子或邻接区域起始密码子等,但必需与编码序列的阅读框相同,以保证整个序列的正确翻译。所述翻译控制信号和起始密码子的来源是广泛的,可以是天然的,也可以是合成的。翻译起始区域可以来自转录起始区域或结构基因。为了便于对转基因植物细胞或植物进行鉴定及筛选,可对所用重组表达载 体进行加工,如加入可在植物中表达的编码可产生颜色变化的酶或发光化合物的基因、具有抗性的抗生素标记物或是抗化学试剂标记基因等。也可不加任何选择性标记基因,直接以逆境筛选转化植株。在本发明中,所述重组表达载体中启动所述基因转录的启动子具体为35S启动子。进一步,所述重组表达载体为将pEarleyGate100载体的attR1和attR2之间的小片段替换为所述基因后得到的重组质粒。更加具体的,所述重组表达载体是将所述基因(序列2的第181-1383位)通过BP反应重组到入门载体上,得到入门质粒;再将入门质粒与目标载体通过LR反应得到的目的重组质粒;所述入门载体具体为pDONR/Zeo(该载体具体为invitrogen公司产品,产品目录号为12535-035);所述目标载体具体为pEarleyGate100(该载体具体为自invitrogen公司产品);所述pEarleyGate100在文献“ZhangL,ZhangL,XiaC,ZhaoG,LiuJ,JiaJ,KongX.AnovelwheatbZIPtranscriptionfactor,TabZIP60,confersmultipleabioticstresstolerancesintransgenicArabidopsis.PhysiolPlant.2015,153(4):538-554.”中公开过。所述表达盒由能够启动所述基因表达的启动子,所述基因,以及转录终止序列组成。所述转基因细胞系为转入所述基因的非繁殖材料。所述蛋白质或所述核酸分子或所述重组载体、表达盒、转基因细胞系或重组微生物在如下任一中的应用也属于本发明的保护范围:(a)调控植物抗逆性;(b)选育抗逆性提高的植物品种。在(a)中,所述调控植物的抗逆性具体体现在:在所述植物体内,若所述基因或所述蛋白的表达量越高,则所述植物的抗逆性越强;若所述基因或所述蛋白的表达量越低,则所述植物的抗逆性越弱。在(b)中,所述选育抗逆性增强的植物品种的方法,具体可包括将所述基因或所述蛋白表达量较高的植株作为亲本进行杂交的步骤。本发明还提供了一种培育抗逆性提高的转基因植物的方法。本发明所提供的培育抗逆性提高的转基因植物的方法,具体可包括如下(a1)和(a2)的步骤:(a1)向受体植物中导入所述蛋白质的编码基因,得到表达所述编码基因的转基因植物;(a2)从步骤(a1)所得转基因植物中得到与所述受体植物相比,抗逆性提高的转基因植物。所述蛋白质在所述转基因植物中的表达量高于所述受体植物;编码所述蛋白质的基因是如下1)-4)中任一的DNA分子:1)序列表中序列2所示的DNA分子;2)序列表中序列2的第181-1383位所示的DNA分子;3)在严格条件下与1)或2)限定的DNA分子杂交且编码抗逆性相关蛋白的DNA分子;4)与1)-3)中任一限定的DNA序列具有90%以上同一性,且编码抗逆性相关蛋白的DNA分子。所述基因具体可通过上述任一所述重组表达载体导入所述受体植物中,得到所述转基因植物。具体可通过使用Ti质粒、Ri质粒、植物病毒载体、直接DNA转化、显微注射、电导、农杆菌介导、基因枪等常规生物学方法将所述重组表达载体转化植物细胞或组织,并将转化的植物组织培育成植株。在本发明中,所述基因是通过重组农杆菌导入的,所述重组农杆菌是将所述重组表达载体导入农杆菌得到的;所述农杆菌具体为农杆菌GV3101-pMP90,该菌在文献“XuZ,KimS,HyeonD,KimD,etal.TheArabidopsisthalianaNACtranscriptionfactorfamily:structure–functionrelationshipsanddeterminantsofANAC019stresssignaling.Biochem.J.2010,426:183-196.”中公开过。在上述应用或方法中,所述抗逆性为如下中的至少一种:抗冻性、抗盐性。在本发明中,所述抗冻性中的“冻”具体体现为-10℃处理3h。在本发明中,所述抗盐性中的“盐”具体体现为150-200mMNaCl处理。在上述应用或方法中,所述植物可为双子叶植物,也可为单子叶植物。如:小麦、烟草或拟南芥等。在本发明中,所述植物为拟南芥,具体为拟南芥Columbia-0亚型。实验证明,将可表达序列表中序列2所示DNA分子(TabZIP14基因)的重组表达载体转化拟南芥得到的转基因植株,与相同条件下的野生型植株相比:在抗冻性实验(-10℃处理3h)中,野生型植株的存活率为16%,转TabZIP14基因的株系的存活率为79%。在抗盐实验中,150mMNaCl处理后,野生型拟南芥的根长明显受到抑制,T3代转TabZIP14基因拟南芥L1、L2和L3株系的根长分别是野生型植株根长的1.4倍、1.3倍和1.4倍,部分野生型的拟南芥叶子呈现白化现象,白化率为37%±0.5%,而T3代转TabZIP14基因拟南芥叶片全部保持绿色;200mMNaCl处理后,T3代转TabZIP14基因拟南芥L1、L2和L3株系的根长比分别是野生型植株根长的1.2倍、1.3倍和1.3倍,野生型植株的叶片几乎全部呈白化现象,白化率为98%±0.4%,而T3代转TabZIP14基因拟南芥叶片基本呈现绿色状态。以上结果说明转TabZIP14基因拟南 芥的抗盐性明显提高。本发明所提供的TabZIP14蛋白及其编码基因在提高植物抗逆性方面具有重要意义,将在培育高抗逆性如强抗冻性和强抗盐性植物品种中发挥重要作用。附图说明图1为PEG胁迫下小麦TabZIP14基因的相对表达量。图2为盐胁迫下小麦TabZIP14基因的相对表达量。图3为低温胁迫下小麦TabZIP14基因的相对表达量。图4为ABA胁迫下小麦TabZIP14基因的相对表达量。图5为T3代转TabZIP14基因拟南芥的RNA水平鉴定结果。图6为低温胁迫下拟南芥的表型。图7为盐胁迫下拟南芥的表型。具体实施方式下述实施例中所使用的实验方法如无特殊说明,均为常规方法。下述实施例中所用的材料、试剂等,如无特殊说明,均可从商业途径得到。KOD-Plus高保真酶:TOYOBO公司产品,产品目录号为KOD-201。入门载体pDONR/Zeo:invitrogen公司产品,产品目录号为12535-035。BPClonaseTMⅡEnzymeMix:invitrogen公司产品,产品目录号为11789-013。目标载体pEarleyGate100:在文献“ZhangL,ZhangL,XiaC,ZhaoG,LiuJ,JiaJ,KongX.AnovelwheatbZIPtranscriptionfactor,TabZIP60,confersmultipleabioticstresstolerancesintransgenicArabidopsis.PhysiolPlant.2015,153(4):538-554.”中公开过,公众可从中国农业科学院作物科学研究所获得。LRClonaseTMⅡEnzymeMix:invitrogen公司产品,产品目录号为11791-019。农杆菌GV3101-pMP90:在文献“XuZ,KimS,HyeonD,KimD,etal.TheArabidopsisthalianaNACtranscriptionfactorfamily:structure–functionrelationshipsanddeterminantsofANAC019stresssignaling.Biochem.J.2010,426:183-196.”中公开过,公众可从中国农业科学院作物科学研究所获得。拟南芥(Arabidopsisthaliana)(Columbia-0亚型):在文献“HeY,LiW,LvJ,JiaY,WangMandXiaG.EctopicexpressionofawheatMYBtranscriptionfactorgene,TaMYB73,improvessalinitystresstoleranceinArabidopsisthaliana.JExpBot,2012,63(3):1511–1522.”中公开过,公众可从中国农业科学院作物科学研究所获得。中国春小麦(ChineseSpringWheat):在文献“KobayashiaF,MaetaaE,TerashimaaA,TakumS.PositiveroleofawheatHvABI5orthologinabioticstressresponseofseedlings.PhysiolPlant,2008,134:74–86.”中公开过,公众可从中国农业科学院作物 科学研究所获得。实施例1、TabZIP14蛋白及其编码基因的发现为了研究小麦bZIP基因在植物抗逆方面的功能,通过生物信息学的方法,从发明人所在实验室已经测序的30000条非冗余的小麦全长cDNA序列中筛选出22个编码小麦bZIP蛋白的基因。通过半定量PCR的方法,对该22个小麦bZIP基因在逆境条件下的表达模式进行研究,筛选出受逆境诱导表达的TabZIP14基因。TabZIP14基因全长cDNA为1576bp,序列如序列表中序列2所示。其开放阅读框为序列2的自5’末端起第181-1383位核苷酸,编码序列表中序列1所示的TabZIP14蛋白,序列1共由400个氨基酸残基组成。实施例2、TabZIP14基因在不同逆境胁迫下的转录水平表达模式一、TabZIP14基因在PEG胁迫下的表达模式对两叶一心期的中国春小麦幼苗置于16.1g/100ml的PEG6000水溶液中进行处理,分别在处理前(0h)、处理后1、3、6、12、24和48小时提取小麦幼苗根组织的RNA,反转录成cDNA,以其cDNA为模板,以上游引物1和下游引物2为引物,进行实时定量PCR分析,得到TabZIP14基因的相对表达量,内参是小麦Tubulin基因。用于检测TabZIP14基因的引物如下:上游引物1:5’-CGGCTTGCTGATGTTAATC-3’;下游引物2:5’-TCGCTTGTGGAGAAGTAAG-3’。用于检测内参Tubulin基因的引物如下:Tubulin-F:5’-TTAGACTTGCGAAGCCAGCA-3’;Tubulin-R:5’-AAATGCCCTTGAGGTTTCCC-3’。结果如图1所示,由图可见TabZIP14基因响应PEG胁迫诱导。二、TabZIP14基因在盐胁迫下的表达模式对两叶一心期的中国春小麦幼苗置于250mMNaCl水溶液中进行处理,分别在处理前(0h)、处理后1、3、6、12、24和48小时提取小麦幼苗根组织的RNA,反转录成cDNA,以其cDNA为模板,以上游引物1和下游引物2为引物,进行实时定量PCR分析,得到TabZIP14基因的相对表达量,内参是小麦Tubulin基因。具体引物序列参见步骤一。结果如图2所示,由图可见TabZIP14基因响应盐胁迫诱导。三、TabZIP14基因在低温胁迫下的表达模式对两叶一心期的中国春小麦幼苗置于4℃处理,分别在处理前(0h)、处理后1、3、6、12、24和48小时提取小麦幼苗叶子组织的RNA,反转录成cDNA,以其cDNA为模板,进行实时定量PCR分析,得到TabZIP14基因的相对表达量,内参是小麦Tubulin基因。具体引物序列参见步骤一。结果如图3所示,由图可见TabZIP14基因响应低温胁迫诱导。四、TabZIP14基因在ABA胁迫下的表达模式对两叶一心期的中国春小麦幼苗置于200μMABA水溶液中进行处理,分别在处理前(0h)、处理后1、3、6、12、24和48小时提取小麦幼苗根组织的RNA,反转录成cDNA,以其cDNA为模板,以上游引物1和下游引物2为引物,进行实时定量PCR分析,得到TabZIP14基因的相对表达量,内参是小麦Tubulin基因。具体引物序列参见步骤一。结果如图4所示,由图可见TabZIP14基因响应ABA诱导表达。实施例3、转TabZIP14基因植株的获得和耐逆性鉴定一、利用Gateway技术构建过表达载体1、attB引物设计上游引物3:5’-GCATGGAGCGCGGCGTCTTC-3’下游引物4:5’-CTACGAGGTCGATCCGGATG-3’2、根据Gateway技术构建表达载体的需要,在上游引物3和下游引物4的5'端分别加入attB1和attB2重组位点(下划线标注部分为attB1和attB2重组位点),分别得到上游引物5和下游引物6。上游引物5:5’-GGGGACAAGTTTGTACAAAAAAGCAGGCTGCATGGAGCGCGGCGTCTTC-3’下游引物6:5’-GGGGACCACTTTGTACAAGAAAGCTGGGTCTACGAGGTCGATCCGGATG-3’3、以中国春小麦的cDNA为模板,用上游引物5和下游引物6进行PCR扩增,得到attBPCR产物,为保证PCR过程的保真性,使用KOD-Plus高保真酶进行PCR反应,PCR反应体系和反应程序如下:4、按照如下体系和程序进行BP重组反应,得到BP反应产物:5、入门质粒的获得将2.5μlBP反应产物加入50μlTOP10感受态细胞进行转化,冰浴30min,42℃热击90s,然后迅速置于冰上2min。加入500μlLB培养液,37℃,210rpm/min复苏50min,复苏液均匀涂布于加有Zeocin(浓度为40μg/ml)的固体LB选择培养基表面,37℃倒置培养过夜。pDONRTM/Zeo载体自身带有致死基因,不能在培养基上生存。通过步骤4的BP重组反应目标基因片段会取代致死基因,最终得到的克隆即为入门克隆(entryclone),质粒为入门质粒,将入门质粒送测序,结果正确。6、按照如下体系和程序进行LR重组反应,得到LR反应产物:7、目标质粒的获得转化过程同步骤5,只是将zeocin替换为卡那霉素(浓度为50μg/ml)。pEarleyGate100载体同样带有致死基因,通过LR重组反应致死基因将会被目标基因片段取代,得到的克隆即为目标克隆(destinationclone),质粒为目标质粒,将目标质粒命名为pEarleyGate100-TabZIP14。pEarleyGate100-TabZIP14的测序结果表明pEarleyGate100-TabZIP14含有序列表中序列2的第181-1383位所示DNA片段。pEarleyGate100-TabZIP14为TabZIP14基因表达载体,启动子是花椰菜花叶病毒35S启动子。二、转TabZIP14基因拟南芥的获得1、将步骤一获得的目标质粒转化农杆菌GV3101-pMP90,得到重组农杆菌。2、拟南芥侵染及转基因植株筛选及鉴定(1)拟南芥的培养将拟南芥(Arabidopsisthaliana)(Columbia-0亚型)种子用灭菌水(含有体积百分含量为10%次氯酸钠以及10%吐温-20的水溶液)摇动消毒15min,在超净工作台中用灭菌水清洗上述消毒的种子至少5次。将清洗完的种子均匀播种在MS培养基上。MS平板于4℃春化3d,然后置于22℃光照培养箱培养一周。待小苗长出四片真叶后移栽至营养钵中培养,保湿2-3d。拟南芥的生长对温度比较敏感,20-22℃为比较适宜的培养温度。当拟南芥植株生长至大部分花蕾处于即将开花状态时,进行农杆菌侵染。(2)拟南芥的侵染采用沾花法用步骤1所得的重组农杆菌对步骤(1)得到的拟南芥进行侵染,将侵染过的拟南芥平放于托盘中,黑暗保湿培养24h,然后放于正常培养条件下培养。拟南芥侵染1周后根据拟南芥的生长状态可再次侵染以提高转化效率,得到转TabZIP14基因拟南芥。(3)阳性转TabZIP14基因拟南芥的初步筛选①收集转TabZIP14基因拟南芥T0代种子,于37℃烘箱中烘干(6-8d),然后4℃春化3d。②将种子直接撒播在营养钵中,22℃保湿培养1周。③待小苗长出4片真叶后,通过喷洒除草剂(basta)进行阳性转TabZIP14基因拟南芥筛选,非阳性转TabZIP14基因拟南芥在喷洒3天后开始出现萎蔫并停止生长,2周后基本死亡。为了将非阳性转TabZIP14基因拟南芥苗彻底除净,可连续喷洒2-3次,每次间隔2-3d。(4)阳性转TabZIP14基因拟南芥的鉴定A.DNA水平鉴定CTAB法提取步骤(3)初步筛选获得的阳性转TabZIP14基因拟南芥叶片的基因组DNA,以其为模板,以TabZIP14基因特异性的正向引物7和反向引物8进行PCR扩增,得到PCR扩增产物,将PCR扩增产物进行琼脂糖凝胶电泳。正向引物7:5’-CAGAAGATGATGATATGGAAGG-3’;反向引物8:5’-TCGCTTGTGGAGAAGTAAG-3’。电泳结果表明,步骤(3)初步筛选获得的阳性转TabZIP14基因拟南芥在PCR扩增之后有约445bp的目的条带,进一步确定为转TabZIP14基因拟南芥。将野生型拟南芥进行上述鉴定实验无目的条带。用上述方法进行鉴定,直至获得T3代纯合转TabZIP14基因拟南芥(以下简称 T3代转TabZIP14基因拟南芥),从所得T3代转TabZIP14基因拟南芥中随机选取3各株系,记为L1、L2和L3。B.RNA水平鉴定对basta除草剂筛选获得的T3代纯合转基因植株,随机选取三株,提取总RNA,并进行第一链cDNA的合成,以上游引物1和下游引物2为引物,进行实时定量PCR分析,得到TabZIP14基因的相对表达量,内参是小麦Tubulin基因。具体引物序列参见实施例2步骤一。结果显示:转TabZIP14基因的T3代纯合植株,利用RT-qPCR检测,结果表明其纯合株系L1,L2和L3中TabZIP14基因的表达比较相近。可以继续后续功能鉴定。转TabZIP14基因的L1、L2和L3株系的RNA水平鉴定结果具体如图5所示。三、转TabZIP14基因拟南芥的抗逆鉴定1、抗冻性分析将野生型拟南芥(WT)以及三个T3代转TabZIP14基因拟南芥(L1,L2,L3)的种子在22℃,12h光照下进行培养,得到3周幼苗,然后将各幼苗在-10℃处理3小时,再恢复培养4天。观察各组拟南芥冷处理前后的幼苗生长状态并统计其存活率。实验重复三次,每次重复每个株系12株。冷处理前后的各幼苗的生长状态如图6所示,由图可以看出,与野生型拟南芥相比,转TabZIP14基因拟南芥的抗冻性明显提高。抗冻处理后,野生型植株的存活率为16%,转TabZIP14的株系存活率为79%(L1、L2和L3这三个株系的存活率均值)。2、抗盐性分析将野生型拟南芥(WT)以及三个T3代转TabZIP14基因拟南芥(L1,L2,L3)的种子在22℃,12h光照下进行垂直培养,得到5天幼苗,将各幼苗转移到含150和200mMNaCl的MS培养基上垂直培养,并分别以在不含NaCl的MS培养基上垂直培养得到的幼苗为对照。观察各组拟南芥幼苗生长状态并统计植株根长和植株白化率(盐胁迫处理后有白化叶片的植株株数与盐胁迫处理前植株株数之比)。实验重复三次,每次重复每个株系8株。各组拟南芥幼苗的状态如图7所示,由图可以看出,150mMNaCl处理后,野生型拟南芥的根长明显受到抑制,T3代转TabZIP14基因拟南芥L1、L2和L3株系的根长分别是野生型植株根长的1.4倍、1.3倍和1.4倍。部分野生型的拟南芥叶子呈现白化现象,白化率为37%±0.5%。而T3代转TabZIP14基因拟南芥叶片全部保持绿色。200mMNaCl处理后,T3代转TabZIP14基因拟南芥L1、L2和L3株系的根长比分别是野生型植株根长的1.2倍、1.3倍和1.3倍。野生型植株的叶片几乎全部呈现白化现在,白化率为98%±0.4%;而T3代转TabZIP14基因拟南芥叶片基本呈现绿色状态。 以上结果说明转TabZIP14基因拟南芥的抗盐性明显提高。当前第1页1 2 3
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1
- 蝎神经毒素AaIT及其编码基因与应用的制造方法与工艺
- 腈水解酶突变体及其编码基因和应用的制造方法与工艺
- 植物抗旱相关蛋白AsTMK1及其编码基因和应用的制造方法与工艺
- 植物抗旱相关蛋白EtSnRK2.2及其编码基因和应用的制造方法与工艺
- OsSAPK7蛋白及其编码基因在提高水稻白叶枯病抗性中的应用的制造方法与工艺
- 一种抗除草剂抗虫融合基因、编码蛋白及应用的制造方法与工艺
- 毛果杨PtrZFP103基因及其编码蛋白和应用的制造方法与工艺
- 大豆bHLH转录因子基因GmFER及其编码蛋白和应用的制造方法与工艺
- 一种花狭口蛙的基因及其编码的多肽和该多肽的用途的制造方法与工艺
- 一种泽陆蛙基因及其编码的多肽和该多肽的用途的制造方法与工艺
- 降低种子重量的白桦AP2基因及其编码蛋白的制造方法与工艺
- 可编码ArtRD蛋白基因及其蛋白和应用的制造方法与工艺
- 蝎神经毒素AaIT及其编码基因与应用的制造方法与工艺
- 腈水解酶突变体及其编码基因和应用的制造方法与工艺
- 植物抗旱相关蛋白AsTMK1及其编码基因和应用的制造方法与工艺
- 植物抗旱相关蛋白EtSnRK2.2及其编码基因和应用的制造方法与工艺
- OsSAPK7蛋白及其编码基因在提高水稻白叶枯病抗性中的应用的制造方法与工艺
- 一种抗除草剂抗虫融合基因、编码蛋白及应用的制造方法与工艺
- 毛果杨PtrZFP103基因及其编码蛋白和应用的制造方法与工艺
- 大豆bHLH转录因子基因GmFER及其编码蛋白和应用的制造方法与工艺
- 腈水解酶突变体及其编码基因和应用的制造方法与工艺
- 植物抗旱相关蛋白AsTMK1及其编码基因和应用的制造方法与工艺
- 植物抗旱相关蛋白EtSnRK2.2及其编码基因和应用的制造方法与工艺
- OsSAPK7蛋白及其编码基因在提高水稻白叶枯病抗性中的应用的制造方法与工艺
- 一种抗除草剂抗虫融合基因、编码蛋白及应用的制造方法与工艺
- 毛果杨PtrZFP103基因及其编码蛋白和应用的制造方法与工艺
- 大豆bHLH转录因子基因GmFER及其编码蛋白和应用的制造方法与工艺
- 一种花狭口蛙的基因及其编码的多肽和该多肽的用途的制造方法与工艺
- 一种泽陆蛙基因及其编码的多肽和该多肽的用途的制造方法与工艺
- 一种水稻MIT1基因、其编码蛋白及应用的制造方法与工艺
- 植物抗旱相关蛋白EtSnRK2.2及其编码基因和应用的制造方法与工艺
- OsSAPK7蛋白及其编码基因在提高水稻白叶枯病抗性中的应用的制造方法与工艺
- 一种抗除草剂抗虫融合基因、编码蛋白及应用的制造方法与工艺
- 毛果杨PtrZFP103基因及其编码蛋白和应用的制造方法与工艺
- 大豆bHLH转录因子基因GmFER及其编码蛋白和应用的制造方法与工艺
- 一种水稻MIT1基因、其编码蛋白及应用的制造方法与工艺
- 蛋白StTrxF、编码基因及其在提高植物淀粉含量中的应用的制造方法与工艺
- 蛋白质及其编码基因在调控植物抗黄萎病性中的应用的制造方法与工艺
- 甘薯耐逆相关蛋白IbFBA2、编码基因、重组载体与应用的制造方法与工艺
- 植物抗旱相关蛋白ZmNAC111及其编码基因与应用的制造方法与工艺
- 植物抗旱相关蛋白EtSnRK2.2及其编码基因和应用的制造方法与工艺
- OsSAPK7蛋白及其编码基因在提高水稻白叶枯病抗性中的应用的制造方法与工艺
- 一种抗除草剂抗虫融合基因、编码蛋白及应用的制造方法与工艺
- 毛果杨PtrZFP103基因及其编码蛋白和应用的制造方法与工艺
- 大豆bHLH转录因子基因GmFER及其编码蛋白和应用的制造方法与工艺
- 一种水稻MIT1基因、其编码蛋白及应用的制造方法与工艺
- 蛋白StTrxF、编码基因及其在提高植物淀粉含量中的应用的制造方法与工艺
- 植物抗旱相关蛋白ZmNAC111及其编码基因与应用的制造方法与工艺
- 核盘菌凝集素SSL‑6蛋白及其编码基因与应用的制造方法与工艺
- 植物抗病毒相关蛋白GmSN1及其编码基因和应用的制造方法与工艺
- 植物抗旱相关蛋白EtSnRK2.2及其编码基因和应用的制造方法与工艺
- OsSAPK7蛋白及其编码基因在提高水稻白叶枯病抗性中的应用的制造方法与工艺
- 一种抗除草剂抗虫融合基因、编码蛋白及应用的制造方法与工艺
- 毛果杨PtrZFP103基因及其编码蛋白和应用的制造方法与工艺
- 大豆bHLH转录因子基因GmFER及其编码蛋白和应用的制造方法与工艺
- 一种水稻MIT1基因、其编码蛋白及应用的制造方法与工艺
- 蛋白StTrxF、编码基因及其在提高植物淀粉含量中的应用的制造方法与工艺
- 蛋白质及其编码基因在调控植物抗黄萎病性中的应用的制造方法与工艺
- 植物抗旱相关蛋白ZmNAC111及其编码基因与应用的制造方法与工艺
- 核盘菌凝集素SSL‑6蛋白及其编码基因与应用的制造方法与工艺