抗CD269(BCMA)人源化抗体的制作方法
本发明涉及结合CD269(BCMA)的人源化抗体或抗体片段,从而破坏CD269和其天然配体(BAFF和APRIL)之间的相互作用,以及它们在治疗浆细胞介导的疾病如多发性骨髓瘤和自身免疫疾病中的用途。
背景技术:
:B细胞成熟抗原(BCMA)是肿瘤坏死因子受体超家族(TNFRSF)的第17个成员。它的天然配体是涉及(通过与进一步的配体相互作用)调节体液免疫、B细胞发育和内稳态的不同方面的B细胞激活因子(BAFF;也称为BLyS或TALL-1,TNFSF13B)以及增殖诱导配体。BCMA在恶性浆细胞上高度表达,所述恶性浆细胞例如是骨髓B细胞非霍奇金淋巴瘤的多发性骨髓瘤(MM),以及比MM更具侵略性并构成浆细胞紊乱的所有病例的约4%的浆细胞白血病(PCL)中的恶性浆细胞。除MM和PLC之外,还在患有霍奇金淋巴瘤的患者的霍奇金和里德-斯特恩伯格细胞(HodgkinandReed-Sternbergcells)上检测到BCMA(Chiu等人(2007)Blood109:729-739)。类似于它在浆细胞上的功能,结合于BCMA的配体显示了调节表达BCMA的多发性骨髓瘤细胞的生长和存活(Novak等人(2004)Blood103:689-694)。BAFF和APRIL通过BCMA的信号传导被认为是恶性浆细胞的促存活因子;因此,BCMA阳性肿瘤细胞的清除和/或配体-受体相互作用的破坏应改善多发性骨髓瘤和自身抗体依赖的自身免疫疾病的治疗结果。目前存在多种可用于治疗多发性骨髓瘤的方法(Raab等人(2009)Lancet374:324-339)。化学疗法导致多数受试者仅能部分控制多发性骨髓瘤;仅极少数进行化学疗法导致完全消退。因此经常应用结合方法,通常涉及另外施以皮质类固醇,例如地塞米松或强的松。然而,皮质类固醇被副作用,例如减少骨密度所困扰。使用某人自己的干细胞(自体的)或使用来自近亲属或相配型的无血缘供体(异体的)的细胞,干细胞移植也被提出。在多发性骨髓瘤中,进行的多数移植是自体类型。虽然不是可治愈的,但是这样的移植在选择的患者中显示出延长生命(Suzuki(2013)JpnJClinOncol43:116-124)。可供选择地,最近沙利度胺和其衍生物已被应用于治疗但也与次佳的成功率和高成本相关。最近蛋白酶抑制剂硼替佐米(PS-341)被批准用于治疗复发性和难治性MM,并且被单独或与已确定的药物联合用于许多临床试验,导致令人鼓舞的临床结果(Richardson等人(2003)NewEnglJMed348:2609-2617;Kapoor等人(2012)SeminHematol49:228-242)。联合治疗的成本是相对高的,并且成功率仍具有显著的改进的余地。由于同时使用多种药物时副作用的积累,治疗选择的联合也不是理想的。用于治疗浆细胞疾病,具体是多发性骨髓瘤的新方法是需要的。特异地靶向浆细胞的能力也对治疗自身免疫疾病具有巨大益处。通常首先使用非类固醇抗炎药(NSAID)或疾病缓解性抗风湿药(DMARD)来治疗轻度形式的自身免疫疾病。通常使用类固醇与强免疫抑制剂如靶向周期中细胞的环磷酰胺(一种细胞毒性剂)治疗涉及由于活动性疾病导致的器官功能障碍的更严重形式的系统性红斑狼疮(SLE)。仅最近,在患有自身免疫疾病的患者的血清中高水平存在的靶向细胞因子BAFF的抗体贝利单抗被食品药品监督管理局(FDA)批准其用于SLE。然而,在人体内仅新形成的B细胞依赖于BAFF以存活,而记忆B细胞和浆细胞对选择性的BAFF抑制不太敏感(Jacobi等人(2010)ArthritisRheum62:201-210)。对于类风湿性关节炎(RA),TNF抑制剂是最先被许可的生物制剂,然后是阿巴西普、利妥昔单抗和托珠单抗以及其他:它们抑制涉及关节炎症和破坏的关键炎症路径,然而由于相对的免疫抑制,以提高感染风险作为代价(Chan等人(2010)NatRevImmunol10:301-316,Keyser(2011)CurrRheumatolRev7:77-87)。尽管批准了这些生物制剂,患SR和SLE的患者常常显示自身免疫标志物的持续性,其最可能与骨髓中存在寿命长的、固着的浆细胞相关,所述浆细胞耐受例如通过利妥昔单抗和高剂量糖皮质激素和环磷酰胺疗法的CD20介导的切除.已经在本领域描述了结合CD269(BCMA)的抗体和其在治疗不同B细胞相关的医学紊乱中的用途。Ryan等人(MolecularCancerTherapeutics,20076(11),3009)描述了通过使用BCMA蛋白的第5至54位氨基酸的肽免疫大鼠获得的抗BCMA抗体。WO2012/163805描述了BCMA结合蛋白,如嵌合抗体和人源化抗体,它们用于阻碍BAFF和/或APRIL与BCMA相互作用的用途以及它们在治疗浆细胞恶性肿瘤例如多发性骨髓瘤的潜在用途。在其中公开的抗体通过使用BCMA蛋白的第4至53位氨基酸的重组肽免疫小鼠获得。WO2010/104949也公开了优选结合BCMA的胞外域的不同抗体以及它们在治疗B细胞介导的医学病症和紊乱中的用途.WO2002/066516和WO2012/066058公开了结合BCMA和另外的靶标两者的二价抗体以及它们在治疗B细胞相关医学紊乱中的潜在用途.在两个公开中的任一个公开中均没有提供关于二价抗体的结合特性和特异性表位的细节。技术实现要素:鉴于现有技术,作为本发明的基础的技术问题是提供适用于治疗与病原性浆细胞相关的人类疾病如多发性骨髓瘤和自身免疫疾病的制剂。该问题通过独立权利要求的特征解决。本发明的优选实施方式通过从属权利要求提供。因此,本发明的目的是提供结合CD269(BCMA),具体是结合CD269(BCMA)的胞外域的表位的人源化抗体或抗体片段。本文公开的抗体包括人源化序列、特别是优选的VL和VH结合区的人源化序列,该人源化序列保持如关于嵌合抗体J22.9-xi描述的适当的配体亲和性。在本发明的不同实施方式中,用于获得所述人源化序列的氨基酸序列修饰可以在原始嵌合抗体J22.9-xi的CDR区或在骨架区发生,其中所述骨架区应理解为在属于免疫球蛋白超家族并且比CDR更低“可变”的蛋白质的可变域的区域。完全惊讶的是本文提供的具体的人源化序列,优选涉及结合的VL和VH区的CDR区,显示了如在实验例中证实的特异性的和强的结合,并保持了原始嵌合抗体J22.9-xi的结合特征,保持的程度为保持它们的希望的治疗效果的程度。因此,本发明涉及包括VH域的抗体或抗体片段,所述VH域包括以下CDR序列:-RYWX1S(H-CDR1;SEQIDNO.15),其中X1:I、F、L、V、Y.C、G、A、S、T);-EINPX2X3STINYAPSLKDK(H-CDR2;SEQIDNo.16),其中X2X3:SS、NS、TS、GS、KS、RS、SD、SN、DE;-SLYX4DYGDAX5DYW(H-CDR3;SEQIDNO.17),其中X4:Y、L、A、V、F、I、W和/或X5:Y、L、F、I、V、A、C,其中所述抗体或其片段特异地结合CD269(BCMA)的胞外域的表位。尤其惊讶的是本文描述的人源化抗体展示出与原始嵌合抗体相比的序列改变,尤其是在所述嵌合抗体的CDR中的序列改变,该人源化抗体保持了针对它们的治疗效果的靶标的足够的结合特性。技术人员不会预料到人源化变体的结合特征会相似于原始嵌合抗体或小鼠抗体。考虑到可变域,尤其是CDR的序列改变,本文证实的人源化序列的有益的结合特征被认为是惊人的技术效果。在部分人源化抗体和完全人源化抗体之间的结合特征的比较也显示了完全人源化序列的改善。这表现出完全预料不到的结果。对嵌合体(部分人源化的)的初始修饰导致在结合亲和性的一些损失。然而,进一步的人源化的引入继而导致增强的结合,其中“完全人源化的”序列显示与原始嵌合体相似的结合特性,从而在进行这样显著的序列修饰而不严重损失结合亲和性之间显示出惊人的技术效果。在优选的实施方式中,如本文描述的抗体或抗体片段的特征在于VH域包括CDR序列RYWIS(SEQIDNO.18)或RYWFS(SEQIDNO.19)。在优选的实施方式中,如本文描述的抗体或抗体片段的特征在于VH域包括CDR序列EINPNSSTINYAPSLKDK(SEQIDNo.20)或EINPSSSTINYAPSLKDK(SEQIDNo.21)。在本发明的进一步的实施方式中,VH域的第54位氨基酸可以涉及任何给定的氨基酸或修饰的氨基酸。如下面的实例中所示,在该残基出的N氨基酸的潜在糖基化不显著破坏抗体特异和较强地结合于靶表位。鉴于该信息,本发明涉及包括如本文描述的CDR2序列的抗体或抗体片段,其中任何给定的氨基酸或修饰的氨基酸可以在CDR2序列中的VH域的第54位氨基酸处出现。在优选的实施方式中,如本文描述的抗体或抗体片段的特征在于,所述VH域包括CDR序列SLYYDYGDAYDYW(SEQIDNO.22)。本发明进一步涉及包括VL域的抗体或抗体片段,所述VL域包括以下CDR序列:-KASQSVX1X2NVA(L-CDR1;SEQIDNO.23),其中X1X2:ES、SS、TS、QS、HS、DH;-SASLRFS(L-CDR2;SEQIDNO24);和/或-QQYNNYPLTFG(L-CDR3;SEQIDNO.25),其中,所述抗体或其片段特异地结合CD269(BCMA)的胞外域的表位。另外,对于LC序列,惊人的是CDR3序列的修饰序列对结合于BCMA靶标不具有显著的有害作用。此外,本发明的抗体当在溶液中,无论是分离或纯化的,或在体外,以及在体内时,在施用后还显示了预料不到的和有益的稳定性特征,这不会从对原始嵌合抗体进行的序列改变而预料到。在优选的实施方式中,如本文描述的抗体或抗体片段的特征在于,VL域包括CDR序列KASQSVDSNVA(SEQIDNO.26)。在优选的实施方式中,如本文描述的抗体或抗体片段包括含有以下序列的VH域:EVQLVESGGGLVQPGGSLRLSCAASGFTFSRYWX1SWVRQAPGKGLVWVGEINPX2X3STINYAPSLKDKFTISRDNAKNTLYLQMNSLRAEDTAVYYCASLYX4DYGDAX5DYWGQGTLVTVSS(SEQIDNO.4),其中X1:I、F、L、V、Y.C、G、A、S、T;X2X3:SS、NS、TS、GS、KS、RS、SD、SN、DE,优选SS;X4:Y、L、A、V、F、I、W;和X5:Y、L、F、I、V、A、C。在优选的实施方式中,如本文描述的抗体或抗体片段的特征在于,所述抗体或抗体片段包括VH域,所述VH域包括根据SEQIDNO.6、SEQIDNO.7、SEQIDNO.8或SEQIDNO.9的序列.在优选的实施方式中,如本文描述的抗体或抗体片段包括VH域,所述VL域包括序列EIVMTQSPATLSVSPGERATLSCKASQSVX1X2NVAWYQQKPGQAPRALIYSASLRFSGIPARFSGSGSGTEFTLTISSLQSEDFAVYYCQQYNNYPLTFGAGTKLELKR(SEQIDNO.12),其中X1X2:ES、SS、TS、QS、HS、DH。在优选的实施方式中,如本文描述的抗体或抗体片段包括VH域,所述VL域包括根据SEQIDNO.14的序列.在优选的实施方式中,如本文描述的抗体或抗体片段包括含有根据SEQIDNO.6、SEQIDNO.7、SEQIDNO.8或SEQIDNO.9的序列的VH域以及含有根据SEQIDNO.14的序列的VH域。本发明进一步涉及如本文描述的抗体或抗体片段,包括VH域,其中所述VH域包括根据以下的序列X1VQLX2X3SGGGLVQPGGSLX4LSCAASGX5X6FX7X8YWZ1SWVRX9APGKGLEWX10GEINPZ2SSTINYAPSLKX11X12FX13ISRDNAKNTLYLQMX14X15X16RX17EDTAX18YYCASLYYDYGDAZ3DYWGQGTX19VTVSS(SEQIDNo.41),其中X1:Q、E;X2:Q、V;X3:Q、E;X4:K、R;X5:I、F;X6:D、T;X7:S、D;X8:R、D;X9:R、Q;X10:I、V;X11:D、G;X12:K、R;X13:I、T;X14:S、N;X15:K、S;X16:V、L;X17:S、A;X18:L、V;X19:S、L;并且其中Z1中的至少一个:I、F、L、V、Y.C、G、A、S、T,优选I或F;Z2:S、N、T、G、K、R、D,优选S和/或Z3:Y、L、F、I、V、A、C,优选Y;并且其中所述抗体或其片段特异地结合CD269(BCMA)的胞外域的表位。该实施方式涵盖不同的人源化抗体,具体是其VH序列,通过在如本文描述的CDR中进行的有利的人源化定义的所有变体。本发明进一步涉及如本文描述的抗体或抗体片段,包括VL域,其中所述VL域包括根据以下的序列DIVMTQSX1X2X3X4X5X6SVGDX7VX8X9TCKASQSVESNVAWYQQKPX10QX11PKX12LIX13SX14X15LRFSGVPARFX16GSGSGTDFTITISX17LQSEDX18AX19YX20CQQYNNYPLTFGAGTKLELKR(SEQIDNo.42),其中X1:Q、P;X2:R、A;X3:F、T;X4:M、L;X5:T、S;X6:T、V;X7:R、E;X8:S、T;X9:V、L;X10:R、G;X11:S、A;X12:A、L;X13:F、Y;X14:A、D;X15:S、D;X16:T、S;X17:N、S;X18:L、F;X19:E、V;X20:F、Y;并且其中所述抗体或其片段特异地结合CD269(BCMA)的胞外域的表位。该实施方式涵盖不同的人源化抗体,具体是其VL序列,通过在如本文描述的CDR中进行的有利的人源化定义的所有变体。关于人源化抗体变体的优选实施方式如本文详细公开的,根据J22.9-xi的本发明的优选实施方式的序列是人源化的以提供用于施用于人受试者的更相容的试剂。生成并测试J22.9-xi的不同人源化序列变体的对人和猕猴CD269(BCMA)两者的结合亲和性和特异性。来自结合测定的结果证实人源化序列保持了嵌合试剂J22.9-xi的需要的结合特性。在下面的序列中,下划线区域表示CDR或推定的CDR。关于人源化VH变体的优选实施方式下面提供了本发明的人源化抗体序列的另外的信息。嵌合序列:HC小鼠(SEQIDNo.1):QVQLQQSGGGLVQPGGSLKLSCAASGIDFSRYWMSWVRRAPGKGLEWIGEINPDSSTINYAPSLKDKFIISRDNAKNTLYLQMSKVRSEDTALYYCASLYYDYGDAMDYWGQGTSVTVSSHC小鼠序列表示最初开发的用于嵌合抗体J22.9-xi的重链可变区(VH),所述嵌合抗体J22.9-xi包括获得自小鼠抗体的VL和VH域,能够结合CD269(BCMA)的胞外域的表位,并且所述VL和VH域分别融合于人CL和CH域。部分人源化序列:HC部分人源化(SEQIDNo.2):EVQLVESGGGLVQPGGSLRLSCAASGFTFDDYWMSWVRQAPGKGLEWVGEINPDSSTINYAPSLKGRFTISRDNAKNTLYLQMNSLRAEDTAVYYCASLYYDYGDAMDYWGQGTLVTVSSHC部分人源化序列表示与本文公开的嵌合抗体相比修饰的氨基酸序列(通过氨基酸替换),其中关于其序列修饰VL和VH结合区以使其更适用于对人施用。人源化VH序列:hHC01(SEQIDNo.3):EVQLVESGGGLVQPGGSLRLSCAASGFTFSRYWMSWVRQAPGKGLVWVGEINPDSSTINYAPSLKDKFTISRDNAKNTLYLQMNSLRAEDTAVYYCASLYYDYGDAMDYWGQGTLVTVSS移除翻译后修饰基序的人源化VH序列:hHC02(SEQIDNo.4):EVQLVESGGGLVQPGGSLRLSCAASGFTFSRYWX1SWVRQAPGKGLVWVGEINPX2X3STINYAPSLKDKFTISRDNAKNTLYLQMNSLRAEDTAVYYCASLYX4DYGDAX5DYWGQGTLVTVSS其中:X1:I、F、L、V、Y.C、G、A、S、T,优选I或F;X2X3:SS、NS、TS、GS、KS、RS、SD、SN、DE,优选SS;X4:Y、L、A、V、F、I、W,优选Y;和/或X5:Y、L、F、I、V、A、C,优选Y;“hHC01”和“hHC02”人源化序列表示新的氨基酸序列,该新的氨基酸序列包括与本文描述的原始嵌合序列和部分人源化序列相比的序列改变。PTM突变是意图移除来自所述蛋白质的潜在有害的翻译后修饰基序,同时保持有利的结合特性。hHC01和hHC02的第1、5、6、19、27、28、34、39、46、48、54、69、84、85、86、88、93、107和/或115位与原始嵌合抗体序列相比优选是突变(替换)的。替换的重要性主要涉及产生的氨基酸而非起源的氨基酸。因此,改变也可以从原始嵌合氨基酸或其他变体如部分人源化序列的对应氨基酸进行。下面的替换与嵌合(SEQIDNo1)序列相比是新的:-HC(VH)序列的氨基酸M34被替换为任何氨基酸,优选I、L、F、V、Y.C、G、A、S、T;-HC(VH)序列的氨基酸E46被替换为V;-HC(VH)序列的氨基酸D54和S55被替换为任何氨基酸组合,优选SS、TS、GS、KS、RS、SD、SN、DE;-HC(VH)序列的氨基酸Y101被替换为任何氨基酸,优选L、A、V、F、I、W;和/或-HC(VH)序列的氨基酸M107被替换为任何氨基酸,优选L、Y、F、I、V、A、C。可在与BCMA直接相互作用需要的那些残基处修饰的序列:hHC03-涉及到与BCMA相互作用的修饰氨基酸(SEQIDNo5):EVQLVESGGGLVQPGGSLRLSCAASGFTFSRYX1MX2WVRQAPGKGLVX3VGX4INPDSSTINYAPSLKDKFTISRDNAKNTLYLQMNSLRAEDTAVYYCASX5X6X7DYGDX8MDYWGQGTLVTVSS其中优选的氨基酸是X1:W、F、Y,优选W;X2:S、T、N、Q、D、E,优选S;X3:W、F、Y,优选W;X4:E、Q,优选E;X5:L、I、V、G、A,优选L;X6:Y、X,优选Y;X7:Y、F、L、I、V、M,优选Y;和/或X8:A、G、V,优选A。“hHC03”人源化序列表示新的氨基酸序列,该新的氨基酸序列包括与原始嵌合序列和部分人源化序列两则相比的氨基酸序列改变。这些序列改变意图反映在结合BCMA靶标的可以被替换的氨基酸中的潜在改变,同时保持有利的结合特性。替换的重要性主要涉及产生的氨基酸而非起源的氨基酸。因此,改变也可以从原始嵌合氨基酸或其他变体的对应氨基酸进行。例如:-HC(VH)序列的氨基酸W33是W、F、Y;-HC(VH)序列的氨基酸S35是S、T、N、Q、D、E;-HC(VH)序列的氨基酸W47是W、F、Y;-HC(VH)序列的氨基酸E50是E、Q;-HC(VH)序列的氨基酸L99是L、I、V、G、A;-HC(VH)序列的氨基酸Y100是Y、X;-HC(VH)序列的氨基酸Y101是Y、F、L、I、V、M;和/或-HC(VH)序列的氨基酸A106A、G、V。通常,当独立于骨架序列作为一个整体考虑时,在人源化过程中对CDR区所做的任何改变也可以被认为是CDR序列的特征。这样修饰的CDR序列可以被认为是在本文描述的整个骨架区中所述修饰的CDR序列的上下文之内,或独立于本文描述的整个骨架区中所述修饰的CDR序列的上下文的本发明的限定性特征。例如,在hHC01至hHC03中以下划线识别的CDR序列可以被认为是独立于周围的骨架序列的本发明的限定性特征。人源化HC(VH)序列的特定例子:hHC04(SEQIDNO6):EVQLVESGGGLVQPGGSLRLSCAASGFTFSRYWISWVRQAPGKGLVWVGEINPNSSTINYAPSLKDKFTISRDNAKNTLYLQMNSLRAEDTAVYYCASLYYDYGDAYDYWGQGTLVTVSShHC05(SEQIDNO7):EVQLVESGGGLVQPGGSLRLSCAASGFTFSRYWFSWVRQAPGKGLVWVGEINPNSSTINYAPSLKDKFTISRDNAKNTLYLQMNSLRAEDTAVYYCASLYYDYGDAYDYWGQGTLVTVSShHC06(SEQIDNO8):EVQLVESGGGLVQPGGSLRLSCAASGFTFSRYWISWVRQAPGKGLVWVGEINPSSSTINYAPSLKDKFTISRDNAKNTLYLQMNSLRAEDTAVYYCASLYYDYGDAYDYWGQGTLVTVSShHC07(SEQIDNO9):EVQLVESGGGLVQPGGSLRLSCAASGFTFSRYWFSWVRQAPGKGLVWVGEINPSSSTINYAPSLKDKFTISRDNAKNTLYLQMNSLRAEDTAVYYCASLYYDYGDAYDYWGQGTLVTVSS比对:对在HC序列的不同替换位置的CLUSTALW(1.83)多序列比对在图8中提供了适当的序列比较。“广义序列”表示HC序列,其中每个X代表对任何给定的氨基酸的潜在氨基酸改变。优选的氨基酸替换是上文描述的用于每个潜在突变位置的那些。关于人源化VL变体的优选实施方式嵌合序列:LC小鼠(SEQIDNo.43):DIVMTQSQRFMTTSVGDRVSVTCKASQSVDSNVAWYQQKPRQSPKALIFSASLRFSGVPARFTGSGSGTDFTLTISNLQSEDLAEYFCQQYNNYPLTFGAGTKLELKRLC小鼠序列表示最初开发的用于嵌合抗体J22.9-xi的轻链的可变区(VL),所述嵌合抗体J22.9-xi包括获得自小鼠抗体的VL和VH域,能够结合CD269(BCMA)的胞外域的表位,并且所述VL和VH域分别融合于人CL和CH域。部分人源化序列:LC部分人源化(SEQIDNO10):DIVMTQSPATLSVSVGDEVTLTCKASQSVDSNVAWYQQKPGQAPKLLIYSDDLRFSGVPARFSGSGSGTDFTLTISSLQSEDFAVYYCQQYNNYPLTFGAGTKLELKRLC部分人源化序列表示与本文发明的实施例中公开的嵌合抗体相比修饰的序列(通过氨基酸替换),其中关于其序列修饰VL和VH结合区以使其更适用于对人施用。人源化VL序列:hLC01(SEQIDNO11):EIVMTQSPATLSVSPGERATLSCKASQSVDSNVAWYQQKPGQAPRALIYSASLRFSGIPARFSGSGSGTEFTLTISSLQSEDFAVYYCQQYNNYPLTFGAGTKLELKR移除翻译后修饰基序的人源化VL序列:hLC02(SEQIDNO12):EIVMTQSPATLSVSPGERATLSCKASQSVX1X2NVAWYQQKPGQAPRALIYSASLRFSGIPARFSGSGSGTEFTLTISSLQSEDFAVYYCQQYNNYPLTFGAGTKLELKR其中:X1X2:ES、SS、TS、QS、HS、DH、优选ES。“hLC01”和“hLC02”人源化序列表示新的氨基酸序列,该新的氨基酸序列包括与本文描述的原始嵌合序列和部分人源化序列两者相比的序列改变。PTM突变是意图移除来自所述蛋白质的潜在有害的翻译后修饰基序,同时保持有利的结合特性。与原始嵌合抗体序列相比,hLC01和hLC02的第1、8、9、10、13、15、17、19、20、21、22、30、41、43、45、49、58、63、70、77、83、85和/或87位优选是突变(替换)的。替换的重要性主要涉及产生的氨基酸而非起源的氨基酸。因此,也可以从原始嵌合氨基酸或其他变体的对应氨基酸进行改变。下面的替换相对于嵌合序列和部分人源化序列是新的:-LC(VL)序列的氨基酸D1被替换为E;-LC(VL)序列的氨基酸V15被替换为P;-LC(VL)序列的氨基酸D17被替换为E;-LC(VL)序列的氨基酸V19被替换为A;-LC(VL)序列的氨基酸T22被替换为S;-LC(VL)序列的氨基酸D30和S31被替换为任何氨基酸组合,优选ES、SS、TS、QS、HS、DH;-LC(VL)序列的氨基酸V58被替换为I;和/或-LC(VL)序列的氨基酸D70被替换为E。可在与BCMA直接相互作用需要的那些残基处在其CDR结合区修饰的序列:hLC03-涉及与BCMA相互作用的修饰氨基酸(SEQIDNO13):EIVMTQSPATLSVSPGERATLSCKASQSVDX1X2VX3WX4QQKPGQAPRALIX5X6AX7X8RX9SGIPARFSGSX10X11GTEFTLTISSLQSEDFAVYYCX12QX13NNX14PX15TFGAGTKLELKR其中优选氨基酸为:X1:S、H、T、N、D、Q;X2:N、E、Q;X3:A、G、V、S、T、L、I;X4:Y、F、L、I、V、A、G;X5:Y、F、L;X6:S、T;X7:S、T、D、N、H、E、Q;X8:L、V、I、M;X9:F、L、I、V、Y、M;X10:G、X;X11:S、X;X12:Q、V、L、I、M;X13:Y、F、L、I、Q;X14:Y、F、R、Q、K;和/或X15:L、I、V、F。“hLC03人源化序列”表示新的氨基酸序列,该新的氨基酸序列包括与原始嵌合序列和部分人源化序列两者相比的氨基酸序列改变。这些序列改变意图反映在结合BCMA靶标的可以被替换的氨基酸中的潜在改变,同时保持有利的结合特性。替换的重要性主要涉及产生的氨基酸而非起源的氨基酸。因此,改变也可以从原始嵌合氨基酸或其他变体的对应氨基酸进行。例如:-LC(VL)序列的氨基酸S31是S、H、T、N、D、Q;-LC(VL)序列的氨基酸N32是N、E、Q;-LC(VL)序列的氨基酸A34是A、G、V、S、T、L、I;-LC(VL)序列的氨基酸Y36是Y、F、L、I、V、A、G;-LC(VL)序列的氨基酸Y49是Y、F、L;-LC(VL)序列的氨基酸S50是S、T;-LC(VL)序列的氨基酸S52是S、T、D、N、H、E、Q;-LC(VL)序列的氨基酸L53是L、V、I、M;-LC(VL)序列的氨基酸F55是F、L、I、V、Y、M;-LC(VL)序列的氨基酸G66是G、X;-LC(VL)序列的氨基酸S67是S、X;-LC(VL)序列的氨基酸Q89是Q、V、L、I、M;-LC(VL)序列的氨基酸Y91是Y、F、L、I、Q;-LC(VL)序列的氨基酸Y94是Y、F、R、Q、K;和/或-LC(VL)序列的氨基酸L96是L、I、V、F。通常,当独立于骨架序列作为一个整体时,对CDR区所做的任何改变也可以被认为是CDR序列的特征。这样修饰的CDR序列可以被认为是在本文描述的整个骨架区中所述修饰的CDR序列的上下文之内,或独立于本文描述的整个骨架区中所述修饰的CDR序列的上下文的本发明的限定性特征。例如,在hLC01至hLC03中以下划线识别的CDR序列可以-以未修饰的或替换的形式-被认为是独立于周围的骨架序列的本发明的限定性特征。人源化LC序列的例子:hLC04(SEQIDNO14):EIVMTQSPATLSVSPGERATLSCKASQSVESNVAWYQQKPGQAPRALIYSASLRFSGIPARFSGSGSGTEFTLTISSLQSEDFAVYYCQQYNNYPLTFGAGTKLELKR比对:对在LC序列内的各个潜在改变位点的CLUSTALW(1.83)多序列比对在图9中提供了适当的序列比较。“广义序列”表示LC序列,其中每个X代表潜在氨基酸改变。优选的氨基酸替换是上文描述的用于每个潜在突变位置的那些。因此,本发明涉及根据hHC01、hHC02、hHC03、hHC04、hHC05、hHC06、hHC07、hLC01、hLC02、hLC03和/或hLC04的人源化序列,或任何给定的其组合。本发明涵盖对任何本文提出的给定的潜在变体残基(如在“广义”序列中识别为X)的潜在修饰的所有可能的组合。通过组合一个或多个这些不同的替换,可以产生展示最初开发的并在本文中证实的嵌合抗体的希望的结合特性的人源化变体。本文描述的抗体或其部分也涵盖与明确公开或通过序列式公开的那些人源化序列具有至少80%,优选90%序列一致性的序列。关于抗体表位的优选实施方式本发明因此涉及结合CD269(BCMA)的分离的抗体或抗体片段,其中抗体结合包括CD269(BCMA)的第13至第32个残基中的一个或多位氨基酸的表位。CD269的第13至第32个残基的氨基酸序列示于SEQIDNo.40。在SEQIDNo.39中提供CD269的N末端序列。CD269的胞外域提供为SEQIDNo.38。包括根据SEQIDNo.38的CD269的胞外域的抗原用于免疫以产生本文描述的小鼠抗体和嵌合抗体的结合特异性。使用整个CD269蛋白或其包括膜结合的或胞内域的片段作为抗体产生期间的抗原,可以产生结合CD269的隐藏域或胞内域的抗体,借此使得这样的制剂不适于或不利于治疗应用。因此本发明的抗体通过它们与CD269的胞外域的结合定义。在胞外域内的特异表位也表示本发明优选的新的并且预料不到的特征。从小鼠抗体或嵌合抗体制备的Fab片段以与纯化的BCMA胞外域的复合结晶,并且解出了复合结构。结构分析揭示了本发明的抗体的表位的细节信息以及其生物相关性。本发明的抗体结合包括胞外域的CD269(BCMA)的第13至第32个残基中的一个或多个氨基酸的表位是有利特性,因为该区域显示了与CD269的两种天然配体BAFF和APRIL的结合位点的显著重叠。至今没有在本领域描述的抗CD269抗体显示了与BAFF和APRIL结合位点的这样的全面重叠。在一种实施方式中,本发明的分离的抗体或抗体片段的特征在于,抗体结合包括CD269(BCMA)的第13、15、16、17、18、19、20、22、23、26、27或32位氨基酸中的一个或多个的表位。在另一种实施方式中,本发明的分离的抗体或抗体片段的特征在于,抗体结合由CD269(BCMA)的第13、15、16、17、18、19、20、22、23、26、27和32位氨基酸组成的表位。这些残基表示直接与本发明的抗体相互作用的氨基酸,正如本文提供的晶体结构数据所鉴定。关于提供CD269的N末端序列的SEQIDNo.39,对这些残基进行了编号。在本发明的一种实施方式中,分离的抗体或抗体片段的特征在于,结合于CD269(BCMA)的抗体破坏BAFF-CD269和/或APRIL-CD269相互作用。本发明的抗体与CD269的胞外域的结合破坏了BAFF-CD269的相互作用。由于APRIL和BAFF的结合位点位于与抗体表位相似的位点,抗体与CD269的结合也阻断APRIL-CD269的相互作用。本发明的抗体的特异表位与APRIL和BAFF的结合位点的比较揭示了在天然配体和本文描述的抗体的结合位点中的全面重叠,所述APRIL和BAFF的晶体结构已经解出并绘制了他们的相互作用。这表示本发明的有利和预料不到的方面,并且使得BAFF-CD269和/或APRIL-CD269的相互作用的破坏可靠并且有效。本发明因此涉及本文描述的分离的抗体或抗体片段,其中抗体通过结合包括CD269(BCMA)的第13、15、17、18、19、22、26、27、30、31、32、33、34、35个残基中的一个或多个氨基酸的表位,尤其由第13、15、17、18、19、22、26、27、32位氨基酸组成的表位破坏APRIL-CD269相互作用。这些氨基酸分别对应于APRIL在CD269上的结合位点,以及结合如本文描述的抗体和APRIL两者的CD269的重叠残基。在另一种实施方式中,本发明因此涉及本文描述的分离的抗体或抗体片段,其中抗体通过结合包括CD269(BCMA)的第13、15、16、17、18、19、22、25、26、27、29、30、31、32、34、35位残基中的一个或多个氨基酸的表位,尤其由第13、15、16、17、18、19、22、26、27、32位氨基酸组成的表位破坏BAFF-CD269相互作用。这些氨基酸分别对应于BAFF在CD269上的结合位点,以及结合如本文描述的抗体和BAFF两者的CD269的重叠残基。虽然在本领域已描述了结合CD269的抗体,其也潜在地破坏APRIL或BAFF与CD269的相互作用,没有提供涉及这样的抗体的特异性表位的相关公开内容.不能假设之前描述的抗体与本发明的抗体一样也结合具有这样的全面重叠的表位。即使APRIL或BAFF与CD269的相互作用已经显示被破坏,这可能由于结合相当不同的表位以及随后发生APRIL或BAFF锚定的位阻而潜在地发生。之前没有记载由现有的抗体导致的APRIL或BAFF与CD269的相互作用破坏的程度。本发明的抗体使有效并可靠的破坏成为可能,这潜在地表示与本领域中描述的那些抗体相比改进的技术效果。可进行体外阻断测定以确定并比较BAFF和/或APRIL破坏,例如使用人BCMA的胞外域与重组BAFF或APRIL。在优选的实施方式中,与本文描述的抗体显示的高亲和性结合的表位特异性,代表了新的和预料不到的技术效果.本质上,J22.9抗体和其人源化变体的例外高的亲和性,不仅提供了天然配体的结合的“破坏”或“阻断”;而且本发明的抗体的超高亲和性,确保了当抗体存在时,本质上完全或几乎完全排除天然配体结合它们的BCMA靶标。如以下实施例中公开的,如本文描述的人源化抗体的亲和性惊人地高并且相对好于在现有技术中尝试的相似方法。在pM范围内的Kd一般被视为在一般实践中不能预见的出色的亲和性。在另一方面,本发明的人源化抗体或抗体片段以高亲和性结合CD269,例如,当通过表面等离子体共振如Biacore测量时,抗体以100nM、90、80、70、60、50、40、30nM或更低的亲和力,以20nM或更低的亲和力,或以15nM或更低的亲和力,或以5nM或更低的亲和力,或以1000pM或更低的亲和力,或以500pM或更低的亲和力,或以100pM或更低,或以80pM或更低,或例如约50pM的亲和力结合于人CD269.在进一步的实施方式中,当通过表面等离子体共振如Biacore测量时,抗体以约1pM至约100nM,或约100pM至约50nM,或约200pM至约20nM结合于人CD269。本发明的进一步优选的实施方式在一种实施方式中,本发明涉及包括通过直接与CD269靶标相互作用的一个或多个氨基酸和/或通过经水相互作用进行相互作用的一个或多个氨基酸定义的氨基酸序列的抗体或抗体片段(见表1至6)。涉及如本文描述的抗体与表位的结合的大量的水相互作用代表了结合的不寻常和惊人的方面。尤其,抗体指向本文描述的特别表位的高亲和性,与涉及抗体与表位之间的结合表面的大量的水相互作用组合,代表了本发明的惊人的和预料不到的方面。本发明因此涉及包括如本文描述的氨基酸序列的抗体或抗体片段,其中所述序列表征为,关于本文公开的嵌合体或关于本文公开的人源化序列变体的对应残基,存在涉及通过根据表5的水桥(waterbridge)与靶标表位的相互作用表面的特异性氨基酸残基,所述特异性氨基酸残基选自包括轻链的Ser31、Asn32、Tyr36、Ser50、Ser52、Gly66、Gln89、Tyr91和/或Tyr94,和/或重链的Trp33、Ser35、Trp47、Glu50、Leu99和/或Tyr101的组。虽然关于抗体的水桥形成的例子是以J22.9-xi嵌合体进行的,但是发明人声称由于与测试的原始嵌合抗体相比在人源化变体中保持了结合特征,使得在本发明的人源化变体中保持了该技术效果。在重链中,仅有的突变的水桥残基为Y101,但它的水相互作用涉及主链(即骨架)原子,并因此可被合理地假定为由于使侧链突变而不会改变;在轻链中,没有涉及水桥的残基的突变。本发明的抗体可以进一步表征为,涉及经水桥与本文描述的抗体相互作用的表位的氨基酸残基。在表5中提供了相关特征。例如在一种实施方式中,本发明因此的特征在于,轻链的残基Ser31与CD269的Thr32经水分子相互作用。本发明的抗体的结合特性的这样的描述旨在用于如表5提供的每个相互作用。此外,在本发明中包括了本文描述的抗体的序列变体,其中涉及“水桥”相互作用的一个或多个残基是修饰的,以便以水“桥”为代价将直接的侧链相互作用“替换”入序列。例如,可以在氨基酸序列中进行突变或改变,从相互作用表面置换水而不显著影响相互作用的亲和性。本发明因此涉及包括如本文描述的氨基酸序列的抗体或抗体片段,其中所述序列被表征为,关于本文公开的嵌合体或关于本文公开的人源化序列变体的对应残基,涉及通过根据表5的水桥与靶标表位的相互作用表面的那些氨基酸残基的序列变异,所述氨基酸残基选自包括轻链的Ser31、Ser31、Asn32、Tyr36、Ser50、Ser52、Gly66、Gln89、Tyr91和/或Tyr94,和/或重链的Trp33、Ser35、Trp47、Glu50、Leu99和/或Tyr101的组.在本文描述的人源化抗体的对应位置的变异可涉及任何给定的氨基酸替换,优选,会有效从相互作用中置换水而保持关于表位亲和性和特异性的相似结合特性的氨基酸替换。在本发明的一种实施方式中,分离的抗体或抗体片段的特征在于,抗体是糖基化的,优选包括N-连接寡糖链,优选在重链的Asn297处。抗体的糖基化是指碳水化合物或聚糖与抗体的连接。N-连接聚糖连接于天冬酰胺或精氨酸侧链的氮上。连接至耙蛋白的碳水化合物链起到不同的功能。例如,除非它们首先进行糖基化,某些蛋白质不正确折叠。而且,连接在蛋白质中的精氨酸的酰胺氮上的多糖对某些分泌的糖蛋白给予稳定性。这种情况下的糖基化对适当折叠不是限制性的需求,而是非糖基化的蛋白可以更快地被降解。如本发明的实施例所证实的,其中公开的抗体的去糖基化导致与糖基化形式的抗体相比治疗效果的下降。惊人的是,糖基化在保持抗体的活性上会起到显著的作用。糖基化因此代表本发明的与预料不到的技术优势相关的优选实施方式。如本文的实施例所证实的,虽然用J22.9-xi-N-聚糖(去糖基化的)处理的动物的总体肿瘤负荷与接受同种型对照抗体的动物没有显著差异,但是与isoAb处理组相比,这些小鼠的寿命显著地提高。由于J22.9-xi-N-聚糖显示为不能诱导ADCC或CDC,该结果表明J22.9-xi与BCMA单独的结合阻碍肿瘤生长。可以合理地考虑,这是由于受体和它的天然配体(APRIL和BAFF)间的相互作用的阻断。本发明的这个方面和本文描述的抗体代表了惊人的技术效果,该技术效果不能从现有技术的抗体中得到。J22.9-xi-N-聚糖(去糖基化的)可以被考虑为描述的试验中用于评估潜在的治疗效果的对照样品,该对照样品能够使抗体与其靶标表位结合,而没有ADCC或CDC的下游效应。本发明的抗体,优选具有糖基化的抗体,因此证明了这样的有效的位点结合,所述结合能够防止(或显著破坏)通过天然配体的结合而导致细胞毒性。本文描述的抗体的该特征没有被描述于本领域中描述的相似的抗体。虽然关于抗体的糖基化的例子用J22.9-xi嵌合体进行,但是发明人声称由于与测试的原始嵌合抗体相比保持了人源化变体的结合特征,因此在本发明的人源化变体中保持了该技术效果。糖基化的优选位置(重链的Asn297)没有与任何突变残基直接连接,并位于人的完整IgG的恒定区。因此,合理假设与嵌合抗体相比,在存在于任何J22.9变体的这个位置的糖基化模式是没有差异的。本发明的用途和功能方面本发明的抗体能够结合本文描述的表位,阻断该表位的天然配体的相互作用并诱导CDC和ADCC。抗体依赖性细胞介导的细胞毒性(ADCC)是通过本发明的产生希望的治疗效果的抗体诱导的一个因素。ADCC是细胞介导的免疫防御机制,其中免疫系统的效应细胞主动地裂解膜表面抗原已被特异性抗原结合的靶细胞。表达CD269的细胞与本发明的抗体结合后,可以诱导ADCC。经典的ADCC是通过自然杀伤(NK)细胞介导的;巨噬细胞、中性粒细胞和嗜酸性粒细胞也可以介导ADCC。由于ADCC依赖于在先的抗体应答,ADCC是适应性免疫应答的一部分。小鼠实验可以表明ADCC是如本文描述的治疗性抗体作用的重要机制。本发明的优选的实施方式涉及如本文描述的分离的抗体或抗体片段,供用作治疗与病原性B细胞存在相关的医学紊乱的药物。在本发明的一种实施方式中,医学紊乱是CD269相关的紊乱,优选与病原性B细胞相关,其优选为浆细胞和/或记忆B细胞的疾病。浆细胞疾病可以是浆细胞的癌症,例如多发性骨髓瘤、浆细胞瘤、瓦尔登斯特伦巨球蛋白血症或浆细胞白血病。浆细胞疾病可以是B淋巴细胞的癌症,如霍奇金病。在本发明的一种实施方式中,医学紊乱是与自身反应性浆细胞和/或自身反应性记忆B细胞相关的自身免疫疾病,如炎症性自身免疫疾病,例如系统性红斑狼疮或类风湿性关节炎。因此,本发明也涵盖用于如本文公开的医学紊乱的治疗方法,优选包括向需要这样的治疗的受试者施用治疗有效量的抗体。本发明的进一步方面涉及包括如本文所述的抗体或抗体片段的抗体药物缀合物(ADC)。抗-CD269抗体药物缀合物“抗CD269ADC”可以被描述为与治疗剂缀合的抗CD269抗体或其片段。在某些实施方式中,ADC包括抗CD269抗体(例如本文描述的J22.9-xi的人源化变体)。当施用于患有CD269表达医学病症如癌症或自身免疫紊乱的受试者时,如本文描述的ADC或ADC衍生物对表达CD269的细胞产生临床有益效果。在一种实施方式中,抗CD269抗体或其衍生物缀合于细胞毒素剂,以致产生的ADC或ADC衍生物表现出对表达CD269的癌细胞的细胞毒性作用,优选当被所述表达CD269的癌细胞占据或内化时。抗CD269的ADC或ADC衍生物优选被内化并积累在表达CD269的细胞中,在所述表达CD269的细胞中ADC或ADC衍生物发挥治疗作用(例如细胞毒性作用)。具体地,对缀合于抗体或抗体衍生物的特别适合的部分是化学治疗剂、前体药物转化酶、放射性同位素或化合物、或毒素。例如,可以将抗CD269抗体或其衍生物与细胞毒性剂如化学治疗剂或毒素(例如细胞抑制剂或杀细胞剂)缀合。本发明的另一方面涉及优选分离的核酸分子,所述核酸分子选自由以下组成的组:a)包括以下的核苷酸序列的核酸分子-其编码根据前述权利要求的任一项所述的分离的抗体或抗体片段,-其编码选自由根据SEQID1至31和41至42的那些序列组成的组中的氨基酸序列,-包括SEQIDNo.32至36的序列或序列片段,b)与根据a)的核苷酸序列互补的核酸分子;c)包括以下核苷酸序列的核酸分子,所述核苷酸序列具有足以在功能上与根据a)或b)的核苷酸序列类似/等价的序列一致性,优选与根据a)或b)的核苷酸序列的序列一致性为至少80%,优选为90%,更优选为95%;d)由于遗传密码的结果简并为根据a)至c)的核苷酸序列的核酸分子;以及e)根据a)至d)的核苷酸序列通过删除、添加、替换、易位、倒位和/或插入进行修饰,并且与根据a)至d)的核苷酸序列在功能上类似/等价的核酸分子。本发明的进一步的方面涉及能够产生本文描述的抗体或抗体片段和/或包括如本文所述的核酸分子的宿主细胞,如细菌细胞或哺乳动物细胞,优选杂交瘤细胞或细胞系。本发明进一步的方面涉及药物组合物,其包含如本文描述的分离的抗体或抗体片段、如本文描述的核酸分子或如本文描述的宿主细胞,以及药学上可接受的载体。本发明的另外的和令人惊讶的方法是如本文公开的抗体的改进的稳定性。所述抗体在适当的条件下可以容易地储存较长时间而结合亲和性没有任何丢失。关于在-80℃或4℃下储存之后的活性稳定性已经进行了适当测试,该测试证实了在前述两种温度下储存之后抗体的出人意料的良好稳定性和活性的保持(图3c)。该改进的稳定性对于嵌合抗体而言是明显的,并且对于其人源化变体而言也是出人意料的。出人意料地,在长期储存条件下,人源化变体的稳定性高于嵌合抗体。如本文描述的抗体的进一步的优点是如在实施例中证实的骨髓瘤细胞的系统性耗竭。之前在现有技术中公开的抗体没有被证实展示出希望的以系统性方式的抗浆细胞作用。针对现有技术的抗体进行的研究仅公开了用骨髓瘤细胞皮下注射并后续处理分离的细胞群。本发明提供了能够在其静脉内注射后系统性耗竭癌性多发性骨髓瘤细胞的抗体,如实施例证实的。靶细胞的有效耗竭代表了除了本发明的抗体的有效特性之外,之前未在现有技术中被描述的技术效果。具体实施方式如本文使用的,抗体通常是指由基本上通过免疫球蛋白基因或免疫球蛋白基因的片段编码的一种或多种多肽组成的蛋白质。在使用术语“抗体”时,术语“抗体片段”也被认为是与之相关的。公认的免疫球蛋白基因包括κ、λ、α、γ、δ、ε和μ恒定区基因,以及无数的免疫球蛋白可变区基因。轻链被分类为κ和λ。重链被分为γ、μ、α、δ或ε,其反过来分别定义免疫球蛋白类别IgG、IgM、IgA、IgD和IgE。基本的免疫球蛋白(抗体)结构单位已知包括四聚体或二聚体。每种四聚体由完全相同的两对多肽链组成,每对具有一条“轻”(L)链(约25kD)和一条“重”(H)链(约50-70kD)。每条链的N-末端定义约100至110或更多个氨基酸的可变区,其主要负责抗体识别。术语“可变轻链”和“可变重链”分别是指轻链和重链的可变区。任选地,抗体或抗体的免疫球蛋白部分可以被化学缀合于或表达为与其他蛋白质的融合蛋白。本发明的抗体意图结合哺乳类的,尤其是人的蛋白靶标。蛋白质名称的使用可以与小鼠或人类形式的蛋白质相对应。“特异性结合”应该如通过本领域技术人员理解的,其中所述技术人员清楚地知道可以同于测试结合和结合特异性的各种实验程序。某些交叉反应或背景结合在许多蛋白-蛋白相互作用中可能不可避免;这不是用于使抗体和表位之间的结合的“特异性”减损。当考虑在理解抗体和表位之间的相互作用中的术语“特异性”时,术语“针对(directedagainst)”也是适用的。本发明的抗体包括但不限于多克隆抗体、单克隆抗体、双特异性抗体、人抗体、人源化抗体或嵌合抗体,单可变区片段(ssFv)、单域抗体(如来自纳米抗体的VHH片段)、单链片段(scFv)、Fab片段、F(ab′)2片段、由Fab表达文库产生的片段、抗独特型抗体和表位结合片段或上述任意的组合,条件是它们保持原始的结合特性。在本发明的方法中也可以使用微型抗体和多价抗体如双抗体、三抗体、四价抗体和肽体(peptabodies)。本发明的免疫球蛋白分子可以是免疫球蛋白分子的任何类(即IgG、IgE、IgM、IgD和IgA)或亚类。因此,本文使用的术语抗体也包括通过整个抗体的修饰或使用重组DNA方法从头合成的抗体和抗体片段。可以通过
技术领域:
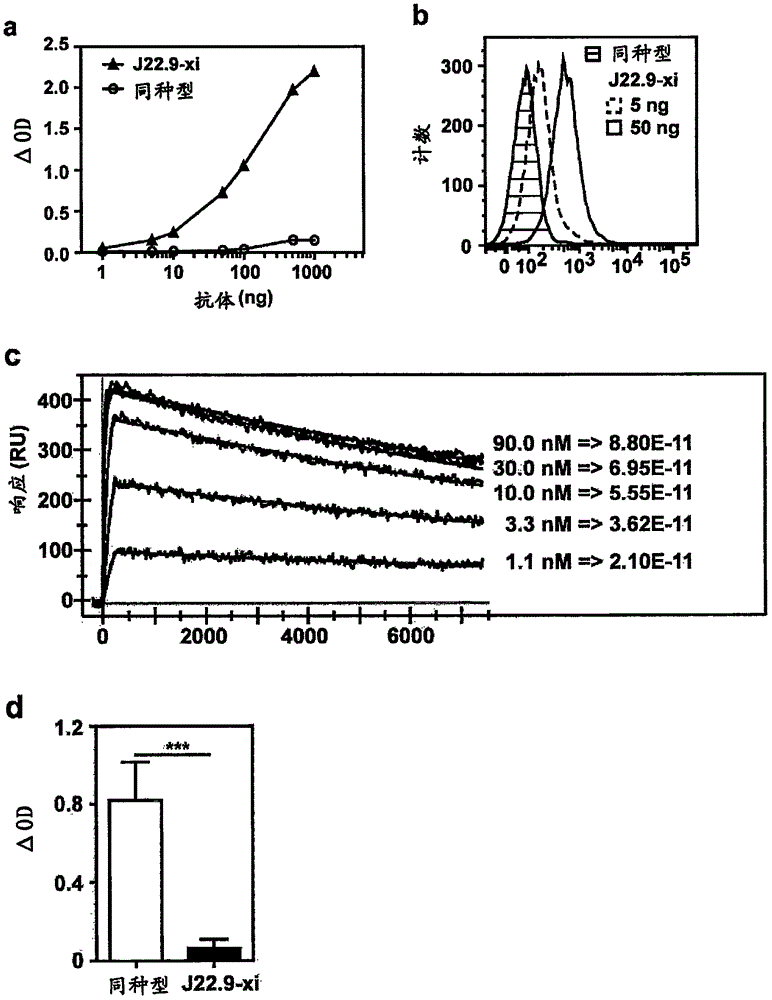
已知的任何方法制备包括本发明的抗体的一个或多个CDR或源自所述抗体的一个或多个CDR的人源化抗体。例如,可以使用四个一般步骤使单克隆抗体人源化。这些步骤是:(1)确定起始抗体轻链可变域和重链可变域的核苷酸序列和预期的氨基酸序列、(2)设计人源化抗体,即决定人源化过程中使用哪一个抗体骨架区、(3)实际的人源化方法/技术和(4)人源化抗体的转染和表达。参见,例如美国专利号4,816,567;5,807,715;5,866,692;6,331,415;5,530,101;5,693,761;5,693,762;5,585,089;6,180,370;5,225,539;6,548,640。术语人源化抗体的意思是免疫球蛋白的骨架区的至少一部分,和任选地涉及结合的CDR区或其他区域的一部分源自或调整为人免疫球蛋白序列。例如,可以例如不按照编码H和L链的小鼠和/或人基因组DNA序列或编码H和L链的cDNA克隆,通过重组DNA技术制备小鼠单克隆抗体的人源化的、嵌合的或部分人源化形式的小鼠单克隆抗体。可以通过由重组DNA技术将非人抗体的CDR区连接至人恒定区来产生人源化形式的小鼠抗体(Queen等人,1989;WO90/07861)。可供选择地,本发明的方法中使用的单克隆抗体可以是人单克隆抗体。例如,可以使用噬菌体展示方法获得人抗体(WO91/17271;WO92/01047)。如本文使用的,人源化抗体也指非人(例如鼠科动物、骆驼、美洲驼、鲨鱼)抗体的形式,其是含有源自非人免疫球蛋白的最少序列的特异性嵌合免疫球蛋白、免疫球蛋白链或其片段(如抗体的Fv、Fab、Fab′、F(ab′)2或其他抗原结合序列)。如本文使用的,人抗体或人源化抗体的意思是具有与由人产生的抗体的氨基酸序列对应的氨基酸序列的抗体和/或使用本领域已知或本文公开的任何用于制备人抗体的技术制备人抗体或人源化抗体。可以通过竞争性结合实验或其他方式选择具有与特定小鼠抗体相同的表位特异性的人抗体。本发明的人源化抗体惊人地在很大程度上分享了小鼠抗体有用的功能特性。也可以提供来自用免疫原性剂免疫的人血清形式的人多克隆抗体。任选地,可以通过使用淀粉样纤维状和/或非纤维状多肽或其片段作为亲和试剂的亲和纯化来浓缩这样的多克隆抗体。单克隆抗体可以获得自根据WO99/60846中描述的技术的血清。本发明进一步涉及如本文描述的抗体或其片段(例如可变区)在识别适用于选择性地结合于靶标的分子或亲和试剂中的用途。根据本发明的亲和试剂、抗体或其片段可以是聚乙二醇化的,其中聚乙二醇化是指聚乙二醇(PEG)聚合物链共价连接至本发明的抗体。可以通过PEG的反应衍生物与靶标分子的温育常规实现聚乙二醇化。对抗体的聚乙二醇化可以潜在地遮盖试剂免遭宿主免疫系统,导致试剂的免疫原性和抗原性降低或试剂的水动力学大小增加,这可以通过减少肾清除率而延长试剂的循环时间。抗体的可变区是指单独的抗体轻链的可变区或抗体重链的可变区,或组合。重链和轻链的可变区各自由通过三个互补决定区(CDR)(又名高变区)连接的四个骨架区(FR)组成.每条链的CDR通过FR很接近地保持在一起,并且与来自其他链的CDR共同形成抗体的抗原结合部位.存在至少两种确定CDR的技术:(1)基于跨物种序列变异性的方法(即Kabat等人.免疫相关蛋白的序列(SequencesofProteinsofImmunologicalInterest),(第五版,1991,国立卫生研究院,马里兰州贝塞斯达));以及(2)基于抗原抗体复合体的晶体学研究的方法(Al-lazikani等人.(1997)J.Molec.Biol.273:927-948)。如本文使用的,CDR可以指通过任一种方法或通过两种方法的组合定义的CDR。在一些实施方式中,本发明提供了抗体,其包括与本发明的抗体的至少1个CDR、至少2个、至少3个或更多个CDR基本上相同的至少1个CDR、至少2个、至少3个或更多个CDR。其他实施方式包括具有与本发明的抗体或源自本发明的抗体的至少2、3、4、5或6个CDR基本上相同的至少2、3、4、5或6个CDR的抗体。在一些实施方式中,至少1、2、3、4、5或6个CDR是与发明的抗体的至少1、2或3个CDR具有至少约85%、86%、87%、88%、89%、90%、95%、96%、97%、98%或99%一致性。理解的是出于本发明的目的,普遍地保留了结合特异性和/或全部活性,虽然相比于所述抗体活性的程度可能变化(可能更大或更小).抗体的半衰期和细胞毒性潜能主要依赖于Fc域与不同Fcγ受体的相互作用。在抗体半衰期的情况下,新生的Fc受体(FcRn)起主要的作用。该受体表达于能够吸收血清蛋白进入其再循环内体中的数种细胞类型和组织上,如单核细胞和血管内皮细胞。在某些内体中,pH降低至接近6,并且在这些条件下,抗体能够结合于FcRn.这种相互作用保护抗体免受降解,直到所述抗体被再次释放入生理pH破坏与受体的结合的血液中(Roopenian和Akilesh(2007)NatRevImmunol7:715-725)。在pH为6下抗体对FcRn的亲和性越高,该抗体的半衰期越长。已知的用于稳定这种相互作用的Fc片段突变总结于Presta(2008,CurrOpinImmunol20:460-470)中。治疗性抗体可以通过结合于它们的靶标时的数种机制起作用。结合本身可以触发信号转导,该信号转导可以导致程序性细胞死亡(Chavez-Galan等人(2009)CellMolImmunol6:15-25)。它也可以通过结合于受体或配体阻断受体与其配体的相互作用。如果对存活重要的信号受到影响,这种干扰可以导致细胞凋亡(Chiu等人(2007)Blood109:729-739)。关于细胞耗竭,存在两个已知的主要效应机制。第一个是对靶细胞的补体依赖性的细胞毒作用(CDC)。存在三种已知的不同途径。然而,在抗体的情况下,CDC的重要途径是经典途径,该途径通过C1q结合于IgG或IgM的恒定区开始(Wang和Weiner(2008)ExpertOpinBiolTher8:759-768)。第二个机制称为抗体依赖性细胞介导的细胞毒性(ADCC)。该效应功能的特征在于募集表达用于抗体的各自的同种型的Fc受体的免疫细胞。ADCC主要通过激活能够单独或作为免疫复合体结合于IgG分子的Fcγ受体(FcγR)介导。小鼠展示三种活化的Fcγ受体(FcγRI、FcγRIII和FcγRIV),并且人展示五种活化的Fcγ受体(FcγRI、FcγRIIA、FcγRIIC、FcγRIIIA和FcγRIIIB)。这些受体表达于天然免疫细胞如粒细胞、单核细胞、巨噬细胞、树突状细胞和自然杀伤细胞,并且因此将天然免疫细胞与适应性免疫系统连接。根据细胞类型,在识别抗体标记的靶细胞时,存在承载FcgR的细胞的数种作用方式。粒细胞通常释放血管活性和细胞毒性物质或化学引诱物,但是也能够进行吞噬作用。单核细胞和巨噬细胞以吞噬作用、氧化爆发、细胞毒性或释放促炎性细胞因子响应,而天然杀伤细胞释放颗粒酶和穿孔素并且也可以通过靶细胞上的FAS与它们的Fas配体的相互作用触发细胞死亡(Nimmerjahn和Ravetch(2008)NatRevImmunol8:34-47;Wang和Weiner(2008)ExpertOpinBiolTher8:759-768;Chavez-Galan等人(2009)CellMolImmunol6:15-25)。也可以通过强化Fc域与激活Fcγ受体(FcγR)的结合来改善抗体依赖性细胞介导的细胞毒性(ADCC)。这也可以通过如Presta中总结的在Fcγ域中的突变来获得(2008,CurrOpinImmunol20:460-470)。改变ADCC的另一种途径是操作存在于每个IgG在Asn297处的糖部分。已知从糖分子的末端的去岩藻糖基化和去除唾液酸以增加抗体的细胞毒性潜能(Anthony和Ravetch(2010)JClinImmunol30Suppl1:S9-14)。保持本发明的所述特性的要求保护的核酸、蛋白质和抗体的序列变体,例如通过要求保护的%序列一致性定义的,也被包含在本发明的范围内。这样的变体显示了可供选择的序列,但是基本上保持相同的结合特性,如靶标特异性,因为提供的特异性序列称为功能性类似物或为功能上类似的.序列一致性涉及当进行序列比对时相同核苷酸或氨基酸的百分比。本领域普通技术人员将领会到,由于遗传密码的简并性,存在许多编码本文描述的多肽的核苷酸序列。这些多核苷酸中的一些对任何天然基因的核苷酸序列具有最小的同源性或序列一致性。尽管如此,本发明特别地考虑了由于密码子使用的差异引起变化的多聚核苷酸。本发明还涵盖落入描述的序列一致性的序列中的删除、替换和其他改变。可通过替换发生的蛋白质序列的修饰也被包括在本发明的范围内。如本文定义的替换是对蛋白质的氨基酸序列进行的修饰,其中一个或多个氨基酸被置换为相同数目的(不同的)氨基酸,产生含有与初级蛋白不同氨基酸序列的蛋白质,优选不显著变更蛋白质的功能。如添加,替换可以是天然的或人工的。在本领域熟知可以在不显著改变蛋白质的功能的情况下进行氨基酸替换。这是特别正确的,当修饰涉及“保守”氨基酸替换,其是一种氨基酸替换为另一种类似特性的氨基酸。这样的“保守”氨基酸可以是天然氨基酸或合成氨基酸,其由于大小、电荷、极性和构象可以被替换而不显著影响蛋白质的结构和功能。频繁地,许多氨基酸可以被保守氨基酸替换而不有害地影响蛋白质的功能。总之,非极性氨基酸Gly、Ala、Val、Ile和Leu;非极性芳香族氨基酸Phe、Trp和Tyr;中性极性氨基酸Ser、Thr、Cys、Gln、Asn和Met;带正电荷的氨基酸Lys、Arg和His;带负电荷的氨基酸Asp和Glu,代表保守氨基酸的组。该清单不是穷举。例如,熟知的是Ala、Gly、Ser和有时候Cys可以互相替换,即使它们属于不同的组。替换变体在移除的抗体分子中具有至少一个氨基酸残基和在其位置插入的不同的残基。最感兴趣的用于替换的突变形成位点包括高变区,但是也考虑FR的改变。如果这样的替换导致生物学活性的改变,那么命名为下表中的“示例性替换”,或下文参考氨基酸类别进一步描述的更大的改变可以被引入并且筛选产物。潜在的氨基酸替换:原始残基优选的保守替换示例性替换的例子Ala(A)ValVal;Leu;IleAsg(R)LysLys;Gln;AsnAsn(N)GlnGln;His;Asp、Lys;ArgAsp(D)GluGlu;AsnCys(C)SerSer;AlaGln(Q)AsnAsn、GluGlu(E)AspAsp;GlnGly(G)AlaAlaHis(H)ArgAsn;Gln;Lys;ArgIle(I)LeuLeu;Val;Met;Ala;Phe;正亮氨酸Leu(L)Ile正亮氨酸;Ile;Val;Met;Ala;PheLys(K)ArgArg;Gln;AsnMet(M)LeuLeu;Phe;IlePhe(F)TyrLeu;Val;Ile;Ala;TyrPro(P)AlaAlaSer(S)ThrThrThr(T)SerSerTrp(W)TyrTyr;PheTyr(Y)PheTrp;Phe;Thr;SerVal(V)LeuIle;Leu;Met;Phe;Ala;正亮氨酸通过选择替换实现抗体的生物学特性的大量修饰,所述替换在它们对保持(a)替换区域的多肽骨架的结构(例如片状或螺旋构象),(b)分子在靶标位点的电荷或疏水性,或(c)大多数的侧链的作用方面显著不同。保守氨基酸替换不限于天然存在的氨基酸,还包括合成氨基酸。通常使用的合成氨基酸是为中性非极性类似物的不同链长的ω氨基酸和环己基丙氨酸;为中性非极性类似物的瓜氨酸和蛋氨酸亚砜,为芳香族中性类似物的苯基甘氨酸;为带负电荷的类似物的磺基丙氨酸和为带正电荷的氨基酸类似物的鸟氨酸。如天然存在的氨基酸,该清单不是穷举的,而仅仅是本领域熟知的替换的示例。可以通过用包括本发明的抗体的编码序列的表达载体转染宿主细胞产生本发明的抗体。通过将抗体的这些编码序列有效地与常规的能够在宿主细胞中控制复制和表达和/或能够从宿主细胞中分泌的调控序列组合来产生表达载体或重组质粒。调控序列包括启动子序列例如CMV启动子和可以源自其他已知抗体的信号序列。相似地,可以产生第二个表达载体,其具有编码互补的抗体轻链或重链的DNA序列。在某些实施方式中,该第二个表达载体与第一个是相同的,除了在相关的编码序列和可选择标记的范围内,如此以尽可能远地确保每个多肽链功能性地表达。可供选择地,抗体的重链和轻链编码序列可以位于单个载体上。通过常规技术用第一和第二载体两者共转染选择的宿主细胞(或通过单个载体简单地转染)以产生本发明的转染的宿主细胞,所述宿主细胞包括重组或合成的轻链和重链两者。然后通过常规技术培养转染的细胞以产生本发明的工程化的抗体。通过适当的测定如ELISA或RIA从培养物中筛选包括重组重链和/或轻链两者的组合的抗体。可以采用相似的常规技术以构建其他抗体。本领域技术人员可以选择适用于在本发明的方法和组合物的构建中采用的克隆和亚克隆步骤的载体。例如,可以使用常规pUC系列的克隆载体。一种载体pUC19是可商购获得的。这样的载体的组件,例如复制子、选择基因、增强子、启动子、信号序列等,可以从商业或天然来源获得,或通过已知的用于指导在选择的宿主中的重组DNA的产物的表达和/或分泌的程序合成。出于该目的,也可以选择其他适当的表达载体,其许多类型已知在本领域中用于哺乳动物、细菌、昆虫、酵母和真菌表达.本发明也涵盖用含有本发明的抗体的编码序列的重组质粒转染的细胞系。用于这些克隆载体的克隆和其他操作的宿主细胞也是常规的。适用于本发明的抗体的表达的宿主细胞或细胞系包括哺乳动物细胞如NS0、Sp2/0、CHO(例如DG44)、COS、HEK,成纤维细胞(例如3T3)和骨髓瘤细胞,例如在CHO或骨髓瘤细胞中表达。可以使用人细胞,因此使分子能够被人糖基化模式修饰。可供选择地,可以采用其他原核或真核细胞系。适合的哺乳动物宿主细胞的选择和转化、培养、扩增、筛选和产物生成及纯化的方法在本领域是已知的。根据本发明,提供了产生本发明的与人CD269结合并中和人CD269活性的抗CD269抗体的方法,所述方法包括以下步骤:提供编码抗体的重链的第一载体;提供编码抗体的轻链的第二载体;用所述第一载体和第二载体转化哺乳动物宿主细胞(例如CHO);在引导抗体从所述宿主细胞中分泌到所述培养基中的条件下培养步骤(c)的宿主细胞;回收步骤(d)中分泌的抗体。一旦表达,可以使用本文描述的方法评估抗体的希望的结合特性。本发明涵盖免疫缀合物(可交换地指“抗体药物缀合物”或“ADC”),所述免疫缀合物包括如本文描述的根据本发明的抗体,所述抗体包括但不限于缀合于一种或多种细胞毒性剂如化学治疗剂、药物、生长抑制剂、毒素(例如蛋白质毒素,细菌、真菌、植物或动物来源的酶活性毒素或其片段)或放射性同位素(即,放射性缀合物)的抗体。用于将治疗剂与蛋白质尤其是抗体缀合(如本发明的抗CD269抗体药物缀合物)的技术是熟知的(见例如,Arnon等人,″用于癌症治疗中药物的靶向免疫的单克隆抗体(MonoclonalAntibodiesForImmunotargetingOfDrugsInCancerTherapy),″单克隆抗体和癌症治疗(MonoclonalAntibodiesAndCancerTherapy)(Reisfeld等人编,AlanR.Liss,Inc.,1985);Hellstrom等人,″用于药物递送的抗体(AntibodiesForDrugDelivery),″控制的药物递送(ControlledDrugDelivery)(Robinson等人编,MarcelDekker,Inc.,第二版.1987);Thorpe,″癌症治疗中的细胞毒性剂的抗体载体(AntibodyCarriersOfCytotoxicAgentsInCancerTherapy:AReview),″单克隆抗体′84:生物和临床应用(MonoclonalAntibodies′84:BiologicalAndClinicalApplications)(Pinchera等人编,1985);″癌症治疗中放射性标记抗体的治疗性用途的分析、结果和未来展望(Analysis,Results,andFutureProspectiveoftheTherapeuticUseofRadiolabeledAntibodyInCancerTherapy),″用于癌症检测和治疗的单克隆抗体(MonoclonalAntibodiesForCancerDetectionAndTherapy)(Baldwin等人编,AcademicPress,1995);以及Thorpe等人,1982,Immunol.Rev.62:119-58。也参见,例如PCT公开WO89/12624.)典型地,ADC或ADC衍生物包括治疗剂和抗CD269抗体或其衍生物之间的连接体区。如上文记载的,在典型的实施方式中,接头在胞内条件下是可切割的,使得在胞内环境下切割接头从抗体释放治疗剂。例如,在一些实施方式中,连接体通过存在于胞内环境的切割剂切割(例如在溶酶体或内体或小窝中)。连接体可以是例如由胞内肽酶或蛋白酶(包括但不限于溶酶体蛋白酶或内体蛋白酶)切割的肽基接头。在其他实施方式中,可切割连接体是pH敏感的,即对在某种pH值下的水解作用敏感。典型地,pH敏感接头是在酸性条件下可水解的。在又一种实施方式中,接头是在还原条件下可裂解的(例如二硫化物接头)。各种二硫化物接头在本领域是已知的(见例如Wawrzynczak等人,免疫缀合物中:癌症的放射性成像和治疗中的抗体缀合物(InImmunoconjugates:AntibodyConjugatesinRadioimageryandTherapyofCancer)(C.W.Vogeled.,OxfordU.Press,1987.也参见美国专利号4,880,935.)。典型地,接头基本上对胞外环境不敏感。在其他的非互相排斥的实施方式中,接头促进细胞内化。在某些实施方式中,当缀合于治疗剂时(即在如本文描述的ADC或ADC衍生物的接头-治疗剂部分的环境中),接头促进细胞内化。在再一种实施方式中,当缀合于治疗剂和抗CD269抗体或其衍生物时(即在如本文描述的ADC或ADC衍生物的环境中),接头促进细胞内化。可以与本组合物和方法使用的多种接头描述于2003年7月31日提交的、名称为“用于治疗癌症、自身免疫疾病或感染疾病的药物缀合物和其用途(DrugConjugatesandTheirUseforTreatingCancer,AnAutoimmuneDiseaseoranInfectiousDisease)”的WO2004010957,以及2002年7月31日提交的、,名称为“用于治疗癌症、自身免疫疾病或感染疾病的药物缀合物和其用途(DrugConjugatesandTheirUseforTreatingCancer,AnAutoimmuneDiseaseoranInfectiousDisease)”的美国临时申请号60/400,403(其公开内容通过引用并入本文中)。在某些实施方式中,免疫缀合物包括如本文描述的抗体,所述抗体包括但不限于抗体和化学治疗剂或其他毒素。可以使用的酶活性毒素和其片段包括白喉毒素A链、白喉毒素的非结合活性片段、外毒素A链(来自绿脓单孢菌(Pseudomonasaeruginosa))、蓖麻毒蛋白A链、相思豆毒蛋白A链、蒴莲根毒蛋白IIA链、α-帚曲霉素、油桐(Aleuritesfordii)蛋白、香石竹毒蛋白、美洲商陆(Phytolacaamericana)蛋白(PAPI、PAPII和PAP-S)、苦瓜(momordicacharantia)抑制剂、麻疯树毒蛋白、巴豆毒蛋白、肥阜草(sapaonariaofficinalis)抑制剂、多花白树毒蛋白、丝林霉素(mitogellin)、局限曲菌素、酚霉素、依诺霉素和单端孢霉烯类(tricothecenes)。各种放射性核素可以用于放射性缀合抗体的产生。本发明的抗体或其片段也可以缀合至一种或多种毒素,所述毒素包括但不限于加利车霉素、美登木素生物碱、多拉司他汀(dolastatins)、奥罗他汀(aurostatins)、单端孢霉烯和CC1065,以及这些毒素的具有毒素活性的衍生物。适合的细胞毒性剂包括但不限于,包括多拉缬氨酸-缬氨酸-多拉异亮氨酸-多拉脯氨酸-苯丙氨酸(MMAF)和单甲基奥瑞他汀E(monomethylauristatinE)(MMAE),以及酯形式的MMAE、DNA小沟结合剂、DNA小沟烷基化剂、烯二炔、莱克西调理素(lexitropsin)、多卡霉素、包括紫杉醇和多西紫杉醇的紫杉烷、嘌呤霉素、多拉司他汀、美登木素生物碱以及长春花生物碱。特异性细胞毒性剂包括拓扑替康、吗啉代阿霉素、根霉素、氰基吗啉代阿霉素,多拉司他汀-10,棘霉素,考布他汀(combretatstatin),杯形生长抑制素(chalicheamicin),美登素,DM-1,DM-4,纺锤菌素.其它适合的细胞毒性剂包括抗微管蛋白剂,如奥瑞他汀,长春花生物碱,鬼臼毒素,紫杉烷,浆果赤霉素衍生物,念珠藻素(cryptophysin),美登木素生物碱,考布他汀(combretastatin)或多拉司他汀。抗微管蛋白剂包括二甲基缬氨酸-缬氨酸-多拉异亮氨酸-多拉脯氨酸-苯丙氨酸-对-亚苯基二胺(AFP),MMAF,MMAE,奥瑞他汀E,长春新碱,长春花碱,长春地辛,长春瑞滨,VP-16,喜树碱,紫杉醇,多西他赛,埃博霉素A,埃博霉素B,诺考达唑,秋水仙素,秋水酰胺(colcimid),雌莫司汀,西马多丁,圆皮海绵内酯,美登素,DM-1,DM-4或五加苷素(eleutherobin)。在一些实施方式中,免疫缀合物包括与多拉他汀或多拉司他汀肽类似物和衍生物奥瑞他汀缀合的抗体(美国专利号5,635,483;5,780,588),多拉他汀和奥瑞他汀已经显示干扰微管动力学、GTP水解以及核和细胞分裂(Woyke等人(2001)抗微生物剂和化学治疗(Antimicrob.AgentsandChemother).45(12):3580-3584)并具有抗癌(美国专利号5,663,149)和抗真菌活性(Pettit等人(1998)抗微生物剂和化学治疗(Antimicrob.AgentsandChemother).42:2961-2965)。多拉他汀和奥瑞他汀(其是多拉他汀的五肽衍生物)药物部分可以通过肽药物部分的N(氨基)末端或C(羧基)末端连接于抗体(WO02/088172)。示例性的奥瑞他汀实施方式包括N-末端连接的单甲基奥瑞他汀药物部分DE和DF,其公开于“能够缀合至配体的单甲基缬氨酸化合物(MonomethylvalineCompoundsCapableofConjugationtoLigands)”,美国专利号7,498,298。如本文使用的,缩写“MMAE”是指单甲基奥瑞他汀E。如本文使用的,缩写“MMAF”是指多拉缬氨酸-缬氨酸-多拉异亮氨酸-多拉脯氨酸-苯丙氨酸。典型地,可以通过在两个或更多个氨基酸和/或肽片段之间形成肽键制备基于肽的药物部分。可以制备这样的肽键,例如,根据肽化学领域熟知的液相合成方法(见Schroder和K.Lubke,″肽(ThePeptides),″第1卷,第76-136页,1965,学术出版社)。美登木素生物碱可以用作偶联于根据本发明的抗体或其片段的活性剂。美登木素生物碱是有丝分裂抑制剂,其通过抑制微管蛋白聚合起作用。从东非灌木齿叶美登木(Maytenusserrata)中首次分离出美登素(美国专利号3,896,111)。随后,发现某些微生物也可以产生美登木素生物碱,如美登醇和C-3美登醇酯(美国专利号4,151,042)。可以从由微生物如束丝放线菌属(Actinosynnema)的发酵产生的安丝菌素前体制备高细胞毒性美登木素生物碱药物。通过化学连接抗体至美登木素生物碱分子制备抗体-美登木素生物碱缀合物而不显著减少抗体或美登木素生物碱分子的生物活性。见例如美国专利号5,208,020。每个抗体分子平均缀合3-4个美登木素生物碱分子,显示了在提高靶细胞细胞毒性而不负面地影响抗体的功能或溶解性方面的有效性,虽然即使一个分子的毒素/抗体也被预期提高细胞毒性高于使用无修饰的抗体。美登木素生物碱在本领域是熟知的,并且可以通过已知的技术合成或从天然来源分离。选择的抗生素的加利车霉素家族的例子可以用作偶联于根据本发明的抗体或其片段的活性剂.抗生素的加利车霉素家族能够在亚皮摩尔浓度下产生双链DNA破坏。对加利车霉素家族的缀合物的制备,见美国专利号5,712,374、5,714,586、5,739,116、5,767,285、5,770,701、5,770,710、5,773,001、5,877,296。另一种可以缀合抗体的抗肿瘤药物是抗叶酸剂QFA。加利车霉素和QFA两者都具有胞内作用部位,并且不容易穿过质膜。因此,这些药剂通过抗体介导的内化的细胞摄取大大提高了它们的细胞毒性作用。其他可以缀合于抗体的抗肿瘤剂包括BCNU、链脲霉素(streptozoicin)、长春新碱和5-氟尿嘧啶,在美国专利号5,053,394、5,770,710中描述的统称为LL-E33288复合物的药剂家族,以及埃斯培拉霉素(esperamicins)(美国专利号5,877,296)。本发明进一步考虑在抗体和具有溶核活性的化合物(例如核糖核酸酶或DNA核酸内切酶如脱氧核糖核酸酶;DNA酶)之间形成免疫缀合物。对于选择性破坏肿瘤,抗体可以包括高放射性的原子。从本发明的意义上来说,药学上可接受的载体可以是在有害意义上不显著干扰本发明的抗体的生物活性的有效性的任何非毒性物质。显然,载体的特征会依赖于给药的途径。这样的组合物可包含除了活性物质和载体之外,在本领域熟知的稀释剂、填充剂、盐、缓冲剂、稳定剂、增溶剂和其他物质。药学上可接受的赋形剂和载体溶液的制剂对本领域技术人员来说是熟知的,如同在多种治疗方案例如口服、非胃肠、静脉内、鼻内和肌内给药和制剂中,用于使用本文描述的具体的组合物的适当的给药和治疗方案的发展。含有活性成分(抗体或抗体片段)的药物,或者称为药物组合物可以是适用于口服的形式,例如片剂、锭剂(troche)、糖锭(lozenge)、水性或油性悬浮液、可分散粉末或颗粒、乳剂、硬或软胶囊、或糖浆剂或酏剂。可以根据制造药用组合物领域已知的任何方法制备用于口服使用的组合物,并且这样的组合物可以含有一种或多种选自由以下组成的组的药剂:甜味剂、调味剂、着色剂和防腐剂以提供药学上美观并可口的制剂.片剂含有与适用于制造片剂的非毒性药学上可接受的赋形剂混合的活性成分。这些赋形剂可以是例如,惰性稀释剂,如碳酸钙、碳酸钠、乳糖、磷酸钙或磷酸钠;制粒剂和崩解剂,例如玉米淀粉或海藻酸;结合剂,例如淀粉、明胶或阿拉伯树胶,以及润滑剂,例如硬脂酸镁、硬脂酸或滑石。片剂可以是无包衣的或它们可以通过已知的技术包衣以延迟在胃肠道中的分解和吸收,从而在更长的一段时间内提供持续的作用。例如可以采用延时物质如单硬脂酸甘油酯或二硬脂酸甘油酯。它们也可以被包衣。本发明也指用于局部施用、口服、吸入或皮肤、皮下或静脉内注射的药用组合物。技术人员知道用于具体的施用形式需要的载体和添加剂。当通过静脉内注射、皮肤注射或皮下注射施用治疗有效量的本发明的活性物质(抗体或抗体片段)时,活性物质可以是无热原的、非胃肠可接受的水溶液的形式。本发明还涉及在治疗具有如本文公开的医学紊乱的受试者中施用治疗相关量的本文描述的抗体。如本文使用的,术语“治疗有效量”的意思是足以显示有意义的患者利益的药物组合物或方法的每种活性组分的总量。本发明的药物组合物中的活性物质的量取决于要治疗的病症的性质和严重性,以及患者已接受的在先治疗的性质。可以施用更大剂量直到患者获得了最优的治疗效果,并且在该点,不再进一步增加剂量。这样的非胃肠可接受的溶液的制备应该注意pH、等渗性和稳定性等,该制备在本领域的技术内。用于静脉内、皮肤或皮下注射的优选的药物组合物除了活性物质外,还应该含有等张溶媒,如氯化钠注射液,林格氏注射液,葡萄糖注射液,葡萄糖和氯化钠注射液,乳酸林格氏注射液或本领域已知的其他溶媒。本发明的药物组合物也含有稳定剂、防腐剂、缓冲剂、抗氧化剂或本领域技术人员已知的其它添加剂。施用的抗体的剂量显然取决于本领域熟知的许多因素,例如抗体的化学性质和药物制剂,和患者的体重、体表、年龄和性别,以及施用的时间和途径。对于成人,剂量示例性地可以为每天0.001μg至1g,优选为每天0.1μg至100mg,更优选为每天1μg至100mg,甚至更优选为每天5μg至10mg。在连续输注中,剂量示例性地为每分钟每千克体重0.01μg至100mg,优选为1μg至10mg。在本发明的另一方面,提供了如本文描述的根据本发明的抗体用于治疗选自以下的B细胞介导的或浆细胞介导的疾病或抗体介导的疾病或紊乱:多发性骨髓瘤(MM)、慢性淋巴细胞白血病(CLL)、非分泌型多发性骨髓瘤、冒烟型多发性骨髓瘤、意义未定的单克隆丙种球蛋白病(MGUS)、孤立性浆细胞瘤(骨、髓外的)、淋巴浆细胞淋巴瘤(LPL)、原发性巨球蛋白血症(Waldenstrom′sMacroglobulinemia)、浆细胞白血病、原发性淀粉样变性(AL)、重链疾病、系统性红斑狼疮(SLE)、POEMS综合征/骨硬化性骨髓瘤、I型和II冷球蛋白血症、轻链沉积病、肺出血肾炎综合征(Goodpasture′ssyndrome)、特发性血小板减少性紫癜(ITP)、急性肾小球肾炎、天疱疮和类天疱疮症紊乱以及获得性大疱性表皮松解(Epidermolysisbullosaacquisita);或任何具有BCMA表达的非霍奇金淋巴瘤B细胞白血病或霍奇金淋巴瘤(HL),或患者产生对重组蛋白替代疗法的中和抗体的任何疾病,其中所述方法包括向所述患者施用治疗有效量的本文描述的抗体的步骤。可以将B细胞紊乱分为B细胞发育/免疫球蛋白产生的缺陷(免疫缺陷)和过度的/不受控制的增殖(淋巴瘤、白血病)。如本文使用的,B细胞紊乱是指两种类型的疾病,并且提供了用抗体治疗B细胞紊乱的方法。在本发明的一个方面,疾病是多发性骨髓瘤还提供了如本文描述的抗体在制造用于治疗如本文描述的疾病和紊乱的药物中的用途.例如在本发明的一个方面,提供了如本文描述的抗体用于治疗或预防响应于BCMA和配体BAFF和APRIL之间的相互作用的调节(如抑制或阻断)的疾病和紊乱的用途。在本发明的一种实施方式中,分离的抗体或抗体片段用于治疗B淋巴细胞癌症,例如霍奇金淋巴瘤.在本发明的一种实施方式中,分离的抗体或抗体片段用于治疗自身免疫疾病,如与炎症相关的医学紊乱,优选具有炎性组分的自身免疫疾病,其中自身免疫疾病选自大动脉炎、巨细胞动脉炎、家族性地中海热、川崎病、结节性多动脉炎、皮肤结节性多动脉炎、肝炎相关的动脉炎、白塞氏综合征、韦格纳肉芽肿病、ANCA-血管炎(ANCA-vasculitidies)、许尔-斯特劳斯综合征(Churg-Strausssyndrome)、显微镜下多血管炎、结缔组织疾病血管炎、紫癜、冷球蛋白血症血管炎、皮肤白细胞碎裂性脉管炎、热带主动脉炎、肉样瘤病、科根综合征、威斯科特-奥尔德里奇综合征(Wiskott-AldrichSyndrome)、麻风结节性动脉炎、CNS的原发性血管炎、血栓闭塞性脉管炎、副肿瘤血管炎(Paraneoplasticateritis)、荨麻疹、Dego病、骨髓增生异常综合征、持久性隆起红斑、高免疫球蛋白D、过敏性鼻炎、支气管哮喘、慢性阻塞性肺疾病、牙周炎、类风湿性关节炎、动脉粥样硬化、淀粉样变性、克罗恩病(MorbusChron)、溃疡性结肠炎、自身免疫性肌炎、糖尿病、多发性硬化症、格林巴利综合征(Guillain-BarreSyndrome)、组织细胞增生症、骨关节炎、过敏性皮炎、牙周炎、慢性鼻窦炎、牛皮癣、牛皮癣关节炎、显微镜下结肠炎、肺纤维化、肾小球肾炎、惠普尔氏病(Whipple′sdisease)、斯蒂尔病(Still′sdisease)、结节性红斑、耳炎、冷球蛋白血症、干燥综合征(Sjogren′ssyndrome)、红斑狼疮、再生障碍性贫血、骨髓纤维瘤、慢性炎症性脱髓鞘性多发性神经病、木村病、系统性硬化症、慢性主动脉周围炎、慢性前列腺炎、特发性肺纤维化、慢性肉芽肿病、特发性贲门失弛缓症、博莱霉素诱导的肺部炎症、阿糖胞苷诱导的肺部炎症、自身免疫性血小板减少症(Autoimmunthrombocytopenia)、自身免疫性中性粒细胞减少症(Autoimmunneutropenia)、自身免疫性溶血性贫血、自身免疫性淋巴细胞减少症、恰加斯氏病(Chagas′disease)、慢性自身免疫性甲状腺炎、自身免疫性肝炎、桥本氏甲状腺炎、萎缩性甲状腺炎(atropicthyroiditis)、甲状腺机能亢进(Gravesdisase)、自身免疫性多腺性综合征、自身免疫性艾迪生综合征(AutoimmuneAddisonSyndrome)、寻常型天疱疮、落叶型天疱疮、疱疹样皮炎、自身免疫性脱发、白癜风、抗磷脂综合征、重症肌无力、僵人综合征、古德帕斯彻综合征(Goodpasture’ssyndrome)、交感性眼炎、毛囊炎、夏普综合征(Sharpsyndrome)和/或伊文思综合征(Evanssyndrome),特别是枯草热、牙周炎、动脉粥样硬化、类风湿性关节炎,优选是类风湿性关节炎或多发性硬化症。序列本发明的优选的抗体序列:优选的核苷酸序列属于CD269(BCMA)的本发明的优选的序列:包括人源化序列修饰的优选的广义氨基酸序列:附图说明通过以本文公开的实施例和附图的示例性方式说明本发明。本文提供的附图代表本发明的具体的实施方式,并且不意图限制本发明的范围。附图被认为提供了增强一个或多个非限制性实施方式的技术支撑的可能和潜在优选实施方式的进一步描述。附图简述:图1:J22.9-xi的体外表征。图2:CD269(BCMA)的结构和J22.9-xiFab:CD269复合体。图3:J22.9-xi的体外细胞毒性。图4:在异种移植NSG小鼠中J22.9-xi的效果。图5:建立的肿瘤的治疗。图6:疾病早期的肿瘤治疗.图7:杂交瘤J22.9的不稳定性。图8:与J22.9-xi比较的HC的人源化序列的序列。图9:与J22.9-xi比较的LC的人源化序列的序列。图10:J22.9-xi的序列优化变体在ELISA中显示相似的结合。图11:J22.9-xi的序列优化变体在流式细胞术中显示相似的结合。图12:SPR原始数据。图13:抗体变体的凝胶电泳.附图的详细描述:图1:J22.9-xi的体外表征。以ELISA(a)或通过使用CD269阳性的MM.1S细胞的流式细胞术(b),J22.9-xi与BCMA浓度依赖性结合。(c)采用指示浓度的BCMA,通过表面等离子体共振测量确定J22.9-xi与BCMA的结合亲和性。(d)J22.9-xi阻断BAFF和吸附于微量滴定板上的BCMA之间的相互作用。图2:(a)CD269(BCMA)识别表面。显示BAFF/APRIL与J22.9-xi的结合表位残基的CD269(BCMA)胞外域的三个视图。上部图中,CD269(BCMA)的结合表面的直接视图:浅灰色暗影表明如从它们的晶体结构辨别的包括BAFF和APRIL的结合表位的所有残基,黑色残基(显示为球体)不接触任一个BAFF、APRIL或J22.9-xi;涉及J22.9-xi结合的表位残基的亚群以表面表示法显示;以球体显示的剩余的浅灰色残基是BAFF和APRIL两个表位的部分但不与J22.9-xi直接接触。中间和下面的图以与上面的图相同的表示法显示,但是分别朝向并远离观察者旋转90°。(b)J22.9-xiFab:CD269复合体的两个视图。J22.9-xi以表面表示法,重链为浅灰色和轻链为暗灰色显示。CD269(BCMA)以结合于J22.9-xi抗原穴区的带状表示法显示。在左侧为Fab:CD269复合体的全视图;在右侧为斜向观察者以显示结合穴的复合体。图3:J22.9-xi的体外细胞毒性。(a)以效应物与靶标的比为20∶1的比使CD269阳性的MM.1S-Luc细胞与人PBMC混合,与指示浓度的J22.9-xi温育4小时。打开符号表明无-N-聚糖的J22.9-xi与来自供体1和供体2的PBMC孵育时的细胞毒性活性。误差线表明SEM.(b)去糖基化不影响J22.9-xi与MM.1S细胞的结合。(c)J22.9-xi在4℃或-80℃下储存3周不影响细胞毒性。图4:在异种移植NSG小鼠中J22.9-xi的效果。(a)每周两次施用200μgJ22.9-xi或对照抗体小鼠以及未处理的对照小鼠随时间的肿瘤发展。(b)在第6至41天之间总的肿瘤负荷((a)的曲线下面积(AUC))。绘制的为平均值与SEM(**P<0.01,***P<0.001,t-检验)。(c)J22.9-xi和同种型对照小鼠的总存活率。使用对数秩(Mantel-Cox)检验计算P值。(d-1)在指示的没有施用治疗性抗体的组中MM.1S-Luc细胞的检测。在最右侧图像下方,数字(41、41、44、40)表明特定的小鼠死亡时注射肿瘤细胞后的天数。(d-2)在第21和28天在指示的组中MM.1S-Luc细胞的检测。背面观。(e)J22.9-xi浓度和肿瘤发展之间的关系。(f)在第6至42天之间的总肿瘤负荷((e)的AUC)。平均值与SEM(**P<0.01,***P<0.001,t-检验)。(g)实验时间线的概览。图5:建立的肿瘤的治疗。(a)每周两次施用200μgJ22.9-xi或对照抗体的小鼠以及未处理的对照小鼠随时间的肿瘤发展。(b)在第8至48天之间总的肿瘤负荷((a)的AUC).绘制的为平均值与SEM(**P<0.01,***P<0.001,t-检验)。(c)J22.9-xi和同种型对照小鼠的总存活率。使用对数秩(Mantel-Cox)检验计算P值。在图5d中提供实验时间线的概览。图6:疾病早期的肿瘤治疗。(a)当用2μg、20μg或200μgJ22.9-xi,或200μg同种型对照抗体或无N-聚糖的J22.9-xi处理时的肿瘤生长过程,以及无肿瘤的肿瘤生长过程。(b)在第9至44天内总的肿瘤负荷((a)的AUC)。显示的是平均值与SEM(**P<0.01,***P<0.001,t-检验)。(c)抗体处理的小鼠和对照异种移植SCID-Beige小鼠的存活率。使用对数秩(Mantel-Cox)检验计算P值。在图6d中提供实验时间线的概览。图7:杂交瘤J22.9的不稳定性。在第1天在BCMA涂覆的微量滴定板上的ELISA中测试的杂交瘤J22.9的上清为与BCMA结合呈阳性。在指示的时间点的后续分析揭示上清的结合能力的减少。在第7、14和21天更换培养基。图8:与J22.9-xi比较的人源化抗体序列的总结。使用标准比对软件进行序列的比较。图9:与J22.9-xi比较的人源化抗体序列的总结。使用标准比对软件进行序列的比较。图10:通过使用人BCMA(hBCMA)或猕猴BCMA(cyBCMA)涂覆的微量滴定板测试嵌合J22.9-xi和人源化变体的结合(J22.9-H对应于人源化序列SEQIDNo.27;J22.9-FSY对应于人源化和PTM修饰的SEQIDNo.28;J22.9-ISY对应于人源化和PTM修饰的SEQIDNo.29)。图11:通过使用人MM细胞系RPMI-8226的流式细胞术测试嵌合J22.9-xi和人源化变体的结合(J22.9-FSY对应于人源化和PTM修饰的SEQIDNo.28;J22.9-ISY对应于人源化和PTM修饰的SEQIDNo.29)。图12:SPR原始数据:通过表面等离子体共振(SPR)谱法测量嵌合J22.9-xi和人源化变体与人BCMA(图12A)和猕猴BCMA(图12B)的结合亲和性。通过胺化学法将IgG固定于ProteonGLH传感器芯片,并在流动相用BCMA测量结合.在图中原始数据的顺序跟对应于图的说明中列出的样品的顺序。图13:抗体变体的凝胶电泳。抗体变体在非还原性SDS-PAGE中电泳跑样(run),并且将抗体变体着色以显示蛋白迁移。实施例通过本文公开的实施例的方式证明本发明.本文提供的实施例仅代表本发明的具体实施方式,并且不意图限制本发明的范围。实施例被认为提供增强一个或多个非限制性实施方式的技术支撑的可能和潜在的优选实施方式的进一步描述。虽然关于抗体-表位复合体的结晶及体内和体外抗肿瘤作用的实施例使用原始的嵌合抗体J22.9-xi进行,但是发明人声称在本发明的人源化变体中保持了这些技术效果,由于与测试的原始嵌合抗体相比在人源化变体中保持了结合特征。因此提供的来自嵌合抗体的数据作为要求保护的人变体的工业适用性和有用性的参考材料和指示提供。初始的生物数据表明J22.9-xi和人源化变体之间的相当的作用。J22.9-xi和BCMA相互作用的结合和阻断特征新的嵌合抗体(J22.9-xi)结合于人CD269(BCMA、TNFRSF17)的胞外域。这最初通过基于人多发性骨髓瘤细胞系MM.1S的ELISA和流式细胞术确定(图1a,b)。使用表面等离子体共振(SPR)确定J22.9-xi与BCMA的亲和性。如图1c显示的平均Kd为54pM。已知BCMA在体内通过与其配体BAFF和/或APRIL的相互作用触发对多发性骨髓瘤和浆细胞的存活重要的信号(MackayF等人(2003)AnnuRevImmunol21:231-264)。因此用人BCMA的胞外域和重组BAFF进行体外阻断测定。J22.9-xi与BCMA的结合明显地阻断受体和其配体BAFF之间的相互作用。使用同种型对照抗体代替J22.9-xi,重组BAFF与BCMA的结合不受影响(图1d)。J22.9-xi-Fab-BCMA复合体的晶体结构揭示与BCMA的广泛结合界面由J22.9-xi制备的Fab片段在与纯化的46个氨基酸残基的BCMA胞外域的复合体中结晶,并且复合体结构以1.9埃的解析度解出。对BCMA的第6至41个残基可观察到高质量的电子密度,显示与J22.9-xi,主要是与抗体的轻链的广泛相互作用(图2B)。该隐藏740.4平方埃并涉及三分之一BCMA残基的界面覆盖了在与APRIL和sTALL1(也称为BAFF)的BMCA复合体的晶体结构中观察到的相同表位的16个残基中的12个残基,包括保守的DxL基序(GordonNC等人(2003),BAFF/BlysS受体3包括编码高度集中的配体结合位点的最小TNF受体样组件(BAFF/BlysSreceptor3comprisesaminimalTNFreceptor-likemodulethatencodesahighlyfocusedligand-bindingsite).Biochemistry42(20):5977-83以及PatelDR等人(2004),工程化APRIL特异的B细胞成熟抗原(EngineeringanAPRIL-specificBCellMaturationAntigen).JBC279(16):16727-35),提供在与BAFF的体外测定中所见的阻断作用的清晰合理化(图2A)。与J22.9-xi的相互作用另外包括与BCMA中的Ala20和Pro23接触的直接侧链,不是被BAFF和APRIL覆盖的结合表位部分的残基,以及数个水介导的氢键。在所有三种结构中BCMA的总体构象是非常相似的,在J22.9-xi和APRIL复合体之间的1.4埃以及J22.9-xi和sTALL1复合体之间的1.5埃的C-αrmsd;J22.9-xiBCMA结合表位(第13至30个残基)的各自的C-αrmsd为0.98和0.88埃。虽然认识到相同的BCMA表位在其核心具有DxL基序,但是J22.9-xi的结合位点非常不同于sTALL1和APRIL的结合位点,如同包括界面的相互作用的结合一样。如可以从图2B和表1和表2中所见,来自J22.9-xi的19个氨基酸(6个来自重链(表1),13个来自轻链(表2))形成了与来自CD269的胞外域的12个残基的直接连接。______________________________________________________表1:J22.9-xi的重链和BCMA之间的氨基酸相互作用的列表。使用软件PDBsum生成这些相互作用列表(LaskowskiRA(2009))。_____________________________________________________表2:J22.9-xi的轻链和BCMA之间的氨基酸相互作用的列表。使用软件PDBsum生成这些相互作用列表(LaskowskiRA(2009))。______________________________________________________表3:涉及CD269:APRIL和CD269:BAFF结合的残基的相互作用列表(通过J22.9不直接接触的残基是下划线的)。使用软件PDBsum生成这些相互作用列表(LaskowskiRA(2009)).______________________________________________________表4:通过J22.9、APRIL和/或BAFF的直接接触结合的CD269靶标的残基。仅通过J22.9直接接触的CD269靶标的残基是下划线的(20、23)。通过J22.9不直接接触的CD269靶标的残基是粗体的(对于APRIL为30、31、33、34、35;对于BAFF为25、29、30、31、34、35)。______________________________________________________表5:J22.9水相互作用(J22.9-xi:H2O:CD269)。使用软件LigPlot(Wallace和Laskowski,欧洲生物信息学研究所)生成表5中的数据(sc=侧链H键;mc=主链H键)在去糖基化后J22.9-xi的强烈细胞毒性功效强烈降低使用转导荧光素酶的MM.1S-Luc细胞系建立基于荧光素酶的细胞毒性测定。在该测定中,因为由培养基中缺乏ATP而使死细胞释放的荧光素酶不能发挥作用,因此仅从活细胞中检测到生物发光。分离来自健康供体的PBMC并与MM.1S-Luc细胞以20∶1的比混合。4小时后测量生物发光。在选择4个未刺激的供体的PBMC制剂下,确定J22.9-xi的体外细胞毒性。细胞毒性潜能在来自不同供体的PBMC之间轻微地变化。在温育的4小时内,细胞裂解在J22.9-xi浓度为125ng/ml下达到18至35%。将J22.9-xi浓度增加至1μg/ml增加细胞裂解高达56%(图3a)。在J22.9-xi用PNGaseF去糖基化后(J22.9-xi-N-聚糖),细胞毒性活性下跌到低于8%,而J22.9-xi-N-聚糖与BCMA阳性MM.1S细胞的结合保持不变(图3a,b)。J22.9-xi减少在异种移植小鼠中的肿瘤负荷并延长存活期我们使用NODscid常见γ链敲除(NSG)缺少功能性B、T和NK细胞群的小鼠。这些用1×107个MM.1S-Luc细胞静脉内注射的小鼠在6周内发展出后肢瘫痪(图4d-1)。第一个症状出现的日期定义处死日期。在尾动脉中注射1×107个MM.1S-Luc后,将小鼠随机分至3个组中。第一组(n=2)不接受处理直到实验结束,而第二组(n=5)和第三组(n=6)分别接受每周两次注射200μg同种型对照或J22.9-xi抗体。从肿瘤细胞注射的日期开始腹腔内(i.p.)施用抗体6周的时间。每周一次使用IVIS谱监测肿瘤生长。在腹腔内注射荧光素后3分钟测量生物发光。从未处理组和接受对照抗体的组中都能发现肿瘤发展的相似过程,而从第6天的第一次测量点已经开始的,用J22.9-xi处理的组显示了显著更低的肿瘤负荷(图4a)。另外,该组显示了在整个监测期期间更小的总体肿瘤荷载(图4b)。同种型对照处理的动物在注射细胞后具有46天的中位存活期。接受J22.9-xi的小鼠平均多存活了26天。这对应于与接受了对照抗体的小鼠比较,55%的扩大的存活率(图4c)。在细胞注射后第28天,在未处理的小鼠和接受同种型对照抗体的小鼠中看到肿瘤细胞大量浸润至脊柱和腹股沟淋巴结内(图4d-2)。将200μg抗体施用于小鼠对应于大约10mg/kg体重。为检测J22.9-xi在更低剂量下的功效,我们将MM.1S-Luc异种移植的小鼠分为4组。第一组(n=7)每周两次接受200μg对照抗体,第二组、第三组(每组n=3)和第四组(n=9)分别每周两次注射2μg、20μg或200μg。如上文描述进行注射和监测。虽然在对照组小鼠中肿瘤如预计发展,但在接受20μg或200μgJ22.9-xi的组中观察到肿瘤生长被显著地限制(图4e,f)。在图4g中提供了实验时间线的概览。在J22.9-xi治疗期间建立的肿瘤的生长停止5周通过将抗体处理的开始推迟至注射肿瘤细胞后5天模仿治疗性施用。将异种移植小鼠分成两组(n=6).动物接受每周两次每次注射200μg同种型对照或J22.9-xi抗体。在细胞注射后第8天完成第一次处理。虽然至第35天在接受J22.9-xi的组(n=5)中不存在可测量的源自肿瘤的生物发光,但在接受同种型对照抗体的动物(n=6)中观察到肿瘤荷载的稳定增长(图5a,b)。来自同种型对照组的小鼠在细胞注射后平均存活56天,而接受J22.9-xi的所有小鼠在第77天时仍然活着(图5c)。在图5d中提供了实验时间线的概览。用J22.9-xi加强早期治疗防止肿瘤生长7周为进一步评估治疗时机对肿瘤生长的作用,从肿瘤细胞注射日开始在连续5天内施用了不同的抗体。在静脉内注射细胞之后,将动物随机分为5组。第一组(n=5)用200μg同种型对照抗体/注射(腹腔注射)处理,而第二组(n=6)接受200μg/注射J22.9-xi-N=聚糖抗体。第三组(n=4)、第四组(n=5)和第五组(n=5)的小鼠分别获得200μg、20μg和2μg/注射J22.9-xi抗体。生物发光测量在细胞注射后第9天开始。至第44天,在任何接受了完整J22.9-xi抗体的组中没有看到源自肿瘤的生物发光。虽然在用J22.9-xi-N-聚糖处理的动物中肿瘤生长是减速的,但是总体肿瘤荷载与接受同种型对照抗体的那些动物没有显著区别(图6a,b)。虽然用J22.9-xi-N-聚糖(去糖基化的)处理的动物的总体肿瘤荷载没有显著不同(图6b),但是与isoAb处理组相比这些小鼠的寿命大幅增加(图6c).因为J22.9-xi-N-聚糖显示不能诱导ADCC或CDC,该结果表明单独的J22.9-xi与BCMA的结合阻碍肿瘤生长。可以合理地认为,这是因为阻断了受体和它的天然配体(APRIL和BAFF)之间的相互作用。在图6d中提供了实验时间线的概览。J22.9-xi的人源化J22.9-xi抗体是基于序列比对和从晶体结构获得的数据人源化的。使用IgBLAST(NCBI)或Clustal(EBI)将可变区序列与它们各自的人同源物比对。在变更前通过目测结构评价每个提出的突变。人源化变体与BCMA靶标的结合使用流式细胞术、ELISA和SPR测试突变体与BCMA的结合。使用表面等离子体共振(ProteOnTMXPR36;Bio-Rad)测量人源化抗体的亲和性。结合数据显示关于人源化抗体变体与如测试的J22.9-xi结合的相同表位的特异性和亲和性的惊人的结果。如在下表中显示,完全惊人的是,如本文描述的人源化抗体展示与原始嵌合抗体的相当的结合特征。SPR数据揭示人源化变体的亲和性与嵌合体的亲和性相似,并且足以根据本文提供的原始嵌合抗体的数据假设它们的临床相关性。技术人员不会预料到通过在嵌合体的人源化期间修饰CDR,维持结合特征至这样的程度。如本文描述的使用BCMA涂覆的微量滴定板(1μg/ml)进行ELISA。如图10中观察到的,所有人源化变体与J22.9-xi相比,使用人和猕猴BCMA的结合是相当的。也使用本文描述的人源化变体进行了流式细胞术,并显示了所有人源化变体与人和猕猴BCMA两者的等价结合(参考图11)还进行了SPR分析,并且测量人源化抗体变体的亲和性。如在下表(表6)中可以观察到的,人源化变体的亲和性(J22.9-H对应于人源化序列SEQIDNo.27;J22.9-FSY对应于人源化和PTM修饰的SEQIDNo.28;J22.9-ISY对应于人源化和PTM修饰的SEQIDNo.29).表6.SPR数据J22.9重链的CDR2中的第54位氨基酸:为了移除人源化J22.9中潜在的翻译后修饰位点,将重链CDR2的残基D54突变为天冬酰胺(N),无意间产生了用于N-连接糖基化的新的潜在修饰位点。含有N54的突变的重链在SDS凝胶上迁移更慢(图13),指示更大的尺寸,并且CDR被糖基化。然而相应的IgG、J22.9-FNY在FACS和ELISA中结合BCMA,并且在与BCMA的复合体中结晶。虽然不完全精确,但是2.7埃解析度的结构显示从N54侧链延伸的清晰的电子密度-与残基的糖修饰一致。惊人的是,侧链这样大的延伸不会破坏与BCMA的结合,并且从这些观察结果中可以预期在这个位置可以容忍多个和不同的氨基酸替换,潜在地还有除了糖之外的衍生化。方法细胞系和培养人多发性骨髓瘤细胞系MM.1S(Greenstein等人(2003)ExpHematol31:271-282)获得自Prof.B.(MDC,柏林,德国)。为了在体内监测肿瘤细胞生长,将荧光素酶和GFP克隆至慢病毒载体系统ViraPower(Invitrogen)的pFU载体内。通过转导细胞的GFP表达,使用荧光激活的单个细胞分选分离单克隆细胞系。在不含酚红,含有10%胎牛血清、100单位/ml的青霉素和100μg/ml链霉素的RPMI-1640中培养细胞系(均来自PAA)。购自加拿大国立研究委员会的HEK293-6E细胞保持于补充7.5mML-谷氨酰胺(PAA)、0.1%普朗尼克F-68(Invitrogen)和25μg/mlG418(Invitrogen)的FreestyleF17培养基(Invitrogen)中。细胞在5%CO2气氛中在110rpm和37℃下生长于锥形烧瓶(Corning)中。抗体产生和纯化为了获得结合BCMA的抗体,使用标准杂交瘤技术。用不完全弗氏佐剂和30μgC末端融合至谷胱甘肽S-转移酶(GST)的人BCMA的胞外域免疫4只BL/6野生型小鼠6次。在细胞融合后,接下来的筛选时段,J22.9杂交瘤显示分泌抗BCMA抗体。由于杂交瘤的不稳定性,分别扩增杂交瘤J22.9的轻链和重链的可变区并克隆在人κ或IgG1恒定域基因上游。通过用1∶2分别编码轻链和重链的DNA质粒混合物瞬时共转染293-6E细胞产生嵌合J22.9-xi抗体。简言之:将293-6E细胞在无血清的FreestyleF17培养基中重悬为1.7×106个细胞/ml,并使用聚乙烯亚胺以终浓度1μg/ml培养基转染。在转染后第2天,向细胞供给含有1%胰蛋白胨N1(OrganoTechnie)的100%转染体积的FreestyleF17。在第7天,通过离心收获细胞,使过滤(0.45μm)的培养基穿过3.5ml蛋白质A琼脂糖柱(Bio-Rad)。用10ml磷酸盐缓冲盐水(PBS)洗涤柱,并通过添加pH3.5的20mM乙酸钠,150mMNaCl洗脱抗体。直接收集2ml的级分于含有pH7.5的100μl1MHEPES的管中以中和。全长IgG的最终产量接近40mg/l培养基因为杂交瘤J22.9丧失了产生/分泌抗BCMA抗体的能力(图8),因此使用PCR扩增重链和轻链的可变区,然后分别克隆在人恒定IgG1和K轻链基因的5’末端。通过用这两种质粒共转染293-6E细胞,产生嵌合J22.9-xi抗体。本发明的抗体的产生因此是内在地困难的,并且不能通过直接的常规方法实现。与J22.9-xi平行地产生由J22.9-xi重链和随机的嵌合κ轻链组成的同种型对照抗体。通过ELISA和流式细胞术显示这种抗体不能结合于BCMA。使用N-糖苷酶F(PNGaseF)(NEB)酶促地移除在J22.9-xi重链的Asn297处的N-连接的寡糖链。将10mgJ22.9-xi与在500μlPBS(pH7.4)中的15,000单位PNGaseF在37℃下温育36小时,然后将缓冲液更换为无菌PBS。通过酶联免疫吸附测定(ELISA)确定J22.9-xi的结合和阻断能力用10μg/ml人BCMA的胞外域涂覆微量滴定板。用连续稀释的1至1000ng范围的J22.9-xi和同种型对照检测涂覆的BCMA。用辣根过氧化物酶(HRP)缀合的山羊抗人二抗(JacksonImmunoResearch,109-035-098,1∶5,000稀释)检测J22.9-xi或同种型对照抗体与涂覆的BCMA的结合。用1μg/ml人或猕猴BCMA(分别为hBCMA或cyBCMA)的胞外域涂覆微量滴定板。用连续稀释的0.26pM至500nM范围的J22.9-xi、J22.9-H、J22.9-ISY和J22.9-FSY检测涂覆的BCMA。用辣根过氧化物酶(HRP)缀合的山羊抗人二抗(JacksonImmunoResearch,109-035-098,1∶5,000稀释)检测抗体与涂覆的BCMA的结合对于阻断实验,在抗体和洗涤之后应用1mg/ml融合于His标签(Biomol)的人重组BAFF,并使用小鼠抗His标签(AbDSerotec,AD1.1.10,1∶5,000稀释,HRP缀合的)抗体检测。使用BDOptEIA试剂A和B(BDBioscience)对所有ELISA进行显色,并用微孔板分光光度计(BioTek)在450nm和570nm处测量。流式细胞术分析对于细胞表面抗原检测实验,使用自制的抗体(J22.9-xi、J22.9-H、J22.9-ISY、J22.9-FSY和同种型对照)以及商业可获得的小鼠抗His标签(AbDSerotec,AD1.1.10,1∶100稀释,AlexaFluor488缀合的)和山羊抗人IgG1(JacksonImmunoResearch,109-116-098,1∶400,PE缀合的)抗体以及融合于His标签(Biomol)的人重组BAFF。在FACSCalibur或FACSCantoII流式细胞仪(BDBioscience)上进行实验。使用Flowjo软件第7.6版(TreeStarInc.)分析数据。Fab和Fab:BCMA复合体的产生通过与胃蛋白酶温育从全长J22.9-xiIgG产生(Fab)2片段。将J22.9-xi通过PD-10缓冲交换柱进入pH3.5的50mM乙酸钠,并且胃蛋白酶以30μg每毫克J22.9-xi添加。在37℃下温育2.5小时足够完全消化可结晶片段(Fc)区,并且通过使胃蛋白酶在PD-10柱上交换至PBS(pH7.2)失活。在5mM乙二胺四乙酸(EDTA)的存在下通过添加2-巯基乙胺(50mM)实现(Fab)2片段还原为单独的Fab。在37℃下温育90分钟,通过用500μM碘乙酰胺烷基化30分钟,然后缓冲液交换为新鲜PBS来阻断还原的半胱氨酸。使Fab片段与1.5摩尔当量的纯化的BCMA组合,并且通过在Superdex7516/60柱上的体积排阻色谱分离复合体。在4至12%SDS聚丙烯酰胺凝胶上分析级分,将含有Fab和BCMA两者的级分合并并浓缩用于结晶试验。Fab:BCMA复合体的结晶向浓缩的复合体补加0.5摩尔当量的纯BCMA以确保饱和,并使浓缩的复合体经受结晶筛选。由商业筛选(Qiagen)使用Gryphon移液机器人(200nl滴)在96孔坐滴式板中辨别初始的Fab:BCMA结晶条件,并在24孔板中以悬滴(2-3μl)优化。将复合体浓缩至8mg/ml,并在21%PEG3350,0.1MBisTrispH6.5下和5mMCuCl2,在20℃下结晶.3天后晶体以薄盘的集群显现,并且在接近7天内获得其最终大小(0.2-0.3mm)。分离集群并且在具有20%甘油作为保护剂的母液中用液氮急速冷冻单独的盘。在柏林亥姆霍兹中心(HelmoltzZentrumBerlin)的BESSY同步加速器从单个晶体收集完整的衍射数据。使用来自依法利珠单抗(3EO9)作为搜索模型的结构的实验阶段通过分子替换以1.9埃的解析度解出结构。使用程序的ccp4套件进行数据处理,使用Phenix(AdamsPD,等人(2010),ActaCryst.D66:213-221)进行结构提纯,使用Coot.(Emsley等人,Acta结晶学D部分-生物结晶学(ActaCrystallographicaSectionD-BiologicalCcystallography),2010,66:486-501)进行建模和评估。使用PyMOL(PyMOL分子图像系统、版本1.5.0.4LLC)成像。体外细胞毒性测定在本测定中,通过在生物发光读出器中测量剩余的活细胞的发光确定J22.9-xi的细胞毒性作用。简言之:通过重力用160ml洗脱缓冲液(含有5mMNa2-EDTA和2.5[w/v]蔗糖的PBS(pH7.4))反冲洗新鲜获得的人滤器血沉棕黄层(FBC)。通过Ficoll梯度离心从洗脱的细胞中分离单核细胞。从中间相取单核细胞并用洗脱缓冲液洗涤两次。红细胞裂解后,将PBMC再次洗涤,计数并通过用RPMI/10%FCSw/o酚红稀释调整至1×107个细胞/ml。将50μlRPMI中的5×104MM.1S-Luc细胞铺于微量滴定板中。在添加100μlPBMC之前10分钟,将MM.1S-Luc细胞与200μl样品体积的J22.9-xi或同种型对照抗体连续稀释液温育。添加靶细胞、抗体和效应细胞后,将微量滴定板在室温(RT)下离心(300×g)2分钟,并在37℃,5%CO2下储存。对照孔用1%TritonX代替抗体处理以完全裂解。在温育4小时后,将含荧光素(250ng/ml)的25μlPBS施用于每个孔中,并且在生物发光读出器(Tecan)中测量活细胞的生物发光。根据下式计算特定的细胞毒性:100-[值(J22.9-xi)-值(总裂解)]/[值(同种型对照)-值(总裂解)]×100。体内研究使用来自Jackson实验室的NOD.Cg-PrkdcscidIl2rgtm1WjlTg(CSF2)2YgyTg(IL3)1YgyTg(KITLG)3YgyJGckRolyJ小鼠(NSG)和来自德国查尔斯河(苏尔茨费尔德,德国)的CB17.Cg-PrkdcscidLystbg/Crl小鼠。用8-14周龄的小鼠进行实验。所有动物研究根据机构和国家的指导方针,在无特定病原体的条件下进行。在涉及治疗已建立的肿瘤和疾病早期的肿瘤治疗的实验例中,使用CB17.Cg-PrkdcscidLystbg/Crl小鼠。本文提到的两种小鼠品系的表型是非常相似的。这些动物不具有功能性B细胞、T细胞和NK细胞。在CB17.Cg小鼠中观察到稍微更慢的肿瘤生长,表明本发明的治疗性抗体的甚至更有希望的作用。通过在第0天在尾动脉静脉内注射1×107MM.1S-Luc细胞诱导多发性骨髓瘤的异种移植模型。在该模型中,未处理的动物在6周内发展出后肢瘫痪。这样的症状的发生指示实验的结束点。对于有效性研究,每周两次或从第0天开始连续5天腹腔内(i.p.)施用抗体。以每次注射2μg、20μg或200μg的剂量提供J22.9-xi抗体;对于同种型对照抗体,使用200μg/注射.在腹腔内注射150μg荧光素后,使用IVIS谱(CaliperLifeSciences)测量MM.1S-Luc细胞的生物发光。每周一次进行测量。在每个时间点,3只未处理的小鼠也施用荧光素。这些动物的总流量值从每个测量结果中减去或在图中显示.为了处理已建立的肿瘤,在注射MM.1S-Luc细胞之后5天开始抗体治疗。在6周的时间内每周两次施用200μgJ22.9-xi或同种型对照抗体。J22.9-xi的人源化将重链和轻链的可变区序列(小鼠)与来自相应的重链和轻链亚型人序列比对以确定产生完全人源化序列变体需要的残基改变。使用J22.9-xi:hBCMA复合体的晶体结构,首先在电脑中评价每个修饰以辨别能够潜在破坏抗体与BCMA的结合的突变。合成每条链的两种完整的J22.9可变区基因,一条链具有原始小鼠序列,一条链具有完全人源化序列(即含有所有必须的人源化突变),各自具有两个添加的限制性酶切位点将基因分为三个盒。在标记潜在的问题突变之后,产生原始小鼠和完全人源化的基因盒的不同的组合,并且表达它们的相应的IgG,并纯化,用BCMA阳性细胞进行FACS分析以评估结合。使用PCR将标记的问题残基单独地突变以证明它们对与BCMA的亲和性的作用,并且通过SPR后续定量评估最终优化的构建体与人和猕猴BCMA两者的结合。SPR使用补加了0.005%吐温20的磷酸盐缓冲盐水(PBST)在ProteonXPR36上进行SPR。根据制造商的说明使用标准胺化学法将15μg/ml浓度的整个IgG固定于ProteonGLH传感器芯片。对于结合实验,PBST中的人或猕猴BCMA用作移动相。从BCMA的多个浓度(hBCMA为0.4至800nM,以及cynoBCMA为2.7至1μM)平行确定的缔合常数(kon)和解离常数(koff)计算结合亲和度(Kd),假定为单位点结合模型。此外,进一步的实验显示本发明的优选实施方式提供了惊人和预料不到的效果,从而以非显而易见的方式解决了本发明的问题。参考文献Adams,P.D.,Afonine,P.V.,Bunkoczi,G.,Chen,V.B.,Davis,I.W.,Echols,N.,Headd,J.J.,Hung,L.W.,Kapral,G.J.,Grosse-Kunstleve,R.W.等人(2010).PHENIX:用于解析大分子结构的综合性的基于Python的系统(PHENIX:acomprehensivePython-basedsystemformacromolecularstructuresolution).ActaCrystallogrDBiolCrystallogr66,213-221.Al-Lazikani,B.,Lesk,A.M.Chothia,C.(1997).免疫球蛋白典型结构的标准构象(Standardconformationsforthecanonicalstructuresofimmunoglobulins).JMolBiol273,927-948.Anthony,R.M.Ravetch,J.V.(2010).IgGFc聚糖的新作用:唾酸IgGFc的抗炎活性(AnovelrolefortheIgGFcglycan:theanti-inflammatoryactivityofsialylatedIgGFcs).JClinImmunol30Suppl1,S9-14.Chanetal.(2010)NatRevImmunol10:301-316Chavez-Galan,L.,Arenas-DelAngel,M.C.,Zenteno,E.,Chavez,R.Lascurain,R.(2009).细胞毒性淋巴细胞诱导的细胞死亡机制(Celldeathmechanismsinducedbycytotoxiclymphocytes).CellMolImmunol6,15-25.Chiu,A.,Xu,W.,He,B.,Dillon,S.R.,Gross,J.A.,Sievers,E.,Qiao,X.,Santini,P.,Hyjek,E.,Lee,J.W.等人(2007).霍奇金淋巴瘤细胞表达TACI和BCMA受体并响应于BAFF和APRIL产生存活和增殖信号(HodgkinlymphomacellsexpressTACIandBCMAreceptorsandgeneratesurvivalandproliferationsignalsinresponsetoBAFFandAPRIL).Blood109,729-739.Gordon,N.C.,Pan,B.,Hymowitz,S.G.,Yin,J.,Kelley,R.F.,Cochran,A.G.,Yan,M.,Dixit,V.M.,Fairbrother,W.J.Starovasnik,M.A.(2003).BAFF/BLyS受体3包括编码高度集中的配体结合位点的最小TNF受体样组件(BAFF/BLySreceptor3comprisesaminimalTNFreceptor-likemodulethatencodesahighlyfocusedligand-bindingsite).Biochemistry42,5977-5983.Greenstein,S.,Krett,N.L.,Kurosawa,Y.,Ma,C.,Chauhan,D.,Hideshima,T.和erson,K.C.Rosen,S.T.(2003).MM.1人多发性骨髓瘤(MM)细胞系的表征:一种阐明对类固醇敏感和耐受的MM细胞的特征、行为和信号的模型系统(CharacterizationoftheMM.1humanmultiplemyeloma(MM)celllines:amodelsystemtoelucidatethecharacteristics,behavior,andsignalingofsteroid-sensitiveand-resistantMMcells).ExpHematol31,271-282.Jacobi,A.M.,Huang,W.,Wang,T.,Freimuth,W.,Sanz,I.,Furie,R.,Mackay,M.,Aranow,C.,Diamond,B.Davidson,A.(2010).在系统性红斑狼疮中长期贝利单抗治疗对B细胞的作用:二期,双盲法,安慰剂对照法,剂量范围的延伸研究(Effectoflong-termbelimumabtreatmentonBcellsinsystemiclupuserythematosus:extensionofaphaseII,double-blind,placebo-controlled,dose-rangingstudy).ArthritisRheum62,201-210.Kapoor,P.,Ramakrishnan,V.和Rajkumar,S.V.(2012).在多发性骨髓瘤中的硼替佐米联合治疗(Bortezomibcombinationtherapyinmultiplemyeloma).SeminHematol49,228-242.Keyser,F.D.(2011).对患有类风湿性关节炎的患者的生物治疗的选择:感染角度(ChoiceofBiologicTherapyforPatientswithRheumatoidArthritis:TheInfectionPerspective).CurrRheumatolRev7,77-87.Laskowski,R.A.(2009).PDBsum新事物(PDBsumnewthings).NucleicAcidsRes37,D355-359.Mackay,F.,Schneider,P.,Rennert,P.Browning,J.(2003).BAFF和APRIL:对B细胞存活的指导(BAFFANDAPRIL:atutorialonBcellsurvival).AnnuRevImmunol21,231-264.Nimmerjahn,F.Ravetch,J.V.(2008).Fcγ受体作为免疫应答的调节剂(Fcgammareceptorsasregulatorsofimmuneresponses).NatRevImmunol8,34-47.Novak,A.J.,Darce,J.R.,Arendt,B.K.,Harder,B.,Henderson,K.,Kindsvogel,W.,Gross,J.A.,Greipp,P.R.Jelinek,D.F.(2004).在多发性骨髓瘤中BCMA,TACI和BAFF-R的表达:生长和存活的机制(ExpressionofBCMA,TACI,andBAFF-Rinmultiplemyeloma:amechanismforgrowthandsurvival).Blood103,689-694.Queen等人,1989;WO90/07861Patel,D.R.,Wallweber,H.J.,Yin,J.,Shriver,S.K.,Marsters,S.A.,Gordon,N.C.,Starovasnik,M.A.Kelley,R.F.(2004).工程化APRIL特异性B细胞成熟抗原(EngineeringanAPRIL-specificBcellmaturationantigen).JBiolChem279,16727-16735.Presta,L.G.(2008).治疗性抗体的分子工程化和设计(Molecularengineeringanddesignoftherapeuticantibodies).CurrOpinImmunol20,460-470.Raab,M.S.,Podar,K.,Breitkreutz,I.,Richardson,P.G.Anderson,K.C.(2009).多发性骨髓瘤(Multiplemyeloma).Lancet374,324-339.Richardson等人(2003)NewEnglJMed348:2609-2617.Ryan等人(MolecularCancerTherapeutics,20076(11),3009)Roopenian,D.C.Akilesh,S.(2007).FcRn:新生Fc受体到达时代(FcRn:theneonatalFcreceptorcomesofage).NatRevImmunol7,715-725.Suzuki,K.(2013).多发性骨髓瘤的目前治疗策略(Currenttherapeuticstrategyformultiplemyeloma).JpnJClinOncol43,116-124.Thorpe等人,1982,Immunol.Rev.62:119-58Wang,S.Y.和Weiner,G.(2008).抗体治疗癌症中的补体和细胞的细胞毒性(Complementandcellularcytotoxicityinantibodytherapyofcancer).ExpertOpinBiolTher8,759-768.Woyke等人(2001)Antimicrob.AgentsandChemother.45(12):3580-3584.当前第1页1 2 3
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1