引起免疫原性反应降低的被修饰细胞的制作方法
相关专利申请的交叉引用
本申请要求2015年1月17日提交的题目为“引起免疫原性反应降低的被修饰细胞”的美国临时专利申请号62/104,709的优先权,所述临时专利申请通过全文引用并入本文。
本公开涉及细胞治疗。更具体地,本公开涉及引起免疫原性反应降低的被修饰细胞。
背景技术:
干细胞具有自我更新和分化的能力。在某些条件下,干细胞可以分化成多种功能细胞。根据发育阶段,干细胞可以分为胚胎干细胞(es细胞)和成体干细胞两类:根据发展的潜力,干细胞可分为三类:全能干细胞(tsc),多能干细胞和单能干细胞。干细胞未分化和未成熟,具有再生人类各种组织和器官的能力。
自20世纪90年代以来,人们已经尝试了几种涉及干细胞的治疗策略从而来治愈疾病。然而,对于临床应用基于干细胞的细胞替代疗法,主要障碍仍然没有克服(例如同种异体移植免疫排斥)。
技术实现要素:
本文的实施方案涉及被修饰细胞,与相应的野生型细胞相比,被修饰细胞的主要组织相容性复合体ii(mhcii)较少。在此情况下,与相应的野生型细胞相比,被修饰细胞具有降低的免疫原性,而且被修饰细胞是被修饰的干细胞或源于该干细胞的细胞。
本文的实施方案进一步涉及制造被修饰干细胞用于生物移植的方法。该方法包括在培养基中培养该被修饰细胞,与相应的野生型细胞相比,被修饰细胞的mhcii较少。在此情况下,与相应的野生型细胞相比,被修饰细胞具有降低的免疫原性,而且被修饰细胞是被修饰的干细胞或源于该被修饰的干细胞的细胞。
本文的实施方案进一步涉及治疗病症的方法。该方法包括向受试者施用治疗有效量的该被修饰细胞,与相应的野生型细胞相比,被修饰细胞的mhcii较少。在此情况下,与相应的野生型细胞相比,被修饰细胞具有降低的免疫原性,而且被修饰细胞是被修饰的干细胞或源于该干细胞的细胞。
在一些实施方案中,与相应的野生型细胞相比,被修饰细胞的mhcii的生物合成或转运途径的一个或多个基因的表达有所降低。
在一些实施方案中,与相应的野生型细胞相比,被修饰细胞上累积的mhcii量减少。
在一些实施方案中,被修饰干细胞的与mhcii的生物合成或运输途径相关的内源基因有破坏。
在一些实施方案中,该破坏包括mhcii类反式激活因子(ciita)的破坏。
在一些实施方案中,该破坏源于至少部分ciita的缺失。
在一些实施方案中,mhcii包含人白细胞抗原ii(hlaii)。
在一些实施方案中,修饰的干细胞包括全能干细胞,多能干细胞,胚胎干细胞,诱导多能干细胞或多能干细胞中的至少一种。
在一些实施方案中,修饰的干细胞包含人胚胎干细胞。
在一些实施方案中,与相应的野生型细胞相比,降低的免疫原性包括由被修饰细胞诱导的炎症反应的死亡水平。
在一些实施方案中,被修饰细胞的核型与相应野生型细胞的核型相同。
在一些实施方案中,被修饰细胞的多能性水平与相应野生型细胞的多能性水平基本相同。
提供该发明内容以用简化的形式介绍以下具体实施方式中进一步描述的精选概念。该发明内容并不旨在标识所要求保护的主题的关键特征或必要特征,也不旨在用于限制所要求保护的主题的范围。
附图说明
参照附图描述具体实施方式。在不同附图中使用相同的参考数字表示类似或相同的项目。
根据与hescs中ciita的破坏相关的实施方案,图1示出了talens针对人ciita外显子2的序列(a),以及用talenx1制造的ciita缺陷型人胚胎干细胞(hesc)的效率(b)。
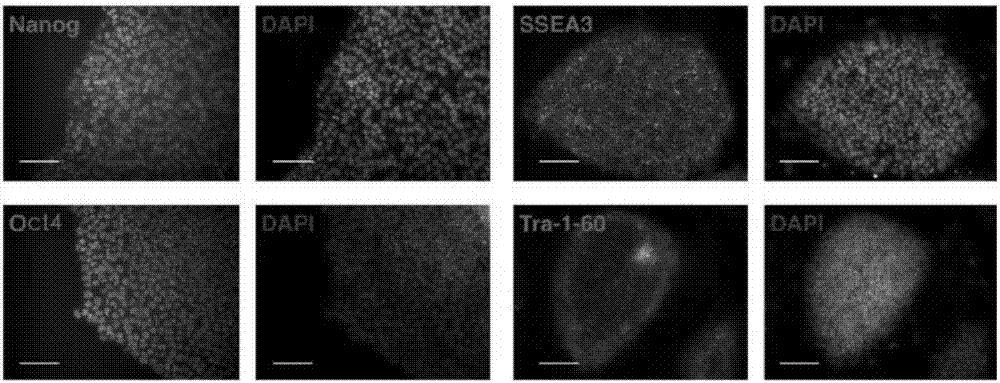
根据在与ciita靶向hescs的多能性相关的实施方案,图2a显示了在ciita-/-hescs中多能标记nanog,oct4,ssea3和tra-1-60的免疫染色。
根据ciita靶向hesc的多能性相关的实施方案,图2b显示了由ciita-/-hescs形成的畸胎瘤中he染色鉴定出三个胚层[(中胚层(左),外胚层(中)和内胚层(右)],比例尺为100μm。
根据与ciita靶向hesc的多能性相关的实施方案,图2c显示了ciita靶向的hesc衍生的eb中分化的标志物表达的rt-pcr分析。
根据与ciita靶向hesc的多能性相关的实施方案,图2d示了ciita杂合和纯合的hesc的核型分析。两组均有两个样本分析,无异常核型。
根据与来自ciita靶向hescs的成纤维细胞中ciita和hlaii类表达相关的实施方案,图3a显示了hescs衍生的成纤维细胞中β2m,ciita,hlaii(dra,dqa,dpa)和ii的rt-pcr分析。用ifn-γ(500u/ml)处理5天。对照组无ifn-γ。将所有组与ciita+/+ifn-γ游离组进行比较。通过t检验评估显着性。数据表示为平均值±sem.n≥3.***p<0.001,**p<0.01。
根据与来自ciita靶向hesc的成纤维细胞中ciita和hlaii类表达相关的实施方案,图3b显示了经过处理的成纤维细胞(如上所述治疗的成纤维细胞)中hlaii和ciita蛋白表达的蛋白质印迹。比例尺100μm。
根据与来自ciita靶向hesc的成纤维细胞中ciita和hlaii类表达相关的实施方案,图3c显示了处理的成纤维细胞(如上所述治疗的成纤维细胞)中hlaii和ciita蛋白表达的免疫染色分析。比例尺100μm。
根据与来自ciita靶向hesc的成纤维细胞中ciita和hlaii类表达相关的实施方案,图3d显示了处理的成纤维细胞中hlai和ii蛋白表达在细胞表面的facs分析。
根据与衍生自ciita靶向hesc的dcs中的hlaii类表达相关的实施方案,图4a显示涉及的来自ciita靶向hesc的dc中cd83,cd86,cd11c,dra,dpa,dqa,ii,ciita,hla-e和β2m的rt-pcr分析。将所有组与ciita+/+hescs组进行比较。
根据来自ciita靶向hesc的dcs中hlaii类表达相关实施方案,图4b示出了dcs中hlaii表达的facs分析,其由的cd83和cd86的共表达来定义,并比较hlaii+的百分比。通过t检验评估显着性。数据表示为平均值±sem,n≥3,***p<0.001,**p<0.01,*p<0.05。
根据与来自teratomas的人成纤维细胞的衍生相关的实施方案,图5显示了源自hesc的成纤维细胞和源自hesc的成纤维细胞表达波形蛋白的形态(a)。将所有组与ciita+/+成纤维细胞组进行比较(gb)。
具体实施方式
除非另有定义,本文所使用的所有技术和科学术语具有如本公开所属的领域的普通技术人员通常理解的相同的含义。虽然在本公开的实践或测试中可以使用与本文描述的方法和材料类似或等同的任何方法和材料,但是描述了优选的方法和材料。对于本公开的目的,下列术语定义如下。
本文使用冠词“一(a)”和“一个(an)”指的是一个或多于一个(即,指的是至少一个)该冠词的语法对象。举例来说,“元件”表示一个元件或多于一个元件。
“约”是指与参考量、水平、值、数量、频率、百分比、量纲、大小、用量、重量或长度相比,量、水平、值、数量、频率、百分比、量纲、大小、用量、重量或长度的变化高达30、25、20、15、10、9、8、7、6、5、4、3、2或1%。
“编码序列”指的是导致基因多肽产物编码的任何核酸序列。与之相比,术语“非编码序列”是指不会导致基因多肽产物编码的任何核酸序列。
在本发明书中,除了文中另有要求,词语“包含(comprise)”、“包含(comprises)”和“包含(comprising)”将被理解为意味包括声明的步骤或因子或步骤组或因子组,但不排除任何其它步骤或因子或步骤组或因子组。
“由……组成”是指包括并且限于短语“由……组成”之后所描述的内容。因此,短语“由……组成”表示所列元素是必要的或强制性的,并且其它元素可以不存在。
“基本上由……组成”是指包括该短语后面列出的任何元素,并且限于对本公开中针对所列元素说明的活性或作用不干扰或无贡献的其它元素。因此,短语“基本上由……组成”表示所列元素是必要的或强制性的,但是其它元素是任选的,并且根据它们是否影响所列元素的活性或作用可以存在或不存在。
术语“互补的”和“互补”指的是通过碱基配对原则相关的多核苷酸(即核苷酸的序列)。例如,序列“a-g-t”与序列“t-c-a”是互补的。互补可以是“部分的”,其中仅一些核酸的碱基根据碱基配对原则进行匹配。或者,核酸之间可存在“完全”或“总体”的互补。核酸链之间的互补程度对核酸链之间杂交的效率和强度具有显著影响。
“对应于”是指(a)具有与参考多核苷酸序列的全部或一部分基本上相同或互补的核酸序列或编码与肽或蛋白质中的氨基酸序列相同的氨基酸序列的多核苷酸;或者(b)具有与参考肽或蛋白质中的氨基酸的序列基本上相同的氨基酸序列的肽或多肽。
“衍生物”是指源自已经通过修饰的基本序列的多肽,例如通过与其它化学部分共轭或络合(例如聚乙二醇化)或者通过如本领域中可理解的后翻译修饰技术。术语“衍生物”还包含其范围内已对亲本序列所做的改变,包含提供用于功能上等同的分子的添加或缺失。
正如在本文中所使用的,术语“功能”和“功能的”等等是指生物的、酶促的或治疗的功能。
“基因”指的是在染色体中占据特异性位点并且由转录和/或翻译调节序列和/或编码区和/或非翻译序列(也就是内含子,5'和3'的非翻译序列)组成的遗传单位。
“同源性”是指相同的或构成保守性取代的氨基酸的百分比数字。可以使用诸如gap的序列对比程序确定同源性(deveraux等,1984,nucleicacidsresearch12,387-395),其通过引用并入本文。在这一方式中,通过将缺口插入对比,与本文中那些引用的序列相似或基本上不同长度的序列将被对比,例如,通过gap所使用的比较算法确定此类缺口。
术语“宿主细胞”包括可以是或已经是本发明任何重组的载体或分离的多核苷酸受体的单个细胞或者细胞培养物。宿主细胞包括单个宿主细胞的子代,并且该子代因正常的、随机的或有意的突变和/或变化而不必与原母细胞完全相同(在形态学上或总dna互补上)。宿主细胞包括使用本发明重组载体或多核苷酸在体内或体外转染或感染的细胞。包含本发明重组载体的宿主细胞为重组宿主细胞。
术语“干细胞”是指在多细胞生物体中发现的生物细胞,其可以分裂(通过有丝分裂)并分化成多种特殊的细胞类型,并可自我更新以产生更多的干细胞。在哺乳动物中,存在两种广泛类型的干细胞:从囊胚的内细胞团中分离的胚胎干细胞和在各种组织中发现的成体干细胞。
组合物(例如,干细胞)的术语“免疫原性”是指组合物诱导免疫反应的能力。例如,当干细胞被移植到受试者时,如果干细胞不接触mhci和/或mhcii,则免疫原性可以减弱。
“分离的”是指基本上或实质上不含通常在其天然状态下附带着的成分的物质。例如,如本文所使用的,“分离的多核苷酸”指的是在天然存在的状态下被来自其两侧的序列所纯化的多核苷酸,例如从正常毗邻于所述片段的序列所移除的dna片段。可选地,如本文所使用的,“分离的肽”或“分离的多肽”等指的是肽或多肽分子从其天然细胞环境中的体外分离和/或纯化,以及与细胞的其它成分关联。
术语“调节”和“改变”包括相对于对照通常在统计学上显著或生理学上显著的数量或程度的“增加”和“提高”以及“减小”或“降低”。在具体的实施例中,相对于未修饰或不同修饰的干细胞,与哺乳动物干细胞移植相关的免疫排斥减小。
“增加”或“增强”的量通常是“统计学上显著”的量,并且可包括本文所描述的量或水平增加1.1、1.2、1.3、1.4、1.5、1.6、1.7、1.8、1.9、2、2.5、3、3.5、4、4.5、5、6、7、8、9、10、15、20、30、40或50倍或更多倍(例如100、500、1000倍)(包括这些数值之间的所有整数和小数点,且大于1,例如1.5、1.6、1.7、1.8等)。
“减小”或“减少”或较少的量通常是“统计学上显著”的量,并且可包括本文所描述的量或水平减小1.1、1.2、1.3、1.4、1.5、1.6、1.7、1.8、1.9、2、2.5、3、3.5、4、4.5、5、6、7、8、9、10、15、20、30、40或50倍或更多倍(例如100、500、1000倍)(包括这些数值之间的所有整数和小数点,且大于1,例如1.5、1.6、1.7、1.8等)。
例如,“从...获得”指的是样品,诸如多核苷酸或多肽从特定源(诸如所需的生物或所需生物内特异性组织)分离或衍生得到的。“从...获得”也可以是指多核苷酸或多肽序列在其中从特定的生物或生物内组织分离或衍生得到的情形。例如,编码本文中所述的参比多肽的多核苷酸序列可以从多种原核或真核生物分离得到或从某些真核生物内特定组织或细胞总分离得到的。
本文所用的术语“可操作地连接的”指将基因放置在启动子的调控下,启动子然后控制基因的转录和任选的基因的翻译。在异源启动子/结构基因组合的构建中,通常优选将遗传序列或启动子定位于到基因转录起始位点的距离大约相同于基因序列或启动子与控制其自然环境的基因之间的距离;控制其自然环境的基因即基因序列或启动子所来源的基因。如本领域已知的,可适应该距离的一些变化而不损失功能。类似地,调节序列元件相对于其控制之下的异源基因的优选位置受其自然环境中元件位置的限定;即其所来源的基因。“组成型启动子”通常是活性的,即在大多数情况下,启动转录。“诱导型启动子”通常只在某些条件下是活性的,诸如在给定的分子因子(例如iptg)或特定的环境条件(例如特定的co2浓度、营养水平、光、热)的存在下。在不存在那种情况下,诱导型启动子通常不允许显著的或可测量水平的转录活性。例如,诱导型启动子可以根据温度、ph值、激素、代谢物(例如乳糖、甘露醇、氨基酸)、光(例如特定波长)、渗透势(例如盐诱导的)、重金属或抗生素等来诱导。许多标准的诱导型启动子对本领域技术人员将是已知的。
术语“多能性”是指es细胞的后代细胞保留多谱系分化潜力的es细胞的能力。在es细胞中维持多能性似乎涉及多个核因子之间的持续相互作用-这实现了一些平衡,其中一些相互作用是抑制或拮抗的,而其他相互作用是积极的或合作的从而促进多能性,这些抑制了分化中涉及的基因。保持多能性的重要因素包括oct4,nanog和sox2。
如本文所使用的,叙述“多核苷酸”或““核酸”是指mrna、rna、crna、rrna、cdna或dna。该术语通常指的是长度为至少10个碱基的核苷酸聚合体形式,所述核苷酸是核糖核苷酸或脱氧核苷酸或任一类型核苷酸的经修饰形式。该术语包括dna和rna的单链和双链形式。
术语“多核苷酸变体”和“变体”等指的是显示与参考多核苷酸序列具有基本序列同一性的多核苷酸或在下文定义的严格条件下与参考序列杂交的多核苷酸。这些术语还包括通过至少一个核苷酸的添加、缺失或取代而区别于参考多核苷酸的多核苷酸。因此,术语“多核苷酸变体”和“变体”包括其中一个或多个核苷酸被添加或缺失,或用不同的核苷酸置换的多核苷酸。在这方面,本领域技术人员很好理解的是,可以对参考多核苷酸进行某些改变,包括突变、添加、缺失和取代,由此所改变的多核苷酸保持参考多核苷酸的生物学功能或活性,或者相对于所述参考多核苷酸具有增加的活性(即,优化)。多核苷酸变体包括,例如,与本文中所描述的参考多核苷酸序列具有至少50%(和至少51%至至少99%,以及在之间的所有整数百分比,例如,90%、95%或98%)序列同一性的多核苷酸。术语“多核苷酸变体”和“变体”还包括天然存在的等位基因变体和编码这些酶的直系同源物。
关于多核苷酸,术语“外源的”指的是在野生型细胞或生物体中并非天然存在但是通常通过分子生物学技术将其引入到细胞中的多核苷酸序列。外源多核苷酸的实例包括载体、质粒,和/或编码所需蛋白质的人造核酸构建体。对于多核苷酸,术语“内源的”或“天然的”指的是可在给定的野生型细胞或生物体中找到的天然存在的多核苷酸序列。同样,从第一生物体分离并且通过分子生物学技术转移到第二生物体的特定多核苷酸序列通常被认为相对于所述第二生物体是“外源的”多核苷酸。在具体的实施例中,多核苷酸序列可以通过分子生物学技术被“引入”到已包含这样的多核苷酸序列的微生物中,例如,克隆一个或多个另外天然存在的多核苷酸序列,并且由此促进所编码的多肽的过表达。
列举的与mhcii的基因有关的“突变”或“缺失”通常指那些干细胞的变化或改变,其使该基因的产物对于糖原的合成和/或储存或给定脂质的生物合成无功能或具有降低的功能。这样的变化或改变的例子包括部分或全部核苷酸对于靶基因的编码或调控序列(例如ciita)的取代,缺失或添加,这破坏、消除、下调或显著减少由该基因编码的多肽的表达,无论是在转录还是翻译水平,和/或其产生相对不活跃的(例如,突变或缺失)或不稳定的多肽。用于产生这些改变或变化的技术,诸如通过与具有可选择标记的载体重组,在本文中示例性给出并且在分子生物学技术领域中是已知的。在具体的实施方案中,基因的一个或多个等位基因,例如两个或全部等位基因,可以在干细胞内突变或缺失。
靶基因的“缺失”也有可能通过打靶基因的mrna实现,如使用领域内已知的多种反义技术(如反义寡核苷酸和sirna)。相应的,当基因的多肽或者编码的酶在修饰过的光合微生物中不表达或者表达量可忽略,靶基因可被认为是“无功能的”。这使得修饰过的干细胞产生或积累比未修饰或者其他不同修饰的干细胞更少的多肽或者酶产物(如mhcii)。
在某些方面,通过在核苷酸水平的突变或改变编码多肽的氨基酸序列,靶基因可以被渲染“非功能性”。这使得一条修饰的多肽被表达,无论是通过修饰该多肽的活性位点、其细胞内定位、其稳定性,或对于本领域技术人员显而易见的其他功能特性,使得其功能或者活性降低(例如,ciita)。参与mhcii生物合成或运输途径的编码序列或一条多肽的这种修饰可以根据本领域已知的技术来完成,如对一个给定的干细胞进行基因组水平的定向诱变,和/或天然的选择(如,定向进化)。
“多肽”、“多肽片段”、“肽”和“蛋白质”在本文中可互换使用,用来指氨基酸残基的聚合物及其变体和合成类似物。因此,这些术语适用于其中一个或多个氨基酸残基是合成的非天然存在的氨基酸的氨基酸聚合物(诸如相应的天然存在的氨基酸的化学类似物),以及适用于天然存在的氨基酸聚合物。在某些方面,多肽可以包括通常催化不同的化学反应的酶多肽,或“酶”。
叙述多肽“变体”指的是通过至少一个氨基酸残基的添加、缺失或取代而区别于参考多肽序列的多肽。在某些实施例中,多肽变体通过一个或多个取代基而区别于参考多肽,所述取代基可以是保守的或非保守的。在某些实施例中,所述多肽变体包含保守性取代基,并且在这方面,本领域中很好理解的是,一些氨基酸可以被改变为具有广泛相似性质的其它氨基酸,而不改变多肽的活性性质。多肽变体还包括其中一个或多个氨基酸被添加或缺失,或用不同的氨基酸残基置换的多肽。
术语“参比序列”一般是指另一序列与之做对比的核酸编码序列或氨基酸序列。本文中所述的所有多肽和多核苷酸序列包括在参比序列中,包括由名称和序列表中描述的那些。
本文中所使用的引述“序列一致性”(或,例如,包括“50%相同的序列”)是指在比较窗口内核甘酸紧接核甘酸的基础上或氨基酸紧接氨基酸的基础上序列相同的程度。因此,可以在比较窗口内通过比较两个最佳比对序列来计算“序列一致性的百分比”,确定在位置上两序列中出现的相同核酸碱基(例如,a、t、c、g、i)或相同氨基酸残基(例如,ala、pro、ser、thr、gly、val、leu、ile、phe、tyr、trp、lys、arg、his、asp、glu、asn、gln、cys和met)的位置的数字以获得配对位置的数字,通过在比较窗口(也就是说窗口大小)内位置的总数字将配对位置的数字划分,以及将结果乘以100以产生序列一致性的百分比。包括了具有与本文所述参比序列(参见,例如,序列表)任意一个至少约50%、55%、60%、65%、70%、75%、80%、85%、90%、95%、97%、98%、99%或100%序列一致性的核苷酸和多肽,其中多肽变体通常保持有该参比多肽的至少一个生物活性。
“统计学显著的”,指的是结果并不是偶然发生的。统计学显著可以通过本领域任何已知的方法来确定。通常用p值来衡量显著性,它表示当原假设为真时所得到的样本观察结果或更极端结果出现的概率。如果所获得的p值小于显著性水平,则该原假设被拒绝。一般情况下,如果p值小于或等于0.05,则认为是显著的。
“大体上”或“基本上”是指几乎完全或完全,例如,95%,96%,97%,98%,99%或更大一些给定的数量。
“转化”是指,从摄取和掺入到宿主细胞基因组中的外源dna得到的;或者是外源基因从一个生物体转移进入另一生物体的基因组中得到的永久性遗传改变。
术语“野生型”是指,基因或基因产物具有从天然来源中分离得到的基因或基因产物的特点。一个野生型的基因或基因产物(如一段多肽)是在野生群体中观察到的最高频率的表型,因此也是在“正常”或“野生型”的基因形式中任意定义的。
本文的实施方案涉及可以通过与主要相容性复合物ii(mhcii)的生物合成或运输途径相关的内源基因的破坏可获得低免疫原性(例如,免疫原性降低)和相容性干细胞。与这些低免疫原性和相容性干细胞分化的细胞不具有组成型和ifn-γ诱导的hlaii,并且可以降低例如t细胞介导的对细胞治疗作用的排斥反应。
人t细胞的激活基于两个信号(tcr-hla信号和共刺激信号)。hla分子由大的基因家族编码并分为i类和ii类。首先,专职或非专职抗原呈递细胞(apc)将蛋白质降解成肽,然后将这些肽加载到hla分子上。然后,cd4+和cd8+t细胞的tcr分别识别hlaii和hlai呈递的肽。同时,这些apc表达一系列共刺激分子(例如cd80和cd86),其将与t细胞的互补分子(例如cd28和细胞毒性t淋巴细胞抗原4(ctla4))相互作用。
需要tcr-hla信号和共刺激信号来激活t细胞。因此,如果抑制任何一种,t细胞不会攻击同种异体移植物。已经证明,表达ctla4-免疫球蛋白融合蛋白(ctla4-ig)和程序性死亡配体-1(pd-l1)的hesc可以通过同时破坏共刺激途径和激活t细胞抑制途径来抑制同种异体免疫应答。这个策略是有用的,但不是普遍适用的。例如,来自hesc的t细胞不能用ctla4-ig和pd-l1的表达来活化。因此,这种方法限制了hesc在临床免疫治疗中的应用,例如hesc衍生的嵌合抗原受体(car)-t,这是癌症治疗中的有效治疗方法。
此外,与小鼠t细胞不同,活化的人t细胞表达hlaii。生产低免疫原性和相容性car-t可能会阻止受体t细胞介导的排斥反应。此外,dcs可以来自没有hlaii的那些hescs。虽然这些dcs不能正常呈递抗原,car技术(car-dcs)和人造hla-肽可以使这些被修饰细胞对癌症更具特异性和敏感性。
在每个有核细胞的表面上发现hlai分子。组成型hlaii分子主要表达于胸腺上皮细胞和专职的apc,包括dcs,b淋巴细胞,单核细胞和巨噬细胞。在炎性细胞因子(例如ifn-γ和tnf-α)的压力下,非专职apc如成纤维细胞和上皮细胞也可以表达hlaii分子,这被称为“诱导hlaii”。每个经典hlai分子结构上由多态性重链(例如hla-a,hla-b和hla-c)组成,其结合相同的轻链β2m。敲除β2m的hescs证明了hlai分子的损失,其使hesc具有避免cd8+t细胞介导的排斥的能力。然而,没有报告已经证明产生具有分化为没有组成型和ifn-γ诱导的hlaii的细胞的能力的hesc。
本文的实施方案涉及被修饰细胞,与相应的野生型细胞相比,被修饰细胞的mhcii较少。在某些情况下,与相应的野生型细胞相比,被修饰细胞具有降低的免疫原性,而且被修饰细胞是被修饰的干细胞或源于该被修饰的干细胞的细胞。
本文的实施方案涉及制造被修饰细胞用于生物移植。该方法包括培养该被修饰细胞,与相应的野生型细胞相比,被修饰细胞的mhcii较少。在某些情况下,与相应的野生型细胞相比,被修饰细胞具有降低的免疫原性,而且被修饰细胞是被修饰的干细胞或源于该被修饰的干细胞的细胞。
本文的实施方案涉及治疗病症的方法。该方法包括向受试者施用治疗有效量的该被修饰细胞,与相应的野生型细胞相比,被修饰细胞的mhcii较少。在某些情况下,与相应的野生型细胞相比,被修饰细胞具有降低的免疫原性,而且被修饰细胞是被修饰的干细胞或源于该被修饰的干细胞的细胞。
在一些实施方案中,该病症可以包括帕金森病,亨廷顿舞蹈病,阿尔茨海默病,肌萎缩性侧索硬化,脊髓性肌萎缩,神经变性疾病精神病以及脊髓损伤,中风,烧伤,心脏病,肝脏疾病,糖尿病,血癌,器官移植缺损和损伤再生的至少一种。
在一些实施方案中,与相应的野生型细胞相比,被修饰细胞的mhcii的生物合成或转运途径的一个或多个基因的表达有所降低。
在一些实施方案中,与相应的野生型细胞相比,被修饰细胞上累积mhcii减少。
在一些实施方案中,修饰的干细胞包括具有与mhcii的生物合成或运输途径相关的内源基因(例如,rfx因子(rfxap,rfx5,rfxank)和ciita)的破坏。在某些实施方案中,该破坏包括mhcii类反式激活因子(ciita)的破坏。例如,由于删除ciita的至少一部分而导致破坏。
hlaii基因由相同的调节复合物调控,由三个rfx因子(rfxap,rfx5,rfxank)和ciita组成。该复合物不仅调节编码经典hlaii分子(hla-dp,hla-dq和hla-dr)的基因,而且调节编码hlaii分子的细胞内运输和肽负载所需的辅助蛋白的基因,包括非经典hlaii分子(不变链(ii),hla-dm和hla-do)。在某些情况下,肿瘤细胞和病毒感染的细胞将通过干扰hlaii合成而逃避cd4+t细胞介导的免疫排斥反应。在这里使用talens技术,通过敲除ciita(hlaii分子的主调节子)来破坏hesc的hlaii分子。ciita的主要功能是调控hlaii,因此它们具有几乎相同的细胞分布。ciita不直接结合dna,但与由环amp应答元件结合蛋白(creb),核因子y复合物(nf-y)和rfx因子(rfx5,rfxank,rfxap)组成的其他元件相互作用。没有功能性ciita的患者患有裸淋巴细胞综合征(bls),其特征在于组织细胞中hlaii的表达不足。mhcii类介导的同种异体反应中,ciita-/-小鼠也受损。
ciita有四个启动子,它们可以以组织特异性方式调节hlaii表达。为了完全针对ciita,talens设计针对的是所有转录公共外显子(外显子2和3)。即使在胚状体(ebs)分化或ifn-γ诱导期间,hescs也不体外表达hlaii和ciita。分别检查了hescs衍生的dc和成纤维细胞上的组成型和诱导的hlaii分子。ciita的缺失可能会显着降低hlaii分子的组成型和诱导型表达。
在一些实施方案中,mhcii包含人白细胞抗原ii(hlaii)。在此情况下,修饰的干细胞包含人胚胎干细胞。
在一些实施方案中,修饰的干细胞包括全能干细胞,多能干细胞,胚胎干细胞,诱导多能干细胞或多能干细胞中的至少一种。
在一些实施方案中,与相应的野生型细胞相比,例如当将被修饰细胞移植到受试者时,降低的免疫原性包括由被修饰细胞诱导的炎症反应的死亡水平。
在一些实施方案中,被修饰细胞的核型与相应野生型细胞的核型相同。
在一些实施方案中,被修饰细胞的多能性水平与相应野生型细胞的多能性水平基本相同。在这些情况下,来自修饰的干细胞分化的细胞(例如成纤维细胞和上皮细胞)不具有组成型和ifn-γ诱导的hlaii。
可以组合上述各种实施方案以提供更进一步的实施方案。如果需要使用各专利、申请和公开的概念以提供其它实施方案,可对各实施方案的特征进行修改。
可以根据上述具体描述对这些实施方案作出这些和其它改变。通常,在随后的权利要求中,所使用的术语应当不被视为将权利要求限于本说明书和权利要求书所公开的具体实施方案,而是应当被视为包含全部可能的实施方案,连同这些权利要求所具有的等价方案的全部范围。因此,权利要求不受本公开内容的限制。
talens破坏hescs中ciita
使用talens在hescs中敲除ciita。ciita的talen是针对外显子2和外显子3设计。选自293t试验中最有效的talen对(2l2和2r2),并用于靶向x1hescs中的ciita(图1)。在一轮中获得了杂合子(ciita+/-)和纯合(ciita-/-)hesc,效率为70%(图1)。
数据表示为平均值±sem。使用graphpadprism5.1(graphpadsoftwareinc.,usa)统计分析数据。通过不配对的student'st检验分析性能变量。除非另有说明,显著是基于*p<0.05。
hescs培养
在用matrigel(becton-dickinson)包被的t25烧瓶(corning)上培养hescs,照射cf1饲养细胞(3×104个细胞/cm2)。将hesc培养在dmem/f12(invitrogen),该培养基含有20%敲除血清替代物(invitrogen),4ng/ml碱性成纤维细胞生长因子(bfgf;invitrogen),2mmol/l1-谷氨酰胺(invitrogen),1%非必需氨基酸(invitrogen)和0.1mmol/lβ-巯基乙醇(sigma-aldrich)。hescs每周大约传播一次。使用胶原酶iv将细胞从进料器中分离为细胞团,在被传送到新制备的饲养细胞之前将其解离成适当的大小。
talens效率检测
设计了ciita的talen靶向外显子2(2l1:gctgaccccctgtgcct(seqidno:1);2l2:gaccccctgtgcctct(seqidno:2);2r1:ctccagccaggtccatct(seqidno:3);2r2:tctccagccaggtccat(seqidno:4))和外显子3(3l1:tcagcaggctgttgt(seqidno:5);3l2:tcagcaggctgttgtgt(seqidno:6);3r1:ccctggtctcttcat(seqidno:7);3r2:aagcctccctggtctt(seqidno:8);3r3:aagcctccctggtct(seqidno:9))。talens用fasttaletalen装配工具(斯丹赛)构建,其活性在293t细胞中得到证实(如前所述)。将构建的talen转染到293t细胞中,并用2μg/ml嘌呤霉素(sigma)选择。选择后收获293t细胞的基因组dna。然后,进行pcr和测序以检查talens的效率。
生成ciita缺陷型hescs
为了制备用于转染的细胞,将收获的hesc接种在用matteig包被的六孔板中,六孔板含有mtesrtm1培养基(stemcelltechnologies)。第二天,通过fugenehd转染试剂(promega)将最有效的talen(2l2和2r2)质粒和egfp-puro质粒(斯丹赛)(1:1:1)转染入hesc。将fugenehd转染试剂/质粒/opti-mem(lifetechnologies)混合物(15μl/6ug/300ul)在室温下孵育15分钟,然后将混合物加入细胞培养物中。两天后将嘌呤霉素加入培养基中。用0.5μg/ml嘌呤霉素选择后,使用tryple(invitrogen)将存活菌落解离成单个细胞,并以500个细胞/cm2的密度接种到cf1包被的平板上。传代后两周,将衍生自单细胞的菌落转移到新鲜的cf1包被的孔中,并行使用直接细胞pcr试剂盒鉴定突变体。
针对ciita的hesc的多能性
hescs的多能性对于其在细胞替代疗法中的应用是必需的。因此,对已建立的ciita靶向hescs的多能性进行了调查。免疫染色显示,ciita-/-hescs对oct4,nanog,tra-1-60和ssea3呈阳性(图2a)。当将ciita-/-hescs注射入非肥胖型糖尿病/严重联合免疫缺陷型(nod/scid)小鼠时,2个月后形成畸胎瘤,在苏木精-伊红(he)染色部分观察到来自3个胚层的组织(图2b)。从ciita-/-hescs衍生的eb进行rt-pcr以显示三个胚层分子标记的表达(图2c)。此外,ciita-/-和ciita+/-hescs均具有正常核型(图2d)。正常核型谱系用于以下的实验。因此,靶向hesc在多能性和核型中没有显示任何差异。rt-pct的引物列于表1。
表格1
来自ciita靶向hescs细胞中的ciita和hlaii表达
对于细胞替代治疗,分化的细胞被直接移植到受试者体内。一些组织细胞(例如专职apc和胸腺上皮细胞)具有hlaii分子和一些其他组织细胞(例如,成纤维细胞和上皮细胞)的组成型表达已经诱导hlaii分子的表达。为了确保hlaii的功能性破坏,研究了从ciita靶向hesc衍生的定义类型的细胞中的两种hlaii表达。
首先检测ifn-γ可诱导的hlaii对hescs来源的成纤维细胞的影响,并进行5天的500uifn-γ处理。使用ccd-1079sk(ccd)细胞系,人成纤维细胞系作为阳性对照。ifn-γ诱导可增加组织细胞中β2m的表达。没有ifn-γ治疗,所有细胞均显示hlaii基因(ciita,dra,dpa,dqa,ii)和β2m的低水平表达。通过ifn-γ处理,所有组中β2m和ciitamrna都增加(图3a)。ciita靶向不影响ciita的转录。ciita+/+和ciita+/-成纤维细胞增加了ccd细胞在ifn-γ处理后的hlaii基因(dra,dpa,dqa,ii)的mrna表达(图3a)。然而,ifn-γ处理的ciita-/-成纤维细胞没有明显增加hlaii基因(dra,dpa,dqa,ii)的mrna表达(图3a)。表明在ifn-γ处理的ciita-/-成纤维细胞中检测到的ciitamrna功能失调,不能转化为功能性蛋白质以调节hlaii的表达(图3a)。通过以下蛋白质印迹和免疫化学数据证明了这一点(图3b,c)。它还表明,ciita+/-成纤维细胞的ciita和hlaii蛋白的水平升高滞后于mrna的增加(图3b,c)。所有组的facs分析显示,在ifn-γ诱导下,细胞表面上几乎没有细胞表达hlaii。ifn-γ诱导后,ccd和ciita+/+成纤维细胞显着增加hlai和ii的表达。然而,ciita+/-和ciita-/-都没有发现明显的hlaii表达的增加(图3d)。
其次,在来自hesc的dcs上,测试了组成型hlaii的表达。为了临床使用方案,选择具有明确化学成分培养基的方案,该培养基不包括血清,饲料以及其他动物的产品。衍生自hescs的dc表达cd83和cd86(图4a,b)。与ciita+/+和ciita+/-dcs相比,ciita-/-dcs中发现较低水平的经典hlaii分子(dra,dqa和dpa)的mrna表达(图4a)。然而,非经典hlaii基因(ii)的mrna表达没有显示任何差异(图4a)。经典hlaii基因(hla-dp,hla-dq和hla-dr)和非经典hlaii基因(hla-dm,hla-do,ii)具有相同的特异性调节模块,可被rfx-ciita综合体识别。该结果表明hla-dr表达完全依赖于ciita,其可能导致ciita靶向细胞中其他hlaii分子的残留表达(图3a,4a)。因此,ii在dcs和成纤维细胞之间具有不同的趋势,并且表明独立于ciita的ii的不同调节途径。ifn-γ诱导的成纤维细胞和dcs中ii的表达可能主要依赖于ciita,而dcs分化持续这么长时间才能激活没有ciita的替代调节途径。似乎ii编码辅助蛋白是hlaii分子的肽加载所必需的,并且不能拯救细胞表面上的dra,dpa和dqa的损失(图3d,4b)。
dcs用cd83和cd86来定义,同时比较了cd83+cd86+dcs中hlaii+细胞的百分比。pbm衍生的dcs显示出这三种标记的高度重叠(图4b)。ciita-/-dcs具有1.98%的hlaii+细胞,而ciita+/+和ciita+/-dcs的hlaii+细胞百分比分别为39.1%和24.8%。
畸胎瘤的人成纤维细胞的畸胎形成和衍生
肌内注射hescs至6-8周nod/scid小鼠(每个位点约5×106个细胞)。约2个月后,肿瘤取样并且进行苏木精-伊红(he)染色。
成纤维细胞样细胞也衍生于畸胎瘤。将tertomas用剪刀切成块,并在补充有10%血清,1%pen-strep和50umβ-甲基乙醇的dmem中培养。经过几次传代之后,贴壁细胞变成均匀的成纤维细胞样细胞。进行细胞形态观察和rt-pcr(图5)。建立了10个细胞系(3个为+/+;3为+/-;4为-/-)。并分析了建立的细胞系中的一些间充质干细胞标记(n>3)。ccd和间充质干细胞(msc)作为对照。那些细胞系更像是成纤维细胞。表明该方法在这些实验中是可重现的。
从hescs衍生人dcs
由先前报道的,dcs与hescs的分化是ebs悬浮培养中逐步通过生长因子来诱导的。在前5天,hescs培养在x-vivotm15培养基(lonza)中,该培养基进一步包括1mm丙酮酸钠,1×非必需氨基酸,2mml-谷氨酰胺,50mm2-巯基乙醇,四种生长因子:包括重组人骨形态发生蛋白-4(rhbmp-4;bd),重组人血管内皮生长因子(rhvegf;r&d),重组人粒细胞巨噬细胞集落刺激因子(rhgm-csf;r&d)和重组人干细胞因子(rhscf;r&d),hescs显示中胚层。从第6天到第10天,除去rhbmp-4,细胞成为造血干细胞(hscs)。从第11天到第15天,除去rhvegf,将hsc变成常见的骨髓祖细胞(cmp)。从第16天到第20天,去除rhscf,逐渐出现并累积单核细胞样细胞作为dc前体。在未来4-6天内,治疗rhgm-csf和重组人白细胞介素4(rhil-4;r&d)将dc前体诱导成为不成熟的dcs(idc)。dcs的成熟需要用混合因子进一步孵育1-2天,混合因子包括rhgm-csf,重组人白细胞介素-1β(rhil1-β;r&d),重组人干扰素γ(rhifn-γ;r&d),前列腺素e2(pge-2;sigma)和重组人肿瘤坏死因子α(rhtnf-α;r&d)。
- 还没有人留言评论。精彩留言会获得点赞!