能够与膜联蛋白‑1结合的特异性结合分子的用途的制作方法
本申请为申请日为2009年12月02日、申请号为200980153335.4、名称为“通过调控膜联蛋白-1对自身免疫疾病的治疗”的中国发明专利申请的分案申请。
本发明属于免疫疾病治疗领域,涉及通过调控膜联蛋白-1(annexin-1)的活性治疗t细胞介导的疾病的方法。
背景技术:
自身免疫疾病是免疫系统功能失常引起的慢性致残病变。在大部分情况中,它们是通过t细胞对存在于抗原呈递细胞(apcs)的mhc分子环境中的自身抗原失控的应答所启动的。已经公开了几种参与自身免疫疾病发病机制的因素,包括环境,遗传和病毒因素,具有一个总体特征:t细胞的过度反应性(hyperresponsivity)。
糖皮质激素(gcs)常用于治疗多种慢性自身免疫疾病,因为它们具有同时阻断固有和适应性免疫应答的能力。近10年的研究,或本发明人和其他研究团队所进行的研究都表明,gc对固有免疫反应的炎症效应是由被称为膜联蛋白-1(anx-a1)的蛋白所介导的。已证实这种蛋白在大量细胞类型中发挥自稳调节,包括中性白细胞(neutrophils),巨噬细胞和内皮细胞。然而,经常被忽视的一方面是anx-a1在适应性免疫反应中的作用。考虑到anx-a1已被提出作为一种gc药理效应的第二信使,令人激动不已。
本发明人之前已指出了anx-a1通过调控t细胞受体(tcr)信号的强度,在t细胞中发挥自稳作用(d'acquistoetal.,blood109:1095-1102,2007)。
而且,本发明人已指出高水平的anx-a1会降低t细胞激活的阈值并偏好分化成th1细胞,而anx-a1缺陷型小鼠显示出受损的t细胞激活和增强的向th2细胞的分化(d'acquistoetal.,eur.j.immunol.37:3131-3142,2007)。
wo2005/027965描述了所发现的通过凋亡中性白细胞递送抗炎症信号至树突细胞的机制,并鉴别了干扰此过程的抗体。特别是,wo2005/027965描述了鉴别作为信号分子的anx-1,据称其被凋亡中性白细胞表达,以抑制树突细胞的激活和成熟。wo2005/027965提出了称为dac5(凋亡细胞检测器nr.5,detectorofapoptoticcell)的抗体通过树突细胞的吞噬作用来识别并阻断呈递于凋亡中性白细胞表面上的anx-1的抗炎症作用。而wo2005/027965指出了通过靶向这种凋亡细胞并通过引发炎症应答将其清除来治疗多种疾病的可能性,但并未论述anx-1在t细胞激活中的作用。
wo2005/027965声称膜联蛋白在正经历凋亡的细胞上表达(参见例如第8页,第6-7和29-30行),以及这种膜联蛋白呈递于这些细胞的表面上(参见例如第6页,第10-11行和第8页,第16-17行)。然而,两个独立的研究(madernaetal.,jimmunol.,174:3727-3733,2005;scannelletal.,jimmunol.,178:4595-4605,2007)显示了凋亡细胞包括中性白细胞会释放膜联蛋白-1,而不会表达蛋白并将其呈递在细胞表面。因为膜联蛋白-1是从细胞释放的,所以无法主张dac5仅仅鉴别在其表面表达蛋白的凋亡细胞,因为抗体也会鉴别释放的膜联蛋白-1。
而且,wo2005/027965主张了将在其细胞膜上表达膜联蛋白-1的凋亡中性白细胞与用lps激活的树突细胞共孵育,导致了tnf-α分泌的抑制和激活标记物cd83,cd86和hla-dr的上调,以及在该培养物中添加dac5转变了表达膜联蛋白-1的凋亡中性白细胞的抑制作用(第5页,第31行至第6页第8行)。来自本发明人的数据(hugginsetal.,fasebj.2008,印刷中)证实了用lps刺激会使树突细胞释放anx-1,因而wo2005/027965所述的dac5会与外化在中性白细胞上的anx-1以及树突细胞释放的膜联蛋白-1结合。而且,本发明人发现了在树突细胞中缺乏膜联蛋白-1会造成成熟/激活标记物的增强表达和炎症细胞因子,例如tnf-α和il-lβ和il-12的产生。因此,wo2005/027965所述的抗体dac5应该会影响树突细胞的成熟和激活,和之后的免疫应答的调控。
为了支持这些,本发明人显示了将anx-a1树突细胞和天然的t细胞在混合淋巴细胞反应(mlr)中共培养,表现出诱导t细胞增殖或il-2和ifn-γ产生的能力明显降低。因而,阻断anx-a1在树突细胞中功能的试剂应该会降低它们刺激强有力的t细胞介导免疫应答的能力。因此,wo2005/027965中涉及的抗体不适合用于治疗在该专利申请中所涉及的疾病。
技术实现要素:
本发明提供了能够与膜联蛋白-1(anx-a1)结合的特异性结合分子在t-细胞介导的疾病的治疗中的用途。
根据本发明的第一方面,提供了能够与膜联蛋白-1(anx-a1)结合的特异性结合分子,其可用于t-细胞介导的疾病的治疗。
本发明人之前阐明了anx-a1调控t细胞受体(tcr)信号的强度,而且高水平的anx-a1降低t细胞激活的阈值并偏好分化成th1细胞。本发明人目前鉴别了膜联蛋白途径,和保障信号,作为治疗t-细胞介导的疾病的封闭靶点。这些疾病包括那些异常的t细胞激活,例如多种自身免疫疾病,以及那些期望的t细胞偏好分化成th1而不是th2细胞的疾病。
本发明利用了能够与膜联蛋白-1(anx-a1)结合的特异性结合分子。
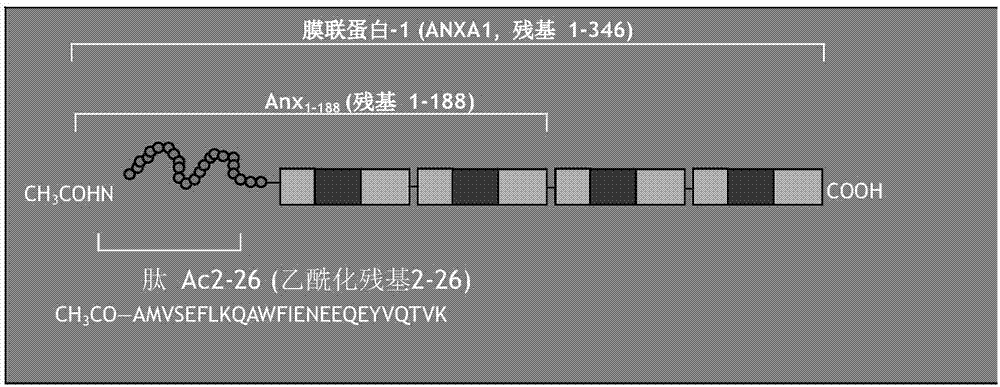
膜联蛋白是一组钙和磷脂结合细胞蛋白,也称为脂皮质蛋白。膜联蛋白家族有13个成员,包括膜联蛋白a1,膜联蛋白a2和膜联蛋白a5。膜联蛋白a1也称为膜联蛋白-1,并在本文中表示为“anx-a1”。膜联蛋白-1(anx-a1)是37-kda蛋白,最初被描述为糖皮质激素活动的介导子。最近几年的证据显示了anx-a1通过调控t细胞受体(tcr)信号的强度在适应性免疫系统特别是t细胞中发挥自稳作用。anx-a1在体内固有免疫系统细胞中充当炎症的内源下调因子。图1a是显示anx-a1三维结构的连续剖面图。
共有八个编码anx-a1的人核苷酸序列。其中,仅有四个被翻译因而有四个anx-a1同工型(isoform),被指定为anxal-002,anxa1-003,anxa1-004和anxa1-006。这些序列可从ensembl网站(www.ensembl.org)获得,被指定为otthumt00000052664(anxal-002),otthumt00000052665(anxal-003),otthumt00000052666(anxa1-004)和otthumt00000052668(anxal-006)。图2a显示了人膜联蛋白-1(anx-a1)的一个同工型anxa1-003的氨基酸((i),seqidno:2)和核苷酸序列((ii),seqidno:3)。而图2b,2c和2d分别显示了同工型anxa1-002,anxa1-004和anxa1-006的氨基酸序列(seqidno:4-6)。如图2所观察到的,同工型anxa1-002,anxa1-004和anxa1-006是anxa1-003的短剪接变体(variant),或是具有少量氨基酸变化的anxa1-003变体。
大量研究表明称为ac.2-26的anx-a1的n末端肽充当了整个蛋白的生物活性替代物(参见例如limetal.,procnatlacadsciusa95,14535-9,1998)。
图1b是膜联蛋白重复序列和该生物活性序列的定位的示意图。肽ac.2-26是具有图2所示anx-a1全长氨基酸序列的氨基酸残基2-26序列的乙酰化肽。肽ac.2-26的序列如图1c(或seqidno:1)所示,如下:
ch3co-amvseflkqawfieneeqeyvqtvk
anx-al及其n末端衍生的生物活性肽通过甲酰肽受体(fpr)家族的成员介导它们的生物作用。anx-al通过直接结合并激活该族的成员之一,甲酰肽样受体-1(formylpeptidereceptorlike-1)(fprl-1),从而发挥其对中性白细胞外渗和固有免疫的反调节(couterregulatory)作用。本发明人之前已经发现了在hranx-a1的存在下刺激t细胞会经由刺激fprl-i增加t细胞激活(d'acquistoetal.,blood109:1095-1102,2007).
本发明利用了能够与膜联蛋白-1(anx-a1)结合的特异性结合分子。特异性结合分子所结合的anx-a1通常是具有图2a所示多肽序列的人anx-a1,或其变体或片段,例如,具有图2b,图2c或图2d所示的多肽序列的人anx-a1的同工型之一。特异性结合分子所结合的人anx-a1片段通常是具有图1c所示序列的多肽。特异性结合分子所结合的anx-a1通常是由图2a所示核苷酸序列编码的。
如本文所用的,术语“变体”是指具有相似氨基酸序列和/或保持相同功能的蛋白。例如,术语“变体”包括含有一个或多个氨基酸添加,删除,置换等等的蛋白或多肽。氨基酸置换通常是保守置换,即用另一个具有大体相似特性的氨基酸置换一个氨基酸,以便整体功能不可能受到严重影响。
因而氨基酸甘氨酸,丙氨酸,缬氨酸,亮氨酸和异亮氨酸通常可以相互置换(具有脂肪族侧链的氨基酸)。在这些可能的置换中,优选地,甘氨酸和丙氨酸可用于相互置换(因为它们具有相对短的侧链),而缬氨酸,亮氨酸和异亮氨酸可用于相互置换(因为它们具有更大的疏水性脂肪族侧链)。通常可用于相互置换的其他氨基酸包括:苯丙氨酸,酪氨酸和色氨酸(具有芳族侧链的氨基酸);赖氨酸,精氨酸和组氨酸(具有碱性侧链的氨基酸);天冬氨酸和谷氨酸(具有酸性侧链的氨基酸);天冬酰胺和谷氨酰胺(具有酰胺基侧链的氨基酸);和半胱氨酸和甲硫氨酸(具有含硫侧链的氨基酸)。
使用三个字母和一个字母代码,氨基酸可做如下表示:甘氨酸(g或gly),丙氨酸(a或ala),缬氨酸(v或val),亮氨酸(l或leu),异亮氨酸(i或ile),脯氨酸(p或pro),苯丙氨酸(f或phe),酪氨酸(y或tyr),色氨酸(w或trp),赖氨酸(k或lys),精氨酸(r或arg),组氨酸(h或his),天冬氨酸(d或asp),谷氨酸(e或glu),天冬酰胺(n或asn),谷氨酰胺(q或gln),半胱氨酸(c或cys),甲硫氨酸(m或met),丝氨酸(s或ser)和苏氨酸(t或thr)。若残基可以是天冬氨酸或天冬酰胺,则可以使用符号asx或b。若残基可以是谷氨酸或谷氨酰胺,则可以使用符号glx或z。提及天冬氨酸包括天冬氨酸盐,提及谷氨酸包括谷氨酸盐,除非文中另有特殊规定。
也可以针对上述涉及的蛋白的氨基酸序列进行氨基酸删除或插入。因而,例如,对多肽活性不具有实质影响,或至少不会消除这种活性的氨基酸可以被删除。这种删除可能是有利的,因为能够在降低多肽整体长度和分子量的同时仍保持其活性。这可以实现降低特定目的所需要的多肽量,例如,能够降低剂量水平。
也可以针对上述融合蛋白的序列进行氨基酸插入。这可以用来改变物质的特性(例如,辅助鉴别,纯化或表达)。
针对上述给定序列的氨基酸变化可使用任何适宜技术进行,例如,使用定点突变或固态合成。
应该认识到在本发明范围内的氨基酸置换或插入可以使用天然存在的或非天然存在的氨基酸进行。无论是使用天然或合成的氨基酸,优选仅存在l-氨基酸。
可以使用程序例如clustal程序来比较氨基酸序列。该程序通过在任一序列适当地插入空白来比较氨基酸序列并寻找最佳比对(optimalalignment)。可以计算氨基酸同一性或相似性(氨基酸类型的同一性和保守性)用于最佳比对。程序如blastx会排列出相似序列的最长延伸并给出拟合值。因而可以获得所发现的几个相似区域的比较,每个具有不同得分。在本发明中还考虑了两种同一性分析的类型。
本文所述的蛋白和多肽的变体应该保持原始蛋白或多肽的功能。可选地,或除了保持原始蛋白或多肽功能之外,本文所述的蛋白和多肽的变体与本文所述的蛋白或多肽(特别是图1c或图2所示的多肽序列)通常具有至少60%的同一性(如上所论述的)。通常地,本发明所用的变体与本文所述蛋白或多肽(特别是图1c或图2所示的多肽序列)具有至少70%,至少80%,至少90%,至少95%,至少97%或至少99%的同一性。
通过以最佳比较为目的的比对序列(如在第一序列中引入缺口以便与序列获得最佳比对),以及在相应位置比较氨基酸残基或核苷酸,来测定两个氨基酸序列或两个核酸序列的百分比同一性。“最佳比对”是能获得最高百分比同一性的两个序列的比对。通过在待比较序列中的相同氨基酸残基或核苷酸的数量来测定百分比同一性(即,%同一性=相同位置的数量/位置的总数量×100)。
可使用本领域技术人员已知的数学算法完成两个序列之间百分比同一性的测定。用于比对两个序列的数学算法的一个示例是karlinandaltschul(1990)proc.natl.acad.sci.usa87:2264-2268的算法,改良为karlinandaltschul(1993)proc.natl.acad.sci.usa90:5873-5877。altschul,etal.(1990)j.mol.biol.215:403-410的nblast和xblast程序引入这种算法。可用nblast程序实施blast核苷酸搜索,得分=100,字长=12以获得与本发明的核酸分子同源的核苷酸序列。可用xblast程序实施blast蛋白搜索,得分=50,字长=3以获得与本发明的蛋白分子同源的氨基酸序列。为了获得用于比较目的的带缺口的比对,可利用altschuletal.(1997)nucleicacidsres.25:3389-3402中所述的缺口blast。可选地,可使用psi-blast实施迭代搜索(iteratedsearch),其检测分子之间的距离关系(ld.)。在利用blast,缺口blast,和psi-blast程序时,可以使用各个程序(如xblast和nblast)的默认参数。参见http://www.ncbi.nlm.nih.gov。用于序列比较的数学算法的另一示例是myersandmiller,cabios(1989)的算法。作为cgc序列比对软件包一部分的align程序(2.0版本)引入了这种算法。本领域已知的用于序列分析的其他算法包括torellisandrobotti(1994)comput.appl.biosci.,10:3-5所述的advance和adam;和pearsonandlipman(1988)proc.natl.acad.sci.85:2444-8所述的fasta。在fasta内,ktup是设定搜索灵敏度和速度的控制选项。
在可选的方法中,变体可以是融合蛋白,其引入了便于纯化的基序,例如通过在所需蛋白或多肽中加上有效标签。可能需要去除“标签”或可能是融合蛋白本身保持足够可用的功能性的情况。
如本文所用的“能够与anx-a1结合的特异性结合分子”是与anx-a1的结合亲和力大于与其他分子的结合亲和力的分子,即其与anx-a1特异性结合。能够与anx-a1结合的特异性结合分子包括抗膜联蛋白-1(anti-anx-a1)抗体和适配体(aptamers)。用于本发明的抗-anx-a1抗体通过阻断t细胞激活而起作用,因而给药它时,可用于治疗t细胞介导的疾病,这些疾病通常是由异常的t细胞激活引起的。
anti-anx-a1抗体可针对例如具有图2所列出的氨基酸序列(通常是图2a所列出的氨基酸序列)的人anx-a1所产生。可选地,anti-anx-a1抗体可以定向于具有图2所列出的氨基酸序列的人anx-a1的一个或多个特定表位(epitope),通常是图2a所列出的氨基酸序列。例如,anti-anx-a1抗体可以定向针对anx-a1的n末端片段,例如来自图2a所列出氨基酸序列的n末端的至少188,100,50或25个氨基酸残基的n末端片段。通常地,用于本发明的anti-anx-a1抗体是针对被称为ac2-26的anx-a1的n末端片段,其具有图1c所示的序列,或针对它的至少6个氨基酸的片段。因此,能够与anx-a1结合的特异性结合分子包括anti-anx-a1抗体,其是针对具有图1c所示序列的anx-a1片段ac2-26或它的至少6个氨基酸的片段的抗体。在这个实施方式中,anti-anx-a1是针对图1c所示序列的片段所产生的,该片段是抗原性的且能够刺激抗体的产生,给药该抗体时,可以用于治疗t细胞介导的疾病,这些疾病通常是由异常的t细胞激活引起的。
如上所述,特异性序列的活性亚片段可以按说明使用。活性亚片段可含有具有图1c所列出序列的被称为ac2-26的anx-a1的n末端片段的至少6个连续氨基酸残基(六肽)的片段或由其所组成,该片段包括以下的一个或多个:
活性亚片段可含有具有图1c所列出序列的被称为ac2-26的anx-a1的n末端片段的大于6个连续氨基酸残基的片段或由其所组成,例如,图1c所列出序列的至少7个,至少8个,至少9个,至少10个,至少11个,至少12个,至少13个,至少14个,至少15个,至少16个,至少17个,至少18个,至少19个,至少20个,至少21个,至少22个,至少23个,至少24个氨基酸。
anti-anx-a1抗体包括单克隆和多克隆抗体。通常地,抗-anx-a1抗体是单克隆抗体。抗-anx-a1抗体可以是商业可供的抗体,例如兔多克隆或鼠单克隆抗体。通常地,抗-anx-a1抗体是人源化的,如以下具体所述。
单克隆抗体可以由杂交瘤产生。它们通常是由骨髓瘤细胞和可产生所需抗体的脾细胞融合形成的,从而形成永生细胞系。公知的kohler&milstein技术(nature256:495-497(1975))或该技术的后续变体都可用于产生符合本发明的单克隆抗体。
多克隆抗体的产生可以通过在适宜的动物宿主(如小鼠,大鼠,荷兰猪,兔,绵羊,山羊或猴)中将anx-a1,或其变体或片段注射至动物中刺激抗体产生。如果需要,可以与anx-a1蛋白一起给药佐剂。公知的佐剂包括弗氏佐剂(完全和不完全)和氢氧化铝。然后借助与anx-a1的结合纯化抗体。
产生可与特定多肽/蛋白结合的单克隆和多克隆抗体的技术现已在本领域中发展成熟,标准免疫学教科书中已有论述,例如,在roittetal.,immunologysecondedition(1989),churchilllivingstone,london。
除了整体抗体,本发明包括能够与本文所述anx-a1结合的抗体的衍生物。因而本发明包括抗体片段和合成构建体。抗体片段和合成构建体的示例由dougalletal在trendsbiotechnol,12:372-379(1994)中给出。
抗体片段包括,例如fab,f(ab')2和fv片段。fab片段如上文roittetal所论述的。fv片段可以被修饰以产生已知作为单链fv(scfv)分子的合成构建体。它包括共价连接可变重链(vh)和可变轻链(vl)区域的肽连接体,其有利于分子的稳定性。该连接体可含有1-20个氨基酸,例如1、2、3或4个氨基酸,5、10或15个氨基酸,或1-20范围内的其他适宜中间数。肽连接体可以由任何大体适宜的氨基酸残基形成,例如甘氨酸和/或丝氨酸。适宜连接体的一个示例是gly4ser。可以使用这种连接体的多聚体,例如二聚体、三聚体、四聚体或五聚体,如(gly4ser)2、(gly4ser)3、(gly4ser)4或(gly4ser)5。然而,在其他实施方式中,不存在肽连接体,vl结构域与vh结构域通过肽键连接。
特异性结合分子可以是单链可变片段(scfv)的类似物。例如,scfv可以与其他特异性结合分子连接(例如其他scfv、fab抗体片段和嵌合(chimeric)igg抗体(如具有人框架(humanframeworks)))。scfv可以与其他scfv连接以便形成多聚体,其是多特异性结合蛋白,例如二聚体、三聚体或四聚体。双特异性scfv有时是指双抗体,三特异性指三抗体,以及四特异性指四抗体。
scfv可使用标准的化学或分子生物学技术通过任何适宜技术制备。在本发明的一种实施方式中,单克隆抗体类似物可由人天然抗体噬菌体展示文库制备成scfv(mccaffertyetal,nature348,552-554(1990);如wo92/01047中所描述)。
可用的其他合成构建体包括互补决定区(cdr)肽。它们是含有抗原结合决定簇的合成肽。也可以使用肽模拟物。这些分子通常在结构上是受限的有机环,其模拟了cdr环的结构且包括与抗原作用的侧链。
合成构建体包括嵌合分子。因而,人源化抗体或其衍生物处于可用于本发明的抗体范围内。人源化抗体的方法是本领域公知的。通过修饰抗体的氨基酸序列可将抗体人源化。人源化抗体的示例是具有人框架区域,但具有啮齿目动物(例如鼠科动物)高变区域的抗体。产生嵌合抗体的方式例如由morrisonetal在pnas,81:6851-6855(1984)以及takedaetal在nature,314:452-454(1985)中论述过。人源化可被实施,例如,由jonesetal在nature,321:522-525(1986);verhoeyenetal在science,239:1534-1536;riechmannetal在nature332:323-327,1988中所描述的。降低本发明特异性结合分子的免疫原性的方法可包括,通过例如定点突变或其他常用分子生物学技术,将cdr移植至适宜的抗体框架支架或可变表面残基重塑(roguskaetalproteineng.9895-904(1996))。
其他可用方法包括鉴别分子内的潜在t-细胞表位,以及随后将其去除,例如通过定点突变(去免疫)。可以按照需要对cdr区域或周围框架序列进行人源化。
合成构建体也包括含有额外部分的分子,该基序能提供具有除抗原结合之外的一些期望特性的分子。例如,该部分可以是标记(如荧光或放射性标记)。可选地,它可以是药物活性剂。
本发明涉及能够与anx-a1结合的特异性结合分子在治疗t细胞介导的疾病中的应用。
本发明可用于治疗多种t细胞介导的疾病。在本文中,“t细胞介导的疾病”表示t细胞在该疾病或病状的致病或发展中发挥作用的任何疾病或病状。t细胞介导的疾病通常是由异常的t细胞激活引起的。因此,这种疾病可通过阻断anx-a1活性防止t细胞激活得到治疗。通常地,本发明所治疗的t细胞介导的疾病是其中th1细胞发挥作用的疾病。
t细胞介导的疾病包括但不限于移植物抗宿主病(graft-versus-hostdisease),移植物排斥(graftrejection),动脉粥样硬化,hiv和/或aids,银屑病和一些自身免疫疾病。根据本发明,可以治疗的自身免疫疾病包括类风湿性关节炎(rheumatoidarthritis,ra),多发性硬化症(ms),系统性红斑狼疮(systemiclupuserythematosus,sle),艾迪生病,格雷夫斯病(grave’sdisease),硬皮病,多发性肌炎(polymyositis),某些形式的糖尿病(例如青少年型糖尿病),自身免疫性葡萄膜视网膜炎(autoimmuneuveoretinitis),溃疡性结肠炎,寻常天疱疮,炎症性肠病和自身免疫性甲状腺炎。t细胞介导的疾病通常是类风湿性关节炎,多发性硬化症,系统性红斑狼疮或动脉粥样硬化。
t细胞介导的疾病通常是类风湿性关节炎。在类风湿性关节炎(ra)中,认为t细胞可识别滑膜中展示抗原的细胞并与其作用。一旦被激活,这些细胞会产生细胞因子和效应分子;这种细胞因子的依次、扩大的产生构成了“细胞因子级联”,其导致了巨噬细胞的激活并诱导炎症过程,最终降解并吸收软骨和骨骼。随着时间推移,可以发生骨腐蚀,软骨毁坏,和关节完整性的完全丧失。最后,可能影响多个器官系统。
在另一实施方式中,t细胞介导的疾病是动脉粥样硬化。炎症在冠状动脉疾病和其他动脉粥样硬化的显示中发挥关键作用。免疫细胞主导着早期动脉粥样硬化病变,它们的效应分子加速了病变的发展,炎症的激活可以引发急性冠状动脉综合征。适应性免疫深度参与动脉粥样硬化形成,因为已显示它与代谢风险因子相互作用以启动、增殖并激活动脉树的损害。
具有高胆固醇血症和快速发展的动脉粥样硬化特征的两个小鼠模型,apoe-/-和低密度脂蛋白受体敲除的小鼠(ldlr-/-),可用于动脉粥样硬化的研究,因为它们模拟了人损害的细胞组成,特别是t淋巴细胞的内容物。在疾病开始之前,淋巴细胞的补充在具有动脉粥样硬化倾向的apoe-/-小鼠的动脉中增加了。
t-淋巴细胞的存在具有功能性结果,因为它们的完全缺乏会减少中度的高胆固醇血症期间损害的形成。分泌cd4+ifn-γ的1型辅助(th1)细胞是斑块(plaque)中发现的主流类型的t细胞,这些t细胞产生致动脉粥样硬化和斑块不稳定的作用。
本发明人目前发现了在人和鼠类的动脉粥样硬化斑块中都表达anx-a1,在小鼠模型中anx-a1表达和ms之间存在关联。
在另一实施方式中,t细胞介导的疾病是系统性红斑狼疮(sle)。目前本发明人发现了anx-a1mrna和蛋白在sle患者的t细胞中的表达水平高于在健康志愿者的t细胞中的。
与anx-a1偏好分化为th1细胞的能力相关的,本发明也可用于例如限制不受控制的保护细胞(th1)针对细胞内病原体的应答,和通过抑制th1分化和偏向th2分化来治疗细胞外感染(th2反应)。
能够与anx-a1结合的特异性结合分子通常与药物可接受载体、赋形剂、介质(vehicle)、佐剂和/或稀释剂配制使用。因而本发明包括含有能够与膜联蛋白-1(anx-a1)结合的特异性结合分子的药物组合物,其可用于治疗t细胞介导的疾病。该药物组合物含有能够与膜联蛋白-1(anx-a1)结合的特异性结合分子和药物可接受载体,赋形剂,介质,佐剂和/或稀释剂。这种组合物可通过药学领域已知的任何方法来制备,例如通过在无菌条件下将活性成分与载体,赋形剂,介质和/或稀释剂混合制备。
适宜的载体,介质,佐剂和/或稀释剂是本领域公知的,包括盐水,磷酸缓冲液(pbs),羧甲基纤维素(cmc),甲基纤维素,羟丙基甲基纤维素(hpmc),右旋糖,脂质体,聚乙烯醇,药用淀粉,甘露醇,乳糖,硬脂酸镁,糖精钠,滑石,纤维素,葡萄糖,蔗糖(和其他糖),碳酸镁,明胶,油,醇类,去污剂,乳化剂或水(优选无菌的)。能够与anx-a1结合的特异性结合分子可配制成液体制剂,其通常是由在适宜的水性或非水性液体载体或例如水,乙醇,甘油,聚乙二醇(peg)或油的载体中的能够与anx-a1结合的特异性结合分子的悬浮液或溶液所组成。
通常地,当能够与anx-a1结合的特异性结合分子是抗体时,抗体被peg化,即共价附着在聚乙二醇上。通常地,这具有降低所述抗体的免疫原性并增长半衰期的作用。
能够与anx-a1结合的特异性结合分子可以单独给药或与其他试剂一起给药。
用于本发明的能够与anx-a1结合的特异性结合分子通常以有效治疗量给药至受试者。这个剂量是可有效改善、消除或预防一种或多种t细胞介导的疾病的症状的量。优选地,待治疗的受试者是人。然而,本发明可同样适用于人或动物药物(veterinarymedicine)。例如,本发明被发现可用于治疗伴侣动物,例如狗和猫,或役用动物(workinganimals)例如赛马。
能够与anx-a1结合的特异性结合分子可通过任何适宜方式给药至受试者。能够与anx-a1结合的特异性结合分子可以全身给药,特别是关节内,动脉内,腹膜内(i.p.),静脉内或肌肉内。然而,能够与anx-a1结合的特异性结合分子也可以通过其他肠道或非肠道途径,例如通过皮下、真皮内、局部(包括口腔,舌下或经皮肤)、口服(包括口腔或舌下)、鼻腔、阴道、肛门、肺或其他适当给药途径来给药。
适合口服给药的药物组合物可呈现为分离单位,例如胶囊或片剂;粉末或颗粒;溶液,糖浆或悬浮液(在水性或非水性液体中;或为可食性泡沫或搅打物(whip);或为乳状液)。适宜用于片剂或硬胶囊的赋形剂包括乳糖,玉米淀粉或其衍生物,硬脂酸或其盐。适宜用于软胶囊的赋形剂包括例如植物油,蜡,脂肪,半固体,或液体多元醇等。用于制备溶液和糖浆,可使用的赋形剂包括例如水,多元醇和糖。用于制备悬浮液,可使用油(如植物油)以提供水包油或油包水型悬浮液。
适合经皮给药的药物组合物可呈现为分离贴片,以便长时间地保持与接受者表皮的直接接触。例如,活性成分可以通过离子渗透从贴片递送,如pharmaceuticalresearch,3(6):318(1986)所概述的。
适合局部给药的药物组合物可配制成软膏,乳膏,悬浮液,洗液,粉末,溶液,糊剂,凝胶,喷剂,气雾剂(aerosols)或油脂剂(oils)。为了侵入眼睛或其他外部组织,例如嘴巴和皮肤,组合物优选应用成局部软膏或乳膏。若配制成软膏,活性成分可与石蜡或易与水混溶的软膏基质使用。可选地,活性成分可与水包油乳膏基质或油包水基质配制成乳膏。适合局部给药至眼睛的药物组合物包括眼睛滴液,其中活性成分溶解或悬浮在适宜载体中,特别是水性溶剂中。适合局部给药至嘴的药物组合物包括锭剂,软糕和漱口剂(mouthwashes)。
适合直肠给药的药物组合物可呈现为栓剂或灌肠剂。
适合鼻腔给药的、其中载体是固体的药物组合物包括粒径范围在例如20-500微米的粗粉,其通过从鼻吸入的方式给药,即从紧靠鼻子的粉末容器中通过鼻腔通道快速吸入。载体是液体的、且作为鼻腔喷剂或鼻腔滴液给药的适宜组合物包括活性成分的水性或油性溶液。
适合通过吸入给药的药物组合物包括细粒粉末或液体喷雾,其可通过多种类型的测量剂量的加压气雾剂,雾化器或吹药器(insufflators)的方式生成。
适合阴道给药的药物组合物可呈现为子宫托,卫生棉条,乳膏,凝胶,糊剂,泡沫或喷雾制剂。
适合非肠道给药的药物组合物包括水性和非水性无菌注射溶液,其可含有抗氧化剂,缓冲剂,抑菌剂和使得制剂与预期接受者的血液基本等渗的溶质;和水性和非水性无菌悬浮液,其可包括悬浮剂和增稠剂。可用于可注射溶液的赋形剂包括例如水,醇类,多元醇,甘油和植物油。组合物可以单位剂量或多剂量容器呈现,例如密封的安瓶和小瓶,并可以冻干(冻干)状态存储,只需要在使用前即时添加所携带的无菌液体,例如注射用水。临时注射溶液和悬浮液可由无菌粉末,颗粒和片剂制备。
药物组合物可含有防腐剂,溶解剂,稳定剂,润湿剂,乳化剂,甜味剂,着色剂,香味剂,盐,缓冲剂,包被剂或抗氧化剂。除了能够与anx-a1结合的特异性结合分子之外,它们也可以含有治疗活性剂。
能够与anx-a1结合的特异性结合分子的给药剂量可根据多个参数,特别是根据所用的能够与anx-a1结合的特异性结合分子;待治疗的患者的年龄,体重和状态;给药途径;和所需要的方案(regimen)来决定。医生能够确定特定患者所需要的给药途径和剂量。
这个剂量可按适当频率重复给药。如果出现副作用可根据正常临床实践减少给药的剂量和/或频率。
对于给药至哺乳动物,特别是人,预期的活性剂日剂量是1μg/kg至10mg/kg体重,通常是约10μg/kg至1mg/kg体重。在任何情况中医生会确定最适合于个体的实际剂量,这取决于以下因素,包括个体的年龄,体重,性别和应答。上述剂量是平均病例的示例。当然,存在应给予更高或更低剂量的情况,这也处于本发明范围内。
在本发明的第二方面,提供了能够与anx-a1结合的特异性结合分子在制备治疗t细胞介导的疾病的药物中的用途。
在本发明的第三方面,提供了治疗t细胞介导的疾病的方法,其包括将治疗量的能够与anx-a1结合的特异性结合分子给药至有需要的受试者。如上所述的,方法治疗的可以是人或动物受试者,本发明扩展至等同于用于人和/或动物药物的治疗方法。
本发明第二和第三方面的优选特征是对第一方面的修正。
附图说明
现在将通过参照以下实施例和附图的方式进一步描述本发明,提供它们的目的仅在于示例说明而不会对本发明构成限制。参照了以下附图,其中:
图1a是显示出四个膜联蛋白重复序列和n末端结构域的膜联蛋白-1结构的缎带图。图1b是膜联蛋白重复序列和生物活性序列、膜联蛋白-1肽ac.2-26定位的示意图。图1c显示了肽ac.2-26的氨基酸序列,其是anx-a1的乙酰化n末端肽片段。
图2a显示了人膜联蛋白-1(anx-a1),同工型anxa1-003的(i)氨基酸序列和(ii)核苷酸序列。图2b显示了人膜联蛋白-1(anx-a1),同工型anxa1-002的氨基酸序列。图2c显示了人膜联蛋白-1(anx-a1),同工型anxa1-004的氨基酸序列。图2d显示了人膜联蛋白-1(anx-a1),同工型anxa1-006的氨基酸序列。
图3显示了人重组膜联蛋白-1(hranx-a1)对t细胞激活的作用。用hranx-a1预处理鼠类cd4+原初细胞(primarycells)之后用不同浓度的抗cd3/cd28激活扩增了细胞增殖(图3a),il-2产生(图3b)和cd25和cd69在细胞表面的表达(图3c和3d)。
图4显示了内源anx-a1介导t细胞增殖。与未刺激t细胞的对照相比,在缺乏anx-a1的t细胞中,用抗cd3、抗cd3/cd28或pma/离子霉素刺激anx-a1+/+或anx-a1-/-t细胞,显示出3h-胸苷引入(分别是图4a,4b和4c)和il-2产生(图4d)的速率降低。
图5显示了在存在或缺乏anx-a1下,激活蛋白-1(ap-1)、核因子-κb(nf-κb)和激活的t细胞的核因子(nfat)的激活(图5a),以及在anx-a1+/+和anx-a1-/-t细胞中,ap-1、nf-κb和nfat激活的对比(图5b)。
图6a显示了按指定次数用anti-cd3/cd28(抗-cd3/cd28)(5.0μg/ml)刺激的t细胞中fprl-1表达的facs分析。图6b显示了用anti-cd3/cd28(5.0μg/ml)刺激之前(对照)或之后t细胞中anx-a1的细胞定位。图6c是anx-al/fprl-1体系在t细胞中的作用的示意图。
图7显示了外源和内源anx-a1介导th1/th2分化。图7a显示了在存在或缺乏hranx-a1下,天然淋巴结t细胞在体外分化成th1(黑柱)或th2(白柱)状态,然后用结合在板上的anti-cd3重复刺激以测量th1或th2细胞因子产生的结果。图7b显示了来自anx-a1+/+和anx-a1-/-小鼠的天然淋巴结t细胞在体外分化成th1(左起第一和第二柱状图)或th2(左起第三和第四柱状图)状态,然后用结合在板上的anti-cd3重复刺激以测量th1或th2细胞因子产生的结果。
图8显示了在胶原诱发性关节炎(cia)模型的免疫阶段,用pbs或hranx-a1治疗12天的dba小鼠的脚掌体积(pawvolume)(图8a)和临床得分(图8b)。图8c是健康对照志愿者(hc)或类风湿性关节炎(ra)患者的cd4+细胞中的anx-a1表达分析。图8d显示了ra患者滑膜组织中anx-a1表达的免疫组织化学分析。
图9显示了全长hranx-a1和n末端肽ac2-26对t细胞激活的作用。
图10显示了在人粥样硬化斑块中anx-a1的表达。在颈动脉内膜剥脱手术期间,从患者中取出的颈动脉粥样硬化斑块中,用小鼠单克隆抗-人anx-a1抗体ib(图10a)或用非免疫igg(图10b)的anx-a1表达的免疫组织化学分析。图像是来自单个患者和具有相似病状的六个不同患者的代表。
图11显示了在鼠类动脉粥样硬化斑块中anx-a1的表达。图11显示了在apoe-/-小鼠主动脉窦(图11a和图11b)和肱动脉(brachiocefalicartery)(图11c)中anx-a1免疫荧光显影。切片用dapi染色以定位核区。所示的结果是来自单个实验和三个独立实验的代表图。原始放大倍数:x200(图11a和图11b),x400(图11c)。
图12显示了在系统性红斑狼疮(sle)患者中的膜联蛋白-1的表达。来自健康人(对照)或系统性红斑狼疮(sle)患者的t细胞中anx-a1表达的rt-pcr(上面组)和western印染(下面组)分析。图中的数量表示从健康人(对照)或系统性红斑狼疮(sle)患者中收集的相同数量(2x106)的t细胞中获得的cdna的体积(μl)或蛋白的量(μg)。
图13显示了t细胞受体(tcr)的激活抑制,根据在与中和单克隆抗体孵育的来自一个捐赠者的人外周t细胞中白介素-2(il-2)产生的测量,其中抗体是针对人重组膜联蛋白-1(anti-anxalmabla)产生的。
图14显示了t细胞受体(tcr)的激活抑制,根据在与中和单克隆抗体孵育的来自不同捐赠者的人外周t细胞中白介素-2(il-2)产生的测量,其中抗体是针对人重组膜联蛋白-1(anti-anxalmabla)产生的。
图15显示了用mog35-55和cfa免疫的c57bl/6小鼠的脊髓切片,其中脊髓是在第12天(0分),第18天(2分)和第20天(4分)取出的。切片用苏木精和伊红(h&e,图15a)或anti-anxal(图15b)染色。对于每个染色,右边组(20x)显示比左边区域的组(4x)更高的放大倍数。结果是3个实验的代表图。
图16显示了用mog35-55和cfa免疫的c57bl/6小鼠的脊髓切片,其中脊髓是在第20天(4分)取出的。切片用anti-anxal和anti-cd3(a)或抗-f4/80(b)染色。对于每个染色,右边组显示右边的两个单独染色的重叠图。结果是3个实验的代表图。
图17显示了用mog35-55和cfa免疫的c57bl/6小鼠的研究结果,23天内每日监测小鼠的eae(a)或体重增加/降低(b)的体征(signs)和症状。结果是平均值±sem(n=10/组)。**p<0.01,是3个实验的代表图。
图18显示了从用mog35-55和cfa免疫并在14天后处死的anx-a1+/+和anx-a1-/-小鼠获得的淋巴结细胞中3h-胸苷的引入(a)和il-2的产生(b)。细胞用mog35-55刺激48小时并用lμci3h-胸苷脉冲12小时。细胞上清液用于测量il-2的产生。结果是平均值±sem(n=4/组)。*p<0.05,**p<0.01,是3个实验的代表图。
图19显示了从用mog35-55和cfa免疫并在14天后处死的anx-a1+/+和anx-a1-/-小鼠获得的脾(a)和淋巴结(b)细胞的总细胞数量。c和d显示了具有抗-cd4fitc和抗-cd8pe的淋巴结细胞的细胞荧光分析。结果是平均值±sem(n=10/组)。**p<0.01,是3个实验的代表图。
图20显示了从用mog35-55和cfa免疫并在14天后处死的anx-a1+/+和anx-a1-/-小鼠获得的淋巴结细胞的细胞上清液中(a)ifn-γ,(b)il-2,(c)tnf-α和(d)il-17的水平。细胞用指定浓度的mog35-55刺激4天而上清液用于细胞因子elisa。结果是平均值±sem(n=4/组)。*p<0.05,**p<0.01,是3个实验的代表图。
图21显示了从用mog35-55和cfa免疫并在22天后处死的anx-a1+/+(a)和anx-a1-/-(b)小鼠获得的脊髓切片的苏木精-伊红染色。对于每个染色,右边组(20x)显示比左边区域组(4x)更高的放大倍数。连续切片用anti-cd3(c)或抗-f4/80(d)染色。图片是具有相似结果的3个独立实验的代表图。
图22显示了从用mog35-55和cfa免疫并在14天后处死的anx-a1+/+(a)和anx-a1-/-(b)小鼠获得的脊髓匀浆的percoll梯度中回收的cd3(a)和f4/80(b)阳性单核细胞的facs分析。散点图和柱状图是来自单个小鼠和n=4个小鼠的2个实验的代表图。柱状图的数量指示了cd3+和f4/80+细胞的百分比。
具体实施方式
实施例1-10
材料和方法
试剂
抗-小鼠cd3(克隆145-2c11),抗-小鼠cd28(克隆37.51),抗-人cd3(克隆okt3),抗-人cd28(克隆cd28.2),偶联pe的抗-cd69(克隆h1.2f3),偶联fitc的抗-cd25(克隆pc61.5),鼠类il-2,il-4,ifn-γ,il-12,抗-il-4(克隆11b11),和抗-ifn-γ(克隆xmgl.2)购买自ebioscience(wembley,英国)。按描述制备了无内毒素的人重组anx-al(hranx-al)。在一些实验中,我们使用变性的hranx-al(95℃5分钟热失活)作为阳性对照。除非另外指明,所有其他试剂都来自sigma-aldrich(stlouis,mo)。
小鼠
balb/c、c57/bl6和dba/1雄性小鼠是从charlesriver实验室(wilmington,ma)获得的。膜联蛋白1敲除小鼠balb/c是在我们实验室生成的并在我们动物工厂中在无病原体条件下培养。此研究使用的所有小鼠都是6-8周龄。按照英国内政部规定(动物操作指南,科学程序法案1986)和欧盟指导守则实施动物实验。
从患者中分离细胞
使用ficoll密度离心(ficoll-paqueplus;amershambiosciences,freiburg,德国)从外周血液中制备外周血单核细胞(pbmcs)。使用阳性选择从外周血中选择cd4+细胞。简而言之,将外周血进行ficoll密度离心(ficoll-paqueplus;amershambiosciences)。通过粘附在血清包被塑料制品上从单核细胞中取出粘附细胞。非粘附细胞与小鼠抗-人cd4抗体(rft4)孵育,用缓冲液(磷酸缓冲液[pbs],0.5%牛血清白蛋白[bsa],2mmedtaph7.2)洗涤,与偶联磁珠的羊抗鼠抗体(miltenyibiotec,auburn,ca)孵育。细胞流经macs柱(miltenyibiotec)并收集cd4+细胞。通过流式细胞计估算细胞纯度。经过10次衰竭(depletions)之后cd3+cd4+细胞的中间百分比是98%(范围97%-99.3%)。剩余细胞重悬在裂解缓冲液(ambion,huntingdon,英国)。
细胞培养
通过阴性选择由淋巴结制备原发鼠类t细胞。简而言之,将腋窝,腹股沟,和肠系膜淋巴结挑出制成单细胞悬浮液,然后洗涤并用ficoll分层。将血沉棕黄层洗涤2次然后与抗体混合物和磁珠孵育,按照生产商的说明(小鼠t-细胞阴性分离试剂盒;dynal,bromborough,英国)。在一些实验中,使用miltenyibioteccd62l+cd4+t-细胞分离试剂盒进一步纯化细胞获得原初cd62l+cd4+t细胞。在含有鼠类il-2(20u/ml)、anti-cd3(5μg/ml)和抗-cd28(5μg/ml)的完全rpmi培养基(10%胎牛血清[fcs],2mml-谷氨酰胺,和每ml100单位的庆大霉素)预包被的6孔板中孵育t细胞,孵育4天创建th0条件。用鼠类il-12(3.4ng/ml;ebioscience),il-2(20u/ml;ebioscience)和抗-il4(克隆11b11;2μg/ml)创建th1条件。用il-4(3000u/ml;peprotech,rockyhill,nj),il-2(20u/ml)和抗-ifn-γ(克隆xmgl.2;2μg/ml)创建th2条件。jurkat细胞来自atcc(manassas,va)并在完全rpmi培养基中培养。
流式细胞计数分析
37℃下,在微量离心管中将纯化的淋巴结t细胞用人重组anx-a1预处理2小时,然后用结合在板上的anti-cd3和抗-cd28刺激,如图所示。16小时之后,细胞用facs缓冲液(含有1%fcs和0.02%nan2的pbs)稀释的偶联pe的抗-cd69(克隆h1.2f3)和偶联fitc的抗-cd25(克隆pc61.5)染色。通过向前和侧向扩散限出完整细胞,并用facscan流式细胞计上的cellquest程序(bectondickinson,franklinlakes,nj)分析。为了分析fprl-1表达,用结合在板上的anti-cd3和抗-cd28多次刺激人外周血t细胞,之后用鼠抗人fprl-1(克隆6c7-3-a;5μg/ml),然后用偶联fitc的抗体进行染色。
细胞增殖实验
37℃下,在微量离心管中将纯化的淋巴结t细胞(105细胞/ml)单独与培养基或与不同浓度的hranx-a1孵育2小时。之后,在96孔板中用结合在板上的anti-cd3和抗cd-28刺激200-μl等份的细胞悬浮液24小时。18小时之后,用lμci(3.7×104bg)的[3h]-胸苷(amershampharmaciabiotech,piscataway,nj)脉冲8小时引入的放射性,并通过自动闪烁计数器(packardinstruments,pangbourne,英国)测量放射性。
电泳迁移率改变试验(electromobilityshiftassay)
按照之前所述的方案,从3×106至5×106细胞中收集核提取物。将核提取物(3μg-5μg)与在20μl结合缓冲液中具有32p末端标记的双链寡核苷酸探针(5×105cpm)的1μg(对于nfat)或2μg(对于nf-κb和ap-1)多聚体(di:dc)孵育,并在150v下,在0.5%tbe的6%聚丙烯酰胺凝胶(29:1交联比)中分离2.5小时。nf-κb和ap-i结合缓冲液(10x)含有100mmtris-hcl,(ph7.5),500mmnacl,10mmedta,50%甘油,10mg/ml白蛋白,30mmgtp,10mmdtt。nfat结合缓冲液(10x)含有100mmhepes(ph7.9),500mmkcl,1mmedta,1mmegta,50%甘油,5mg/ml白蛋白,1%nonidetp-40,10mmdtt。nf-κb和ap-1双链寡核苷酸探针是来自promega,而nfat来自santacruzbiotechnology(santacruz,ca)。
western印染分析
如图所示,孵育淋巴结t细胞。在37℃孵育不同时间段之后,将细胞在冰冷的裂解缓冲液(1%np-40,20mmtrisph7.5,150mmnacl,1mmmgcl2,1mmegta,0.5mmpmsf,1μm抑酶肽,1μm亮抑酶肽,1μm胃酶抑素,50mmnaf,10mmna4p2o7,和1mmnavo4,1mmβ-甘油磷酸)中裂解。将细胞裂解物在4℃13/226g(13000)rpm离心5分钟,收集上清液并在sds-10%聚丙烯酰胺凝胶中电泳。转膜后,在4℃将膜与用含有tween-20和5%无脂奶粉的tris-缓冲液(ttbs:0.13mnacl;2.68mmkcl;0.019mtris-hcl;0.001%vol/voltween-20;ph7.4)稀释的抗体孵育过夜。对于使用抗-perkl/2和抗-akt的实验,用补充了50mmnaf和牛血清白蛋白(5%)的ttbs缓冲液替代牛奶。对于每种情况,使用从相同数量细胞中获得的等量提取物。按照生产商的说明通过增强的化学发光(ecl;amershampharmaciabiotech)进行蛋白的免疫印染和显影。为了获得细胞溶质和膜成分,首先收集细胞并在冰冷的pbs中洗涤,然后在300g简单离心2分钟。将获得的细胞沉淀在裂解缓冲液(20mmtris-hcl,ph7.5;在裂解缓冲液所列的蛋白酶抑制剂)中裂解,并通过25-规格注射器至少5次以确保裂解充分。然后将悬浮液在300g离心2分钟,收集上清液,在800g再离心45分钟(4℃)。在该阶段,收集上清液(细胞溶质成分),并将沉淀(膜成分)重悬在含有1%(vol/vol)tritonx-100的裂解缓冲液中。整个实验中所有成分都保持在冰上。
细胞因子elisa
对于th1/th2细胞因子产生的分析,在24孔板中,将在偏移条件(skewingconditions)中分化4天并在完全rpmi培养基中静置1天之后获得的th0/th1/th2细胞(106/ml)用结合在板上的anti-cd3(5μg/ml)刺激8h。收集培养物上清液,并使用th1/th2panelelisa试剂盒(ebioscience)分析ifn-γ、1l-2、il-4和il-10的含量。il-13试剂盒也购买自ebioscience。
实施例1-人重组膜联蛋白-1(hranx-a1)对t细胞激活的作用
在缺乏或存在不同浓度的hranx-a1下,将鼠类原初淋巴结t细胞用5.0(■),2.5(▲)和1.25(▼)μg/ml的anti-cd3/cd28刺激24h,然后用3h-胸苷脉冲以测量增殖。结果如图3a所示。
图3b显示了在缺乏或存在不同浓度的hranx-a1下,主要从用anti-cd3/cd28(1.25μg/ml)刺激24h的鼠类原初淋巴结t细胞中所产生的il-2。
在缺乏(上面组)或存在(下面组)hranx-a1(600nm)下,用浓度为1.25μg/ml(左柱),2.5μg/ml(中柱)和5.0μg/ml(右柱)的anti-cd3/cd28(1.25μg/ml)刺激鼠类原初淋巴结t细胞12h,然后通过facs分析cd25和cd69表达。结果如图3c所示。
在图3d中,在150(x),300(*)和600(o)nm的hranx-a1存在下,用指定浓度的anti-cd3/cd28刺激鼠类原初淋巴结t细胞12h,然后通过facs分析cd25(左图)和cd69(右图)表达。
在所有实验中,数值是n=3-4只小鼠的平均值±s.e.。*p<0.05;**p<0.01。
结果显示了鼠类原初cd4+原发细胞用hranx-a1预处理之后用不同浓度的anti-cd3/cd28激活,增强了细胞增殖(图3a),il-2产生(图3b)和cd25和cd69在细胞表面的表达(图3c和d)。
实施例2-内源anx-a1介导t细胞增殖
图4显示了:与未刺激t细胞的对照相比,(a)anti-cd3(5.0μg/ml)(b)anti-cd3/cd28(5.0μg/ml)或(c)pma(20ng/ml)和离子霉素(2ng/ml)诱导的野生型和anx-al缺陷的t细胞增殖,表示为3h-胸苷引入的百分比。在一些实验中,存在小鼠重组il-2(20ng/ml)也可激活细胞。数值是n=4-5的平均值±s.e.。对il-2刺激的anx-al+/+细胞,p<0.01;对anti-cd3或anti-cd3/cd28或pma/离子霉素刺激的anx-al+/+细胞,**p<0.01;对anti-cd3或anti-cd3/cd28或pma/离子霉素刺激的anx-al-/-细胞,§§p<0.01。
图4d显示了从用anti-cd3,anti-cd3/cd28(5.0μg/ml)或pma(20ng/ml)和离子霉素(2ng/ml)刺激24h的鼠类原初淋巴结t细胞中所产生的il-2。数值是n=4-5只小鼠的平均值±s.e.。**p<0.01。
结果显示了与未刺激t细胞的对照相比,在缺乏anx-a1的t细胞中,anti-cd3,anti-cd3/cd28或pma/离子霉素对anx-al+/+或anx-al-/-t细胞的刺激表现出3h-胸苷引入(分别是图4a,4b和4c)和il-2产生(图4d)的速率降低。
实施例3-在存在或缺乏anx-a1下ap-i、nf-κb和nfat的激活
对于内源和外源anx-a1如何介导t细胞激活进行了研究。分析了在hranx-a1存在下的刺激细胞中的三个主要的t细胞转录激活因子,称为激活蛋白-1(ap-1),核因子-κb(nf-κb)和激活t细胞的核因子(nfat)。
图5a是在存在或缺乏指定浓度的hranx-a1下,用anti-cd3/cd28(1.25μg/ml)刺激的t细胞中ap-1,nf-κb和nfat激活的电泳迁移率改变试验。图5b显示了在用anti-cd3/cd28(5.0μg/ml)刺激的anx-a1+/+和anx-a1-/-t细胞中ap-1、nf-κb和nfat激活的对比。
结果证实了所有三个转录因子都增加激活(图5a)。相反地,与它们的对照相比,anx-a1-/-t细胞显示出这些转录因子的减少激活(图5b)。
实施例4-fprl-1和anx-a1在t细胞中的外化
我们研究了t细胞是否表达anx-a1的受体,甲酰肽受体样-1(fprl-1)。特异性单克隆抗fprl-1抗体对未刺激的人外周血t(pbt)细胞的facs染色证实了没有表达受体。然而,anti-cd3/cd28的刺激诱导了fprl-1在1小时之内外化,之后在细胞表面以稳定的稳态表达(图6a)。有趣的是,对anx-a1观察到相同模式。因而,anx-a1在人pbt中分布的分析证实了蛋白平均地分布在胞质和膜之间。然而,当细胞被anti-cd3/cd28刺激时,观察到膜中anx-a1的积累。
然后蛋白输出至膜外侧,并释放胞外环境中。与此模式一致,当我们对来自用anti-cd3/cd28刺激的人pbt的培养上清液中的anx-a1进行免疫沉淀时,我们观察到与未刺激细胞的对照相比anx-a1的增加释放(图6b)。综合而言,这些观察证实了通过tcr信号增加了anx-a1释放,并伴随着其受体的上调。
在生理条件下,anx-a1/fprl-1整合tcr以介导tcr信号的强度。然而,在病理条件下,例如在ra或系统性红斑狼疮中(未公开数据),其中蛋白以更高水平表达,这可导致由于tcr信号阈值的降低而增加的t细胞激活(图6c)。
实施例5-外源和内源anx-a1介导th1/th2分化
最近的研究假定了tcr信号强度会影响符合th1/th2效应细胞的t细胞谱系。增加或减少在用hranx-al(图3和5a)或anx-a1-/-细胞(图4和5b)处理的t细胞中的tcr信号,然后我们搜寻确定不同水平的anx-al是否会影响t细胞分化成th1或th2细胞。
在存在或缺乏hranx-al(600nm)下,原初淋巴结t细胞在体外分化成th1(黑柱)或th2(白柱)状态,然后用结合在板上的anti-cd3(5.0μg/ml)重复刺激8h以测量产生的th1或th2细胞因子。结果如图7a所示。数值是n=4-5只小鼠的平均值±s.e.。**p<0.01
如图7a所示的,在存在hranx-al下,原初t细胞(cd441o,cd62lhi)分化成(anti-cd3/cd28,il2,il12和抗-il4)或th2(anti-cd3/cd28,il2,il4和抗-ifnγ)状态增加了il2和ifnγ产生,并在anti-cd3重复刺激的基础上伴随着il4和il10释放的减少。
来自anx-a1+/+和anx-a1-/-小鼠的原初淋巴结t细胞在体外分化成th1(左起第一和第二柱状图)或th2(左起第三和第四柱状图)状态,然后用结合在板上的anti-cd3(5.0μg/ml)重复刺激8h以测量产生的th1或th2细胞因子。结果如图7b所示。数值是n=4-5只小鼠的平均值±s.e.。**p<0.01
如图7b所示的,对于内源蛋白也获得了相似的发现:分析了来自anx-a1+/+或anx-a1-/-小鼠的分化th1/th2细胞中的th1/th2细胞因子产生,与敲除的小鼠相比,野生型小鼠产生了更高水平的il2和ifnγ,而il4和il13的产生具有相反的曲线。
实施例6-anx-a1和类风湿性关节炎
为了证明hranx-a1在体内增加了t细胞激活,我们使用慢性自身免疫疾病的小鼠模型,胶原诱发性关节炎(cia)的dba小鼠模型。用胶原(在原初细胞分化成th效应细胞期间)免疫之后,每天给小鼠注射hranx-a1共12天,之后基于抗原攻击分析疾病的发展。图8显示了用pbs(100μl)或hranx-al(1μgs.c.一天两次)处理的小鼠的脚掌体积(图8a)和临床得分(图8b)。在21天通过提高胶原获得了疾病开始的同步,从22天开始临床信号显现(疾病开始的第1天)。数值是6-8只小鼠的平均值±s.e.。各组使用曼-惠特尼检验进行比较。*p<0.01
如图8a和8b所观察到的,与用pbs介质处理的小鼠相比,用hranx-al处理的小鼠恶化了关节炎的体征和症状,证实了高水平的anx-al会影响t细胞激活和分化,这些作用会影响ra小鼠模型的疾病发展。
为了研究这些研究之间的临床相关性,分析了来自ra患者的cd4+外周t细胞和滑膜cd3+细胞中的anx-al表达。图8c显示了结果。中间数值如水平线指示的,显示了曼-惠特尼检验的p值。*p<0.01。如图8c所示的,与健康对照志愿者(hc)的细胞相比,racd4+细胞表达了高水平的anx-almrna和蛋白(数据未显示)。
还使用绿色和红色荧光标签的二抗血清进行了荧光免疫组织化学,如图8d每组所示的。在ra患者滑膜组织中anx-al表达的免疫组织化学分析揭示了cd3+细胞的高度共区域化。因此,考虑到ra患者的cd4细胞表达更高水平的anx-al,可以得出结论这种蛋白的失调表达可促进该疾病的发展。
实施例7-全长hranx-a1和n末端肽ac2-26对t细胞激活的作用
研究了hranx-a1的n末端肽(肽ac2-26)和全长hranx-a1对t细胞激活的作用。在全长hranx-al(300nm)或anx-al衍生的n-末端肽ac.2-26(l00μm)的存在或缺乏下,用0.6,1.25或2.5μg/mlanti-cd3/cd28刺激鼠类原初淋巴结t细胞中产生的il-2,进行24h。
发现了n-末端肽ac.2-26保持了全长蛋白的大部分生物活性,即增加il-2的产生(图9)和t细胞增殖(数据未显示)。
实施例8-anx-a1和动脉粥样硬化
为了研究在人动脉粥样硬化斑块中是否表达anx-a1,将在颈动脉内膜剥脱手术期间从患者中取出的颈动脉粥样硬化斑块切片用小鼠单克隆抗-人anx-a1抗体(mablb)染色。pepinskyetalfebsletters261:247-252,1990描述了该抗体的制备。简而言之,用处于完全弗氏佐剂中的膜联蛋白-1(是指pepinskyetal中的脂皮质蛋白-1)进行腹膜内注射balb/c小鼠进行免疫。在14和28天对动物增加处于不完全弗氏佐剂中的膜联蛋白-1。6周之后,取出检验血液并筛选阻断膜联蛋白-1活性的抗体。抗血清显示出抗膜联蛋白活性的小鼠脾细胞与sp3×ag8细胞融合产生杂交瘤。杂交瘤培养物用于分析上清液中可沉淀放射标记的膜联蛋白-1的抗体,通过有限稀释将产生沉淀50%以上的输入计数的抗体的杂交瘤进行亚克隆。大部分有希望的品系在降植烷预处理小鼠中生成腹水,并在蛋白a琼脂糖上使用pierce结合和洗脱缓冲体系对单克隆抗体进行亲和纯化。
如图10所示的,在斑块内可观察到对anx-a1的致密且清楚的染色,这证实了渗入这些组织内的炎症表达了高水平的anx-a1。
在apoe-/-小鼠中也进行了相似的分析。通过共聚焦显微镜对10月龄的apoe-/-小鼠的主动脉窦和肱动脉(bca)中的anx-a1进行定位,以确定anx-a1的表达和空间分布。非动脉粥样硬化的动脉组织缺乏免疫反应性的anx-a1(数据未显示)。相反地,主动脉窦和bca的动脉粥样硬化的斑块对anx-a1染色牢固(图11)。
在主动脉窦(图11a)和bca(图11b)的纤维盖附近和在主动脉窦中斑块的坏疽核心附近(图11b)检测到anx-a1的明显免疫反应性。这些结果证实了在人和鼠类动脉粥样硬化斑块中都表达anx-a1,这表明了其表达可有效影响斑块的稳定性。
实施例9-anx-a1和系统性红斑狼疮(sle)
对anx-a1生物学功能的临床研究将针对该蛋白的自身抗体的存在和自身免疫疾病包括系统性红斑狼疮(sle)、类风湿性关节炎和炎性肠病的发展联系起来。鉴于这些发现,我们假定了这些自身抗体的生成可能是由于这些患者体内膜联蛋白-1不受控制的表达。为了证实这个假定,分析了从健康志愿者和sle患者收集的t细胞中膜联蛋白-1的表达水平。在slet细胞中膜联蛋白-1mrna和蛋白的表达水平明显高得多(图12)。因而,这些结果支持了在slet细胞中,和在患有其他自身免疫疾病的患者t细胞中的膜联蛋白-1的表达增加可能负责这些疾病所述的th1细胞因子水平增加,由此表示自身免疫疾病发展的风险因子的假定。
实施例10-通过anti-anx-a1抗体抑制t细胞激活
将纯化的人外周血t细胞与anti-cd3和抗-cd28抗体(5μg/ml)的混合物孵育以激活t细胞受体(tcr):所发生的如图13中通过白介素-2(il-2)的明显产生所证实,其是对t细胞激活和分化关键的细胞因子。
然后将细胞与不同浓度(1.0、0.1、0.01和0.001微克/ml)的针对人重组膜联蛋白1(mabla)产生的中和小鼠单克隆抗体孵育。该抗体的产生如pepinskyetalfebsletters261:247-252,1990和实施例8所描述的。
用mabla处理产生了il-2产生(图13)和细胞增殖(数据未显示)的浓度依赖型抑制。igg可用作在1.0、0.1、0.01和0.001微克/ml浓度的对照且在所有浓度都无效。结果显示了膜联蛋白-1的阻断作用似乎在低浓度的特异性单克隆抗体mabla存在下更有效。
在所有情况中,数据是三次重复测量的平均值±se。*p<0.01
然后将来自不同捐赠者的纯化的人外周血t细胞与不同浓度(1μg/ml)的anti-cd3和抗-cd28抗体混合物孵育以激活t细胞受体(tcr)。再次,所发生的如图14中通过白介素-2(il-2)的产生所证实,但是水平更低,由于所用anti-cd3和抗-cd28抗体的浓度更低。
再次地,将细胞与不同浓度(1.0、0.1、0.01和0.001微克/ml)的针对人重组膜联蛋白-1(mabla)产生的中和小鼠单克隆抗体孵育。用mab1a处理产生il-2产生的浓度依赖型抑制(图14)。igg可用作在1.0微克/ml浓度的对照且无效。此外,结果显示,除了mab1a的1.0微克/ml浓度外,膜联蛋白-1的阻断作用似乎在低浓度的特异性单克隆抗体mabla存在下更有效。
在所有情况中,数据是三次重复测量的平均值±se。*p<0.01
这些实验证实了内源的膜联蛋白1在特异性刺激物的存在下促进t细胞激活。此外,膜联蛋白1途径的阻断对t细胞激活的衰减高达50%。抑制达到约50%的最大值的事实表明了,如所主张的,“正常和持家”免疫不受与anx-a1特异性结合的分子进行治疗的t细胞介导的疾病的影响。
实施例11-anx-a1和多发性硬化症(ms)
材料和方法
试剂
合成髓鞘少突胶质细胞糖蛋白肽(mog)33-55(mevgwyrspfsrvvhlyrngk)并用cambridgeresearchbiochemicals(billingham,uk)纯化。含有结核分枝杆菌(mycobacteriumtuberculosis)h37a的完全弗氏佐剂购买自difco,同时百日咳(bordetellapertussis)毒素来自sigma-aldrichco(poole,uk)。除非另有说明,所有其他试剂都来自sigma-aldrichco。
小鼠
雄性anx-a敲除小鼠之前有描述hannonetal,fasebj,17:253-255,2003;roviezzoetal,j.physiolpharmacol53:541-553,2002)(9-11周龄),在c57bl/6本底回交超过10代,并喂养在b&k动物培养工厂(hull,uk)中。年龄和性别匹配的对照c57bl/6小鼠可用作所有实验的对照。按照英国内政部规定(动物法案1986)和欧盟导则,将动物保持在标准条件下并维持在22±1℃的12h/12h亮/暗循环中。
eae的诱导
在第0天用300μl含有300μgmog35-55的pbs与等体积含有300μg热致死结核分枝杆菌h37ra的cfa的乳液联合皮下免疫小鼠。第0和2天在两侧注射乳液,之后腹膜内注射在100μl盐水中的百日咳毒素(500ng/100μl)。每日观察小鼠的eae体征和体重减轻。疾病严重性评分成6个级别:0=无疾病;1=部分无力的尾部(partialflaccidtail);2=完全无力的尾部;3=后肢张力减退;4=部分后肢麻痹;5=完全后肢瘫痪;6=濒死或死亡动物。
细胞增殖试验
在96孔板中,将从用mog33-55免疫14天的小鼠中获得的淋巴结细胞(105细胞/200μl)用mog33-55(50-100μg/200μl)刺激48h。在最后12h期间,培养物用lμci[3h]-胸苷(amershampharmaciabiotech,buckinghamshire,uk)脉冲,并通过自动闪烁记数器(packardinstrumentcompany,inc.,illinois,us)测量引入的放射活性。
细胞因子elisa
将从用mog33-55免疫14天的小鼠中获得的淋巴结细胞(106细胞/ml)用mog33-55(100μg/ml)刺激4天。收集细胞上清液,并按照生产商说明使用elisa试剂盒(ebioscience,dorset,uk)分析ifn-γ、il-2、il-17a和tnf-α含量。
从脊髓分离炎症细胞
小鼠用co2处死。使用附有21-规格针的注射器通过流体静力压用pbs将脊髓从脊柱中取出。将组织切成小片并使用1ml无菌注射器的活塞使其通过细胞滤器(70nm;bdfalcon)。将单细胞悬浮液在390×g离心10分钟,并重悬在20ml含有30%percoll(sigma)的pbs中,覆盖在10ml含有70%percoll的pbs上。在390×g离心20分钟之后,从中间相取出单核细胞,洗涤,并重悬在facs缓冲液(含有1%fcs和0.02%nan2的pbs)中,用于进一步分析。
流式细胞计数
在4℃,将来自percoll纯化的脊髓组织或ficoll纯化的淋巴结的细胞样品重悬在含有cd16/cd32fcγiir阻断抗体(克隆93;ebioscience)的facs缓冲液30分钟。之后,细胞悬浮液用偶联fitc的anti-cd3(1:100;克隆1452c11)或抗-f4/80(1:100;克隆bmt)进行标记,同时在4℃将淋巴结细胞用抗-cd4-fitc(1:500;克隆l3t4)和抗-cd8(1:1000;克隆ly-2)染色30分钟,之后使用cellquest软件(bectondickinson)通过facscalibur进行分析。每个样品中分析至少104个细胞,根据用无关igg的同种型获得的染色来确定阳性和阴性种群。
组织学
将脊髓组织切开并在4%中性缓冲福尔马林中固定48h,然后与含有edta(0.1mm在pbs中)的脱钙溶液孵育14天,之后进行石蜡包埋。依据疾病严重性,对在不同时间点取样的石蜡包埋切片实施组织学评估。用二甲苯将脊髓切片(5μm)去石蜡化,并用苏木精和伊红(h&e)染色以评估炎症。在冷冻切片上,使用anti-anxal(稀释度1:500;zymed,invitrogen)和偶联抗-兔ig辣根过氧化物酶(hrp)抗体(稀释度1:500;dako)实施对anxa1的染色。如之前所述地,使用偶联fitc的anti-cd3(1:100;克隆1452cl1)或抗-f4/80(1:100;克隆bmt)对anxal和cd3或f4/80进行双重染色。也可用苏木精对切片进行复染色。在所有情况中,每个动物评估最少≥3个切片。使用imagepro图像分析软件包获得相位差数码图像。
统计学分析
使用prism软件(graphpad软件)来运行所有检验。用双尾、非配对样本t检验来实施细胞频率,增殖和细胞因子产生的统计学评估。应用anova分析eae临床等级。<0.05的p值在统计学上被认为是显著的。低于0.05的p-值被认为是显著的。数据表示为每组n个样品的平均值±s.e.m。
结果
anxa1表达与实验性自身免疫脑脊髓炎(eae)的严重性相关联
在用mog35-55免疫诱导ms的小鼠模型eae中评估了脊髓内容物中anxa1水平与cns中单核细胞的渗透程度之间的关联。mog35-55诱导的eae是cns的自身免疫脱髓鞘的模型,其被广泛用于研究负责ms发展的病理机制。为了这个目标,收集在疾病的不同阶段即第12天(0分),第18天(2分)和第20天(4分)的,用mog35-55肽免疫的野生型小鼠的脊髓和脑,并用苏木精和伊红染色一起对anxa1进行免疫组织化学。
如图15所示的,在无病征小鼠的诱导阶段期间收集的脊髓组织显示出对anxa1弱染色(0分,分别是图15a和b)。然而,随着临床体征的开始和渗入cns的炎症的显现,在髓膜四周观察到anxa1免疫染色的分散斑块(2分,分别是图15a和b)。随着疾病的进展,可观察到渗入斑块的anxa1阳性细胞数量增加(4分,分别是图15a和b),这表明表达高水平anxa1的炎症细胞的渗透与疾病的严重性是相关的。
为了辨别脊髓中的anxa1免疫反应性的细胞来源,anti-anxal和anti-cd3(t细胞标记)或抗-f4/80(巨噬细胞标记)对切片实施了双重免疫荧光染色。在eae峰值的小鼠脊髓切片中检测到大量的渗入t细胞和巨噬细胞(分别是图16a和b,中间)。然而,相同切片中的anxa1染色显示出与t细胞和巨噬细胞部分共区域化,而不存在对一种或其他细胞类型的特别偏好(分别是图16a和b,右边组)。
anxa1-/-小鼠发展了受损的eae
因为anxa1表达在eae峰值时上调,所以研究了该蛋白对eae发展的作用。在第0天用在cfa中的mog35-55肽s.c.免疫了anxa1+/+和anxa1-/-小鼠,然后在第0天和第2天i.v.注射百日咳毒素。免疫12天之后anxa1+/+和anxa1-/-小鼠都开始发展eae,约20天达到疾病顶峰。然而,与anxa1+/+相比,anxa1-/-小鼠降低了疾病水平(图17a)。有趣的是,这只在疾病后阶段显现和明显,即第18-23天和之前。
对于eae动物模型的研究已证实了疾病的急性期与体重减轻是同时的,可能是由于食欲减退和缺乏流体摄入。免疫小鼠的体重测量与临床得分的严重性相关联,从开始第18天,与anxa1+/+对照相比,anxa1-/-小鼠显示出体重减轻(图17b)。进一步比较eae在anxa1+/+和anxa1-/-小鼠中的发展显示了在死亡率和疾病最高得分上都降低了,而在发病率或疾病开始中没有区别(表1)。
表1:在anxa1+/+和anxa1-/-小鼠中mog35-55诱导的eae的临床参数(平均值±sem,n=10/组)
**p<0.01,3个实验的代表,§eae临床得分等于或大于1.
anxa1-/-小鼠中对mog35-55的体外回忆应答
t细胞在eae的发展中发挥关键作用,而anxa1-/-t细胞具有应答anti-cd3/cd28刺激的受损能力。鉴于这些发现,研究了在anxa1-/-小鼠中eae的降低发展是否与对抗原刺激的更低应答相关。在体外用mog35-55刺激免疫之后14天收集的来自anxa1+/+和anxa1-/-小鼠淋巴结细胞。与野生型小鼠相比,当用mog35-55刺激时,anxa1-/-淋巴结细胞显示出增殖速率的降低和il-2产生水平的降低(分别是图18a和b)。用脾细胞获得了相似的结果(数据未显示)。
关于细胞增殖的这些结果反映了免疫小鼠的脾脏和引流淋巴结回收的细胞数量。与对照相比,来自相同动物的ficoll纯化脾和淋巴结单核细胞的总细胞计数揭示了在anxa1-/-小鼠中明显降低(分别是图19a和b),而在cd4或cd8阳性细胞百分比上没有可测量到的变化(分别图19c和d)。
在anxa1-/-小鼠中降低的mog35-55特异性th1和th17细胞因子应答
使用来自mog35-55免疫的c57/bl6小鼠的引流淋巴结细胞的研究显示了在th1和th17细胞因子产生上的明显变化。用mog35-55再激发96h的anxa1-/-淋巴结细胞中细胞因子产生的分析显示了,与野生型细胞相比,th1细胞因子ifn-γ,il-2,和tnf-α产生的降低(图20a-c)。相似地,th17标记产物il-17的测量揭示了,与野生型相比,在anxa1-/-中这种细胞因子水平的降低(图20d)。
eae期间anxa1-/-小鼠神经系统中的t细胞渗透
从开始18天anxa1-/-小鼠中减少的eae体征促使我们去研究是否存在神经-病理关联。分析了在第18天或22天收集的anxa1+/+和anxa1-/-处理的小鼠的脊髓用于炎症组织学证据。发现了与anxa1+/+动物相比,在anxa1-/-小鼠中检测到免疫细胞渗透数量的减少。(图21a和b)
在anxa1-/-小鼠中减少的炎症组织学体征与渗透至cns的cd3和f4/80阳性细胞数量的减少相关(分别是图21c和d)。通过facs测量从第18天脊髓组织分离的cd3和f4/80阳性白细胞的百分比,确认了这些定性分析。与免疫组织化学结果一致,与anxa1+/+小鼠相比,anxa1-/-小鼠中具有分别约为60%和80%的t细胞和巨噬细胞的减少(分别是22a和b)。
结果显示了在疾病顶峰时小鼠脊髓中表达膜联蛋白-1的细胞有明显的积累。因此,膜联蛋白-1表达和eae发展之间存在关联。
此外,结果显示了膜联蛋白-1缺陷的小鼠更少发展严重的eae。因此消除膜联蛋白-1表达限制了eae(ms的小鼠模型)的发展。
sequencelisting
<110>玛丽皇后和威斯特-弗尔德学院(queenmary&westfield
college)
<120>能够与膜联蛋白-1结合的特异性结合分子的用途
<130>i44208kst
<150>gb0822011.3
<151>2008-12-02
<160>6
<170>patentinversion3.3
<210>1
<211>25
<212>prt
<213>artificial
<220>
<223>thesequenceissynthesized
<400>1
alametvalserglupheleulysglnalatrppheilegluasnglu
151015
gluglnglutyrvalglnthrvallys
2025
<210>2
<211>346
<212>prt
<213>artificial
<220>
<223>thesequenceissynthesized
<400>2
metalametvalserglupheleulysglnalatrppheilegluasn
151015
glugluglnglutyrvalglnthrvallysserserlysglyglypro
202530
glyseralavalserprotyrprothrpheasnproserseraspval
354045
alaalaleuhislysalailemetvallysglyvalaspglualathr
505560
ileileaspileleuthrlysargasnasnalaglnargglnglnile
65707580
lysalaalatyrleuglngluthrglylysproleuaspgluthrleu
859095
lyslysalaleuthrglyhisleuglugluvalvalleualaleuleu
100105110
lysthrproalaglnpheaspalaaspgluleuargalaalametlys
115120125
glyleuglythraspgluaspthrleuilegluileleualaserarg
130135140
thrasnlysgluileargaspileasnargvaltyrargglugluleu
145150155160
lysargaspleualalysaspilethrseraspthrserglyaspphe
165170175
argasnalaleuleuserleualalysglyaspargsergluaspphe
180185190
glyvalasngluaspleualaaspseraspalaargalaleutyrglu
195200205
alaglygluargarglysglythraspvalasnvalpheasnthrile
210215220
leuthrthrargsertyrproglnleuargargvalpheglnlystyr
225230235240
thrlystyrserlyshisaspmetasnlysvalleuaspleugluleu
245250255
lysglyaspileglulyscysleuthralailevallyscysalathr
260265270
serlysproalaphephealaglulysleuhisglnalametlysgly
275280285
valglythrarghislysalaleuileargilemetvalserargser
290295300
gluileaspmetasnaspilelysalaphetyrglnlysmettyrgly
305310315320
ileserleucysglnalaileleuaspgluthrlysglyasptyrglu
325330335
lysileleuvalalaleucysglyglyasn
340345
<210>3
<211>1041
<212>dna
<213>artificial
<220>
<223>thesequenceissynthesized
<400>3
atggcaatggtatcagaattcctcaagcaggcctggtttattgaaaatgaagagcaggaa60
tatgttcaaactgtgaagtcatccaaaggtggtcccggatcagcggtgagcccctatcct120
accttcaatccatcctcggatgtcgctgccttgcataaggccataatggttaaaggtgtg180
gatgaagcaaccatcattgacattctaactaagcgaaacaatgcacagcgtcaacagatc240
aaagcagcatatctccaggaaacaggaaagcccctggatgaaacactgaagaaagccctt300
acaggtcaccttgaggaggttgttttggctctgctaaaaactccagcgcaatttgatgct360
gatgaacttcgtgctgccatgaagggccttggaactgatgaagatactctaattgagatt420
ttggcatcaagaactaacaaagaaatcagagacattaacagggtctacagagaggaactg480
aagagagatctggccaaagacataacctcagacacatctggagattttcggaacgctttg540
ctttctcttgctaagggtgaccgatctgaggactttggtgtgaatgaagacttggctgat600
tcagatgccagggccttgtatgaagcaggagaaaggagaaaggggacagacgtaaacgtg660
ttcaataccatccttaccaccagaagctatccacaacttcgcagagtgtttcagaaatac720
accaagtacagtaagcatgacatgaacaaagttctggacctggagttgaaaggtgacatt780
gagaaatgcctcacagctatcgtgaagtgcgccacaagcaaaccagctttctttgcagag840
aagcttcatcaagccatgaaaggtgttggaactcgccataaggcattgatcaggattatg900
gtttcccgttctgaaattgacatgaatgatatcaaagcattctatcagaagatgtatggt960
atctccctttgccaagccatcctggatgaaaccaaaggagattatgagaaaatcctggtg1020
gctctttgtggaggaaactaa1041
<210>4
<211>346
<212>prt
<213>artificial
<220>
<223>thesequenceissynthesized
<400>4
metalametvalserglupheleulysglnalatrppheilegluasn
151015
glugluglnglutyrvalglnthrvallysserserlysglyglypro
202530
glyseralavalserprotyrprothrpheasnproserseraspval
354045
alaalaleuhislysalailemetvallysglyvalaspglualathr
505560
ileileaspileleuthrlysargasnasnalaglnargglnglnile
65707580
lysalaalatyrleuglngluthrglylysproleuaspgluthrleu
859095
lyslysalaleuthrglyhisleuglugluvalvalleualaleuleu
100105110
lysthrproalaglnpheaspalaaspgluleuargalaalametlys
115120125
glyleuglythraspgluaspthrleuilegluileleualaserarg
130135140
thrasnlysgluileargaspileasnargvaltyrargglugluleu
145150155160
lysargaspleualalysaspilethrseraspthrserglyaspphe
165170175
argasnalaleuleuserleualalysglyaspargsergluaspphe
180185190
glyvalasngluaspleualaaspseraspalaargalaleutyrglu
195200205
alaglygluargarglysglythraspvalasnvalpheasnthrile
210215220
leuthrthrargsertyrproglnleuargargvalpheglnlystyr
225230235240
thrlystyrserlyshisaspmetasnlysvalleuaspleugluleu
245250255
lysglyaspileglulyscysleuthralailevallyscysalathr
260265270
serlysproalaphephealaglulysleuhisglnalametlysgly
275280285
valglythrarghislysalaleuileargilemetvalserargser
290295300
gluileaspmetasnaspilelysalaphetyrglnlysmettyrgly
305310315320
ileserleucysglnalaileleuaspgluthrlysglyasptyrglu
325330335
lysileleuvalalaleucysglyglyasn
340345
<210>5
<211>204
<212>prt
<213>artificial
<220>
<223>thesequenceissynthesized
<400>5
metasnleuileleuargtyrthrpheserlysmetalametvalser
151015
glupheleulysglnalatrppheilegluasnglugluglnglutyr
202530
valglnthrvallysserserlysglyglyproglyseralavalser
354045
protyrprothrpheasnproserseraspvalalaalaleuhislys
505560
alailemetvallysglyvalaspglualathrileileaspileleu
65707580
thrlysargasnasnalaglnargglnglnilelysalaalatyrleu
859095
glngluthrglylysproleuaspgluthrleulyslysalaleuthr
100105110
glyhisleuglugluvalvalleualaleuleulysthrproalagln
115120125
pheaspalaaspgluleuargalaalametlysglyleuglythrasp
130135140
gluaspthrleuilegluileleualaserargthrasnlysgluile
145150155160
argaspileasnargvaltyrargglugluleulysargaspleuala
165170175
lysaspilethrseraspthrserglyasppheargasnalaleuleu
180185190
serleualalysglyaspargsergluaspphegly
195200
<210>6
<211>115
<212>prt
<213>artificial
<220>
<223>thesequenceissynthesized
<400>6
metalametvalserglupheleulysglnalatrppheilegluasn
151015
glugluglnglutyrvalglnthrvallysserserlysglyglypro
202530
glyseralavalserprotyrprothrpheasnproserseraspval
354045
alaalaleuhislysalailemetvallysglyvalaspglualathr
505560
ileileaspileleuthrlysargasnasnalaglnargglnglnile
65707580
lysalaalatyrleuglngluthrglylysproleuaspgluthrleu
859095
lyslysalaleuthrglyhisleuglugluvalvalleualaleuleu
100105110
lysthrpro
115
- 还没有人留言评论。精彩留言会获得点赞!