结合OX40的抗体及其用途的制作方法
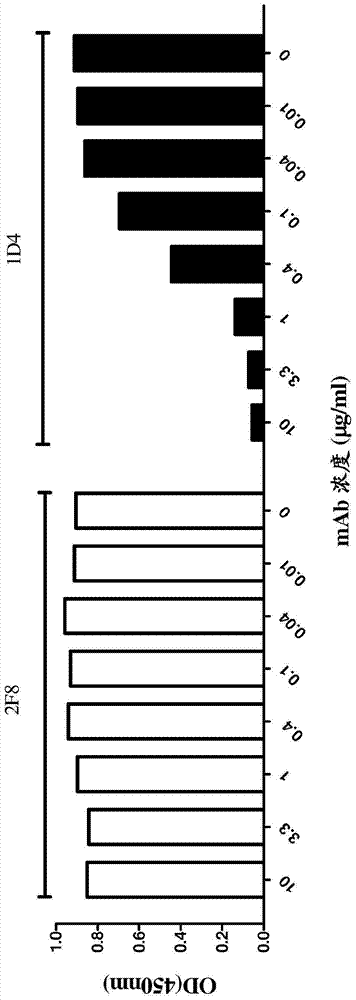
结合OX40的抗体及其用途相关申请本申请要求于2011年7月11日提交的美国临时申请号61/506,491的权利,其全部内容通过引用全文整合到本文中。发明领域本发明涉及结合人OX40的拮抗剂抗体或其片段。更具体而言,本发明涉及结合人OX40的拮抗剂抗体或其片段,其包括包含SEQIDNO:1的氨基酸序列的重链CDR1,和/或包含SEQIDNO:2的氨基酸序列的重链CDR2,和/或包含SEQIDNO:3的氨基酸序列的重链CDR3;和/或包括包含SEQIDNO:4的氨基酸序列的轻链CDR1,和/或包含SEQIDNO:5的氨基酸序列的轻链CDR2,和/或包含SEQIDNO:6的氨基酸序列的轻链CDR3。发明背景OX40是TNFR受体超家族的成员,并在1987年首次鉴别为在大鼠的活化CD4+T细胞上表达的50kDa糖蛋白(PatersonDJ等人,(1987)Mol.Immunol.24:1281-90)。OX40的胞外配体结合结构域由3个完整的富含半胱氨酸的结构域(CRD)和第4个部分的C-末端CRD组成(BodmerJL等人,(2002)TrendsBiochem.Sci.27:19-26)。OX40的配体是OX40L(CD252),3拷贝的OX40结合3聚体的配体,形成OX40-OX40L复合物(CompaanDM&HymowitzSG(2006)Structure,14:1321-1330)。OX40是膜结合型受体;然而也检测到了可溶性异构体(TaylorL&SchwarzH(2001)J.Immunol.Methods,255:67-72)。与CD28不同,OX40不是在新生T细胞上组成型表达的,而是在T细胞受体(TCR)结合(engagement)后诱导的。OX40是次级共刺激分子,在激活后24至72小时表达;它的配体OX40L也不在静息的抗原呈递细胞上表达,而在所述细胞激活后激活。OX40主要由激活的CD4+T细胞表达,以及由激活的CD8+T细胞有限程度的表达(Salek-ArdakaniS等人,(2006)Curr.Immunol.Rev.2:37-53)。发明概述本发明一般涉及结合人OX40的拮抗剂抗体或其片段,其制备和使用的方法,包括治疗OX40介导的病症的方法。本发明的结合人OX40的拮抗剂抗体或其片段是拮抗型抗体,不表现出激动剂效应和/或在结合后不激活人OX40。在一个方面,本发明提供了结合人OX40的拮抗剂抗体或其片段,其包括包含SEQIDNO:1的氨基酸序列的重链CDR1,和/或包含SEQIDNO:2的氨基酸序列的重链CDR2,和/或包含SEQIDNO:3的氨基酸序列的重链CDR3;和/或包括包含SEQIDNO:4的氨基酸序列的轻链CDR1,和/或包含SEQIDNO:5的氨基酸序列的轻链CDR2,和/或包含SEQIDNO:6的氨基酸序列的轻链CDR3。在另一个方面,本发明提供了结合人OX40的拮抗剂抗体或其片段,其包括包含SEQIDNO:7的氨基酸序列的重链可变区序列。在另一个方面,本发明提供了结合人OX40的拮抗剂抗体或其片段,其包括这样的重链可变框架区,所述重链可变框架区是选自IGHV2-70*10(SEQIDNO:19)、IGHV2-70*01(SEQIDNO:20)、IGHV2-70*13(SEQIDNO:21)、IGHV2-5*09(SEQIDNO:22)和IGHV2-70*11(SEQIDNO:23)的人基因的产物或源自所述人基因。在另一个方面,本发明提供了拮抗剂抗体或其片段,其包括包含SEQIDNO:32的氨基酸序列的重链序列,其中重链可变框架区包括相对于相应小鼠抗体的相应重链可变框架区的至少一个氨基酸修饰。在另一个方面,本发明提供了结合人OX40的拮抗剂抗体或其片段,其包括包含SEQIDNO:8的氨基酸序列的轻链可变区序列。在另一个方面,本发明提供了结合人OX40的拮抗剂抗体或其片段,其包括这样的轻链可变框架区,所述轻链可变框架区是选自IGKV3-11*01(SEQIDNO:24)、IGKV1-39*01(SEQIDNO:25)、IGKV1D-39*01(SEQIDNO:26)、IGKV3-11*02(SEQIDNO:27)和IGKV3-20*01(SEQIDNO:28)的人基因的产物或源自所述人基因。在另一个方面,本发明提供了拮抗剂抗体或其片段,其包括这样的轻链可变框架区,所述轻链可变框架区是IGKV3-11*01(SEQIDNO:24)的人基因的产物或源自所述人基因,其中轻链可变框架区包括相对于相应小鼠抗体的轻链可变区的相应框架区的至少一个氨基酸修饰。在另一个方面,本发明提供了结合人OX40的拮抗剂抗体或其片段,其包括选自SEQIDNO:32、33、34、35、36、37和38的重链序列。在另一个方面,本发明提供了结合人OX40的拮抗剂抗体或其片段,其包括选自SEQIDNO:39、40、41、42、43、44、45、46、47、48和49的轻链序列。在另一个方面,本发明提供了结合人OX40的拮抗剂抗体或其片段,其包括:(a)包含SEQIDNO:37或38的氨基酸序列的重链序列;和(b)包含SEQIDNO:47的氨基酸序列的轻链序列。在另一个方面,本发明提供了结合人OX40的拮抗剂抗体或其片段,其包括包含选自SEQIDNO:58、59、79和80的氨基酸序列的重链可变区。在另一个方面,本发明提供了结合人OX40的拮抗剂抗体或其片段,其包括包含选自SEQIDNO:60、86、87和89的氨基酸序列的轻链可变区。在另一个方面,本发明提供了结合人OX40的拮抗剂抗体或其片段,其包括:(a)包含SEQIDNO:58或59的氨基酸序列的重链可变区;和(b)包含SEQIDNO:60的氨基酸序列的轻链可变区。在另一个方面,本发明提供了结合人OX40的拮抗剂抗体或其片段,其中抗体包含人IgG4Fc区,其中抗体不具有Fc介导的细胞毒活性。在另一个方面,本发明提供了结合人OX40的拮抗剂抗体或其片段,其中抗体包含人IGHG1Fc区,其中抗体具有细胞毒机制,例如抗体依赖性细胞毒作用(ADCC)。在优选的方面,结合人OX40的拮抗剂抗体或其片段具有无岩藻糖基化的IGHG1Fc区,并表现出增强的Fc介导的细胞毒机制,如ADCC。在另一个方面,本发明还描述了以与相应嵌合抗体类似的亲和力结合人OX40的拮抗型人源化抗体或其片段,例如保留了相应嵌合抗体的至少75%的OX40结合亲和力(KD),或与相应嵌合抗体比较时,具有至少等价的或更高的OX40结合亲和力(KD)。在另一个方面,本发明提供了这样的拮抗剂抗体或其片段,其结合位于人OX40胞外区的第二结构域中的表位。本发明还提供了编码结合人OX40的抗体及其片段的分离的核酸、载体,和包含所述核酸或载体的宿主细胞。还提供了包含所述拮抗剂抗体或其片段以及可药用载体的组合物,和包括与治疗剂连接的所述拮抗剂抗体或其片段的免疫缀合物。本发明还提供了治疗OX40介导的病症的方法。在一个方面,在同种异体活化的T细胞激活和增殖的体外模型(混合型淋巴细胞反应;MLR)中,拮抗剂抗体或其片段有效的抑制了两个不同个体(应答者)的MLR,EC50值是约100ng/mL。此外,在异种移植物抗宿主反应中,拮抗型抗体或其片段有力的抑制了GVHD反应,所述反应是在人类患者中移植骨髓后观察到的同种异体移植物抗宿主病(GVHD)的模型。本发明还提供了试剂盒和制品,其包括所述抗体或其片段、用于治疗OX40介导的病症的组合物或免疫缀合物。附图简介图1:(A)固定化重组人OX40-his的直接结合ELISA。通过直接ELISA测量嵌合2F8和1D4抗体与人OX40的结合。在96孔板中,4℃孵育各浓度(10至0.01mg/ml)的1D4(黑色直方图)和2F8(白色直方图)与2mg/ml包被的重组人OX40-his标记的蛋白质过夜。通过辣根过氧化物酶(HRP)缀合的抗人抗体,检测每种抗体与OX40的结合。(B)固定化重组人OX40-Fc的竞争性ELISA。通过阻断ELISA评估嵌合1D4和2F8对OX40/OX40L相互作用的抑制效应。在96孔板中,4℃孵育各浓度(10至0.01mg/ml)的1D4(黑色直方图)和2F8(白色直方图)与2mg/ml包被的重组人OX40-Fc标记的蛋白质过夜。5分钟后,向每个孔添加固定浓度的生物素化重组人OX40L(0.04mg/ml),并在室温孵育30分钟。使用链霉抗生物素-HRP检测OX40L与OX40的结合。图2:通过3H胸苷整合测量的单向混合淋巴细胞反应(MLR)。直条显示了至少3次重复的平均3H胸苷整合(计数)±平均值的标准误。显示了同种型对照(曲妥珠单抗)和阳性对照(依法珠单抗)。效应子仅代表效应子细胞。效应子+靶代表了省略抗体时的测量值。图3:嵌合1D4抗体的流式细胞仪分析。(A)对人活化的外周血单核细胞(PBMC)和HPB-ALL细胞的染色。直方图曲线(Histogramplots)显示了荧光强度(X轴)和相对细胞数(%最大事件——Y轴)。标出了染色的细胞类型。在测量前,用PHA和IL-2激活人PBMC持续48h。(B)对活化的食蟹猴(cynomologus)PBMC染色。通过流式细胞仪评估嵌合1D4抗体与食蟹猴OX40的结合。从收集自食蟹猴的全血中分离外周血单核细胞(PBMC),在存在10mg/mlPHA和100U/mlrhuIL-2的条件下,培养3x106细胞50小时。用25mg/ml的对照抗体(上图(i))或生物素化的绵羊抗人OX40抗体(中图(ii))或生物素化嵌合1D4抗体(下图(iii))孵育活化的PBMC。用链霉抗生物素-APC检测每种抗体与食蟹猴OX40的结合。图4:表面等离振子共振测量抗OX40抗体。数据表示为应答数(缩写为RU;Y轴)vs时间(X轴)。图4A-VH1/VL1抗体vs1D4嵌合体。图4B-基于VH1、VH2和VH3的人源化抗体(如所示)vs1D4嵌合体。图4C-低结合子的例子:VH4/VL4、VH5/VL4、VH5/VL5和VH5/VL6。图4D-弱结合子(VH5/VL9和VH4/VL9)和良好结合子(VH6/VL9和VH7/VL9)的例子。图4E-基于VH7的人源化抗体。图4F-VH6/VL9具有超过1D4嵌合体和人源化变体VH7/VL9的最佳结合特性。图5:序列比对。1D4的重链(图5A)或轻链(图5B)可变区与选定的IMGT种系框架(IGHV2-70*10(SEQIDNO:19)和IGKV3-11*01(SEQIDNO:24))和回复突变的可变区变体的比对,所述回复突变的可变区变体是VH1(SEQIDNO:29)、VH2(SEQIDNO:77)、VH3(SEQIDNO:78)、VH4(SEQIDNO:79)、VH5(SEQIDNO:80)、VH6(SEQIDNO:58)、VH7(SEQIDNO:59)、VL1(SEQIDNO:30)、VL2(SEQIDNO:81)、VL3(SEQIDNO:82)、VL4(SEQIDNO:83)、VL5(SEQIDNO:84)、VL6(SEQIDNO:85)、VL7(SEQIDNO:86)、VL8(SEQIDNO:87)、VL9(SEQIDNO:60)、VL10(SEQIDNO:88)、VL11(SEQIDNO:89)。图6:使用差异扫描量热法,测量人源化抗OX40抗-抗体VH6/VL9FAB片段的热稳定性。数据表示为过量摩尔热容(缩写为Cp[kcal/mol/℃];Y轴)vs温度(X轴)。图7:表位表征。该图显示了人源化抗OX40抗-抗体VH6/VL9表位,其是基于实施例7所述的ELISA测定结果的。图8:通过3H胸苷整合测量的混合淋巴细胞反应(MLR)。图8A和8B显示了来自两个不相关供体的混合淋巴细胞反应的结果。通过3H胸苷整合测量增殖。图表显示了每种条件的绝对计数值±SEM。应答者细胞是未处理的PBMC,刺激子细胞是丝裂霉素-处理的PBMC。用混合有异源刺激子PBMC的应答者细胞进行含测试抗体的所有条件。阳性对照是依法珠单抗(抗LFA-1抗体)。图9:异种移植物vs宿主反应模型。该图显示了每个提及条件的8只动物的组内百分比存活。载体:仅PBS。垂直虚线表示了最后一天治疗。在未接受PBMC的2只照射过的对照动物的组中没有观察到死亡率和症状(未显示)。发明详述本发明涉及结合人OX40的拮抗剂抗体及其片段。本文中使用的术语―人OX40”包括人OX40的变体、同种型和物种同源物。因此,在某些情况下,本发明的抗体可与人以外的物种的OX40交叉反应。在某些实施方案中,抗体可以完全对一种或多种人OX40蛋白特异并且可以不表现出物种或其它类型的非人交叉反应。示例性的人OX40的完整氨基酸序列具有Swiss-Prot登录号P43489(TNR4_HUMAN;SEQIDNO:12)。OX40也被称为CD134、TNFRSF4、ACT35或TXGP1L。人OX40被EntrezGene称为GeneID:7293,被HGNC称为HGNC:11918。OX40可以被称为CD134(134分化簇)。OX40可以由被称为TNFRSF4/OX40的基因编码。本文中―人OX40”的使用涵盖了人OX40的所有已知的或仍未被发现的等位基因和多态形式。如未另外具体指出,则术语―人OX40”、―OX40”或―OX40受体‖在本文中是等价使用的,意指―人OX40”。术语―OX40配体‖或―OX40L”在本文中是等价使用的,包括OX40配体,尤其是人OX40配体。OX40L是TNF超家族的成员,也被称为gp34或CD252。OX40L也被称为CD252(252分化簇),具有序列数据库登录号P23510(Swiss-Prot)或Q6FGS4(Uniprot)。OX40L在活化的B细胞、T细胞、树突状细胞和内皮细胞的表面表达。本文中使用的术语―结合人OX40的抗体或其片段”包括结合人OX40(例如分离形式的人OX40)的抗体或其片段,其亲和力(KD)为500nM或更低,优选200nM或更低,更优选150nM或更低,更优选120nM或更低,甚至更优选110nM或更低。术语―结合人OX40的抗体或其片段”包括抗体或其抗原结合片段。术语―拮抗型抗体‖或―拮抗剂抗体‖在本文中是等价使用的,包括能够抑制和/或中和OX40的生物学信号传递活性的抗体,例如通过阻断OX40与OX40配体的结合或实质上降低OX40与OX40配体的结合,因此抑制或降低由OX40触发的信号传递通路,和/或抑制或降低OX40介导的细胞应答样淋巴细胞增殖、细胞因子表达或淋巴细胞存活。本文中使用的术语―抗体‖包括完整的抗体和其任何抗原结合片段或单链。―抗体‖指包括通过二硫键相互连接的至少两条重链(H)和两条轻链(L)的糖蛋白,或其抗原结合片段。每条重链都包括重链可变区(本文中缩写为VH)和重链恒定区。重链恒定区包括三个结构域CH1、CH2和CH3。每条轻链都包括轻链可变区(本文中缩写为VL)和轻链恒定区。轻链恒定区包括一个结构域CL。VH和VL区可以进一步细分为被称为互补决定区(CDR)的超变区,其序列超变和/或参与抗原识别和/或通常形成结构性限定的环,分散在被称为框架区(FR或FW)的较保守的区域中。每个VH和VL都包括3个CDR和4个FW,按下列顺序从氨基端向羧基端排列:FW1、CDR1、FW2、CDR2、FW3、CDR3、FW4。FW1、FW2、FW3和FW4的氨基酸序列都构成本文所述VH或VL的“非CDR区‖或―未延伸的CDR区”。本文中使用的术语“重链可变框架区‖可包括一个或多个(例如,1、2、3和/或4个)重链框架区序列(例如,框架区1(FW1)、框架区2(FW2)、框架区3(FW3)和/或框架区4(FW4))。优选的,重链可变框架区包括FW1、FW2和/或FW3,更优选FW1、FW2和FW3。本文中使用的术语“轻链可变框架区‖可包括一个或多个(例如,1、2、3和/或4个)轻链框架区序列(例如,框架区1(FW1)、框架区2(FW2)、框架区3(FW3)和/或框架区4(FW4))。优选的,轻链可变框架区包括FW1、FW2和/或FW3,更优选FW1、FW2和FW3。重链和轻链的可变区含有与抗原相互作用的结合结构域。抗体的恒定区可介导免疫球蛋白与宿主组织或因子的结合,包括免疫系统的各种细胞(例如,效应子细胞)和经典补体系统的第一组分(C1q)。根据恒定区遗传确定,将抗体分类,也被称为同种型。人的恒定轻链分为κ(Cκ)和λ(Cλ)轻链。重链分为μ、δ、γ、α或ε,并将抗体同种型分别定义为IgM、IgD、IgG、IgA和IgE。因此,本文中使用的“同种型‖意指通过其恒定区的化学和抗原特征定义的免疫球蛋白的任何类别和/或亚类。已知的人免疫球蛋白同种型是IgG1(IGHG1)、IgG2(IGHG2)、IgG3(IGHG3)、IgG4(IGHG4)、IgA1(IGHA1)、IgA2(IGHA2)、IgM(IGHM)、IgD(IGHD)和IgE(IGHE)。所谓的人免疫球蛋白假γIGHGP基因代表了额外的人免疫球蛋白重链恒定区基因,该额外的基因已被测序,由于改变的转换区而不编码蛋白质(BensmanaM等人,(1988)NucleicAcidsRes.16(7):3108)。尽管具有改变的转换区,人免疫球蛋白假γIGHGP基因具有所有的重链恒定结构域(CH1-CH3)的和铰链区开放阅读框。其重链恒定结构域的所有的开放阅读框编码与具有预测的结构特征的所有人免疫球蛋白恒定结构域良好比对的蛋白质结构域。该额外的假γ同种型在本文中被称为IgGP或IGHGP。已报道了其他的假免疫球蛋白基因,例如人免疫球蛋白重链恒定结构域εP1和P2假基因(IGHEP1和IGHEP2)。IgG类是最常用于治疗目的的。在人中,该类别包括IgG1、IgG2、IgG3和IgG4亚类。在小鼠中,该类别包括亚类IgG1、IgG2a、IgG2b、IgG2c和IgG3。本文中使用的术语“嵌合抗体‖包括这样的抗体,其中可变区序列源自一个物种,而恒定区序列源自另一个物种,例如可变区序列源自小鼠抗体,而恒定区序列源自人抗体的抗体。本文中使用的术语“人源化抗体‖或―人源化抗OX40抗体‖包括这样的抗体,其中源自另一种哺乳动物物种(例如小鼠)的种系的CDR序列被移植到人的框架序列上。在人的框架序列和源自另一种哺乳动物物种的种系的CDR序列中也可以进行额外的框架区修饰。本文中使用的术语―Fab”或―Fab区‖包括包含VH、CH1、VL和CL免疫球蛋白结构域的多肽。Fab可以指分离的该区域,或者位于全长抗体或抗体片段环境中的该区域。本文中使用的术语―Fc”或―Fc区‖包括包含除第一恒定区免疫球蛋白结构域以外的抗体恒定区的多肽。因而,Fc指IgA、IgD和IgG的最后两个恒定区免疫球蛋白结构域,和IgE和IgM的最后三个恒定区免疫球蛋白结构域,和这些结构域N端的柔性铰链。对于IgA和IgM,Fc可包括J链。对于IgG,Fc包括免疫球蛋白结构域Cγ2和Cγ3(Cγ2和Cγ3),和在Cγ1(Cγ1)和Cγ2(Cγ2)之间的铰链。虽然Fc区的边界可以改变,但人IgG重链Fc区通常定义为包含残基C226或P230至其羧基端,其中编号是根据EU编号系统。对于人IgG1,Fc区在本文定义为包含残基P232至其羧基端,其中编号是根据EU编号系统(EdelmanGM等人,(1969)ProcNatlAcadSciUSA,63(1):78-85)。Fc可以指分离的该区域,或者位于Fc多肽(例如抗体)环境中的该区域。本文中的术语―铰链‖或―铰链区‖或―抗体铰链区‖包括包含在抗体的第一和第二恒定结构域之间的氨基酸的柔性多肽。本文所述的―铰链区‖是长度为6-62个氨基酸的序列区,仅存在于IgA、IgD和IgG中,涵盖了桥接两条重链的半胱氨酸。结构上,IgGCH1结构域终止于EU220位,IgGCH2结构域始于残基EU237位。因而,对于IgG,本文中抗体铰链定义为包括221(IgG1的D221)至231(IgG1的A231)位,其中编号是根据EU编号系统(EdelmanGM等人,见上文)。本文中使用的术语―亲本抗体‖或―亲本免疫球蛋白‖包括未修饰的抗体,所述抗体之后经修饰产生变体。所述亲本抗体可以是天然存在的抗体,或者天然存在的抗体的变体或改造版本。亲本抗体可以指抗体本身,包含所述亲本抗体的组合物,或其编码氨基酸序列。本文中使用的―亲本抗OX40抗体‖意指结合人OX40的抗体或免疫球蛋白,并经修饰产生变体。本文中使用的―相应小鼠抗体‖意指结合人OX40的小鼠抗体或免疫球蛋白,可以经修饰生成变体,尤其是本文公开的小鼠抗体1D4。本文中使用的术语―变体抗体‖或―抗体变体‖包括由于相比亲本的至少一个氨基酸修饰,而不同于亲本抗体序列的抗体序列。本文中的变体抗体序列优选的具有与亲本抗体序列至少约80%,最优选至少约90%,更优选至少约95%的氨基酸序列同一性。抗体变体可以指抗体本身,包含所述抗体变体的组合物,或编码其的氨基酸序列。本文中的术语―氨基酸修饰‖包括在多肽序列中的氨基酸取代、插入和/或缺失。本文中的―氨基酸取代‖或―取代‖意指用另一种氨基酸替换亲本多肽序列中特定位置上的氨基酸。例如,取代R94K指这样的变体多肽,其中94位的精氨酸被赖氨酸替换,此情况为重链可变框架区变体。对于前例,94K表示用赖氨酸取代94位。出于本文的目的,通常通过斜杠分隔多个取代。例如,R94K/L78V指包含取代R94K和L78V的双变体。本文中使用的“氨基酸插入‖或―插入‖意指在亲本多肽序列中的特定位置添加氨基酸。例如,插入-94表示在94位的插入。本文中使用的―氨基酸缺失‖或―缺失‖意指去除亲本多肽序列中特定位置上的氨基酸。例如R94-表示缺失94位的精氨酸。本文中使用的术语―保守修饰‖或―保守序列修饰‖意指不显著影响或改变含有所述氨基酸序列的抗体的结合特征的氨基酸修饰。此类保守修饰包括氨基酸取代、插入和缺失。可通过本领域已知的标准技术将修饰导入本发明的抗体中,例如定点诱变和PCR介导的诱变。保守的氨基酸取代是用具有相似侧链的氨基酸残基替换氨基酸残基的取代。本领域已经定义了具有相似侧链的氨基酸残基家族。这些家族包括含碱性侧链的氨基酸(例如,赖氨酸、精氨酸、组氨酸)、酸性侧链(例如、天冬氨酸、谷氨酸)、不带电的极性侧链(例如,甘氨酸、天冬酰胺、谷氨酰胺、丝氨酸、苏氨酸、酪氨酸、半胱氨酸、色氨酸)、非极性侧链(例如,丙氨酸、缬氨酸、亮氨酸、异亮氨酸、脯氨酸、苯丙氨酸、甲硫氨酸)、β分支侧链(例如,苏氨酸、缬氨酸、异亮氨酸)和芳香族侧链(例如,酪氨酸、苯丙氨酸、色氨酸、组氨酸)。因而,可以用其他相同侧链家族的氨基酸残基替换本发明抗体的CDR区中或框架区中的一个或多个氨基酸残基,并可以测试所改变的抗体(变体抗体)保留的功能。对于所有的人免疫球蛋白重链恒定结构域,都是根据“EU编号系统‖编号(EdelmanGM等人,(1969)ProcNatlAcadSciUSA,63(1):78-85)。对于人κ免疫球蛋白轻链恒定结构域(IGKC),根据“EU编号系统”编号(Edelman等人,见上文)。对于人λ免疫球蛋白轻链恒定结构域(IGLC1、IGLC2、IGLC3、IGLC6和IGLC7),根据“Kabat编号系统‖编号(KabatEA等人,(1991)Sequencesofproteinsofimmunologicalinterest.,第5版,USDepartmentofHealthandHumanServices,NIH公开号91-3242),如DariavachP等人,(1987)ProcNatlAcadSciUSA,84(24):9074-8和FrangioneB等人,(1985)ProcNatlAcadSciUSA,82(10):3415-9所述。术语―可变结构域‖指介导抗原结合和定义特定抗体与特定抗原特异性的结构域。在天然存在的抗体中,抗原结合位点由2个定义特异性的可变结构域组成:1个位于重链(VH),另一个位于轻链(VL)。在一些情况下,特异性可专门位于2个结构域中的1个结构域中,如可见于骆驼科的重链抗体中的单结构域抗体。V区长度通常是约110个氨基酸,由15-30个氨基酸的、被称为框架区(FR)的相对不变的氨基酸序列区段和长度9-12个氨基酸的、被称为―超变区‖的非常易变的较短区域组成,所述框架区被―超变区‖分割开。天然重链和轻链的可变结构域包括4个大部分采用β片层构象的FR,通过3个形成环的超变区连接。由FR将每条链中的超变区与其他链的超变区紧密的保持在一起,有助于形成抗体的抗原结合位点(参见KabatEA等人,见上文)。本文中使用的术语―超变区‖指负责抗原结合的抗体的氨基酸残基。超变区一般包括来自―互补决定区‖或―CDR‖的氨基酸残基,后者是最高的序列可变性和/或参与抗原识别。根据Kabat(KabatEA等人,见上文)对所有的可变结构域编号。本文使用和涵盖了多个CDR定义。Kabat定义是基于序列可变性的并是最常用的定义(KabatEA等人,见上文)。Chothia相反指结构环的位置(ChothiaC&LeskAM(1987)J.Mol.Biol.196:901-917)。AbM定义是Kabat和Chothia定义之间的妥协,由OxfordMolecular的AbM抗体建模软件使用(MartinACR等人,(1989)Proc.NatlAcad.Sci.USA,86:9268–72;MartinACR等人,(1991)MethodsEnzymol.203:121–153;PedersenJT等人,(1992)Immunomethods,1:126–136;ReesAR等人,(1996)InSternbergM.J.E.(编著),ProteinStructurePrediction.OxfordUniversityPress,Oxford,141–172)。接触定义是最近引入的(MacCallumRM等人,(1996)J.Mol.Biol.262:732-745),基于对ProteinDatabank中可利用的复杂结构的分析。(theinternationalImMunoGeneTicsinformation)对CDR的定义(http://www.imgt.org)是基于对所有种类的所有免疫球蛋白和T细胞受体V区的IMGT编号的(theinternationalImMunoGeneTicsinformationLefrancMP等人,(1991)NucleicAcidsRes.27(1):209-12;RuizM等人,(2000)NucleicAcidsRes.28(1):219-21;LefrancMP(2001)NucleicAcidsRes.29(1):207-9;LefrancMP(2003)NucleicAcidsRes.31(1):307-10;LefrancMP等人,(2005)Dev.Comp.Immunol.29(3):185-203;KaasQ等人,(2007)BriefingsinFunctionalGenomics&Proteomics,6(4):253-64)。本发明中讨论的所有互补决定区(CDR)优选是根据定义的。上述CDR中每一个的可变结构域残基如下(根据KabatEA等人编号,见上文):LCDR1:27-32;LCDR2:50-52;LCDR3:89-97;HCDR1:26-35;HCDR2:51-57和HCDR3:93-102。本文中使用的VL区的―非CDR区‖包括氨基酸序列:1-26(FR1)、33-49(FR2)、53-88(FR3)和98-约107(FR4)。本文中使用的VH区的―非CDR区‖包括氨基酸序列:1-25(FR1)、36-50(FR2)、58-92(FR3)和103-约113(FR4)。本发明的CDR可包括基于上述定义的―延伸的CDR”,并具有如下的可变结构域:LCDR1:24-36、LCDR2:46-56、LCDR3:89-97、HCDR1:26-36、HCDR2:47-65、HCDR3:93-102。这些延伸的CDR是根据Kabat等人,见上文编号的。本文使用的VL区的―非延伸CDR区‖包括氨基酸序列:1-23(FR1)、37-45(FR2)、57-88(FR3)和98-约107(FR4)。本文使用的VH区的―非延伸CDR区‖包括氨基酸序列:1-25(FR1)、37-46(FR2)、66-92(FR3)和103-约113(FR4)。本文使用的术语―全长抗体‖包括构建天然生物学形态的抗体的结构,包括可变区和恒定区。例如,在大部分哺乳动物中,包括人和小鼠,IgG类的全长抗体是四聚体,并由2对相同的两条免疫球蛋白链组成,每对具有一条轻链和一条重链,每条轻链包含免疫球蛋白结构域VL和CL,每条重链包括免疫球蛋白结构域VH、CH1(Cγ1)、CH2(Cγ2)和CH3(Cγ3)。在一些哺乳动物中,例如骆驼和羊驼(llama),IgG抗体可以仅由两条重链组成,每条重链包含与Fc区连接的可变结构域。抗体片段包括但不限于:(i)由VL、VH、CL和CH1结构域组成的Fab片段,包括Fab’和Fab’-SH,(ii)VH和CH1结构域组成的Fd片段,(iii)由单个抗体的VL和VH结构域组成的Fv片段;(iv)由单个可变区组成的dAb片段(Ward等人,1989,Nature341:544-546);(v)F(ab')2片段,包含2个连接的Fab片段的二价片段;(vi)单链Fv分子(scFv),其中VH结构域和VL结构域通过肽接头连接,所述接头允许两个结构域关联形成抗原结合位点(Bird等人,1988,Science242:423-426;HustonJS等人,1988,Proc.Natl.Acad.Sci.U.S.A.85:5879-5883);(vii)双特异性单链Fv二聚体(PCT/US92/09965);(viii)―双抗体‖或―三链抗体‖,通过基因融合构建的多价或多特异性片段(TomlinsonI&HollingerP,2000,MethodsEnzymol.326:461-479;WO94/13804;Holliger等人,1993,Proc.Natl.Acad.Sci.U.S.A.90:6444-6448);和(ix)与相同或不同抗体遗传融合的scFv(Coloma&Morrison,1997,NatureBiotechnology15,159-163)。本文中使用的术语―效应子功能‖包括由抗体Fc区与Fc受体或配体相互作用导致的生物化学事件。效应子功能包括FcγR-介导的效应子功能,例如ADCC(抗体依赖性细胞介导的细胞毒作用)和ADCP(抗体依赖性细胞介导的吞噬作用),和补体介导的效应子功能,例如CDC(补体依赖性细胞毒作用)。可以通过改变(即,增强或降低),优选增强抗体与效应子分子(例如Fc受体或补体组分)的亲和力,改变抗体的效应子功能。一般通过修饰效应子分子结合位点来改变结合亲和力,在此情况下,恰当的是以合适的方式定位目标位点和修饰至少一部分位点。还可以设想抗体与效应子分子的结合位点中的改变不必显著改变整体的结合亲和力,可以改变相互作用的几何形状,使效应子机制在非生产性结合中无效。还可以设想通过修饰不直接参与效应子分子结合而参与效应子功能实施的位点,改变效应子功能。通过改变抗体的效应子功能,可以控制免疫应答的多个方面,例如增强或抑制免疫系统的多种反应,对诊断和治疗具有潜在的有益效果。本文中使用的术语―OX40介导的病症‖包括例如变态反应、哮喘、COPD、类风湿关节炎、牛皮癣和与自身免疫和炎症相关的适应症。本文中使用的术语―对象‖包括任何人或非人动物。术语“非人动物‖包括所有的脊椎动物,例如哺乳动物和非哺乳动物,例如非人灵长类、绵羊、狗、猫、马、牛、鸡、两栖类、爬行类等。优选的对象是人。抗OX40抗体在第一个方面,本发明提供了结合人OX40的拮抗剂抗体或其片段,其包括包含SEQIDNO:1的氨基酸序列的重链CDR1,和/或包含SEQIDNO:2的氨基酸序列的重链CDR2,和/或包含SEQIDNO:3的氨基酸序列的重链CDR3;和/或包括包含SEQIDNO:4的氨基酸序列的轻链CDR1,和/或包含SEQIDNO:5的氨基酸序列的轻链CDR2,和/或包含SEQIDNO:6的氨基酸序列的轻链CDR3。在一些实施方案中,结合人OX40的拮抗剂抗体或其片段包括包含SEQIDNO:13的氨基酸序列的延伸的重链CDR1,和/或包含SEQIDNO:14的氨基酸序列的延伸的重链CDR2,和/或包含SEQIDNO:15的氨基酸序列的延伸的重链CDR3;和/或包括包含SEQIDNO:16的氨基酸序列的延伸的轻链CDR1,和/或包含SEQIDNO:17的氨基酸序列的延伸的轻链CDR2,和/或包含SEQIDNO:18的氨基酸序列的延伸的轻链CDR3。优选的,结合人OX40的拮抗剂抗体或其片段包括包含SEQIDNO:1的氨基酸序列的重链CDR1,包含SEQIDNO:2的氨基酸序列的重链CDR2,包含SEQIDNO:3的氨基酸序列的重链CDR3;和/或包含SEQIDNO:4的氨基酸序列的轻链CDR1,包含SEQIDNO:5的氨基酸序列的轻链CDR2,包含SEQIDNO:6的氨基酸序列的轻链CDR3。更优选的,结合人OX40的拮抗剂抗体或其片段包括包含SEQIDNO:1的氨基酸序列的重链CDR1,包含SEQIDNO:2的氨基酸序列的重链CDR2,包含SEQIDNO:3的氨基酸序列的重链CDR3;和包含SEQIDNO:4的氨基酸序列的轻链CDR1,包含SEQIDNO:5的氨基酸序列的轻链CDR2,和包含SEQIDNO:6的氨基酸序列的轻链CDR3。本领域普遍已知CDR3结构域不依赖于CDR1和/或CDR2结构域,单独就可以确定抗体对同族抗原的结合特异性,基于共同的CDR3序列可以可预测的产生具有相同结合特异性的多种抗体。参见例如KlimkaA等人,Br.J.Cancer83(2):252-260(2000)(描述了仅使用鼠抗CD30抗体Ki-4的重链可变结构域CDR3生产人源化抗CD30抗体);BeiboerSH等人,J.Mol.Biol.296:833-849(2000)(描述了仅使用亲本鼠MOC-31抗EGP-2抗体的重链CDR3序列生产重组的上皮糖蛋白-2(EGP-2)抗体);RaderC等人,Proc.NatlAcad.SciU.S.A.95:8910-8915(1998)(描述了使用鼠抗整联蛋白αvβ3抗体LM609的重链和轻链可变CDR3结构域的一组人源化抗整联蛋白αvβ3抗体,其中每个抗体成员在CDR3结构域之外都包括不同的序列,并能够以与亲本鼠抗体同样高或更高的亲和力结合与亲本鼠抗体相同的表位);Barbas等人,J.Am.Chem.Soc.116:2161-2162(1994)(公开了CDR3结构域为抗原结合提供了最重要的贡献)。因此,本发明提供了结合人OX40的抗体及其片段,其包括一个或多个重链和/或轻链CDR3结构域,特别是包括包含SEQIDNO:3的氨基酸序列的重链CDR3,和/或包含SEQIDNO:6的氨基酸序列的轻链CDR3,其中所述抗体能够结合人OX40。在一些实施方案中,本发明的抗体包括来自非人抗体的一个或多个重链和/或轻链CDR3结构域,所述抗体(a)能够与相应的亲本非人(例如鼠)抗体竞争结合;(b)保留相应的亲本非人(例如鼠)抗体的功能特征;(c)结合与相应的亲本非人(例如鼠)抗体结合的表位相同的表位;和/或(d)具有与相应的亲本非人(例如鼠)抗体相似的结合亲和力。在另一个方面,本发明提供了结合人OX40的拮抗剂抗体或其片段,其包括包含SEQIDNO:7的氨基酸序列的重链可变区序列。在另一个方面,本发明提供了结合人OX40的拮抗剂抗体或其片段,其包括包含SEQIDNO:8的氨基酸序列的轻链可变区序列。在一些实施方案中,结合人OX40的拮抗剂抗体或其片段包括包含SEQIDNO:7的氨基酸序列的重链可变区序列,和包含SEQIDNO:8的氨基酸序列的轻链可变区序列。在另一个方面,本发明提供了结合人OX40的拮抗剂抗体或其片段的变体。因此,本发明提供了抗体或其片段,具有这样的重链和/或轻链可变区序列的非CDR区的氨基酸序列,其与亲本拮抗剂抗体的重链或轻链之重链和/或轻链可变区序列(例如分别如SEQIDNO:7或SEQIDNO:8的重链和轻链可变区序列)的非CDR区的氨基酸序列至少80%相同(具有至少80%氨基酸序列同一性)。本发明还提供了这样的抗体或其片段,具有重链和/或轻链可变区序列的非延伸CDR区的氨基酸序列,所述序列与亲本拮抗剂抗体的重链或轻链之重链和/或轻链可变区序列的非延伸CDR区的氨基酸序列至少80%相同。优选的,重链和/或轻链可变区序列的非CDR区或非延伸CDR区的氨基酸序列同一性是至少85%,更优选至少90%,最优选至少95%,特别是96%,更特别97%,甚至更特别98%,最特别99%,包括例如80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%和100%。本文中关于氨基酸序列的同一性或同源性定义为在比对序列和为了实现最大百分比序列同一性,在必要时导入空位后,候选序列中与结合人OX40的拮抗剂抗体或其片段相同的氨基酸残基的百分比。因而,可以通过通常用于比较两条多肽的氨基酸位置相似性的标准方法,来确定序列同一性。使用计算机程序例如BLAST或FASTA,比对两条多肽各自氨基酸的最佳匹配(沿一条或两条序列的全长或者沿一条或两条序列预定的部分)。程序提供了默认的开放罚分和默认的空位罚分,评分矩阵例如PAM250(标准评分矩阵;参见DayhoffMO等人,(1978),inAtlasofProteinSequenceandStructure,第5卷,第3期增补)可与计算机程序结合使用。例如,百分比同一性可计算为:相同匹配的总数乘以100,再除以匹配跨度中较长序列的长度和为了比对两条序列向较长序列中导入的空位数之和。在一些实施方案中,本发明因而提供了结合人OX40的拮抗剂抗体或其片段,其中所述抗体或其片段包括与SEQIDNO:19、20、21、22或23的框架区序列至少70%相同的重链可变框架区序列,和/或与SEQIDNO:24、25、26、27和28的框架区序列至少60%相同的轻链可变框架区序列。在一些实施方案中,本发明提供了结合人OX40的拮抗剂抗体或其片段,其中所述抗体或其片段包括与SEQIDNO:19的框架区序列至少74%相同的重链可变框架区序列,和/或与SEQIDNO:24的框架区序列至少65%相同的轻链可变框架区序列。在另一个方面,本发明提供了结合人OX40的拮抗剂抗体或其片段,其包括如上文所述的重链和/或轻链CDR,并还包括这样的重链可变框架区,所述重链可变框架区是选自IGHV2-70*10(SEQIDNO:19)、IGHV2-70*01(SEQIDNO:20)、IGHV2-70*13(SEQIDNO:21)、IGHV2-5*09(SEQIDNO:22)和IGHV2-70*11(SEQIDNO:23)的人基因的产物或源自所述人基因;优选这样的重链可变框架区,所述重链可变框架区是IGHV2-70*10(SEQIDNO:19)的人基因的产物或源自所述人基因。重链可变框架区可包括一个或多个(例如,1、2、3和/或4个)重链框架区序列(例如,框架1(FW1)、框架2(FW2)、框架3(FW3)和/或框架4(FW4)),其存在于这些人基因的产物中或源自这些人基因。优选的,重链可变区框架包括FW1、FW2和/或FW3,更优选FW1、FW2和/或FW3,其存在于选自IGHV2-70*10(SEQIDNO:19)、IGHV2-70*01(SEQIDNO:20)、IGHV2-70*13(SEQIDNO:21)、IGHV2-5*09(SEQIDNO:22)和IGHV2-70*11(SEQIDNO:23)的人基因的产物中或源自所述人基因。本文使用的重链框架区序列包括FW1(位置1至位置25)、FW2(位置36至位置49)、FW3(位置66至位置94)和FW4(位置103至位置113),其中利用Kabat中提出的编号系统表示所述氨基酸位置。在一些实施方案中,本发明提供了抗体或其片段,其中抗体或其片段包括这样的重链可变框架区,所述重链可变框架区是IGHV2-70*10(SEQIDNO:19)的人基因的产物或源自所述人基因,其中所述重链可变框架区包括来自相应鼠抗体的相应重链可变框架区的至少一个氨基酸修饰。在一些实施方案中,本发明提供了抗体或其片段,其中抗体或其片段包括包含SEQIDNO:32的氨基酸序列的重链序列,其中重链可变框架区包括来自相应鼠抗体的相应重链可变框架区的至少一个氨基酸修饰。优选的,氨基酸修饰包括在选自23、35b、48、50、60和62的氨基酸位置上的氨基酸取代,更优选在选自23、35b、50、60和62的氨基酸位置上的氨基酸取代,最优选在氨基酸位置35b的氨基酸取代,其中根据Kabat编号表示每组成员的氨基酸位置。具体而言,氨基酸修饰包括选自23S、35bG、48L、50H、60N和62A的氨基酸取代,优选选自T23S、S35bG、I48L、R50H、S60N和S62A的氨基酸取代,而S35bG是最优选的氨基酸取代,其中根据Kabat编号表示每组成员的氨基酸位置。在另一个方面,本发明提供了结合人OX40的拮抗剂抗体或其片段的变体,其包括这样的轻链可变框架区,所述轻链可变框架区是选自IGKV3-11*01(SEQIDNO:24)、IGKV1-39*01(SEQIDNO:25)、IGKV1D-39*01(SEQIDNO:26)、IGKV3-11*02(SEQIDNO:27)和IGKV3-20*01(SEQIDNO:28)的人基因的产物或源自所述人基因;优选是IGKV3-11*01(SEQIDNO:24)的人基因的产物或源自所述人基因的轻链可变框架区。轻链可变框架区可包括一个或多个(例如,1、2、3和/或4个)轻链框架区序列(例如,框架1(FW1)、框架2(FW2)、框架3(FW3)和/或框架4(FW4)),其存在于这些人基因的产物中或源自这些人基因。优选的,轻链可变区框架包括FW1、FW2和/或FW3,更优选FW1、FW2和/或FW3,其存在于选自IGKV3-11*01(SEQIDNO:24)、IGKV1-39*01(SEQIDNO:25)、IGKV1D-39*01(SEQIDNO:26)、IGKV3-11*02(SEQIDNO:27)和IGKV3-20*01(SEQIDNO:28)的人基因的产物中或源自所述人基因。本文使用的轻链框架区序列包括FW1(位置1至位置23)、FW2(位置35至位置49)、FW3(位置57至位置88)和FW4(位置98至位置108),其中利用Kabat中提出的编号系统表示所述氨基酸位置。在一些实施方案中,本发明提供了抗体或其片段,其中抗体或其片段包括这样的轻链可变框架区,所述轻链可变框架区是IGKV3-11*01(SEQIDNO:24)的人基因的产物或源自所述人基因,其中所述轻链可变框架区包括来自相应鼠抗体的相应轻链可变框架区的至少一个氨基酸修饰。在一些实施方案中,本发明提供了抗体或其片段,其中抗体或其片段包括包含SEQIDNO:39的氨基酸序列的轻链序列,其中轻链序列的轻链可变框架区包括来自相应鼠抗体的相应轻链可变框架区的至少一个氨基酸修饰。优选的,氨基酸修饰包括在选自1、33、34、46、47、54、56和71的氨基酸位置上的氨基酸取代和/或氨基酸位置31上的缺失,更优选在选自33、34、46、47、54、56和71的氨基酸位置上的氨基酸取代和/或氨基酸位置31上的缺失,最优选氨基酸位置46和/或47的氨基酸位置上的氨基酸取代,其中根据Kabat编号表示每组成员的氨基酸位置。具体而言,氨基酸修饰包括选自1Q、33M、34H、46P、47W、54L、56S和71Y的氨基酸取代和/或T31的缺失,优选选自1Q、33M、34H、46P、47W、54L、56S和71Y的氨基酸取代,更优选选自33M、34H、46P、47W和71Y的氨基酸取代,而46P、47W是特别优选的,其中根据Kabat编号表示每组成员的氨基酸位置。在一些实施方案中,结合人OX40的拮抗剂抗体或其片段包括这样的重链可变框架区和轻链可变框架区,所述重链可变框架区是选自V2-70*10(SEQIDNO:19)、V2-70*01(SEQIDNO:20)、V2-70*13(SEQIDNO:21)、V2-5*09(SEQIDNO:22)和V2-70*11(SEQIDNO:23)的人基因的产物或源自所述人基因,所述轻链可变框架区是选自V3-11*01(SEQIDNO:24)、IGKV1-39*01(SEQIDNO:25)、IGKV1D-39*01(SEQIDNO:26)、IGKV3-11*02(SEQIDNO:27)和IGKV3-20*01(SEQIDNO:28)的人基因的产物或源自所述人基因;优选的重链可变框架区是V2-70*10(SEQIDNO:19)的人基因的产物或源自所述人基因,轻链可变框架区是V3-11*01(SEQIDNO:24)的人基因的产物或源自所述人基因。本发明还涵盖了存在于上述不同的人基因的产物中或源自所述人基因的重链可变区框架区,和/或存在于上述不同的人基因的产物中或源自所述人基因的轻链可变区框架区的组合。人重链和轻链可变区基因的种系DNA序列可见于―VBase”人种系序列数据库中(可获得自因特网的www.mrccpe.cam.ac.uk/vbase),以及Kabat,EA等人,见上文;Tomlinson,IM等人,(1992)J.MoI.Biol.227:776-798和Cox,JPL等人,(1994)Eur.J.Immunol.24:827-836。如另一个实例,人重链和轻链可变区基因的种系DNA序列可见于Genbank数据库。在另一个方面,本发明还提供了结合人OX40的拮抗剂抗体或其片段,其中至少一个重链CDR和/或至少一个轻链CDR包括至少一个氨基酸修饰。可以实施定点诱变或PCR介导的诱变,来导入修饰,并可以在体外或体内测定中评估对抗体结合的影响或者其他的目标功能特性。优选的导入保守修饰。修饰可以是氨基酸取代、添加或缺失,但优选是取代。通常,在CDR区中实施不超过5个,优选不超过4个,更优选不超过3个,甚至更优选不超过2个,最优选不超过1个的氨基酸修饰。在某些实施方案中,框架序列可用于改造可变区,产生变体抗体。本发明的变体抗体包括对VH和/或VK中的框架区残基进行了修饰的变体,例如改善抗体的特性。通常,进行此类框架区修饰来减少抗体的免疫原性。例如,一种方法是―回复突变‖一个或多个框架区残基为相应的鼠序列,或者―回复突变‖一个或多个框架区残基为相应的种系序列。因而在另一个方面,本发明提供了结合人OX40的拮抗剂抗体或其片段,其中所述拮抗剂抗体或其片段的至少一个重链可变区的框架区包括来自相应鼠抗体的重链可变区的相应框架区的至少一个氨基酸修饰。优选的,所述氨基酸修饰是氨基酸取代。通常,在框架区内实施不超过6个,优选不超过5个,优选不超过4个,更优选不超过3个,甚至更优选不超过2个,最优选不超过1个的氨基酸修饰。在一些实施方案中,本发明提供了结合人OX40的拮抗剂抗体或其片段,其中重链可变区的框架区的氨基酸修饰包括在选自23、35b、48、50、60和62的氨基酸位置上的氨基酸取代,其中根据Kabat编号表示每组成员的氨基酸位置。重链可变区的框架区的优选的氨基酸取代位于选自23、35b、50、60和62的氨基酸位置上。重链可变区的框架区的更优选的氨基酸取代是选自23S、35bG、48L、50H、60N和62A的氨基酸取代,而35bG是重链可变区的框架区的最优选的氨基酸取代。本发明提供了结合人OX40的拮抗剂抗体或其片段,其中所述拮抗剂抗体或其片段的至少一个轻链可变区的框架区包括来自相应鼠抗体的轻链可变区的相应框架区的至少一个氨基酸修饰。优选的,所述氨基酸修饰是氨基酸取代和/或氨基酸缺失。通常,在框架区内实施不超过6个,优选不超过5个,优选不超过4个,更优选不超过3个,甚至更优选不超过2个,最优选不超过1个的氨基酸修饰。在一些实施方案中,本发明提供了人源化抗体或其片段,其中轻链可变区的框架区的氨基酸修饰包括在选自1、33、34、46、47、54、56和71的氨基酸位置上的氨基酸取代和/或氨基酸位置31上的缺失。更优选的,轻链可变区的框架区的氨基酸修饰包括在Y31的缺失和/或选自1Q、33M、34H、46P、47W、54L、56S和71Y的氨基酸取代,其中根据Kabat编号表示每组成员的氨基酸位置。最优选的,轻链可变区的框架区的氨基酸修饰包括在T31的缺失和/或选自33M、34H、46P、47W和71Y的取代,而46P,和/或L47W是特别优选的。在一些实施方案中,本发明的人源化抗体或其片段可包括上述重链可变区序列的框架区的氨基酸修饰,和上述轻链可变区序列的框架区的氨基酸修饰。本发明还提供了结合人OX40的拮抗型抗体或其片段,其包括选自SEQIDNO:29、58、59、77、78、79和80的重链可变区,优选选自SEQIDNO:58、59、79和80,更优选选自SEQIDNO:58和59。本发明还提供了结合人OX40的拮抗型抗体或其片段,其包括选自SEQIDNO:30、60、81、82、83、84、85、86、87、88和89的轻链可变区,优选选自SEQIDNO:60、86、87和89,更优选SEQIDNO:60。在一些实施方案中,结合人OX40的拮抗型抗体或其片段包括选自SEQIDNO:29、58、59、77、78、79和80的重链可变区,和选自SEQIDNO:30、60、81、82、83、84、85、86、87、88和89的轻链可变区。考虑到每种上述重链和轻链可变区序列都可以结合人OX40,可以―混合和匹配‖重链和轻链可变区序列,来产生本发明的抗OX40结合分子。可以使用例如实施例中描述的结合测定,来测试此类―混合和匹配‖的抗体的OX40结合。在一些实施方案中,结合人OX40的拮抗型抗体或其片段包括选自SEQIDNO:58和59的重链可变区,和选自SEQIDNO:60和89的轻链可变区。在更优选的实施方案中,结合人OX40的拮抗型抗体或其片段包括包含SEQIDNO:58的氨基酸序列的重链可变区,和包含SEQIDNO:60的氨基酸序列的轻链可变区;包含SEQIDNO:58的氨基酸序列的重链可变区,和包含SEQIDNO:89的氨基酸序列的轻链可变区;包含SEQIDNO:59的氨基酸序列的重链可变区,和包含SEQIDNO:60的氨基酸序列的轻链可变区;包含SEQIDNO:59的氨基酸序列的重链可变区,和包含SEQIDNO:89的氨基酸序列的轻链可变区。最优选的是这样的结合人OX40的拮抗型抗体或其片段,其包括选自SEQIDNO:58和59的重链可变区,和包含SEQIDNO:60的氨基酸序列的轻链可变区本申请还提供了结合人OX40的拮抗型抗体或其片段,其包括选自SEQIDNO:32、33、34、35、36、37和38的重链序列,优选选自SEQIDNO:35、36、37和38,更优选选自SEQIDNO:37和38。本申请还提供了结合人OX40的拮抗型抗体或其片段,其包括选自SEQIDNO:39、40、41、42、43、44、45、46、47、48和49的轻链序列,优选选自SEQIDNO:45、46、47和49,更优选SEQIDNO:47。在一些实施方案中,结合人OX40的拮抗型抗体或其片段包括选自SEQIDNO:32、33、34、35、36、37和38的重链序列,和选自SEQIDNO:39、40、41、42、43、44、45、46、47、48和49的轻链序列。考虑到每种上述重链和轻链可变区序列都可以结合人OX40,可以―混合和匹配‖重链和轻链可变区序列,来产生本发明的抗OX40结合分子。可以使用例如实施例中描述的结合测定,来测试此类―混合和匹配‖的抗体的OX40结合。在一些实施方案中,结合人OX40的拮抗型抗体或其片段包括选自SEQIDNO:37和38的重链序列,和选自SEQIDNO:47和49的轻链序列。在更优选的实施方案中,结合人OX40的拮抗型抗体或其片段包括包含SEQIDNO:37的氨基酸序列的重链序列,和包含SEQIDNO:47的氨基酸序列的轻链序列;包含SEQIDNO:37的氨基酸序列的重链序列,和包含SEQIDNO:49的氨基酸序列的轻链序列;包含SEQIDNO:38的氨基酸序列的重链序列,和包含SEQIDNO:47的氨基酸序列的轻链序列;包含SEQIDNO:38的氨基酸序列的重链序列,和包含SEQIDNO:49的氨基酸序列的轻链序列。最优选的是这样的结合人OX40的拮抗型抗体或其片段,其包括选自SEQIDNO:37和38的重链序列,和包含SEQIDNO:47的氨基酸序列的轻链序列。在本发明的一个实施方案中,拮抗剂抗体或其片段是鼠抗体、嵌合抗体或人源化抗体,优选人源化抗体,更优选单克隆鼠抗体、单克隆嵌合抗体或单克隆人源化抗体。本发明还提供了结合人OX40的单价抗体或其片段,即,由单抗原结合臂组成的抗体。本发明还提供了结合人OX40的抗体的片段,所述片段选自Fab、Fab′、Fab′-SH、Fd、Fv、dAb、F(ab')2、scFv、双特异性单链Fv二聚体、双抗体、三链抗体和与相同或不同抗体遗传融合的scFv。优选的片段是scFv、双特异性单链Fv二聚体和双抗体。本发明还提供了结合人OX40的全长抗体。本发明还提供了结合人OX40的抗体或其片段,其还包括重链和/或轻链恒定区,特别是人重链和/或人轻链恒定区。人重链恒定区可选自IgG1(IGHG1)、IgG2(IGHG2)、IgG3(IGHG3)、IgG4(IGHG4)、IgA1(IGHA1)、IgA2(IGHA2)、IgM(IGHM)、IgD(IGHD)或IgE(IGHE)构成的人免疫球蛋白的组,而人重链恒定区IgG,特别是IgG1(IGHG1)是优选的。人轻链恒定区可选自κ或λ恒定区构成的人免疫球蛋白的组,而人κ恒定区是优选的。在一些优选的实施方案中,所述结合人OX40的拮抗型抗体或其片段包括人IgG1(IGHG1)重链恒定结构域和人轻链κ恒定结构域。除了在框架区或CDR区进行修饰外,或作为其备选,还可以改造本发明的抗体以包括Fc区内的修饰,通常改变抗体的一种或多种功能特性,例如血清半衰期、补体固定、Fc受体结合,和/或抗原依赖性细胞的细胞毒作用。此外,可以化学修饰(例如,可以在抗体上连接一个或多个化学部分)或修饰本发明的抗体,改变它的糖基化。这些方案中的每一种如下详细描述。如下概述的Fc区内的修饰是根据Fc区内的残基的EU编号。在一个实施方案中,修饰CH1的铰链区,使得改变铰链区内的半胱氨酸残基数,例如增加或减少。该方法进一步描述在Bodmer等人的美国专利号5,677,425中。改变CH1的铰链区内的半胱氨酸残基数,以例如促进轻链和重链装配,或增加或减少抗体的稳定性。在另一个实施方案中,突变抗体的Fc铰链区,以减少抗体的生物学半衰期。更具体而言,在Fc铰链片段的CH2-CH3结构域交界区中导入一个或多个氨基酸突变,使得抗体相比天然的Fc铰链结构域SpA结合具有受损的葡萄球菌A蛋白(SpA)结合。该方法进一步描述在Ward等人的美国专利号6,165,745中。在另一个实施方案中,修饰抗体以增加其生物学半衰期。多种方法都是可以的。例如,可以导入一种或多种下列突变:T252L、T254S、T256F,如Ward等人的美国专利号6,277,375所述。可选的,为了增加生物学半衰期,可以在CH1或CL区中改变抗体,以含有取自IgG的Fc区的CH2结构域的2个环中的补救受体(salvagereceptor)结合表位,如Presta等人的美国专利号5,869,046和6,121,022所述。在其他实施方案中,通过用不同的氨基酸残基替换至少一个氨基酸残基来改变Fc区,以改变抗体的效应子功能。例如,可以用不同的氨基酸残基替换选自氨基酸残基234、235、236、237、297、318、320和322的一个或多个氨基酸,使得抗体对效应子配体具有改变的亲和力,但保留亲本抗体的抗原结合能力。改变亲和力的效应子配体可以是例如Fc受体或补体的C1组分。该方法进一步描述在Winter等人的美国专利号5,624,821和5,648,260中。在另一个实例中,可以用不同的氨基酸残基替换选自氨基酸残基329、331和322的一个或多个氨基酸,使得抗体具有改变的C1q结合和/或降低或消除的补体依赖性细胞毒作用(CDC)。该方法进一步描述在Idusogie等人的美国专利号6,194,551中。在另一个实例中,改变CH2结构域的N端区域中氨基酸位置231至238中的一个或多个氨基酸残基,从而改变抗体固定补体的能力。该方法进一步描述在Bodmer等人的PCT公开WO94/29351中。在另一个实例中,修饰Fc区以增加抗体介导的抗体依赖性细胞毒作用(ADCC)的能力,和/或增加抗体对Fcγ受体的亲和力,通过修饰下列位置上的一个或多个氨基酸:238、239、248、249、252、254、255、256、258、265、267、268、269、270、272、276、278、280、283、285、286、289、290、292、293、294、295、296、298、301、303、305、307、309、312、315、320、322、324、326、327、329、330、331、333、334、335、337、338、340、360、373、376、378、382、388、389、398、414、416、419、430、434、435、437、438或439。该方法进一步描述在Presta的PCT公开WO00/42072中。本发明还提供了结合人OX40的拮抗型抗体或其片段,其包括人重链和/或轻链恒定区,其中人重链恒定区包括包含来自人IgG4(IGHG4)的CH1区、铰链区、CH2区和CH3区的同种型变体,其中铰链区包括第228位丝氨酸取代为脯氨酸。优选的,包含同种型变体的人源化抗体是全长抗体。特别优选的结合人OX40的人源化抗体或其片段包括包含SEQIDNO:57的氨基酸序列的重链序列和包含SEQIDNO:47的氨基酸序列的轻链序列,所述抗体或其片段包括包含人IgG4(IGHG4)的CH1、具有S228P取代的人IgG4(IGHG4)的铰链区和人IgG4(IGHG4)的CH2和CH3的同种型变体。已发现相比包含来自人IgG1(IGHG1,通常是天然的人IgG1)的人重链恒定区的结合人OX40的拮抗型抗体或其片段(即,相比仅在修饰的重链恒定区不同于同种型变体的结合人OX40的拮抗型抗体或其片段),同种型变体没有表现出Fc介导的细胞毒作用机制,如ADCC。本发明还提供了结合人OX40的拮抗型抗体或其片段,其包括人IgGFc区,其中与人IgGFc区连接的成熟核心碳水化合物结构缺少岩藻糖(在本文中被可选的称为―无岩藻糖基化‖)。优选的,抗体包括人IgG1(IGHG1)Fc区,其中与人IgG1(IGHG1)Fc区连接的成熟核心碳水化合物结构缺少岩藻糖。更优选是包括人IgG1(IGHG1)Fc区的全长抗体,其中与人IgG1(IGHG1)Fc区连接的成熟核心碳水化合物结构缺少岩藻糖。从WO03/035835已知,与人IgGFc区连接的成熟核心糖类结构缺乏岩藻糖可增强ADCC。因而,在其它实施方案中,本发明的拮抗型抗体或其片段包括这样的人IgG1(IGHG1)Fc区,其中与所述人IgG1(IGHG1)Fc区连接的成熟核心糖类结构缺乏岩藻糖,而所述缺乏岩藻糖的抗体相比不缺乏岩藻糖的亲本人源化抗体或其片段表现出增强的ADCC。产生缺乏岩藻糖的抗体的方法是例如(a)使用改造或突变的宿主细胞,所述宿主细胞是岩藻糖代谢缺陷的,使得其对其中表达的蛋白质岩藻糖基化能力降低(或不能岩藻糖基化);(b)在阻止或降低岩藻糖基化的条件下培养细胞;(c)翻译后去除岩藻糖(例如,使用岩藻糖苷酶);(d)翻译后添加理想的糖类,例如,在重组表达未糖基化的糖蛋白之后;或者(e)纯化糖蛋白,选择没有岩藻糖基化的产物。优选使用的是WO10/095031的实施例14描述的方法,例如Longmore等人(1982)Carbohydr.Res.365-92或Imai-Nishiya等人,(2007),BMCBiotechnol.7,84中描述的方法。本发明还提供了结合人OX40的拮抗剂抗体或其片段,其结合与包括包含SEQIDNO:7的氨基酸序列的重链可变序列和/或包含SEQIDNO:8的氨基酸序列的轻链可变序列的抗体所识别的表位相同的表位。本发明还提供了人OX40的特定区域或表位,特别是人OX40受体胞外结构域的区域或表位,其被本发明提供的抗体结合,特别是被包括包含SEQIDNO:7的氨基酸序列的重链可变序列和/或包含SEQIDNO:8的氨基酸序列的轻链可变序列的抗体结合。可以通过本领域已知的任何合适的表位定位方法,与本发明提供的任一种抗体结合,来鉴别人OX40多肽的这一特定区域或表位。这类方法的例子包括在源自OX40的具有不同长度的肽中筛选结合本发明抗体的最小片段,所述片段可以特异性结合含有被抗体识别的表位序列的抗体。可以合成或通过蛋白质水解消化OX40多肽,来生产OX40肽。可以通过例如质谱分析鉴别结合抗体的肽。在另一个实例中,可以使用NMR光谱或X射线晶体衍射鉴别被本发明的抗体结合的表位。在需要时,一经鉴别,可以使用结合本发明抗体的表位片段作为免疫原,获得结合相同表位的其他拮抗剂抗体。抗OX40抗体特性评估抗体针对例如人OX40的结合能力的标准测定是本领域已知的,包括例如ELISA、Western印迹、RIA和流式细胞仪分析。合适的测定详细描述在实施例中。也可以通过本领域已知的标准测定来评估抗体的结合动力学(例如,结合亲和力如KD),例如Scatchard或系统分析。可以通过本领域已知的标准竞争性测定来评估相对结合亲和力Ki。在另一方面,本发明提供了结合人OX40的拮抗型抗体或其片段,以剂量依赖性的方式,比重组人源化抗体依法利珠单抗更高的程度阻断人混合型淋巴细胞反应(MLR)。重组人源化抗体依法利珠单抗结合淋巴细胞功能相关抗原1的CD11a亚基。可以根据实施例3进行和测量MLR。在另一方面,本发明提供了结合人OX40的拮抗剂抗体或其片段,其还能够识别食蟹猴OX40。根据实施例4,使拮抗型抗OX40抗体与人和食蟹猴外周血单核细胞(PBMC)结合并进行测量,且在图3中示出。发现抗体识别在人和食蟹猴的活化的淋巴细胞表面表达的OX40,表示该抗体具有交叉反应特性。在另一方面,本发明提供了结合人OX40,特别是分离形式的人OX40的拮抗型抗体或其片段,如实施例5和6中详述的以及图4中示意的,其与被用作分析物的重组单价人OX40受体胞外结构域(SEQIDNO:11)的亲和力(KD)为500nM或更低,优选200nM或更低,更优选150nM或更低,更优选120nM或更低,甚至更优选110nM或更低,例如通过仪器(GEHealthcareEuropeGmbH,Glattbrugg,Switzerland)对偶联蛋白A的CM5研究级传感芯片(GEHealthcareEuropeGmbH,Glattbrugg,Switzerland;BR-1000-14)上捕获的抗体进行表面等离振子共振(SPR)测量。本文使用的―单价‖涉及使用OX40受体的亲和力测量时,指人OX40受体结构域(如胞外结构域),没有人为二聚化或如其原本应该的那样多聚化的,例如当结构域与免疫球蛋白Fc区氨基端融合时。在优选的方面,本发明提供了保留相应嵌合抗体的至少75%的OX40结合亲和力(KD)的人源化抗体或其片段。优选的,结合人OX40的人源化抗体或其片段与相应的嵌合抗体具有等价的亲和力。“等价的亲和力‖意指在相应嵌合抗体的OX40结合亲和力的±10%范围内的亲和力值。更优选的,本发明提供了具有比相应嵌合抗体更高的亲和力的结合人OX40的人源化抗体或其片段。在本发明的优选的方面,提供了结合人OX40的拮抗型抗体或其片段,如实施例5和6中详述的以及图4中示意的,其与被用作分析物的重组单价人OX40受体胞外结构域(SEQIDNO:11)的结合亲和力(KD)为110nM或更低,优选100nM或更低,更优选90nM或更低,更优选80nM或更低,甚至更优选70nM或更低,例如通过仪器(GEHealthcareEuropeGmbH,Glattbrugg,Switzerland)对偶联蛋白A的CM5研究级传感芯片(GEHealthcareEuropeGmbH,Glattbrugg,Switzerland;BR-1000-14)上捕获的抗体进行表面等离振子共振(SPR)测量。本发明的另一方面提供了具有良好热稳定性的结合人OX40的拮抗型抗体或其片段。在优选的实施方案中,结合人OX40的拮抗型人源化抗体或其片段,具有热稳定温度大于70℃,优选大于75℃,更优选大于80℃,甚至更优选大于85℃的FAB片段。为了分析FAB片段热稳定性,使用差异扫描量热法,在全长IgG中鉴别FAB片段的中点融解温度。这类量热测量法是本领域技术人员已知的,并可根据例如Garber&Demarest(2007),BBRC,355:751-7进行,如实施例6中进一步描述的和图6所示。在另一方面,本发明描述了结合人OX40胞外区上的表位的拮抗型抗体或其片段。如实施例7所述和图7所示,在人和大鼠序列之间交换OX40胞外区的四个结构域中的一个或多个,并生成Fc融合蛋白。然后,实施结合ELISA,测试拮抗型人源化抗体对人OX40胞外区、大鼠OX40胞外区和四种人-大鼠嵌合蛋白的反应性。因而本发明提供了定位在人OX40胞外区的第二结构域中的拮抗型抗体或其片段。本发明还提供了可用于抑制免疫反应的拮抗型抗体或其片段。在用作自体反应性T细胞活化和增殖的体外模型(O’FlahertyE等人,(2000)Immunology,100(3):289-99;DuPontB&HansenJA(1976)Adv.Immunol.23:107-202)的MLR(参见实施例8)中测试拮抗型人源化抗OX40抗体的效果。混合来自两个不相关供体的PBMC,导致激活T细胞和T淋巴细胞增殖。此外,在该测定中测试3种不同的拮抗型人源化抗OX40抗体模式:IgG1(IGHG1)模式、无岩藻糖基化IgG1(IGHG1)模式和IgG4(IGHG4)模式,进一步确定细胞毒作用机制(如ADCC)对MLR抑制的贡献。拮抗型人源化抗OX40抗体在2个不同的个体(应答者)中都有效抑制了MLR,EC50值是约100ng/mL。然而,结果表现出依赖使用的抗体模式的差异。在第一个个体(应答者1)中,IgG1(IGHG1)和IgG4(IGHG4)抗体模式有效的抑制了T细胞反应性,表示对于这一个体,细胞毒作用机制不是关键性的。对于第二个个体(应答者2),IgG1(IGHG1)模式实现了超过60%的抑制,而IgG4(IGHG4)模式仅微弱的阻断了MLR。在两个个体中,无岩藻糖基化IgG1(IGHG1)模式都非常有效的抑制MLR。上述结果表示,在一些个体中,阻断OX40激活可以有效抑制MLR,且额外的细胞毒作用机制可以极大的增强该效果。因此,对于治疗患OX40介导的病症的患者,施用结合人OX40且具有增强的细胞毒作用机制的拮抗型抗体或其片段是特别有效的,其中所述病症表现为不依赖于患者的OX40共刺激状态,例如具有低OX40表达水平的患者。本发明的优选的实施方案提供了结合人OX40的拮抗型人源化抗体,用于治疗患OX40介导的病症的患者。此外,患者可具有低的OX40表达水平。优选的,结合人OX40的拮抗型人源化抗体包括IgG1(IGHG1)区。更优选的,结合人OX40的拮抗型人源化抗体包括无岩藻糖基化的IgG1区。在本发明的另一方面,在异种移植物抗宿主反应中证实拮抗型抗体或其片段的效果,其中用人PBMC重建SCID小鼠。该反应提供了用于在人类患者骨髓移植后观察到的同种异体移植物抗宿主病(GVHD)的模型。在该模型中,人PBMC,特别是T淋巴细胞,对小鼠宿主细胞发动强烈的应答,产生严重的炎性症状。如实施例9所述和图9及表10所示,结合人OX40的拮抗型人源化抗体在1mg/kg剂量时强烈抑制GVHD反应。令人惊讶的是,该抗体表现出比更好的效果,是被认可的GVHD疗法(XhaardA等人,(2011)Bull.Cancer,98(8):889-99;SimpsonD(2001)ExpertOpin.Pharmacother.2(7):1109-17)。因此,在优选的实施方案中,本发明提供了结合人OX40的拮抗型抗体或其片段,其在治疗GVHD中是有效的。优选的,向对象施用抗体导致存活中位数(天)比施用载体时改善4倍。更优选的,向对象施用抗体导致存活中位数(天)比施用改善2倍。因此,本发明提供了结合人OX40的拮抗型抗体或其片段,其在治疗GVHD的患者和/或抑制GVHD时比更有效。此外,已报道了激动型抗OX40结合抗体在同种异体小鼠GVHD模型(ValzasinaB等人,(2005)Blood,105(7):2845-51;BlazarBR等人,(2003)Blood,101(9):3741-8)中使GVHD恶化,因此,从实施例9可以概括:本发明的拮抗型抗体及其片段在结合人OX40后不表现出激动效果,因为在模型中没有观察到GVHD恶化。因此,本发明提供了结合后不表现出激动活性的结合人OX40的人源化抗体或其片段。核酸、载体和宿主细胞本发明还提供了编码结合人OX40的抗体及其片段的分离的核酸、载体以及包含所述核酸或载体的宿主细胞。核酸可位于完整细胞中、细胞裂解液中或者以部分纯化的或基本纯化的形式。当通过标准技术,包括碱/SDS处理、CsCl分带、柱层析、琼脂糖凝胶电泳和其他本领域普遍已知的技术,与其他细胞组分或其他污染物(例如,其他的细胞核酸或蛋白质)纯化分离时,核酸是―分离的‖或―使得基本纯的‖,参见例如F.Ausubel,等人编著,(1987)CurrentProtocolsinMolecularBiology,GreenePublishingandWileyInterscience,NewYork。本发明的核酸可以是例如DNA或RNA,可以含有或不含内含子序列。在优选的实施方案中,核酸是cDNA分子。可以使用标准的分子生物学技术获得本发明的核酸,例如可以通过标准的PCR扩增或cDNA克隆技术,获得编码抗体的轻链和重链或者编码VH和VL区段的cDNA。对于从免疫球蛋白基因文库获得的抗体(例如,使用噬菌体展示技术),可以从文库回收编码抗体的一种或多种核酸。向宿主细胞中导入外源核酸的方法是本领域普遍已知的,并可随所使用的宿主细胞而变化。技术包括但不限于葡聚糖介导的转染、磷酸钙沉淀、氯化钙处理、聚乙烯亚胺介导的转染、聚凝胺(polybrene)介导的转染、原生质体融合、电穿孔、病毒或噬菌体感染、在脂质体中封装多核苷酸和直接显微注射DNA到核中。在哺乳动物细胞的情况下,转染可以是瞬时或稳定的。本发明的优选的核酸分子是编码选自SEQIDNO:32、33、34、35、36、37和38的重链序列,和/或选自SEQIDNO:39、40、41、42、43、44、45、46、47、48和49的轻链序列的那些核酸分子。本发明的优选的核酸分子是编码选自SEQIDNO:29、58、59、77、78、79和80的重链可变区,和/或选自SEQIDNO:30、60、81、82、83、84、85、86、87、88和89的轻链可变区的那些核酸分子。本发明的优选的核酸分子是编码SEQIDNO:8的轻链可变区,和/或SEQIDNO:7的重链可变区的那些核酸分子;例如,包含SEQIDNO:9的核酸序列的重链可变区编码DNA,和/或包含SEQIDNO:10的核酸序列的轻链可变区编码DNA。本发明的更优选的核酸分子是编码SEQIDNO:58或59的重链可变区,和/或SEQIDNO:60的轻链可变区的那些核酸分子;例如,包含SEQIDNO:61或62的核酸序列的重链可变区编码DNA,和/或包含SEQIDNO:63的核酸序列的轻链可变区编码DNA是最优选的。一旦获得编码VH和VL区段的DNA片段,则可以通过标准的重组DNA技术进一步操作这些DNA片段,例如将可变区基因转化为全长抗体链基因,或转化为对应于上文所述片段的片段基因,如Fab片段基因或转化为scFv基因。在这些操作中,将VL或VH编码DNA片段有效的连接至编码另一种蛋白质(例如抗体恒定区或柔性接头)的另一种DNA片段上。该上下文中使用的术语―有效连接‖意指连接两个DNA片段,使得所述两个DNA片段编码的氨基酸序列保持读码框连续。通过将VH编码DNA与另一种编码重链恒定区(CH1、CH2和CH3)的DNA分子有效连接,可以将编码VH区的分离的DNA转化为全长重链基因。这些人重链恒定区基因的序列是本领域已知的(参见例如,Kabat,E.A.等人,见上文),可以通过标准PCR扩增获得涵盖了这些区域的DNA片段。重链恒定区可以是IgG1(IGHG1)、IgG2(IGHG2)、IgG3(IGHG3)、IgG4(IGHG4)、IgA1(IGHA1)、IgA2(IGHA2)、IgM(IGHM)、IgD(IGHD)或IgE(IGHE)恒定区,但最优选是IgG1(IGHG1)恒定区。对于Fab片段重链基因,可以将VH编码DNA与另一种只编码重链CH1恒定区的DNA分子有效连接。通过将VL编码DNA与另一种编码轻链恒定区CL的DNA分子有效连接,可以将编码VL区的分离的DNA转化为全长轻链基因(以及Fab轻链基因)。这些人轻链恒定区基因的序列是本领域已知的(参见例如,Kabat,E.A.等人,见上文),可以通过标准PCR扩增获得涵盖了这些区域的DNA片段。在优选的实施方案中,轻链恒定区可以是κ或λ恒定区,优选κ恒定区。为了产生scFv基因,可以将VH和VL编码DNA片段与另一种编码柔性接头的片段有效连接,例如,编码氨基酸序列(GIy4-Ser)3的片段,使得VH和VL序列可以作为连续的单链蛋白质表达,其中VL和VH区通过柔性接头连接(参见例如,Bird等人,(1988)Science242:423-426;Huston等人,(1988)Proc.Natl.Acad.ScLUSA85:5879-5883;McCafferty等人,(1990)Nature348:552-554)。已开发了各种技术生产抗体的抗体片段。传统上,这些片段源自完整抗体的蛋白质水解消化(参见例如,MorimotoK等人,JournalofBiochemicalandBiophysicalMethods,24:107-117(1992);和Brennan等人,Science,229:81-3(1985))。然而,现在可以通过重组宿主细胞直接生产这些片段。例如,可以从上文讨论的抗体噬菌体文库中分离抗体片段。可选的,可以从大肠杆菌中直接回收Fab′-SH片段,并将其化学偶联形成F(ab′)2片段(CarterP等人,Bio/Technology,10:163-167(1992))。根据另一种方法,可以从重组宿主细胞培养物中直接分离F(ab′)2片段。生产抗体片段的其他技术对熟练的技术人员而言是显而易见的。在其他实施方案中,选定的抗体是单链Fv片段(scFv),参见例如WO1993/16185;美国专利号5,571,894和美国专利号5,587,458。抗体片段还可以是―线性抗体‖,例如美国专利号5,641,870所述。为了表达蛋白质,可以将编码本发明抗体的核酸整合到载体中,优选表达载体中。多种表达载体可用于蛋白质表达。表达载体可包括自我复制的染色体外载体,或整合到宿主基因组中的载体。构建的表达载体与宿主细胞类型是可相容的。因而,可用于本发明的载体(优选表达载体)包括但不限于使蛋白质能够在哺乳动物细胞、细菌、昆虫细胞、酵母和体外系统中表达的那些载体。如本领域已知的,多种表达载体是可商业或其他方式获得的,可用于本发明中来表达抗体。表达载体通常包括与控制或调控序列、可选择的标志物、任何融合配偶体和/或其他元件有效连接的蛋白质。本文中,―有效连接‖意指核酸置于与另一种核酸序列的功能性关联中。术语―调控序列‖意在包括启动子、增强子和控制抗体链基因转录或翻译的其他表达控制元件(例如,多聚腺苷酸化信号)。此类调控序列描述在例如Goeddel(GeneExpressionTechnology,MethodsinEnzymology185,AcademicPress,SanDiego,CA(1990))中。一般而言,这些表达载体包括与编码抗体的核酸有效连接的转录和翻译调控核酸,且通常对用于表达所述蛋白质的宿主细胞是恰当的。一般而言,转录和翻译调控序列可包括启动子序列、核糖体结合位点、转录起始和终止序列、翻译起始和终止序列,和增强子或激活子序列。还如本领域已知的,表达载体通常含有选择基因或标志物,允许选择含有表达载体的转化的宿主细胞。选择基因是本领域普遍已知的,可随使用的宿主细胞而变化。例如,通常可选择的标志物基因赋予已导入载体的宿主细胞对药物的抗性,例如G418、潮霉素或氨甲喋呤。优选的可选择的标志物基因包括二氢叶酸还原酶(DHFR)基因(用于用氨甲喋呤选择/扩增的dhfr-宿主细胞中)和neo基因(用于G418选择)。用于在克隆或表达本文载体中的DNA的合适的宿主细胞是原核细胞、酵母或更高等的真核细胞。用于该目的的合适的原核细胞包括真细菌,包括革兰氏阴性生物或革兰氏阳性生物,例如肠杆菌科(Enterobacteriaceae)如埃希氏菌属(Escherichia)例如大肠杆菌、肠杆菌属(Enterobacter)、克雷伯氏菌属(Klebsiella)、变形杆菌属(Proteus)、沙门氏菌属(Salmonella)例如鼠伤寒沙门氏菌(Salmonellatyphimurium)、沙雷菌属(Serratia)例如粘质沙雷菌(Serratiamarcescans),和志贺菌属(Shigella),以及芽孢杆菌属(Bacilli)例如枯草芽孢杆菌(B.subtilis)和地衣芽孢杆菌(B.licheniformis)、假单胞菌属(Pseudomonas)例如铜绿假单胞菌(P.aeruginosa)和链霉菌属(Streptomyces)。合适的大肠杆菌克隆宿主包括大肠杆菌294(ATCC31,446)、大肠杆菌B、大肠杆菌X1776(ATCC31,537)和大肠杆菌W3110(ATCC27,325)。除原核细胞外,真核微生物例如丝状真菌或酵母是合适的克隆或表达宿主。在低等真核宿主微生物中,酿酒酵母(Saccharomycescerevisiae)或普通的贝克酵母是最常用的。然而,多种其他属、种和菌株通常是可获得且有效的,例如粟酒裂殖酵母(Schizosaccharoriycespombe);克鲁维酵母属(Kluyveromyces)宿主,包括乳酸克鲁维酵母(K.lactis)、脆壁克鲁维酵母(K.fragilis)(ATCC12,424)、保加利亚克鲁维酵母(K.bulgaricus)(ATCC16,045)、威克克鲁维酵母(K.wickeramii)(ATCC24,178)、K.WaItH(AJCC56,500)、果蝇克鲁维酵母(K.drosopmarum)(ATCC36,906)、耐热克鲁维酵母(K.thermotolerans)或K.marxianusyarrowia(EP402226);巴斯德毕赤酵母(Pichiapastoris)(EP183,070);念珠菌属(Candida);里氏木霉(Trichodermareesia(EP244234));粗糙脉孢菌(Neurosporacrassa);许旺酵母属(Schwanniomyces)例如许旺酵母(Schwanniomycesoccidentalis);和丝状真菌,包括链孢霉属(Neurospora)、青霉属(Penicillium)、Tolypocladium或曲霉属(Aspergillus)宿主例如构巢曲霉(A.nidulans)或黑曲霉(A.niger)。用于表达本发明的人源化抗体的合适的宿主细胞源自多细胞生物。无脊椎动物细胞的实例包括plaril和昆虫细胞。已鉴别了多种杆状病毒菌株和变体,与相应的许可昆虫宿主细胞,所述细胞来自宿主例如草地贪夜蛾(Spodopterafrugiperda)(毛虫)、埃及伊蚊(Aedesaegypti)(蚊子)、白纹伊蚊(Aedesalbopictus)(蚊子)、黑腹果蝇(Drosophilamelanogaster)(果蝇)和家蚕(Bombyxmori)。用于转染的多种病毒菌株是公众可获得的,例如苜蓿尺蛾(Autographacalifornica)NPV的L-1变体和家蚕NPV的Bm-5菌株,此类病毒可特别用于转染草地贪夜蛾细胞。还可以利用棉花、谷物、土豆、大豆、碧冬茄属、西红柿和烟草的植物细胞培养物作为宿主。用于表达本发明的重组抗体的宿主细胞优选的是哺乳动物宿主细胞,包括中国仓鼠卵巢细胞(CHO细胞)(包括dhfr-CHO细胞,描述在Urlaub和Chasin,(1980)Proc.Natl.Acad.ScLUSA77:4216-4220中,与DHFR可选择标志物一起使用,例如R.J.Kaufman和P.A.Sharp(1982)J.MoI.Biol159:601-621所述)、NSO骨髓瘤细胞、COS细胞和SP2细胞。特别的,为了与NSO骨髓瘤细胞一起使用,另一种优选的表达系统是GS基因表达系统,公开在WO87/04462(Wilson)、WO89/01036(Bebbington)和EP338841(Bebbington)中。当将重组的抗体基因导入哺乳动物宿主细胞中时,通过培养宿主细胞一定时间来生产抗体,所述时间足以允许抗体在所述宿主细胞中的表达,或更优选的,允许抗体分泌到所述宿主细胞生长的培养基中。可以在多种培养基中培养用于生产结合人OX40的抗体的宿主细胞。可商购的培养基例如Ham'sF10(Sigma-AldrichChemieGmbH,Buchs,Switzerland)、最低必需培养基(MEM,Sigma-AldrichChemieGmbH)、RPMI-1640(Sigma-AldrichChemieGmbH,Basel,Switzerland)和Dulbecco'sModifiedEagle'sMedium(DMEM;Sigma-AldrichChemieGmbH)适合培养宿主细胞。可以使用标准的蛋白质纯化方法,从培养基中回收抗体。抗体可与融合配偶体有效连接,使得能够进行表达的蛋白质的靶向、纯化、筛选、展示等。融合配偶体可通过接头序列与抗体序列连接。接头序列一般包括少量氨基酸,通常少于10个,但也可以使用更长的接头。通常,选择柔性且抗降解的接头序列。对本领域技术人员显而易见的是,多种序列的任一种都可用作接头。例如,普通的接头序列包括氨基酸序列GGGGS。融合配偶体可以是靶向序列或信号序列,指导抗体和任何相关的融合配偶体到达所需要的细胞位置或胞外基质中。如本领域已知的,某些信号序列可以将蛋白质靶向分泌到生长基质中,或位于细胞的内膜和外膜之间的周质空间中。融合配偶体还可以是编码使得能够纯化和/或筛选的肽或蛋白质的序列。此类融合配偶体包括但不限于多聚组氨酸标签(His-tag)(例如H6和H10或其它与固定化金属亲和层析(IMAC)系统一起使用的标签(例如,Ni+2亲和柱))、GST融合物、MBP融合物、Strep-标签、细菌酶BirA的BSP生物素化靶序列,和由抗体靶向的表位标签(例如c-myc标签、flag标签等)。对本领域技术人员显而易见的是,此类标签可用于纯化、筛选或两者。构建和生产抗体通过使用普遍已知的和常规的操作规程免疫动物,获得针对OX40多肽生成的抗体,即,向动物,优选非人动物施用多肽,参见例如实验免疫学手册(WeirDM(编著),第4卷,BlackwellScientificPublishers,Oxford,England,1986)。可以免疫多种暖血动物,如兔、小鼠、大鼠、绵羊、牛、骆驼或猪。然而,小鼠、兔、猪和大鼠通常是最合适的,特别是小鼠。可以通过技术人员已知的重组DNA技术生产抗体。此外,可以通过酶促或化学切割天然存在的抗体,来生产抗体。可以通过将来自非人动物(例如小鼠)的VH和/或VL区的一个或多个CDR或其部分转移至人VH和/或VL区的一个或多个框架区上,来构建本发明的人源化抗体。任选的,当减少抗体的免疫原性和/或维持结合亲和力需要或理想时,可以用相应的非人(例如小鼠)残基替换位于VH和/或VL区中的人框架残基。任选的,可以用人残基替换位于CDR中的非人氨基酸残基。可以基于如上所述制备的非人单克隆抗体的序列,来制备本发明的嵌合或人源化抗体。可以从目的非人杂交瘤中获得编码重链和轻链免疫球蛋白的DNA,并使用标准的分子生物学技术将其改造为含有非鼠(例如人)免疫球蛋白序列。例如,为了产生嵌合抗体,可以使用本领域已知的方法将鼠可变区与人恒定区连接(参见例如,Cabilly等人的美国专利号4,816,567)。为了产生人源化抗体,可以使用本领域已知的方法将鼠CDR区插入到人框架中(参见例如,Winter的美国专利号5,225,539,和Queen等人的美国专利号5,530,101;5,585,089;5,693,762和6,180,370)。可以构建本发明的人源化抗体,其中基于在潜在的受体分子可变区和鼠抗体重链可变区之间的同源性的考虑,来选择重链可变区的人受体分子。种系候选人受体分子优选是降低潜在的免疫原性的。种系数据库由读取时穿过重链FW3区的末端且部分至CDR3序列的抗体序列构成。为了选择FW4区,可以搜索源自选定的种系分子的成熟抗体序列数据库,或使用源自人类供体的选定的种系分子的抗体序列。人受体分子优选选自与鼠供体分子相同的重链类型,而且是与鼠供体分子的可变区相同的规范结构类型。针对重链可变区选择人受体分子的次要考虑是规避鼠供体分子和人受体分子之间在CDR长度上的同源性。优选通过搜索V-BASE数据库的同源性,来选择人受体抗体分子,但也可以使用其他数据库如Kabat和公开的NCBI数据库。可以构建本发明的人源化抗体,其中基于在潜在的受体分子可变区和鼠抗体轻链可变区之间的同源性的考虑,来选择轻链可变区的人受体分子。种系候选人受体分子优选是降低潜在的免疫原性的。种系数据库由读取时穿过重链FW3区的末端且部分至CDR3序列的抗体序列构成。为了选择FW4区,可以搜索源自选定的种系分子的成熟抗体数据库,或使用源自人类供体的选定的种系分子的抗体序列。人受体分子优选选自与鼠供体分子相同的轻链类型,且是与小鼠供体分子的可变区相同的规范结构类型。针对轻链可变区选择人受体分子的次要考虑包括鼠供体分子和人受体分子之间在CDR长度上的同源性。优选通过V-BASE数据库的同源性搜索,来选择人受体抗体分子,但也可以使用其他数据库如Kabat和公开的NCBI数据库。人源化非人抗体的方法如本文所述,包括下文实施例6。本发明提供了生产结合人OX40的拮抗型抗体或其片段的方法,包括培养包含分离的核酸(编码结合人OX40的拮抗型抗体或其片段)或包含含有分离的核酸(编码结合人OX40的拮抗型抗体或其片段)的载体的宿主细胞,使得核酸表达并生产抗体。优选分离所述抗体。可以使用上文所述的宿主细胞、核酸和载体。可以通过例如本领域普遍已知的(例如,Morrison,S.(1985)Science229:1202)和上文进一步概述的重组DNA技术和基因转染方法的组合,获得核酸表达。例如,为了表达抗体或其抗体片段,可以通过标准的分子生物学技术(例如使用表达目标抗体的杂交瘤的PCR扩增或cDNA克隆)获得编码部分或全长的轻链和重链DNA,并可将所述DNA插入到载体中,例如表达载体。选择的表达载体和表达控制序列与所使用的表达宿主细胞是可相容的。可以将抗体轻链基因和抗体重链基因插入到分离的载体中,或更通常的,两种基因都插入到同一表达载体中。通过标准方法(例如,连接抗体基因片段和载体上互补的限制性位点,或者如果不存在限制性位点时平端连接)将抗体基因插入到表达载体中。通过将本文所述的抗体轻链和重链可变区插入到已编码所需同种型的重链恒定区和轻链恒定区的表达载体中,使得VH区段与载体内的CH1区段有效连接,VK区段与载体内的CL区段有效连接,将其用来产生任何抗体同种型的全长抗体基因。抗OX40抗体的表征和纯化可以使用测量与人OX40的结合的测定,和/或测量阻断OX40与其配体OX40L结合的能力的测定,实施抗体筛选。结合测定的实例是ELISA,特别是使用固定在平板上的人OX40和人Fc的融合蛋白,和应用缀合的二抗检测与融合蛋白结合的抗OX40抗体。阻断测定的例子是基于流式细胞的测定,测量对OX40配体融合蛋白与人CD4细胞上的OX40的结合的阻断。使用荧光标记的二抗,检测与细胞结合的OX40配体融合蛋白的量。该测定搜索由于上清液中的抗体阻断配体融合蛋白与OX40结合的信号降低。阻断测定的另一例子是通过测量胸苷整合,测量由包被在平板上的OX40配体融合蛋白介导的新生人T细胞共刺激的阻断。作为评估抗OX40抗体的功能活性(例如T细胞活化的降低)的测定,可以使用如实施例3和8所述的人混合淋巴细胞反应(MLR)。可以以本领域技术人员已知的多种方式分离或纯化本发明的抗体。标准的纯化方法包括层析技术,包括在大气压下进行的离子交换层析、疏水相互作用层析、亲和层析、大小排阻层析或凝胶过滤层析,和反相层析,或使用系统例如FPLC和HPLC在高压下进行。纯化方法还包括电泳的、免疫学的、沉淀、透析和层析聚焦技术。超滤和透析过滤技术也可与蛋白质浓缩结合使用。为了纯化OX40抗体,可以在例如用于单克隆抗体纯化的转瓶中生长选定的宿主细胞。在用蛋白A琼脂糖(Pharmacia,Piscataway,NJ)的亲和层析之前,可以过滤和浓缩上清液。可以通过凝胶电泳和高效液相层析检测洗脱的IgG,以保证纯度。因而本发明的优选的抗体是结合人OX40的分离的和/或纯化的抗体。免疫缀合物在另一个方面,本发明提供了与治疗剂连接的结合人OX40的拮抗剂OX40抗体或其片段,所述治疗剂例如细胞毒素、药物(例如免疫抑制剂)或放射性毒素。此类缀合物在本文中称为―免疫缀合物‖。包括一种或多种细胞毒素的免疫缀合物被称为―免疫毒素‖。细胞毒素或细胞毒性剂包括对细胞致命(例如杀死细胞)的任何试剂。实例包括紫杉醇(taxol)、细胞松弛素B(cytochalasinB)、短杆菌肽D(gramicidinD)、溴化乙锭(ethidiumbromide)、依米丁(emetine)、丝裂霉素(mitomycin)、依托泊苷(etoposide)、替尼泊苷(tenoposide)、长春新碱(vincristine)、长春碱(vinblastine)、秋水仙碱(colchicin)、多柔比星(doxorubicin)、柔红霉素(daunorubicin)、二羟基炭疽菌素二酮(dihydroxyanthracindione)、米托蒽醌(mitoxantrone)、光神霉素(mithramycin)、放线菌素D(actinomycinD)、1-脱氢睾酮(1-dehydrotestosterone)、糖皮质激素(glucocorticoids)、普鲁卡因(procaine)、丁卡因(tetracaine)、利多卡因(lidocaine)、普萘洛尔(propranolol),和嘌呤霉素(puromycin)及其类似物或同源物。治疗剂还包括例如抗代谢剂(例如,氨甲蝶呤(methotrexate)、6-巯基嘌呤(6-mercaptopurine)、6-硫代鸟嘌呤(6-thioguanine)、阿糖胞苷(cytarabine)、5-氟尿嘧啶氨烯咪胺(5-fluorouracildecarbazine))、烷化剂(例如,氮芥(mechlorethamine、塞替派苯丁酸氮芥(thioepachlorambucil)、美法仑(melphalan)、卡莫司汀(carmustine)(BSNU)和洛莫司汀(lomustine)(CCNU)、环磷酰胺(cyclothosphamide)、白消安(busulfan)、二溴甘露醇(dibromomannitol)、链脲霉素(streptozotocin)、丝裂霉素C(mitomycinC)和顺式-二氯二胺铂(II)(cis-dichlorodiamineplatinum(II)(DDP)顺铂(cisplatin)、蒽环类(anthracyclines)(例如,柔红霉素(daunorubicin)(原道诺霉素(daunomycin)和多柔比星(doxorubicin)、抗生素(例如,更生霉素(dactinomycin)(原放线菌素(actinomycin))、博来霉素(bleomycin)、光神霉素(mithramycin)和安曲霉素(anthramycin)(AMC))和抗有丝分裂剂(例如,长春新碱(vincristine)和长春碱(vinblastine))。可以与本发明抗体连接的治疗性细胞毒素的其他实例包括多卡米星(duocarmycins)、加利车霉素(calicheamicins)、美坦辛(maytansines)和auristatins,及其衍生物。加利车霉素抗体缀合物的实例是可商购的(AmericanHomeProducts)。可以使用本领域可获得的接头技术连接本发明的抗体与细胞毒素。用于缀合细胞毒素与抗体的接头类型的实例包括但不限于腙类、硫醚、酯、二硫化物和含肽接头。可以选择例如对溶酶体区室内的低pH切割敏感的或对蛋白酶切割敏感的接头,所述蛋白酶例如在肿瘤组织中优先表达的蛋白酶,如组织蛋白酶(例如,组织蛋白酶B、C、D)。关于细胞毒素类型、接头类型和将治疗剂与抗体缀合的方法的进一步讨论,还参见Saito,G等人,(2003)Adv.DrugDeliv.Rev.55:199-215;Trail,P.A等人,(2003)CancerImmunol.Immunother.52:328-337;Payne,G.(2003)CancerCell3:207-212;Allen,T.M.(2002)Nat.Rev.Cancer2:750-763;Pastan,I.和Kreitman,R.J.(2002)Curr.Opin.Investig.Drugs3:1089-1091;Senter,P.D.和Springer,CJ.(2001)Adv.DrugDeliv.Rev.53:247-264。本发明的抗体还可与放射性同位素连接,产生细胞毒性的放射药物,也被称为放射性免疫缀合物。可与用于诊断或治疗性抗体缀合的放射性同位素的实例包括但不限于:碘131、铟111、钇90和镥177。本领域已建立了制备放射性免疫缀合物的方法。放射性免疫缀合物的实例是可商购的,包括(EDECPharmaceuticals)和(CorixaPharmaceuticals),相似的方法可用于制备使用本发明抗体的放射性免疫缀合物。本发明的抗体免疫缀合物可用于修饰给定的生物学应答,而药物部分不被视为限制于经典化学治疗剂。例如,药物部分可以是具有理想生物学活性的蛋白质或多肽。此类蛋白质可包括例如酶促活性毒素,或其活性片段,例如相思豆毒蛋白(abrin)、蓖麻毒蛋白A(ricinA)、假单胞菌外毒素(pseudomonasexotoxin)或白喉毒素(diphtheriatoxin);蛋白质例如肿瘤坏死因子或干扰素-γ;或生物学应答修饰剂,例如淋巴因子、白介素-1(IL-1)、白介素-2(IL-2)、白介素-6(IL-6)、粒细胞巨噬细胞集落刺激因子(GM-CSF)、粒细胞集落刺激因子(G-CSF)或其它生长因子。连接此类治疗剂与抗体的技术是普遍已知的,参见例如Arnon等人,"MonoclonalAntibodiesForImmunotargetingOfDrugsInCancerTherapy",inMonoclonalAntibodiesAndCancerTherapy,Reisfeld等人(编著),第243-56页(AlanR.Liss,Inc.1985);Hellstrom等人,"AntibodiesForDrugDelivery",inControlledDrugDelivery(第2版),Robinson等人(编著),第623-53页(MarcelDekker,Inc.1987);Thorpe,"AntibodyCarriersOfCytotoxicAgentsInCancerTherapy:AReview",inMonoclonalAntibodies'84:BiologicalAndClinicalApplications,Pinchera等人(编著),第475-506页(1985);"Analysis,Results,AndFutureProspectiveOfTheTherapeuticUseOfRadiolabeledAntibodyInCancerTherapy",inMonoclonalAntibodiesForCancerDetectionAndTherapy,Baldwin等人(编著),第303-16页(AcademicPress1985)和Thorpe等人,Immunol.Rev.,62:119-58(1982)。在另一个方面,本发明提供了结合人OX40的拮抗剂OX40抗体或其片段,与治疗剂共同施用,例如细胞毒素、药物(例如免疫抑制剂)或放射性毒素。药物组合物在另一个方面,本发明提供了组合物,例如药物组合物,其包括本发明的拮抗剂抗体或其片段和可药用载体。此类组合物可包括一种本发明的抗体和/或免疫缀合物或其组合(例如,两种或多种不同的抗体或免疫缀合物),和/或治疗剂,例如上述细胞毒素、药物(例如免疫抑制剂)或放射性毒素。例如,本发明的药物组合物包括结合靶抗原的不同表位,或具有互补活性的抗体(或免疫缀合物)的组合。本发明的药物组合物还可以在组合疗法中施用,即与其它试剂组合。例如,组合疗法可包括与至少一种其它的抗炎剂或免疫抑制剂组合的本发明的拮抗剂OX40抗体。本文中使用的―可药用载体‖包括任意的和所有的溶剂、分散基质、包衣、抗菌剂和抗真菌剂、等渗的和延迟吸附的介质,以及诸如此类生理学相容的。优选的,载体是适合静脉内、肌内、皮下、不经肠道的、脊柱的或表皮施用的(例如,通过注射或输注)。根据施用途径,活性化合物(即,抗体或免疫缀合物)可包被在材料中,保护所述化合物免受可能灭活所述化合物的酸和其它天然条件的作用。可药用载体包括无菌的含水溶液或分散液,和用于即时制备的无菌的可注射溶液或分散液的无菌粉末。此类基质和介质用于药学活性物质的用途是本领域已知的。除非是与所述活性化合物不相容的任何常规基质或介质,否则可考虑其在本发明的药物组合物中的用途。补充的活性化合物也可被掺入到所述组合物中。在另一个方面,本发明提供了包含免疫缀合物和可药用载体的组合物,所述免疫缀合物包括与治疗剂连接的结合人OX40的拮抗剂抗体或其片段。可如上文所述使用免疫缀合物和治疗剂。在另一个方面,本发明提供了包含本发明的拮抗剂抗体或其片段的组合物,所述拮抗剂抗体或其片段还包括另一种药学活性剂。优选的,另一种药学活性剂是:a)人OX40的另一种拮抗剂,b)镇痛剂和c)免疫抑制剂中的一种或多种,例如糖皮质激素,如泼尼松。本发明的药物组合物还可包括可药用抗氧化剂。可药用抗氧化剂的实例包括:(1)水溶性抗氧化剂,例如抗坏血酸、盐酸半胱氨酸、硫酸氢钠、焦亚硫酸钠、亚硫酸钠等;(2)油溶性抗氧化剂,例如棕榈酰抗坏血酸酯(ascorbylpalmitate)、丁基化羟基苯甲醚(BHA)、丁基羟基甲苯(BHT)、卵磷脂、棓酸丙酯、α生育酚等;和(3)金属螯合剂,例如柠檬酸、乙二胺四乙酸(EDTA)、山梨糖醇、酒石酸、磷酸等。可用于本发明的药物组合物中的合适的含水和不含水载体的实例包括水、乙醇、多元醇(例如甘油、丙二醇、聚乙二醇等)及其合适的混合物、植物油例如橄榄油,和可注射的有机酯例如油酸乙酯。例如通过使用包被材料(如卵磷脂),通过维持所需的颗粒大小(分散体的情形下)和通过使用表面活性剂,可以维持恰当的流动性。这些组合物还可以含有佐剂,例如防腐剂、湿润剂、乳化剂和分散剂。可以通过上文的灭菌程序和通过包含各种抗菌剂和抗真菌剂来确保防止存在微生物,所述抗菌剂和抗真菌剂例如对羟基苯甲酸酯、三氯叔丁醇、山梨酸苯酚(phenolsorbicacid)等。组合物中包括等渗剂也是理想的,例如糖、氯化钠等。此外,通过包括延迟吸附的试剂,例如单硬脂酸铝和明胶,可以产生可注射的药物形式的延长吸附。治疗和其他用途本发明的拮抗剂抗体具有多种体外和体内的诊断和治疗性用途,涉及OX40介导的病症的诊断和治疗。例如,可以在体外或离体条件下,将这些分子施用给培养的细胞,或例如在体内条件下,施用给人类对象,来治疗、预防和诊断多种OX40介导的病症。优选的对象是人,包括患OX40活性介导的病症(OX40介导的病症)的患者。本发明的拮抗剂抗体可有效用于治疗不依赖于其OX40共刺激状态的患者。更优选的对象是人类,包括表达低OX40水平的患者。出于本发明的目的,―患者‖包括人和其他动物,优选哺乳动物和最优选人。因而,本发明的抗体同时具有人类治疗和兽医应用。本发明中的术语―治疗‖意在包括对疾病或病症的治疗性治疗,以及预防性或抑制性的手段。因而,例如,在疾病发动前成功的施用抗体,导致疾病的治疗。作为另一个实例,在疾病的临床表征后成功的施用抗体,来抗争疾病的症状,包括疾病的治疗。―治疗‖还涵盖了在疾病出现后,为了消除所述疾病而施用抗体。在发动后或出现临床症状后成功的施用抗体,伴随临床症状的可能的减轻以及也许疾病的缓解,包括了疾病的治疗。上述―需要治疗的‖包括已患所述疾病或病症的,以及倾向于患所述疾病或病症的哺乳动物,包括要被预防疾病或病症的那些。在特定的实施方案中,拮抗剂抗体在体内用于治疗、预防或诊断多种OX40介导的疾病。因而,本发明提供了在对象中治疗OX40介导的病症的方法,所述方法包括向对象施用治疗有效量的拮抗型抗体或其片段。示例性的OX40介导的病症包括感染(病毒性、细菌性、真菌性和寄生虫性)、与感染相关的内毒性休克、关节炎、类风湿性关节炎、哮喘、慢性阻塞性肺病(COPD)、盆腔炎性疾病、阿兹海默病、炎性肠病、克氏病、溃疡性结肠炎、佩罗尼氏病、乳糜泄(coeliacdisease)、胆囊疾病、潜毛窦、腹膜炎、牛皮癣、脉管炎、术后粘连、中风、I型糖尿病、莱姆病、关节炎、脑膜脑炎、自身免疫性葡萄膜炎、中枢和外周神经系统的免疫介导的炎性病症,如多发性硬化、狼疮(例如系统性红斑狼疮)和格-巴二氏综合征(Guillain-Barrsyndrome)、异位性皮炎、自身免疫性肝炎、纤维性肺泡炎、Grave氏病、IgA肾病、特发性血小板减少性紫癜、Meniere氏病、天疱疮、原发性胆汁性肝硬化、结节病、硬皮病、韦格纳氏肉芽肿(Wegener'sgranulomatosis)、胰腺炎、创伤(手术)、移植物抗宿主病(GVHD)、移植物排斥、心血管疾病,包括缺血性疾病,如心肌梗塞和动脉粥样硬化、血管内凝血(intravascularcoagulation)、骨吸收、骨质疏松、骨关节炎、牙周炎、胃酸过少(hypochlorhydia)和视神经脊髓炎(neuromyelitisoptica)。其他示例性的OX40介导的病症包括感染(病毒性、细菌性、真菌性和寄生虫性)、与感染相关的内毒性休克、关节炎、类风湿性关节炎、哮喘、支气管炎、流感、呼吸道合胞病毒、肺炎、慢性阻塞性肺病(COPD)、特发性肺纤维化(IPF)、隐源性致纤维性肺泡炎(CFA)、原发性纤维性间质性肺炎(idiopathicfibrosinginterstitialpneumonia)、肺气肿、盆腔炎性疾病、阿兹海默病、炎性肠病、克氏病、溃疡性结肠炎、佩罗尼氏病、乳糜泄、胆囊疾病、潜毛窦(Pilonidaldisease)、腹膜炎、牛皮癣、脉管炎、术后粘连、中风、I型糖尿病、莱姆病、关节炎、脑膜脑炎、自身免疫性葡萄膜炎、中枢和外周神经系统的免疫介导的炎性病症,如多发性硬化、狼疮(例如系统性红斑狼疮)和格-巴二氏综合征(Guillain-Barrsyndrome)、异位性皮炎、自身免疫性肝炎、纤维性肺泡炎、Grave氏病、IgA肾病、特发性血小板减少性紫癜、Meniere氏病、天疱疮、原发性胆汁性肝硬化、结节病、硬皮病、韦格纳氏肉芽肿(Wegener'sgranulomatosis)、胰腺炎、创伤(手术)、移植物抗宿主病(GVHD)、移植物排斥、心血管疾病,包括缺血性疾病,如心肌梗塞和动脉粥样硬化、血管内凝血(intravascularcoagulation)、骨吸收、骨质疏松、骨关节炎、牙周炎、胃酸过少和视神经脊髓炎(neuromyelitisoptica)。用本发明的抗体治疗的优选的OX40介导的病症选自多发性硬化、类风湿关节炎、结肠炎、牛皮癣、哮喘、COPD、IPF、移植物抗宿主病(GVHD)、动脉粥样硬化和糖尿病。用本发明的抗体治疗的特别优选的OX40介导的病症是移植物抗宿主病(GVHD)。本发明还提供了用于治疗疼痛、特别是与炎症相关的疼痛的抗体。在一个实施方案中,本发明的抗体可用于检测OX40的水平,或在其膜表面含有OX40的细胞的水平,所述水平之后可与某些疾病症状关联。可选的,抗体可用于抑制或阻断OX40功能,其反过来可与某些疾病症状的预防或缓解相关,从而提示OX40作为疾病的介质。这可以通过在允许抗体和OX40之间形成复合体的条件下,将样品和对照样品与OX40抗体接触来实现。在样品和对照中检测在抗体和OX40之间形成的任何复合体并比较。根据本发明抗体与OX40的特异性结合,本发明抗体可用于特异性检测细胞表面表达的OX40表达,例如可用于检测具有低OX40表达水平的患者。本发明的抗体还可用于通过免疫亲和纯化来纯化OX40。因而本发明还提供了用于检测具有低OX40表达水平的患者的体外筛选方法,包括步骤:(a)从患者血样中纯化外周血单核细胞(PBMC);(b)将PBMC进行流式细胞仪分析;和(c)确定CD4+和/或CD8+T细胞中的OX40阳性细胞数,及将其与对照水平比较。在优选的实施方案中,通过与对照水平相比时,OX40阳性细胞中的表达水平增加至多10%,更优选至多20%,甚至更优选至多30%,表示低的OX40表达水平。KotaniA等人,(2001)Blood,98:3162-4和XiaoyanZ等人,(2005)Clin.Exp.Immunol.143:110-6.中进一步详细描述了确定OX40表达的方法。在另一个实施方案中,可以初步测试本发明抗体与治疗或体外诊断用途相关的结合活性。例如,可以使用流式细胞仪测定,来测试本发明的组合物。本发明还提供了使用拮抗剂抗体或其片段作为药物的用途,和拮抗剂抗体或其片段在制备用于治疗OX40介导的病症的药物中的用途。在其他实施方案中,本发明提供了拮抗剂抗体或其片段,其用作为药物。本发明还提供了拮抗剂抗体或其片段,用在治疗OX40介导的病症的方法中。OX40介导的病症如上文所述。本发明的拮抗剂抗体特别可用于治疗不依赖于患者的OX40共刺激状态的OX40介导的病症。在优选的实施方案中,拮抗剂抗体或其片段可用于在表达低水平OX40的患者中治疗OX40介导的病症。如前所述,本发明的拮抗剂OX40抗体可以和一种或其它多种治疗剂共同施用,例如细胞毒性剂、放射毒性剂或免疫抑制剂。抗体可以与药剂连接(作为如上所述免疫复合物)或者可以与药剂分别施用。在后一种情况下(分别施用),可以在药剂之前、之后或同时施用抗体,或者可以与其它已知的疗法共同施用抗体,所述疗法例如抗癌疗法,例如放射。对于施用抗体,剂量范围为宿主体重的从约0.0001至100mg/kg,更常见从0.01至10mg/kg。示例性的治疗方案需要每周一次、每两周一次、每三周一次、每四周一次、每月一次、每3月一次或每3至6个月一次施用。通常在多种情况施用所述抗体。单次剂量之间的间隔可以是例如每周、每月、每三月或每年。间隔还可以是不规律的,通过测量血液的抗体水平与患者中的靶抗原提示。在一些方法中,调节剂量获得约1-1000μg/ml的血浆抗体浓度,而在一些方法中,约25-300μg/ml。可选的,可以以持续释放的制剂施用抗体,在此情况下需要较低的施用频率。剂量和频率根据患者中的抗体的半衰期而变化。施用的剂量和频率可以根据所述治疗是预防性还是治疗性而改变。在预防性应用中,在较长时间段中,以相对较不频繁的间隔,施用相对较低的剂量。一些患者在其余生中持续接受治疗。在治疗性应用中,有时需要以相对较短的间隔施用相对较高的剂量,直到降低或终止疾病的进展。活性成分的实际剂量(即,本发明的药物组合物中的抗体)是可以改变的,从而获得活性成分的这样的量,对于特定的患者、组合物和施用模式,所述量有效的实现理想的治疗响应,而对患者没有毒性。选定的剂量水平取决于多种药物动力学因子,包括所使用的本发明特定组合物的活性、施用途径、施用时间、所使用的特定抗体的排泄速率、治疗的持续时间、与所使用的特定组合物组合使用的其他药物、化合物和/或材料、年龄、性别、体重、病况、健康状态和受治患者之前的药物史等本领域普遍已知的因素。“治疗有效量‖的本发明的抗OX40抗体优选导致疾病症状严重程度的减少、疾病无症状期的频率和持续期的增加,和/或预防由于病痛导致的损伤或残疾。可以在预测人类功效的动物模型系统中评估化合物治疗OX40介导的病症的能力。可选的,可以通过检验化合物抑制细胞生长的能力,来评估组合物的该特性,可以通过熟练的技术人员已知的测定在体外测量此类抑制作用。本领域的普通技术人员将能够基于如下因素确定所述量,例如对象的尺寸、对象症状的严重程度,和所选定的特定组合物或施用途径。可以利用本领域已知的各种方法中的一种或多种,通过一种或多种施用途径,来施用本发明的抗体或组合物。如熟练的技术人员所理解的,施用的途径和/或方式将根据所需的结果而改变。优选的施用途径包括静脉内、肌内、皮内、腹膜内、皮下、脊柱的或其它不经肠道的施用途径,例如通过注射或输注。更优选的施用途径是静脉内或皮下的。本文中使用的词组―不经肠道的施用‖意指除肠和局部施用以外的施用方式,通常通过注射,包括但不限于静脉内、肌内、动脉内、硬膜内、囊内、眶内、心内、皮内、腹膜内、经气管的、皮下、表皮下、关节内(intraarticular)、囊下(subcapsular)、蛛网膜下(subarachnoid)、脊柱内(intraspinal)、硬膜外(epidural)和胸骨内的注射和输注。可选的,可以通过非不经肠道的途径施用本发明的抗体,例如局部的、表皮的或粘膜的施用途径,例如鼻内、口服、阴道的、经直肠的、舌下的或局部的。制品和试剂盒在本发明的另一个实施方案中,提供了用于治疗OX40介导的病症的、包括本发明的拮抗剂抗体或其片段、组合物或免疫缀合物的制品。制品可包括容器,和与容器相关联或在其上的标签或包装内置物。合适的容器包括例如瓶、管状瓶或注射器。可以用多种材料形成容器,例如玻璃或塑料。容器保留对治疗疾病有效的组合物,可具有无菌的入口(例如,容器可以是静脉内用溶液包或管状瓶,具有可被皮下注射针头穿刺的瓶塞)。组合物中的至少一种活性剂是本文所述的拮抗剂抗体。标签或包装内置物可指示所述组合物可用于治疗的选择的疾病,例如癌症。在一个实施方案中,标签或包装内置物可指示包括拮抗剂抗体的所述组合物可用于治疗OX40介导的病症。此外,制品可包括(a)其中含有组合物的第一容器,其中所述组合物包括本文的拮抗剂抗体,和(b)其中含有组合物的第二容器,其中所述组合物包括拮抗剂抗体以外的治疗剂。本发明的这一实施方案中的制品可进一步包括包装内置物,其指示第一和第二组合物可组合的用于治疗OX40介导的疾病或病症。此类治疗剂可以是上述章节中描述的任何附属疗法(例如,血栓溶解剂、抗血小板剂、化疗剂、抗血管发生剂、抗激素化合物、保心药和/或哺乳动物免疫功能的调节剂,包括细胞因子)。可选的或额外的,制品可还包括包含可药用缓冲液的第二(或第三)容器,例如抑菌性注射用水(BWFI)、磷酸缓冲盐溶液、林格液(Ringer'ssolution)和葡萄糖溶液。还可包括其他对商业和用户观点而言理想的材料,包括其他的缓冲液、稀释剂、滤器、针头和注射器。还落入本发明范围内的包括包含本发明的抗体、组合物或免疫缀合物和使用说明的试剂盒。试剂盒可进一步含有一种或多种其他试剂,例如免疫抑制剂、细胞毒性剂或放射毒性剂,或一种或多种其他的本发明的拮抗剂抗体(例如,结合OX40抗原中不同于第一拮抗剂抗体的表位的拮抗剂抗体,其具有补充活性)。不需进一步描述,可以相信本领域的普通技术人员可以利用上述说明和下列示例性实施例,制造和利用本发明的物质和实施要求保护的方法。下列工作实施例供促进本发明的实践所用,并非被视为对本发明其余内容的任何方式的限制。实施例实施例1:生成和筛选小鼠抗人OX40抗体为了生产重组人OX40-Fc蛋白,从imaGenes购买了人TNFRSF4的cDNA(克隆编号:RZPDB737H0329D;Berlin,Germany)。使用该cDNA作为模板,PCR扩增人TNFRSF4胞外结构域的DNA编码区(SEQIDNO:11)。在分离的PCR反应中,通过PCR扩增人IgG1的Fc区(EU位置223-451),添加5’GSGGG接头和3’SA-6xHis接头和限制性位点用于克隆。然后,使用重叠延伸PCR,用侧翼引物融合所得到的两种产物,添加限制性位点用于后续克隆入基于Invitrogen的质粒pcDNA3.1(-)(InvitrogenAG,Basel,Switzerland,货号V795-20)的经修饰哺乳动物表达载体中,所述质粒含有人CMV启动子和美国专利5924939所述的Ig供体受体片段(第一内含子)、OriP序列(KoonsMD等人,(2001)JVirol.75(22):10582-92)、SV40增强子和与胃泌素终止子融合的SV40polyA,如KimD,等人,(2003)Biotechnol.Prog.19(5):1620-2所述。该重组质粒允许在哺乳动物细胞中表达人TNFRSF4胞外结构域-Fc融合蛋白,并通过人TNFRSF4蛋白的天然信号肽驱动分泌到细胞培养基中。为了生产重组蛋白质,使用jetPEITM转染试剂(Polyplus-transfectionS.A.,Strasbourg,France;distributor:Brunschwig,Basel,Switzerland),将上述重组载体转染到适应悬浮的HEK293细胞(ATCC编号CRL1573)中。在5天后收集细胞培养上清液,并使用在FPLC系统(GEHealthcareEuropeGmbH,Glattbrugg,Switzerland)上操作的蛋白A亲和纯化柱(HiTrapProteinA琼脂糖柱;GEHealthcareEuropeGmbH,Glattbrugg,Switzerland)进一步纯化。为了生产重组人OX40-his蛋白,通过PCR扩增人TNFRSF4的胞外区(SEQIDNO:11),添加3’GSG-6xHis接头和限制性位点用于克隆。之后,将PCR产物克隆到上述修饰型pcDNA3.1(-)质粒中。该重组质粒允许在哺乳动物细胞中表达人OX40-his蛋白,通过人TNFRSF4的天然信号肽驱动分泌到细胞培养基中。为了生产蛋白质,使用jetPEITM转染试剂(Polyplus-transfectionS.A.,Strasbourg,France;distributor:Brunschwig,Basel,Switzerland),将上述重组载体转染到适应悬浮的HEK293细胞(ATCC编号CRL1573)中。在转染5天后收集细胞培养上清液,并使用在FPLC系统(GEHealthcareEuropeGmbH,Glattbrugg,Switzerland)上操作的Ni2+-NTA亲和纯化柱(HiTrapNi2+-NTA琼脂糖柱;GEHealthcareEuropeGmbH,Glattbrugg,Switzerland)纯化。通过SDS-PAGE判断,重组人OX40-Fc和OX40-his蛋白是95%纯的,并在使用前进一步交换缓冲液为磷酸盐缓冲液(PBS)。将溶解在PBS中的重组人OX40-Fc蛋白与等体积的Stimune佐剂(Prionics,Switzerland,ref:7925000)混合,并制备乳状液。将乳状液转移到0.5mL胰岛素注射器(BDPharmingen,Allschwil,Switzerland)中,并用50μg乳化蛋白在后足垫、尾基部和颈部皮下免疫BALB/c动物(Harlan,Netherlands)。2周后,用相同的抗原量和相同的注射途径重复免疫。使用包被了重组人OX40-his蛋白的平板,通过直接ELISA评估在免疫的小鼠血清中存在的循环型抗人OX40抗体。将一系列稀释度(1:100至1:109)的不同小鼠血清添加到平板中,并使用山羊抗小鼠H+L完整分子-HRP(Sigma-AldrichChemieGmbH,Buchs,Switzerland)检测结合的抗体。在处死前3天,在表现出最佳抗人OX40-IgG血清滴度的动物中,用50μg不含佐剂的抗原进行最后一次皮下加强。使动物安乐死,收集腹股沟、腋下、手臂、腘和髋部的淋巴结,用2个25G针头,在DNA酶(RocheDiagnostics(Schweiz)AG,Rotkreuz,Switzerland)和胶原酶(RocheDiagnostics(Schweiz)AG,Rotkreuz,Switzerland)溶液中分散淋巴结结构,制备单细胞悬浮液。用聚乙二醇1500(RocheDiagnostics(Schweiz)AG,Rotkreuz,Switzerland),以比例7:1(融合配偶体与收获的淋巴结细胞)融合单细胞悬浮液与骨髓瘤细胞系X63AG8.653(小鼠BALB/c骨髓瘤细胞系;ATCC登录号:CRL1580;KearneyJF等人,(1979)J.Immunol.123(4):1548-1550)。将融合的细胞种植到含有小鼠巨噬细胞的96孔平底平板中,所述平板含有补充了10%胎牛血清(FBS,PAALaboratories,Pasching,Austria)、2mML-谷氨酰胺、100U/ml(BiochromAG,Germany)青霉素、100μg/ml链霉素(BiochromAG,Germany)、10mMHEPES(InvitrogenAG,Basel,Switzerland)、50μMβ-巯基乙醇(Sigma-AldrichChemieGmbH,Buchs,Switzerland)、HAT(Sigma-AldrichChemieGmbH,Buchs,Switzerland)和1%生长因子(Hybridokine,Interchim/Uptima,France)的DMEM-10培养基(InvitrogenAG,Basel,Switzerland)。通过ELISA从融合物中筛选约800个孔,寻找所存在的识别人OX40和阻断人OX40L与其受体结合的小鼠IgG。扩展阳性细胞,并进行2轮亚克隆。收集细胞,克隆和测序重链和轻链。实施例2:从杂交瘤细胞中克隆和测序抗OX40抗体的VH和VL链对于每个阳性的选定杂交瘤,制备总RNA,逆转录成cDNA,通过PCR分别扩增VH和VL基因。将这些PCR产物连接到拯救载体(pDrive载体;QIAGENAG,Hombrechtikon,Switzerland;货号231124)中,允许对单个PCR产物DNA测序,确定选定杂交瘤的单克隆性或多克隆性。该载体允许在含有IPTG和X-gal的LB琼脂平板上进行蓝白选择(由于LacZα–肽对X-gal的降解,没有插入物的克隆是蓝色的)。从阳性(白色)细菌克隆制备重组质粒,并使用载体骨架特异性的标准DNA测序引物(M13rev、M13fwd、T7或SP6)测序。最终将DNA序列亚克隆到用于在哺乳动物细胞中重组表达目标抗体的表达载体中。分离RNA根据生产商的操作规程,使用QIAGEN的RNeasyMiniKit(QIAGENAG,Hombrechtikon,Switzerland;货号74106),从2-10x106细胞中分离总RNA;使用NanoDropND-1000分光光度计(WITECAG,Littau,Switzerland)定量样品。一步RT-PCR将上述总RNA制品进一步逆转录成cDNA,使用2个不同的简并引物的混合物,通过PCR扩增VH和VL片段,每种简并引物的混合物都允许回收小鼠免疫球蛋白重链可变片段和可变重链连接区的所有不同亚类,或回收所有的小鼠免疫球蛋白轻链κ可变片段和可变轻链κ连接区。用于逆转录和扩增的引物通过Microsynth(Balgach,Switzerland)合成,并HPLC纯化(表1-4)。使用QIAGEN一步RT-PCR试剂盒(QIAGENAG,Hombrechtikon,Switzerland;货号210212)同时实施逆转录和PCR扩增。由于技术使用特定的引物,将每种mRNA样品重复处理2份,允许分别逆转录和扩增VH或VL片段。将2μg总RNA溶解在不含RNA酶的水中,至终体积30μl,与10μl的QIAGENOneStepRT-PCR缓冲液的5x储液、2μl浓度为10mM的dNTP混合物,3μl浓度为10μM的引物混合物和2μl的QIAGENOneStepRT-PCREnzymeMix混合。然后将最终混合物放在PCR管中,并在PCR热循环仪(BioRadiCyclerversion4.006,Bio-RadLaboratoriesAG,Reinach,Switzerland)中循环,循环使用下列设置:50℃,30min95℃,15min40个循环:94℃,30sec55℃,30sec72℃,1min然后72℃,10min保持在4℃pDrive克隆在2%琼脂糖凝胶上运行PCR产物。在DNA电泳后,从琼脂糖凝胶上切下目标片段(~450bp),并使用Macherey-NagelNucloSpinExtractII试剂盒250(Macherey-Nagel,Oensingen,Switzerland;货号740609.250)进一步抽提。为了DNA测序,将抽提的PCR产物克隆到上述拯救载体(pDrive载体,QIAGENAG,Hombrechtikon,Switzerland;货号231124)中,并转化到E.coliTOP10菌株(InvitrogenAG,Basel,Switzerland;货号C404006)中。Miniprep抽提在1.5ml补充了100μg/ml青霉素的LuriaBertani(LB)培养基中,37℃(振荡250RPM)培养阳性克隆过夜,接种在Macherey-NagelSquare-wellBlock平板(Macherey-Nagel,Oensingen,Switzerland;货号740488.24)上。第二天,使用NucleoSpinMulti-8Plasmid试剂盒(Macherey-Nagel,Oensingen,Switzerland;货号740620.5),实施DNAminiprep抽提。测序和序列分析将样品送去DNA测序服务公司Fasteris(Plan-les-Ouates,Switzerland),进行DNA测序。使用标准引物:M13rev、M13fwd、T7、SP6(表5)。为了分析DNA序列,使用CloneManager9专业版(Scientific&EducationalSoftware,NC,USA)和BioEditSequenceAlignmentEditor(Hall,TA(1999)Nucl.Acids.Symp.Ser.41:95-98)。克隆表达载体用于重组嵌合抗体表达为了在哺乳动物细胞中重组表达,使用基于装配的PCR方法,将分离的鼠VH和VL片段装配成嵌合免疫球蛋白。这些嵌合抗体由重链和轻链组成,在所述重链中,鼠重链可变结构域与人IgG1重链恒定结构域(γ1、铰链区、γ2和γ3区)融合,在所述轻链中,鼠轻链可变结构域与人κ恒定结构域(Cκ)融合。将PCR装配的鼠可变部分和人恒定部分克隆到修饰型哺乳动物表达载体中,所述载体是基于实施例1提及的Invitrogen的修饰型pcDNA3.1(-)载体,差别在于使用人免疫球蛋白轻链κ前导肽驱动蛋白质分泌。为了免疫球蛋白候选物的蛋白质生产,将等量的重链和轻链载体DNA共转染到适应悬浮的HEK293细胞(ATCC编号CRL1573)中。在转染5天后收集细胞培养上清液,并使用在FPLC系统上操作的蛋白A亲和纯化柱(HiTrap蛋白A琼脂糖柱)(都来自GEHealthcareEuropeGmbH,Glattbrugg,Switzerland)纯化。表1:引物MixVH–反向表2:引物MixVH–正向表3:引物MixVL–反向表4:引物MixVL–正向表5:测序引物实施例3:抗人OX40抗体的生物学表征OX40-特异性抗体检测ELISA:通过直接ELISA确定杂交瘤和重组抗体候选者的抗体滴度、特异性和生产。简而言之,用100μl在PBS中2μg/ml的重组人OX40-his包被96孔微滴度板(CostarUSA,distributorVWRAG,Nyon,Switzerland),OX40-his蛋白的产生参见实施例1。在4℃孵育平板过夜,然后用PBS2%BSA(牛血清白蛋白,PAALaboratories,Pasching,Austria)在室温(RT)封闭1小时。去除封闭溶液,加入杂交瘤上清液或纯化的抗体。在RT孵育平板30分钟,然后用PBS0.01%Tween-20(Sigma-AldrichChemieGmbH,Buchs,Switzerland)洗涤9次,加入1:1000稀释的辣根过氧化物酶(HRP)标记的山羊抗小鼠H+L-检测抗体(Sigma-AldrichChemieGmbH,Buchs,Switzerland)。为了检测具有人Fc的重组嵌合抗体(参见实施例2),使用1:1000稀释的HRP标记的兔抗人IgG抗体(Sigma-AldrichChemieGmbH,Buchs,Switzerland)作为检测抗体。在室温(RT)孵育平板30分钟,用PBS0.01%Tween-20洗涤9次,向平板加入TMB底物(Bio-radLaboratoriesAG,Reinach,Switzerland),通过6分钟后添加H2SO4终止反应。然后通过显微平板读板器读取450nm吸光值(Biotek,USA;distributor:WITTECAG,Littau,Switzerland)。图1A显示了嵌合1D4抗体和嵌合2F8抗体识别OX40-his包被的蛋白。OX40L阻断ELISA:如下生成重组人OX40配体蛋白(OX40L):从imaGenes(Berlin,Germany)购买人TNFSF4的cDNA(克隆名称:IOH46203),用侧翼的限制性位点扩增人TNFSF4配体的胞外部分(氨基酸51-183)(根据UniprotQ6FGS4序列编号)。之后,将得到的在其5’涵盖ASA接头和8-Histag序列的PCR产物克隆到Invitrogen的修饰型pREP4载体版本(InvitrogenAG,Basel,Switzerland)中,该载体携带了CMV启动子、牛生长激素多聚腺苷酸化,和驱动重组蛋白分泌的鼠VJ2C前导肽。为了生产重组蛋白质,使用jetPEITM转染试剂(Polyplus-transfectionS.A.,Strasbourg,France;distributor:Brunschwig,Basel,Switzerland),将重组载体转染到适应悬浮的HEK293细胞(ATCC编号CRL1573)中。在5天后收集细胞培养上清液,并使用在FPLC系统(GEHealthcareEuropeGmbH,Glattbrugg,Switzerland)上操作的蛋白A亲和纯化柱(HiTrap蛋白A琼脂糖柱;GEHealthcareEuropeGmbH,Glattbrugg,Switzerland)纯化。为了确定生成的抗OX40抗体是否可以阻断OX40L与OX40受体的结合,研发了阻断ELISA。用100μl在PBS中2μg/ml的重组人OX40-Fc(参见实施例1),包被96孔微滴度板(Costar,USA;distributorVWRAG,Nyon,Switzerland)。在4℃孵育平板过夜,然后用PBS2%BSA在RT封闭1小时。去除封闭溶液,向平板加入杂交瘤上清液或纯化的抗体。5分钟后,向每个孔加入50μl的0.04mg/ml生物素化的重组人OX40L。在RT孵育平板60分钟,然后用PBS0.01%Tween-20洗涤9次,加入1:2000稀释的HRP-链霉抗生物素(Sigma-AldrichChemieGmbH,Buchs,Switzerland)。在RT孵育平板30分钟,用PBS0.01%Tween-20洗涤9次,向平板加入TMB底物(Bio-radLaboratoriesAG,Reinach,Switzerland),6分钟后添加H2SO4终止反应。然后通过显微平板读板器读取450nm吸光值(Biotek,USA;distributor:WITTECAG,Littau,Switzerland)。图1B显示了嵌合1D4抗体能够以剂量依赖性的方式阻断OX40和OX40L之间的相互作用,而嵌合2F8抗体不能阻断OX40和OX40L之间的相互作用。人混合淋巴细胞反应(MLR)将2个不同供体的血液收集到3个10mL以柠檬酸作为抗凝剂的S-Monovette(Sarstedt,Nümbrecht,Germany)中。使用1号供体的细胞作为效应子细胞,使用2号供体的细胞作为靶细胞。按生产商的说明,使用50mLBlood-Sep-Filter管(供应商:Brunschwig,Basel,Switzerland),纯化2个供体的PBMC(外周血单核细胞)。用不含FBS的RoswellParkMemorialInstitute(RPMI,PAALaboratories,Pasching,Austria)培养基洗涤细胞2次。用50μg/ml丝裂霉素C(Sigma-AldrichChemieGmbH,Buchs,Switzerland)在37℃孵育靶细胞30分钟。用不含FBS的RPMI洗涤细胞3次,并按1x106细胞/mL重悬在RPMI,其含10%FBS(PAALaboratories,Pasching,Austria)、2mML-谷氨酰胺(Lonza,Leuven,Belgium)、100U/ml青霉素、100μg/ml链霉素(BiochromAG,Berlin,Germany)中。在96孔U型底显微平板(TPP,Trasadingen,Switzerland)中,按每个孔100μl的终体积分配50’000个靶细胞和80’000个效应子细胞。向孔内加入100μl抗体稀释液。平板在5%CO2温箱中37℃孵育7天。在开始MLR7天后,用0.5μCi的3H胸苷(PerkinElmer)脉冲细胞。脉冲18小时后,收获细胞,并在Wallacβ计数器上定量整合的放射性。图2显示了嵌合1D4抗体能够以剂量依赖性的方式,比阳性对照更高程度的阻断MLR。实施例4:流式细胞仪对抗人OX40抗体与人和其他动物的活化外周血单核细胞(PBMC)的结合人细胞从LaChaux-de-Fonds,Switzerland的血液收集中心(CentredeTransfusionSanguineetLaboratoiredeSérologie,rueSophie-Mairet29,CH-2300),收集含有人白细胞的过滤器。通过用60mL含有10U/mLliquemin(DrossapharmAG,Lucern,Switzerland)的PBS反冲洗,从过滤器移出细胞。然后,按生产商的说明,用50mLBlood-Sep-Filter管(供应商:Brunschwig,Basel,Switzerland)纯化PBMC。用含FBS(PAALaboratories,Pasching,Austria)的RoswellParkMemorialInstitute(RPMI,PAALaboratories,Pasching,Austria)培养基洗涤细胞3次。按3x106细胞/mL将细胞重悬在含RPMI,10%FBS(PAALaboratories,Pasching,Austria)、2mMUltraglutamine(Lonza,Leuven,Belgium)、100U/ml青霉素、100μg/ml链霉素(BiochromAG,Berlin,Germany)、10μg/ml植物凝聚素(PHA;Sigma-AldrichChemieGmbH,Buchs,Switzerland)+100U/mLrHuIL-2(Proleukin,Novartis,Basel,Switzerland)的24孔板(TPP,Trasadingen,Switzerland)中。48小时后,收集细胞,并如下所述通过流式细胞仪分析。在RPMI,10%FBS中培养HPB-ALL细胞(T急性淋巴细胞性白血病细胞系,来自DeutscheSammlungvonMikroorganismenundZellkulturenGmbH,Braunschweig,Germany)。将2x105细胞分散在96孔V型底板(TPP,Trasadingen,Switzerland)中,在1300rpm离心3分钟;丢弃上清液,收集细胞,并如下所述通过流式细胞仪分析。将如上制备的PBMC和HPB-ALL细胞重悬在50μl含有5μg/mL嵌合1D4抗体,或5μg/mL恰当的同种型对照,或20μlPE标记的商业抗人OX40抗体(克隆L106,BDBiosciences,Allschwil,Switzerland的FACS缓冲液(PBS,2%FBS,10%Versene(Invitrogen,USA))中。在冰上孵育细胞30分钟,洗涤2次,并重悬在50μlFACS缓冲液中。使用1/200稀释的抗人IgG-藻红蛋白-PE(BDBiosciences,Allschwil,Switzerland)检测嵌合1D4抗体和同种型对照抗体。在冰上孵育细胞15分钟,洗涤1次,重悬在400μlFACS缓冲液中,并在FACS仪器上分析(Cyan,BeckmanCoulterInternationalS.A.,Nyon,Switzerland)。食蟹猴原代细胞将食蟹猴的全血(获得自ProfessorEricRouiller,LaboratoryofNeurophysiology,UniversityofFribourg,Fribourg,Switzerland)收集到柠檬酸试管(BDBiosciences,Allschwil,Switzerland)中。将2mLPBS与3mL血液混合,并将混合物置于10ml的85:15Ficoll:PBS混合物(GEHealthcareEuropeGmbH,Glattbrugg,Switzerland)的顶层。在室温下不间断的离心样品20分钟。收集PBMC层,用PBS洗涤3次。将细胞按3x106细胞/mL重悬在Dulbecco’sModifiedEagle培养基(DMEM,PAALaboratories,Pasching,Austria)中,其含10%FBS(PAALaboratories,Pasching,Austria)、非必需氨基酸(PAALaboratories,Pasching,Austria)、1mM丙酮酸钠(PAALaboratories,Pasching,Austria)、2mMUltraglutamine(Lonza,Belgium)、100U/ml青霉素(BiochromAG,Germany)、100μg/ml氯霉素(BiochromAG,Germany)。在24孔平板(TPP,Trasadingen,Switzerland)中分配1mL细胞悬浮液,并加入10ug/mlPHA(PHA/M,Sigma-AldrichChemieGmbH,Buchs,Switzerland)、100U/mLrHuIL-2(Proleukin,Novartis,Basel,Switzerland)。在5%CO2温箱中37℃孵育细胞50小时。收集活化的PBMC,并重悬在PBS/2.5%FBS(FACS缓冲液)中。在96孔V型底平板中,按50μlFACS缓冲液分配五万个细胞,并向孔中加入25μg/ml的生物素化抗人OX40嵌合1D4抗体、或生物素化同种型对照抗体、或在绵羊中产生的生物素化商业抗人OX40(BDBiosciences,Allschwil,Switzerland)。在冰上孵育样品20分钟,然后用冰冷的FACS缓冲液洗涤细胞2次,然后在冰上用1:20稀释的链霉抗生物素-PE(BDBiosciences,Allschwil,Switzerland)孵育细胞15分钟。用FACS缓冲液洗涤细胞1次,然后重悬在300μl的FACS缓冲液中。向每个样品添加体积2μl的碘化丙啶(PI;Sigma-AldrichChemieGmbH,Buchs,Switzerland),排除死细胞。通过流式细胞仪分析细胞(Cyan,BeckmanCoulterInternationalS.A.,Nyon,Switzerland)。图3A和3B显示了嵌合1D4抗体能够识别分别在人和食蟹猴的活化淋巴细胞表面上表达的OX40,因而提供了对药物研发高度需要的交叉反应特性。实施例5:通过表面等离振子共振(SPR)分析嵌合1D4抗体与人OX40受体胞外结构域的动力学结合亲和力常数使用实施例1所述的重组组氨酸标记的人OX40受体胞外结构域作为分析物,在蛋白A捕获的抗体上测量动力学结合亲和力常数(KD)。在BIAcore2000(GEHealthcare-BIAcore,Uppsala,Sweden)上室温进行测量,并用BiaEvaluation软件(BIAcore;v4.1)分析。通过注射35μl的1:1N-羟基硫代琥珀酰亚胺(NHS)/1-乙基-3-[3-二甲基氨基丙基]碳化二亚胺盐酸盐(EDC)溶液(v/v;5μl/min流速;流路1和2),激活CM5研究级传感芯片(GEHealthcareEuropeGmbH,Glattbrugg,Switzerland;BR-1000-14)。将蛋白A(ref.P7837;Sigma-AldrichChemieGmbH,Buchs,Switzerland)在醋酸缓冲液pH4.5(GE,BR-1003-50;低于pI一个pH单位)中稀释为50μg/ml的终浓度,之后通过在流路1和2注射35μl(5μl/min),将蛋白A固定在前述激活的CM5传感芯片上;这对应于约1500应答单位(RU)。然后,通过注射35μl乙醇胺溶液(5μl/min),将蛋白A-CM5传感芯片去活化。最后,注射2次10μl甘氨酸溶液(GE,ref.BR-1003-54;10mM;pH1.5),释放未交联的蛋白A分子。在亲和力测量前,通过用不同的流速向定量的蛋白A捕获的抗体上注射固定浓度的分析物(5、15、30、50、75μl/min;2min),实施质量传递限制试验。对不同流速的结合速率(on-rate)斜率分析表示了质量传递。为了测量亲和力,在HBS-EP缓冲液(GE,ref.BR-1001-88;0.01MHepes,0.15MNaCl,EDTA3mM,0.005%SurfactantP20,pH7.4)中,将储藏在1xPBS缓冲液中的嵌合1D4抗体稀释至15nM的终浓度。之后,将10μl该稀释储液注射在蛋白ACM5芯片的流路2(30μl/min)上,达到200-250RU。在这一捕获步骤后,以30μl/min的流速,在流路1和2(流路1用作参照)注射不同浓度(50nM至0.4μM)的重组组氨酸标记的人OX40受体胞外结构域。在每种结合事件后,用甘氨酸缓冲液pH1.5注射1min(10μl/min),再生表面。测量值(传感图(sensorgram):fc2-fc1)与具有质量传递的2:1二价分析物模型最佳拟合。为了说明在每次开始测量时蛋白A捕获抗体的实验误差的原因,在所有拟合中将Rmax值设为局部。解离时间是至少300-600秒。实施重复测量2次,包括0浓度样品作为参照。Chi2值表示每个点的实验数据和参照数据之间的方差总和;而剩余的点表示在拟合的每个点的实验数据和参照数据之间的差异。Chi2值和剩余值都用于评估实验数据和每个结合模型之间的拟合质量。用固定在蛋白A传感芯片上的捕获的嵌合1D4抗人OX40抗体重复测量2次,以重组组氨酸标记的人OX40受体胞外结构域作为分析物。KD值在91至116nM之间,Chi2值<1.25。实施例6:小鼠单克隆抗体1D4的人源化本文描述了人源化抗人OX40小鼠抗体1D4,包括人受体框架的选择、回复突变和基本保留和/或改善人CDR移植受体框架结合特性的突变。设计改造的可变区使用同源性匹配来选择移植1D4CDR的人受体框架。可使用数据库(例如,人和小鼠免疫球蛋白基因座的种系可变基因的数据库(IMGT数据库(theinternationalImMunoGeneTicsinformationLefrancMP等人,(1999)NucleicAcidsRes.27(1):209-12;RuizM等人,(2000)NucleicAcidsRes.28(1):219-21;LefrancMP(2001)NucleicAcidsRes.29(1):207-9;LefrancMP(2003)NucleicAcidsRes.31(1):307-10;LefrancMP等人,(2005)Dev.Comp.Immunol.29(3):185-203;KaasQ等人,(2007)BriefingsinFunctionalGenomics&Proteomics,6(4):253-64)或VBASE2(RetterI.等人,2005,NucleicAcidsRes.,33,DatabaseissueD671-D674)或Kabat数据库(JohnsonG.等人,2000,NucleicAcidsRes.,28,第214-218页))或出版物(例如,Kabat等人,见上文))鉴别鼠重链和轻链V区所属的人亚族,并确定用作受体分子的最适人种系框架。在这些用作受体的亚族中选择重链和轻链可变序列(VH和VL)可基于序列同源性和/或CDR1和CDR2区的结构匹配,来帮助在移植后保留六个CDR的恰当的相对呈现。例如,IMGT数据库的使用提示在1D4重链可变结构域框架和人重链可变结构域亚类2的成员之间的良好的同源性。对种系序列观察到了CDR和框架序列的最高同源性和同一性:IGHV2-70*10(SEQIDNO:19)、IGHV2-70*01(SEQIDNO:20)、IGHV2-70*13(SEQIDNO:21)、IGHV2-5*09(SEQIDNO:22)和IGHV2-70*11(SEQIDNO:23);其在达到CDR3的整个序列都具有高于73%的序列同一性,IGHV2-70*10、IGHV2-70*01和IGHV2-70*13具有74%序列同一性;而IGHV2-5*09和IGHV2-70*11分别具有73.5%和73%的序列同一性。使用同一方法,1D4轻链可变结构域序列表现出与人轻链可变结构域κ亚类3的成员的良好同源性。对种系序列观察到了CDR和框架序列的最高同源性和同一性:IGKV3-11*01(SEQIDNO:24)(65.3%同一性)、IGKV1-39*01(SEQIDNO:25)(64.9%同一性)、IGKV1D-39*01(SEQIDNO:26)(64.9%同一性)、IGKV3-11*02(SEQIDNO:27)(64.2%同一性)和IGKV3-20*01(SEQIDNO:28)(62.5%同一性)。作为人源化过程的起点,选择人IGHV2-70*10(SEQIDNO:19)和IGKV3-11*01(SEQIDNO:24)可变结构域作为1D4CDR的受体。由于在其框架一区中与1D4的优良同源性,因此超过其他人重链可变结构域,选择IGHV2-70*10。制备第一个人γ1同种型的人源化抗体(见下文)。抗体涵盖了人-小鼠杂交重链可变结构域和人-小鼠杂交轻链可变结构域。杂交重链可变结构域是基于人重链可变结构域IGHV2-70*10的,其中种系CDR1和2分别被1D4重链CDR1和2替换。从上述IMGT检索中鉴别出与人受体框架最匹配的JH片段序列。得到的人-小鼠杂交重链可变序列(具有人IGHV2-70*10框架区、1D4小鼠CDR和与人受体最佳匹配的JH)在本文中被称为重链可变结构域VH1,具有SEQIDNO:29。类似的,用于该第一个人源化抗体候选物的人-小鼠杂交轻链可变结构域具有人IGKV3-11*01框架区、1D4小鼠CDR和与人受体最佳匹配的JK,在本文中被称为轻链可变结构域VL1,具有SEQIDNO:30。所述涵盖VH1和VL1的第一个人源化抗体在本文中被缩写为VH1/VL1抗体。生产第一人源化抗体原型由GENEARTAG(Regensburg,Germany)以scFv模式合成VH1和VL1的编码DNA序列(cDNA),从而允许单个DNA序列涵盖两个可变结构域(SEQIDNO:31)。通过PCR从该scFv构建体重新得到单个可变结构域cDNA,并使用PCR装配技术进一步在上游装配其相应的恒定结构域cDNA序列。最后,将完整的重链和轻链cDNA连接到独立的载体中,所述载体基于携带CMV启动子和牛生长激素多聚腺苷酸化信号的修饰型pcDNA3.1载体(Invitrogen,CA,USA)。轻链特异性载体通过使用BamHI和BsiWI限制性酶位点,将目标轻链可变结构域cDNA连接到κ轻链恒定结构域cDNA之前,允许表达人κ同种型轻链;同时使用BamHI和SalI限制性酶位点改造重链特异性载体,允许将目标重链可变结构域cDNA连接到编码人IGHG1CH1、IGHG1铰链区、IGHG1CH2和IGHG1CH3恒定结构域的cDNA序列之前。在两个重链和轻链表达载体中,分泌是由含有BamHI位点的小鼠VJ2C前导肽驱动的。BsiWI限制性酶位点位于κ恒定结构域中;而SalI限制性酶位点则可见于IGHG1CH1结构域中。通过使用聚乙稀亚胺(PEI,Sigma,Buchs,Switzerland),共转染等量的重链和轻链载体到适应悬浮的HEK293-EBNA1细胞(目录编号:CRL-10852)中,瞬时生产VH1/VL1抗体。通常,用含有50μg编码重链的表达载体和50μg编码轻链的表达载体的DNA-PEI混合物,转染100ml密度为0.8-1.2百万细胞/ml的细胞悬浮液。当编码抗体基因的重组表达载体被导入宿主细胞中时,进一步培养细胞为期4至5天来生产抗体,以允许分泌到培养基(EX-CELL293,HEK293-无血清培养基,Sigma,Buchs,Switzerland)中,所述培养基补充了0.1%普流罗尼酸(pluronicacid)、4mM谷氨酰胺和0.25μg/ml遗传霉素。使用重组的蛋白A层流基质(GEHealthcareEuropeGmbH,Glattbrugg,Switzerland),从无细胞上清液中纯化VH1/VL1抗体,并在测定前缓冲液交换为磷酸缓冲盐溶液。如实施例5所述,通过SPR测量与人OX40的结合。经移植的人框架的回复突变由于从1D4小鼠抗体直接移植的CDR导致不与人OX40结合的候选物(表6和图4),所以开始其中用鼠残基替换人残基的诱变。被称为回复突变的该过程是单克隆抗体人源化中最不可预测的程序。必需鉴别和选择小鼠抗体中需要保留的关键框架残基,从而在保留亲和力的同时使人源化抗体的潜在免疫原性最小化。表7、表8和图5显示了在小鼠和人抗体框架间不同的残基(Kabat编号)。可影响CDR构象或可变结构域间包装的残基是特别令人感兴趣的,因其可能对抗体亲和力具有最大影响。为了鉴别可影响大部分CDR构象和/或可变结构域间包装的残基,使用设为自动模式的结构同源性-建模服务器SWISS-MODEL(ArnoldK等人,(2006)Bioinformatics,22(2):195-201;http://swissmodel.expasy.org),计算可变结构域的VH1-VL1对的3D模型。模型分析允许基于位置对CDR区和/或重链-轻链可变结构域包装的推定影响,选择一小组位置。该小组位置由以下可变重链位置和下列可变轻链位置组成,可变重链位置:23、35b、48、50、60和62;下列可变轻链位置:1、33、34、46、47、54、56和71(Kabat编号)。除上述回复突变外,在一些候选物中缺失可见于VH1/VL1抗体中的轻链位置Y31。使用标准诱变和上述方法,在VH1/VL1抗体序列中,制备基于重链和轻链取代的各种组合的其他人源化候选物。如实施例5所述,通过SPR测定人源化抗体候选物的结合亲和力。表6显示了基于上述单取代或取代组合的一些人源化抗体的产量和结合特性。在所显示的28个抗体中,9个候选物没有表现出与人OX40的结合,另外的9个一组则具有弱至差的结合。根据SPR,基于VL9的人源化抗体表现出与人OX40最一致至弱的结合。仅2个抗体——VH6/VL9和VH7/VL9表现出与人OX40良好的结合。两种人源化抗体都在可变重链位置23、35b、50、60和62,以及可变轻链位置33、34、46、47和71(Kabat编号)上具有回复突变。除上述回复突变外,VH6/VL9和VH7/VL9都受益于去除轻链位置31。令人惊讶的是,H7/VL9具有比1D4嵌合抗体和VH6/VL9变体更好的针对人OX40的亲和力。表9中概括了这些人源化抗体的结合亲和力。通过差异扫描量热法分析选定的人源化抗OX40抗体的热稳定性使用差异扫描量热法(DSC)测量人源化抗体的热稳定性。单克隆抗体融解曲线是其同种型的特征(GarberE&DemarestSJ(2007)Biochem.Biophys.Res.Commun.355:751-7),然而,即使在全长IgG中,也可以轻易的鉴别FAB片段的中点融解温度。使用这类FAB部分的中点融解监控人源化候选物的单克隆稳定性。在VP-DSC差异扫描量热仪(MicroCal,Northampton,UK)上进行量热法测量。细胞体积是0.128ml,加热速率是200℃/h,过压保持在65p.s.i.。所有抗体都使用PBS(pH7.4)中1mg/ml的浓度。通过与含有相同缓冲液但省略了抗体的重复2次的样品比较,估计抗体的摩尔热容。使用标准的方法分析部分摩尔热容和融解曲线。在使用软件Originv7.0中的Non-TwoState模型进一步分析前,将热图进行基线校正和浓度归一化。人源化变体VH6/VL9FAB片段76.3℃表现出单相变(singletransition),形状和幅度与协作解折叠一致,这一般是在紧密折叠的FAB片段中观察到的,表示改造过程对于保留FAB稳定性是成功的。总而言之,人源化变体表现出良好的热稳定性。表6:人源化抗人OX40抗体表6(续):人源化抗人OX40抗体表7:比较1D4和人受体重链可变IGHV2-70,10框架表8:比较1D4和人受体轻链可变IGKV3-11*01框架表9:选定的人源化和嵌合抗OX40抗体的结合特征实施例7:人源化抗OX40抗体的表位表征为了表征人源化抗OX40抗体的表位,使用多个人-大鼠OX40嵌合蛋白,绘制VH6/VL9抗体的人OX40胞外区的定义结构域。制备人-大鼠OX40嵌合蛋白和ELISA根据实施例1所述方法,将大鼠和人-大鼠OX40蛋白配制成Fc融合蛋白。为了ELISA,在高结合性96孔板(Coastar)上包被2μg/mL在PBS中的OX40蛋白,4℃过夜。在用VH6/VL9抗体或同种型对照抗体孵育前,用PBS2%牛血清白蛋白(BSA)封闭平板。然后洗涤平板,并用山羊抗人IgF(ab’)2片段特异性-HRP(JacksonImmunoResearchEuropeLtd,Newmarket,UK)孵育。在洗涤后,用TMB底物(Bio-RadLaboratoriesAG,Reinach,Switzerland)孵育平板,揭示抗体结合。通过加入2MH2SO4终止反应,在SynergyHT2分光光度计(Biotek,USA;供应商:WITTECAG,Littau,Switzerland)上读取450nM处的光密度(OD450nM)。结果不论其来源,OX40胞外区分为4种结构模块,称为结构域1、2、3和4(CompaanDM&HymowitzSG(2006)Structure,14(8):1321-30)。通过交换人和大鼠序列之间的4个结构域的一个或多个,构建对应于人OX40的胞外区的嵌合OX40蛋白(人TNFRSF4的氨基酸29-214,根据UniprotP43489序列编号)。例如,嵌合RHRROX40蛋白对应于大鼠OX40胞外区,其中第二结构域被相应的人结构域序列替换。实施结合ELISA,测试VH6/VL9抗体对人OX40胞外区(缩写为HHHH,SEQIDNO:11)、大鼠OX40胞外区(缩写为RRRR,SEQIDNO:52)和四种人-大鼠嵌合蛋白:RHRR(SEQIDNO:53)、HRRR(SEQIDNO:54)、HHRR(SEQIDNO:55)和RRHH(SEQIDNO:56)的反应性。图7显示了该ELISA的结果。作为该表位定位实验的先决条件,VH6/VL9抗体显示出与人OX40蛋白而不与大鼠OX40蛋白结合,表示与大鼠OX40没有交叉反应。发现VH6/VL9抗体结合RHRR和HHRR,而不结合HRRR或RRHH,表示VH6/VL9表位定位在人OX40胞外区的第二结构域中。实施例8:VH6/VL9抗体通过杀死和阻断机制阻断人混合淋巴细胞反应在单边同种异体混合淋巴细胞反应(MLR)中,测试VH6/VL9抗体抑制体外免疫反应的效力。MLR是同种反应性T细胞活化和增殖的体外模型(O’FlahertyE等人,(2000)Immunology,100(3):289-99;DuPontB&HansenJA(1976)Adv.Immunol.23:107-202)。当混合来自2个不相关供体的外周血单核细胞(PBMC)时,T细胞通过识别同种异体主要组织相容性(MHC)分子而激活。该活化导致T淋巴细胞增殖。MLR反应广泛用于证实T细胞靶向的免疫抑制药物的效果(BromelowKV等人,(2001)J.Immunol.Methods,247(1-2):1-8)。免疫抑制药物(如环孢霉素)主要通过抑制T细胞激活而发挥作用。除了测试VH6/VL9抗体的阻断效应外,还研究了细胞毒作用机制(如抗体依赖性细胞毒作用(ADCC))对MLR抑制的作用。该测定中试验了VH6/VL9抗体的3种不同的抗体模式:IGHG1模式(在本文中被称为VH6/VL9)、无岩藻糖基化的IGHG1(IgG1)模式(在本文中被称为无岩藻糖基化的VH6/VL9)和IGHG4(IgG4)模式(在本文中被称为VH6/VL9IGHG4S228P)。已知IGHG1(IgG1)抗体胜任细胞毒作用机制,如ADCC。已知由于对细胞毒性细胞(如天然杀伤细胞(NK细胞))上表达的FcγRIIIa更高的亲和力,无岩藻糖基化的IGHG1抗体表现出增强的ADCC活性(MizushimaT等人,(2011)GenesCells,16(11):1071-80)。相反,已知IGHG4(IgG4)抗体不具有这类Fc-介导的细胞毒作用机制,如ADCC。VH6/VL9抗体的配制在实施例6中描述的重链特异性载体中,用编码IGHG4CH1、具有S228P取代的IGHG4铰链区、IGHG4CH2和IGHG4CH3恒定结构域的cDNA序列,替换编码IGHG1CH1、IGHG1铰链区、IGHG1CH2和IGHG1CH3恒定结构域的cDNA序列,实现具有取代S228P的IGHG4免疫球蛋白配制。通过标准的PCR诱变技术,向人IGHG4重链cDNA模板中导入取代S228P。得到的重链具有SEQIDNO:57。按WO2010/095031公开的实施例14描述的操作规程,生产无岩藻糖基化的VH6/VL9IGHG1抗体。混合淋巴细胞反应将2个不同的人类供体的血液收集到3个10mL以柠檬酸作为抗凝剂的S-Monovette(Sarstedt,Nümbrecht,Germany)中。按生产商的说明,使用50mLBlood-Sep-Filter管(供应商:Brunschwig,Basel,Switzerland),纯化2个人类供体的外周血单核细胞(PBMC)。用不含FBS的RoswellParkMemorialInstitute(RPMI,PAALaboratories,Pasching,Austria)培养基洗涤细胞2次。用50μg/ml丝裂霉素C(Sigma-AldrichChemieGmbH,Buchs,Switzerland)在37℃孵育30分钟制备2个供体的刺激子细胞。然后,用不含FBS的RPMI洗涤细胞3次,并按1x106细胞/mL重悬在RPMI中,其含10%FBS(PAALaboratories,Pasching,Austria)、2mML-谷氨酰胺(Lonza,Leuven,Belgium)、100U/ml青霉素和100μg/ml链霉素(BiochromAG,Berlin,Germany)。在96孔U型底显微平板(TPP,Trasadingen,Switzerland)中,将应答者细胞重悬在RPMI,10%FBS、L-谷氨酰胺、100U/ml青霉素和100μg/ml链霉素中。按每个孔100μl的终体积分配50’000个刺激子细胞和80’000个应答者细胞。向孔内加入100μl抗体稀释液(或仅培养基)。平板在5%CO2温箱中37℃孵育7天。在最后18个小时中,用0.5μCi的3H胸苷(PerkinElmer,BaselSwitzerland)脉冲细胞。在滤网式(filtermat)过滤器(PerkinElmer)上收获细胞,并在Wallacβ计数器(PerkinElmer)上定量整合的放射性。结果图8显示的结果证实了VH6/VL9抗体能够有效抑制2个不同个体(应答者)的MLR,EC50值约100ng/mL。结果还显示了依赖于所使用的抗体模式的不同的应答,在不同个体的MLR中观察到了阻断和细胞毒作用机制贡献的差异。IGHG1和IGHG4模式有效抑制了应答者1(图8A)的T细胞反应性,表示细胞毒作用机制对应答者1不是关键的。相反,IGHG4模式仅能够在高浓度时不良阻断应答者2的MLR(图8B),而在较低浓度时非常快的丢失了其效力,而IGHG1模式能够实现大于60%的抑制,暗示对于应答者2,杀伤机制对大部分的抑制效果负责。该作用模式的差异可能源自这样的事实,即,MLR反应中的T细胞激活、增殖和存活是依赖于个体之间的OX40共刺激信号而改变的,取决于同种异体反应性的程度和其他可能的共刺激信号。对于较弱依赖于OX40来源的共刺激信号的个体,通过ADCC机制消除活化型T细胞是VH6/VL9的主要作用机制。令人惊讶的是,无岩藻糖基化的IGHG1模式在两个应答者中都表现出非常强的抑制MLR的能力。这一观察结果强调了这样的事实:即使阻断机制足以实现抑制MLR,增加或增强杀伤机制也可以改善抗OX40抗体的抑制作用。当不论患者的OX40共刺激状态而治疗OX40介导的病症时,这类增强作用是特别有效的,例如当患者具有低的OX40表达水平时。实施例9:VH6/VL9抗体阻断异种移植物抗宿主病异种移植物抗宿主反应(GVH)是用于在人类患者在骨髓移植后观察到的同种异体移植物抗宿主病(GVHD)的模型。GVH反应是由移植物免疫细胞介导的急性免疫应答,是同种异体或异种MHC识别导致的对宿主环境的攻击(MurphyWJ等人,(1996)Semin.Immunol.8(4):233-41)。T淋巴细胞是GVH反应的主要效应子细胞。在基于用人PBMC重建的SCID小鼠的异种GVHD模型中试验VH6/VL9抗体的免疫抑制能力。在该模型中,人PBMC,主要是T淋巴细胞对小鼠宿主细胞发动了强烈的应答。反应显著导致了严重的皮肤和肠道炎症,伴有减重。该模型的最相关的读出是动物的存活。方法在腹腔内用三千万个人PBMC重建前,亚致死的辐射动物(SCID小鼠)。还通过每周2次注射TMβ1抗体,耗尽小鼠的鼠NK细胞。自PBMC注射2天前开始,每周5次以连续剂量静脉内给予VH6/VL9抗体、或载体的治疗。用10mg/kg或1mg/kg的载体(PBS)或VH6/VL9抗体,或8mg/kg的(人IgG1的Fc组分与人可溶性TNF受体2融合的融合蛋白,Amgen-Pfizer)治疗动物。每周3次检查动物,并对GVHD症状评分,包括减重、腹泻、毛皮问题和一般行为。如果认为症状太严重,则基于伦理学方法处死动物。结果图9显示,即使在1mg/kg的较低剂量下,VH6/VL9抗体也非常有效的抑制GVHD反应。令人惊讶的是,VH6/VL9抗体展现出比改良的效果,是公认的人类GVHD疗法(XhaardA等人,(2011)Bull.Cancer,98(8):889-99;SimpsonD(2001)ExpertOpin.Pharmacother.2(7):1109-17)。用1或10mg/kg的VH6/VL9抗体治疗的动物的中位存活时间比载体治疗组长四倍(表10),比长2倍。此外,这一结果强调VH6/VL9抗体不具有激动剂活性,因为已报道过在同种异体小鼠GVHD模型中,激动型抗OX40抗体使GVHD恶化(ValzasinaB等人,(2005)Blood,105(7):2845-51;BlazarBR等人,(2003)Blood,101(9):3741-8),而本研究中没有观察到该事件。表10:标注的治疗组的中位存活时间(天)载体:仅PBS。1D4:GBR830-1D4抗体;是临床产品。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1