基因G20E03、其编码的蛋白及其在提高烟草植株抗大豆孢囊线虫病中的应用的制作方法
本发明属于烟草抗病技术领域,尤其涉及一种基因g20e03、其编码的蛋白及其在提高烟草植株抗大豆孢囊线虫病中的应用。
背景技术:
大豆孢囊线虫(heteroderaglycines,soybeancystnematode,scn)是植物内寄生线虫,主要侵染大豆(glycinemax)等豆科植物,烟草(nicotianatabacum)是其非寄主。但近年来,在山东地区发现一个特殊的能够侵染烟草的scn群体(tobaccopopulationofsoybeancystnematode,scnt)。利用形态学及分子生物学等鉴定手段,将山东省寄生烟草的孢囊线虫鉴定为大豆孢囊线虫(heteroderaglycines)。受scnt侵染的烟草根系,其分叉变多,后期会着生小米粒大小的白色或黄色球形颗粒(孢囊线虫的雌虫),部分受侵染根系出现褐色坏死,最后整条根腐烂、干枯。孢囊线虫危害烟草的根系,不仅会导致烟叶产量损失和品质下降,而且会加重烟草上其他病害(如烟草枯萎病)的发生,已成为严重制约我国山东、河南等地烟区生产的重要因素之一。
目前防治孢囊线虫的方法主要有轮作、化学防治和选育抗病品种,但是由于轮作的局限性和化学防治给生态环境造成的巨大污染,以及抗线虫种质资源的匮乏,寻找高效的、可持续的大豆孢囊线虫防治方法是目前植物寄生线虫研究领域的重点。
通过基因技术获得抗病植株是防治农作物病害的有效手段,然而,由于现有技术对于scnt的研究仅停留在对比scn与scnt的致病差异性上,由于scnt特殊的致病力,使得通过基因技术筛选得到强致病因子并获得抗大豆孢囊线虫病的烟草植株仍存在很高的技术难度。
技术实现要素:
针对现有技术存在的不足之处,本发明所要解决的技术问题是克服现有技术难点,系统运用基因技术筛选得到强致病因子并获得抗大豆孢囊线虫病的烟草植株,从而有效解决烟草病害的技术问题,提供一种能够大量表达烟草大豆孢囊线虫病效应蛋白基因的dsrna,及有效防治烟草大豆孢囊线虫病的方法。
为解决所述技术问题,本发明采用的技术方案为:
本发明提出基因g20e03,所述基因g20e03的基因全长序列如seqidno.1所示。
优选的,所述基因g20e03的基因全长序列包括5’端非编码区、3’端非编码区和cdna区,所述cdna区的基因序列如seqidno.2所示。
优选的,所述基因g20e03由以下方法克隆得到:
使用trizol法提取大豆孢囊线虫烟草群体的2龄幼虫rna,并反转录为cdna;
根据scnt的转录组测序结果设计引物对g20e03-part-f和g20e03-part-r,以上述cdna为模板进行pcr扩增,得到pcr产物,对所述pcr产物进行纯化及t-a连接,得到连接产物,将所述连接产物转入大肠杆菌中,随机挑取单菌落,利用所述引物对g20e03-part-f和g20e03-part-r对所述单菌落进行pcr筛选,得到阳性单克隆菌落并测序;
根据上述的g20e03基因部分序列,设计5’端非编码区和3’端非编码区基因的特异引物g20e03-5’-r和g20e03-3’-f,pcr扩增,5’端扩增出260bp的片段,3’端扩增出666bp的片段;
对上述序列进行拼接,克隆得到所述基因g20e03的基因全长序列。
本发明另一方面提供了以上任一技术方案所述的基因g20e03编码的蛋白,所述蛋白的氨基酸序列如seqidno.3所示。
本发明还提供了上述蛋白在提高烟草植株抗大豆孢囊线虫病中的应用。
本发明还提供了以上任一技术方案所述的基因g20e03在提高烟草植株抗大豆孢囊线虫病中的应用。
优选的,包括以下步骤:通过转基因技术获得基因g20e03的rnai转基因烟草植株,在转基因烟草植株中表达基因g20e03的dsrna,使大豆孢囊线虫基因g20e03沉默,缓解基因g20e03对烟草植株的免疫抑制作用,提高烟草植株的抗大豆孢囊线虫病性。
优选的,所述通过转基因技术获得基因g20e03的rnai转基因烟草植株包括以下步骤:
构建重组表达载体pfgc5941-rnai-g20e03;
将重组表达载体pfgc5941-rnai-g20e03转化农杆菌lba4404,得到重组农杆菌;
通过农杆菌介导的叶盘法转化烟草,进行组织培养,分别利用引物g20e03-part-f和g20e03-part-r、bar-f和bar-r检测均为阳性的转基因烟草植株即为g20e03的rnai转基因烟草植株。
优选的,所述构建重组表达载体pfgc5941-rnai-g20e03包括以下步骤:
以大豆孢囊线虫cdna为模板,采用race法克隆g20e03基因,得到完整的基因序列;
设计引物对g20e03-orf-r-f和g20e03-orf-r-r,pcr扩增得到含xbal和bamhi酶切位点的orf反向序列,分别使用限制性内切酶xbal和bamhi进行酶切,纯化回收后,连接至经限制性内切酶xbal和bamhi酶切的植物干扰载体pfgc5941,获得重组表达载体pfgc5941-g20e03;
设计引物对g20e03-orf-f-f和g20e03-orf-f-r,pcr扩增得到含ncoi和swai酶切位点的orf正向序列,分别使用限制性内切酶ncoi和swai对重组表达载体pfgc5941-g20e03进行酶切,纯化回收后,连接至经限制性内切酶ncoi和swai酶切的载体pfgc5941-g20e03,获得重组表达载体pfgc5941-rnai-g20e03。
优选的,通过转基因技术获得基因g20e03的rnai转基因烟草植株还包括:
将t0代转基因烟草种子采用种子穴盘育苗,并分别利用特异性引物g20e03-part-f和g20e03-part-r、bar-f和bar-r进行pcr检测,得到t1代转基因烟草;
按照上述方法逐代培养,得到纯合的转基因株系。
与现有技术相比,本发明的有益效果在于:
本发明提供了一种能够提高烟草植株抗大豆孢囊线虫病的基因g20e03及其编码的蛋白,并证明了该基因编码的蛋白在大豆孢囊线虫寄生烟草中发挥重要作用,能够抑制烟草免疫反应,通过转基因技术实现对基因g20e03的沉默表达,从而提高烟草抗孢囊线虫的能力,对培育抗大豆孢囊线虫的烟草品种奠定了良好的理论和实践基础。
附图说明
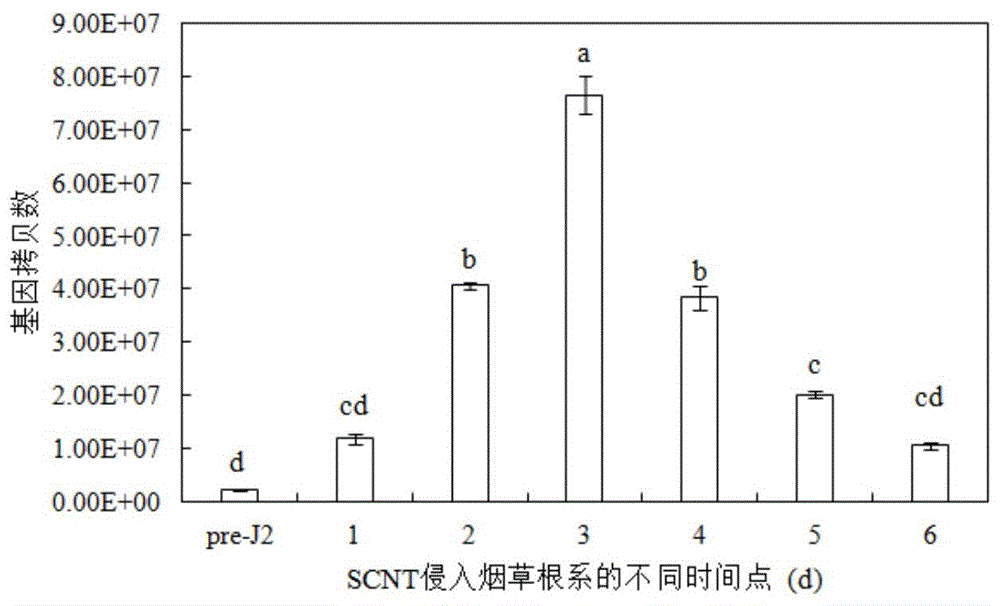
图1为g20e03基因在侵入寄主前后不同时间点下的表达量,其中,pre-j2为scnt侵染前2龄幼虫,小写字母代表差异显著性p<0.05;
图2为扩增g20e03基因部分序列的电泳结果,其中,m:dl2000marker,1:g20e03部分片段;
图3为race克隆g20e03基因电泳结果,其中,a:扩增g20e03基因5’区序列的电泳结果,m:dl2000marker,1:g20e035’片段,b:扩增g20e03基因3’区序列的电泳结果,m:dl2000marker,2:g20e033’片段;
图4为扩增g20e03基因orf区序列的电泳结果,其中,m:dl2000marker,1:g20e03orf片段;
图5为g20e03免疫抑制功能的验证;
图6为scnt侵入烟草后的g20e03基因沉默效率检测,其中,wt为野生型烟草,l17和l26为rnai株系烟草;
图7为野生型和干涉株系l17和l26接种scnt35d后的孢囊和雌虫数量,其中,wt、l17和l26分别代表接种scnt35d的野生型和rnai株系。
具体实施方式
下面将结合附图对本发明具体实施例中的技术方案进行详细、完整的描述。显然,所描述的实施例仅仅是本发明总的技术方案的部分具体实施方式,而非全部的实施方式。基于本发明的总的构思,本领域普通技术人员所获得的所有其他实施例,都落于本发明保护的范围。
本发明提出基因g20e03,所述基因g20e03的基因全长序列如seqidno.1所示。
在一优选实施例中,所述基因g20e03的基因全长序列包括5’端非编码区、3’端非编码区和cdna区,所述cdna区的基因序列如seqidno.2所示。
在一优选实施例中,所述基因g20e03由以下方法克隆得到:
使用trizol法提取大豆孢囊线虫烟草群体的2龄幼虫rna,并反转录为cdna;
根据scnt的转录组测序结果设计引物对g20e03-part-f和g20e03-part-r,以上述cdna为模板进行pcr扩增,得到pcr产物,对所述pcr产物进行纯化及t-a连接,得到连接产物,将所述连接产物转入大肠杆菌中,随机挑取单菌落,利用所述引物对g20e03-part-f和g20e03-part-r对所述单菌落进行pcr筛选,得到阳性单克隆菌落并测序;
根据上述的g20e03基因部分序列,设计5’端非编码区和3’端非编码区基因的特异引物g20e03-5’-r和g20e03-3’-f,pcr扩增,5’端扩增出260bp的片段,3’端扩增出666bp的片段;
对上述序列进行拼接,克隆得到所述基因g20e03的基因全长序列。
上述技术方案中,以g20e03-part-f和g20e03-part-r作为特异性引物对,进行pcr扩增获得cdna片段,以g20e03-5’-r和g20e03-3’-f作为特异性引物对,进行pcr扩增获得5’端非编码区片段及3’端非编码区片段,需要说明的是,上述引物对并非常规选择得到的引物对,而是根据scnt2龄幼虫的转录组测序获得的序列进行特异性引物的设计。
本发明另一方面提供了以上任一技术方案所述的基因g20e03编码的蛋白,所述蛋白的氨基酸序列如seqidno.3所示。
本发明还提供了上述蛋白在提高烟草植株抗大豆孢囊线虫病中的应用。在转基因烟草植株内的基因g20e03的dsrna,通过线虫取食进入线虫体内,引发所述基因的失活,显著降低了孢囊线虫的寄生能力,从而抑制孢囊线虫的侵染、寄生、繁殖和传播,进而提高烟草植株的抗病性。效应蛋白在植物寄生线虫和寄主互作过程中发挥重要作用,既决定着线虫对寄主的致病强度,也是植株抗性产生与否的关键因子。线虫合成的效应蛋白通过口针注入寄主植物体内,从而有利于线虫的侵染、寄生和生长发育。需要说明的是上述基因g20e03编码的蛋白g20e03的氨基酸序列如seqidno.3所示。同时,经过一个或一个以上氨基酸残基的取代和/或缺失和/或添加并且与大豆孢囊线虫寄生相关的衍生蛋白,也在本发明的范畴之内。
本发明还提供了以上任一技术方案所述的基因g20e03在提高烟草植株抗大豆孢囊线虫病中的应用。需要说明的是,现有技术利用形态学及分子生物学等鉴定手段,将山东省寄生烟草的孢囊线虫鉴定为大豆孢囊线虫(heteroderaglycines),该群体为scnt,且针对scn与scnt的致病差异性上进行了研究,但是,在此基础上能够获得scnt的强致病基因,仍然需要付出大量的创造性的劳动,包括:首先通过比较转录组学得到13个差异表达的编码效应蛋白的基因,并从这些基因中确定最有可能导致烟草植株感染大豆孢囊线虫病的基因;然后,研究该基因是否调控寄主植物的免疫反应;为进一步明确效应蛋白g20e03对scnt侵染烟草能力的影响,还开展了g20e03基因的rnai转基因植物的研究。具体的:
首先经过比较转录组学得到13个差异表达的编码效应蛋白的基因,利用荧光定量pcr研究scnt2龄幼虫的上述13个基因在线虫侵染寄主不同时间下的表达量。将scnt的2龄幼虫分别接种烟草1、2、3、4、5、6d后,分离烟草根系的2龄幼虫,提取其rna,荧光定量pcr检测scnt侵入根系的2龄幼虫13个基因的绝对表达量。结果表明,与侵染前2龄幼虫(pre-j2)相比较,烟草根系中的2龄幼虫的g20e03基因表达量在第2、3、4、5、6d时的表达量显著高于侵染前的表达量。其中侵染3d时的表达量最高,较pre-j2相比,高达33.15倍之多(如图1所示)。由此推测该基因与线虫早期侵染寄主相关。
在此研究基础上,为确认该基因是否调控寄主植物的免疫反应,首先利用race法克隆该基因全长,获得该基因的全序列(如图2、3、4所示);其次构建含g20e03基因cdna序列的pgr107载体,并将重组载体转化到谷枯菌中,分别用1ml注射器将阴性对照gfp基因与g20e03基因分别在本氏烟叶背面的左右两侧高压渗透,7d后,可以观察到在注射gfp的区域有明显的坏死(programmedcelldeath,pcd);而注射g20e03的叶片显示为没有任何坏死或相对较小的坏死(如图5所示)。以上结果表明了g20e03能够抑制烟草的免疫反应。
为进一步确认效应蛋白g20e03对scnt侵染烟草能力的影响,本发明还开展了g20e03基因的rnai转基因植物的研究。本发明成功构建了g20e03基因的rnai植物表达载体,转化农杆菌lba4404后,通过烟草叶盘法将该重组载体转入烟草,在烟草中产生了g20e03的dsrna,该植株能显著抑制大豆孢囊线虫的寄生,获得了强抗孢囊线虫的烟草,为防治烟草大豆孢囊线虫病提供了坚实的理论基础和实践指导。
与上述效应基因g20e03杂交且编码与大豆孢囊线虫寄生相关蛋白的dna分子,或者与上述大豆孢囊线虫效应基因g20e03有90%以上同源性且编码大豆孢囊线虫寄生相关蛋白的dna分子,也在本发明的保护范围之内。
在一优选实施例中,包括以下步骤:通过转基因技术获得基因g20e03的rnai转基因烟草植株,在转基因烟草植株中表达基因g20e03的dsrna,使大豆孢囊线虫基因g20e03沉默,缓解基因g20e03对烟草植株的免疫抑制作用,提高烟草植株的抗大豆孢囊线虫病性。inplantarna干扰(rnainterference,rnai)是通过在植物寄主体内表达具发夹结构的dsrna,线虫通过口针取食寄主,从而进入线虫体内,诱发靶标基因的rnai效应,导致靶标基因表达沉默,从而对靶标基因所在线虫的生长、发育和繁殖产生影响,甚至会产生致死作用。由于大豆孢囊线虫专性寄生的特点,rnai作为一种反向遗传学技术,在线虫基因功能的研究中得到广泛的应用。
在一优选实施例中,所述通过转基因技术获得基因g20e03的rnai转基因烟草植株包括以下步骤:
构建重组表达载体pfgc5941-rnai-g20e03;
将重组表达载体pfgc5941-rnai-g20e03转化农杆菌lba4404,得到重组农杆菌;
通过农杆菌介导的叶盘法转化烟草,进行组织培养,分别利用引物g20e03-part-f和g20e03-part-r、bar-f和bar-r检测均为阳性的转基因烟草植株即为g20e03的rnai转基因烟草植株。
所述pfgc5941是一种植物rnai载体,将含有大豆孢囊线虫烟草群体效应基因g20e03的正反向序列分别克隆到植物表达载体中,重组表达载体转化农杆菌后,通过烟草叶盘法转化到烟草体内,可以在烟草中产生所述基因的dsrna,当孢囊线虫侵染该植株后,dsrna进入孢囊线虫体内,导致所述基因的mrna降解,从而引发所述基因的失活,显著降低了孢囊线虫的寄生能力,从而抑制孢囊线虫的侵染、寄生、繁殖和传播。
在一优选实施例中,所述构建重组表达载体pfgc5941-rnai-g20e03包括以下步骤:
以大豆孢囊线虫cdna为模板,采用race法克隆g20e03基因,得到完整的基因序列;
设计引物对g20e03-orf-r-f和g20e03-orf-r-r,pcr扩增得到含xbal和bamhi酶切位点的orf反向序列,分别使用限制性内切酶xbal和bamhi进行酶切,纯化回收后,连接至经限制性内切酶xbal和bamhi酶切的植物干扰载体pfgc5941,获得重组表达载体pfgc5941-g20e03;
设计引物对g20e03-orf-f-f和g20e03-orf-f-r,pcr扩增得到含ncoi和swai酶切位点的orf正向序列,分别使用限制性内切酶ncoi和swai对重组表达载体pfgc5941-g20e03进行酶切,纯化回收后,连接至经限制性内切酶ncoi和swai酶切的载体pfgc5941-g20e03,获得重组表达载体pfgc5941-rnai-g20e03。
在一优选实施例中,通过转基因技术获得基因g20e03的rnai转基因烟草植株还包括:
将t0代转基因烟草种子采用种子穴盘育苗,并分别利用特异性引物g20e03-part-f和g20e03-part-r、bar-f和bar-r进行pcr检测,得到t1代转基因烟草;
按照上述方法逐代培养,得到纯合的转基因株系。
为了更清楚详细地介绍本发明实施例所提供的基因g20e03、其编码的蛋白及其在提高烟草植株抗大豆孢囊线虫病中的应用,下面将结合具体实施例进行描述。
烟草k326:购自中国农业科学院烟草研究所;大豆孢囊线虫烟草群体:青岛农业大学植物线虫研究室保存;载体pfgc5941、农杆菌菌株gv3101和lba4404均为青岛农业大学植物线虫研究室保存。
实施例1
13个编码效应蛋白的基因在侵入寄主前后不同时间点下的表达量分析
1、烟草植株的培育
采用种子穴盘育苗,并用营养土对烟草种子覆盖,置于温室中培养,光照16h/黑暗8h,温度25℃,湿度65%。待幼苗长至具有2-3片真叶时将其移栽至装有200g灭菌砂土的培养钵中(培养钵为塑料水杯,杯口直径6.5cm、杯底直径4.5cm、杯高7.5cm)。
2、scnt的2龄幼虫的接种
线虫接种:具体步骤为:在烟草幼苗根际周围用玻璃棒打4个孔,孔深至根系水平,每孔接入约500条scnt的2龄幼虫(scntj2),每杯共接种2000条scntj2,用水淋冲后覆盖灭菌土封严,烟苗置于温室内培育。
3、荧光定量pcr检测13个编码效应蛋白的基因在侵入寄主前后不同时间点下的表达量分析
分别接种烟草1、2、3、4、5、6d后,分离烟草根系的2龄幼虫,trizol法提取其rna,荧光定量pcr检测scnt侵入根系的2龄幼虫13个基因的相对表达量。结果表明,与侵染前2龄幼虫(pre-j2)相比较,烟草根系中的2龄幼虫的g20e03基因表达量在第2、3、4、5、6d时的表达量显著高于侵染前的表达量。其中侵染3d时的表达量最高,较pre-j2相比,高达21.22倍之多。由此推测该基因在scnt寄生烟草中发挥重要作用,与线虫早期侵染寄主相关,从而调控寄主植物的免疫反应。
实施例2
大豆孢囊线虫烟草群体g20e03蛋白及其编码基因的克隆
1、使用trizol法提取大豆孢囊线虫烟草群体的2龄幼虫rna,并反转录为cdna。
2、g20e03基因部分片段的获得
根据转录组测序结果的g20e03基因序列设计引物对g20e03-part-f和g20e03-part-r,扩增g20e03基因的部分片段。pcr扩增体系:cdna2μl,两条引物各2μl,buffer2.5μl,dntp2μl,taqase0.5μl,ddh2o14μl。pcr扩增程序:94℃5min,30×(94℃45s,55℃30s,72℃1min),72℃5min,4℃保存。琼脂糖电泳检测扩增结果如图2所示。
pcr产物纯化及t-a连接:将pcr产物利用qiagen公司胶回收试剂盒qiaquickgelextrationkit(cat.nos.28704),扩增产物进行胶回收,并经1%琼脂糖凝胶电泳检测回收纯化的pcr产物,并将回收产物连接到载体pmdtm18-t上,16℃连接16h,连接体系见表1。
表1连接体系
连接产物转化大肠杆菌及测序:将连接好的g20e03-part-pmdtm18-t重组载体利用全式金公司的感受态细胞(trans5achemicallycompetentcelllot#j2421103)通过冻融法转入大肠杆菌中:于冰上向50μl感受态细胞中加入10μl载体连接产物,冰浴30min,42℃热激45s,冰上2min,再向离心管中加入500μl无菌的lb液体培养基(不含抗生素),混匀后振荡培养1h(37℃,200rpm),细菌复苏后,将菌液梯度涂板,37℃倒置过夜培养,挑取单个转化子于lb液体培养基(含抗生素)培养5-6h(菌液od值大约为1.0),利用引物对g20e03-part-f和g20e03-part-r进行pcr筛选阳性克隆,并将阳性克隆菌液送至生工测序,同时将阳性菌液加甘油后-80℃超低温保存。
3、g20e03基因5’和3’端的扩增
根据转录组测序结果设计g20e03基因部分片段的“5’和3’”基因特异引物(g20e03-5’-r和g20e03-3’-f),根据
4、g20e03基因orf序列的获得
序列拼接获得g20e03基因全长序列,如seqidno.1所示。设计g20e03orf区引物(g20e03-orf-f和g20e03-orf-r),扩增g20e03基因的orf。pcr扩增体系和程序同实施例2第2部分内容。琼脂糖电泳检测扩增结果如图4所示,得到597bp大小的dna片段。
实施例3
g20e03基因抑制植物免疫功能的研究
1、植物材料的培养
取一定量的沙子和基质,按照一定比例混合到一起,180℃灭菌60min。分装于各个托盘约三分之二处,将烟草种子分别播种入各孔中,每隔2天浇一次营养液,当幼苗长出6-8片真叶时为试验的最佳时期。
2、重组载体的构建
根据g20e03基因的cdna序列,设计带有smai酶和sali酶双酶切位点的g20e03(g20e03-gr-f和g20e03-gr-r)引物,进行pcr扩增并纯化;双酶切上述pcr纯化产物及pgr107载体,1%琼脂糖凝胶电泳分离,切胶回收目的条带,使g20e03与pgr107载体连接(t4连接),构建g20e03-pgr107重组载体。连接产物转化大肠杆菌后送至生工测序。测序正确后转化农杆菌gv3101。
3、含重组载体g20e03-pgr107的农杆菌菌液液注射烟草
采用高压渗透法,利用1ml注射器将阴性对照pgr107-gfp与g20e03-pgr107菌液分别在本氏烟叶背面的左右两侧进行注射(pgr107-bax在24h后注射),7d后,观察烟草表型。每次注射10株烟草叶片,三次重复,共计注射30个烟草叶片。如图5所示,可以观察到在注射gfp的区域有明显的坏死(programmedcelldeath,pcd);而注射g20e03的叶片显示为没有任何坏死或相对较小的坏死。该结果表明了g20e03能够抑制烟草的免疫反应。
实施例4
inplantarnai方法沉默g20e03基因分析其功能
1、构建植物干涉重组表达载体pfgc5941-rnai-g20e03
(1)反向序列的插入
pcr扩增得到含xbal和bamhi酶切位点的orf反向序列,分别使用限制性内切酶xbal和bamhi进行酶切,纯化回收后,连接至经限制性内切酶xbal和bamhi酶切的植物干扰载体pfgc5941,获得重组表达载体pfgc5941-g20e03;
(2)正向序列的插入
pcr扩增得到含ncoi和swai酶切位点的orf正向序列,分别使用限制性内切酶ncoi和swai对重组表达载体pfgc5941-g20e03进行酶切,纯化回收后,连接至经限制性内切酶ncoi和swai酶切的载体pfgc5941-g20e03,获得重组表达载体pfgc5941-rnai-g20e03;
2、农杆菌感受态制备
(1)挑取农杆菌lba4404单菌落于5mllb液体培养基中(利福平rif5μl,单位25ng/μl),28℃,220rpm,振荡过夜培养;
(2)取2ml菌液转入50mllb液体培养基(不含抗生素)中,继续培养使菌液浓度达到od600=0.5左右(约4-6h);
(3)转入无菌离心管中,冰浴30min,4℃,5000rpm,5min,弃上清;
(4)用预冷的50mmol/lcacl2将残留的lb冲洗干净;
(5)加入10ml冰预冷的50mmol/lcacl2重悬菌体,冰浴30min;
(6)4℃,5000rpm,离心5min,弃上清;
(7)加入2ml冰预冷的50mmol/lcacl2重悬菌体。
3、重组表达载体转化农杆菌
(1)将5μl提取的重组质粒dna加入至200μl农杆菌感受态细胞中,轻轻混匀;
(2)冰浴30min,立即转入液氮速冻10min,迅速置于37℃热激水浴5min,立即取出后再冰浴2min;
(3)加入500μllb液体培养基中,28℃,130rpm,4-5h后,将菌液移至lb固体培养基中(加入1mlamp终浓度50mg/l和0.5ml利福平终浓度25mg/l)均匀涂布整个平板,梯度涂板(50μl,100μl,150μl,200μl)。28℃培养36-48h。
(4)挑取单菌落进行液体培养加入1mlamp终浓度50mg/l和0.5ml利福平终浓度25mg/l,pcr鉴定并测序。
4、农杆菌介导的烟草叶盘法转化烟草
(1)烟草无菌苗的培养
转基因所用烟草(nicotianatabacum)品种为k326。先用70%的乙醇浸泡烟草种子30s,无菌水冲洗5遍,再用3%的次氯酸钠溶液浸泡,其间不断振荡,5min后用无菌水冲洗3遍后,播种于gm培养基上(每盘10粒,共30粒),25℃,16h光照/8h黑暗,湿度65%培养。
(2)农杆菌的培养、侵染及共培养
从yep平板(rifr,strr,kanr)上挑取含有重组质粒的农杆菌单菌落,于yep(50mg/lstr,25mg/lrif和50mg/lkan)液体培养基中,28℃,220rpm振荡培养至od600为0.6-0.8;4℃,5000rpm离心10min,弃上清,收集菌体,用等体积50mlccm液体培养基重悬菌体;取烟草无菌苗的幼嫩、健壮叶片,去主脉,用无菌打孔器打成小圆饼后浸入制备好的菌液中侵染5min,并用无菌滤纸迅速吸干叶盘表面的多余菌液,叶盘背面向上平放在加盖一层滤纸的ccm培养基上,25℃,黑暗共培养3d。
(3)不定芽的诱导
培养之后,依次用无菌水洗、sim+cef(50mg/l)各洗5min,充分洗掉叶盘表面的农杆菌后放在吸水纸上,吸干多余水分;将叶盘背面向下,放在芽诱导培养基sim上,叶盘边缘轻压入培养基中,25℃,16h光照培养;侵染叶盘不断膨大,大约10d后长出绿色的愈伤,约两周后,切下带芽的叶缘,插入到sim培养基中继续培养。
(4)诱导生根
待不定芽长到大约3cm左右时,用无菌的解剖刀将叶缘长出的幼芽从基部与叶盘切开,选取形态正常生长旺盛的抗性苗,插入到rm培养基中进行诱导生根,25℃,16h光照/8h黑暗培养。
(5)再生植株的移栽与培养
待根系发育健壮后,选取根系长得烟草幼苗,将其从瓶中取出,用自来水洗净根上的琼脂,移入土中栽培,移栽前两天用保鲜膜覆盖使其保持一定的湿度,定期浇营养液,同时注意烟草开花时要用纸袋将花套袋,防止花粉在植株间传播。烟草结种后,收获变褐变硬的烟草种子,晒干后,将每株烟草的种子装到一个无菌的1.5ml离心管中,编号并于4℃保存。
上述培养基成分具体如下:
gm:ms+3%sucrose(蔗糖)+0.8%agar(琼脂),ph5.8
ccm:gm+1mg/l6-ba+0.2mg/lnaa+0.1mmas,ph5.8
sim:gm+1mg/l6-ba+0.2mg/lnaa+3mg/lppt+50mg/lcef,ph5.8
rm:1/2ms+3%sucrose(蔗糖)+0.8%agar(琼脂)+15mg/lkan+50mg/lcef,ph5.8
5、g20e03基因功能分析
(1)t0代转基因株系的鉴定
将t0代转基因烟草种子采用种子穴盘育苗,同时培育野生型烟草(烟草品种:k326作对照),置于温室中定期浇营养液,温度25℃,相对湿度65%,同时对其进行病虫害防治。
根据试剂盒(e.z.n.a.tmplantdnaminikit,omegad3485-0150preps)说明书对野生型和转基因烟草的幼嫩叶片进行dna提取。以提取的dna为模板,利用特异性引物(bar-f和bar-r)进行pcr检测,pcr反应体系:dntpmixtureμl,10×buffer2.μl,bar-f/bar-r各μl,ddh2o1μl,dna模板μl,extaq0.μl。pcr反应条件:94℃min,30×(94℃45s,55℃30s,72℃1min),72℃5min。
(2)转基因纯系烟草的培育
采用种子穴盘育苗,对阳性烟草苗t1代种子进行育苗,同等条件下培育野生型烟草(烟草品种k326作对照),并用营养土对烟草种子覆盖,置于温室中,定期浇营养液(温度30℃,相对湿度为80%),同时对其进行病虫害防治,鉴定为阳性的株系继续培养,获得t2代转基因株系,采用上述培育和鉴定方法,直至获得纯合的转基因株系;
(3)g20e03基因沉默效率的检测
培育野生型和转基因株系,待幼苗长至6-8片真叶时,将其移栽至培养钵中(塑料杯,杯口直径6.5cm、杯底直径4.5cm、杯高7.5cm),分别接种scnt的2龄幼虫于野生型植株和干涉植株的根系中(l17和l26),每株接种1000条,35d后,参照实施1的方法,检测g20e03基因的沉默效率,如图6所示,结果表明g20e03基因在l17和l26株系的拷贝数显著低于其在野生型植株的拷贝数,分别降低了53%和57%。
(4)g20e03基因沉默显著降低了scnt对烟草的致病性
培育野生型和两个转基因株系,每个处理和对照选取生长一致的植株各5株,每株接种1000条scnt的2龄幼虫,3次重复试验,35天后统计土内的孢囊和根上的雌虫数量,如图7所示,结果表明,l17和l26株系的scnt孢囊和雌虫数量明显低于野生型,分别降低了51%和63%。上述结果表明了scnt在侵染烟草的过程中,g20e03基因表达量的下降,显著降低了scnt对烟草植株的致病性。具体作用机制还有待于鉴定该基因与寄主植物体内的互作蛋白后进一步揭示。
所用引物序列见表2:
表2所用引物名称及序列
<110>青岛农业大学
<120>基因g20e03、其编码的蛋白及其在提高烟草植株抗大豆孢囊线虫病中的应用
<160>3
<170>patentinversion3.5
<210>1
<211>917
<212>dna
<213>基因g20e03的全基因序列
<400>1
acagatttgaacatatcaccttctatttaaaccctaattaaaacacacctaatttatccattaattagtccttttaatttgcaataaattaaaacctcatttcgcttttttccatctactaaaaatttcttaatggcattcgtcattccaagtttattcgctttctttttgttcgcttcattggcagcaacacaaaactcgtaccaaaagaaatctaacaatgaaccgaaggtctttaagggtgtcaaaaacggatgtaaattgataaactgtaccgatggtcgaacctgcggcattcgagttgggctcgcaaagtttggcaacagaaattttgagaaatttgattttccaaaatgtataacaaatgaaacggaactaaacatgaacacggacaacgatggaaatgcgcacattgttcctaatggaccagggtgccaaatgatacaatgttctgttggctaccaatgtcaagttcgaatttcaatttccaagcttggcaatttgccatatgctcagtcaattggaaccttcccgcaatgcgtggggcctaatgtaagcttcaactcatcaagttcaattatcgagaatggcccagggtgtgaccaacttccgacgaaatgtgacgaaggcacaaaatgcgtaactgccgtcggaattgcgaaatatggcaatttgccatggtcgcaagtaaagaattcgattatttttttgttttttgatcgcttattctagtttttctcgcccttctgtacatgaaacgatgaaaagcgctgcaactgttttatattcagaggagccaaaaaacgacggaaaatttatttttacctaatttaacaactcttgtgagtaaatttgtgatttaaaatgtattgtgatttatgattttttggtaaactaaaattcatttaaaaaaaaaa
<210>2
<211>600
<212>dna
<213>基因g20e03的cdna基因序列
<400>2
atggcattcgtcattccaagtttattcgctttctttttgttcgcttcattggcagcaacacaaaactcgtaccaaaagaaatctaacaatgaaccgaaggtctttaagggtgtcaaaaacggatgtaaattgataaactgtaccgatggtcgaacctgcggcattcgagttgggctcgcaaagtttggcaacagaaattttgagaaatttgattttccaaaatgtataacaaatgaaacggaactaaacatgaacacggacaacgatggaaatgcgcacattgttcctaatggaccagggtgccaaatgatacaatgttctgttggctaccaatgtcaagttcgaatttcaatttccaagcttggcaatttgccatatgctcagtcaattggaaccttcccgcaatgcgtggggcctaatgtaagcttcaactcatcaagttcaattatcgagaatggcccagggtgtgaccaacttccgacgaaatgtgacgaaggcacaaaatgcgtaactgccgtcggaattgcgaaatatggcaatttgccatggtcgcaagtaaagaattcgattatttttttgttttttgatcgcttattctag
<210>3
<211>199
<212>prt
<213>基因g20e03编码的氨基酸序列
<400>3
metalaphevalileproserleuphealaphepheleuphealaserleualaalathrglnasnsertyrglnlyslysserasnasngluprolysvalphelysglyvallysasnglycyslysleuileasncysthraspglyargthrcysglyileargvalglyleualalyspheglyasnargasnpheglulyspheasppheprolyscysilethrasngluthrgluleuasnmetasnthraspasnaspglyasnalahisilevalproasnglyproglycysglnmetileglncysservalglytyrglncysglnvalargileserileserlysleuglyasnleuprotyralaglnserileglythrpheproglncysvalglyproasnvalserpheasnserserserserileilegluasnglyproglycysaspglnleuprothrlyscysaspgluglythrlyscysvalthralavalglyilealalystyrglyasnleuprotrpserglnvallysasnserileilepheleuphepheaspargleuphe
- 还没有人留言评论。精彩留言会获得点赞!
- OTUD5的单克隆抗体制备及其在癌症治疗中的用途的制造方法与工艺
- 一种胱氨酸蛋白酶抑制剂C测定试剂盒的制造方法与工艺
- 氮氧化物作为新型组蛋白去乙酰化酶抑制剂的抗肿瘤应用的制造方法与工艺
- 一种硼替佐米水溶性药用组合物及其制备方法和用途与制造工艺
- 初级癌症疗法后使用的含硼蛋白酶体抑制剂的制造方法与工艺
- 斯氏副柔线虫半胱氨酸蛋白酶抑制剂Pj_CPI基因克隆、重组表达方法及应用与制造工艺
- 一种制备自噬小体型瘤苗的培养基添加剂及其制备方法与制造工艺
- 一种高产脯氨酸蛋白酶的黑曲霉菌株的制造方法与工艺
- N-萘烷基取代的含氮杂环衍生物、制备方法及其抗肿瘤用途与制造工艺
- 作为BET蛋白抑制剂的1H‑吡咯并[2,3‑c]吡啶‑7(6H)‑酮和吡唑并[3,4‑c]吡啶‑7(6H)‑酮的制造方法与工艺