针对PARP抑制剂的应答的预测和靶向C‑MET和PARP1的组合疗法的制作方法
背景技术:
本申请要求2014年9月29日提交的美国临时申请号62/057,037的优先权,其全部内容以引用的方式并入本文中。
本发明大体上涉及医药和肿瘤学领域。更具体地说,本发明涉及用于鉴别和治疗对parp抑制剂有抗性的癌症的方法。
背景技术
癌细胞中高含量的活性氧物质(ros)可引起氧化性dna破坏,导致基因组不稳定性和肿瘤发生(irani等人,1997;trachootham等人,2009;radisky等人,2005;lindahl,1993)。ros诱导的dna损伤(例如单链断裂)将parp1募集到病变部位以通过parp1上的聚adp-核糖基化(par基化)和其靶蛋白精心安排dna修复过程(luo和kraus,2012;gibson和kraus,2012)。已经在临床试验中广泛评估了parp抑制剂,因为在brca突变型癌细胞中发现了parp抑制作用的合成致死性,所述brca突变型癌细胞的双链dna破坏的修复机制存在缺陷(farmer等人,2005;bryant等人,2005)。
此外,还在三阴性乳腺癌(tnbc)的临床试验中研究了parp抑制剂,因为三阴性乳腺癌据报道具有brca特性(hampson等人,2010;turashvili等人,2011),例如brca突变、brca1启动子的甲基化和brca路径的失调。tnbc是起初对化学疗法作出应答的乳腺癌的一个侵袭性亚型,但是大多数tnbc最终产生抗性。此外,不同于其它乳腺癌亚型(例如对于雌激素受体(er)和/或her2呈阳性的乳腺癌亚型,针对其可获得特异性抑制剂),没有获批准的用于治疗tnbc的靶向疗法。超过100个测试parp抑制剂的临床试验在进行中,并且美国食品和药物管理局最近批准了parp抑制剂奥拉帕尼(olaparib)(lynparzatm,astrazeneca)用于治疗brca突变卵巢癌患者,并且关于这种靶向不同癌症类型的药物有许多临床试验正在进行中。虽然在携带brca突变的tnbc癌症患者中报道了振奋人心的结果(tutt等人,2010),但是这些结果在另一个试验中并没有观察到(gelmon等人,2011)。这些临床观察提出了一个重要问题,就是如何增加tnbc或其它癌症类型中的应答率。因此,需要用于增加对parp抑制剂的应答率和鉴别将对parp抑制剂作出应答的患者群组的方法以便在治疗期间将患者分层。
技术实现要素:
在这里显示受体酪氨酸激酶c-met与为dna修复所必需的parp1在tyr907结合并使parp1磷酸化,因此c-met使癌细胞对parp抑制产生抗性。组合c-met和parp1抑制剂在体外和乳腺癌异种移植肿瘤模型中产生了协同效应。parp1ptyr907具有作为生物标志物以预测抗性和使患者分层以最大化parp抑制剂的治疗益处的潜力。此外,确定了用于通过抑制c-met介导的parp1在y907的磷酸化来增加tnbc中的parp抑制剂应答率的潜在治疗策略,从而为用于具有高c-met表达且不对parp抑制作出应答的患者的组合疗法提供理论基础。
在第一实施方案中,本发明提供一种预测患者的癌症对parp1抑制剂的抗性的方法,其包括分析癌症样品以确定所述样品中parp1tyr907的磷酸化状态。在一些方面中,如果parp1tyr907被磷酸化,那么预测所述癌症对parp1抑制剂有抗性。
在各个方面中,分析可以包括测量parp1tyr907的磷酸化水平。在特定方面中,分析可以包括将所述样品与特异性结合磷酸化parp1tyr907的抗体接触。在一些方面中,分析可以包括进行蛋白质印迹法、elisa、免疫沉淀、放射免疫分析或免疫组织化学分析。
在一些方面中,所述方法可以进一步包括如果parp1tyr907被磷酸化,那么将所述患者鉴别为患有对parp1抑制剂有抗性的癌症。在特定方面中,鉴别可以包括报告所述患者是否患有对parp1抑制剂有抗性的癌症。在特定方面中,报告可以包括准备书面报告或口头报告。在特定方面中,所述方法可以进一步包括向所述患者、医生、医院或保险商报告。
在特定方面中,将所述患者鉴别为患有对parp1抑制剂疗法有抗性的癌症可以进一步包括将所述患有癌症的患者鉴别为parp1抑制剂与met抑制剂的组合治疗的候选对象。在一些方面中,所述parp1抑制剂是奥拉帕尼、abt-888(维利帕尼(veliparib))、bsi-201(依尼帕尼(iniparib))、bmn673、瑞卡帕布(rucaparib)(ag-014699、pf-01367338)、ag14361、ino-1001、a-966492、pj34或mk-4827。在一些方面中,所述met抑制剂可以是incb28060、arq197(替凡替尼(tivantinib))、amg458、gsk1363089(xl880或福来替尼(foretinib))、e7050(戈瓦替尼(golvatinib))、mk-2461、bms-777607、jnj-38877605、xl184(卡博替尼(cabozantinib))、amg337、arq197、mgcd265、pf04217903或pf02341066(克唑替尼(crizotinib))。
在一个实施方案中,提供一种治疗癌症患者的方法,其包括用治疗有效量的parp1抑制剂与met抑制剂的组合治疗被确定患有表达tyr907磷酸化parp1的癌症的患者。在各个方面中,所述癌症可以是乳腺癌、肾癌、肺癌或卵巢癌。在特定方面中,乳腺癌可以是三阴性乳腺癌。
在一些方面中,所述parp1抑制剂是奥拉帕尼、abt-888(维利帕尼)、bsi-201(依尼帕尼)、bmn673、瑞卡帕布(ag-014699、pf-01367338)、ag14361、ino-1001、a-966492、pj34或mk-4827。在一些方面中,所述met抑制剂可以是incb28060、arq197(替凡替尼)、amg458、gsk1363089(xl880或福来替尼)、e7050(戈瓦替尼)、mk-2461、bms-777607、jnj-38877605、xl184(卡博替尼)、amg337、arq197、mgcd265、pf04217903或pf02341066(克唑替尼)。
在一些方面中,所述parp1抑制剂可以与所述met抑制剂基本上同时给予。在特定方面中,所述患者可以先前已经历至少一轮抗癌疗法。在一个方面中,所述患者可以是人。
在各个方面中,所述方法可以进一步包括施用第二抗癌疗法,例如外科疗法、化学疗法、放射疗法、冷冻疗法、激素疗法、毒素疗法、免疫疗法或细胞因子疗法。
在一个实施方案中,提供一种用于选择用于癌症患者的药物疗法的方法,其包括(a)分析来自所述患者的癌症样品以确定所述样品中parp1tyr907的磷酸化状态;和(b)如果确定parp1tyr907被磷酸化,那么选择parp1抑制剂与met抑制剂的组合。
在一个实施方案中,提供一种使癌症对基于parp1抑制剂的抗癌疗法敏感的方法,其包括给予患有所述癌症的患者有效量的met抑制剂。在一些方面中,所述方法可以进一步包括给予受试对象基于parp1抑制剂的抗癌疗法。在一些方面中,所述基于parp1抑制剂的抗癌疗法可以与所述met抑制剂基本上同时给予。在一些方面中,所述parp1抑制剂是奥拉帕尼、abt-888(维利帕尼)、bsi-201(依尼帕尼)、bmn673、瑞卡帕布(ag-014699、pf-01367338)、ag14361、ino-1001、a-966492、pj34或mk-4827。在一些方面中,所述met抑制剂可以是incb28060、arq197(替凡替尼)、amg458、gsk1363089(xl880或福来替尼)、e7050(戈瓦替尼)、mk-2461、bms-777607、jnj-38877605、xl184(卡博替尼)、amg337、arq197、mgcd265、pf04217903或pf02341066(克唑替尼)。
在一个实施方案中,提供一种包含parp1抑制剂和met抑制剂的组合物,其用于治疗患者的癌症。在一些方面中,所述parp1抑制剂是奥拉帕尼、abt-888(维利帕尼)、bsi-201(依尼帕尼)、bmn673、瑞卡帕布(ag-014699、pf-01367338)、ag14361、ino-1001、a-966492、pj34或mk-4827。在一些方面中,所述met抑制剂可以是incb28060、arq197(替凡替尼)、amg458、gsk1363089(xl880或福来替尼)、e7050(戈瓦替尼)、mk-2461、bms-777607、jnj-38877605、xl184(卡博替尼)、amg337、arq197、mgcd265、pf04217903或pf02341066(克唑替尼)。
在一个实施方案中,提供一种包含特异性地结合tyr907磷酸化parp1蛋白质的抗体的组合物。在特定方面中,抗体特异性地结合对应于序列admvsksanpychtsqgd(seqidno:1)的parp1多肽。
在一个实施方案中,提供一种包含多肽的组合物,所述多肽包含与免疫原融合或缀合的parp1蛋白质的至少8个连续氨基酸,其中parp1蛋白质的至少8个连续氨基酸包括tyr907且其中tyr907被磷酸化。在一些方面中,所述多肽可以包含序列admvsksanychtsqgd(seqidno:1)。在各个方面中,所述免疫原可以是钥孔血蓝蛋白(klh)、牛血清白蛋白(bsa)、蓝色载体蛋白、智利鲍鱼血蓝蛋白(concholepasconcholepashemocyanin)(cch)或卵清蛋白(ova)。
如本文中所使用,关于指定组分的“基本上不含”在本文中用于意味指定组分中没有组分被有目的地配制成组合物和/或仅作为污染物存在或以痕量存在。由组合物的任何无意污染产生的指定组分的总量因此远远低于0.05%,优选低于0.01%。最优选的是其中用标准分析方法无法检测到指定组分的量的组合物。
如本说明书中所使用,“一”可以意味着一或多个。如本文的权利要求书中所使用,当结合词语“包含”使用时,词语“一”可以意味着一个或多于一个。
权利要求书中所用的术语“或”,除非明确指出只指代替代物或替代物是互相排斥的,否则用于意指“和/或”,虽然本公开支持只指代替代物和指代“和/或”的定义。如本文中所使用,“另一个”可以意味着至少第二或更多。
贯穿本申请,术语“约”用于指示一个值包括装置的内在误差变化、用于确定值的方法、或存在于研究受试对象中的变化。
本发明的其它目标、特征和优点将根据以下详细说明变得显而易知。然而,应了解虽然指示了本发明的优选实施方案,但是详细说明和具体实施例仅以说明的方式被提供,因为根据这个详细说明在本发明的精神和范围内的各种变化和修改将对于所属领域技术人员变得显而易知。
附图说明
以下图式形成本说明书的部分并且被包括在内以进一步展示本发明的特定方面。本发明可以参考这些图中的一或多个以及本文呈现的具体实施方案的详细说明被更好地理解。
图1a-k.ros诱导c-met和parp1的结合。(图1a)用8-ohdg抗体对乳腺癌患者的组织微阵列染色。展示代表性图像。通过皮尔逊卡方检验(p=0.001)进行相关分析。(图1b)用8-ohdg抗体染色的乳腺癌细胞的定量。通过axiovision软件测量荧光强度。(图1c)乳腺癌细胞用8-ohdg抗体染色并通过荧光显微法筛选。通过axiovision软件测量荧光强度。所示的定量是平均值±sem。*p<0.05,t检验。(图1d)乳腺癌细胞用于分离总dna并接受elisa分析以测量8-ohdg含量。(图1e)乳腺癌细胞用10μm的dcf-da孵育30分钟。通过axiovision软件测量荧光强度。显示了定量。(图1f)乳腺癌细胞用10μm的dcf-da孵育30分钟。通过荧光显微法筛选图像并通过axiovision软件测量荧光强度。所示的定量是平均值±sem。*p<0.05,t检验。(图1g)用抗par、parp1和微管蛋白抗体对乳腺癌裂解产物进行蛋白质印迹法。蛋白质印迹法是典型的三重复实验。定量展示在右侧。(图1h)用抗par抗体对乳腺癌细胞系进行蛋白质印迹法。所示的定量数据是平均值±sem。蛋白质印迹法是典型的三重复实验(n=3)。*p<0.05,t检验。(图1i)mda-mb-231细胞用20μm亚砷酸钠(as)处理18小时。内源parp1和c-met的结合通过ip和蛋白质印迹法检测。(图1j)进行duolink分析以检测parp1和c-met的共定位。柱形图,10μm。展示代表性图像。误差棒代表标准差。*p<0.05,t检验。(图1k)mda-mb-231细胞用h2o2或h2o2加克唑替尼预处理进行处理。用指定抗体对细胞溶质部分和核部分进行免疫沉淀和蛋白质印迹分析。l:长期暴露;s:短期暴露。误差棒代表标准差。*p<0.05,t检验。
图2a-r.c-met调节针对parp抑制剂的抗性。(图2a)mda-mb-231细胞中通过两种不同的shrna(shmet-a和shmet-b)的c-met敲低的蛋白质印迹分析。(图2b-c)c-met敲低细胞用parp抑制剂(abt-888和ag014699)处理并接受细胞活力分析。数据被显示为三重复实验(n=3)的平均值±sem。*p<0.05,t检验。(图2d)mda-mb-231细胞用abt-888和克唑替尼或福来替尼处理并经细胞活力分析。(图2e)mda-mb-231细胞用ag014699和克唑替尼或福来替尼处理并经克隆形成细胞存活分析。显示了定量。(图2f)c-met敲低细胞用ag014699处理并经软琼脂集落形成分析以确定锚定非依赖性细胞生长(anchorage-independentcellgrowth)。显示了定量。(图2g)mda-mb-231细胞中3’utr(shmet-c)处的c-met敲低以及野生型(wt)和激酶死亡(kd)c-met的再表达的蛋白质印迹分析。(图2h)具有3’utrc-met敲低和野生型或激酶死亡(kd)c-met的再表达的mda-mb-231细胞用ag014699处理并经克隆形成细胞存活分析。显示了定量。(图2i)mcf-7细胞(mcf-7/c-met)中的c-met表达的蛋白质印迹分析。(图2j-k)mcf-7/c-met细胞用ag014699处理并经克隆形成细胞存活分析和软琼脂分析。(图2l)在brac1突变型tnbc细胞(mda-mb-436和hcc1937)中parp抑制剂的ic50。由蛋白质印迹分析测量的c-met表达以图显示。(图2m)在hcc1937细胞中通过pgipzshrna的c-met敲低的蛋白质印迹分析。(图2n)具有c-met敲低细胞的hcc1937用parp抑制剂(ag014699和abt-888)处理并经细胞活力分析。(图2o)在mda-mb-436细胞中的野生型和激酶死亡(kd)c-met表达的蛋白质印迹分析。(图2p)过表达c-met的mda-mb-436细胞用parp抑制剂(ag014699和abt-888)处理并经细胞活力分析。(图2q-r)brca1和brca2敲低的mda-mb-231和mda-mb-157细胞用ag014699处理并接受细胞活力分析。误差棒代表标准差。cri,克唑替尼;ft,福来替尼。*p<0.05,**p>0.05,ranova。
图3a-i.c-met通过在y907磷酸化parp1介导parp1功能。(图3a)mda-mb-231细胞用abt-888、福来替尼或其组合处理。γh2ax通过免疫荧光共聚焦显微法检测。显示了定量。(图3b)c-met或对照敲低mda-mb-231细胞以及再表达野生型(wt)或激酶死亡(kd)突变体的c-met敲低细胞用指定药物处理18小时。γh2ax通过免疫荧光共聚焦显微法检测。显示了定量。(图3c)hek293t细胞用v5-parp1和flag-c-met表达质粒转染,并且细胞用10μmh2o2处理15分钟。parp1用v5抗体免疫沉淀,随后用4g10(抗磷-酪氨酸抗体)进行蛋白质印迹法。(图3d)具有由靶向3’utr的shrna敲低的内源parp1的mda-mb-231细胞和再表达arp1野生型或y907突变体的敲低细胞的蛋白质印迹分析。(图3e)表达parp1野生型或y907突变体的稳定细胞用h2o2处理并用甲酰胺基嘧啶dna糖基化酶(fpg)孵育,随后进行彗星分析以评估受损dna。尾部的dna指示受损dna。显示了定量。(图3f)表达parp1野生型或y907突变体的稳定细胞用h2o2(20μm)处理30分钟。通过par抗体检测聚-adp核糖基化。(图3g)mda-mb-231细胞用h2o2或h2o2加克唑替尼或福来替尼预处理进行处理。使用指定抗体对细胞裂解产物进行蛋白质印迹分析。(图3h)具有或不具有c-met敲低的parp1敲低细胞中的野生型或突变体的再表达用parp抑制剂ag014699处理并经克隆形成分析。显示了代表性图像和定量。(图3i)在parp1敲低细胞中具有野生型或突变型的再表达且具有或不具有c-met敲低的细胞用50μmabt-888处理18小时。γh2ax通过免疫荧光共聚焦显微法检测。显示了定量。误差棒代表标准差。*p<0.05,t检验。**p>0.05。abt,abt-888;cri,克唑替尼;ft,福来替尼。
图4a-s.靶向tnbc中的parp1和c-met的临床相关性和潜在治疗策略。(图4a)tnbc患者中的parp1的py907与c-met之间的关联,和针对parp1-py907和c-met的在人乳腺癌的组织微阵列中的免疫组织化学染色的代表性图像。通过皮尔逊卡方检验(p=0.021)进行相关分析。(图4b)通过细胞活力分析测量tnbc细胞系(mba-md-231和hcc1937)中的c-met和parp抑制的协同效应。(图4c和4e)通过mba-md-231细胞和hcc1937细胞中的克隆形成细胞存活分析测量c-met抑制剂克唑替尼(cri)和parp抑制剂ag014699(ag)的协同效应。(图4d和4f)通过mba-md-231细胞和hcc1937细胞中的克隆形成细胞存活分析测量c-met和parp抑制剂(ag014699和克唑替尼)的协同效应。数据显示为三重复实验(n=3)的平均值±sem。*p<0.05,t检验。(图4g)通过mba-md-231细胞和hcc1937细胞中的软琼脂分析测量c-met抑制剂克唑替尼(cri)和parp抑制剂ag014699(ag)的协同效应。(图4h)通过细胞活力分析测量的a1034细胞的c-met和parp抑制的协同效应。(图4i)a1034细胞用h2o2或克唑替尼预处理加h2o2处理,并用指定抗体对细胞裂解产物进行蛋白质印迹法。(图4j)mda-mb-231细胞接种到裸小鼠的乳腺脂肪垫中(每组n=10)。克唑替尼(5mg/kg)、ag014699(5mg/kg)或组合每周口服5天,持续指定时间。在指定时间测量肿瘤大小。(图4k)mda-mb-231细胞接种到裸小鼠的乳腺脂肪垫中(每组n=10)。福来替尼(5mg/kg)、abt-888(25mg/kg)或组合每周口服5天,持续指定时间。在指定时间测量肿瘤大小。(图l-o)mda-mb-231异种移植肿瘤组织的tunel、ki67、γh2ax和parp1-py907染色。显示了定量。(图4p)通过细胞活力分析测量的mcf-7/c-met细胞的c-met和parp抑制的协同效应。(图4q)mcf-7/c-met细胞用h2o2或克唑替尼预处理加h2o2处理,并用指定抗体对细胞裂解产物进行蛋白质印迹法。(图4r)具有c-met的异位表达的mcf-7细胞接种到裸小鼠的乳腺脂肪垫中(每组n=10)。克唑替尼(5mg/kg)、ag014699(5mg/kg)或组合每周口服5天,持续指定时间。在指定时间测量肿瘤大小。(图4s)h1993细胞被皮下注射入雌性裸小鼠的右腹中。克唑替尼(5mg/kg)、ag014699(5mg/kg)或组合每周口服5天,持续指定时间。在指定时间测量肿瘤大小。误差棒代表偏差。fa,受影响的部分。ag,ag014699;abt,abt-888;cri,克唑替尼;ft,福来替尼。*p<0.05,t检验。
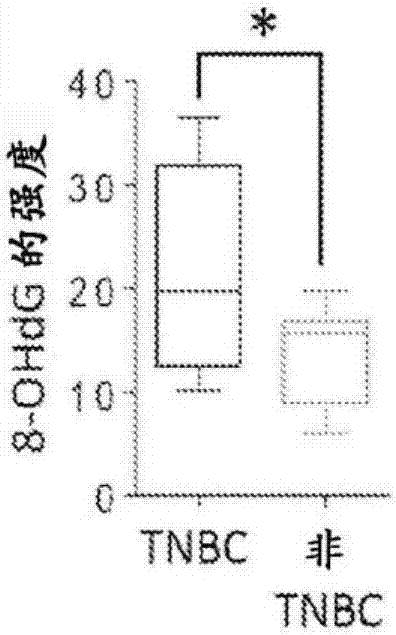
图5.在tnbc中8-ohdg和dcf的含量较高。将乳腺癌细胞(4×103)接种到96孔板上并用10μm的dcf-da处理30分钟。通过酶标仪测量dcf并用graphpadprism分析。显示三重复实验的定量。误差棒代表标准差。*p<0.05,t检验。
图6a-b.erbb3、met和flt3的tcga数据库分析。(图6a)tcga侵袭性乳腺癌患者群的erbb3、met和flt3mrna表达的聚类分析。区分tnbc和其它人乳腺癌(n=1050)的erbb3、met和flt3的分层聚类分析。根据erbb2、esr1和pgr(分别编码her2、er和pr)的表达,通过mrna亚型将乳腺癌患者分类成非tnbc患者(n=819)和tnbc患者(n=231)。显示了通过esr1、pgr和erbb2mrna子类型化从tnbc患者和非tnbc患者中的tnbc相关激酶的原始mrna表达水平和log2转换的mrna表达水平所产生的箱形图。(图6b)乳腺癌细胞系中的c-met表达。用指定抗体对细胞裂解产物进行蛋白质印迹法。***p<0.05,t检验。
图7a-g.c-met与parp1结合。(图7a-b)用v5-parp1和flag-c-met转染hek293t细胞,然后用10μmh2o2处理15分钟。进行免疫沉淀(ip)和蛋白质印迹法以检测parp1与c-met的结合。(图7c)用10μmh2o2处理mda-mb-231细胞15分钟。进行ip/蛋白质印迹法以检测parp1与c-met的结合。(图7d)用10μmh2o2处理hcc1937细胞15分钟。进行ip/蛋白质印迹法以检测parp1与c-met的结合。(图7e-f)用flag-c-met转染mda-mb-436和mcf7细胞,然后用10μmh2o2处理15分钟。进行ip/蛋白质印迹法以检测parp1与c-met的结合。(图7g)用h2o2处理具有shctrl的hela细胞和动力蛋白ic敲低细胞。用指定抗体对细胞进行蛋白质印迹法以实现细胞分级分离。
图8a-m.c-met的下调使细胞对parp抑制剂敏感并减低ros。(图8a)通过细胞活力分析测量的具有c-met敲低的mda-mb-231细胞中的abt-888的ic50。(图8b)c-met敲低细胞用parp抑制剂azd2281处理并经细胞活力分析。右图展示azd2281的ic50。误差棒代表标准差。(图8c)通过细胞活力分析测量的具有c-met敲低的mda-mb-231细胞中的ag014699的ic50。(图8d)mda-mb-231细胞用ag014699和克唑替尼或福来替尼处理并经克隆形成细胞存活分析。展示代表性图像。(图8e-h)c-met敲低细胞用parp抑制剂、abt-888、ag014699、azd2281或bai-201处理并经软琼脂集落形成分析以确定锚定非依赖性细胞生长。显示了代表性图像和ic50。误差棒代表标准差。(图8i)mda-mb-231和hcc1937细胞用c-met抑制剂、克唑替尼(cri)或福来替尼(ft)预处理2小时。细胞然后用10μm的dcf-da处理30分钟。通过酶标仪在495exnm/529emnm的光谱下测量dcf含量。误差棒代表标准差。(图8j)通过plkoshrna得到的c-met敲低mda-mb-231和hcc1937细胞用10μm的dcf-da孵育30分钟。然后通过酶标仪在495exnm/529emnm的光谱下测量dcf含量。误差棒代表标准差。(图8k-l)再表达野生型c-met或激酶死亡c-met的c-met敲低(3’utr)mda-mb-231细胞用abt-888或ag014699处理并经克隆形成细胞存活分析。展示代表性图像。误差棒代表标准差。*p<0.05,ranova。(图8m)具有3’utrc-met敲低和野生型c-met或激酶死亡c-met的再表达的mda-mb-231细胞用parp抑制剂(abt888)处理并经细胞活力分析以确定细胞存活。图的右侧展示敲低细胞和恢复细胞的ic50。数据显示为平均值±s.e.m.且图像是典型的三重复实验(n=3)(*p<0.05,t检验)。
图9a-i.c-met的过表达增加细胞对parp抑制剂的抗性。(图9a-d)过表达c-met的mcf7细胞用parp抑制剂、abt-888、ag014699或azd2281处理,并经细胞活力分析以评估细胞生长和ic50。(图9e-f)过表达c-met的mcf7细胞或对照mcf7细胞用abt-888或ag014699处理并经克隆形成细胞存活分析。展示代表性图像。误差棒代表标准差。(图9g-i)过表达c-met的mcf7细胞或对照mcf7细胞用abt-888、ag014699或bsi-201处理并经软琼脂集落形成分析以确定锚定非依赖性细胞生长。展示代表性图像。误差棒代表标准差。*p<0.05,ranova。
图10a-j.c-met对于parp抑制剂应答是重要的。(图10a)具有c-met敲低的hcc1937细胞用parp抑制剂abt-888处理并经细胞活力分析。在图的右侧显示hcc1937细胞中的abt-888的ic50。误差棒代表标准差。(图10b)通过细胞活力分析测量的具有c-met敲低的hcc1937中的ag014699的ic50。(图10c)表达野生型c-met、激酶死亡(kd)c-met或对照载体的mda-mb-436细胞用abt-888处理并经细胞活力分析。在图的右侧显示mda-mb-436中的abt-888的ic50。误差棒代表标准差。(图10d)通过细胞活力分析测量的具有c-met过表达的mda-mb-436中的ag014699的ic50。(图10e-f)mda-mb-231和mda-mb-436是c-met表达敲低型或过表达c-met。用指定抗体对细胞进行蛋白质印迹法。(图10g-h)brca1或brca2敲低的mda-mb-231和mda-mb-157细胞用abt-888处理并经细胞活力分析。误差棒代表标准差。(图10i-j)brca1或brca2敲低的mda-mb-231细胞用abt-888处理并经克隆形成细胞存活分析。误差棒代表标准差。*p<0.05,**p>0.05,ranova。
图11a-f.c-met参与dna损伤应答。(图11a)c-met敲低mda-mb-231细胞和对照mda-mb-231细胞用h2o2处理指定时间并用甲酰胺基嘧啶dna糖基化酶(fpg)孵育。通过彗星分析评定dna损伤。展示代表性图像。在右侧显示dna损伤程度的定量。(图11b)c-met敲低mda-mb-231细胞和对照mda-mb-231细胞用h2o2和hu/arac处理以诱导并允许dna损伤的积累。hu/arac和h2o2用新鲜培养基替换并孵育指定时间。通过彗星分析评定损伤的dna。显示dna损伤程度的定量。(图11c)将经h2o2处理的荧光素酶质粒转染到c-met敲低mda-mb-231细胞中,24小时后测量荧光素酶活性。数据被显示为三重复实验的平均值±sem(n=3)(*p<0.05,t检验)。(图11d)c-met野生型(wt)表达和激酶死亡突变型(kd)表达的蛋白质印迹分析。(图11e)过表达c-met的mcf7细胞和对照mcf7细胞用h2o2处理指定时间。显示通过彗星分析得到的损伤dna的定量。(图11f)过表达c-met的mcf7细胞用h2o2处理或用克唑替尼预处理15分钟。通过彗星分析评定dna损伤。显示dna损伤程度的定量。误差棒代表标准差。*p<0.05,t检验。
图12a-f.c-met介导parp1在y907上的磷酸化。(图12a)gst融合parp1野生型和parp1y907f和y986f突变体的库马斯蓝染色。(图12b)gst融合野生型parp1和parp1y907f和y986f突变体用[γ-32p]-atp和经过纯化的c-met孵育。通过放射自显影术使parp1中的放射标记atp可见。通过蛋白质印迹法使总gst可见。(图12c)斑点杂交分析用于评估针对parp1的磷酸化y907(py907)的特异性抗体。(图12d)通过蛋白质印迹分析表征parp1敲低mda-mb-231细胞中的抗py907-parp1抗体。也通过共聚焦显微法表征抗体。(图12e)再表达野生型parp1或y907fparp1的parp1敲低mda-mb-231细胞用h2o2处理30分钟并经蛋白质印迹分析。(图12f)将mda-mb-231细胞的c-met表达敲低并用h2o2处理30分钟,并用指定抗体进行蛋白质印迹分析。
图13a-e.由c-met介导的parp1在y907上的磷酸化调节parp抑制剂应答。(图13a)再表达野生型parp1、y907fparp1或y907eparp1的parp1敲低mda-mb-231细胞用h2o2(20μm)处理30分钟。细胞裂解产物接受parp酶活性分析。数据被显示为三重复实验(n=3)的平均值±sem。*p<0.05,t检验。(图13b)通过parp酶活性分析测量的野生型和突变型y907parp之间的abt-888的ic50。(图13c)再表达野生型parp1或突变型parp1的c-met敲低mda-mb-231细胞的蛋白质印迹分析。(图13d-e)来自图13c的对照细胞和c-met敲低细胞用parp抑制剂(abt-888或ag014699)处理并经细胞活力分析。误差棒代表标准差。*p<0.05,ranova。
图14a-c.人乳腺癌肿瘤组织中的py907的临床相关。(图14a)通过乳腺癌患者肿瘤组织的ihc染色进行肽竞争分析以表征py907-parp1抗体。(图14b)非tnbc组织微阵列中的c-met与py907-parp1之间的相关分析。p<0.05,皮尔逊卡方检验。(图14c)tnbc组织微阵列中的8-ohdg与py907-parp1之间的相关分析。通过皮尔逊卡方检验(p=0.019)进行相关分析。图14b和图14c显示阳性情况和阴性情况的代表性染色图像。
图15a-g.c-met表达乳腺癌细胞系中parp抑制剂与c-met抑制剂的组合治疗的协同效应。(图15a)mcf10a细胞用parp抑制剂与c-met抑制剂的组合进行治疗。通过基于chou–talalay方法计算联合指数(ci)确定协同效应。(图15b)通过mba-md-231细胞和hcc1937细胞中的软琼脂分析测量c-met抑制剂克唑替尼(cri)和parp抑制剂ag014699(ag)的协同效应。展示代表性图像。(图15c)mba-md-231细胞用c-met抑制剂福来替尼和parp抑制剂abt-888处理。使用克隆形成细胞存活分析测量细胞生长。通过chou–talalay方法确定ft与abt组合的ci值。显示了代表性图像和定量。误差棒代表标准差。(图15d-f)用parp抑制剂与c-met抑制剂处理a1471和bt549细胞,并通过细胞活力分析测量组合治疗的协同效应。(图15e-g)用h2o2或克唑替尼预处理加h2o2处理a1471和bt549细胞,并用指定抗体对细胞裂解产物进行蛋白质印迹分析。误差棒代表标准差。abt,abt-888;ft,福来替尼;ag,ag014699;cri,克唑替尼。
图16a-b.parp与c-met的组合治疗抑制肿瘤生长。(图16a)将a1034细胞注射入裸小鼠的乳腺脂肪垫中。克唑替尼(5mg/kg)、ag014699(5mg/kg)或组合每周口服5天,持续指定时间。在指定时间测量肿瘤大小。(图16b)将hcc1937细胞注射入裸小鼠的乳腺脂肪垫中。克唑替尼(5mg/kg)、ag014699(5mg/kg)或组合每周口服5天,持续指定时间。在指定时间测量肿瘤大小。误差棒代表标准差。*p<0.05,t检验。
图17a-i:小鼠中c-met与parp1的组合抑制的临床化学和体重分析。(图17a-b)ag014699和克唑替尼对于具有mda-mb-231异种移植肿瘤的裸小鼠的肾功能和肝功能的治疗效果。(图17c)ag014699和克唑替尼治疗之前和ag014699和克唑替尼治疗之后的裸小鼠体重。(图17d-e)abt-888和福来替尼对于具有mda-mb-231异种移植肿瘤的裸小鼠的肾功能和肝功能的治疗效果。(图17f)abt-888和福来替尼治疗之前和abt-888和福来替尼治疗之后的裸小鼠体重。(图17g-h)ag014699和克唑替尼对于具有a1034同基因小鼠肿瘤的fvb小鼠的肾功能和肝功能的治疗效果。(图17i)ag014699和克唑替尼治疗之前和ag014699和克唑替尼治疗之后的fvb小鼠体重。ast,天冬氨酸转氨酸。alt,丙氨酸转氨酶。ag,ag014699;cri,克唑替尼;abt,abt-888;ft,福来替尼;ast,天冬氨酸转氨酸。alt,丙氨酸转氨酶。误差棒代表标准差。
图18a-c.c-met异位表达乳腺癌和nsclc细胞系中parp抑制剂与c-met抑制剂的组合治疗的协同效应。(图18a)具有c-met异位表达的mcf-7细胞用parp抑制剂与c-met抑制剂的组合处理。通过基于chou–talalay方法计算联合指数(ci)确定协同效应。(图18b)用parp抑制剂和c-met抑制剂处理肺癌细胞系h1993和a549,并通过细胞活力分析测量组合治疗的协同效应。(图18c)用h2o2或克唑替尼预处理加h2o2处理h1993和a549细胞,并用指定抗体对细胞裂解产物进行蛋白质印迹分析。abt,abt-888;ft,福来替尼;ag,ag014699;cri,克唑替尼。
图19.提议的模型。ros激活c-met使parp1在tyr907上磷酸化。磷酸化parp1增强其dna修复活性,从而造成parp抑制剂抗性。
具体实施方式
三阴性乳腺癌(tnbc)是一种乳腺癌的高侵袭性亚型,其缺乏雌激素受体(er)的表达、孕酮受体(pr)的表达和her2的过表达或扩增(carey等人,2010)。虽然一些tnbc患者最初对化学疗法有应答,但是其中大多数最终产生抗性(liedtke等人,2008)。目前没有针对tnbc的有效靶向疗法。最近,聚(adp-核糖)聚合酶(parp)抑制剂已作为tnbc患者的有前景的治疗剂出现(anders等人,2010;rouleau等人,2010)。当parp抑制剂依尼帕尼与化学疗法组合时,2期研究报道了改善的临床益处(o’shaughnessy等人,2011);然而,随后的3期研究指出其并不提供显著的总体存活或无进展存活益处(o’shaughnessy等人,2011),并且观察到抗性和低应答率(lord和ashworth,2013)。因此,迫切需要鉴别用于分层患者的潜在生物标志物以增加parp抑制剂治疗的应答率。在这里显示受体酪氨酸激酶c-met与为dna修复所必需的parp1在ityr907(y907)上结合并使parp1磷酸化,从而使tnbc细胞对parp抑制产生抗性。c-met和parp1抑制剂的组合在体外和乳腺癌异种移植肿瘤模型中产生了协同效应。本研究鉴别了生物标志物(py907,parp1),其可以潜在地预测抗性并分层患者以针对具有高c-met表达且不对parp抑制作出应答的患者最大化parp抑制剂和合理组合疗法的治疗益处。parp和c-met抑制剂的组合可用作治疗tnbc的有前景的治疗策略。除乳腺癌以外,这些发现也可为其它癌症类型中的parp抑制提供新的研究途径。
i.本发明的方面
利用作为治疗tnbc的当前治疗策略的化学疗法和放射疗法,50%的患者最初对一线疗法有应答,并且早期应答者具有良好预后。然而,另外50%的患者的疾病在一线疗法后仍残留,这些患者具有不良预后且导致更坏结果。针对这种类型的侵袭性更高且等级更高的乳腺癌缺少特异性靶向疗法是临床上的一个主要挑战。有趣的是,即使fda最近批准了奥拉帕尼用于治疗brca突变型卵巢癌患者,具有野生型brca的一些患者依然显示对于这种抑制剂的应答(tutt等人,2010;gelmon等人,2011;ledermann等人,2014),表明其它机制可以调节parp抑制剂应答。因此,了解tnbc的特征和开发将会使tnbc患者受益的机制驱动型靶标疗法是必要的。在这里,揭示了c-met使parp1在y907上磷酸化且调节parp1活性和针对parp抑制的应答所借助的机制(图18a-c)。c-met抑制与parp抑制的组合协同抑制体外细胞生长和体内肿瘤生长。值得注意的是,py907-parp1具有作为预测针对parp抑制剂的抗性和分层患者以从parp抑制获得最大益处的生物标志物的潜力。
值得注意的是,临床研究已经指示大约一半的tnbc患者过表达c-met(zagouri等人,2013;castaldi等人,2010)。有趣的是,不仅在tnbc中,而且在非tnbc中也观察到c-met与py907-parp表达之间的正相关(图4a)。由于parp抑制剂目前在临床试验中用于tnbc,因此聚焦于这个乳腺癌亚型。大约三分之一(图4a的24/77)的对py907/c-met呈阳性的tnbc患者可能对单独的parp抑制剂有抗性,并且可以从使用py907/c-met作为生物标志物的c-met抑制与parp抑制的组合治疗受益。
parp不仅用于tnbc,另外还广泛用于其它癌症类型的临床试验中,并且c-met是在多种癌症类型中过表达的原癌基因(baccaccio和comoglio,2006)。非tnbc(图14b)中观察到的c-met与py907之间的正相关和来自mcf-7细胞(图4p-r和图18a)或nsclc(图4s和图18b-c)中c-met的异位表达的结果表明,c-met在肿瘤中过表达的其它类型的癌症的患者也可受益于这种组合疗法。
ii.治疗方法
本发明的特定方面可用于鉴别和/或治疗基于parp1的tyr907的磷酸化状态的疾病或病症。本发明的其它方面使具有癌症的受试对象对parp抑制剂的治疗敏感。
如本文中所使用的术语“受试对象”或“患者”是指实施本发明方法的任何个体。患者一般是人,但是如所属领域技术人员所了解的,患者也可以是动物。因此,在患者的定义内包括其它动物,包括哺乳动物,例如啮齿动物(包括小鼠、大鼠、仓鼠和豚鼠)、猫、狗、兔、农场动物(包括牛、马、山羊、绵羊、猪等)和灵长动物(包括猴子、黑猩猩、猩猩和大猩猩)。
“治疗”是指对受试对象施用或应用治疗剂或对受试对象实施某种程序或方式以获得疾病或健康相关病状的治疗益处。举例来说,治疗可以包括施用化学疗法、免疫疗法、放射疗法、进行外科手术或其任何组合。
本申请中通篇使用的术语“治疗益处”或“治疗有效”是指通过这种病状的医学治疗促进或增强受试对象的良好状态的任何东西。这包括(但不限于)疾病的征兆或症状的频率或严重性的降低。举例来说,癌症治疗可例如包括肿瘤侵袭性的降低、癌症生长速率的降低或转移的预防。癌症治疗还可指延长具有癌症的受试对象的存活期。
包括组合疗法的方法和组合物增强治疗或预防效果,和/或增加另一种抗癌疗法或抗过度增殖疗法的治疗效果。治疗性和预防性的方法和组合物能以可有效实现期望效果的组合量提供。组织、肿瘤或细胞可以与包含一或多种药剂的一或多种组合物或医药制剂接触,或者组织、肿瘤和/或细胞与两种或更多种不同的组合物或制剂接触。另外,预期此类组合疗法可以和化学疗法、放射疗法、外科疗法或免疫疗法结合使用。
术语“接触”和“暴露”在应用于细胞时在本文中用于描述将治疗构建体和化学治疗剂或放射治疗剂递送给靶细胞或直接毗邻靶细胞放置的过程。举例来说,为了实现细胞杀死,两种药剂以可有效杀死细胞或防止细胞分裂的组合量递送给细胞。
本文中所述的方法适用于治疗癌症。一般来说,术语“癌症”和“癌性”是指或描述典型特征为细胞生长失调的哺乳动物的生理疾病。更具体地说,使用任何一或多种parp抑制剂或其变异体且结合本文中提供的方法治疗的癌症包括(但不限于)实体肿瘤、转移癌或非转移癌。在特定实施方案中,癌症可以起源于膀胱、血液、骨头、骨髓、脑、乳房、结肠、食道、十二指肠、小肠、大肠、结肠、直肠、肛门、牙龈、头、肾、肝、肺、鼻咽、颈、卵巢、胰腺、前列腺、皮肤、胃、睾丸、舌头或子宫。
癌症尤其可以是以下组织学类型,但不限于:恶性肿瘤;癌;淋巴瘤;胚细胞瘤;肉瘤;未分化癌;巨细胞和梭形细胞癌;小细胞癌;小细胞肺癌;非小细胞肺癌;乳头状癌;鳞状细胞癌;淋巴上皮癌;基底细胞癌;毛母质癌;移行细胞癌;乳头状移行细胞癌;腺癌;恶性促胃液素瘤;胆管癌;肝细胞癌;组合型肝细胞癌和胆管癌;小梁状腺癌;腺样囊性癌;腺瘤性息肉腺癌;家族性结肠息肉病腺癌;实体癌;恶性类癌瘤;细支气管肺泡癌;乳头状腺癌;嫌色细胞癌;嗜酸细胞癌;嗜酸性腺癌;嗜碱细胞癌;透明细胞腺癌;颗粒细胞癌;滤泡状腺癌;乳头状和滤泡状腺癌;非包裹性硬化癌;肾上腺皮质细胞癌;子宫内膜癌;皮肤附件癌;大汗腺腺癌;皮脂腺癌;盯聍腺腺癌;粘液表皮样癌;囊腺癌;乳头状囊腺癌;乳头状浆液性囊腺癌;粘液性囊腺癌;粘液腺癌;印戒细胞癌;浸润性导管癌;髓样癌;小叶癌;炎性癌;乳腺佩吉特氏病;腺泡细胞癌;腺鳞癌;具有鳞状细胞化生的腺癌;恶性胸腺瘤;恶性卵巢间质肿瘤;恶性泡膜细胞瘤;恶性粒层细胞瘤;恶性睾丸母细胞瘤;塞尔托利氏细胞癌;恶性莱迪希细胞瘤;恶性脂质细胞瘤;恶性副神经节瘤;恶性乳腺外副神经节瘤;嗜铬细胞瘤;血管球肉瘤;恶性黑素瘤;无色素性黑素瘤;浅表扩散性黑素瘤;巨大色素痣恶性黑素瘤;恶性雀斑样痣黑素瘤;肢端雀斑样痣黑素瘤;结节性黑素瘤;上皮样细胞黑素瘤;恶性蓝痣;肉瘤;纤维肉瘤;恶性纤维组织细胞瘤;粘液肉瘤;脂肪肉瘤;平滑肌肉瘤;横纹肌肉瘤;胚胎性横纹肌肉瘤;腺泡状横纹肌肉瘤;间质肉瘤;恶性混合瘤;米勒管混合瘤;肾母细胞瘤;肝母细胞瘤;癌肉瘤;恶性间质瘤;恶性布伦纳瘤;恶性叶状肿瘤;滑膜肉瘤;恶性间皮瘤;无性细胞瘤;胚胎癌;恶性畸胎瘤;恶性卵巢甲状腺瘤;绒毛膜癌;恶性中肾瘤;血管肉瘤;恶性血管内皮瘤;卡波西氏肉瘤;恶性血管外皮细胞瘤;淋巴管肉瘤;骨肉瘤;皮质旁骨肉瘤;软骨肉瘤;恶性软骨母细胞瘤;间胚叶性软骨肉瘤;骨巨细胞瘤;尤文肉瘤;恶性牙源性肿瘤;成釉细胞牙肉瘤;恶性成釉细胞瘤;成釉细胞纤维肉瘤;恶性松果体瘤;脊索瘤;恶性神经胶质瘤;室管膜瘤;星形细胞瘤;原浆性星形细胞瘤;纤维型星形细胞瘤;星形母细胞瘤;胶质母细胞瘤;少突神经胶质瘤;成少突神经胶质细胞瘤;原始神经外胚叶;小脑肉瘤;节细胞神经母细胞瘤;成神经细胞瘤;视网膜母细胞瘤;嗅神经源性肿瘤;恶性脑膜瘤;神经纤维肉瘤;恶性神经鞘瘤;恶性颗粒细胞瘤;b细胞淋巴瘤;恶性淋巴瘤;霍奇金病;霍奇金氏;低等级/滤泡性非霍奇金淋巴瘤;副肉芽肿;恶性小淋巴细胞淋巴瘤;恶性弥散性大细胞淋巴瘤;恶性滤泡性淋巴瘤;蕈样肉芽肿;套细胞淋巴瘤;华氏巨球蛋白血症;其他指定的非霍奇金淋巴瘤;恶性组织细胞增多病;多发性骨髓瘤;肥大细胞肉瘤;免疫增生性小肠疾病;白血病;淋巴细胞性白血病;浆细胞白血病;红白血病;淋巴肉瘤细胞性白血病;骨髓性白血病;嗜碱粒细胞性白血病;嗜酸性粒细胞性白血病;单核细胞性白血病;肥大细胞白血病;巨核母细胞白血病;髓样肉瘤;慢性淋巴细胞白血病(cll);急性淋巴细胞白血病(all);毛细胞白血病;慢性成髓细胞白血病;和毛细胞白血病。
患者的有效应答或患者对治疗的“应答性”是指被赋予处在疾病或病症风险下或罹患疾病或病症的患者的临床或治疗益处。这种益处可以包括细胞应答或生物应答、完全应答、部分应答、稳定疾病(无发展或复发)或存在延后复发的应答。举例来说,有效应答可以是被诊断患有癌症的患者的肿瘤大小变小或无进展存活期。
聚(adp-核糖)聚合酶1在促进dna修复、控制rna转录、介导细胞死亡和调节免疫应答中具有关键作用。parp1抑制剂是酶parp1的一组药理学抑制剂(参看np_001609.2,其以引用的方式并入本文中)。在各种临床前癌症模型和人临床试验中,已经显示parp1抑制剂可通过增加癌细胞的凋亡、限制肿瘤生长、减少转移和延长荷瘤受试对象的存活期来增强放射和化学疗法的作用(wo2007/084532;donawho等人,2007;kummar等人,2009)。举例来说,parp1抑制剂包括(但不限于)奥拉帕尼(azd-2281)、维利帕尼(abt-888)、依尼帕尼(bsi-201)、瑞卡帕布(ag014699、pf-01367338)、ag14361、ino-1001、a-966492、pj34、mk-4827、cep9722、bnm-673、3-氨基苯甲酰胺,和美国专利号7,928,105、美国专利号8,124,606、美国专利号8,236,802、美国专利号8,450,323、wo2006/110816、wo2008/083027和wo2011/014681中公开的那些。
失调的c-met受体酪氨酸激酶途径在肿瘤形成、生长、维持和发展中起作用(birchmeier等人,2003;boccaccio等人,2006;christensen等人,2005)。c-met在显著比例的人癌症中过表达,并且通常和不良临床结果有关,例如侵袭性更高的疾病、疾病发展、肿瘤转移和患者存活期缩短。c-met抑制剂是受体酪氨酸激酶c-met的一组药理学抑制剂。c-met受体酪氨酸激酶抑制剂包括(但不限于)incb28060(wo2008/064157)、arq197(替凡替尼)、amg458、gsk1363089(xl880或福来替尼)、e7050(戈瓦替尼)、mk-2461、bms-777607、jnj-38877605、xl184(卡博替尼)、amg337、arq197、mgcd265、pf04217903和pf02341066(克唑替尼)。
关于肿瘤疾病治疗,视肿瘤疾病的阶段而定,肿瘤疾病治疗包括以下疗法中的一种或组合:用于移除肿瘤组织的外科手术、放射疗法和化学疗法。其它治疗方案可以和抗癌剂的施用组合,例如治疗组合物和化学治疗剂。举例来说,想要用此类抗癌剂治疗的患者也可以接受放射疗法和/或可以进行外科手术。
对于疾病的预防或治疗,治疗组合物(例如parp抑制剂或c-met抑制剂)的适当剂量将取决于欲治疗的疾病类型(如上文所定义)、疾病的严重性和过程、药剂的施用是用于预防目的还是治疗目的、先前疗法、患者的临床病史和对药剂的应答和医生的判断。药剂适合一次性施用给患者,或经过一系列治疗施用给患者。
组合治疗
包括组合疗法的方法和组合物增强治疗或预防效果,和/或增加另一种抗癌疗法或抗过度增殖疗法的治疗效果。治疗性和预防性的方法和组合物能以可有效实现期望效果(例如杀死癌细胞和/或抑制细胞过度增殖)的组合量提供。组织、肿瘤或细胞可以与包含一或多种药剂的一或多种组合物或医药制剂接触,或者组织、肿瘤和/或细胞与两种或更多种不同的组合物或制剂接触。另外,预期此类组合疗法可以和放射疗法、外科疗法或免疫疗法结合使用。
组合施用可以包括以同一个剂型同时施用两种或更多种药剂、以分开的剂型同时施用、和分开施用。也就是说,本发明的治疗组合物和另一种治疗剂可以一起配制成同一个剂型并同时施用。或者,本发明的治疗组合物和另一种治疗剂可以同时施用,其中这两种药剂存在于分开的制剂中。在另一个替代方案中,在施用治疗剂后可以紧接着施用另一种治疗剂,反过来也一样。在分开施用方案中,本发明的治疗组合物和另一种治疗剂可以间隔几分钟、或间隔几小时、或间隔几天施用。
第一抗癌治疗可以在第二抗癌治疗之前、期间、之后或以各种组合施用。给药的间隔可以在从同时到几分钟到几天到几周的范围内。在与第二治疗分开向患者提供第一治疗的实施方案中,一般将确保在每个递送时间之间的显著时间段不会结束,使得两种化合物仍然能够对患者发挥有利的组合作用。在这些情况下,预期可以在彼此间隔约12小时到24小时或72小时内且更具体地在彼此间隔约6-12小时内为患者提供第一疗法和第二疗法。在一些情况下,显著延长治疗的时间周期可能是合乎需要的,其中在对应的给药之间间隔几天(2、3、4、5、6或7)到几周(1、2、3、4、5、6、7或8)。
在特定实施方案中,治疗过程将持续1-90天或更长(这个范围包括间隔的天数)。预期一种药剂可以在第1天到第90天(这个范围包括间隔的天数)中的任一天或其任何组合时给予,并且另一种药剂在第1天到第90天(这个范围包括间隔的天数)中的任一天或其任何组合时给予。在一天(24小时的周期)内,患者可被给予一次或多次药剂施用。此外,在治疗过程后,预期存在不施用抗癌治疗的时间周期。这个时间周期可以持续1-7天和/或1-5周和/或1-12个月或更长(这个范围包括间隔的天数),这取决于患者的状况,例如其预后、力量、健康等。预期治疗周期将在必要时重复。
可以使用各种组合。对于下文的实例来说,parp抑制剂和/或c-met抑制剂是“a”并且另一种抗癌疗法是“b”。
a/b/ab/a/bb/b/aa/a/ba/b/bb/a/aa/b/b/bb/a/b/b
b/b/b/ab/b/a/ba/a/b/ba/b/a/ba/b/b/ab/b/a/a
b/a/b/ab/a/a/ba/a/a/bb/a/a/aa/b/a/aa/a/b/a
本发明的任何化合物或疗法对于患者的施用将遵循用于施用此类化合物的一般方案,同时考虑药剂的毒性(如果有的话)。因此,在一些实施方案中,存在监测可归因于组合疗法的毒性的步骤。
1.化学疗法
根据本发明可使用多种多样的化学治疗剂。术语“化学疗法”是指使用药物治疗癌症。“化学治疗剂”用于表示在癌症治疗中施用的化合物或组合物。这些药剂或药物通过其在细胞内的作用模式(例如其是否影响细胞周期和在哪个阶段影响细胞周期)被分类。或者,药剂可以基于其直接交联dna、插入dna中或通过影响核酸合成来诱导染色体畸变和有丝分裂畸变的能力被表征。
化学治疗剂的实例包括烷基化剂,例如噻替派和环磷酰胺;烷基磺酸盐,例如白消安、英丙舒凡和哌泊舒凡;氮杂环丙烷,例如苯佐替派、卡波醌、美美妥替哌和乌瑞替哌;乙撑亚胺和甲基三聚氰胺,包括六甲蜜胺、三亚乙基三聚氰胺、三亚乙基磷酰胺、三亚乙基硫代磷酰胺和三羟甲基三聚氰胺;多聚乙酰(尤其是布拉它辛和布拉它辛酮);喜树碱(包括合成类似物托泊替康);苔藓虫素;苔藓抑素;cc-1065(包括其阿多来新、卡折来新和比折来新合成类似物);隐藻素(尤其是隐藻素1和隐藻素8);多拉司他汀;倍癌霉素(包括合成类似物kw-2189和cb1-tm1);软珊瑚醇;水鬼蕉碱;匍枝珊瑚醇;软海绵素;氮芥,例如苯丁酸氮芥、萘氮芥、胆磷酰胺、雌莫司汀、异环磷酰胺、二氯甲基二乙胺、盐酸氧氮芥、美法仑、新氮芥、苯芥胆甾醇、泼尼莫司汀、曲洛磷胺和尿嘧啶氮芥;亚硝基脲,例如卡莫司汀、吡葡亚硝脲、福莫司汀、洛莫司汀、尼莫司汀和雷莫司汀;抗生素,例如烯二炔类抗生素(例如卡奇霉素,尤其是卡奇霉素γ1i和卡奇霉素ωi1);达内霉素,包括达内霉素a;二膦酸盐,例如氯膦酸盐;埃斯培拉霉素;以及新制癌菌素发色团和相关色素蛋白烯二炔抗生素发色团、阿克拉霉素、放线菌素、authrarnycin、重氮丝氨酸、博莱霉素、放线菌素c、carabicin、洋红霉素、嗜癌菌素、色霉素、放线菌素d、正定霉素、地托比星、6-重氮-5-氧代-l-正亮氨酸、阿霉素(包括吗啉基-阿霉素、氰基吗啉基-阿霉素、2-吡咯啉基-阿霉素和去氧阿霉素)、表柔比星、依索比星、伊达比星、麻西罗霉素、丝裂霉素(例如丝裂霉素c)、霉酚酸、诺拉霉素、橄榄霉素、培洛霉素、波非罗霉素、嘌呤霉素、三铁阿霉素、罗多比星、链黑菌素、链佐星、杀结核菌素、乌苯美司、净司他丁和佐柔比星;抗代谢物,例如甲氨蝶呤和5-氟尿嘧啶(5-fu);叶酸类似物,例如二甲叶酸、蝶罗呤和三甲曲沙;嘌呤类似物,例如氟达拉滨、6-巯基嘌呤、硫咪嘌呤和硫代鸟嘌呤;嘧啶类似物,例如安西他滨、阿扎胞苷、6-氮杂尿苷、卡莫氟、阿糖胞苷、二去氧尿苷、去氧氟尿苷、依诺他滨和氟尿苷;雄激素,例如7β,17α-二甲睾酮、2-甲氧睾酮丙酸酯、环硫雄醇、美雄烷和睾内酯;抗肾上腺药物,例如米托坦和曲洛司坦;叶酸补充剂,例如亚叶酸;醋葡醛内酯;醛磷酰胺糖苷;氨基乙酰丙酸;恩尿嘧啶;安吖啶;优布希;比生群;达曲沙;地磷酰胺;地美可辛;亚胺醌;依氟鸟氨酸;依利醋铵;埃博霉素;依托格鲁;硝酸镓;羟基脲;香菇多糖;氯尼达明;美登素类化合物,例如美登木碱和安丝菌素;米托胍腙;米托蒽醌;莫匹达谋;nitraerine;喷司他丁;蛋氨氮芥;吡柔比星;洛索蒽醌;鬼臼酸;2-乙基酰肼;甲基苄肼;psk多糖复合物;雷佐生;根霉素;西佐喃;螺旋锗;细交链孢菌酮酸;三乙撑亚胺苯醌;2,2',2"-三氯三乙胺;单端孢霉烯(尤其是t-2毒素、疣孢菌素a、杆孢菌素a和蛇形菌素);聚氨酯;长春地辛;氮烯唑胺;甘露醇氮芥;二溴甘露醇;二溴卫矛醇;哌血生;gacytosine;阿糖胞苷("ara-c");环磷酰胺;紫杉烷类,例如紫杉醇和多西他赛吉西他滨;6-硫代鸟嘌呤;巯基嘌呤;铂配位络合物,例如顺铂、奥沙利铂和卡铂;长春碱;铂;依托泊苷(vp-16);异环磷酰胺;米托蒽醌;长春新碱;长春瑞滨;诺安托;替尼泊苷;依达曲沙;道诺霉素;氨基蝶呤;希罗达;伊班膦酸盐;伊立替康(例如cpt-11);拓扑异构酶抑制剂rfs2000;二氟甲基鸟氨酸(dmfo);类视黄醇,例如视黄酸;卡培他滨;卡铂、甲基苄肼、普卡霉素、吉西他滨、诺维本、法尼基-蛋白转移酶抑制剂、反铂和任何上述物质的药学上可接受的盐、酸或衍生物。
2.放射疗法
引起dna损伤并已被广泛使用的其它因素包括通常被称为γ射线、x射线和/或放射性同位素对肿瘤细胞的定点递送的那些。也涵盖dna损伤因素的其它形式,例如微波、质子束照射(美国专利5,760,395和4,870,287)和uv照射。最有可能的是,所有这些因素都影响对于dna、对于dna前体、对于dna复制和修复和对于染色体组装和维持的大范围的损伤。x射线的剂量范围从长时期(3到4周)的50到200伦琴的每日剂量到2000到6000伦琴的单次剂量不等。放射性同位素的剂量范围差别很大,并且取决于同位素的半衰期、所发射的辐射的强度和类型、和癌细胞的摄取。
3.免疫疗法
所属领域技术人员将了解,另外的免疫疗法可以和本发明的方法组合或结合使用。在癌症治疗的情况下,免疫治疗剂一般依赖于使用免疫效应细胞和分子靶向和破坏癌细胞。利妥昔单抗
在免疫疗法的一个方面中,肿瘤细胞必须具有能够被靶向、即不存在于大多数其它细胞上的某种标志物。存在许多肿瘤标志物,并且其中的任一个可能都适合于本发明的情况下的靶向。常见肿瘤标志物包括cd20、癌胚抗原、酪氨酸酶(p97)、gp68、tag-72、hmfg、唾液酸化的路易斯抗原、muca、mucb、plap、层粘连蛋白受体、erbb和p155。免疫疗法的一个替代方面是组合抗癌效果与免疫刺激效果。也存在免疫刺激分子,包括:细胞因子,例如il-2、il-4、il-12、gm-csf、γ-ifn;趋化因子,例如mip-1、mcp-1、il-8;和生长因子,例如flt3配体。
当前处在研究中或使用中的免疫疗法的实例是免疫佐剂,例如牛分枝杆菌、恶性疟原虫、二硝基氯苯和芳香族化合物(美国专利5,801,005和5,739,169;hui和hashimoto,1998;christodoulides等人,1998);细胞因子疗法,例如干扰素α、β和γ、il-1、gm-csf和tnf(bukowski等人,1998;davidson等人,1998;hellstrand等人,1998);基因疗法,例如tnf、il-1、il-2和p53(qin等人,1998;austin-ward和villaseca,1998;美国专利5,830,880和5,846,945);和单克隆抗体,例如抗cd20、抗神经节苷脂gm2和抗p185(hollander,2012;hanibuchi等人,1998;美国专利5,824,311)。预期一或多种抗癌疗法可以和本文中描述的抗体疗法一起使用。
4.外科手术
大约60%的癌症患者将接受特定类型的外科手术,包括预防性手术、诊断性手术或分期手术、根治性手术和姑息性手术。根治性手术包括切除术,其中所有或部分的癌组织被物理上去除、切除和/或破坏并且可以和其它疗法结合使用,例如本发明的治疗、化学疗法、放射疗法、激素疗法、基因疗法、免疫疗法和/或替代性疗法。肿瘤切除术是指物理去除至少一部分肿瘤。除了肿瘤切除术以外,外科手术治疗包括激光手术、冷冻手术、电手术和显微控制的手术(莫氏手术)。
在切除部分或所有的癌细胞、癌组织或肿瘤后,在体内可以形成空腔。治疗可以通过灌注、直接注射或接受另一种抗癌疗法的区域的局部应用来实现。所述治疗可以重复,例如每1、2、3、4、5、6或7天重复一次,或每1、2、3、4和5周重复一次,或每1、2、3、4、5、6、7、8、9、10、11或12个月重复一次。这些治疗也可以具有不同的剂量。
5.其它药剂
预期其它药剂可以和本发明的特定方面组合使用以提高治疗的治疗功效。这些其它药剂包括影响细胞表面受体和gap接合的上调的药剂、细胞抑制剂和分化剂、细胞粘附抑制剂、增加过度增殖细胞对凋亡诱导剂的敏感性的药剂、或其它生物药剂。通过提高gap接合的数量增加细胞内信号传导将会增强对于邻近过度增殖细胞群体的抗过度增殖作用。在其它实施方案中,细胞抑制剂或分化剂可以和本发明的特定方面组合使用以提高治疗的抗过度增殖功效。预期细胞粘附抑制剂可以提高本发明的功效。细胞粘附抑制剂的实例是点状粘附激酶(fak)抑制剂和洛伐他汀。进一步预期增加过度增殖细胞对凋亡的敏感性的其它药剂(例如抗体c225)可以和本发明的特定方面组合使用以提高治疗功效。
实施例
包括以下实施例用于证明本发明的优选实施方案。所属领域技术人员应了解,以下实施例中公开的技术代表发明人所发现的用于在本发明的实践中很好地发挥作用的技术,因此可被视为构成其实践的优选模式。然而,根据本公开,所属领域技术人员应了解,公开的具体实施方案可以作出许多改变,并且仍然获得相同或相似结果而不会脱离本发明的精神和范围。
材料和方法
化学品和抗体.过氧化氢(#216763)、放线菌酮(#c4859)和亚砷酸钠溶液(#35000)从sigma-aldrich(st.louis,mo)获得。检测微管蛋白(#t5168)、flag(#f3165)和肌动蛋白(#a2066)的抗体也从sigma-aldrich(st.louis,mo)获得。检测γh2ax(#05-636)和抗-磷酸酪氨酸(#05-321,4g10)的抗体从emdmillipore(billerica,ma)获得。用于蛋白质印迹法的检测gst融合蛋白(#sc-53909)、ha标签(#sc-805)和parp1(#sc-7150)的抗体从santacruzbiotechnology(santacruz,ca)获得。用于免疫沉淀(ip)和用于检测c-met(#8198)和磷酸化c-met(#3077)的针对parp1(#9532)的抗体从cellsignalingtechnology(danvers,ma)获得。针对8-羟基-2’-去氧鸟苷(8-ohdg)的抗体从genoxcorporation(baltimore,md)获得。所有的荧光标记第二抗体都从invitrogen(carlsbad,ca)获得。小鼠抗磷酸化parp1y907抗体是在中国医科大学分子医学中心针对磷酸化合成肽(admvsksan-yp-chtsqgd;seqidno:1)产生的。用于蛋白质印迹法的辣根过氧化物酶(hrp)缀合的第二抗体从ebioscience(sandiego,ca)获得。c-met激酶抑制剂:克唑替尼(#c-7900)和福来替尼(#f-4185),从lclaboratories(woburn,ma)获得。parp抑制剂abt-888(维利帕尼,#ct-a888)、ag014699(瑞卡帕布,#ct-ag01)和bsi201(依尼帕尼,#ct-bsi201)从chemietek(indianapolis,in)获得。ad2281(奥拉帕尼,#s1060)从selleckchemicals(houston,tx)获得。
细胞培养和处理.所有的细胞系都从美国菌种保藏中心(atcc)获得并被维持在补充有10%胎牛血清和抗生素的达尔伯克氏改良伊格培养基(dmem)/f12或rpmi-1640中。a1034和a1471小鼠细胞系由moragpark博士(mcgilluniversity)馈赠。使用
质粒和转染.对于c-met或parp1的稳定敲低和c-met或parp1过表达研究,用pgipzshrna(对照)载体(thermofisherscientific,rockford,il)或plkoshrna载体(sigma-aldrich,st.louis,mo)和pcdh-neo载体(systembiosciences,mountainview,ca)转染乳腺癌细胞。表1提供关于实验中所用的shrna的详细信息。
表1.shrna序列.
8-ohdgelisa分析.通过使用dneasyblood&tissuekit(qiagen,valencia,ca)从乳腺癌细胞纯化总dna。通过使用8-ohdgelisa试剂盒(abcam,boston,ma)测量乳腺癌细胞中的8-羟基-2-去氧鸟苷(8-ohdg)含量。计算每个细胞系中8-ohdg含量的平均值±s.d.。
ros检测.将细胞接种到12孔板或96孔板中。在过夜生长后,在pbs中用10μm2’,7’-二氯荧光素二乙酸酯(dcfda)孵育细胞1小时。洗涤细胞并用pbs替换培养基。在利用495exnm/529emnm光谱的蔡司显微镜下测量2’,7’-二氯荧光素(dcf)。计算每个细胞系中的来自5个图像的dcf强度的平均值±s.d.。细胞也接种到96孔板中。在过夜孵育后,在pbs中用10μm2’,7’-二氯荧光素二乙酸酯(dcfda)处理细胞。孵育1小时后,用pbs替换培养基。在利用495exnm/529emnm光谱的酶标仪下测量2’,7’-二氯荧光素(dcf)。计算每个细胞系中dcf含量的平均值±s.d.。
受体酪氨酸激酶抗体阵列.humanphospho-rtkarraykit(ary001b)从r&dsystems(minneapolis,mn)购得。对于parp1相关蛋白研究,改良制造商的方案。简言之,用亚砷酸钠(as)处理具有内源parp1敲低和ha标记野生型parp1的再表达的mda-mb-231细胞以诱导ros。遵照方案的指示,用阵列膜孵育细胞裂解产物。使用hrp缀合ha(#26183-hrp;thermofisherscientific,rockford,il)让膜显影。使用gs-800校准型密度计(bio-radlaboratories,hercules,ca)定量膜的密度。
分层聚类和显示.如先前所述(hsu等人,2014),用clusterandtreeview(eisen等人,1998)程序分析聚类。简言之,所用的分层聚类算法是密切地基于sokal等人(1958)的平均连接方法。对于靶标受体酪氨酸激酶的任何集合,通过使用平均连接聚类来计算上对角相似矩阵。通过如先前所述(sokal等人,1958)计算树状图来确定这个算法。通过基于测得的荧光比对每个细胞着色而以图表表示热图。对数比0(比值1.0指示基因未改变)被着色成黑色,正值对数比被着色成红色,负值对数比被着色成绿色。
免疫沉淀和免疫印迹法.对于免疫沉淀(ip),用1μg的小鼠或兔igg和20μl的蛋白g-琼脂糖(roche,indianapolis,in)在4℃下将如文中所述经过处理的细胞的裂解产物预清除1小时。在轻微搅拌下,用1μg的第一抗体或抗-igg抗体在4℃下孵育预清除的裂解产物。进一步在4℃下用蛋白g-琼脂糖孵育裂解产物30分钟。收集蛋白g-琼脂糖沉淀并在4℃下洗涤三次。如先前所述(du等人,2014)对免疫沉淀物进行十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(sds-page)和蛋白质印迹分析。
共聚焦显微分析.对于固定细胞,如先前所述(eisen等人,1998)进行共聚焦显微分析。简言之,如文中所述的那样处理腔室玻片(labtek,scottsvalley,ca)上生长的细胞。在用冰冷pbs洗涤后,将细胞固定,渗透化,并用第一抗体和荧光标记的第二抗体孵育。使用具有63x/1.4物镜的zeisslsm710激光扫描显微镜(carlzeiss,thornwood,ny)检查免疫染色细胞。使用zen和axiovison(carlzeiss)和imagej软件程序(nationalinstitutesofhealth,bethesda,md)进行数据分析。计算每个处理组中的来自5个图像的指定抗体的荧光强度的平均值±s.d.。
duolink分析.将细胞接种到腔室玻片中并用文中所述的相关试剂处理。然后将细胞固定,渗透化,并用3%bsa封闭。第一抗体与细胞和一对寡核苷酸标记的抗体(pla探针)一起孵育。根据制造商的方案(duolinkassaykit,sigma-aldrich)进行连接和扩增,随后安装玻片以供在共聚焦显微镜下测量。计算每个处理组中的来自20个细胞的pla信号强度的平均值±s.d.。
细胞分级分离.如先前所述(eisen等人,1998)的那样制备细胞溶质部分和核部分。简言之,将细胞用冰冷的磷酸盐缓冲盐水(pbs)洗涤,用低渗缓冲液[10mmtris-hcl,ph8.0,10mmkcl,0.5%np-40,2mmmgcl2,0.5mm二硫苏糖醇(dtt),1mmpmsf,和0.15u/ml抑酶肽]溶胀,然后使用dounce匀浆器均质化。通过在600×g下离心5分钟使核沉淀,并收集上清液作为细胞溶质部分。将核沉淀溶解在netn缓冲液[10mmtris-hclph8.0,150mmnacl,1%np-40,1%脱氧胆酸钠,0.1%sds,0.5mm二硫苏糖醇(dtt),1mmpmsf,和0.15u/ml抑肽酶]中并声波降解,随后在16,000×g下离心5分钟。
细胞活力分析:将细胞(1,500)接种到96孔板中并用指定抑制剂处理72小时。然后在新鲜培养基中用100μm刃天青孵育细胞1小时。通过荧光酶标仪在560exnm/590emnm的光谱下测量细胞活力。相对于来自三个独立实验的经dmso处理的对照,存活曲线被表示为平均值±s.d.。
克隆形成细胞存活分析.将细胞接种到12孔板或24孔板中。在过夜孵育后,用抑制剂处理细胞,随后孵育8天。将集落固定并用0.5%结晶紫染色,洗涤,干燥并成像。用甲醇从集落分解出结晶紫并在540nm下测量。基于540nm下的吸光度,相对于来自三个独立实验的经dmso处理的对照,存活曲线被表示为百分率±s.d.。
荧光素酶修复分析.用h2o2破坏荧光素酶质粒。将经纯化的损伤质粒转染到细胞中。在转染后24小时测量荧光素酶活性。
软琼脂锚定非依赖性细胞生长分析.将含有dmem/f12培养基、10%fbs和0.5%琼脂的细胞生长培养基的基层铺在6孔板中(每孔1.5ml)。在基层凝固后,接种含有dmem/f12培养基、10%fbs和0.35%琼脂糖的顶层(每孔1.5ml)和细胞。将培养基(1ml)添加到每个孔中并每3天更换一次。培养4周后,用0.005%结晶紫对集落染色。用imagej软件对集落计数。相对于来自三个独立实验的经dmso处理的对照,存活曲线被表示为平均值±s.d.。
药物组合的协同定量.通过细胞活力分析、克隆形成细胞存活分析、或软琼脂锚定非依赖性细胞生长分析测量细胞生长。通过计算联合指数(ci)的chou–talalay方法(chou,2010)确定协同效应。
患者组织样品和免疫组织化学染色.人乳腺癌组织微阵列(#brc2281,#brc1021)从pantomics(richmond,ca)获得。人肿瘤组织标本从接受乳腺癌手术切除作为主要治疗的患者获得,所述手术切除在md安德森癌症中心或马偕纪念医院(台湾省台北市)在1995年与2009年之间在md安德森的伦理审查委员会所批准的指导方针下进行,并且在所有病例中在报名时从患者获得知情同意书(hsu等人,2014)。用针对8-ohdg、c-met和py907-parp1的第一抗体孵育组织微阵列,用生物素缀合的第二抗体和亲合素-过氧化物酶检测,并通过氨基乙基咔唑色原体显现。用acis(dakonorthamerica,carpinteria,ca)分析图像。为了验证p-y907-parp1抗体在ihc中的特异性,通过用p-y907-parp1抗体对人乳腺癌样品染色来进行肽竞争分析,所述p-y907-parp1抗体经过模拟物或磷酸化y907-parp1-肽、非磷酸化y907-parp1-肽或另一种磷酸化酪氨酸肽p-y986-parp1封闭。患者肿瘤样品的ihc染色的程序是类似的。
彗星分析.进行彗星分析以检测dna链断裂。如文中所述的那样处理细胞。收集细胞并用pbs洗涤,然后与1%低熔点琼脂糖(sigma,vii型)混合。将细胞和琼脂糖的混合物铺到用0.5%琼脂糖预涂布的载玻片上并用盖玻片在冰上覆盖。5分钟后,轻轻移开盖玻片,涂覆第三层0.5%琼脂糖,然后把盖玻片放回原位。在琼脂糖凝固后,移走盖玻片并将载玻片浸在预冷却的裂解缓冲液(2.5mnacl,0.1medta,10mmtris-hclph7.7,1%tritonx-100,1%dmso)中过夜。将载玻片在水中洗涤三次历时5分钟,并用甲酰胺基嘧啶dna糖基化酶(fpg)(2u/载玻片,enzymatics,beverly,ma)在37℃下孵育1小时。然后将载玻片在碱性电泳缓冲液(50mmnaoh,1mmedta,1%dmso,ph12.8)中在4℃孵育30分钟,随后在22v下电泳20分钟。用0.4mtris-hclph7.0中和载玻片,并用pbs中的2μm碘化丙啶染色30分钟。用荧光显微镜成像彗星尾,并使用imagej软件分析。计算每个处理组中的来自20个细胞的尾部中的dna强度的平均值±s.d.。
体外激酶分析.通过诱导异丙基β-d-1-硫代半乳糖吡喃糖苷(iptg)表达重组gst-wt-parp1片段、gst-y907f-parp1片段和gst-y986f-parp1片段,并用谷胱甘肽琼脂糖小球纯化。用冷pbs洗涤3次后,用500μl1×激酶缓冲液悬浮小球,其中留出50μl用于gst蛋白质印迹法。将小球离心,并在30℃下将100μmatp、0.5μg人重组活性c-met蛋白和50μci[γ-32p]-atp添加到50μl激酶缓冲液中持续15-30分钟。通过在sds加载染料中在100℃加热5分钟来终止激酶反应。对样品进行两个相同的sds-page分析。一个用于gst融合parp1蛋白质的库马斯蓝染色。干燥第二个凝胶并将其用于通过放射自显影检测底物的磷酸化。
parp酶活性分析.通过使用来自emdmillipore的商业分析试剂盒测量parp1酶活性。遵照制造商的方案并进行细微改动,将500ng总裂解产物添加到每个反应中。parp抑制剂abt-888的剂量范围是从0.01μm到1,000μm。用底物孵育后确定野生型和突变体的parp酶活性,通过酶标仪测量。
小鼠异种移植模型.所有的动物程序都在md安德森癌症中心在机构动物管理和使用委员会(iacuc)的批准下(协议号10-14-07231)进行。在6-8周大的雌性裸小鼠的乳腺脂肪垫中注射0.5×106mda-mb-231细胞、2×106hcc1937细胞或5×106mcf-7细胞。在6-8周大的雌性fbv的乳腺脂肪垫中注射0.5×106a1034细胞。将h1993细胞皮下注入6-8周大的雌性裸小鼠的右腹中。对于mda-mb-231和a1034细胞异种移植小鼠模型,在注射后用ivisimagingsystem对小鼠成像5天,并分到处理组中(n=10)。将克唑替尼(5mg/kg)和福来替尼(5mg/kg)、ag014699(5mg/kg)和abt-888(25mg/kg)溶解在50mm乙酸钠水溶液(ph4)中,并作为单一药剂或以组合形式施用。在处理2周后,使用ivisimagingsystem评定肿瘤大小。对小鼠注射100μl的d-荧光素(xenogen;15mg/ml的pbs溶液)。10分钟后,用氧气和异氟烷的混合物(inhalationanesthesiasystem;matrixmedical,orchardpark,ny)麻醉小鼠,并用ivisimagingsystem成像。在各实验之间维持成像参数以用于比较分析。在最后一次处理之后收集肿瘤样品,并通过免疫荧光染色分析。对于毒性评定,在处理之前和之后对小鼠称重。在处理后收集血液样品。使用肝标志酶丙氨酸转氨酶(alt)和天冬氨酸转氨酶(ast)和肾标志副产物肌酐和血液尿素氮的生化分析评估治疗毒性。所有的体内实验都在每种条件下用10只小鼠进行。没有使用统计学方法预先确定样品大小。
统计分析.除非另外指出,否则每个样品都是一式三份地分析。每个实验至少重复三次。所有的误差棒都代表标准差。学生t检验用于比较两组独立样品。重复测量anova分析用于评估剂量曲线应答期间的统计显著性。使用皮尔逊卡方检验分析相关。
实施例1-ros诱导c-met和parp1的结合
ros对dna造成的氧化损伤的标志物8-羟基去氧鸟苷(8-ohdg)的含量在tnbc中高于非tnbc中,如乳腺癌组织微阵列的ihc染色(图1a)所示。与乳腺癌患者的肿瘤组织中的观察结果一致,8-ohdg的含量在tnbc细胞系中也高于非tnbc细胞系中,如免疫荧光染色和elisa分析(图1b-d)所见的。ros标志物2',7'-二氯荧光素(dcf)在大多数tnbc细胞系中也高于非tnbc细胞系中(图1e-f和图5)。然后确认tnbc中更高的ros和氧化dna损伤是否开启parp1活性。事实上,tnbc细胞的par化比非tnbc细胞要高(图1g-h)。总地来说,这些结果表明ros的tnbc相关增加可以导致更高的parp1活性以修复由细胞ros诱导的dna损伤。
受体酪氨酸激酶是成药靶标,其通常在tnbc中过表达并已知在肿瘤发生期间起关键作用(casaletto和mcclatchey,2012;speers等人,2009;hsu等人,2014)。为了研究在ros诱导的氧化应激下调节parp1应答的基础分子机制和鉴别潜在的成药搭档,搜寻可以在ros刺激时与parp1结合且在tnbc也被激活的受体酪氨酸激酶。为此,用亚砷酸钠(as)处理mda-mb-231tnbc细胞以诱导ros,然后用抗体阵列(humanphospho-rtkarray)处理细胞裂解产物以鉴别parp1相关激酶。进一步使由as(截留比3.0;表2)诱导且基于从tcga数据库的分层聚类分析(图6a)与tnbc有关的靶标的范围缩窄,并鉴别c-met(hgfr)作为最优候选。作为原癌基因,c-met已显示与tnbc的较差患者存活有关(speers等人,2009;hsu等人,2014;zagouri等人,2013)并且由ros激活(fischer等人,2004)。tnbc中的c-met表达高于非tnbc乳腺癌细胞系中的c-met表达也暗示c-met的重要性(图6b),因此第一次研究c-met是否调节tnbc中的parp1功能。在hek293t细胞(图7a-b)和mda-mb-231细胞(图1i和7c)中分别进一步验证c-met与parp1之间的外源性和内源性结合。此外,在例如hcc1937(内源性,图7d)和mda-mb-436或mcf7(具有c-met的异位表达,图7e-f)等其它细胞系中检测到c-met与parp1之间的相互作用,结果表明其相互作用在氧化应激下通过h2o2和as处理而增强。从duolink分析观察到类似结果(图1j)。此外,细胞分级分离显示h2o2处理以激酶依赖方式增强核中的c-met与parp1之间的结合(图1k)。该研究进一步证明c-met通过类似于需要马达蛋白动力蛋白的egfr核转运(du等人,2014)的机制移位到核中(图7g)。总的来说,这些发现暗示氧化应激诱导c-met的核转运和c-met与parp1的相互作用。dna损伤应答中连接ros与核c-met和parp1的环节提出一个有趣的问题,即c-met是否也调节parp1功能。如果这样的话,c-met也改变针对parp抑制的细胞应答。
表2.parp1相关的受体酪氨酸激酶.
实施例2-c-met调节针对parp抑制剂的抗性
为了确定c-met在对于parp抑制的应答中是否起作用,在三种不同的parp抑制剂奥拉帕尼(azd2281,获得美国食品和药物管理局(fda)批准)、维利帕尼(abt-888)和瑞卡帕布(ag014699)存在下检查细胞生长和集落形成能力,这些parp抑制剂已经在临床试验中评估过。事实上,敲低c-met使mda-mb-231细胞对parp抑制剂更敏感,如降低的细胞活力所指示的(图2a-c和图8a-c)。与敲低c-met一致,c-met被抑制剂克唑替尼和福来替尼抑制也增强细胞对parp抑制剂的敏感性(图2d-e和8d)。此外,当敲低c-met时,锚定非依赖性细胞生长也降低(图2f和8e-h)。与先前发现(jagadeeswaran等人,2007)一致,c-met被shrna或小分子抑制剂抑制都导致ros减少(图8i-j),表明c-met激活和ros的反馈调节可以参与parp1介导的dna损伤应答和parp抑制剂应答。
为了进一步研究parp抑制剂应答期间的c-met功能,野生型c-met和激酶死亡(kd)突变型c-met在内源c-met敲低mda-mb-231细胞中再表达(图2g和8m),且发现只有野生型c-met的再表达增加克隆形成细胞存活(图2h和图8k-l)。异位表达c-met的mcf-7细胞(图2i)在parp抑制下具有增加的细胞活力(图9a-d)、克隆形成能力(图2j和9e-f)和锚定非依赖性细胞生长(图2k和9g-i)。这些结果与观察到的结果一致,其中c-met的下调增加细胞对parp抑制剂的敏感性。值得注意的是,这里对于体外分析使用的剂量与先前研究中使用的剂量是相当的(lehmann等人,2011;anders等人,2013)。总的来说,这些结果表明c-met减弱对于parp抑制剂的应答。
虽然brca突变和缺陷被认为是对于parp抑制剂的应答的主要预测标志物(farmer等人,2005;bryant等人,2005),但是并非所有的具有brca突变的患者都对parp抑制很好地作出应答(lord和ashworth,2013)。举例来说,虽然mda-mb-436细胞和hcc1937细胞都具有brca突变,但是mda-mb-436细胞对parp抑制敏感,而hcc1937细胞有抗性(lehmann等人,2011)(图2l)。基于现有结果,推测在brca突变型tnbc细胞中观察到的parp抑制剂应答的差别归因于c-met的差异表达。事实上,蛋白质印迹分析指示对parp抑制具有更高抗性的hcc1937细胞表达的c-met的含量显著高于mda-mb-436细胞(图2l)。与这个结果一致,敲低c-met使hcc1937细胞对parp抑制更敏感(图2m-n和10a-b),而增加mda-mb-436细胞中野生型c-met而不是激酶死亡突变型c-met的异位表达减弱mda-mb-436细胞对parp抑制的应答(图2o-p和10c-d)。值得注意的是,c-met的敲低或异位表达对brca1/2蛋白含量不具有影响(图10e-f)。为了进一步研究在parp抑制剂应答中都起重要作用的brca1/2与c-met之间的关系,在具有高(mda-mb-231)和低(mda-mb-157)c-met表达的一对野生型brca1和brca2细胞系中敲低brca1和brca2表达(图2q),然后用parp抑制剂处理所述野生型brca1和brca2细胞系。有趣的是,敲低brca1或brca2只使表达低含量的c-met的细胞(图2r-s和10g-j)敏感化。总的来说,这些结果表明增强的c-met激酶表达即使在brca失活下仍然使细胞对parp抑制剂有抗性,且为这个长期谜题提供分子解释。
实施例3-c-met通过在y907上磷酸化parp1介导parp1功能
接下来,确认c-met是否激活parp1功能。为了解决这个问题,将具有或不具有c-met敲低的mda-mb-231细胞暴露于h2o2,并进行彗星分析以评估dna损伤程度。具有c-met敲低的细胞具有比对照细胞更高的尾部强度,这指示增加的氧化dna损伤(图11a)。为了确定dna损伤的增加是否是降低的dna修复的结果,进行dna修复分析,其中用hu/arac(羟基脲/1-fl-d-阿糖呋喃糖胞嘧啶)处理细胞以积累被h2o2损伤的dna,然后在不同时间点后回收,此时测量修复dna的百分率。与对照细胞相比,c-met敲低细胞在dna修复方面的效率较低(图11b)。荧光素酶修复分析的结果还显示,当用被h2o2损伤的荧光素酶质粒转染细胞时,在没有c-met的情况下dna修复功能有所下降(图11c)。与c-met敲低一致,用克唑替尼抑制c-met也增加细胞对parp抑制剂abt-888的敏感性,如γh2ax集落形成(dna损伤的一种标志物)的增强所指示的(图3a)。dna修复也需要作为mcf-7细胞中的野生型c-met而不是激酶死亡c-met的表达的c-met的激酶活性,以降低h2o2诱导的dna损伤,其通过用c-met抑制剂预处理而恢复(图11d-f)。mcf-7细胞中c-met的异位表达也降低abt-888诱导的γh2ax集落形成(dna损伤的一种标志物)。此外,具有c-met敲低的mda-mb-231细胞在abt-888处理下的γh2ax集落形成比具有载体对照的mda-mb-231细胞更高(图3b,左上),所述γh2ax集落形成在野生型c-met的再表达情况下减弱,而在载体对照(图3b,左下)、激酶死亡c-met或野生型c-met加c-met抑制剂、克唑替尼预处理下(图3b,右上)未减弱。这些发现一起表明c-met通过激酶依赖机制增强响应于氧化dna损伤的parp1的dna修复功能。
由于c-met和parp1在体内在物理上结合(图1f-g和7a-f)并且c-met的激酶活性增强响应于dna损伤或parp抑制的dna修复,因此预期作为酪氨酸激酶的c-met也可在氧化应激下使parp1磷酸化。事实上,在flag标记的c-met和v5标记的parp1共转染后,h2o2诱导parp1酪氨酸磷酸化(图3c)。使用软件程序(networkinv2.0)(linding等人,2007),位于parp1的催化区中的h-y-e基序上的tyr907(y907)(ruf等人,1996)被预测为唯一的c-met磷酸化位点。体外激酶分析也显示,与y986f突变体(h-y-e区域中的另一个tyr残基)相比,通过掺入γ-32p引起的parp1磷酸化在y907f突变体中显著降低(图12a-b)。这些结果表明y907是真正的c-met磷酸化位点。
因为y907位于parp1的催化区中,所以接下来确认y907磷酸化是否影响parp1的功能。野生型、y907f(不能磷酸化的)或y907e(磷酸化模拟物)突变型parp1在parp1敲低的mda-mb-231细胞(图3d)中稳定表达,并且通过彗星分析测量h2o2诱导的dna损伤。不出所料,parp1敲低细胞相比对照细胞具有更多的dna损伤(图3e,第2组对比第3组)。野生型parp1(图3e,第4组)而不是y907f突变体(图3e,第5组)的再表达降低内源parp1耗尽细胞中的dna损伤,并且表达y907e突变体(磷酸化模拟物)的细胞具有最少量的dna损伤(图3e,第6组)。为了确定parp1的y907的磷酸化是否影响其活性,比较野生型parp1与突变型parp1的par化(par)水平。如图3f所示,表达野生型parp1的细胞响应于h2o2而具有增加的par。有趣的是,虽然磷酸化模拟物y907e突变体的par含量高于不可磷酸化的y907f突变体,但是这两者响应于h2o2都相对没有变化,表明突变体不再对h2o2敏感。为了进一步研究在y907上磷酸化的parp1的功能重要性,产生特异性抗体以检测磷酸化y907(py907)(图12c-f)。克唑替尼和福来替尼都消除由h2o2诱导的在y907上parp1的磷酸化(图3g)。这些结果表明h2o2诱导的野生型parp1活性需要y907磷酸化。
因为c-met削弱parp抑制的作用(图2),所以确认是否由c-met介导的parp1的y907的磷酸化影响parp抑制剂应答。表达经h2o2或未经h2o2处理的野生型parp1的细胞或表达y907f或y907eparp1突变体的细胞在增加的abt-888浓度下经parp酶活性分析以测量ic50。磷酸化模拟物y907e突变体的活性类似于经h2o2处理的野生型parp1的活性(较高ic50),而不可磷酸化的y907f突变体的活性类似于没有h2o2的野生型parp1(较低ic50)(图13a-b)。这些结果指示parp1的y907的磷酸化削弱abt-888的抑制作用。
接下来,为了确定parp1y907磷酸化在功能上是否受c-met介导,在再表达野生型parp1和y907突变型parp1的parp1敲低细胞中敲低c-met(图13c)并用abt-888或不用abt-888处理这些细胞。通过γh2ax集落形成分析评估这些细胞中的dna损伤程度。如图3i所示,只有野生型parp1对c-met敲低作出应答,如增加的γh2ax集落所指示(图3i,插图1对比插图4)。然而,敲低c-met对表达y907f或y907e突变体的细胞不具有明显影响。在对照细胞和c-met敲低细胞中,响应于abt-888,不可磷酸化的y907f突变体(图3i,插图2对比插图5)具有高于磷酸化模拟物y907e突变体(图3i,插图3对比插图6)的γh2ax集落形成。此外,克隆形成细胞存活分析(图3h)和细胞活力分析(图13d-e)也显示一致的结果,其中c-met敲低使表达野生型parp1的细胞对parp抑制剂abt-888和ag014699更加敏感。克隆形成细胞存活和细胞活力在c-met存在下在表达野生型parp1的细胞和表达y907e的细胞之间(左边,图3h和13d-e)以及在不存在c-met的情况下在表达野生型parp1的细胞和表达y907f的细胞之间(右边,图3h和13d-e)是相似的。这些发现一起提供强有力的证据支持parp1y907的磷酸化以c-met依赖方式削弱parp抑制剂应答和dna损伤。
实施例4-在tnbc中靶向parp1和c-met的临床相关性和潜在治疗策略
在临床患者样品中首先表征针对py907的特异性抗体(图14a)。接下来,通过免疫组织化学(ihc)染色法测量的在乳腺癌组织微阵列中的py907-parp1表达证实tnbc和非tnbc中py907-parp1与c-met表达之间的正相关(图4a和14b)。同样,高ros(8-ohdg)也和高py907-parp1表达有关(图14c)。这些结果表明细胞内ros可以是c-met依赖方式的parp1y907磷酸化的关键诱导物,其削弱parp抑制剂应答。此外,这种机制提供潜在临床应用,其中c-met抑制可以增强对于parp抑制的敏感性。为了确定其可行性,首先检查c-met抑制剂(福来替尼和克唑替尼)和parp抑制剂(abt-888和ag-014699)的组合功效。abt-888/福来替尼和ag-014699/克唑替尼组合这两者在mda-mb-231和hcc1937tnbc细胞中都显示协同细胞生长抑制(图4b),但在mcf10a乳腺上皮细胞中不显示协同细胞生长抑制(图15a)。ag-014699/克唑替尼的组合处理还抑制克隆形成(图4c和4e)和锚定非依赖性生长(图4g和15b)。对于abt-888/福来替尼组合观察到对克隆形成细胞存活的类似抑制作用(图15c)。在另一个乳腺癌细胞系bt549中也观察到c-met和parp1的合成抑制(图15d)。c-met抑制消除h2o2诱导的y907-parp1磷酸化(图15e)。除了人乳腺癌细胞系之外,在来源于组成性地表达活性人c-met的tnbc转基因小鼠模型(knight等人,2013)的两种小鼠乳腺肿瘤细胞系中评估组合治疗的效果。用c-met和parp抑制剂组合治疗协同性地抑制小鼠肿瘤细胞生长(图4h和15f)。此外,在这些小鼠细胞系中,py907-parp1被h2o2刺激并被c-met抑制所消除(图4i和15g)。
接下来,在体内针对已建立的tnbc异种移植模型评估parp和c-met抑制剂的组合。在mda-mb-231异种移植肿瘤模型中,组合治疗(ag014699/克唑替尼和abt-888/福来替尼)导致显著降低的肿瘤生长(图4j-k)。在mda-mb-231异种移植肿瘤模型中,在使用转基因小鼠细胞系a1034和另一种tnbc细胞系hcc1937的协同模型中也观察到类似结果。ag014699和克唑替尼的组合显著抑制肿瘤生长(图16a-b)。在从最后一次治疗后24小时内的小鼠收获的mda-mb-231异种移植肿瘤组织中观察到增加的细胞凋亡(tunel染色,图4l)、降低的细胞增殖(ki67染色,图4m)和更多的dna损伤(γh2ax染色,图4n)。果然,parp1y907磷酸化也被克唑替尼抑制(图4o)。此外,临床化学分析(图17a、b、d、e、g、h)以及小鼠体重(图17c、f、i)表明动物的总体健康没有受到ag-014699/克唑替尼和abt-888/福来替尼的组合治疗的不利影响。这些结果表明这个组合在体外和体内对tnbc都是有效的。
为了将这些发现拓展到其它癌症类型,在具有c-met低表达的非tnbc细胞系mcf-7中异位表达c-met。mcf-7/c-met显示和tnbc细胞系类似的应答。在图4p和14a中,只有具有c-met异位表达的mcf-7响应于c-met和parp抑制剂的组合治疗才具有协同效应,而具有载体对照或激酶死亡突变体的mcf-7细胞不具有协同效应。更多的机制研究揭示,在具有c-met表达的mcf-7细胞中h2o2增加了通过c-met抑制剂预处理而消除的parp1的y907的磷酸化(图4q)。此外,在两种非小细胞肺癌细胞系(具有c-met高表达的h1993和具有c-met低表达的a549)中观察到类似结果(图18b),其中只在具有c-met高表达的h1993中观察到c-met抑制剂和parp抑制剂组合的协同效应。更高的c-met导致更高的可被c-met抑制阻断的响应于h2o2处理的parp1y907磷酸化(图18c)。ag014699和克唑替尼的组合治疗抑制h1993肺癌和mcf/c-met乳腺癌肿瘤模型中的肿瘤生长(图4r-s)。总的来说,c-met介导的parp1磷酸化控制具有c-met表达的tnbc和其它癌症类型中的parp抑制剂应答。这些结果表明py907-parp1可以用作生物标志以在临床上针对单一parp抑制剂或c-met抑制剂和parp抑制剂的组合治疗对患者分层。
***
本文中公开和要求的所有方法都可以根据本公开进行和实施而不需要过度实验。虽然已经参考优选实施方案描述了本发明的组合物和方法,但是对于所属领域技术人员显而易见的是,可以对本文中所述的方法和步骤和方法的步骤的顺序应用变化而不会脱离本发明的概念、精神和范围。更具体地说,显而易见的是,在化学和生理学这两者上都相关的特定药剂可以替代本文所述的药剂,同时仍将实现相同或类似结果。对于所属领域技术人员显而易见的所有这些类似替代和修改都被认为处于随附权利要求书所限定的本发明的精神、范围和概念内。
参考文献
以下参考文献在其提供示例性程序细节或本文所阐述内容的其它补充细节的程度上以引用方式被具体地并入本文中。
美国专利号7,928,105
美国专利号8,124,606
美国专利号8,236,802
美国专利号8,450,323
wo2006/110816
wo2007/084532
wo2008/064157
wo2008/083027
wo2011/014681
anders等人,聚(adp-核糖)聚合酶抑制:三阴性乳腺癌的“靶向”疗法.(poly(adp-ribose)polymeraseinhibition:“targeted”therapyfortriple-negativebreastcancer.)clinicalcancerresearch,16:4702-4710,2010.
anders等人,聚乙二醇化脂质体阿霉素在乳腺癌的颅内模型中的药代动力学和功效.(pharmacokineticsandefficacyofpegylatedliposomaldoxorubicininanintracranialmodelofbreastcancer.)plosone,8:e61359,2013.
birchmeier等人,met、转移、运动性等等.(met,metastasis,motilityandmore.)nat.rev.mol.cell.biol.,4:915-925,2003.
boccaccio和comoglio,侵袭性生长:癌症和干细胞的met驱动型遗传程序.(invasivegrowth:amet-drivengeneticprogrammeforcancerandstemcells.)nat.rev.cancer,6:637-645,2006.
bryant等人,用聚(adp-核糖)聚合酶的抑制剂特异性杀死brca2缺陷肿瘤.(specifickillingofbrca2-deficienttumourswithinhibitorsofpoly(adp-ribose)polymerase.)nature,434:913-917,2005.
carey等人,三阴性乳腺癌:疾病实体或方便标题?(triple-negativebreastcancer:diseaseentityortitleofconvenience?)naturereviews.clinicaloncology,7:683-692,2010.
casaletto和mcclatchey,发育和癌症中的受体酪氨酸激酶的空间调控.(spatialregulationofreceptortyrosinekinasesindevelopmentandcancer.)naturereviewscancer,12:387-400,2012.
castaldi等人,康复设施的使用有多适当?通过基于aep协定的评估工具评定.(howappropriateistheuseofrehabilitationfacilities?assessmentbyanevaluationtoolbasedontheaepprotocol.)j.prev.med.hyg.,51:116-120,2010.
chou,使用chou-talalay方法的药物组合研究和其协同量化.(drugcombinationstudiesandtheirsynergyquantificationusingthechou-talalaymethod.)cancerres.,70:440-446,2010.
christensen等人,作为人癌症标靶的c-met和治疗干预的抑制剂的表征.(c-metasatargetforhumancancerandcharacterizationofinhibitorsfortherapeuticintervention.)cancerlett.,225:1-26,2005.
donawho等人,abt-888,一种口服活性聚(adp-核糖)聚合酶抑制剂,其增强dna破坏剂在临床前肿瘤模型中的功效.(abt-888,anorallyactivepoly(adp-ribose)polymeraseinhibitorthatpotentiatesdna-damagingagentsinpreclinicaltumormodels.)clin.cancerres.,13:2728-2737,2007.
du等人,突触融合蛋白6介导的高尔基体易位在通过依赖于微管的运输的egfr的核功能中起重要作用.(syntaxin6-mediatedgolgitranslocationplaysanimportantroleinnuclearfunctionsofegfrthroughmicrotubule-dependenttrafficking.)oncogene,33:756-770,2014.
eisen等人,全基因组表达模式的聚类分析和显示.(clusteranalysisanddisplayofgenome-wideexpressionpatterns.)proc.natl.acad.sci.u.s.a.,95:14863-14868,1998.
farmer等人,靶向brca突变细胞中的dna修复缺陷以作为治疗策略.(targetingthednarepairdefectinbrcamutantcellsasatherapeuticstrategy.)nature,434:917-921,2005.
fischer等人,活性氧物质介导人癌细胞中的g蛋白偶合受体和表皮生长因子受体对met受体的反式激活.(reactiveoxygenspeciesmediatemetreceptortransactivationbygprotein-coupledreceptorsandtheepidermalgrowthfactorreceptorinhumancarcinomacells.)j.biol.chem.,279:28970-28978,2004.
foulkes等人,三阴性乳腺癌.(triple-negativebreastcancer.)thenewenglandjournalofmedicine,363:1938-1948,2010.
gastaldi等人,met致癌基因和基底样乳腺癌:要当心的另一个罪犯?(themetoncogeneandbasal-likebreastcancer:anotherculprittowatchoutfor?)breastcancerresearch,12:208,2010.
gelmon等人,复发型高级别严重或低分化卵巢癌或三阴性乳腺癌的患者中的奥拉帕尼:2期,多中心,开放标签,非随机研究.(olaparibinpatientswithrecurrenthigh-gradeserousorpoorlydifferentiatedovariancarcinomaortriple-negativebreastcancer:aphase2,multicentre,open-label,non-randomisedstudy.)lancetoncol.,12:852-861,2011.
gibson和kraus,对于聚(adp-核糖)和parp的分子和细胞功能的新的领悟.(newinsightsintothemolecularandcellularfunctionsofpoly(adp-ribose)andparps.)nat.rev.mol.cellbiol.,13:411-424,2012.
hampson等人,用于确定临床试验受试对象中的利妥昔单抗药代动力学的elisa的验证.(validationofanelisaforthedeterminationofrituximabpharmacokineticsinclinicaltrialssubjects.)j.immunol.methods,360:30-38,2010.
hsu等人,作为三阴性乳腺癌的治疗标靶的pkc-α、cdk6和met的定义.(definitionofpkc-alpha,cdk6,andmetastherapeutictargetsintriple-negativebreastcancer.)cancerres.,74:4822-4835,2014.
irani等人,由ras转化型成纤维细胞中的氧化剂介导的有丝分裂信号传导.(mitogenicsignalingmediatedbyoxidantsinras-transformedfibroblasts.)science,275:1649-1652,1997.
jagadeeswaran等人,hgf/c-met路径的激活造成小细胞肺癌细胞的活性氧物质产生和运动性.(activationofhgf/c-metpathwaycontributestothereactiveoxygenspeciesgenerationandmotilityofsmallcelllungcancercells.)americanjournalofphysiology.lungcellularandmolecularphysiology,292:l1488-1494,2007.
knight等人,met与p53损失协同作用以诱导具有低claudin乳腺癌的特征的乳腺肿瘤.(metsynergizeswithp53losstoinducemammarytumorsthatpossessfeaturesofclaudin-lowbreastcancer.)proc.natl.acad.sci.u.s.a.,110:e1301-1310,2013.
kummar等人,具有晚期恶性肿瘤的患者中的聚(adp-核糖)聚合酶抑制剂abt-888的0期临床试验.(phase0clinicaltrialofthepoly(adp-ribose)polymeraseinhibitorabt-888inpatientswithadvancedmalignancies.)j.clin.oncol.,27:2705-2711,2009.
ledermann等人,铂敏感复发性严重卵巢癌患者的奥拉帕尼维持疗法:随机2期试验中的brca状态的结果的预先计划回顾性分析.(olaparibmaintenancetherapyinpatientswithplatinum-sensitiverelapsedserousovariancancer:apreplannedretrospectiveanalysisofoutcomesbybrcastatusinarandomisedphase2trial.)thelancetoncology,15:852-861,2014.
lehmann等人,人三阴性乳腺癌亚型的鉴别和用于筛选靶向疗法的临床前模型.(identificationofhumantriple-negativebreastcancersubtypesandpreclinicalmodelsforselectionoftargetedtherapies.)j.clin.invest.,121:2750-2767,2011.
liedtke等人,三阴性乳腺癌患者对于新辅助疗法的应答和长期生存率.(responsetoneoadjuvanttherapyandlong-termsurvivalinpatientswithtriple-negativebreastcancer.)journalofclinicaloncology,26:1275-1281,2008.
lindahl,dna的一级结构的不稳定性和衰变.(instabilityanddecayoftheprimarystructureofdna.)nature,362:709-715,1993.
linding等人,体内磷酸化网络的系统发现.(systematicdiscoveryofinvivophosphorylationnetworks.)cell,129:1415-1426,2007.
liu等人,egfr表达与三阴性乳腺癌的降低的无疾病生存率产生关联:基于组织微阵列的回顾性分析.(egfrexpressioncorrelateswithdecreaseddisease-freesurvivalintriple-negativebreastcancer:aretrospectiveanalysisbasedonatissuemicroarray.)medicaloncology,29:401-405,2012.
lord和ashworth,对于靶向brca突变癌症的疗法的抗性的机制.(mechanismsofresistancetotherapiestargetingbrca-mutantcancers.)nat.med.,19:1381-1388,2013.
luo和kraus,关于par和parp:通过聚(adp-核糖)和parp-1的细胞应激信号传导.(onparwithparp:cellularstresssignalingthroughpoly(adp-ribose)andparp-1.)genesdev.,26:417-432,2012.
nowsheen等人,人三阴性乳腺癌细胞中的egfr与parp抑制之间的合成致死相互作用.(syntheticlethalinteractionsbetweenegfrandparpinhibitioninhumantriplenegativebreastcancercells.)plosone,7:e46614,2012.
o'shaughnessy等人,转移性三阴性乳腺癌中的依尼帕尼加化学疗法.(iniparibpluschemotherapyinmetastatictriple-negativebreastcancer.)thenewenglandjournalofmedicine,364:205-214,2011.
o'shaughnessy等人,转移性三阴性乳腺癌(tnbc)中的依尼帕尼(bsi-201)与吉西他滨/卡铂(g/c)的组合的随机iii期研究.(arandomizedphaseiiistudyofiniparib(bsi-201)incombinationwithgemcitabine/carboplatin(g/c)inmetastatictriple-negativebreastcancer(tnbc).)journalofclinicaloncology,29:abstr1007,2011.
radisky等人,rac1b和活性氧物质介导mmp-3诱导的emt和基因组不稳定性.(rac1bandreactiveoxygenspeciesmediatemmp-3-inducedemtandgenomicinstability.)nature,436:123-127,2005.
rouleau等人,parp抑制:parp1等.(parpinhibition:parp1andbeyond.)naturereviewscancer,10:293-301,2010.
ruf等人,鸡的聚(ad-核糖)聚合酶的催化片段的结构.(structureofthecatalyticfragmentofpoly(ad-ribose)polymerasefromchicken.)proc.natl.acad.sci.u.s.a.,93:7481-7485,1996.
sokal等人,用于评估系统关系的统计方法(astatisticalmethodforevaluatingsystematicrelationships),universityofkansas,1958.
speers等人,鉴别用于治疗雌激素受体阴性乳腺癌的新颖激酶标靶.(identificationofnovelkinasetargetsforthetreatmentofestrogenreceptor-negativebreastcancer.)clin.cancerres.,15:6327-6340,2009.
trachootham等人,通过ros介导的机制靶向癌细胞:放射治疗方法?(targetingcancercellsbyros-mediatedmechanisms:aradicaltherapeuticapproach?)nat.rev.drugdiscov.,8:579-591,2009.
turashvili等人,在3992例乳腺癌组织微阵列系列中作为预后生物标志的p-钙粘附蛋白表达.(p-cadherinexpressionasaprognosticbiomarkerina3992casetissuemicroarrayseriesofbreastcancer.)(p-cadherinexpressionasaprognosticbiomarkerina3992casetissuemicroarrayseriesofbreastcancer.)mod.pathol.,24:64-81,2011.
turner等人,散发性癌中的'brcaness'的标志.(hallmarksof'brcaness'insporadiccancers.)naturereviewscancer,4:814-819,2004.
tutt等人,具有brca1或brca2突变和晚期乳腺癌的患者中的口服聚(adp-核糖)聚合酶抑制剂奥拉帕尼:概念验证试验.(oralpoly(adp-ribose)polymeraseinhibitorolaparibinpatientswithbrca1orbrca2mutationsandadvancedbreastcancer:aproof-of-concepttrial.)lancet,376:235-244,2010.
zagouri等人,高met表达是三阴性乳腺癌患者的不利预后因子.(highmetexpressionisanadverseprognosticfactorinpatientswithtriple-negativebreastcancer.)br.j.cancer,108:1100-1105,2013.
序列表
<110>得克萨斯大学体系董事会
<120>针对parp抑制剂的应答的预测和靶向c-met和parp1的组合疗法
<130>utfc.p1248wo
<140>pct/us2015/052966
<141>2015-09-29
<150>us62/057,037
<151>2014-09-29
<160>13
<170>patentinversion3.5
<210>1
<211>17
<212>prt
<213>人工序列
<220>
<223>合成多肽
<400>1
alaaspmetvalserlysseralaasntyrcyshisthrserglngly
151015
asp
<210>2
<211>19
<212>dna
<213>人工序列
<220>
<223>合成多聚核苷酸
<400>2
ccatccagaatgtcattct19
<210>3
<211>18
<212>dna
<213>人工序列
<220>
<223>合成多聚核苷酸
<400>3
gcattaaagcagcgtatc18
<210>4
<211>18
<212>dna
<213>人工序列
<220>
<223>合成多聚核苷酸
<400>4
gcattaaagcagcgtatc18
<210>5
<211>19
<212>dna
<213>人工序列
<220>
<223>合成多聚核苷酸
<400>5
tgtgttgtatggtcaataa19
<210>6
<211>21
<212>dna
<213>人工序列
<220>
<223>合成多聚核苷酸
<400>6
ccttcagaaggttgctgagta21
<210>7
<211>21
<212>dna
<213>人工序列
<220>
<223>合成多聚核苷酸
<400>7
tggaaagatgttaagcattta21
<210>8
<211>19
<212>dna
<213>人工序列
<220>
<223>合成多聚核苷酸
<400>8
ttcatttctaatacctgcc19
<210>9
<211>19
<212>dna
<213>人工序列
<220>
<223>合成多聚核苷酸
<400>9
ttaagtcacataatcgatc19
<210>10
<211>19
<212>dna
<213>人工序列
<220>
<223>合成多聚核苷酸
<400>10
ttcagtacaattaggtggg19
<210>11
<211>19
<212>dna
<213>人工序列
<220>
<223>合成多聚核苷酸
<400>11
ttgttcagcagattccatg19
<210>12
<211>19
<212>dna
<213>人工序列
<220>
<223>合成多聚核苷酸
<400>12
tctttaagacagctaagag19
<210>13
<211>19
<212>dna
<213>人工序列
<220>
<223>合成多聚核苷酸
<400>13
tattaaatgactctttggc19
- 还没有人留言评论。精彩留言会获得点赞!
- 一种靶向锚定剂及其制备方法和应用与流程
- 一种超低分子量插层抑制剂及其制备方法与流程
- 具有胶原特异结合能力的神经再生抑制分子的拮抗蛋白CBD-PlexinB1LBD及应用的制作方法与工艺
- 具有胶原特异结合能力的神经再生抑制分子的拮抗蛋白CBD-EphA4LBD及应用的制作方法与工艺
- 一种KDM1A小分子抑制剂在抑制肿瘤细胞生长和转移中的应用的制作方法与工艺
- 一种抑制多药耐受基因表达的靶向抗肿瘤纳米药物的制作方法与工艺
- MALT1靶向抑制物在制备MALT1依赖性肿瘤治疗药物中的应用的制作方法与工艺
- 作为ASGPR靶向剂的被取代的‑6,8‑二氧杂双环[3.2.1]辛烷‑2,3‑二醇化合物的制作方法与工艺
- 小分子C‑Myc抑制剂的制作方法与工艺
- 一种RNA抑制剂及其在肿瘤中的应用的制作方法与工艺
- 靶向PD-1的多肽及其应用的制造方法与工艺
- 靶向同位素生产系统的制造方法与工艺
- 在制备68GA‑螯合物官能化的靶向剂中用作金属抑制剂的单糖、二糖或多糖的制造方法与工艺
- 用于引导和靶向按需物质递送的纳米结构的载体的制造方法与工艺
- 通过PROSTRATIN靶向K‑RAS介导的信号转导通路和恶性肿瘤的制造方法与工艺
- 肿瘤靶向核磁共振/荧光双模态成像造影剂及其制备和应用的制造方法与工艺
- pH氧化还原双敏感的PAMAM靶向纳米给药载体及其制备方法与制造工艺
- 具有卵巢肿瘤靶向D构型多肽及其基因递释系统的制造方法与工艺
- 靶向survivin纳米抗体及其制备方法和应用与制造工艺
- 用于PSMA靶向的成像和放射疗法的金属/放射性金属标记的PSMA抑制物的制造方法与工艺
- 阻断UBE2M介导Neddylation修饰在肾细胞癌治疗中的应用的制造方法与工艺
- 无载体共组装肿瘤靶向抗癌纳米药物及其制备方法与应用与流程
- 一种用于判断靶向单克隆抗体治疗肿瘤疗效的预测分子的制造方法与工艺
- 一种靶向CD47的免疫检查点抑制剂药物组合物及其制备方法与流程
- 具有淋巴靶向功能的生物自组装纳米晶注射剂及制备方法与流程
- 用于检测肝癌的基因标志物及其用途的制造方法与工艺
- 特异性shRNA筛选及其靶向Ang‑2基因抑制肺癌细胞的验证方法与流程
- 一种多功能自动化生物细胞提取分离控制系统及采用该系统实现的细胞提取分离方法与流程
- 一种融合蛋白及光敏剂复合物及其制备方法和应用与流程
- 具有可视化和显示光斑边界功能的光动力治疗系统的制造方法与工艺
- 肿瘤靶向核磁共振/荧光双模态成像造影剂及其制备和应用的制造方法与工艺
- pH氧化还原双敏感的PAMAM靶向纳米给药载体及其制备方法与制造工艺
- 具有卵巢肿瘤靶向D构型多肽及其基因递释系统的制造方法与工艺
- 靶向survivin纳米抗体及其制备方法和应用与制造工艺
- 用于PSMA靶向的成像和放射疗法的金属/放射性金属标记的PSMA抑制物的制造方法与工艺
- 一种线粒体靶向的粘度荧光探针及其制备方法和应用与制造工艺
- 一种兼具荧光可视、磁靶向和pH敏多功能的复合空心微球药物载体及其制备方法与制造工艺
- 一种用于修饰微泡的多肽以及靶向GBM的药物制剂的制造方法与工艺
- 白果内酯作为脑靶向增效剂在制备防治脑肿瘤药物的应用的制造方法与工艺
- 机器人辅助精确靶向穿刺软组织目标柔性针夹紧固定装置的制造方法
- 一种用于判断靶向单克隆抗体治疗肿瘤疗效的预测分子的制造方法与工艺
- 一种检测肺癌相关基因热点突变的试剂盒及其应用的制造方法与工艺
- 一种为肺癌提供离体个体化药物测试的方法及培养基与流程
- 一种谷胱甘肽/超声双重刺激响应纳米聚合物囊泡、其制备方法及应用与流程
- 一种多重刺激响应交联型聚合物纳米水凝胶、其制备方法及应用与流程
- 一种靶向CD47的免疫检查点抑制剂药物组合物及其制备方法与流程
- 具有淋巴靶向功能的生物自组装纳米晶注射剂及制备方法与流程
- 用于检测肝癌的基因标志物及其用途的制造方法与工艺
- 特异性shRNA筛选及其靶向Ang‑2基因抑制肺癌细胞的验证方法与流程
- 一种具有靶向功能仿病毒结构高分子囊泡及其制备与应用的制造方法与工艺