芳香化合物的生产的制作方法
本领域涉及在生产例如(+)-雪松醇或(-)-罗汉柏烯的萜烯的方法中使用的核酸、酶、载体和细胞。
背景技术:
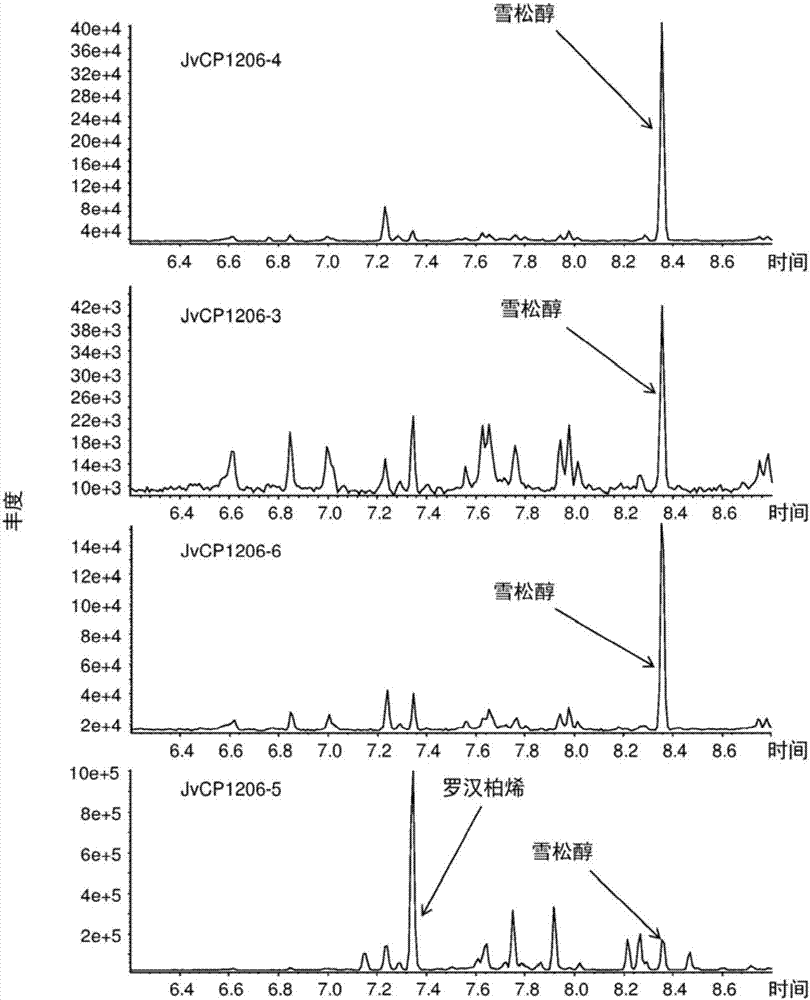
:在大多数生物体(微生物、动物和植物)中发现了萜烯。这些化合物由称为异戊二烯单元的五碳单元构成并且通过存在于它们结构中的这些单元的数目进行归类。因此,单萜、倍半萜和二萜是分别含有10、15和20个碳原子的萜烯。例如,在植物界广泛地发现了倍半萜。许多倍半萜分子因为它们的风味和芳香特性以及它们的美容、医药和抗菌效果而公知。已经鉴定出许多倍半萜烃类和倍半萜类。萜烯的生物合成生产涉及称为萜合酶的酶。倍半萜合酶存在于植物界,并使用底物法尼基焦磷酸(fpp),但它们具有不同的产物特性。已经克隆了编码倍半萜合酶的基因和cdna,并表征了相应的重组酶。(+)-雪松醇目前的来源是含有雪松油的松柏类植物。(-)-罗汉柏烯目前的来源是例如刺柏(juniperuscedrus)和罗汉柏(thujopsisdolabrata)的松柏类植物。技术实现要素:本文提供了来自北美圆柏(juniperusvirginiana)、金塔柏(platycladusorientalis'beverleyensis')或侧柏(platycladusorientalis)的分离的,其包含(+)-雪松醇或(-)-罗汉柏烯合酶。本文还提供了来自北美圆柏(juniperusvirginiana)、金塔柏(platycladusorientalis'beverleyensis')或侧柏(platycladusorientalis)的分离的核酸分子,其编码(+)-雪松醇或(-)-罗汉柏烯合酶。本文还提供了生产(+)-雪松醇或(-)-罗汉柏烯的方法,包括:a.使非环状法呢基二磷酸酯(fpp)前体与具有从(+)-雪松醇合酶活性和(-)-罗汉柏烯合酶活性中选出的活性的多肽接触,其中该多肽包含:i.与从seqidno:1、seqidno:2、seqidno:3、seqidno:4、seqidno:13和seqidno:14中选出的多肽具有至少70%、75%、80%、85%、90%、95%、98%和/或99%的序列同一性的氨基酸序列;或者ii.从seqidno:1、seqidno:2、seqidno:3、seqidno:4、seqidno:13和seqidno:14中选出的氨基酸序列;从而生产从(+)-雪松醇和(-)-罗汉柏烯中选出的化合物;以及b.可选地,分离(+)-雪松醇和(-)-罗汉柏烯,条件是当该多肽包含:i.与从seqidno:1、seqidno:2、seqidno:3和seqidno:13中选出的序列具有至少70%、75%、80%、85%、90%、95%、98%和/或99%的序列同一性的氨基酸序列;或者ii.从seqidno:1、seqidno:2、seqidno:3和seqidno:13中选出的氨基酸序列时;所生产的化合物为(+)-雪松醇,而不存在(-)-罗汉柏烯。本文还提供了一种多肽,其中该多肽包含:a.与从seqidno:1、seqidno:2、seqidno:3、seqidno:4、seqidno:13和seqidno:14中选出的多肽具有至少70%、75%、80%、85%、90%、95%、98%和/或99%的序列同一性的氨基酸序列;或者b.从seqidno:1、seqidno:2、seqidno:3、seqidno:4、seqidno:13和seqidno:14中选出的氨基酸序列。本文还提供了一种核酸,其编码如上所述的多肽。本文还提供了一种核酸,其包含:a.与从seqidno:5、seqidno:6、seqidno:7、seqidno:8、seqidno:9、seqidno:10、seqidno:11、seqidno:12、seqidno:15、seqidno:16、seqidno:17、seqidno:18和seqidno:19中选出的核苷酸序列有至少70%、75%、85%、90%、95%、98%和/或99%相似或至少70%、75%、80%、85%、90%、95%、98%和/或99%相同的核苷酸序列;或者b.从seqidno:5、seqidno:6、seqidno:7、seqidno:8、seqidno:9、seqidno:10、seqidno:11、seqidno:12、seqidno:15、seqidno:16、seqidno:17、seqidno:18和seqidno:19中选出的核苷酸序列。附图说明图1.北美圆柏(juniperusvirginiana)幼苗(1~2岁龄)的地表和地下部分的gcms分析。表示了(+)-雪松醇的峰。图2.通过四种不同的北美圆柏(j.virginiana)倍半萜合酶jvcp1206-4、jvcp1206-3、jv1206-6和jvcp1206-5在体外测定中产生的倍半萜混合物的gcms分析。表示了对应于(+)-雪松醇和(-)-罗汉柏烯的峰。图3.通过表达四种不同的北美圆柏(j.virginiana)倍半萜合酶jvcp1206-4、jvcp1206-3、jv1206-6和jvcp1206-5的工程细菌细胞在体内产生的倍半萜混合物的gcms分析。表示了对应于(+)-雪松醇和(-)-罗汉柏烯的峰。图4.由重组北美圆柏(j.virginiana)倍半萜合酶产生的(+)-雪松醇和(-)-罗汉柏烯的结构。图5.金塔柏(p.orientalis'beverleyensis')叶子二氯甲烷提取物的gc/ms色谱图(仅显示倍半萜的区域)。箭头表示(+)-雪松醇的峰。图6.图5中(+)-雪松醇的峰的质谱图。图7.侧柏(p.orientalis)叶子二氯甲烷提取物的gc/ms色谱图(仅显示倍半萜的区域)。箭头表示(-)-罗汉柏烯的峰。图8.图7中(-)-罗汉柏烯的峰的质谱图。图9.porb1的大肠杆菌表达实验的gc/ms色谱图(仅显示倍半萜的区域)。箭头表示(+)-雪松醇的峰。图10.图9中(+)-雪松醇的峰的质谱图。图11.por2-3-5的大肠杆菌表达实验的gc/ms色谱图(仅显示倍半萜的区域)。箭头表示(-)-罗汉柏烯的峰。图12.图11中(-)-罗汉柏烯的峰的质谱图。具体实施方式对于本文的说明书以及所附的权利要求书,除非另有说明,使用的“或”是指“和/或”。类似地,“包含(comprise)”、“包含(comprises)”、“包含(comprising)”、“包括(include)”、“包括(includes)”和“包括(including)”可以互换使用并且不旨在限制。还应当理解的是,在各个实施方案的描述使用术语“包含”的情况下,本领域技术人员能够理解,在一些具体情形中,可以替代地使用语言“基本上由……组成”或“由……组成”来描述一个实施方案。在一个实施方案中,本文提供的方法包括以下步骤:用编码具有(+)-雪松醇合酶或(-)-罗汉柏烯合酶活性的多肽的核酸转化宿主细胞或非人生物体,其中该多肽包含:a.与从seqidno:1、seqidno:2、seqidno:3、seqidno:4、seqidno:13和seqidno:14中选出的多肽具有至少70%、75%、80%、85%、90%、95%、98%和/或99%的序列同一性的氨基酸序列;或者b.从seqidno:1、seqidno:2、seqidno:3、seqidno:4、seqidno:13和seqidno:14中选出的氨基酸序列;并在允许生产该多肽的条件下培养该宿主细胞或生物体。在另一个实施方案中,本文提供的方法还包括在有助于生产(+)-雪松醇或(-)-罗汉柏烯的条件下,培养能够生产fpp并能被转化以表达多肽的非人宿主生物体或细胞,其中该多肽包含:a.与从seqidno:1、seqidno:2、seqidno:3、seqidno:4、seqidno:13和seqidno:14中选出的序列具有至少70%、75%、80%、85%、90%、95%、98%和/或99%的序列同一性的氨基酸序列;或者b.从seqidno:1、seqidno:2、seqidno:3、seqidno:4、seqidno:13和seqidno:14中选出的氨基酸序列。在另一个实施方案中,本文提供一种表达载体,其包含本文所述的核酸。在另一个实施方案中,本文提供一种非人宿主生物体或细胞,其被转化以含有本文所述的至少一种核酸,使得其异源表达或过表达本文所述的至少一种多肽。在一个实施方案中,本文提供的非人宿主生物体是植物、原核生物或真菌。在一个实施方案中,本文提供的非人宿主是微生物,特别是细菌或酵母。在一个实施方案中,本文提供的非人生物体是大肠杆菌,而所述酵母是酿酒酵母(saccharomycescerevisiae)。在一个实施方案中,本文提供的非人生物体是酿酒酵母。在一个实施方案中,细胞是原核细胞。在另一个实施方案中,细胞是细菌细胞。在一个实施方案中,细胞是真核细胞。在一个实施方案中,真核细胞是酵母细胞或植物细胞。在另一个实施方案中,本文提供的方法还包括使用化学或生物化学合成或两者的组合将(+)-雪松醇加工成衍生物。在另一个实施方案中,本文提供的方法还包括使(+)-雪松醇与至少一种酶接触以生产(+)-雪松醇衍生物。在另一个实施方案中,本文提供的方法包括使用化学或生物化学合成或两者的组合将(-)-罗汉柏烯转化为(-)-罗汉柏烯衍生物。在另一个实施方案中,本文提供的方法还包括使(-)-罗汉柏烯与至少一种酶接触以生产罗汉柏烯衍生物。多肽催化特定倍半萜(例如(+)-雪松醇合酶或(-)-罗汉柏烯合酶)合成的能力可以通过进行如本文提供的实施例中详述的酶测定来简单地证实。多肽也意味着包括截短的多肽,只要它们保持它们的(+)-雪松醇合酶活性和/或(-)-罗汉柏烯合酶活性。如下文所述,通过修饰本文所述的序列获得的核苷酸序列可以使用本领域已知的任何方法获得,例如通过引入任何类型的突变,例如缺失、插入或取代突变。这些方法的例子在说明书中与变体多肽和制备它们的方法有关的部分中列举。当产生这两个序列的比对时,两个肽或核苷酸序列之间的同一性百分比是两个序列中相同的氨基酸或核苷酸残基的数目的函数。相同残基被定义为两个序列中在比对的给定位置中相同的残基。本文使用的序列同一性的百分比是从最佳比对中通过将两个序列之间相同的残基数除以最短序列中的残基总数并乘以100计算得到的。最佳比对是同一性百分比最高可能性的比对。可以将间隙引入到一个或两个序列中的比对的一个或多个位置中以获得最佳比对。然后将这些间隙考虑为用于计算序列同一性百分比的不相同的残基。用于确定氨基酸或核酸序列同一性百分比的比对可以使用计算机程序以及例如在互联网上可公开获得的计算机程序以多种方式实现。优选地,可使用设定为默认参数可从nationalcenterforbiotechnologyinformation(ncbi)于http://www.ncbi.nlm.nih.gov/blast/bl2seq/wblast2.cgi获得的blast程序(tatiana等,femsmicrobiollett.,1999,174:247-250,1999)来获得肽或核苷酸序列的最佳比对并计算序列同一性的百分比。使用的缩写bp碱基对kb千碱基dna脱氧核糖核酸cdna互补dnadtt二硫苏糖醇fpp法尼基焦磷酸gc气相色谱iptg异丙基-d-硫代半乳糖-吡喃糖苷lb溶源性肉汤ms质谱mva甲羟戊酸pcr聚合酶链反应rna核糖核酸mrna信使核糖核酸mirna微rnasirna小干扰rnarrna核糖体rnatrna转移rna定义术语“多肽”是指连续聚合的氨基酸残基例如至少15个残基,至少30个残基,至少50个残基的氨基酸序列。在本文提供的一些实施方案中,多肽包含作为酶、其片段或其变体的氨基酸序列。术语“分离的”多肽是指通过任何方法或本领域已知方法(包括重组、生物化学和合成法)的组合从其天然环境中取出的氨基酸序列。术语“蛋白”是指任何长度的氨基酸序列,其中氨基酸被共价肽键所连接,并且包括寡肽、肽、多肽和全长蛋白,不管天然生成的还是合成的。术语“(+)-雪松醇合酶”、“(-)-罗汉柏烯合酶”、“(+)-雪松醇合酶活性”、“(-)-罗汉柏烯合酶活性”、“(+)-雪松醇合酶蛋白”以及“(-)-罗汉柏烯合酶蛋白”指的是能够将法呢基二磷酸酯(fpp)转化为(+)-雪松醇或(-)-罗汉柏烯的酶。术语“生物学功能”、“功能”、“生物学活性”或“活性”是指(+)-雪松醇合酶和(-)-罗汉柏烯合酶催化从fpp至(+)-雪松醇和(-)-罗汉柏烯的形成的能力。可互换使用的术语“核酸序列”、“核酸”和“多核苷酸”是指核苷酸的序列。核酸序列可以是任何长度的单链或双链脱氧核糖核苷酸或核糖核苷酸,并且包括基因、外显子、内含子、有义和反义互补序列、基因组dna、cdna、mirna、sirna、mrna、rrna、trna、重组核酸序列、分离的和纯化的天然产生的dna和/或rna序列、合成dna和rna序列、片段、引物和核酸探针的编码和非编码序列。本领域技术人员了解,rna的核酸序列相同于dna序列,差异在于胸腺嘧啶(t)被替代为尿嘧啶(u)。“分离的核酸”或“分离的核酸序列”被定义为一种核酸或核酸序列,其所处的环境与天然产生的核酸或核酸序列所处的环境不同。如本文使用的如应用于核酸的术语“天然产生的”是指在自然的细胞中发现的核酸。例如,存在于能够从自然源中分离出的生物体(例如生物体的细胞)中的核酸序列,并且该核酸序列未经人类在实验室进行特意的改造,那么该核酸序列是天然产生的。“重组核酸序列”是使用实验室方法(分子克隆)将来自多于一个源的遗传物质组合在一起所生成的核酸序列,由此创造出不是天然产生并且不能以其它方式在生物有机体中发现的核酸序列。“重组dna技术”是指一种用于制备重组核酸序列的分子生物学程序,例如描述于由weigel和glazebrook编的laboratorymanuals,2002coldspringharborlabpress;和sambrook等的1989coldspringharbor,ny:coldspringharborlaboratorypress。术语“基因”是指一种dna序列,其包含可操作地连接到适当调控区域(例如启动子)的被转录为rna分子(例如细胞中的mrna)的区域。因此,基因可以包含多个可操作地连接的序列,诸如启动子、5'前导序列(包含例如参与翻译初始化的序列)、cdna或基因组dna的编码区、内含子、外显子和/或3'非翻译序列(包含例如转录终止位点)。“嵌合基因”是指通常不能在自然物种中找到的任何基因,特别是这样一种基因,其中核酸序列存在在性质上彼此不相关联的一个或多个部分。例如,启动子在性质上与转录区的部分或全部或与另一调控区不相关联。术语“嵌合基因”应当被理解为包括表达构建体,其中启动子或转录调控序列被可操作地连接到一个或多个编码序列或反义(即有义链的反向互补链)或反向重复序列(有义和反义,由此rna转录物在转录后形成双链rna)。术语“嵌合基因”还包括这样的基因,其通过组合一个或多个编码序列的部分以产生新基因而获得。“3'urt”或“3'非翻译序列”(也称为“3'未翻译区”或“3'末端”)是指在基因编码序列的下游发现的核酸序列,其包含例如转录终止位点和(在大多数但非全部的真核mrna中)多聚腺苷酸化信号诸如aauaaa或其变体。在转录终止后,mrna转录物可以在多聚腺苷酸化信号的下游被切去并且可以添加poly(a)尾,其参与了mrna向翻译位点例如胞浆的运输。“基因的表达”包括基因的转录和mrna向蛋白质的翻译。过表达是指在转基因细胞或生物体中,以mrna水平、多肽和/或酶活性测量的基因产物的生产超过了相似遗传背景的非转化细胞或生物体中的生产水平。如本文中使用的“表达载体”是指使用分子生物学方法和重组dna技术工程化以将外来或外源dna递送到宿主细胞中的核酸分子。表达载体典型地包括适合转录核苷酸序列所需的序列。编码区通常编码目的蛋白,但是也可以编码rna,例如反义rna、sirna等。如本文所使用的“表达载体”包括任何线性的或环状的重组载体,包括但不限于病毒载体、噬菌体和质粒。本领域技术人员根据表达系统能够选择适合的载体。在一个实施方案中,表达载体包括可操作地连接到至少一个调控序列(其控制转录、翻译、起始和终止,诸如转录启动子、操纵子和增强子)或mrna核糖体结合位点和可选地包括至少一个选择标记的核酸。当调控序列功能性地涉及本文实施方案的核酸时,核苷酸序列是“可操作地连接的”。“调控序列”是指确定本文实施方案的核酸序列的表达水平、并且能够调控可操作地连接到该调控序列的核酸序列的转录速率的核酸序列。调控序列包含启动子、增强子、转录因子、启动子元素等。“启动子”是指控制编码序列的表达的核酸序列,其提供rna聚合酶用的结合位点以及适合转录所需的其它因子,包括但不限于转录因子结合位点、抑制子和活化子蛋白结合位点。术语启动子的含义包括术语“启动子调控序列”。启动子调控序列可以包括可能影响转录、rna加工或相关编码核酸序列的稳定性的上游和下游元素。启动子包括天然来源的和合成的序列。编码核酸序列通常位于启动子相对于以转录起始位点为起始的转录方向的下游。术语“组成型启动子”是指不受调控的启动子,其允许与其可操作地连接的核酸序列的持续转录。如本文使用的术语“可操作地连接”是指处于功能性关系的多核苷酸元素的连接。当核酸与另一核酸序列处于功能性关系时,那么该核酸是“可操作地连接的”。例如,如果启动子或者转录调控序列能够影响编码序列的转录,那么该启动子或者转录调控序列是可操作地连接到该编码序列的。可操作地连接意味着被连接的dna序列通常是邻接的。与启动子序列有关的核苷酸序列相对于要被转化的宿主生物体或细胞(例如植物、细菌或酵母细胞)可以是同源或异源来源的。所述序列还可以是完全或部分合成的。不管来源如何,与启动子序列相关的核酸序列将根据在结合到本文实施方案的多肽后所连接的启动子性质而表达或沉默。相关核酸在所有时间或替代地在特定时间在整个生物体中或在特定组织、细胞或细胞室中可以编码需要表达或抑制的蛋白。此种核苷酸序列特别地编码将所需表型性状赋予给由其改变或转化的宿主细胞或生物体的蛋白质。更特别地,相关核苷酸序列引起在生物体中生产(+)-雪松醇合酶或(-)-罗汉柏烯合酶。特别地,该核苷酸序列编码(+)-雪松醇合酶或(-)-罗汉柏烯合酶。“靶肽”是指一种氨基酸序列,其将蛋白质或多肽靶向细胞内细胞器(即,线粒体或质体)或细胞外空间(分泌信号肽)。编码靶肽的核酸序列可以被融合到编码蛋白或多肽的氨基末端(例如n-末端)的核酸序列,或者可以被用来替换原有的靶多肽。术语“引物”是指短的核酸序列,其被杂交到模板核酸序列并且被用于与该模板互补的核酸序列的聚合。如本文所使用的术语“宿主细胞”或“转化细胞”是指被改变以含有至少一种核酸分子(例如,编码所需蛋白或核酸序列的重组基团,该核酸序列在转录后产生有用于生产(+)-雪松醇和/或(-)-罗汉柏烯的(+)-雪松醇合酶蛋白或(-)-罗汉柏烯合酶蛋白)的细胞(或生物体)。宿主细胞特别地是细菌细胞、真菌细胞或植物细胞。宿主细胞可以含有重组基因,其已经被整合到宿主细胞的核或细胞器的基因组中。替代地,宿主可以含有染色体外的重组基因。同源序列包括直系或旁系同源序列。鉴别直系或旁系的方法包括现有技术中已知且在本文中描述的进化方法、序列相似性和杂交方法。旁系同源物来源于基因复制,其产生具有相似序列和相似功能的两种或更多种基因。旁系同源物典型地聚簇在一起并且通过基因在相关植物物种内的复制而形成。使用成对blast分析或在基因家族的进化分析过程中使用程序诸如clustal在类似基因的组中发现旁系同源物。在旁系同源物中,共有序列可以是相关基因中序列的识别特性并且具有基因的类似功能。直系或直系同源序列是彼此相似的序列,因为它们被发现在由共同的祖先传下的物种中。例如,具有共同祖先的植物物种已知含有相同类似序列和功能的许多酶。本领域技术人员能够识别直系同源序列并预测直系同源的功能,例如通过使用clustal或blast程序构建一个物种的基因族的进化树。术语“选择性标记物”是指在表达时能够被用来选择包括该选择性标记物的(多个)细胞的任何基因。以下描述了选择性标记物的例子。本领域技术人员了解不同的抗生素、杀真菌剂、营养缺陷型或除草剂选择标记物可适用于不同的目标物种。术语“生物体”是指任何非人多细胞或单细胞生物体。诸如植物或微生物。特别地,微生物是细菌、酵母、藻类或真菌。术语“植物”被互换地用来包括植物细胞,包括植物原生质体、植物组织、产生再生植株的植物细胞组织、或植物的部分、或植物器官诸如根、茎、叶、花、花粉、胚珠、胚、果实等。任何植物均可以用来实施本文实施方案的方法。在体外要与无环焦磷酸酯(例如fpp)接触的多肽可以通过使用标准蛋白或酶提取技术从表达它的任何有机体中提取得到。如果宿主生物体是将本文实施方案的多肽释放到培养基中的单细胞生物体或细胞,那么可以简单地从所述培养基收集所述多肽,例如,通过离心,可选地随后进行洗涤步骤和再悬浮于适合的缓冲液中。如果所述生物体或细胞将多肽累积于其细胞中,那么可以通过破裂或裂解所述细胞并且进一步从细胞裂解液中提取多肽来获得所述多肽。然后可以以分离形式或与其它蛋白质(例如从培养的细胞或微生物得到的粗蛋白质提取物)一起将具有(+)-雪松醇合酶活性和/或(-)-罗汉柏烯合酶活性的多肽以最佳ph悬浮在缓冲溶液中。如果足够,可以加入盐、dtt、无机阳离子和其他种类的酶辅因子,以优化酶的活性。可以将前体fpp加入到多肽悬浮液中,然后将其在最佳温度,例如15~40℃,特别是25~35℃,更特别是于30℃温育。温育后,可以通过标准分离程序,例如溶剂萃取和蒸馏,可选地在从溶液中除去多肽之后,从所温育的溶液中分离(+)-雪松醇和/或(-)-罗汉柏烯。根据另一具体实施方案,上述实施方案中的任一种的方法在体内进行。一方面,一个实施方案包括在有助于生产(+)-雪松醇或(-)-罗汉柏烯的条件下,培养能够生产fpp并能被转化以表达至少一种多肽的非人宿主生物体或细胞,其中该多肽包含与从seqidno:1、seqidno:2、seqidno:3、seqidno:4、seqidno:13和seqidno:14中选出的序列有至少70%相同的氨基酸序列,并且具有(+)-雪松醇合酶活性或(-)-罗汉柏烯合酶活性。根据一个更具体的实施方案,该方法还包括用编码多肽的至少一种核酸转化能够生产fpp的非人生物体或细胞,从而所述生物体表达所述多肽,其中该多肽包含与从seqidno:1、seqidno:2、seqidno:3、seqidno:4、seqidno:13和seqidno:14中选出的序列有至少70%相同的氨基酸序列,并且具有(+)-雪松醇合酶活性或(-)-罗汉柏烯合酶活性。本文提供的这些实施方案是特别有利的,因为可以在体内实施该方法而不预先分离多肽。该反应直接发生在经转化以表达所述多肽的生物体或细胞内。根据一个更具体的实施方案,本文使用的至少一种核酸包含通过修饰seqidno:5、seqidno:6、seqidno:7、seqidno:8、seqidno:9、seqidno:10、seqidno:11、seqidno:12、seqidno:15、seqidno:16、seqidno:17、seqidno:18和seqidno:19或其互补序列而已经获得的核苷酸序列。所述生物体或细胞旨在“表达”多肽,前提是所述生物体或细胞经转化以含有编码所述多肽的核酸,这种核酸被转录至mrna并且在所述宿主生物体或细胞中发现所述多肽。术语“表达”涵盖“异源表达”和“过表达”,后者是指mrna、多肽和/或酶活性的水平高于在非转化的生物体或细胞中所测量的水平。随后,在致力于此种经转化的非人宿主生物体或细胞的说明书部分中,将描述用于转化非人宿主生物体或细胞的适合方法的更详细说明。特定的生物体或细胞是指当其天然地产生fpp时、或当其不天然地产生fpp但是在使用本文所述的核酸转化之前或与使用所述核酸一起被转化以生产fpp时,“能够产生fpp”。经转化以比天然存在的生物体或细胞产生更高量的fpp的生物体或细胞也被“能够生产fpp的生物体或细胞”所涵盖。用于转化生物体(例如微生物)以便它们生产fpp的方法在本领域中是已知的。为了在体内进行本文的实施方式,在有助于(+)-雪松醇合酶和/或(-)-罗汉柏烯合酶的生产的条件下培养宿主生物体或细胞。因此,如果宿主是转基因植物,那么例如提供最佳的生长条件,诸如最佳光、水和营养条件。如果宿主是单细胞生物体,那么有助于(+)-雪松醇合酶和/或(-)-罗汉柏烯合酶的生产的条件可以包含向宿主的培养基中添加适合的辅因子。此外,可以选择培养基以便最大化(+)-雪松醇合酶活性和/或(-)-罗汉柏烯合酶活性。在下列实施例中以更详细的方式描述了最佳的培养条件。适合于在体内进行本文实施方案的方法的非人宿主生物体可以是任何非人多细胞或单细胞生物体。在一个特定的实施方案中,用来在体内进行本文实施方案的非人宿主生物体是植物、原核生物或真菌。可以使用任何植物、原核生物或真菌。特别有用的植物是天然产生高量萜烯的那些。在一个更具体的实施方案中,用来在体内进行本文实施方式的方法的非人宿主生物体是微生物。可以使用任何微生物,但根据甚至更具体的实施方案,所述微生物是细菌或酵母。更特别地,所述细菌是大肠杆菌(e.coli)并且所述酵母是酿酒酵母(saccharomycescerevisiae)。这些生物体的一些天然不生产或仅少量生产fpp。为了适合于进行本文实施方案的方法,这些生物体必须经转化以生产所述前体或大量生产所述前体。如上所说明的,它们可以在使用根据任一上述实施方案所述的核酸修饰之前或同时地被如此转化。也可以使用分离的高等真核细胞以代替完整生物体作为宿主,以在体内实施本文实施方案的方法。合适的真核细胞可以是任何非人细胞,但是特别是植物或真菌细胞。在另一具体的实施方案中,多肽包含:c.与从seqidno:1、seqidno:2、seqidno:3、seqidno:4、seqidno:13和seqidno:14中选出的多肽具有至少70%、75%、80%、85%、90%、95%、98%和/或99%的序列同一性的氨基酸序列;或者d.从seqidno:1、seqidno:2、seqidno:3、seqidno:4、seqidno:13和seqidno:14中选出的氨基酸序列。根据另一个具体实施方案,在本文所述的任一实施方案中使用的或由本文所述的核酸编码的具有(+)-雪松醇合酶活性和/或(-)-罗汉柏烯合酶活性的该至少一种多肽包含一种氨基酸序列,其是通过遗传工程获得的seqidno:1、seqidno:2、seqidno:3、seqidno:4、seqidno:13或seqidno:14的变体,条件是所述变体保持了其(+)-雪松醇合酶活性和/或(-)-罗汉柏烯合酶活性。如本文所用,多肽旨在是包含本文确定的氨基酸序列的多肽或肽片段以及截短的或变体多肽,前提是它们保持了如上所定义的它们的(+)-雪松醇合酶活性和/或(-)-罗汉柏烯合酶活性,并且它们与seqidno:1、seqidno:2、seqidno:3、seqidno:4、seqidno:13或seqidno:14的相应片段享有至少确定百分比的同一性。本文所述的多肽的片段可以占例如本文所述的多肽氨基酸序列的至少50%、60%、70%、80%、90%、95%或99%。变体多肽的例子是由交替mrna剪接事件或本文所述多肽的蛋白水解切割产生的天然存在的蛋白。归因于蛋白水解的变种包括例如在不同类型的宿主细胞中表达后n-或c-末端的差异,这是由于从本文实施方案的多肽蛋白水解去除一个或多个末端氨基酸引起的。本文实施方案还涵盖了如下文所描述的,通过天然或人工突变本文实施方案的核酸获得的核酸所编码的多肽。由在氨基和羧基末端融合额外的肽序列产生的多肽变体也可以用在本文实施方案的方法中。特别地,此种融合能够增强多肽的表达,有用于蛋白的纯化或改善多肽在期望环境或表达系统中的酶活性。例如,此种额外的肽序列可以是信号肽。因此,本文包括使用变体多肽(诸如通过与其它寡肽或多肽融合所获得的那些和/或连接到信号肽的那些)的方法。由与另一种功能肽(诸如来自萜烯生物合成路径的另一种蛋白)融合产生的多肽也可以有利地被用于本文实施方案的方法中。根据一个具体实施方案,多肽包含与从seqidno:1、seqidno:2、seqidno:3、seqidno:4、seqidno:13和seqidno:14中选出的序列有至少70%,特别是至少75%,特别是至少80%,特别是至少85%,特别是至少90%,特别是至少95%,特别是至少98%,甚至更特别是99%相同的氨基酸序列。根据一个具体实施方案,多肽包含与从seqidno:1、seqidno:2、seqidno:3、seqidno:4、seqidno:13和seqidno:14中选出的序列有至少75%,特别是至少80%,特别是至少85%,特别是至少90%,特别是至少95%,特别是至少98%,甚至更特别是99%相同的氨基酸序列。根据一个具体实施方案,多肽包含与从seqidno:1、seqidno:2、seqidno:3、seqidno:4、seqidno:13和seqidno:14中选出的序列有至少80%,特别是至少85%,特别是至少90%,特别是至少95%,特别是至少98%,甚至更特别是99%相同的氨基酸序列。根据一个具体实施方案,多肽包含与从seqidno:1、seqidno:2、seqidno:3、seqidno:4、seqidno:13和seqidno:14中选出的序列有至少85%,特别是至少90%,特别是至少95%,特别是至少98%,甚至更特别是99%相同的氨基酸序列。根据一个具体实施方案,多肽包含与从seqidno:1、seqidno:2、seqidno:3、seqidno:4、seqidno:13和seqidno:14中选出的序列有至少90%,特别是至少95%,特别是至少98%,甚至更特别是99%相同的氨基酸序列。根据一个具体实施方案,多肽包含与从seqidno:1、seqidno:2、seqidno:3、seqidno:4、seqidno:13和seqidno:14中选出的序列有至少95%,特别是至少98%,甚至更特别是99%相同的氨基酸序列。根据一个具体实施方案,多肽包含与从seqidno:1、seqidno:2、seqidno:3、seqidno:4、seqidno:13和seqidno:14中选出的序列有至少98%,甚至更特别是99%相同的氨基酸序列。根据一个具体实施方案,多肽包含与从seqidno:1、seqidno:2、seqidno:3、seqidno:4、seqidno:13和seqidno:14中选出的序列有至少99%相同的氨基酸序列。根据一个具体实施方案,多肽包含与从seqidno:1有至少70%,特别是至少75%,特别是至少80%,特别是至少85%,特别是至少90%,特别是至少95%,特别是至少98%,甚至更特别是99%相同的氨基酸序列。根据一个具体实施方案,多肽包含与从seqidno:2有至少70%,特别是至少75%,特别是至少80%,特别是至少85%,特别是至少90%,特别是至少95%,特别是至少98%,甚至更特别是99%相同的氨基酸序列。根据一个具体实施方案,多肽包含与从seqidno:3有至少70%,特别是至少75%,特别是至少80%,特别是至少85%,特别是至少90%,特别是至少95%,特别是至少98%,甚至更特别是99%相同的氨基酸序列。根据一个具体实施方案,多肽包含与从seqidno:4有至少70%,特别是至少75%,特别是至少80%,特别是至少85%,特别是至少90%,特别是至少95%,特别是至少98%,甚至更特别是99%相同的氨基酸序列。根据一个具体实施方案,多肽包含与从seqidno:13有至少70%,特别是至少75%,特别是至少80%,特别是至少85%,特别是至少90%,特别是至少95%,特别是至少98%,甚至更特别是99%相同的氨基酸序列。根据一个具体实施方案,多肽包含与从seqidno:14有至少70%,特别是至少75%,特别是至少80%,特别是至少85%,特别是至少90%,特别是至少95%,特别是至少98%,甚至更特别是99%相同的氨基酸序列。一方面,当如本文所述多肽与fpp接触时,具有(+)-雪松醇合酶活性和/或(-)-罗汉柏烯合酶活性的多肽可具有对(+)-雪松醇或(-)-罗汉柏烯产物的特定选择性。本文所使用的对(+)-雪松醇或(-)-罗汉柏烯产物的选择性是指与倍半萜产物的总量相比,所生产的(+)-雪松醇或(-)-罗汉柏烯产物的量,并且通常表示为百分比。对于特定的基因表达系统,例如大肠杆菌表达系统可以给出选择性。一方面,多肽可以生产(+)-雪松醇作为主要的倍半萜产物。例如,多肽可以具有约70~90%,例如70%或更多,72%或更多,73%或更多,74%或更多,75%或更多,78%或更多,79%或更多,82%或更多,84%或更多,86%或更多或88%或更多的(+)-雪松醇的选择性。这样的选择性可以在例如在本实施例中描述的大肠杆菌表达系统中获得。一方面,多肽可以生产(+)-雪松醇,而不存在(-)-罗汉柏烯。一方面,多肽可以生产(-)-罗汉柏烯作为主要的倍半萜产物。例如,多肽可以具有约15~60%,例如18%或更多,20%或更多,25%或更多,26%或更多,30%或更多,35%或更多,40%或更多,44%或更多,45%或更多,50%或更多,53%或更多,55%或更多,57%或更多的(-)-罗汉柏烯产物的选择性。这样的选择性可以在例如在本实施例中描述的大肠杆菌表达系统中获得。一方面,多肽可以生产(-)-罗汉柏烯,而不存在(+)-雪松醇,或者除了(-)-罗汉柏烯之外还可以生产(+)-雪松醇,但是量较少。一方面,本文描述的包含如下序列的多肽生产(+)-雪松醇作为主要的倍半萜产物,并可具有如上所述的(+)-雪松醇的选择性:(i)与从seqidno:1、seqidno:2、seqidno:3和seqidno:13中选出的序列具有至少70%、75%、80%、85%、90%、95%、98%和/或99%的序列同一性的氨基酸序列;或者(ii)从seqidno:1、seqidno:2、seqidno:3和seqidno:13中选出的氨基酸序列;这种多肽可以生产(+)-雪松醇,而不存在(-)-罗汉柏烯。例如,具有seqidno:13中的氨基酸序列的本文所述的porb1多肽可以在大肠杆菌表达系统中获得约88%的(+)-雪松醇选择性而不存在(-)-罗汉柏烯。例如,具有seqidno:1中的氨基酸序列的本文所述的jvcp1206-4多肽可以在大肠杆菌表达系统中获得约75%的(+)-雪松醇选择性而不存在(-)-罗汉柏烯。例如,具有seqidno:3中的氨基酸序列的本文所述的jvcp1206-6多肽可以在大肠杆菌表达系统中获得约84%的(+)-雪松醇选择性而不存在(-)-罗汉柏烯。一方面,本文描述的包含如下序列的多肽生产(-)-罗汉柏烯作为主要的倍半萜产物,并可具有如上所述的(-)-罗汉柏烯的选择性:(i)与从seqidno:4和seqidno:14中选出的序列具有至少70%、75%、80%、85%、90%、95%、98%和/或99%的序列同一性的氨基酸序列;或者(ii)从seqidno:4和seqidno:14中选出的氨基酸序列;这种多肽例如可以生产(-)-罗汉柏烯,而不存在(+)-雪松醇,或者除了(-)-罗汉柏烯之外还可以生产(+)-雪松醇,但是量较少。例如,具有seqidno:14中的氨基酸序列的本文所述的por2-3-5多肽可以在大肠杆菌表达系统中获得约57%的(-)-罗汉柏烯选择性。例如,具有seqidno:4中的氨基酸序列的本文所述的jvcp1206-5多肽可以在大肠杆菌表达系统中获得约26%的(-)-罗汉柏烯选择性。如上所述,编码本文实施方案的多肽的核酸是当该方法在体内进行时修饰旨在使用的非人宿主生物体或细胞的有用工具。因此,本文还提供了编码根据任何上述实施方案的多肽的核酸。根据一个具体实施方案,核酸包含与从seqidno:5、seqidno:6、seqidno:7、seqidno:8、seqidno:9、seqidno:10、seqidno:11、seqidno:12、seqidno:15、seqidno:16、seqidno:17、seqidno:18或seqidno:19或其互补序列中选出的序列有至少70%、特别是至少75%、特别是至少80%、特别是至少85%、特别是至少90%、特别是至少95%、特别是至少98%和特别是至少99%相同的核苷酸序列。根据一个具体实施方案,核酸包含与从seqidno:5、seqidno:6、seqidno:7、seqidno:8、seqidno:9、seqidno:10、seqidno:11、seqidno:12、seqidno:15、seqidno:16、seqidno:17、seqidno:18或seqidno:19或其互补序列中选出的序列有至少75%、特别是至少80%、特别是至少85%、特别是至少90%、特别是至少95%、特别是至少98%和更特别是至少99%相同的核苷酸序列。根据一个具体实施方案,核酸包含与从seqidno:5、seqidno:6、seqidno:7、seqidno:8、seqidno:9、seqidno:10、seqidno:11、seqidno:12、seqidno:15、seqidno:16、seqidno:17、seqidno:18或seqidno:19或其互补序列中选出的序列有至少80%、特别是至少85%、特别是至少90%、特别是至少95%、更特别是至少98%和甚至更特别是至少99%相同的核苷酸序列。根据一个具体实施方案,核酸包含与从seqidno:5、seqidno:6、seqidno:7、seqidno:8、seqidno:9、seqidno:10、seqidno:11、seqidno:12、seqidno:15、seqidno:16、seqidno:17、seqidno:18或seqidno:19或其互补序列中选出的序列有至少85%、特别是至少90%、特别是至少95%、更特别是至少98%和甚至更特别是至少99%相同的核苷酸序列。根据一个具体实施方案,核酸包含与从seqidno:5、seqidno:6、seqidno:7、seqidno:8、seqidno:9、seqidno:10、seqidno:11、seqidno:12、seqidno:15、seqidno:16、seqidno:17、seqidno:18或seqidno:19或其互补序列中选出的序列有至少90%、特别是至少95%、更特别是至少98%和甚至更特别是至少99%相同的核苷酸序列。根据一个具体实施方案,核酸包含与从seqidno:5、seqidno:6、seqidno:7、seqidno:8、seqidno:9、seqidno:10、seqidno:11、seqidno:12、seqidno:15、seqidno:16、seqidno:17、seqidno:18或seqidno:19或其互补序列中选出的序列有至少95%、更特别是至少98%和甚至更特别是至少99%相同的核苷酸序列。根据一个具体实施方案,核酸包含与从seqidno:5、seqidno:6、seqidno:7、seqidno:8、seqidno:9、seqidno:10、seqidno:11、seqidno:12、seqidno:15、seqidno:16、seqidno:17、seqidno:18或seqidno:19或其互补序列中选出的序列有至少98%和甚至更特别是至少99%相同的核苷酸序列。根据一个具体实施方案,核酸包含与从seqidno:5、seqidno:6、seqidno:7、seqidno:8、seqidno:9、seqidno:10、seqidno:11、seqidno:12、seqidno:15、seqidno:16、seqidno:17、seqidno:18或seqidno:19或其互补序列中选出的序列有至少99%相同的核苷酸序列。根据一个具体实施方案,核酸包含与seqidno:5或其互补序列有至少70%、特别是至少75%、特别是至少80%、特别是至少85%、特别是至少90%、特别是至少95%、特别是至少98%和特别是至少99%相同的核苷酸序列。根据一个具体实施方案,核酸包含与seqidno:6或其互补序列有至少70%、特别是至少75%、特别是至少80%、特别是至少85%、特别是至少90%、特别是至少95%、特别是至少98%和特别是至少99%相同的核苷酸序列。根据一个具体实施方案,核酸包含与seqidno:7或其互补序列有至少70%、特别是至少75%、特别是至少80%、特别是至少85%、特别是至少90%、特别是至少95%、特别是至少98%和特别是至少99%相同的核苷酸序列。根据一个具体实施方案,核酸包含与seqidno:8或其互补序列有至少70%、特别是至少75%、特别是至少80%、特别是至少85%、特别是至少90%、特别是至少95%、特别是至少98%和特别是至少99%相同的核苷酸序列。根据一个具体实施方案,核酸包含与seqidno:9或其互补序列有至少70%、特别是至少75%、特别是至少80%、特别是至少85%、特别是至少90%、特别是至少95%、特别是至少98%和特别是至少99%相同的核苷酸序列。根据一个具体实施方案,核酸包含与seqidno:10或其互补序列有至少70%、特别是至少75%、特别是至少80%、特别是至少85%、特别是至少90%、特别是至少95%、特别是至少98%和特别是至少99%相同的核苷酸序列。根据一个具体实施方案,核酸包含与seqidno:11或其互补序列有至少70%、特别是至少75%、特别是至少80%、特别是至少85%、特别是至少90%、特别是至少95%、特别是至少98%和特别是至少99%相同的核苷酸序列。根据一个具体实施方案,核酸包含与seqidno:12或其互补序列有至少70%、特别是至少75%、特别是至少80%、特别是至少85%、特别是至少90%、特别是至少95%、特别是至少98%和特别是至少99%相同的核苷酸序列。根据一个具体实施方案,核酸包含与seqidno:15或其互补序列有至少70%、特别是至少75%、特别是至少80%、特别是至少85%、特别是至少90%、特别是至少95%、特别是至少98%和特别是至少99%相同的核苷酸序列。根据一个具体实施方案,核酸包含与seqidno:16或其互补序列有至少70%、特别是至少75%、特别是至少80%、特别是至少85%、特别是至少90%、特别是至少95%、特别是至少98%和特别是至少99%相同的核苷酸序列。根据一个具体实施方案,核酸包含与seqidno:17或其互补序列有至少70%、特别是至少75%、特别是至少80%、特别是至少85%、特别是至少90%、特别是至少95%、特别是至少98%和特别是至少99%相同的核苷酸序列。根据一个具体实施方案,核酸包含与seqidno:18或其互补序列有至少70%、特别是至少75%、特别是至少80%、特别是至少85%、特别是至少90%、特别是至少95%、特别是至少98%和特别是至少99%相同的核苷酸序列。根据一个具体实施方案,核酸包含与seqidno:19或其互补序列有至少70%、特别是至少75%、特别是至少80%、特别是至少85%、特别是至少90%、特别是至少95%、特别是至少98%和特别是至少99%相同的核苷酸序列。本文实施方案的核酸可以被定义为包括单链形式或双链形式的脱氧核糖核苷酸或核糖核苷酸聚合物(dna和/或rna)。术语“核苷酸序列”应当被理解为包含多核苷酸分子或独立片段形式的寡核苷酸分子或作为较大核酸的组件。本文实施方案的核酸还涵盖某些分离的核苷酸序列,包括基本上不含内源性污染物质的那些分离的核苷酸序列。本文实施方案的核酸可以是截短的,前提是其编码如上所述的本文所涵盖的多肽。在一个实施方案中,本文实施方案的核酸可以天然存在于植物例如北美圆柏(juniperusvirginiana)、金塔柏(platycladusorientalis'beverleyensis')或侧柏(platycladusorientalis)或其他物种中,或者通过修饰seqidno:5、seqidno:6、seqidno:7、seqidno:8、seqidno:9、seqidno:10、seqidno:11、seqidno:12、seqidno:15、seqidno:16、seqidno:17、seqidno:18或seqidno:19或其互补序列而获得。突变可以是这些核酸的任何种类的突变,诸如点突变、缺失突变、插入突变和/或移码突变。可以制备变体核酸以便使其核苷酸序列适应特定的表达系统。例如,如果氨基酸由特定的密码子编码,已知细菌表达系统能更有效地表达多肽。由于遗传密码的简并性,多于一个密码子可以编码相同的氨基酸序列,多个核酸序列能够编码相同的蛋白或多肽,所有这些dna序列均被涵盖在本文实施方案中。在适当的情况下,编码(+)-雪松醇合酶和/或(-)-罗汉柏烯合酶的核酸序列可以被优化以增加在宿主细胞中的表达。例如,可以使用特定的密码子通过宿主来合成本文实施方案的核苷酸以提高表达。用于转化适于在体内进行本文实施方案的方法的宿主生物体或细胞的另一重要工具是表达载体,其包含根据本文实施方案的任一实施方案所述的核酸。因此,本文还提供此种载体。如下文进一步披露的,本文提供的表达载体可以被用在用于制备遗传转化的宿主生物体和/或细胞的方法中,用在含有本文实施方案的核酸的宿主生物体和/或细胞中,以及用在用于制备具有(+)-雪松醇合酶活性和(-)-罗汉柏烯合酶活性的多肽的方法中。经转化以含有本文实施方案的至少一种核酸以便异源表达或过表达本文实施方案的至少一种多肽的重组非人宿主生物体和细胞,也是实施本文实施方案的方法的非常有用的工具。因此,本文提供此种非人宿主生物体和细胞。根据任一上述实施方案所述的核酸可以被用来转化非人宿主生物体和细胞,并且所表达的多肽可以是任何上述的多肽。本文实施方案的非人宿主生物体可以是任何非人多细胞或单细胞生物体。在特定的实施方案中,所述非人宿主生物体是植物、原核生物或真菌。任何植物、原核生物或真菌均适于根据本文提供的方法进行转化。特别有用的植物是天然生产高量萜烯的那些植物。在更特别的实施方案中,所述非人宿主生物体是微生物。任何微生物均适合在本文中使用,但根据甚至更特别的实施方案,所述微生物是细菌或酵母。更特别地,所述细菌是大肠杆菌,而所述酵母是酿酒酵母。除了完整的生物体之外,分离的高等真核细胞也可被转化。作为高等真核细胞,我们在此指的是除酵母细胞之外的任何非人真核细胞。特别是,高等真核细胞是植物细胞或真菌细胞。通过附接共价或非共价连接到多肽主链的修饰基团,变体也可以不同于本文实施方案的多肽。通过引入n-连接的或o-连接的糖基化位点、和/或添加半胱氨酸残基,所述变体还包括与本文提供的多肽不同的多肽。本领域技术人员能够了解如何改造氨基酸序列和保留生物活性。可以使用各种方法确定任何(+)-雪松醇合酶和/或(-)-罗汉柏烯合酶蛋白、变体或片段的功能性或活性。例如,在植物、细菌或酵母细胞中瞬时或稳定的过表达可以被用来测试蛋白是否具有活性,即从fpp前体生产(+)-雪松醇和/或(-)-罗汉柏烯。可以在微生物表达系统(诸如本文提供的实施例描述的检定)中评估(+)-雪松醇合酶活性和/或(-)-罗汉柏烯合酶活性。本文的实施方案提供本文实施方案的多肽,其被用于在体外或体内使fpp前体与本文实施方案的多肽接触而生产(+)-雪松醇和/或(-)-罗汉柏烯的方法中。本文还提供一种分离的重组或合成多核苷酸,其编码本文提供的多肽或变体多肽。本文提供的实施方案包括但不限于cdna、基因组dna和rna序列。在本文中,将编码(+)-雪松醇合酶和/或(-)-罗汉柏烯合酶或其变体的任何核酸序列称为(+)-雪松醇合酶和/或(-)-罗汉柏烯合酶编码序列。本领域技术人员清楚的是,基因(包括本文实施方案的多核苷酸)可通过本领域已知的方法基于可获得的核苷酸序列信号(诸如在所附的序列表中发现的)进行克隆。这些包括例如设计表示此种基因的侧翼序列的dna引物,所述基因的一种是在有义取向上产生的并且其初始化有义链的合成,而另一种以反向互补的方式创造并且产生反义链。诸如在聚合酶链反应中使用的那些热稳定的dna聚合酶通常被用来进行此种实验。替代地,表示基因的dna序列可以被化学合成并且随后导入到dna载体分子中,所述dna载体分子可以通过例如相容性细胞(诸如大肠杆菌)来繁殖。本文提供的是由seqidno:5、seqidno:6、seqidno:7、seqidno:8、seqidno:9、seqidno:10、seqidno:11、seqidno:12、seqidno:15、seqidno:16、seqidno:17、seqidno:18或seqidno:19的突变获得的核酸序列,这种突变可以被常规地制得。对于本领域技术人员言清楚的是,可以向这些的dna序列中导入一个或多个核苷酸的突变、缺失、插入和/或替换。为了测试根据本文实施方案所述的变体dna序列的功能,目标序列被可操作地连接到选择性或筛选性标记基因,并且在原生质体的瞬时表达分析中或在稳定转化植物中测试报告基因的表达。本领域技术人员能够理解,能够推动表达的dna序列被构建为模块。因此,来自较短dna片段的表达水平可能不同于来自最长片段的表达水平,并且能够彼此不同。本文还提供了编码本文提供的(+)-雪松醇合酶和/或(-)-罗汉柏烯合酶蛋白的核酸序列的功能性等同物,即在严格条件下杂交到seqidno:5、seqidno:6、seqidno:7、seqidno:8、seqidno:9、seqidno:10、seqidno:11、seqidno:12、seqidno:15、seqidno:16、seqidno:17、seqidno:18或seqidno:19的核酸序列的核苷酸序列。本领域技术人员能够想到用于识别其它有机体中同源序列的方法以及用于检测同源序列之间的序列同一性百分比的方法(在本文的定义部分确定)。本文提供的替代实施方案提供一种改变宿主细胞中基因表达的方法。例如,在某些背景下(例如,在昆虫叮咬或蛰伤之后或在暴露于一定的温度下),在宿主细胞或宿主有机体中可以增强或过表达或诱导本文实施方案的多核苷酸。本文提供的多核苷酸的表达的改变还会产生“异位表达”,其在改变的以及在对照或野生型有机体中是一种不同的表达模式。表达的改变是由本文实施方案的多肽与外源性或内源性调节剂的接触而发生的或者是由于多肽的化学改造导致的。所述术语还指本文实施方案的多核苷酸的改变的表达模式,其被改变至低于检测水平或者完全被抑制活性。在一个实施方案中,在单一宿主中共同表达多种(+)-雪松醇合酶和/或(-)-罗汉柏烯合酶编码核酸序列,特别地在不同启动子的控制下进行。替代地,在单个转化载体上可以存在多种(+)-雪松醇合酶和/或(-)-罗汉柏烯合酶蛋白编码核酸序列,或者可以同时使用单独的载体进行共转化并且选择包含两个嵌合基因的转化体。本文实施方案的编码(+)-雪松醇合酶和/或(-)-罗汉柏烯合酶蛋白的核酸序列可以被插入到表达载体中和/或被包含在插入到表达载体中的嵌合基因中,以便在宿主细胞或宿主有机体中生产(+)-雪松醇合酶和/或(-)-罗汉柏烯合酶蛋白。用于将转基因插入到宿主细胞的基因组中的载体在本领域中是众所周知的,并且包括质粒、病毒、粘粒和人工染色体。其中插入了嵌合基因的二元或共整合载体也被用来转化宿主细胞。本文提供的实施方案提供重组表达载体,其包含编码(+)-雪松醇合酶和/或(-)-罗汉柏烯合酶的核酸序列、或可操作地连接到缔合核酸序列(诸如例如启动子序列)的包含编码(+)-雪松醇合酶和/或(-)-罗汉柏烯合酶的核酸序列的嵌合基因。或者,启动子序列可以已经存在于载体中,以便待转录的核酸序列插入到该启动子序列的载体下游。载体通常被工程化以具有复制起点、多克隆位点和选择性标记物。一方面,(+)-雪松醇和(-)-罗汉柏烯可由合酶产物纯化而得。通过本文所述的任何方法生产的(+)-雪松醇和(-)-罗汉柏烯可以转化为衍生物,例如但不限于烃、酯、酰胺、糖苷、醚、环氧化物、醛、酮、醇、二醇、缩醛或缩酮。可以通过化学方法例如但不限于氧化,还原,烷基化,酰化,脱水和/或重排来获得(+)-雪松醇和(-)-罗汉柏烯衍生物。(+)-雪松醇和(-)-罗汉柏烯的化学转化的例子可见于charless.cell.afragrantintroductiontoterpenoidchemistry.theroyalsocietyofchemistry,2003.page163-172;g.ohloff,w.pickenhagen,p.kraft.scentandchemistry–themolecularworldofodors,verlaghelveticachimicaacta,zurich,2011,page172-174;us007615525、us20120077722、us3845132或wo2005083045。或者,可以使用生化方法通过使(+)-雪松醇或(-)-罗汉柏烯与酶例如但不限于氧化还原酶、单加氧酶、双加氧酶、转移酶接触来获得(+)-雪松醇和(-)-罗汉柏烯衍生物。生物化学转化可以在体外使用分离的酶或在体内使用全细胞进行。例如,生产(+)-雪松醇和(-)-罗汉柏烯的相同宿主生物或细胞可以被工程化以表达生产衍生物所需的酶。(+)-雪松醇和(-)-罗汉柏烯的生物化学转化的例子可见于abraham,w.r.,p.washausen,andk.kieslich.1987.z.naturforsch.42c,414-419;takigawah.,kubotah.,sonoharah.,okudam.,tanakas.,fujikuray.anditos.novel.1993,environmicrobiol.59(5),1336–1341;lamare,v.,j.d.fourneron,andr.furstoss.1987,tetrahedronlett.28,6269-6272;lamare,v.,andr.furstoss.1990,tetrahedron46.4109-132;sakamakih1,kitanakas,chaiw,hayashiday,takagiy,horiuchica.2001.j.nat.prod.64(5).630-631。本文进一步提供的是选自表1所列化合物的(+)-雪松醇衍生物。表1(雪松醇衍生物的例子)本文进一步提供的是选自表2所列化合物的(-)-罗汉柏烯衍生物。表2(罗汉柏烯衍生物的例子)本文还提供根据本文所述方法生产的包含(+)-雪松醇和(-)-罗汉柏烯或其衍生物的产品。以下实施例仅是说明性的,并不意图限制本文提供的权利要求或实施方案的范围。实施例1北美圆柏(juniperusvirginiana)植物材料和根转录组测序。北美圆柏(juniperusvirginiana)的种子从b&tworldseeds(aigues-vives,france)获得。种子直接在0.5升盆中的土壤中发芽。收集一至两岁龄的植物用于分析代谢物的组成和转录组分析。将植物从盆中取出,根部用自来水冲洗。将地表部分和根分离并在液氮中冷冻。首先使用waringblender(waringlaboratory,torrington,usa)在液氮中将组织大致切碎,然后使用研钵和研杵研磨成细粉末。用过量的mtbe(甲基叔丁基醚)提取地表和地下部分的样品,并通过gcms进行分析。分析在连接到agilent5975质谱检测器的agilent6890系列gc系统上进行。gc配备0.25mm内径×30mdb-1ms毛细管柱(agilent)。载气为he,恒定流速为1ml/min。初始烘箱温度为50℃(保持1分钟),然后以10℃/分钟的梯度升至300℃。产品的鉴定是基于质谱和保留指数与真实标准和内部数据库的比较。分析表明,雪松醇只存在于根部,而不是在地表部分(图1)。因此,取北美圆柏(j.virginiana)植物的根用于转录组分析。按照kolosova等人(kolosovan,millerb,ralphs,ellisbe,douglasc,ritlandk,andbohlmannj,isolationofhigh-qualityrnafromgymnospermandangiospermtrees.j.biotechniques,36(5),821-4,2004)所描述的程序提取总rna,具有以下修改。将10毫升体积的提取缓冲液用于1克磨碎的组织,并用2%(w/v)的pvp(聚乙烯吡咯烷酮,sigma-aldrich)补充提取缓冲液。对于ctab(十六烷基三甲基溴化铵,sigma-aldrich)提取,将核酸沉淀物(pellet)重悬于2mlte缓冲液(10mmtris-hcl,ph8,1mmedta)中,并用2ml5mnacl和1ml10%ctab进行提取。对于异丙醇沉淀,核酸沉淀物溶解在500μlte中。将最终的rna沉淀物重新悬浮于无菌蒸馏水中。使用illuminatotalrna-seq技术和illuminahiseq2000测序仪对根转录组进行测序。共生成了16.2百万个2×100bp配对读出物(pairedreads)。使用velvetdenovo基因组汇编器(http://www.ebi.ac.uk/~zerbino/velvet/)和oases软件(http://www.ebi.ac.uk/~zerbino/oases/)组装读出物。共组装了46,644个平均大小为1,241bp的重叠群(contigs)。使用tblastn算法(altschul等,j.mol.biol.215,403-410,1990)搜索重叠群并使用已知倍半萜合酶的氨基酸序列作为查询。这种方法允许检测138种不同的萜合酶编码序列。在进一步分选数据后,根据与已知倍半萜合酶的氨基酸序列同源性,保留17个全长序列。实施例2北美圆柏(juniperusvirginiana)倍半萜合酶的功能表达。密码子优化版本的选定的推定萜编码序列在体外合成并克隆到pj411表达质粒(dna2.0,menlopark,ca,usa)中。在krx大肠杆菌细胞(promega)中进行北美圆柏(j.virginiana)萜合酶的异源表达。将转化细胞的单个菌落用于接种5mllb培养基。在37℃下温育5至6小时后,将培养物转移到20℃培养箱中,并保持1小时以达平衡。然后通过加入1mmiptg和0.2%鼠李糖来诱导蛋白质的表达,并将培养物在20℃下温育过夜。第二天,通过离心收集细胞,重悬于0.1体积的50mmmopsoph7,10%甘油中,通过超声处理来裂解。通过离心(以20,000g进行30分钟)清除提取物,并将含有可溶性蛋白质的上清液用于进一步实验。将含有重组蛋白的粗制大肠杆菌蛋白提取物用于表征酶活性。在10~100μm的法呢基二磷酸酯(fpp,sigma)和0.1~0.5mg粗蛋白的存在下,在2ml的50mmmopsoph7,10%甘油、1mmdtt、10mmmgcl2中进行测定。将管在30℃下温育12~24小时,并用一体积的戊烷萃取两次。在氮气流下浓缩后,通过gc和gc-ms分析提取物,并与对照蛋白质的测定提取物进行比较。通过如实施例1所述的gcms进行由酶形成的产物的分析。在这些条件下,除了几种其他倍半萜产物之外,四种重组萜合酶还生产出雪松醇。因此,jvcp1206-3、jvcp1206-4和jvcp1206-6生产出萜的混合物,其中雪松醇占所生产的总倍半萜化合物的至少70~80%。jvcp1206-5酶生产了一种混合物,其中(-)-罗汉柏烯是主要产物,而雪松醇占总倍半萜化合物的10%(图2)。实施例3使用重组的北美圆柏(juniperusvirginiana)倍半萜合酶在工程细胞中体内生产(+)-雪松醇和(-)-罗汉柏烯合酶。为了评估雪松醇和罗汉柏烯在异源细胞中的体内生产,用包含实施例2中鉴定的四种北美圆柏(j.virginiana)倍半萜合酶之一的pj411(pj411-jvcp1206-4、pj411-jvcp1206-3、pj411-jvcp1206-6和pj411-jvcp1206-5)质粒转化大肠杆菌细胞,并评价来自内源fpp池的倍半萜的生产。为了提高细胞的生产力,异源fpp合酶和来自完整异源甲羟戊酸(mva)途径的酶也在相同的细胞中表达。包含fpp合酶基因和完整mva途径的基因的表达质粒的构建描述于专利wo2013064411或schalk等人(2013)j.am.chem.soc.134,18900-18903。简言之,表达质粒制备成含有由编码完整甲羟戊酸途径的酶的基因组成的两个操纵子。由大肠杆菌乙酰乙酰辅酶a硫解酶(atob)、金黄色葡萄球菌hmg-coa合酶(mvas)、金黄色葡萄球菌hmg-coa还原酶(mvaa)和酿酒酵母fpp合酶(erg20)基因组成的第一合成操纵子为在体外合成(dna2.0,menlopark,ca,usa),并连接到ncoi-bamhi消化的pacycduet-1载体(invitrogen)中,得到pacyc-29258。从肺炎链球菌(atccbaa-334)的基因组dna扩增出含有甲羟戊酸激酶(mvak1)、磷酸甲羟戊酸激酶(mvak2)、甲羟戊酸二磷酸脱羧酶(mvad)和异戊烯基二磷酸异构酶(idi)的第二操纵子,连接进入pacyc-29258的第二多克隆位点,提供质粒pacyc-29258-4506。因此,该质粒含有编码从乙酰辅酶a到fpp的生物合成途径的所有酶的基因。将krx大肠杆菌细胞(promega)与质粒pacyc-29258-4506和质粒pj411-jvcp1206-4、pj411-jvcp1206-3、pj411-jvcp1206-6或pj411-jvcp1206-5共转化。在羧苄青霉素(50μg/ml)和氯霉素(34μg/ml)lb-琼脂糖平板上选择所转化的细胞。使用单个菌落接种补充有相同抗生素的5ml液体lb培养基。将培养物在37℃下温育过夜。第二天,用0.2ml过夜培养物接种2ml补充有相同抗生素的tb培养基。在37℃下温育6小时后,将培养物冷却至28℃,向每个管中加入0.1mmiptg和0.2%鼠李糖。培养物在28℃下温育48小时。然后将培养物用2体积的mtbe萃取两次,将有机相浓缩至500μl,并通过如上实施例1所述的gc-ms进行分析。在这种体内条件下,四种倍半萜合酶生产与体外测定中(+)-雪松醇相同比例的倍半萜混合物:对于jvcp1206-3、jvcp1206-4和jvcp1206-6为70~80%的(+)-雪松醇,对于jvcp1206-5为10%(图3)。使用jvcp1206-5,(-)-罗汉柏烯是所生产出的倍半萜混合物的主要产物。使用这些工程化的大肠杆菌细胞,使用较多(1l)培养物来生产较大量的由这些酶生产的倍半萜产物混合物。通过快速色谱法在硅胶柱上从产物混合物中纯化出(+)-雪松醇。通过nmr分析获得足够的量来确认结构。使用brukeravance500mhz光谱仪测量旋光度。[α]d20=+10.6°(0.85%,chcl3)的值符合文献,并证实了(+)-雪松醇的生产。实施例4四种北美圆柏(j.virginiana)雪松醇合酶的序列比较。使用clustalw程序对四种北美圆柏(j.virginiana)雪松醇合酶的氨基酸序列进行比对,并从比对中推导出序列同一性。四种雪松醇合酶的序列同一性如下表所示。jvcp1206-3jvcp1206-4jvcp1206-6jvcp1206-5jvcp1206-3id9798.693.4jvcp1206-497id98.492.2jvcp1206-698.698.4id93.7jvcp1206-592.492.293.7id实施例5雪松木植物材料来源和叶片转录组测序。从中国浙江省杭州市收集了金塔柏(platycladusorientalis'beverleyensis')和侧柏(platycladusorientalis)植物材料。为了确定金塔柏(p.orientalis'beverleyensis')(样品编号:pnli20141232)和侧柏(p.orientalis)(样品编号:pnli20141243)是否含有(+)-雪松醇和(-)-罗汉柏烯,分别用二氯甲烷提取它们的新鲜叶子来做化学分析。通过gc/ms分析提取物,gc/ms分析参数描述如下:agilent6890系列gc系统配有db1-ms柱30m×0.25mm×0.25μm膜厚(p/n122-0132,j&wsciencesinc.,folsom,ca)并与5975系列质谱联用。载气为恒定流量0.7ml/min的氦气。注射为分流(1:25)模式,注射器温度设定在250℃。烘箱温度程序升温从50℃(保持5分钟)以5℃/分钟升至300℃,然后以50℃/分钟升至340℃并保持3分钟。产品鉴定基于质谱和保留指数。gc/ms分析表明,金塔柏(p.orientalis'beverleyensis')的叶子在其总挥发倍半萜中含有37%的(+)-雪松醇(图5和图6),而侧柏(p.orientalis)的叶子在其总挥发倍半萜中含有11%的(-)-罗汉柏烯(图7和图8)。将金塔柏(p.orientalis'beverleyensis')和侧柏(p.orientalis)的新鲜叶片用于转录组分析。使用rneasyplantminikit(qiagen,germany)提取总rna。使用ultratmrnalibraryprepkitforillumina(neb,usa)和truseqpeclusterkit(illumina,usa)处理这些总rna样品,然后在illuminahiseq2500测序仪上测序。对于金塔柏(p.orientalis'beverleyensis')和侧柏(p.orientalis)分别生成了17百万个和22.6百万个2×150bp配对末端读出物。使用trinity(http://trinityrnaseq.sf.net/)软件分别组装来自金塔柏(p.orientalis'beverleyensis')和侧柏(p.orientalis)的读出物。从金塔柏(p.orientalis'beverleyensis')和侧柏(p.orientalis)分别获得了带有1564bp的n50的58300个独立基因(unigenes)和带有1602bp的n50的62252个独立基因。这些独立基因用interproscan软件(http://www.ebi.ac.uk/tools/pfa/iprscan/)进行注释。使用来自现有技术的(+)-雪松醇合酶和(-)-罗汉柏烯合酶的序列来搜索来自金塔柏(p.orientalis'beverleyensis')和侧柏(p.orientalis)的潜在(+)-雪松醇合酶和(-)-罗汉柏烯合酶。这种方法为每种物种提供了两种新的推定的倍半萜合酶序列,包括来自金塔柏(p.orientalis'beverleyensis')的porb1和来自侧柏(p.orientalis)的por2-3-5。如实施例6所述评价了porb1和por2-3-5的酶活性。实施例6porb1和por2-3-5的功能表达和表征。首先使用smartertmracecdnaamplificationkit(clontech)将用rneasyplantminikit(qiagen,germany)提取的总rna逆转录成cdna,然后将该产物用作基因克隆的模板。通过使用正向引物(5'-tttaagtgcttctgcgatg-3'(seqidno:20))和反向引物(5'-acatctaggtttgtgcctt-3'(seqidno:21))从金塔柏(p.orientalis'beverleyensis')的cdna扩增出porb1。认为por2-3-5被错误地组装,因此使用基因特异性反向引物(5'-atcgccatctccagtgtg-3'(seqidno:22))连同由smartertmracecdnaamplificationkit(clontech)提供的universalprimeramix用于克隆por2-3-5的5'末端序列,从中设计出用于全长克隆的正向引物。然后使用正向引物(5'-ctttagtgcttctgtgatg-3'(seqidno:23))和反向引物(5'-catacaagtttgtgcctca-3’(seqidno:24))从侧柏(p.orientalis)的cdna扩增出por2-3-5。通过遵循大肠杆菌的遗传密码子频率来优化porb1和por2-3-5的序列并合成。将ndei的限制性位点加入到porb1和por2-3-5二者的5'末端,同时将kpni加入到3'末端。将porb1和por2-3-5亚克隆到pj401(dna2.0)质粒中或petduet-1(novagen)质粒中,用于随后在大肠杆菌中表达。将krx大肠杆菌细胞(promega)与质粒pacyc/scmva(含有编码异源甲羟戊酸途径的基因)、质粒pj401-porb1、petduet-porb1、pj401-por2-3-5和petduet-por2-3-5分别共转化。为了构建pacyc/scmva质粒,将8个生物合成基因分为2个合成操纵子,称为“上”和“下”甲羟戊酸(mva)途径。作为上mva途径,创建了由atob编码的来自大肠杆菌的乙酰乙酰辅酶a硫解酶、hmg-coa合酶和分别由erg13和erg19编码的来自酿酒酵母的hmg-coa还原酶的截短形式组成的合成操纵子。该操纵子将初级代谢物乙酰辅酶a转化为(r)-甲羟戊酸。作为“下”甲羟戊酸途径,创建了第二合成操纵子,其编码甲羟戊酸激酶(erg12,酿酒酵母)、磷酸甲羟戊酸激酶(erg8,酿酒酵母)、磷酸甲羟戊酸脱羧酶(mvd1,酿酒酵母)、异戊烯基二磷酸异构酶(idi,大肠杆菌)和法呢基焦磷酸(fpp)合酶(ispa,大肠杆菌)。最后,将来自酿酒酵母(erg20)的第二种fpp合成酶引入到上途径操纵子中,以改进类异戊二烯c5单元(ipp和dmapp)至法呢基焦磷酸(fpp)的转化。在噬菌体t7启动子(pacycduet-1,invitrogen)的控制下,将每个操纵子亚克隆到低拷贝表达质粒的多重克隆位点之一。在含有卡那霉素(终浓度50μg/ml)和氯霉素(终浓度34μg/ml)的lb-琼脂平板上选择所共转化的细胞。使用单个菌落接种补充有相同抗生素和葡萄糖(终浓度0.4%w/v)的5ml液体lb培养基,用500μl癸烷覆盖。培养物在37℃和200rpm摇动下温育过夜。第二天,用0.3ml的过夜培养物接种补充有相同抗生素和甘油(终浓度6%w/v)的2mltb培养基,并用200μl癸烷覆盖。在37℃下温育6小时并以200rpm振荡后,将培养物冷却至25℃保持1小时,并向每个管中加入iptg(终浓度0.1μm)和鼠李糖(终浓度0.02%w/v)。将培养物在25℃和180rpm摇动下温育另外48小时。然后用1体积的mtbe萃取培养物,加入50μl的2mg/ml的异长叶烯作为内标,然后通过gc/ms分析样品。gc/ms分析使用与实施例5中所述相同的系统。载气为氦气,恒定流量1.0ml/min。注射为不分流模式,注射器温度设定在250℃。烘箱温度程序升温从80℃以10℃/分钟升至220℃,然后以30℃/分钟升至280℃并保持1分钟。产品鉴定基于质谱和保留指数。gc/ms分析表明,porb1生产(+)-雪松醇作为主要产品,选择性为78%~88%(图9和图10),而por2-3-5生产(-)-罗汉柏烯作为主要产品,选择性为45%~55%(图11和图12)。实施例7雪松醇合酶的序列比较。使用clustalw程序对porb1和por2-3-5的氨基酸序列以及四种北美圆柏(j.virginiana)雪松醇合酶的氨基酸序列进行比对,并从比对中推导出序列同一性。合酶之间的序列同一性如下表所示。porb1por2-3-5jvcp1206-3jvcp1206-4jvcp1206-5jvcp1206-6查询序列porb1id74.1076.1676.3377.3676.16por2-3-574.27id76.5976.4279.1776.25-序列表–seqidno:1jvcp1206-4,氨基酸序列.msnlkgdhissvssipahafnewgdafvqsmempygepeyreraetlvkqvkillkemqtgdgdlierlemvdalqclgieryfqaeikealdyvyrswdgtvgiglgcnsatkhlnatalglrvlrlhrydvspdtlynfkdntgefvlcgenkvsndedtnkeekvmrsmlnllrlsslafpgeiimeeaqafstrylkelleisgdtfnrsfikeveyaltyewprtftrweawnfieicdldndrledkrilqlakldfnilqfqyklemknlsswwvesgisnlvatrarhieylfwavastdemefsssrialakttaiitvmddifddyatleylkcisdaisknwdvsiienipnnlktcfefisktvhqmaidatkyqgrdmmpfitkawadyieacfeearwkltgyfptydeymksaelcvgfgqiflssgllaspnlcdddiekiyldksrffklmrvcmrliddindfederlhgkiasaiacykgdhpncseseainqiitlnnkllreltreffksnmnflewqkicvnstrgvqffyifrdgftyshkeikqqifkilvdpikmseqidno:2jvcp1206-3,氨基酸序列.msnlkgdhissvssipahafnewgdafvqsmempygepeyreraetlvkqvkillkemqtgdgdlierlemvdalqclgieryfqaeikealdyvyrswdgtvgiglgcnsatkhlnatalglrvlrlhrydvspdtlhnfkdntgkfvltgenkdnndedtnkeekvmrsilnlfrlsslafpgeiimeeakafstrylkelleisrdtfnrsfikeveyaltyewprtftrweawnfieicdldndrledkrilqlakldfnilqfqyklemknlsswwvesgisnlvatrarhieylfwavastdemefsssrialakttaiitvmddifddyatleylkcisdaisknwdvsiienipnnlktcfefisktvhqmaidatkyqgrdmmpfitkawadyieacfeearwkltgyfptydeymksaelcvgfgqiflssgllaspnlcdddiekiyldksrffklmrvcmrliddindfederlhgkiasaiacykgdhpncseseainqivmlnnkllreltreflksnmnflewekicvnstrgvqfcyifgdgftyshkeikqqifkilvnpikvseqidno:3jvcp1206-6,氨基酸序列.msnlkgdhissvssipahafnewgdafvqsmempygepeyreraetlvkqvkillkemqtgdgdlierlemvdalqclgieryfqaeikealdyvyrswdgtvgiglgcnsatkhlnatalglrvlrlhrydvspdtlhnfkdntgkfvltgenkdnndedtnkeekvmrsilnlfrlsslafpgeiimeeakafstrylkelleisrdtfnrsfikeveyaltyewprtftrweawnfieicdldndrledkrilqlakldfnilqfqyklemknlsswwvesgisnlvatrarhieylfwavastdemefsssrialakttaiitvmddifddyatleylkcisdaisknwdvsiienipnnlktcfefisktvhqmaidatkyqgrdmmpfitkawadyieacfeearwkltgyfptydeymksaelcvgfgqiflssgllaspnlcdddiekiyldksrffklmrvcmrliddindfederlhgkiasaiacykgdhpncseseainqiitlnnkllreltreffksnmnflewqkicvnstrgvqffyifrdgftyshkeikqqifkilvdpikmseqidno:4jvcp1206-5,氨基酸序列.msnlkgdhissvssipahafnewgdafvqsmempygepeyreraetlvkqvkillkemqtgdgdlierlemvdalqclgieryfqaeikealdyvyrswdgtvgiglgcnsatkhlnatalglrvlrlhrydvspdtlhnfkdntgkfvltgenkdnndedtnkeekvmrsilnlfrlsslafpgeiimeeakafstrylkelleisrdtfnrsfikeveyaltyewprtftrwearnfieicdldndrlkdkrilelakldfnilqfqyqlemknlsrwwvesgisnlvatrersieylfwavtstdelefsssriahakcttiitimddifddyatleqlkcivdaisknwdvsiienipnnlktcfefvsktvhelaidateyqgrdmmpfitkawtdygeacfeqacwkvkgyfptyneyikcaelsvafgpillhtallaspdlcdddiekiyldksrffklmrvcmrliddindfederlhgkiasaiacykgdhpncseseainqiitlnnkllreltreffksnmnflewqkicvnstrgvqffyifrdgftyshkeikqqifkilvdpikmseqidno:5jvcp1206-4,野生型cdna序列.atgtcgaatttgaaaggagaccacatttcttctgtttcttccattccagcccatgcttttaatgagtggggcgatgcttttgttcaatctatggagatgccgtacggggaacctgaataccgtgaacgtgctgaaacacttgtgaaacaagtcaaaatcttgttaaaagaaatgcaaactggagatggtgatctaatcgagcggcttgagatggttgatgctttgcaatgccttggcattgagcgatattttcaggctgagattaaagaagctcttgattacgtttaccgctcttgggatggaactgtgggaataggattaggctgcaacagtgctacaaagcatttgaatgccacagctttgggactcagagtacttcgactccatcgttatgacgtctctccagacacgttgtacaatttcaaggacaatactggcgagttcgtcctctgtggagaaaataaagtgagtaacgatgaggatactaataaggaagagaaagtgatgagaagtatgctcaacctgttaagactatccagtttggcattccctggagaaatcattatggaagaggctcaagcatttagcactagatatcttaaagaattattagaaatttctggagatacatttaacaggagttttattaaagaggtggagtatgctcttacatatgaatggcctcgaacctttactagatgggaggcgtggaatttcatagagatctgtgatttagataatgacaggttggaagacaaaaggattttacagcttgcaaaattggattttaatatactacaatttcaatataagttggagatgaaaaatctgtcaagttggtgggttgaatctggcatctccaatctggttgcaacaagggcccgacatattgaatatcttttttgggcagttgcttctacagatgagatggagttttctagtagtagaatagctcttgcaaagaccaccgcaattattacagtaatggatgacatttttgatgactatgcaacacttgagtatctcaaatgtatttcagatgccatttctaaaaattgggatgtttctattatagaaaatattcccaacaacttgaagacatgttttgaatttatttctaaaacagttcatcaaatggcaatagatgctactaaatatcaaggacgtgacatgatgccttttattacaaaagcgtgggcagattatatagaagcctgctttgaggaggcacgctggaaactgacaggatattttccaacctacgatgagtacatgaaatctgctgaactatgtgttggatttggacagatatttttatctagtgggctactagcatctcctaatttatgtgatgatgatattgagaagatataccttgacaaatctagattctttaaactcatgcgagtgtgtatgcggttgattgatgatataaatgattttgaggatgagaggctccatggaaagattgcctcagctattgcttgttacaagggtgatcatccaaattgttcagaaagcgaggccatcaatcaaatcatcacgctcaataataaattattgagagaattgacaagagaattttttaaatcaaatatgaattttcttgaatggcaaaagatatgtgtcaatagtaccagaggagtacaatttttctatatatttagagatgggtttacatattctcacaaggagatcaagcagcagatatttaaaatccttgttgatccaataaaaatgtagseqidno:6jvcp1206-3,野生型cdna序列.atgtcgaatttgaaaggagaccacatttcttctgtttcttccattccagcccatgcttttaatgagtggggcgatgcttttgttcaatctatggagatgccgtacggggaacctgaataccgtgaacgtgctgaaacacttgtgaaacaagtcaaaatcttgttaaaagaaatgcaaactggagatggtgatctaatcgagcggcttgagatggttgatgctttgcaatgccttggcattgagcgatattttcaggctgagattaaagaagctcttgattacgtttaccgctcttgggatggaactgtgggaataggattaggctgcaacagtgctacaaagcatttgaatgccacagctttgggactcagagtacttcgactccatcgttatgacgtctctccagacacgttgcacaatttcaaggacaatactgggaagttcgtcctcactggagaaaataaagacaataacgatgaagatactaataaggaagagaaagtgatgagaagtattctcaacctgttcagactatccagtttggcattccctggagaaattattatggaagaggctaaagcatttagcactagatatcttaaagaattattagaaatttctagagatacatttaacaggagttttattaaagaggtggagtatgctcttacatatgaatggcctcgaacctttactagatgggaggcgtggaatttcatagagatctgtgatttagataatgacaggttggaagacaaaaggattttacagcttgcaaaattggattttaatatactacaatttcaatataagttggagatgaaaaatctgtcaagttggtgggttgaatctggcatctccaatctggttgcaacaagggcccgacatattgaatatcttttttgggcagttgcttctacagatgagatggagttttctagtagtagaatagctcttgcaaagaccaccgcaattattacagtaatggatgacatttttgatgactatgcaacacttgagtatctcaaatgtatttcagatgccatttctaaaaattgggatgtttctattatagaaaatattcccaacaacttgaagacatgttttgaatttatttctaaaacagttcatcaaatggcaatagatgctactaaatatcaaggacgtgacatgatgccttttattacaaaagcgtgggcagattatatagaagcctgctttgaggaggcacgctggaaactgacaggatattttccaacctacgatgagtacatgaaatctgctgaactatgtgttggatttggacagatatttttatctagtgggctactagcatctcctaatttatgtgatgatgatattgagaagatataccttgacaaatctagattctttaaactcatgcgagtgtgtatgcggttgattgatgatataaatgattttgaggatgagaggctccatggaaagattgcctcagctattgcttgttacaagggtgatcatccaaattgttcagaaagtgaggccatcaatcaaatcgtcatgctcaataataaattattgagagaattgacaagagaatttttaaaatcaaatatgaattttcttgaatgggaaaagatatgtgtcaatagtacaagaggggtacaattttgctatatatttggagatgggtttacatattctcacaaggagatcaagcaacagatatttaaaattcttgtcaatccaataaaagtgtagseqidno:7jvcp1206-6,野生型cdna序列.atgtcgaatttgaaaggagaccacatttcttctgtttcttccattccagcccatgcttttaatgagtggggcgatgcttttgttcaatctatggagatgccgtacggggaacctgaataccgtgaacgtgctgaaacacttgtgaaacaagtcaaaatcttgttaaaagaaatgcaaactggagatggtgatctaatcgagcggcttgagatggttgatgctttgcaatgccttggcattgagcgatattttcaggctgagattaaagaagctcttgattacgtttaccgctcttgggatggaactgtgggaataggattaggctgcaacagtgctacaaagcatttgaatgccacagctttgggactcagagtacttcgactccatcgttatgacgtctctccagacacgttgcacaatttcaaggacaatactgggaagttcgtcctcactggagaaaataaagacaataacgatgaagatactaataaggaagagaaagtgatgagaagtattctcaacctgttcagactatccagtttggcattccctggagaaattattatggaagaggctaaagcatttagcactagatatcttaaagaattattagaaatttctagagatacatttaacaggagttttattaaagaggtggagtatgctcttacatatgaatggcctcgaacctttactagatgggaggcgtggaatttcatagagatctgtgatttagataatgacaggttggaagacaaaaggattttacagcttgcaaaattggattttaatatactacaatttcaatataagttggagatgaaaaatctgtcaagttggtgggttgaatctggcatctccaatctggttgcaacaagggcccgacatattgaatatcttttttgggcagttgcttctacagatgagatggagttttctagtagtagaatagctcttgcaaagaccaccgcaattattacagtaatggatgacatttttgatgactatgcaacacttgagtatctcaaatgtatttcagatgccatttctaaaaattgggatgtttctattatagaaaatattcccaacaacttgaagacatgttttgaatttatttctaaaacagttcatcaaatggcaatagatgctactaaatatcaaggacgtgacatgatgccttttattacaaaagcgtgggcagattatatagaagcctgctttgaggaggcacgctggaaactgacaggatattttccaacctacgatgagtacatgaaatctgctgaactatgtgttggatttggacagatatttttatctagtgggctactagcatctcctaatttatgtgatgatgatattgagaagatataccttgacaaatctagattctttaaactcatgcgagtgtgtatgcggttgattgatgatataaatgattttgaggatgagaggctccatggaaagattgcctcagctattgcttgttacaagggtgatcatccaaattgttcagaaagcgaggccatcaatcaaatcatcacgctcaataataaattattgagagaattgacaagagaattttttaaatcaaatatgaattttcttgaatggcaaaagatatgtgtcaatagtaccagaggagtacaatttttctatatatttagagatgggtttacatattctcacaaggagatcaagcagcagatatttaaaatccttgttgatccaataaaaatgtagseqidno:8jvcp1206-5,野生型cdna序列.atgtcgaatttgaaaggagaccacatttcttctgtttcttccattccagcccatgcttttaatgagtggggcgatgcttttgttcaatctatggagatgccgtacggggaacctgaataccgtgaacgtgctgaaacacttgtgaaacaagtcaaaatcttgttaaaagaaatgcaaactggagatggtgatctaatcgagcggcttgagatggttgatgctttgcaatgccttggcattgagcgatattttcaggctgagattaaagaagctcttgattacgtttaccgctcttgggatggaactgtgggaataggattaggctgcaacagtgctacaaagcatttgaatgccacagctttgggactcagagtacttcgactccatcgttatgacgtctctccagacacgttgcacaatttcaaggacaatactgggaagttcgtcctcactggagaaaataaagacaataacgatgaagatactaataaggaagagaaagtgatgagaagtattctcaacctgttcagactatccagtttggcattccctggagaaattattatggaagaggctaaagcatttagcactagatatcttaaagaattattagaaatttctagagatacatttaacaggagttttattaaagaggtggagtatgctcttacatatgaatggcctcgaacctttactagatgggaggcccggaatttcatagaaatctgtgatttagataatgacaggttgaaagataaaaggattttagagcttgcaaaattggattttaatatactacaatttcaatatcagctggagatgaaaaatctctcaaggtggtgggttgaatctggcatctccaatctagttgcaacaagggagcgatctattgaatatcttttttgggcagttacttctacagatgagttggaattttctagtagtagaatagctcatgcaaagtgcaccacaataattacaataatggatgatatttttgatgactatgcaacacttgagcaactcaaatgtattgtagatgccatttcaaaaaattgggatgtttctattatagagaatatacccaataacttgaagacatgctttgaatttgtttctaaaacagttcatgaattggcaatagatgctactgaatatcaaggacgtgacatgatgccttttattacaaaagcgtggacagattatggagaagcttgctttgagcaggcatgctggaaagtgaaaggatattttccaacctacaatgagtacataaagtgtgctgaattaagtgttgcatttggaccgatattgttacatactgcactactagcatctcccgatttatgcgatgatgatattgagaagatataccttgacaaatctagattctttaaactcatgcgagtgtgtatgcggttgattgatgatataaatgattttgaggatgagaggctccatggaaagattgcctcagctattgcttgttacaagggtgatcatccaaattgttcagaaagcgaggccatcaatcaaatcatcacgctcaataataaattattgagagaattgacaagagaattttttaaatcaaatatgaattttcttgaatggcaaaagatatgtgtcaatagtaccagaggagtacaatttttctatatatttagagatgggtttacatattctcacaaggagatcaagcagcagatatttaaaatccttgttgatccaataaaaatgtagseqidno:9jvcp1206-4,密码子优化的cdna序列.atgagcaatttgaaaggcgatcacatcagcagcgtatctagcattccggcacatgcattcaatgaatggggcgacgcctttgttcagagcatggaaatgccgtacggtgagccggaatatcgcgagcgtgcggagactctggtcaaacaagtgaagattctgctgaaagagatgcaaaccggtgacggcgacttgattgaacgtctggagatggtggatgcgctgcaatgcctgggtattgagcgttatttccaagcggagattaaagaggcgctggattacgtgtaccgtagctgggacggcacggtgggcatcggtctgggttgcaactcggccaccaagcatctgaacgctaccgctctgggcctgcgtgttctgcgcctgcatcgttatgatgtgagccctgacaccttgtataactttaaggacaataccggcgaatttgtcctgtgtggtgagaacaaagttagcaatgatgaagatactaacaaagaagagaaggttatgcgcagcatgttgaatttgctgcgcctgagctctttggcttttccgggtgagatcatcatggaagaagcgcaggcgtttagcacccgttatctgaaagaactgctggagatctctggcgacacctttaatcgtagcttcatcaaagaggtcgagtacgcgctgacctatgaatggccacgtaccttcacccgctgggaagcatggaatttcattgaaatttgtgacctggacaacgaccgtctggaagataagcgtatcctgcagctggcgaagctggacttcaacatcctgcagtttcagtacaagctggagatgaagaatctgagcagctggtgggttgagagcggtatttccaacttggtcgcgacgcgtgcgcgccacatcgagtacttgttttgggcggtcgcgtctacggacgagatggagttttccagctcccgtatcgccctggcgaaaaccacggctattatcaccgttatggatgacattttcgatgattacgcgacgctggagtacctgaaatgtatttccgacgccattagcaagaattgggatgtcagcattattgaaaacatcccgaacaatctgaaaacgtgcttcgagttcattagcaaaacggtgcaccagatggccattgatgcgacgaagtatcagggccgtgacatgatgccgtttatcactaaggcctgggctgattacattgaagcctgtttcgaagaagcacgctggaagctgacgggttacttcccgacctatgatgagtacatgaaaagcgcggaactgtgcgtgggtttcggtcagatttttctgagctcgggcctgttggcaagcccgaatttgtgtgatgacgatattgagaagatttacctggataaaagccgtttcttcaagctgatgcgcgtttgcatgcgtctgatcgatgacatcaacgacttcgaggacgaacgtctgcacggtaagatcgcaagcgcaatcgcatgctataagggtgaccacccgaattgcagcgaaagcgaggcaattaaccaaatcatcaccttgaacaataaactgctgcgcgaactgacccgcgagtttttcaagagcaatatgaactttctggagtggcagaaaatctgtgtgaactccacccgtggtgtccaattcttctatatctttcgtgatggttttacctactctcacaaagagattaaacaacaaatcttcaaaattctggttgacccgatcaagatgtaaseqidno:10jvcp1206-3,密码子优化的cdna序列.atgagcaatttgaaaggcgatcacatcagcagcgtatctagcattccggcacatgcattcaatgagtggggtgatgcgttcgtccaaagcatggaaatgccgtatggtgagccggagtaccgtgaacgtgctgaaacgctggttaaacaagtgaagattctgctgaaagaaatgcagaccggcgatggtgacctgatcgaacgcctggagatggtggacgcactgcaatgtctgggtattgagcgttactttcaagccgagatcaaagaagcgctggactacgtgtaccgcagctgggatggcaccgtcggtattggtctgggttgcaatagcgcgaccaagcacctgaatgcaacggcgctgggtctgcgcgttctgcgcctgcaccgctatgatgttagcccggatactctgcataacttcaaggataacacgggtaagtttgtcctgacgggcgagaacaaagacaataacgacgaagatactaacaaagaagagaaggttatgcgttccattctgaatctgtttcgtttgagctccctggcatttccgggcgagatcattatggaagaggctaaagcgttctctactcgttacctgaaagaactgctggaaatcagccgcgacaccttcaatcgtagcttcatcaaagaggttgagtatgctttgacctacgagtggcctcgcacctttacgcgttgggaagcgtggaatttcatcgaaatttgcgacctggacaacgaccgtctggaagataagcgtatcttgcagctggcaaagctggacttcaatatcctgcaatttcagtacaaactggaaatgaagaatctgtccagctggtgggtcgagagcggtattagcaacctggtggcgacgcgtgcgcgtcatatcgaatacttgttctgggcggtcgccagcacggacgagatggagttcagcagctctcgtattgccctggcaaagaccaccgcaattatcaccgtgatggatgacattttcgatgactacgcgaccctggagtacctgaaatgtatttcggatgcgatcagcaagaactgggatgtttccattattgaaaacattccgaacaacctgaaaacctgttttgagtttatcagcaaaaccgttcaccagatggcgatcgatgctacgaaatatcagggtcgtgacatgatgccattcattacgaaggcgtgggccgactatattgaggcatgtttcgaagaagcgcgttggaagctgacgggctactttccgacctacgacgagtatatgaagagcgcggaattgtgcgttggttttggtcagatctttctgagctctggcctgttggcttccccgaatctgtgcgacgacgacattgagaaaatctatttggacaagtcccgcttcttcaagctgatgcgtgtttgtatgcgcttgatcgatgacattaacgatttcgaggatgagcgtctgcacggcaaaatcgccagcgccatcgcctgctataaaggcgaccatccgaattgtagcgagtctgaggcgatcaaccagatcgtgatgctgaataacaaattgctgcgcgaactgacccgcgagttcctgaagagcaatatgaactttctggagtgggagaagatttgcgtgaacagcacccgtggtgtgcaattctgctacatttttggcgatggttttacctatagccacaaagaaatcaaacaacagatctttaagattctggtcaatccgatcaaggtctaaseqidno:11jvcp1206-6,密码子优化的cdna序列.atgagcaatttgaaaggcgatcacatcagcagcgtatctagcattccggcacatgcattcaatgagtggggtgacgcgtttgtgcagagcatggaaatgccgtatggtgaaccggaatatcgtgagcgtgctgaaaccctggtgaagcaagtcaagattctgttgaaagaaatgcaaaccggcgacggtgatctgatcgagcgcctggagatggttgatgcgctgcagtgtctgggtattgagcgctattttcaagccgagatcaaagaagcgctggattacgtttatcgtagctgggatggcacggttggtattggcctgggctgcaatagcgcgaccaagcacctgaacgctaccgcgctgggtctgcgcgtgttgcgtttgcaccgctacgacgtttcgccggatactctgcataactttaaagataatacgggcaaattcgtcctgacgggtgagaacaaagataacaacgatgaggacacgaacaaagaagaaaaagtcatgcgctccatcctgaatctgtttcgtctgagcagcctggcttttcctggcgagatcattatggaagaagcgaaggcgtttagcacccgttacctgaaagaactgttggagatcagccgtgataccttcaaccgtagctttatcaaagaggtggagtacgcgctgacctacgagtggccgcgtacctttacccgttgggaagcctggaatttcattgagatctgcgacctggataacgaccgtctggaagataagcgtattctgcaattggcgaaactggacttcaatattctgcagttccagtacaagctggagatgaagaatctgtccagctggtgggttgagagcggtatcagcaacctggtcgcgacgcgtgcacgtcatatcgagtacctgttttgggcggtcgctagcacggacgaaatggagtttagctccagccgcattgcactggccaagaccactgcaatcattaccgtgatggatgatatctttgacgattacgcgaccttggagtatctgaaatgcatctctgacgcgatcagcaagaactgggacgttagcattattgaaaacattccgaataacttgaaaacgtgttttgagttcattagcaaaactgttcaccaaatggcaatcgacgccaccaaatatcagggccgtgacatgatgccgtttatcaccaaggcctgggcagactacatcgaggcatgctttgaagaagctcgctggaaactgacgggttatttcccgacctacgatgagtacatgaagtccgccgagctgtgcgtcggcttcggtcagattttcctgtcgagcggtctgctggcaagcccaaatctgtgtgacgacgacattgaaaagatttacttggacaagagccgctttttcaagctgatgcgtgtgtgtatgcgtctgattgatgacattaacgatttcgaggacgaacgcctgcacggtaagatcgcgtccgccattgcgtgctacaagggcgaccatccgaattgctctgaatctgaagcgattaaccaaatcatcaccctgaacaataaactgctgcgtgagttgacccgtgagttcttcaagtctaacatgaattttctggagtggcagaagatttgtgttaatagcacgcgcggtgtgcaattcttctatatcttccgcgatggtttcacgtatagccacaaagagatcaagcagcagattttcaaaatcctggtggacccgatcaaaatgtaaseqidno:12jvcp1206-5,密码子优化的cdna序列.atgagcaatttgaaaggcgatcacatcagcagcgtatctagcattccggcacatgcattcaacgagtggggcgacgctttcgtgcaatctatggagatgccgtatggtgagccggagtaccgtgagcgtgcggaaacgctggtgaaacaagttaagatcctgctgaaagagatgcagaccggtgatggcgatctgattgaacgtctggagatggtcgatgcgctgcaatgcctgggtatcgaacgttacttccaggcggagatcaaagaggccctggactatgtttaccgtagctgggatggcacggtcggtattggtctgggttgcaacagcgcgacgaaacacctgaacgcgacggctctgggtctgcgcgttctgcgcctgcaccgttacgatgtcagcccggacacgctgcataactttaaggacaatacgggcaaatttgtgctgactggtgaaaacaaagataacaacgacgaggataccaataaagaagaaaaggtcatgcgttccatcctgaatttgttccgcctgagcagcttggcctttccgggcgagatcattatggaagaagcgaaggcgtttagcacccgttatctgaaagaactgctggaaattagccgcgacacctttaaccgcagctttatcaaagaagtcgaatacgccctgacctacgagtggccgcgtacctttacccgttgggaagcgcgtaatttcattgaaatctgtgatttggataatgaccgtctgaaggataagcgtatcctggagctggcgaagctggactttaacattttgcagttccaatatcagttggagatgaaaaatctgagccgctggtgggtggagagcggtattagcaacttggttgccactcgtgagcgttccattgaatacctgttctgggcggtcacgtctaccgacgaactggagtttagctctagccgcatcgcgcacgcgaaatgcaccacgatcatcaccatcatggatgatatctttgacgattatgcaaccctggagcaactgaagtgtattgtggacgctatttcgaagaactgggacgtttccatcattgagaacattccgaataatctgaaaacctgtttcgagttcgtgagcaaaaccgttcacgagctggcaattgatgccaccgagtatcaaggtcgtgacatgatgccgttcatcaccaaggcctggaccgattatggtgaagcatgtttcgagcaggcttgctggaaggtgaagggttactttcctacctacaacgagtatatcaagtgcgcagaactgagcgtcgcctttggcccgattctgctgcatacggcgctgttggcgagcccagacctgtgcgacgatgacattgagaaaatctatttggacaagtcgcgcttctttaaactgatgcgcgtttgtatgcgcctgattgacgacattaatgacttcgaggatgagcgcttgcacggcaagattgcaagcgcgattgcatgctacaagggtgatcatccgaattgcagcgaatccgaggcaatcaaccagatcattactctgaacaataaactgctgcgtgaactgacgcgtgagttctttaagagcaatatgaattttctggaatggcagaagatttgtgttaactccacccgtggcgttcagttcttctacatcttccgtgacggtttcacctacagccacaaagaaatcaaacagcaaatcttcaaaatcctggtggacccgatcaagatgtaaseqidno:13porb1,氨基酸序列msnlmgdhisslssipsnafnqwddafiqsmetpygepeyreraetlakeikiflkdmqsgggdgdlierleivdalqclgidryfqaeikaaldyvyncwdesvgiglgsqsatkdlnatalalrvfrlnrydvsadtlkyfkdnngrfvlcgdnkdnndednskeekvmrsmlnllrlsslafpaeivmeeakafssrylkellgksgdtskksflkeveyaliyewprtfirwearnfieiyeldnerlkekrilelakldfnilqfhyklemknlsswwveseiskliatrersieyllwaissmdelehsssrialakitslitilddifddyatfeqlkcirdaifkgwdvsiienipnnwkrcvefvfktihqltidatdyqgrdmmpfvskawedyveacfeqarwklkgyfptyneyikiagkcvgfgpfslhsailaspnlcdddiqkiyldksrfyqlmrvamrliddihdfeeerlhgkmasaiscymadhpncsekeamnhiielnnevlkeltreflkpsmifhewekifvnstrgvqffyvhgdgftythkeikhqilkiivdpiki---------------------------------------------------seqidno:14por2-3-5,氨基酸序列mstlegdniysvsslpahafnewedasvqsmemsygepeyreraetlvkevkillkemhtgdgdlierlemvdalqclgiyryfqaeikqaldyvyscwdgnvgiglgsesptqhlnatalgirvlrlhrydvsadtlknfkdkngqfvlcggnndnndeeekvmrsmlnlfrlssvaipgemvleeakafssrylkellensgdtvkrsfikeveyaltyewpitfdrwealnfieiydlnnerlmdkrilelaklnfnilqfqyklemknlsswwaksgiskllavrersieylfwaitsveelelsssrialvkcttvitivddifddyatfeqlqcitdaiskdwdvsllenipsnlktslefvsktihelamdatkyqgrdmmpfvtkawldytnacfeqarwkvtgyfpsyneyikaaelsvafgpillhtalaaspilcdediekiyldksrfyhimrvsmrltddihdfederlhgkmasaiscykgdhpncseeeainnivtlnnellkemireffkpnshylewekicvnstrgigffyifgdgftyshkeikeqifkiivnpikvseqidno:15porb1,编码dna序列(野生型)atgtctaatttgatgggagatcacatttcttctctttcttccattccatccaatgctttcaatcagtgggacgatgcgtttattcaatctatggagacgccatacggggaacctgaataccgtgaacgtgctgaaacacttgctaaggaaataaaaatctttttaaaagacatgcaatctggaggtggagatggcgatctaatcgagcggcttgagattgttgacgccttgcaatgcctcggaatagatcgttattttcaggctgaaataaaagcggctcttgattacgtttataactgttgggatgaaagtgtggggataggattagggagccaaagtgctacaaaggatttgaatgctacagctttagcacttcgagtgtttcgacttaatcgttatgatgtgtctgcagacacgttgaagtatttcaaggataataatgggcggttcgtactctgtggagacaataaagacaacaacgacgaggataatagcaaagaagaaaaagtgatgagaagtatgctcaacctgttaagactttccagtttggcatttcctgcagaaatcgttatggaagaggctaaagcattcagttctagatatcttaaagaactattaggaaaatctggagatacatctaagaaaagttttcttaaagaggtggagtatgcccttatatatgaatggcctcgaacatttattagatgggaggcacgaaatttcatagaaatctatgaactagataatgagaggttaaaagagaaaaggattttagaacttgcgaaattggattttaacatactacaatttcactacaagctagagatgaaaaatctctcaagttggtgggttgaatctgaaatctccaagctaattgcaacaagagaacgatccattgaatatcttttgtgggcaattagttctatggatgaattggagcattctagtagtagaatagctcttgcaaaaatcacatcacttatcacaatattggatgatatttttgatgactatgcaacatttgagcaactcaaatgcattagggatgccatttttaaaggttgggatgtttctatcatagaaaacattcccaacaactggaaaagatgcgtggaatttgtttttaaaacaattcatcaattgacaatagatgctactgattatcaagggcgtgacatgatgccttttgtttcaaaagcgtgggaagattatgtggaagcctgctttgagcaggcacgatggaaattgaaaggatattttccaacctacaatgagtacataaagatagctggaaaatgtgtagggtttggacccttttctttacattctgccatactagcatctccaaatttatgtgatgatgatattcagaagatataccttgataaatctagattttatcaactcatgcgagtggctatgaggttaattgatgatatacacgactttgaggaagagagactccatggaaagatggcctcagctatttcttgttatatggctgatcatccaaattgttcagagaaagaggcaatgaatcatatcatcgaactaaataatgaagtattgaaggaattgacaagagaatttttaaaaccaagtatgatatttcatgagtgggagaagatatttgtcaattctactcgaggagtacaatttttctatgtacatggtgatggatttacatatacgcataaggagatcaagcatcagatactaaaaattattgtcgatccaataaaaatctag---------------------------------------------------seqidno:16por2-3-5,编码dna序列(野生型)atgtcgactttggaaggagacaacatttattctgtttcttccttaccagcccatgcttttaatgagtgggaagatgcttctgttcaatctatggagatgtcatacggggaacctgaataccgtgaacgtgctgaaacacttgtgaaagaagtaaaaatcttgttgaaagaaatgcacactggagatggcgatctaatcgagcggcttgagatggttgatgcattgcaatgccttggaatttatcgatactttcaggctgagattaaacaagctcttgattacgtttacagctgctgggatggaaatgtggggataggattaggctccgagagtcctacacagcatttgaatgccacagctttgggaatcagagtactgcgactccatcgttatgatgtgtctgcagacacgttgaagaatttcaaggacaaaaatgggcagttcgtactctgtggaggaaataatgacaataacgatgaggaagagaaagtgatgagaagtatgctcaacctgttcagactttccagtgtggcaattcctggagaaatggttctggaagaggctaaagcatttagcagtagatatcttaaagaattattagaaaattctggagatacagttaagagaagttttattaaagaggtggagtatgctcttacctatgaatggcctataacttttgatagatgggaggcactgaatttcatagaaatctatgatttaaataatgagaggttgatggacaaaaggatattagagcttgcaaaattgaattttaatatactacaatttcaatacaagttggagatgaaaaatctctcaagttggtgggctaaatctggcatctcgaaactacttgcagtaagggagcgatccattgaatatcttttttgggcaattacttctgtagaagaattggagctttctagtagtagaatagctcttgtaaagtgcacaacagttattacaatagtggatgatatttttgatgactatgcaacatttgagcaactccaatgtattacagatgctatctctaaagattgggatgtttctcttttagaaaacattcccagcaacttgaagacaagcttggaatttgtttcaaaaacaattcatgagttggcaatggatgctactaaatatcaagggcgtgacatgatgccttttgttacaaaagcgtggttagattacacgaacgcctgctttgagcaagcacgatggaaagtgactggttattttccaagctacaatgagtacataaaggctgctgaattaagtgtagcatttggaccgatattgttacatactgccctagcagcatctcctattttatgcgatgaagatattgagaagatataccttgataaatctagattctatcatatcatgcgagtgtctatgcggttgactgatgatatacatgattttgaggatgagaggctgcatggaaagatggcttcagctatttcttgttataagggtgatcatccaaattgttcagaagaagaggcaataaataatattgtcaccctcaataatgaattattgaaggaaatgataagggaattttttaaaccaaatagtcattatcttgaatgggaaaagatatgtgtcaatagtactagaggaataggatttttctatatatttggagatgggtttacatattctcacaaggaaatcaaggagcagatatttaaaattattgttaatccaataaaagtgtag---------------------------------------------------seqidno:17porb1,编码dna序列(通过大肠杆菌的genscript遗传密码子频率优化)atgtccaacctgatgggcgatcatattagctctctgagttccatcccgtccaacgcttttaatcagtgggatgacgcgttcattcaatcaatggaaaccccgtatggtgaaccggaataccgtgaacgcgctgaaacgctggcgaaagaaatcaaaatcttcctgaaagatatgcagtctggcggtggcgacggcgatctgattgaacgtctggaaatcgtggacgccctgcagtgcctgggtattgatcgctattttcaagcagaaatcaaagcggccctggactatgtttacaactgttgggatgaatcggtcggtattggcctgggttcccaatcagccaccaaagatctgaacgcaacggctctggcgctgcgtgtgtttcgcctgaatcgttatgacgtttctgcggataccctgaaatacttcaaagataacaacggccgtttcgttctgtgcggtgacaacaaagataacaacgacgaagataactctaaagaagaaaaagtcatgcgtagtatgctgaatctgctgcgcctgtcatcgctggctttcccggcggaaattgtcatggaagaagccaaagcatttagctctcgctatctgaaagaactgctgggcaaaagcggtgataccagcaaaaaatcttttctgaaagaagtggaatacgccctgatttacgaatggccgcgcacgttcatccgttgggaagcacgcaacttcatcgaaatctacgaactggacaacgaacgtctgaaagaaaaacgcattctggaactggcgaaactggattttaacatcctgcagttccattacaaactggaaatgaaaaacctgagttcctggtgggtggaatctgaaattagtaaactgatcgctacccgtgaacgctccattgaatatctgctgtgggcgatctcatcgatggatgaactggaacacagctctagtcgtattgctctggcgaaaatcacctcactgattacgatcctggatgacatttttgatgactacgctaccttcgaacagctgaaatgcattcgtgacgcgatcttcaaaggctgggatgttagtattatcgaaaacatcccgaacaattggaaacgctgtgtggaatttgttttcaaaacgattcatcagctgaccatcgacgctacggattatcaaggtcgtgacatgatgccgtttgtcagcaaagcatgggaagattatgtggaagcctgtttcgaacaggcacgctggaaactgaaaggctactttccgacctataacgaatacattaaaatcgccggtaaatgcgttggctttggtccgttctccctgcactcagccattctggcatctccgaatctgtgtgatgacgatatccagaaaatctacctggataaaagtcgtttctaccaactgatgcgtgtcgcgatgcgcctgattgacgatatccatgattttgaagaagaacgcctgcacggcaaaatggcctcggcaattagctgctatatggccgatcatccgaactgtagcgaaaaagaagcaatgaatcacattatcgaactgaacaatgaagtgctgaaagaactgacccgtgaatttctgaaaccgtcgatgatcttccatgaatgggaaaaaatcttcgttaacagcacgcgcggtgtccagtttttctatgtgcacggcgacggtttcacctacacgcataaagaaatcaaacaccaaatcctgaaaattatcgttgatccgattaaaatctaa---------------------------------------------------seqidno:18porb1,编码dna序列(通过大肠杆菌的dna2.0遗传密码子频率优化)atgtctaatttgatgggtgatcacatttcgagcctgagcagcattccgagcaacgcattcaatcagtgggatgacgcattcatccagtcgatggaaaccccgtatggtgagccggagtaccgtgagcgtgcggaaaccctggcaaaagaaatcaagatttttctgaaagacatgcagagcggcggcggcgatggcgatctgatcgagcgtttggaaatcgtggatgcgctgcaatgcctgggtatcgaccgttacttccaagccgagatcaaagctgccctggactacgtttataattgttgggacgagtctgttggcattggtctgggtagccagagcgccactaaagatctgaacgcaacggcgctggcgctccgtgttttccgcttgaaccgttacgacgtcagcgcggacaccttaaagtatttcaaagataacaacggtcgttttgtgctgtgtggcgataataaagacaacaatgacgaagataacagcaaagaagaaaaagtcatgcgcagcatgctgaatttgctgcgtctgagcagcctggcgtttcctgctgagattgtcatggaagaagcaaaggcctttagctctcgttatctgaaagaactgctgggtaagagcggcgataccagcaaaaagtcgtttttgaaagaagtggagtacgcactgatttatgagtggccgcgtaccttcatccgctgggaggcacgcaactttatcgagatctacgaactggacaacgaacgcctgaaagaaaagcgtatcttggaactggcgaaactggacttcaacattctgcagttccactataaactggagatgaagaatttgtcctcctggtgggtggagtccgagatcagcaagctgattgcgacgcgtgagcgtagcattgagtatctgctgtgggctattagcagcatggacgaactggagcactccagcagccgtatcgccctggcgaagattacctctctgattaccattctggatgatatttttgacgactacgcgacctttgagcaactgaagtgcatccgcgacgccatcttcaagggctgggatgttagcatcattgagaacatcccgaacaattggaaacgttgtgttgaatttgtctttaagacgattcatcaactgaccatcgacgctacggactaccagggtcgcgacatgatgccgttcgtgagcaaagcgtgggaagattatgttgaggcgtgcttcgagcaagcgcgttggaagctgaagggttactttccgacgtacaacgaatacatcaagatcgcgggtaaatgcgtcggtttcggtccattctcccttcatagcgcgattttggcgagcccgaacctgtgcgatgacgacatccaaaagatctatctggataagagccgtttttatcaattgatgcgcgtcgcgatgcgtctgattgacgacattcacgactttgaagaggaacgcctgcacggtaaaatggcctccgcgatcagctgctacatggcagatcacccgaactgttcagagaaagaggcaatgaaccacattattgagttgaataatgaagtcctgaaagaactgacccgtgagttcctgaaaccgagcatgatcttccatgagtgggaaaagatctttgtgaatagcacgcgcggtgtgcaattcttttacgttcacggcgatggcttcacctacacgcataaagaaatcaagcatcagattctgaagattatcgtggacccgattaagatttaa---------------------------------------------------seqidno:19por2-3-5,编码dna序列(通过大肠杆菌的genscript遗传密码子频率优化)atgagcaccctggaaggcgacaacatctacagcgtgagcagcctgccggcgcacgcgttcaacgagtgggaagatgcgagcgttcagagcatggagatgagctacggtgaaccggaatatcgtgagcgtgcggaaaccctggtgaaggaagttaaaatcctgctgaaggagatgcacaccggtgacggcgatctgattgagcgtctggaaatggtggacgcgctgcaatgcctgggcatctaccgttattttcaggcggaaattaaacaagcgctggactacgtgtatagctgctgggatggcaacgttggtatcggtctgggtagcgagagcccgacccagcacctgaacgcgaccgcgctgggtattcgtgtgctgcgtctgcaccgttacgacgttagcgcggataccctgaagaacttcaaggataaaaacggtcaatttgtgctgtgcggtggcaacaacgacaacaacgatgaggaagagaaagttatgcgtagcatgctgaacctgttccgtctgagcagcgtggcgatcccgggtgaaatggttctggaagaggcgaaggcgtttagcagccgttatctgaaagagctgctggaaaacagcggtgacaccgtgaagcgtagcttcatcaaagaggttgaatacgcgctgacctatgagtggccgattaccttcgatcgttgggaagcgctgaactttatcgagatttacgacctgaacaacgaacgtctgatggataagcgtatcctggagctggcgaaactgaacttcaacattctgcagtttcaatataagctggaaatgaaaaacctgagctcctggtgggcgaagagcggcatcagcaaactgctggcggttcgtgagcgtagcatcgaatacctgttttgggcgattaccagcgtggaagagctggagctgagcagcagccgtatcgcgctggttaagtgcaccaccgtgatcaccattgttgacgatattttcgacgattatgcgacctttgaacagctgcaatgcatcaccgacgcgattagcaaagactgggatgtgagcctgctggagaacatcccgagcaacctgaagaccagcctggaattcgttagcaaaaccattcacgagctggcgatggacgcgaccaagtaccagggtcgtgatatgatgccgtttgtgaccaaagcgtggctggattacaccaacgcgtgcttcgagcaagcgcgttggaaggtgaccggctattttccgagctacaacgaatatatcaaagcggcggagctgagcgttgcgttcggtccgatcctgctgcacaccgcgctggcggcgagcccgattctgtgcgacgaggatatcgaaaagatttacctggacaaaagccgtttctatcacatcatgcgtgttagcatgcgtctgaccgacgatattcacgactttgaggatgaacgtctgcacggcaagatggcgagcgcgattagctgctacaaaggtgatcacccgaactgcagcgaagaggaagcgatcaacaacattgtgaccctgaacaacgagctgctgaaggaaatgatccgtgagttctttaaaccgaacagccactatctggagtgggaaaagatttgcgttaacagcacccgtggcatcggtttcttttacattttcggcgacggttttacctatagccacaaggagatcaaagaacagattttcaagatcattgtgaacccgatcaaagtttaaseqidno:20正向引物tttaagtgcttctgcgatgseqidno:21反向引物acatctaggtttgtgccttseqidno:22基因特异性反向引物atcgccatctccagtgtgseqidno:23正向引物ctttagtgcttctgtgatgseqidno:24反向引物catacaagtttgtgcctca当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1