嵌合蛋白的制作方法
发明领域
本发明涉及在过继性细胞疗法(act)中有用的嵌合蛋白。所述嵌合蛋白可以起自杀基因作用,使得表达所述嵌合蛋白的细胞能够被删除。本发明还提供编码此类嵌合蛋白的核酸,包含此类核酸的细胞及其治疗用途。
发明背景
过继性细胞疗法
过继免疫疗法是已经建立且不断发展的治疗方法。在同种异体造血干细胞移植(hsct)的背景中,经常给予供体淋巴细胞输注(dli)以治疗血液恶性肿瘤的复发。肿瘤浸润性淋巴细胞(til)在治疗转移性黑色素瘤中是有效的。对t细胞的遗传工程化大大增加了t细胞疗法的范围和效力:t细胞受体转移允许靶向细胞内癌症抗原,而嵌合抗原受体(car)允许靶向表面癌症或谱系特异性抗原。已经使用两种方法观察了临床反应,并且正在进行许多进一步的试验。
过继性免疫疗法后可发生急性不良事件。移植物抗宿主病(gvhd)是dli的常见且严重的并发症。工程化t细胞的施用也导致毒性。例如,在针对黑色素瘤抗原的天然t细胞受体转移研究中已经报道了中靶脱肿瘤毒性(on-targetoff-tumourtoxicity);再定向(re-directed)至肾细胞癌抗原碳酸酐酶ix(caix)的t细胞产生出乎意料的肝毒性。cd19car疗法后报道了免疫激活综合征。最后,载体诱导的插入诱变导致淋巴增殖性病症的理论风险。这些毒性的发生率和严重性是不可预知的。此外,与其不良事件通常以治疗剂的半衰期减退的治疗性蛋白或小分子相反,t细胞植入并且复制,这潜在地导致升高且暴发性毒性。
自杀基因
自杀基因是遗传编码的机制,其允许在面对不可接受的毒性时选择性地破坏过继性转移的细胞,例如t细胞。已经在临床研究中测试了两种自杀基因:单纯疱疹病毒胸苷激酶(hsv-tk)和诱导型胱天蛋白酶9(icasp9)。
单纯疱疹病毒i来源的胸苷激酶(hsv-tk)基因已经用作供体t细胞输注的体内自杀开关,用于治疗造血干细胞移植后的复发性恶性肿瘤和埃巴病毒(ebv)淋巴增殖。然而,对引起移植物抗宿主病的t细胞的破坏是不完全的,并且使用更昔洛韦(或类似物)作为前药来激活hsv-tk,排除了更昔洛韦作为巨细胞病毒感染的抗病毒药物的施用。此外,即使在免疫抑制的人类免疫缺陷病毒和骨髓移植患者中,hsv-tk导向的免疫应答也导致hsv-tk转导的细胞的消除,损害了输注的t细胞的持续性并因此降低其功效。
在原始icasp9分子中利用了胱天蛋白酶9背后的活化机制。胱天蛋白酶被活化所需要的所有,是克服胱天蛋白酶同二聚化的能量障碍。同二聚体经历构象变化,且二聚体对之一的蛋白水解结构域变得有活性。在生理上,这通过胱天蛋白酶9的card结构域结合到apaf-1而发生。在icasp9中,apaf-1结构域被修饰的fkbp12替代,所述修饰的fkbp12已经突变以选择性地结合二聚化化学诱导剂(cid)。cid的存在导致同二聚化和活化。icasp9基于与人fk506结合蛋白(fkbp)融合的经修饰的人胱天蛋白酶9(straathofetal.,(2005)blood105:4247-4254)。其使得能够在称为ap1903的小分子cid的存在下进行条件性二聚化。ap1903是一种实验性药物,并认为是生物惰性的,因为它不与野生型fkbp12相互作用。然而,使用该试剂的临床经验限于非常少数的患者(distasi,a.etal.(2011)n.engl.j.med.365,1673–1683;andiuliucci,j.d.etal.(2001)j.clin.pharmacol.41,870–879)。ap1903也是一种相对较大的极性分子,且不太可能穿过血脑屏障。
在一种替代方法中,使用复杂策略可以通过小分子活化执行剂(executioner)胱天蛋白酶,该策略涉及将烟草蚀刻病毒(tev)蛋白水解位点导入胱天蛋白酶3或6或7和与分开的tev蛋白酶(其在雷帕霉素的存在下重组)共表达(morganetal(2014)methodsenzymol.544:179-213)。这由于一些原因对于临床上有用的自杀开关来说是不满意的策略:首先需要三种分开的蛋白质,这是高度复杂的:分别是修饰的胱天蛋白酶和拆分的tev蛋白酶的两个组分;其次,tev组分是异种的(xenogeneic),可能具有免疫原性;最后,该策略仅活化蛋白酶敏感的胱天蛋白酶分子,其是顶端(apical)胱天蛋白酶下游的且相比顶端(apical)胱天蛋白酶较不敏感。
已经描述了基于fas的cid活化的自杀基因(amaraetal(1999)hum.genether.10,2651–2655)。这也取决于该cid用于激活,并且由于它不直接活化细胞凋亡级联,因此逃避(通过fas抗性)是可能的。
基于替换实验性cid需求的标准药物的同二聚化体系将是有吸引力的替代方案。然而,没有同二聚化小分子药物可用。
已经提出了其它自杀基因,例如全长cd20,当在t细胞上表达时可以使t细胞对通过治疗性抗cd20抗体利妥昔单抗(rituximab)的裂解易感(introna,m.etal.(2000)hum.genether.11,611–620)。关于抗体识别这一主题还描述了其它自杀基因,例如:rqr8使t细胞对cd20易感,但比全长cd20分子更紧凑(philip,b.etal.(2014)blooddoi:10.1182/blood-2014-01-545020);egfr的截短形式(huegfrt)使得细胞对通过抗egfrmab的裂解易感(wang,x.etal.(2011)blood118,1255–1263);以及在细胞表面上表达的myc表位标签使得细胞对使用抗myc抗体的裂解易感(kiebacketal(2008)proc.natl.acad.sci.u.s.a.105,623–628)。这些抗体依赖性方法的主要限制是它们依赖于起作用的高局部浓度的治疗性抗体的生物可用度。已知例如裂解性抗体对于大块疾病(bulkydisease)并不特别有效,并且基于抗体的自杀基因的限制是在没有达到高抗体浓度的细胞驻留者(resident)将会逃逸。此外,在某些情况下:例如由cart细胞诱导的严重的巨噬细胞活化综合征或细胞因子风暴(cytokinestorm);由单克隆抗体诱导的额外的免疫活化可能对自杀基因活化尝试治疗的临床情况是有害的。
因此,需要与上述缺点不相关的替代性自杀基因。
附图简述
图1-显示rapcasp9的不同方法的草图。(a)双重构建体,其中两个分子单独表达。每个分子具有分别与fkbp12或frb融合的casp9的催化结构域。(b)单一构建体,其中fkbp12和frb直接融合在一起,然后通过柔性接头与casp9的催化结构域融合。在此取向上,自身异二聚化应当是不可能的。(c)单一构建体,其中胱天蛋白酶9的催化结构域侧翼有frb和fkbp12。这里,自身异二聚化可以发生,因此预计这种重复(iteration)不会很好地发挥功能。(d)双重构建体,其中半胱氨酸蛋白酶9的催化结构域融合到fkbp12融合和分开的小蛋白质,其是共表达的两拷贝frb的融合物。
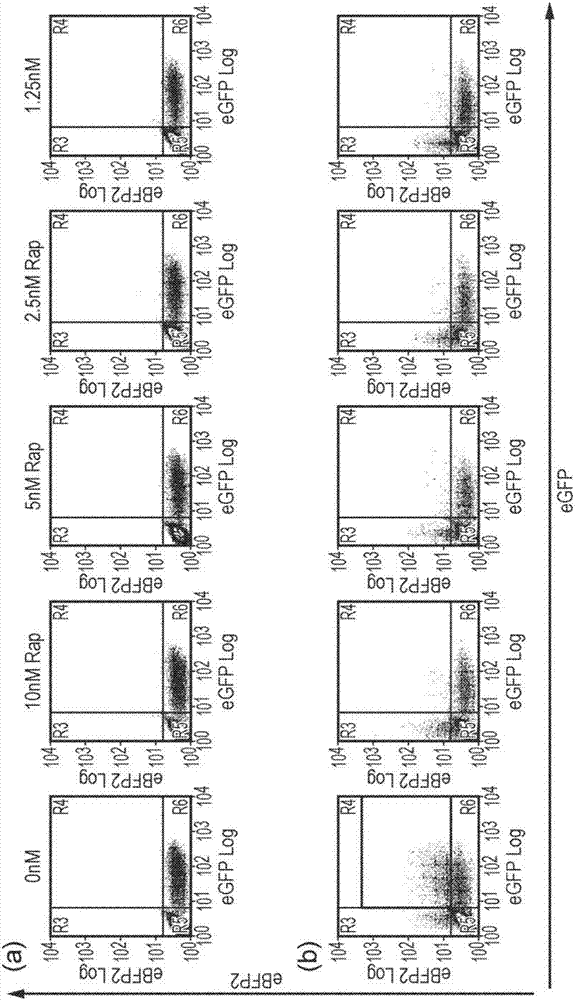
图2-证明了可以用异二聚化剂(heterodimerizer)活化胱天蛋白酶9。t细胞用单独egfp转导(图2a),或用fkbp12-dcasp9(共表达egfp)和frb-dcasp9(共表达ebfp2)共转导(图2b)。有意地仅部分转导t细胞,使得非转导的t细胞作为内部对照起作用。然后将t细胞暴露于降低浓度的雷帕霉素。48小时后,细胞用膜联蛋白-v和7aad染色,并通过流式细胞术分析,观察表达荧光蛋白的活细胞比例。即使在最低浓度的雷帕霉素的存在下也非常有效地删除表达egfp和ebfp2两者的t细胞。
图3-rapcasp9变体的功能。(a)使用单独的egfp转导t细胞;(b)使用分别与egfp和ebfp2共表达的fkbp12-casp9和frb-casp9双重转导t细胞;(c)使用frb-fkbp12-casp9转导t细胞以及(d)使用frb-casp9-fkbp12转导t细胞和(e)fbp12-casp9-2a-frb-frbw转导。仅转导细胞的部分,阴性细胞作为内部阴性对照起作用。将t细胞暴露于2.5nm雷帕霉素48小时。然后用膜联蛋白-v和7aad染色t细胞,并通过流式细胞术分析。如通过膜联蛋白-v和7aad染色测定,在活细胞上显示了egfp对ebfp2。
图4-雷帕霉素和雷帕霉素类似物(rapalogs)。a)雷帕霉素;b)c-20-methyllyrlrapamycin(marap);c)c16(s)-丁基磺胺雷帕霉素(c16(s)-butylsulfonamidorapamycin)(c16-bs-rap);d)c16-(s)-3-甲基吲哚雷帕霉素(c16-(s)-3-mehylindolerapamycin)(c16-irap);和e)c16-(s)-7-甲基吲哚雷帕霉素(c16-(s)-7-methylindolerapamycin)(ap21976/c16-airap)。
图5-实施例3中测试的构建体的总结。
图6-实施例3的门控(gating)策略总结。
图7、8和9-研究显示用各种浓度的雷帕霉素孵育后用图5中所示构建体转染的jurkat细胞的杀伤。
图10-总结图7、8和9所示的facs数据的图。
图11-比较雷帕霉素对替西罗莫司(temsirolimus)存在下的jurkat细胞杀伤的图。
发明概述
本发明人已经开发了新的自杀基因,其在二聚化化学诱导剂(cid)例如雷帕霉素或雷帕霉素类似物的存在下二聚化。
雷帕霉素和雷帕霉素类似物通过在mtor的frb结构域和fkbp12之间形成界面诱导异二聚化。该结合导致fkbp12阻断对mtor活性位点的接近,从而抑制其功能。尽管mtor是一种非常大的蛋白,但对于与雷帕霉素相互作用所要求的mtor的精确小区段是已知的并可以使用。
本发明人已经显示了可以使用由雷帕霉素介导的异二聚化来诱导胱天蛋白酶的同二聚化。具体而言,他们已经令人惊讶地显示了可以创建多结构域分子,其包括(i)mtor的frb结构域;(ii)fkbp12;和(iii)胱天蛋白酶,使用该分子的一个拷贝的frb结构域和该分子的另一个拷贝的fkb12结构域之间的异二聚化以引起胱天蛋白酶结构域的同二聚化。
因此,在本发明的第一个方面的第一个实施方案中,本发明提供具有以下式的嵌合蛋白:
ht1-ht2-casp
其中
casp是胱天蛋白酶结构域;
ht1是第一异二聚化结构域;和
ht2是第二异二聚化结构域
且其中,在二聚化化学诱导剂(cid)的存在下,嵌合蛋白的相同对相互作用使得来自一个嵌合蛋白的ht1与来自另一个嵌合蛋白的ht2异二聚化,导致两个胱天蛋白酶结构域的同二聚化。
构造为使得ht1不以任何显著程度与相同嵌合蛋白中的ht2异二聚化。
胱天蛋白酶结构域可以包含选自下组的引发剂(initiator)胱天蛋白酶:胱天蛋白酶-8、胱天蛋白酶-9和胱天蛋白酶-10,或选自胱天蛋白酶-3和胱天蛋白酶-7的执行剂(executioner)胱天蛋白酶。
在本发明的第一个方面的此第一个实施方案的多结构域蛋白中,一个异二聚化结构域可以包含fk506结合蛋白(fkbp),而另一个异二聚化结构域可以包含mtor的frb结构域。
对于该异二聚化结构域组合,适合的cid是雷帕霉素或雷帕霉素类似物。
在本发明的第一个方面的第二个实施方案中,提供了包含胱天蛋白酶和异二聚化结构域的嵌合蛋白,所述异二聚化结构域包含fk506结合蛋白(fkbp),和包含胱天蛋白酶结构域和异二聚化结构域的嵌合蛋白,所述异二聚化结构域包含mtor的frb结构域。
在本发明的该方面的第三个实施方案中,提供以下两种蛋白:
ht1-casp和ht2-ht2
其中ht1-casp是包含胱天蛋白酶结构域(casp)和第一异二聚化结构域(ht1)的嵌合蛋白;且ht2-ht2是包含两个或更多个第二异二聚化结构域(ht2)的界面蛋白;并且
其中在二聚化化学诱导剂(cid)的存在下,嵌合蛋白ht1-casp9的对相互作用使得来自每个嵌合蛋白的ht1与来自界面(interfacing)蛋白的ht2结构域异二聚化,导致两个胱天蛋白酶结构域的同二聚化。
在本发明的该方面的第四个实施方案中,提供了具有以下式的嵌合蛋白:
ht1-casp-ht2
其中
casp是胱天蛋白酶结构域;
ht1是第一异二聚化结构域;和
ht2是第二异二聚化结构域
且其中,在二聚化化学诱导剂(cid)的存在下,嵌合蛋白的相同对相互作用使得来自一个嵌合蛋白的ht1与来自另一个嵌合蛋白的ht2异二聚化,导致两个胱天蛋白酶结构域的同二聚化。
关于本发明的第一个方面的该第四个实施方案,在一个异二聚化结构域包含fk506结合蛋白(fkbp)而另一个二聚化结构域包含mtor的frb结构域,且cid为雷帕霉素或其衍生物的情况下,则可以使用小于5nm的浓度,例如1-3nm或约1nm以引起两个胱天蛋白酶结构域的同二聚化。
嵌合蛋白可以包含与fkbp12融合的胱天蛋白酶结构域,且界面蛋白可以是两个或更多个frb结构域的融合。这两个或更多个frb结构域作为界面起作用,使两个fkbp12-casp结构域在一起。
在第二个方面,本发明提供核酸序列,其编码根据本发明的第一个方面的嵌合蛋白。
核酸序列可以以核酸构建体的形式,其包含多个核酸序列。例如,构建体可以包含一个或多个根据本发明的第二个方面的核酸序列和编码t细胞受体(tcr)或嵌合抗原受体(car)的核酸序列。
核酸构建体可以包含:
i)编码嵌合蛋白的第一核酸,所述嵌合蛋白包含胱天蛋白酶结构域和异二聚化结构域,所述异二聚化结构域包含fk506结合蛋白(fkbp);和
ii)编码嵌合蛋白的第二核酸,所述嵌合蛋白包含胱天蛋白酶结构域和异二聚化结构域,所述异二聚化结构域包含mtor的frb结构域。
还提供了具有以下结构的核酸构建体:
ht1-casp-coexpr-ht2-ht2
其中
casp是编码胱天蛋白酶结构域的核酸序列;
ht1是编码第一异二聚化结构域的核酸序列;
ht2是编码第二异二聚化结构域的核酸序列;和
coexpr是允许ht1-casp和ht2-ht2共表达的核酸序列,
其中所述核酸构建体的表达导致嵌合蛋白ht1-casp和界面蛋白ht2-ht2的产生,且其中在二聚化化学诱导剂(cid)的存在下,所述嵌合蛋白ht1-casp9的对相互作用,使得来自每个嵌合蛋白的ht1与来自界面蛋白的ht2结构域异二聚化,导致两个胱天蛋白酶结构域的同二聚化。
ht1可以包含fk506结合蛋白(fkbp)且ht2可以包含mtor的frb结构域。
核酸构建体还可以包含编码t细胞受体(tcr)或嵌合抗原受体(car)的核酸序列。
在第三个方面,本发明提提供载体,其包含根据本发明的第二个方面的核酸序列或核酸构建体。
载体还可以包含感兴趣的核苷酸,例如编码嵌合抗原受体或t细胞受体的核苷酸序列,使得当载体用于转导靶细胞时,靶细胞共表达根据本发明的第一个方面的嵌合蛋白和嵌合抗原受体或t细胞受体。
在本发明的第四个方面,提供细胞,其表达根据本发明的第一个方面的嵌合蛋白。
细胞可以包含:
i)第一嵌合蛋白,所述第一嵌合蛋白包含胱天蛋白酶结构域和异二聚化结构域,所述异二聚化结构域包含fk506结合蛋白(fkbp);和
ii)第二嵌合蛋白,所述第二嵌合蛋白包含胱天蛋白酶结构域和异二聚化结构域,所述异二聚化结构域包含mtor的frb结构域。
还提供表达两种蛋白的细胞:
ht1-casp和ht2-ht2
其中ht1-casp是包含胱天蛋白酶结构域(casp)和第一异二聚化结构域(ht1)的嵌合蛋白;且ht2-ht2是包含两个第二异二聚化结构域(ht2)的界面蛋白;且
其中在二聚化化学诱导剂(cid)的存在下,所述嵌合蛋白ht1-casp9的对相互作用,使得来自每个嵌合蛋白的ht1与来自界面蛋白的ht2结构域异二聚化,导致两个胱天蛋白酶结构域的同二聚化。
细胞可以包含根据本发明的第二个方面的核酸序列或构建体。
细胞可以例如是造血干细胞,淋巴细胞或t细胞。
还提供了制备根据本发明的第四个方面的细胞的方法,其包括使用根据本发明的第三个方面的载体转导或转染细胞的步骤。
还提供删除根据本发明的第四个方面的细胞的方法,其包括将细胞暴露于二聚化化学诱导剂(cid)的步骤。
cid可以是雷帕霉素或雷帕霉素类似物。
还提供用于预防或治疗受试者中的疾病的方法,其包括向所述受试者施用根据本发明的第四个方面的细胞的步骤。
该方法可以包括以下步骤:
(i)使用根据根据本发明的第二个方面的载体转导或转染从受试者分离的细胞样品,和
(ii)将转导的/转染的细胞施用至患者。
方法可以用于治疗癌症。
还提供用于预防和/或治疗受试者中病理性免疫反应的方法,所述病理性免疫反应由将根据本发明的第四个方面的细胞施用至受试者引起,所述方法包括向所述受试者施用雷帕霉素或雷帕霉素类似物的步骤。
病理性免疫反应可以选自下组:移植物抗宿主病;中靶脱肿瘤毒性(on-target,off-tumourtoxicity);免疫活化综合征;和淋巴组织增殖性病症。
用于治疗或预防受试者中的疾病的方法可以包括以下步骤:
(i)将根据本发明的第四个方面的细胞施用至所述受试者;
(ii)监控所述受试者的病理性免疫反应的形成;和
(iii)如果所述受试者显示正在形成或已经形成病理性免疫反应的体征,则向所述受试者施用雷帕霉素或雷帕霉素类似物。
还提供了根据本发明的第四个方面的细胞,其用于造血干细胞移植、淋巴细胞输注或过继细胞转移。
还提供雷帕霉素或雷帕霉素类似物,其用于预防或治疗由向受试者施用根据本发明的第四个方面的细胞引起的病理性免疫反应。
因此,本发明提供了自杀基因,其面对不可接受的毒性时允许选择性破坏过继输注的细胞,且其由雷帕霉素和/或其类似物活化。
雷帕霉素是具有充分了解的性质,极好的生物利用度和分布体积、且广泛可用的标准药物。雷帕霉素也不加重所治疗的病症,事实上,由于它是一种免疫抑制剂,其可能对不需要的毒性以及其自杀基因功能具有有益的影响。
发明详述
嵌合蛋白
本发明涉及嵌合蛋白,其作为自杀基因发挥作用。通过施用二聚化化学诱导剂(cid)例如雷帕霉素或雷帕霉素类似物可以在体内或体外删除表达该嵌合蛋白的细胞。
嵌合蛋白可以具有以下式:
ht1-ht2-casp
其中
casp是胱天蛋白酶结构域;
ht1是第一异二聚化结构域;和
ht2是第二异二聚化结构域。
嵌合蛋白可以具有以下式:
ht1-ht2-l-casp
其中casp、ht1和ht2如上文所定义,且l是任选的接头。
构造应当使得ht1不与相同嵌合蛋白分子中的ht2显著异二聚化,但当两个嵌合蛋白在二聚化化学诱导剂(cid)的存在下在一起时,来自一个嵌合蛋白的ht1与来自另一个嵌合蛋白的ht2异二聚化,导致两个胱天蛋白酶的同二聚化。
构造使得ht1不以任何显著的程度与相同嵌合蛋白中的ht2异二聚化。例如,在表达根据本发明的第一方面的该实施方案的嵌合蛋白的细胞中,cid的存在应当导致比相同嵌合蛋白内的异二聚化更大比例的两个嵌合蛋白之间的二聚化。在细胞或细胞群体中或在溶液中相同分子内的异二聚化的嵌合蛋白的量在cid的存在下可以是与分开的嵌合蛋白分子异二聚化的嵌合蛋白的量的小于50%、40%、30%、20%、10%、5%或1%。
嵌合蛋白可以包含如seqidno.1所示的序列:
seqidno.1(frb-fkbp12-l3-dcasp9)
在以上序列中“fkbp12”是指fkbp12的序列;“dcasp9”是指casp9的催化结构域;“l1”是一重复接头(onerepeatlinker);“fmd-2a”是口蹄疫2a样肽erav;“frb”是mtor的frb结构域;“l3”是二重复接头(tworepeatlinker);且“frbw”是密码子摆动的frb。
在第二个实施方案中,本发明提供“两分子”自杀基因系统,其中cid是雷帕霉素或雷帕霉素类似物。
因此,本发明还提供i)嵌合蛋白,其包含胱天蛋白酶结构域和异二聚化结构域,所述异二聚化结构域包含fk506结合蛋白(fkbp12);和ii)嵌合蛋白,其包含胱天蛋白酶结构域和异二聚化结构域,所述异二聚化结构域包含mtor的frb结构域。
当细胞例如t细胞表达这些嵌合蛋白两者时,雷帕霉素或雷帕霉素类似物的存在导致i)的包含fkbp的结构域的与ii)的包含frb的结构域异二聚化,从而导致i)和ii)的胱天蛋白酶的同二聚化。
在本发明的该实施方案中,嵌合蛋白可以包含如seqidno.2或3所示的序列。
seqidno.2(fkbp12-dcasp9)
seqidno.3(frb-dcasp9)
在第三个实施方案中,本发明提供了备选的“两分子”方法,其具有比第二个实施方案更小的足迹(footprint)。这里,ht1与胱天蛋白酶融合,以及第二分子包含共表达的ht2-ht2融合物。在cid的存在下,ht2-ht2将两个个ht1-casp分子带到一起。实际上,这可以通过共表达fkbp12-casp9与frb-frb并使用雷帕霉素活化来实施。方便地,这些组分可以与口蹄疫2a样肽共表达。编码第二ht2(例如frb)的序列可以进行密码子摆动以防止重组。
seqidno.4(fkbp12-dcasp9-2a-frb-frbw)
在上述序列中,“fkbp12”指代fkbp12;“dcasp9”是casp9的催化结构域;“l1”是一重复接头;“fmd-2a”是口蹄疫2a样肽erav;“frb”是mtor的frb结构域;“l2”是二重复接头;且“frbw”是密码子摆动的frb。
胱天蛋白酶
胱天蛋白酶,或半胱氨酸-天冬氨酸蛋白酶或半胱氨酸依赖性天冬氨酸定向蛋白酶是半胱氨酸蛋白酶家族,其在凋亡中发挥重要功能。
已经在人类中鉴定了12种胱天蛋白酶。存在两种类型的凋亡胱天蛋白酶:引发剂胱天蛋白酶和执行剂胱天蛋白酶。引发剂胱天蛋白酶,如胱天蛋白酶-2、胱天蛋白酶-8、胱天蛋白酶-9和胱天蛋白酶-10,切割效应胱天蛋白酶的非活性的原形式,从而活化它们。执行剂胱天蛋白酶,例如胱天蛋白酶-3、胱天蛋白酶-6和胱天蛋白酶-7,然后切割细胞内的其它蛋白底物,以触发凋亡过程。
本发明的第一个方面的嵌合蛋白的胱天蛋白酶结构域可以包含选自胱天蛋白酶-2;胱天蛋白酶-8、胱天蛋白酶-9和胱天蛋白酶-10的引发剂胱天蛋白酶;或者选自胱天蛋白酶-3、胱天蛋白酶-6和胱天蛋白酶-7的执行剂胱天蛋白酶。
特别地,本发明的第一个方面的嵌合蛋白的胱天蛋白酶结构域可以包含胱天蛋白酶-9。胱天蛋白酶-9是关键的引发剂胱天蛋白酶,因此其活化是对于凋亡诱导非常敏感的触发子(trigger)。此外,同二聚化是活化所需要的全部,而不是同二聚化和蛋白水解切割。
全长胱天蛋白酶-9具有如seqidno.5所示的序列。
seqidno.5(胱天蛋白酶-9)
1mdeadrrllrrcrlrlveelqvdqlwdallsselfrphmiediqragsgsrrdqarqlii
61dletrgsqalplfiscledtgqdmlasflrtnrqaaklskptlenltpvvlrpeirkpev
121lrpetprpvdigsggfgdvgaleslrgnadlayilsmepcghcliinnvnfcresglrtr
181tgsnidceklrrrfssphfmvevkgdltakkmvlallelaqqdhgaldccvvvilshgcq
241ashlqfpgavygtdgcpvsvekivnifngtscpslggkpklffiqacggeqkdhgfevas
301tspedespgsnpepdatpfqeglrtfdqldaisslptpsdifvsystfpgfvswrdpksg
361swyvetlddifeqwahsedlqslllrvanavsvkgiykqmpgcfnflrkklffkts
可以截短胱天蛋白酶-9,例如去除胱天蛋白酶募集结构域。截短的胱天蛋白酶-9如seqidno.6所示。
seqidno.6(截短的胱天蛋白酶-9,缺少card结构域)
gfgdvgaleslrgnadlayilsmepcghcliinnvnfcresglrtrtgsnidceklrrrfsslhfmvevkgdltakkmvlallelaqqdhgaldccvvvilshgcqashlqfpgavygtdgcpvsvekivnifngtscpslggkpklffiqacggeqkdhgfevastspedespgsnpepdatpfqeglrtfdqldaisslptpsdifvsystfpgfvswrdpksgswyvetlddifeqwahsedlqslllrvanavsvkgiykqmpgcfnflrkklffkts
本发明的第一个方面的嵌合蛋白可以包含seqidno.5或seqidno.6或其保留了二聚化从而触发凋亡的能力的片段或变体。
变体胱天蛋白酶可以与seqidno.5或6具有至少80%、85%、90%、95%、98%或99%序列同一性。
可以通过程序例如blast(其在http://blast.ncbi.nlm.nih.gov可免费获得)容易地确定两条肽序列之间的百分比同一性。
在体内,蛋白酶胱天蛋白酶9是称为凋亡体(apoptosome)的多组分通路的中心参与者,所述凋亡体控制胚胎发生期间的细胞删除,以及触发细胞死亡以及致死细胞损害,例如离子辐射或化疗药物的生理应答。胱天蛋白酶9的功能是通过有限的蛋白水解生成活性形式的胱天蛋白酶3和7,并从而将凋亡信号传输至执行阶段。然而,胱天蛋白酶9在其紧密相关物中的不寻常之处在于大亚基和小亚基之间的蛋白水解不将潜伏的酶原(zymogen)转化为催化形式。事实上,对于活化所必需的是同二聚化。
异二聚化结构域
大环内酯类雷帕霉素和fk506通过诱导细胞蛋白的异二聚化发挥作用。每个药物以高亲和力结合fkbp12蛋白,创建药物-蛋白复合物,其随后分别结合并失活mtor/frap和钙依赖磷酸酶(calcineurin)。已经定义了mtor的fkbp-雷帕霉素结合(frb)结构域并作为分离的89个氨基酸的蛋白部分应用,其可以与感兴趣的蛋白融合。然后雷帕霉素可以诱导frb融合物与fkbp12或与fkbp12融合的蛋白的接近。
在本发明的语境中,异二聚化结构域之一(ht1或ht2)可以是或包含frb或其变体,且另一个异二聚化结构域(ht2或ht1)可以是或包含fkbp12或其变体。
雷帕霉素具有理想的二聚化剂的几个特性:当结合至fkbp12时,其对于frb具有高亲和力(kd<1nm),且对于mtor的frb结构域具有高特异性。雷帕霉素是一种有效的治疗性免疫抑制剂,其在哺乳动物中具有有利的药代动力学和药效学概况。具有不同的药代动力学和动态特性的雷帕霉素的药理学类似物例如依维莫司(everolimus)、替西罗莫司(temsirolimus)和德福莫林(deforolimus)(benjaminetal,naturereviews,drugdiscovery,2011)也可以根据临床设置使用。
为了防止雷帕霉素结合并失活内源性mtor,可以修饰接触frb的雷帕霉素的表面。对frb结构域的补偿突变以形成容纳“突起的”雷帕霉素的表面(burface)仅恢复与frb突变体而不与内源性mtor蛋白的二聚化相互作用。
bayle等人(chembio;2006;13;99-107)描述了多种雷帕霉素类似物,或“rapalog”,以及其相应的经修饰的frb结合结构域。例如,bayle等人(2006)描述了以下雷帕霉素类似物:c-20-methyllyrlrapamycin(marap);c16(s)-丁基磺胺雷帕霉素(c16-bs-rap)和c16-(s)-7-甲基吲哚雷帕霉素(ap21976/c16-airap),如图3中所示,与每种各自的相应互补结合结构域组合。其它雷帕霉素/雷帕霉素类似物包括西罗莫司(sirolimus)和他克莫司(tacrolimus)。
嵌合蛋白的异二聚化结构域可以是或包含如seqidno:7至seqidno:11所示的序列之一,或其变体。
seqidno7-fkbp12结构域
mgvqvetispgdgrtfpkrgqtcvvhytgmledgkkfdssrdrnkpfkfmlgkqevirgweegvaqmsvgqrakltispdyaygatghpgiipphatlvfdvellkle
seqidno8-mtor的野生型frb区段
masrilwhemwhegleeasrlyfgernvkgmfevleplhammergpqtlketsfnqaygrdlmeaqewcrkymksgnvkdltqawdlyyhvfrriskles
seqidno9-2098处具有t至l取代的frb,其允许结合ap21967
masrilwhemwhegleeasrlyfgernvkgmfevleplhammergpqtlketsfnqaygrdlmeaqewcrkymksgnvkdllqawdlyyhvfrriskles
seqidno10-mtor的frb区段,其在完全mtor的2098处具有t至h取代和残基2101处的f取代为w,其以相对于野生型降低的亲和力结合雷帕霉素
masrilwhemwhegleeasrlyfgernvkgmfevleplhammergpqtlketsfnqaygrdlmeaqewcrkymksgnvkdlhqafdlyyhvfrriskles
seqidno11-mtor的frb区段,其在完全mtor的残基2095处具有k至p取代,其以降低的亲和力结合雷帕霉素
masrilwhemwhegleeasrlyfgernvkgmfevleplhammergpqtlketsfnqaygrdlmeaqewcrkymksgnvpdltqawdlyyhvfrriskles
变体序列可以与seqidno.7至11具有至少80%、85%、90%、95%、98%或99%序列同一性,条件是该序列提供了有效的二聚化系统。也就是说,条件是该序列促进两个嵌合蛋白的充分共定位以允许两个胱天蛋白酶结构域的同二聚化。
示于seqidno.8的“野生型”frb结构域包含人mtor的氨基酸2025-2114。使用人类mtor的氨基酸编号系统,本发明的嵌合蛋白的frb序列可以包含在以下一个或多个位置处的氨基酸取代:2095、2098、2101。
本发明的嵌合蛋白中使用的变体frb可以包含在位置2095、2098和2101处的以下氨基酸之一:
2095:k、p、t或a
2098:t、l、h或f
2101:w或f
bayle等人(如上文)描述了以下frb变体,其根据位置2095、2098和2101处的氨基酸进行注释(参见表1):ktw、plf、klw、plw、tlw、alw、ptf、atf、ttf、klf、plf、tlf、alf、ktf、khf、kff、klf。这些变体能够以不同程度结合雷帕霉素和雷帕霉素类似物,如bayle等人的表1和图5a中所示。本发明的嵌合蛋白可以包含这些frb变体之一。
接头
可以包括接头以空间上分开胱天蛋白酶结构域和异二聚化结构域。
在本发明的第一个方面的第一个实施方案中,嵌合蛋白包含两个异二聚化结构域,其保持在以下构造中,使得它们在cid的存在下在单个分子中不能彼此异二聚化,但一个分子上的ht1能够与具有相同异二聚化结构域的另一个嵌合分子上的ht2异二聚化(图1b)。在其中ht1和ht2在胱天蛋白酶结构域侧翼的设计(ht1-casp-ht2)中,活化不如其中ht1和ht2连接到一起的设计,表明防止来自单个分子的ht1和ht2对单个cid的非产生性结合的重要性。
在该实施方案中,接头(l1)应当提供足够的柔性,使得催化结构域能够同二聚化,但非如此大量的柔性,使得不能克服同二聚化的能量障碍(图1)。例如,接头长度可以少于15个,少于10个或5-15个或5-10个氨基酸。
在本发明的第一个方面的第二个实施方案中,嵌合蛋白包含单个异二聚化结构域,其在cid的存在下能够与第二嵌合蛋白上的互补异二聚化结构域异二聚化。
在备选的构造中,可以在具有长接头(l2)的单个分子上提供两个异二聚化结构域,提供具有以下式的构建体:
ht1-casp1-l2-ht2-casp2
ht和casp结构域在接头的每一侧可以以任一顺序。
在该实施方案中,接头l2可以赋予足够的柔性,使得第一异二聚化结构域能够与第二异二聚化结构域异二聚化;并使得对应“第一嵌合蛋白”的分子的部分中的胱天蛋白酶结构域能够与对应“第二嵌合蛋白”的分子的部分中的胱天蛋白酶结构域同二聚化。
在本发明的第一个方面的第三个实施方案中,casp与单个异二聚化结构域融合,但第二分子是另一个异二聚化结构域的两个或多个拷贝的融合。可以共表达两种分子。在这种情况下,第二分子起界面作用,在cid的存在下将两个或多个casp结构域带到一起。在这种情况下,两个或更多个拷贝的异二聚化结构域必须以如下方式融合,使得允许casp9结构域的接近足以活化它们。
界面蛋白可以是多聚体的,包含多于两个ht2结构域。例如,可以在单个界面蛋白中组合多个ht2结构域,其使用多聚化接头例如卷曲螺旋结构域。
在该实施方案中,界面蛋白可以具有式ht2-l2-ht2,或ht2-l2,其中l2是螺旋卷曲结构域。
螺旋卷曲是一种结构基序,其中2至7个α螺旋缠绕在一起,类似绳索的股。螺旋卷曲结构域的结构是本领域公知的,例如如lupas&gruber(advancesinproteinchemistry;2007;70;37-38)所述。
螺旋卷曲通常含有疏水(h)和带电荷(c)的氨基酸残基的重复的模式,hxxhcxc,称为七元重复(heptadrepeat)。七元重复中的位置通常标记为abcdefg,其中a和d是疏水位置,通常被异亮氨酸、亮氨酸或缬氨酸占据。将具有该重复模式的序列折叠为α螺旋二级结构导致疏水残基被呈现为“条纹(stripe)”,其以左手的方式轻柔盘绕螺旋,形成两亲性结构。对于两个此类螺旋将它们自身排列在细胞质中最有利的方式是在将疏水性链彼此缠绕,夹在亲水性氨基酸之间。因此,疏水性表面的埋藏为寡聚化提供了热力学驱动力。螺旋卷曲界面中的包装非常紧密,在a和d残基的侧链之间具有几乎完全范德华力接触。
含有螺旋卷曲结构域的蛋白的实例包括但不限于驱动蛋白马达蛋白(motorprotein),丁型肝炎δ抗原,古细菌盒c/dsrnp核心蛋白,软骨低聚基质蛋白(comp),甘露糖结合蛋白a,富含丝氨酸的卷曲螺旋蛋白1(coiled-coilserine-richprotein1),多肽释放因子2,snap-25,snare,lac阻遏物或载脂蛋白e。
二聚化化学诱导剂(cid)
二聚化化学诱导剂(cid)可以是任何分子,其诱导具有相同的ht1和ht2结构域上的单独的嵌合分子上的ht1和ht2之间的异二聚化。
cid可以是雷帕霉素或雷帕霉素类似物(“rapalog”),其具有相对于雷帕霉素改进的或不同的药代动力学或药效学特定,但具有相同的广泛作用机制。cid可以是具有对互补fkbp12或frb的工程化特异性的改变的雷帕霉素,例如如图4中所示。bayle等人(2006,如上文)描述了在c16和/或c20处功能化的多种雷帕霉素类似物。
第一类中的此类雷帕霉素类似物的例子包括西罗莫司(sirolimus)、依维莫司(everolimus)、替西罗莫司(temsirolimus)和德福莫林(deforolimus)。第二类中的雷帕霉素类似物的例子包括c-20-methyllyrlrapamycin(marap);c16(s)-丁基磺胺雷帕霉素(c16-bs-rap);c16-(s)-3-甲基吲哚雷帕霉素(c16-irap);和c16-(s)-7-甲基吲哚雷帕霉素(ap21976/c16-airap)。
在cid的存在下胱天蛋白酶结构域的同二聚化可以导致胱天蛋白酶活化,其比在cid不存在下发生的胱天蛋白酶活化高2、5、10、50、100、1000或10000倍。
雷帕霉素是一种强力的免疫抑制剂。雷帕霉素的类似物(rapalogues)用于日常临床用途。现代雷帕霉素类似物具有极好的生物可用性和分布体积。尽管它们是强力的免疫抑制剂,短剂量(以活化自杀基因)应当具有最小限度的副作用。此外,与单抗的施用不同,雷帕霉素和类似物的药理学效果在自杀基因要求活化的临床场景,如脱肿瘤毒性(off-tumortoxicity)或免疫超活化综合征中可以完全是有利的。
核酸序列
本发明的第二个方面提供核酸序列,其编码根据本发明的嵌合蛋白。
如本文所使用的,术语“多核苷酸”、“核苷酸”和“核酸”意图彼此同义。
本领域的技术人员将理解,由于遗传密码的简并性,许多不同的多核苷酸和核酸可以编码相同的多肽。此外,应当理解理解技术人员可以使用常规技术,进行核苷酸取代以反映在其中表达多肽的任何具体宿主微生物的密码子选择,所述核苷酸取代不影响本文所述的多核苷酸编码的多肽序列。
根据本发明的第二方面的核酸可以包含dna或rna。它们可以是单链的或双链的。它们也可以是在它们中包括合成的或修饰的核苷酸的多核苷酸。对寡核苷酸的许多不同类型的修饰在本领域中是已知的。这些包括甲基膦酸酯和硫代磷酸酯主链,在分子的3’和/或5’末端添加吖啶或多聚赖氨酸链。为了如本文所述的用途的目的,应当理解可以通过本领域可用的任何方法修饰多核苷酸。为了提高感兴趣的多核苷酸的体内活性或寿命可以进行此类修饰。
关于核苷酸序列,术语“变体”、“同源物”或“衍生物”包括在对序列的一个(或多个)核酸的任何取代、变异、修饰、置换、缺失或添加。
在本发明的该方面的第一个实施方案中,提供核酸序列,其编码具有以下式的嵌合蛋白:
ht1-ht2-l-casp
ht1是第一异二聚化结构域;和
ht2是第二异二聚化结构域。
l是任选的接头;
casp是胱天蛋白酶结构域;
核酸序列可以编码如seqidno.1所示的嵌合蛋白序列或其变体。
例如,核苷酸序列可以包含如seqidno.12所示的序列
seqidno.12(frb-fkbp12-l3-casp9)
atggcttctagaatcctctggcatgagatgtggcatgaaggcctggaagaggcatctcgtttgtactttggggaaaggaacgtgaaaggcatgtttgaggtgctggagcccttgcatgctatgatggaacggggcccccagactctgaaggaaacatcctttaatcaggcctatggtcgagatttaatggaggcccaagagtggtgcaggaagtacatgaaatcagggaatgtcaaggacctcctccaagcctgggacctctattatcatgtgttccgacgaatctcaaagctcgagtatagcggcggcggcagcctggagggcgtgcaggtggagaccatcagcccaggcgacggcagaaccttccccaagagaggccagacctgcgtggtgcactataccggcatgctggaggacggcaagaagttcgacagcagccgcgaccgcaataagcccttcaagttcatgctgggcaagcaggaggtgatcagaggctgggaggagggcgtggcccagatgagcgtgggccagagagccaagctgaccatcagccccgactacgcctatggcgccaccggccaccccggcatcatcccaccccacgccaccctggtgtttgatgtggagctgctgaagctggagtccggcggaggcgggtctggaggaggcggcagcggcggcggcgggtcaggcgtggatggcttcggcgacgtgggagccctggagagcctgagaggcaacgccgatctggcctacatcctgagcatggagccctgtggccactgcctgatcatcaacaacgtgaacttctgccgggagagcggcctgcggacccggaccggcagcaacatcgactgcgagaagctgaggaggcgcttctcctccctgcactttatggtggaggtgaaaggcgatctgactgccaagaaaatggtgctggccctgctggagctggcccagcaggaccacggagccctggattgctgtgtggtggtgatcctgtcccacggctgccaggccagccacctgcagttccccggagccgtgtacggcaccgacggctgtcccgtgtccgtggagaagatcgtgaacatcttcaacggcacctcctgcccctccctgggcggcaagcccaagctgttctttatccaggcctgtggcggcgagcagaaggaccacggctttgaggtggccagcacctcccccgaggacgagagcccaggcagcaaccccgagcccgacgccacccccttccaggagggcctgcgcaccttcgaccagctggacgccatcagcagcctgcccacccccagcgacatcttcgtgagctacagcacctttcccggcttcgtgagctggcgcgatcccaagtccggctcttggtatgtggagaccctggacgacatctttgagcagtgggctcatagcgaggacctgcagagcctgctgctgcgcgtggccaatgccgtgagcgtgaagggcatctacaagcagatgccaggctgcttcaacttcctgcggaagaagctgttcttcaagaccagcgcctcctga
在本发明的该方面的第二个实施方案中,提供编码具有以下式的嵌合蛋白的核酸序列:ht1-l-casp
其中
ht1是异二聚化结构域。
l是任选的接头;和
casp是胱天蛋白酶结构域;
核酸序列可以编码如seqidno.2或3所示的嵌合蛋白序列或其变体。
例如,核苷酸序列可以包含如seqidno.13或14所示的序列
seqidno.13(fkbp12-dcasp9)
atgctggagggcgtgcaggtggagaccatcagcccaggcgacggcagaaccttccccaagagaggccagacctgcgtggtgcactataccggcatgctggaggacggcaagaagttcgacagcagccgcgaccgcaataagcccttcaagttcatgctgggcaagcaggaggtgatcagaggctgggaggagggcgtggcccagatgagcgtgggccagagagccaagctgaccatcagccccgactacgcctatggcgccaccggccaccccggcatcatcccaccccacgccaccctggtgtttgatgtggagctgctgaagctggagtccggaggcggctccggcgtggatggcttcggcgacgtgggagccctggagagcctgagaggcaacgccgatctggcctacatcctgagcatggagccctgtggccactgcctgatcatcaacaacgtgaacttctgccgggagagcggcctgcggacccggaccggcagcaacatcgactgcgagaagctgaggaggcgcttctcctccctgcactttatggtggaggtgaaaggcgatctgactgccaagaaaatggtgctggccctgctggagctggcccagcaggaccacggagccctggattgctgtgtggtggtgatcctgtcccacggctgccaggccagccacctgcagttccccggagccgtgtacggcaccgacggctgtcccgtgtccgtggagaagatcgtgaacatcttcaacggcacctcctgcccctccctgggcggcaagcccaagctgttctttatccaggcctgtggcggcgagcagaaggaccacggctttgaggtggccagcacctcccccgaggacgagagcccaggcagcaaccccgagcccgacgccacccccttccaggagggcctgcgcaccttcgaccagctggacgccatcagcagcctgcccacccccagcgacatcttcgtgagctacagcacctttcccggcttcgtgagctggcgcgatcccaagtccggctcttggtatgtggagaccctggacgacatctttgagcagtgggctcatagcgaggacctgcagagcctgctgctgcgcgtggccaatgccgtgagcgtgaagggcatctacaagcagatgccaggctgcttcaacttcctgcggaagaagctgttcttcaagaccagcgcctcctga
seqidno.14.(frb-dcasp9)
atggcttctagaatcctctggcatgagatgtggcatgaaggcctggaagaggcatctcgtttgtactttggggaaaggaacgtgaaaggcatgtttgaggtgctggagcccttgcatgctatgatggaacggggcccccagactctgaaggaaacatcctttaatcaggcctatggtcgagatttaatggaggcccaagagtggtgcaggaagtacatgaaatcagggaatgtcaaggacctcctccaagcctgggacctctattatcatgtgttccgacgaatctcaaagctcgagtatagcggcggcggcagcggcgtggatggcttcggcgacgtgggagccctggagagcctgagaggcaacgccgatctggcctacatcctgagcatggagccctgtggccactgcctgatcatcaacaacgtgaacttctgccgggagagcggcctgcggacccggaccggcagcaacatcgactgcgagaagctgaggaggcgcttctcctccctgcactttatggtggaggtgaaaggcgatctgactgccaagaaaatggtgctggccctgctggagctggcccagcaggaccacggagccctggattgctgtgtggtggtgatcctgtcccacggctgccaggccagccacctgcagttccccggagccgtgtacggcaccgacggctgtcccgtgtccgtggagaagatcgtgaacatcttcaacggcacctcctgcccctccctgggcggcaagcccaagctgttctttatccaggcctgtggcggcgagcagaaggaccacggctttgaggtggccagcacctcccccgaggacgagagcccaggcagcaaccccgagcccgacgccacccccttccaggagggcctgcgcaccttcgaccagctggacgccatcagcagcctgcccacccccagcgacatcttcgtgagctacagcacctttcccggcttcgtgagctggcgcgatcccaagtccggctcttggtatgtggagaccctggacgacatctttgagcagtgggctcatagcgaggacctgcagagcctgctgctgcgcgtggccaatgccgtgagcgtgaagggcatctacaagcagatgccaggctgcttcaacttcctgcggaagaagctgttcttcaagaccagcgcctcctga
在第二个实施方案中,可以以构建体的形式提供核酸序列,所述构建体编码两种嵌合蛋白。
构建体可以编码具有以下式的多肽:
ht1-l2-casp-coexpr-ht2-l2-casp
其中
ht1是第一异二聚化结构域;
l1和l2是任选的接头,其可以是相同的或不同的;
coexpr是使得能够共表达以下两种蛋白的序列:ht1-l1-casp和ht2-l2-casp;
ht2是第二异二聚化结构域;和
casp是胱天蛋白酶结构域。
在存在编码相同或相似的序列,例如两个胱天蛋白酶结构域的核酸序列的情况下,序列之一可以进行密码子摆动以避免同源重组。
在第三个实施方案中,提供核酸序列,其编码具有以下式的序列:
ht1-casp-coexpr-ht2-ht2
其中
casp是胱天蛋白酶结构域;
ht1是第一异二聚化结构域;
coexpr是使得蛋白ht1-casp和ht2-ht2能够共表达的序列,例如切割位点;和
ht2是第二异二聚化结构域,其在二聚化化学诱导剂(cid)的存在下与ht1异二聚化。
在编码第二蛋白ht2-ht2的序列中,编码ht2的序列之一可以进行密码子摆动以避免同源重组。
根据第三个实施方案的核酸构建体可以具有如seqidno.15所示的序列。
seqidno.15(fkbp12-casp9-2a-frb-frbw)
atgctggagggcgtgcaggtggagaccatcagcccaggcgacggcagaaccttccccaagagaggccagacctgcgtggtgcactataccggcatgctggaggacggcaagaagttcgacagcagccgcgaccgcaataagcccttcaagttcatgctgggcaagcaggaggtgatcagaggctgggaggagggcgtggcccagatgagcgtgggccagagagccaagctgaccatcagccccgactacgcctatggcgccaccggccaccccggcatcatcccaccccacgccaccctggtgtttgatgtggagctgctgaagctggagtccggaggcggctccggcgtggatggcttcggcgacgtgggagccctggagagcctgagaggcaacgccgatctggcctacatcctgagcatggagccctgtggccactgcctgatcatcaacaacgtgaacttctgccgggagagcggcctgcggacccggaccggcagcaacatcgactgcgagaagctgaggaggcgcttctcctccctgcactttatggtggaggtgaaaggcgatctgactgccaagaaaatggtgctggccctgctggagctggcccagcaggaccacggagccctggattgctgtgtggtggtgatcctgtcccacggctgccaggccagccacctgcagttccccggagccgtgtacggcaccgacggctgtcccgtgtccgtggagaagatcgtgaacatcttcaacggcacctcctgcccctccctgggcggcaagcccaagctgttctttatccaggcctgtggcggcgagcagaaggaccacggctttgaggtggccagcacctcccccgaggacgagagcccaggcagcaaccccgagcccgacgccacccccttccaggagggcctgcgcaccttcgaccagctggacgccatcagcagcctgcccacccccagcgacatcttcgtgagctacagcacctttcccggcttcgtgagctggcgcgatcccaagtccggctcttggtatgtggagaccctggacgacatctttgagcagtgggctcatagcgaggacctgcagagcctgctgctgcgcgtggccaatgccgtgagcgtgaagggcatctacaagcagatgccaggctgcttcaacttcctgcggaagaagctgttcttcaagaccagcgcctcccagtgcaccaattatgctttgcttaagctggcaggcgatgtggaatcaaacccgggtcctggggtacaggtggagaccatctctcctggcgacgggagaacatttcctaaaaggggccaaacatgcgtggttcactataccggtatgctggaggatggcaaaaaagtagactccagccgggatagaaacaaaccctttaagttcatgctgggtaagcaggaagttatacggggctgggaagagggagtagctcagatgtctgtgggccagagggccaagctgaccatctcaccggactacgcctacggcgctaccggccaccctggcattataccaccccatgcaactctcgtcttcgatgttgagttgctcaaactggaatcaggcggaggcgggtctggaggaggcggcagcatgctggagggcgtgcaggtggagaccatcagcccaggcgacggcagaaccttccccaagagaggccagacctgcgtggtgcactataccggcatgctggaggacggcaagaagttcgacagcagccgcgaccgcaataagcccttcaagttcatgctgggcaagcaggaggtgatcagaggctgggaggagggcgtggcccagatgagcgtgggccagagagccaagctgaccatcagccccgactacgcctatggcgccaccggccaccccggcatcatcccaccccacgccaccctggtgtttgatgtggagctgctgaagctggagtcctga
具有高度相似性的核酸序列,例如胱天蛋白酶序列或frb序列可以进行密码子摆动以避免重组。
核酸构建体
本发明还提供核酸构建体,其包含:
i)编码嵌合蛋白的第一核酸序列,所述嵌合蛋白包含胱天蛋白酶结构域和异二聚化结构域,所述异二聚化结构域包含fk506结合蛋白(fkbp);和
ii)编码嵌合蛋白的第二核酸序列,所述嵌合蛋白包含胱天蛋白酶结构域和异二聚化结构域,所述异二聚化结构域包含mtor的frb结构域。
本发明还提供核酸构建体,其包含编码一种或多种嵌合蛋白的核酸序列和进一步感兴趣的核酸(noi)。该noi可以例如编码t细胞受体(tcr)或嵌合抗原受体(car)。
核酸序列可以通过允许两种或更多种核酸序列的共表达的序列连接。例如,构建体可以包含内部启动子,内部核糖体进入序列(ires)序列或编码切割位点的序列。切割位点可以是自切割的,使得当多肽产生时,其立即切割为离散的蛋白而不需要任何外部切割活性。
多种自切割位点是已知的,包括口蹄疫病毒(fmdv)2a自切割肽,其具有如seqidno.16或17所示的序列。
seqidno.16
raegrgslltcgdveenpgp
或者
seqidno17
qctnyallklagdvesnpgp
共表达序列可以是内部核糖体进入序列(ires)。共表达序列可以是内部启动子。
t细胞受体(tcr)
t细胞受体或tcr是t细胞表面上发现的分子,其负责识别结合主要组织相容性复合物(mhc)分子的抗原。tcr和抗原之间的结合是相对低亲和力的并且是简并的:许多tcr识别相同的抗原且许多抗原被相同的tcr识别。
tcr由两条不同的蛋白链组成,即其是异二聚体。在95%的t细胞中,其由alpha(α)和beta(β)组成,而在5%的t细胞中,这由gamma和delta(γ/δ)链组成。该比率在个体发育期间和疾病状态中改变。
当tcr与抗原性肽和mhc(肽/mhc)接合时,t淋巴细胞通过由相关酶、共受体、特化的衔接头分子,以及活化或释放的转录因子介导的一系列生物化学事件活化。
本发明的核酸构建体或载体可以包含编码tcrα链、tcrβ链、tcrγ链或tcrδ链的核酸序列。其可以例如包含编码tcrα链的核酸序列和编码tcrβ链的核酸序列;或编码tcrγ链的核酸序列或编码tcrδ链的核酸序列。两个核酸序列可以通过使得能够共表达两条tcr链的序列连接,例如内部启动子、ires序列或切割位点例如自切割位点。
嵌合抗原受体(car)
感兴趣的核酸序列(noi)可以编码嵌合抗原受体(car)。
经典的car是嵌合i型跨膜蛋白,其将细胞外抗原识别结构域(结合剂)与细胞内信号传导结构域(胞内域(endodomain))连接。结合剂通常是衍生于单克隆抗体(mab)的单链可变片段(scfv),但其可以基于包含抗原识别位点的其它形式例如配体。为了将结合剂从膜分开并允许其适当定向,间隔区结构域可以是必要的。使用的常见间隔区结构域是igg1的fc。取决于抗原,更紧密的间隔区可以足够,例如来自cd8α的茎部和甚至仅单独的igg1铰链。跨膜结构域将蛋白锚定在细胞膜中并将间隔区连接至胞内域,所述胞内域可以包含细胞内信号传导结构域或与细胞内信号传导结构域结合。
早期的car设计具有来源于fcεr1或cd3ζ的γ链的细胞内部分的胞内域。因此,这些第一代受体传输免疫信号1,其足够触发t细胞杀伤关联靶细胞,但不能完全活化t细胞以增殖或存活。为了克服这一限制,构建了复合信号传导域:将t细胞共刺激分子的细胞内部分融合至cd3ζ的细胞内部分,产生了第二代受体,其在抗原识别后能够同时传输活化和共刺激信号。最通常使用的共刺激结构域是cd28的共刺激结构域。这提供了最强力的共刺激信号-即免疫信号2,其触发t细胞增殖。已经描述了一些受体,其包括tnf受体家族胞内域,例如紧密相关的ox40和41bb,其传输存活信号。现在已经描述了甚至更强力的第三代car,其具有能够传输活化,增殖和存活信号的胞内信号传导域。
可以使用例如逆转录病毒载体将编码car的核酸转移至t细胞。以这种方式,可以生成大量抗原特异性t细胞用于用于过继细胞转移。当car结合靶抗原时,这导致活化信号传输至上面表达有它的t细胞。因此,car将t细胞的特异性和细胞毒性引导至表达靶向抗原的细胞。
载体
在第三个方面,本发明提供载体,其包含本发明的核酸序列或核酸构建体。
本发明还提供载体,或载体的试剂盒,其包含一种或多种本发明的核酸序列或核酸构建体和任选地一种或多种另外的感兴趣的核酸序列(noi)。此类载体可以用于将核酸序列或核酸构建体引入宿主细胞,使得其表达一种或多种根据本发明的第一个方面的嵌合蛋白,和任选地一种或多种其它的感兴趣的蛋白(poi)。该试剂盒还可以包含cid。
载体可以是例如质粒或病毒载体,例如逆转录病毒载体或慢病毒载体,或基于转座子的载体或合成的mrna。
载体可以能够转染或转导t细胞。
noi可以例如编码嵌合抗原受体或t细胞受体,使得当载体用于转导靶细胞时,靶细胞共表达嵌合蛋白和嵌合抗原受体或t细胞受体。
细胞
本发明还涉及包含根据本发明的第一个方面的嵌合蛋白的细胞。
细胞可以表达根据本发明的第一个方面的第一个实施方案的具有两个异二聚化结构域的嵌合蛋白。
细胞可以表达根据本发明的第一个方面的第二个实施方案的两种嵌合蛋白,一种表达胱天蛋白酶结构域和异二聚化结构域,所述异二聚化结构域包含fk506结合蛋白(fkbp);和一种表达胱天蛋白酶结构域和异二聚化结构域,所述异二聚化结构域包含mtor的frb,
还提供表达以下两种蛋白的细胞:
ht1-casp和ht2-ht2
其中ht1-casp是包含胱天蛋白酶结构域(casp)和第一异二聚化结构域(ht1)的嵌合蛋白;且ht2-ht2是包含两个第二异二聚化结构域(ht2)的界面蛋白
使得在二聚化化学诱导剂(cid)的存在下,嵌合蛋白ht1-casp9的对相互作用使得来自每个嵌合蛋白的ht1与来自界面蛋白的ht2结构域异二聚化,导致两个胱天蛋白酶结构域的同二聚化(参见图1d)。
细胞可以是例如免疫细胞如t细胞或自然杀伤(nk)细胞。
细胞可以是干细胞例如造血干细胞。
t细胞或t淋巴细胞是淋巴细胞的类型,其在细胞介导的免疫中发挥中心作用。通过细胞表面上的t细胞受体(tcr)的存在,它们可以与其它淋巴细胞例如b细胞和自然杀伤细胞(nk细胞)区分。
辅助性t辅助细胞(th细胞)在免疫学过程中辅助其他白细胞,包括b细胞成熟为浆细胞和记忆b细胞,以及活化细胞毒性t细胞和巨噬细胞。th细胞在其表面上表达cd4。th细胞当通过抗原呈递细胞(apc)表面上的mhcii类分子给它们呈递肽抗原时被激活。这些细胞可以分化成几种亚型之一,包括th1,th2,th3,th17,th9或tfh,其分泌不同的细胞因子以促进不同类型的免疫应答。
细胞毒性t细胞(tc细胞或ctl)破坏病毒感染的细胞和肿瘤细胞,并且也涉及移植物排斥。ctl在其表面表达cd8。这些细胞通过与存在于所有有核细胞表面上的mhci类结合的抗原结合而识别其靶标。通过调节性t细胞分泌的il-10,腺苷和其他分子,可以将cd8+细胞失活至无变应性状态,这防止自身免疫疾病,如实验性自身免疫性脑脊髓炎。
记忆t细胞是抗原特异性t细胞的子集,该细胞在感染已经消退之后长期持续。它们在再次暴露于其关联抗原后迅速扩增为大量效应t细胞,从而为免疫系统提供了针对过去感染的“记忆”。记忆t细胞包括三种亚型:中央记忆t细胞(tcm细胞)和两种类型的效应记忆t细胞(tem细胞和temra细胞)。记忆细胞可以是cd4+或cd8+。记忆t细胞通常表达细胞表面蛋白cd45ro。
以前称为抑制物t细胞的调节性t细胞(treg细胞)对于维持免疫耐受性至关重要。它们的主要作用是关闭t细胞介导的免疫朝向免疫反应的结束,以及抑制逃避胸腺中阴性选择过程的自身反应性t细胞。
已经描述了两种主要类型的cd4+treg细胞-天然发生的treg细胞和适应性treg细胞。
天然存在的treg细胞(也称为cd4+cd25+foxp3+treg细胞)出现在胸腺中,并与已经与发育中的t细胞和已经用tslp激活的髓样(cd11c+)和浆细胞样(cd123+)树突状细胞之间的相互作用相关。天然存在的treg细胞可以通过称为foxp3的细胞内分子的存在与其它t细胞区分开来。foxp3基因的突变可以阻止调节性t细胞发育,导致致命的自身免疫性疾病ipex。
适应性treg细胞(也称为tr1细胞或th3细胞)可以在正常免疫应答期间起源。
自然杀伤细胞(或nk细胞)是形成先天免疫系统的一部分的一类细胞溶解性细胞。nk细胞以不依赖于mhc的方式对来自病毒感染的细胞的先天信号提供快速应答。
nk细胞(属于先天性淋巴样细胞群)被定义为大颗粒淋巴细胞(lgl),并构成从产生b和t淋巴细胞的共同淋巴样祖细胞分化的第三种细胞。已知nk细胞在骨髓,淋巴结,脾,扁桃体和胸腺中分化和成熟,在那里它们进入循环。
干细胞是未分化的细胞,其可以分化为特化的细胞。在哺乳动物中,存在两大类型的干细胞:胚胎干细胞,其是从胚泡的内细胞团分离,和成体干细胞,其在各种组织中存在。在成年生物体中,干细胞和祖细胞充当机体的修复系统,补充成体组织。在发育中的胚胎中,干细胞可以分化成所有的特化细胞-外胚层,内胚层和中胚层(见诱导的多能干细胞)-但还维持再生器官如血液,皮肤或肠组织的正常周转。
在人类中存在三种已知的可及的自体成体干细胞来源:
1.骨髓,其需要通过收获来提取,即钻入骨中。
2.脂肪组织,其需要通过抽脂术提取。
3.血液,这需要通过单采血液成分(apheresis)提取,其中血液从供体采集并通过机器,该机器提取干细胞并将血液的其它部分返回供体。
成体干细胞经常在药物疗法中使用,例如在骨髓移植中。现在干细胞可以人工生长并转化(分化)为具有与多种组织例如肌肉或神经的细胞一致的特征的特化细胞类型。通过体细胞核移植或去分化生成的自体胚胎干细胞和胚胎细胞系也可以用于生成特化的细胞类型用于细胞疗法。
造血干细胞(hsc)是血细胞,其产生所有其它的血细胞,并来源于中胚层。它们定位于在大多数骨骼的核心中含有的红骨髓。
它们产生髓样细胞(单核细胞和巨噬细胞,嗜中性粒细胞、嗜碱性粒细胞、嗜酸性粒细胞、红细胞、巨核细胞/血小板、树突状细胞),和淋巴样谱系(t细胞、b细胞、nk细胞)。造血组织含有具有长期和短期再生能力的细胞,和定型(committed)多能、寡能和单能祖细胞。
hsc是异质性群体。存在三类干细胞,通过血液中它们的淋巴样对髓样后代的比率(l/m)区分。髓样偏好的(my-bi)hsc具有低的l/m比率(0和3之间),而淋巴样偏好的(ly-bi)的hsc显示大的比率(>10)。第三类由平衡的(bala)hsc组成,其l/m比率在3和10之间。仅髓样偏好的和平衡的hsc具有持久的自我更新特性。
本发明的表达嵌合蛋白的细胞可以是上述任何细胞类型。
表达一种或多种根据本发明的第一个方面的嵌合蛋白的t或nk细胞可以从患者自己的外周血(第一方)离体产生,或在来自供体外周血的造血干细胞移植物(第二方)的设置中产生,或自不相关供体的外周血(第三方)产生。
或者,表达一种或多种根据本发明的第一个方面的嵌合蛋白的t或nk细胞可以衍生自诱导型祖细胞或胚胎祖细胞离体分化为t细胞。或者,可以使用保留其溶解功能并且可以起治疗剂作用的永生化t细胞系。
在所有这些实施方案中,通过诸如使用病毒载体转导或使用dna或rna转染等手段,引入编码嵌合蛋白或每种嵌合蛋白和任选的noi的dna或rna来生成表达嵌合蛋白的细胞。
本发明的细胞可以是来自受试者的离体t或nk细胞。t或nk细胞可以来自外周血单核细胞(pbmc)样品。t或nk细胞可以在用编码一种或多种根据本发明的第一个方面的嵌合蛋白的核酸转导之前进行活化和/或扩增,例如通过使用抗cd3单克隆抗体处理。
本发明的t或nk细胞可以如下制备:
(i)从受试者或上文列出的其它来源分离含t或nk细胞的样品;和
(ii)使用编码一种或多种根据本发明的第二个方面的核酸序列转导或转染t或nk细胞。
本发明还提供试剂盒,其包含t或nk细胞和cid,所述t或nk细胞包含一种或多种根据本发明的第一个发明的嵌合蛋白。
药物组合物
本发明还涉及药物组合物,其含有多种根据本发明的第四个方面的细胞。药物组合物可以另外包含药学上可接受的载体,稀释剂或赋形剂。药物组合物可以任选地包含一种或多种另外的药学活性多肽和/或化合物。此类配制剂可以例如为适合用于静脉输注的形式。
方法
本发明还提供用于制备根据本发明的第四个方面的细胞的方法,其包括使用根据本发明的第三个方面的载体转导或转染细胞的步骤。
载体可以是例如逆转录病毒载体或慢病毒载体。
本发明还提供用于删除根据本发明的第四个方面的细胞的方法,其包括将细胞暴露于cid例如雷帕霉素或雷帕霉素类似物的步骤。细胞可以在体内或离体暴露于cid。在cid诱导的胱天蛋白酶结构域的同二聚化后,可以通过由胱天蛋白酶活化诱导的凋亡引起细胞的删除。
可以以药物组合物的形式施用cid。药物组合物可以另外包含药学上可接受的载体,稀释剂或赋形剂。药物组合物可以任选地包含一种或多种另外的药学活性多肽和/或化合物。此类配制剂可以为例如适合用于静脉输注的形式。
本发明还提供用于预防和/或治疗受试者中由对受试者施用根据本发明的第四个方面的细胞引起的病理性免疫反应的方法,其包括向所述受试者施用cid例如雷帕霉素或雷帕霉素类似物的步骤。
病理性免疫反应可以选自下组:移植物抗宿主病;中靶脱肿瘤毒性(on-target,off-tumourtoxicity);免疫活化综合征;和淋巴组织增殖性病症。
本发明还提供用于治疗或预防受试者中的疾病的方法,其包括向所述受试者施用根据本发明的第四个方面的细胞的步骤。该细胞可以以如上文定义的药物组合物的形式。
所述方法可以包括以下步骤:
(i)使用根据本发明的第三个方面的载体转导或转染从受试者分离的细胞样品,和
(ii)将转导的/转染的细胞施用至患者。
用于治疗疾病的方法涉及本发明的细胞的治疗性用途。本文中,可以将细胞施用至具有已经存在的疾病或状况的受试者,以减轻,降低或改善至少一种与该疾病相关的症状和/或减慢、降低或阻断疾病的进展。
用于预防疾病的方法涉及本发明的免疫细胞的预防性用途。本文中,可以将此类细胞施用至尚未感染疾病和/或没有显示疾病的任何症状的受试者,以预防或削弱疾病的原因或降低或防止与疾病相关的至少一种症状的形成。该受试者可以具有形成疾病的素因或认为具有形成疾病的风险。
本发明提供的用于治疗疾病的方法可以涉及监控疾病的进展和监控任何毒性活性和调整施用至受试者的cid的剂量,以提供可接受水平的疾病进展和毒性活性。
监控疾病的进展意味着随时间评估与疾病相关的症状以确定其是否在降低/改善或增加/恶化。
毒性活性指在将本发明的细胞施用至受试者后由其引起的副作用。毒性活性可以包括例如免疫毒性、胆汁毒性和呼吸窘迫综合征。
特别地,本发明提供用于治疗受试者中的疾病的方法,其包括以下步骤:
(i)将根据本发明的第四个方面的细胞施用至受试者;
(ii)监控所述受试者的病理性免疫反应的形成;和
(iii)如果所述受试者显示正在形成或已经形成病理性免疫反应的体征,则向所述受试者施用雷帕霉素或雷帕霉素类似物。
本发明提供本发明的细胞,其用于治疗和/或预防疾病。
该细胞可以例如用于造血干细胞移植、淋巴细胞输注或过继细胞转移。
本发明还涉及本发明的细胞在制备用于治疗和/或预防疾病的药物中的用途。
本发明还提供用于治疗和/或预防毒性活性的cid剂,其能够诱导根据本发明的第一个方面的嵌合蛋白二聚化。
本发明还提供cid试剂,其用于活化细胞中的根据本发明的第一个方面的嵌合蛋白的胱天蛋白酶结构域对。
通过本发明的细胞和方法治疗和/或预防的疾病可以是感染,例如病毒感染。
本发明的方法还可以用于控制病理性免疫应答,例如在自身免疫性疾病、变态反应和移植物抗宿主排斥中。
在本发明的细胞表达tcr或car的情况下,它们可以用于治疗癌性疾病,例如膀胱癌、乳腺癌、结肠癌、子宫内膜癌、肾癌(肾细胞)、白血病、肺癌、黑色素瘤、非霍奇金淋巴瘤、胰腺癌、前列腺癌和甲状腺癌。
本发明的表达tcr/car的细胞可以能够杀伤靶细胞,例如癌细胞。
本发明还提供雷帕霉素或雷帕霉素类似物,用于预防或治疗受试者中由对受试者施用根据本发明的第四个方面的细胞引起的病理性免疫反应。
本发明的细胞可以用于任何细胞疗法,其中将经修饰或未修饰的细胞施用至患者。细胞治疗的一个例子是在cd34+干细胞移植后的过继性t细胞转移。干细胞转移后施用t细胞有助于加速患者受体免疫系统的重建。当匹配的相关或不相关的供体不可用时,或者该疾病对于广泛的供体搜索来说太具攻击性时,使用hla单倍体相同的(haploidentical)家族供体可以是有效的。此类供体可以是父母,兄弟姐妹或二级亲属。此类输注可以增强免疫恢复,并且从而降低病毒感染和消除复发性白血病细胞。然而,供体干细胞移植物中同种异体反应性(alloreactive)t细胞的共存可以导致抑制物抗宿主病(gvhd),其中供体细胞针对接受体起反应,这可以逐渐损伤接受体的皮肤、肠、肝和其它器官。
细胞疗法的其它实例包括使用天然(native)细胞或遗传工程改造以表达异源基因的细胞。这些治疗用于许多病症,包括血液病症,但这些治疗方法可以具有负面的副作用。在另一种方法中,可以分化成许多类型的成熟细胞的未成熟祖细胞(例如间充质基质细胞)可用于通过置换患病细胞的功能来治疗病症。本发明提供了快速且有效的机制,以去除细胞疗法中使用的供体细胞可能的负面影响。
本发明提供在供体t细胞移植后,降低人类患者中移植物抗宿主病的作用的方法,其包括用根据本发明的载体转导或转染供体细胞培养物中的人类供体t细胞;向患者施用转导或转染的t细胞;随后检测患者中存在或不存在移植物抗宿主病;以及向检测到移植物抗宿主病存在的患者施用二聚化化学诱导剂(cid)。t细胞可以是非同种异体耗竭的(non-allodepleted)。
本发明提供了干细胞移植的方法,包括向人类患者施用单倍体相同的干细胞移植物;以及向所述患者施用单倍体相同的供体t细胞,其中使用根据本发明的载体转染或转导单倍体相同的供体细胞培养物中的t细胞。
细胞可以是供体细胞培养物中的非同种异体耗竭的人类供体t细胞。
本发明还提供了干细胞移植的方法,包括向人类患者施用单倍体相同的干细胞移植物;以及向所述患者施用非同种异体耗竭的供体t细胞,其中使用根据本发明的载体转染或转导单倍体相同的供体细胞培养物中的t细胞。
单倍体相同的干细胞移植物可以是cd34+单倍体相同的干细胞移植物。人类供体t细胞可以与患者的t细胞单倍体相同的。患者可以患有可以通过干细胞移植减轻的任何疾病或病症。患者可以患有癌症,例如实体瘤或血液或骨髓的癌症。患者可以患有血液或骨髓疾病。患者可以患有镰状细胞性贫血或异染性脑白质营养不良。
可以从骨髓样品或外周血制备供体细胞培养物。可以从供体外周血单核细胞制备供体细胞培养物。在一些实施方案中,在转染或转导之前从供体细胞培养物同种异体耗竭供体t细胞。在施用至患者之前,转导或转染的t细胞可以在il-2的存在下培养。
将通过实施例的方式进一步描述本发明,这些实施例旨在帮助本领域普通技术人员实施本发明,且不意图以任何方式限制本发明的范围。
实施例
实施例1-产生表达嵌合蛋白的t细胞
使用不同的构建体转导t细胞。对于双分子rapcasp9(图1a),使用两种载体转导t细胞:一种编码通过内部核糖体进入位点的方式与绿色荧光蛋白egfp共表达的fkbp12-casp9,而另一种编码与蓝色荧光蛋白ebfp2共表达的frb-casp9。对于一分子rapcasp9(图1b),使用编码相应的与egfp共表达的rapcasp9的仅一种载体转导t细胞。提供fkb12-casp9和frb-frbw的构建体在三顺反子(tri-cistronic)基因盒中编码,其中使用fmd-2a样肽共表达fkb12-casp9和frb-frbw,且egfp与ires共表达。有意地仅部分转导t细胞,使得细胞培养物内,细胞的部分保持未转导以起内部阴性对照作用。作为进一步的对照,仅使用编码egfp的载体转导t细胞以排除雷帕霉素对转导的细胞的非特异性效果。
实施例2-测试使用雷帕霉素删除表达嵌合蛋白的细胞
将t细胞暴露不同浓度的雷帕霉素,并孵育48小时。此后,t细胞使用膜联蛋白-v和7add染色并通过流式细胞术分析。通过在活细胞上门控,和询问表达荧光蛋白的细胞,可以清楚地测量转导的和非转导的群体的存活。双重frb-casp9和fkbp12-casp9方法产生仅对双重阳性细胞的有效删除,如预期的。fkbp-frb-casp9构建体产生对单阳性细胞的有效删除。fkbp12-casp9-frb构建体产生最小删除。fkbp12-casp9/frb-frbw产生对单阳性细胞的有效删除。对照产生无特异性删除(图2和图3)。
实施例3-测试构建体的扩展组
生成了如图5中所示的构建体并转导至jurkat细胞中。将转导的细胞与非转导的(nt)细胞混合以在群体中具有两种构建体阳性和阴性的细胞。以0、1、10、100和1000nm的浓度添加雷帕霉素,并将细胞孵育24小时。收获后,细胞使用pi和膜联蛋白v染色并通过facs分析。结果示于图6至9并总结于图10中。
具有根据本发明的第一个方面的第一个实施方案定义的构造的构建体,即mp20244,在该测定中表现非常好,在高于并包括1nm的雷帕霉素浓度下产生对转导的细胞的非常有效的杀伤。
具有根据本发明的第一个方面的第二个实施方案定义的构造的构建体,即mp20206和mp20207,也表现非常好,在高于并包括1nm的雷帕霉素浓度下产生对转导的细胞的非常有效的杀伤。
具有根据本发明的第一个方面的第三个实施方案定义的构造的构建体,即mp20265,也表现良好,在1nm雷帕霉素下产生一些杀伤,并在10nm和以上的雷帕霉素浓度下产生有效杀伤。
具有根据本发明的第一个方面的第四个实施方案定义的构造的构建体,即mp20263、mp20264和mp21067在1nm雷帕霉素下表现良好,但在较高浓度的雷帕霉素下杀伤不太有效。
实施例4-使用替西罗莫司测试构建体
在与实施例3中描述的实验的等同实验中,使用雷帕霉素和替西罗莫司(雷帕霉素类似物)处理表达如图5中所示的构建体的细胞。
与实施例3中概述的实验一样,将转导的jurkat细胞与非转导(nt)混合,产生含有表达构建体的细胞和非转导的细胞两者的群体。
以2x105细胞每孔的浓度的细胞保持未处理,或使用以下浓度的雷帕霉素或替西罗莫司处理:0.01、0.1、1、10nm(雷帕霉素或替西罗莫司)。
细胞孵育24h,然后对于膜联蛋白v和pi染色并通过facs分析。结果如图11所示。
在替西罗莫司的存在下,使用图5中所示的多种构建体观察到了jurkat细胞杀伤的等同的模式,如先前已经在存在雷帕霉素的情况下观察到的。
特别地,具有根据本发明第一个方面的第一个实施方案定义的构造的构建体mp20244;和具有根据本发明的第一个方面的第二个实施方案定义的构造的构建体对mp20206和mp20207,两者均表现良好。两者都在高于和包括1nm的所有浓度的替西罗莫司下有效杀伤转染的细胞。
上述说明书中提到的所有出版物通过引用并入本文。在不脱离本发明的范围和精神的情况下,本发明所述的方法和系统的各种修改和变化对于本领域技术人员将是显而易见的。尽管已经结合具体的优选实施例描述了本发明,但是应当理解,所要求保护的本发明不应被不适当地限于这些具体实施例。实际上,对于分子生物学或相关领域的技术人员显而易见的用于实施本发明描述的模式的各种修改意图在所附权利要求的范围内。
序列表
<110>ucl商务股份有限公司
synconapartnersllp
<120>嵌合蛋白
<130>p106629pct
<150>gb1503133.9
<151>2015-02-24
<160>17
<170>patentinversion3.5
<210>1
<211>517
<212>prt
<213>人工序列
<220>
<223>嵌合蛋白frb-fkbp12-l3-dcasp9
<400>1
metalaserargileleutrphisglumettrphisgluglyleuglu
151015
glualaserargleutyrpheglygluargasnvallysglymetphe
202530
gluvalleugluproleuhisalametmetgluargglyproglnthr
354045
leulysgluthrserpheasnglnalatyrglyargaspleumetglu
505560
alaglnglutrpcysarglystyrmetlysserglyasnvallysasp
65707580
leuleuglnalatrpaspleutyrtyrhisvalpheargargileser
859095
lysleuglutyrserglyglyglyserleugluglyvalglnvalglu
100105110
thrileserproglyaspglyargthrpheprolysargglyglnthr
115120125
cysvalvalhistyrthrglymetleugluaspglylyslyspheasp
130135140
serserargaspargasnlysprophelysphemetleuglylysgln
145150155160
gluvalileargglytrpglugluglyvalalaglnmetservalgly
165170175
glnargalalysleuthrileserproasptyralatyrglyalathr
180185190
glyhisproglyileileproprohisalathrleuvalpheaspval
195200205
gluleuleulysleugluserglyglyglyglyserglyglyglygly
210215220
serglyglyglyglyserglyvalaspglypheglyaspvalglyala
225230235240
leugluserleuargglyasnalaaspleualatyrileleusermet
245250255
gluprocysglyhiscysleuileileasnasnvalasnphecysarg
260265270
gluserglyleuargthrargthrglyserasnileaspcysglulys
275280285
leuargargargpheserserleuhisphemetvalgluvallysgly
290295300
aspleuthralalyslysmetvalleualaleuleugluleualagln
305310315320
glnasphisglyalaleuaspcyscysvalvalvalileleuserhis
325330335
glycysglnalaserhisleuglnpheproglyalavaltyrglythr
340345350
aspglycysprovalservalglulysilevalasnilepheasngly
355360365
thrsercysproserleuglyglylysprolysleuphepheilegln
370375380
alacysglyglygluglnlysasphisglyphegluvalalaserthr
385390395400
serprogluaspgluserproglyserasnprogluproaspalathr
405410415
propheglngluglyleuargthrpheaspglnleuaspalaileser
420425430
serleuprothrproseraspilephevalsertyrserthrphepro
435440445
glyphevalsertrpargaspprolysserglysertrptyrvalglu
450455460
thrleuaspaspilephegluglntrpalahissergluaspleugln
465470475480
serleuleuleuargvalalaasnalavalservallysglyiletyr
485490495
lysglnmetproglycyspheasnpheleuarglyslysleuphephe
500505510
lysthrseralaser
515
<210>2
<211>402
<212>prt
<213>人工序列
<220>
<223>嵌合蛋白fkbp12-dcasp9
<400>2
metleugluglyvalglnvalgluthrileserproglyaspglyarg
151015
thrpheprolysargglyglnthrcysvalvalhistyrthrglymet
202530
leugluaspglylyslyspheaspserserargaspargasnlyspro
354045
phelysphemetleuglylysglngluvalileargglytrpgluglu
505560
glyvalalaglnmetservalglyglnargalalysleuthrileser
65707580
proasptyralatyrglyalathrglyhisproglyileilepropro
859095
hisalathrleuvalpheaspvalgluleuleulysleuglusergly
100105110
glyglyserglyvalaspglypheglyaspvalglyalaleugluser
115120125
leuargglyasnalaaspleualatyrileleusermetgluprocys
130135140
glyhiscysleuileileasnasnvalasnphecysargglusergly
145150155160
leuargthrargthrglyserasnileaspcysglulysleuargarg
165170175
argpheserserleuhisphemetvalgluvallysglyaspleuthr
180185190
alalyslysmetvalleualaleuleugluleualaglnglnasphis
195200205
glyalaleuaspcyscysvalvalvalileleuserhisglycysgln
210215220
alaserhisleuglnpheproglyalavaltyrglythraspglycys
225230235240
provalservalglulysilevalasnilepheasnglythrsercys
245250255
proserleuglyglylysprolysleuphepheileglnalacysgly
260265270
glygluglnlysasphisglyphegluvalalaserthrserproglu
275280285
aspgluserproglyserasnprogluproaspalathrprophegln
290295300
gluglyleuargthrpheaspglnleuaspalaileserserleupro
305310315320
thrproseraspilephevalsertyrserthrpheproglypheval
325330335
sertrpargaspprolysserglysertrptyrvalgluthrleuasp
340345350
aspilephegluglntrpalahissergluaspleuglnserleuleu
355360365
leuargvalalaasnalavalservallysglyiletyrlysglnmet
370375380
proglycyspheasnpheleuarglyslysleuphephelysthrser
385390395400
alaser
<210>3
<211>392
<212>prt
<213>人工序列
<220>
<223>嵌合蛋白frb-dcasp9
<400>3
metalaserargileleutrphisglumettrphisgluglyleuglu
151015
glualaserargleutyrpheglygluargasnvallysglymetphe
202530
gluvalleugluproleuhisalametmetgluargglyproglnthr
354045
leulysgluthrserpheasnglnalatyrglyargaspleumetglu
505560
alaglnglutrpcysarglystyrmetlysserglyasnvallysasp
65707580
leuleuglnalatrpaspleutyrtyrhisvalpheargargileser
859095
lysleuglutyrserglyglyglyserglyvalaspglypheglyasp
100105110
valglyalaleugluserleuargglyasnalaaspleualatyrile
115120125
leusermetgluprocysglyhiscysleuileileasnasnvalasn
130135140
phecysarggluserglyleuargthrargthrglyserasnileasp
145150155160
cysglulysleuargargargpheserserleuhisphemetvalglu
165170175
vallysglyaspleuthralalyslysmetvalleualaleuleuglu
180185190
leualaglnglnasphisglyalaleuaspcyscysvalvalvalile
195200205
leuserhisglycysglnalaserhisleuglnpheproglyalaval
210215220
tyrglythraspglycysprovalservalglulysilevalasnile
225230235240
pheasnglythrsercysproserleuglyglylysprolysleuphe
245250255
pheileglnalacysglyglygluglnlysasphisglyphegluval
260265270
alaserthrserprogluaspgluserproglyserasnproglupro
275280285
aspalathrpropheglngluglyleuargthrpheaspglnleuasp
290295300
alaileserserleuprothrproseraspilephevalsertyrser
305310315320
thrpheproglyphevalsertrpargaspprolysserglysertrp
325330335
tyrvalgluthrleuaspaspilephegluglntrpalahisserglu
340345350
aspleuglnserleuleuleuargvalalaasnalavalservallys
355360365
glyiletyrlysglnmetproglycyspheasnpheleuarglyslys
370375380
leuphephelysthrseralaser
385390
<210>4
<211>651
<212>prt
<213>人工序列
<220>
<223>嵌合蛋白fkbp12-dcasp9-2a-frb-frbw
<400>4
metleugluglyvalglnvalgluthrileserproglyaspglyarg
151015
thrpheprolysargglyglnthrcysvalvalhistyrthrglymet
202530
leugluaspglylyslyspheaspserserargaspargasnlyspro
354045
phelysphemetleuglylysglngluvalileargglytrpgluglu
505560
glyvalalaglnmetservalglyglnargalalysleuthrileser
65707580
proasptyralatyrglyalathrglyhisproglyileilepropro
859095
hisalathrleuvalpheaspvalgluleuleulysleuglusergly
100105110
glyglyserglyvalaspglypheglyaspvalglyalaleugluser
115120125
leuargglyasnalaaspleualatyrileleusermetgluprocys
130135140
glyhiscysleuileileasnasnvalasnphecysargglusergly
145150155160
leuargthrargthrglyserasnileaspcysglulysleuargarg
165170175
argpheserserleuhisphemetvalgluvallysglyaspleuthr
180185190
alalyslysmetvalleualaleuleugluleualaglnglnasphis
195200205
glyalaleuaspcyscysvalvalvalileleuserhisglycysgln
210215220
alaserhisleuglnpheproglyalavaltyrglythraspglycys
225230235240
provalservalglulysilevalasnilepheasnglythrsercys
245250255
proserleuglyglylysprolysleuphepheileglnalacysgly
260265270
glygluglnlysasphisglyphegluvalalaserthrserproglu
275280285
aspgluserproglyserasnprogluproaspalathrprophegln
290295300
gluglyleuargthrpheaspglnleuaspalaileserserleupro
305310315320
thrproseraspilephevalsertyrserthrpheproglypheval
325330335
sertrpargaspprolysserglysertrptyrvalgluthrleuasp
340345350
aspilephegluglntrpalahissergluaspleuglnserleuleu
355360365
leuargvalalaasnalavalservallysglyiletyrlysglnmet
370375380
proglycyspheasnpheleuarglyslysleuphephelysthrser
385390395400
alaserglncysthrasntyralaleuleulysleualaglyaspval
405410415
gluserasnproglyproglyvalglnvalgluthrileserprogly
420425430
aspglyargthrpheprolysargglyglnthrcysvalvalhistyr
435440445
thrglymetleugluaspglylyslysvalaspserserargasparg
450455460
asnlysprophelysphemetleuglylysglngluvalilearggly
465470475480
trpglugluglyvalalaglnmetservalglyglnargalalysleu
485490495
thrileserproasptyralatyrglyalathrglyhisproglyile
500505510
ileproprohisalathrleuvalpheaspvalgluleuleulysleu
515520525
gluserglyglyglyglyserglyglyglyglysermetleuglugly
530535540
valglnvalgluthrileserproglyaspglyargthrpheprolys
545550555560
argglyglnthrcysvalvalhistyrthrglymetleugluaspgly
565570575
lyslyspheaspserserargaspargasnlysprophelysphemet
580585590
leuglylysglngluvalileargglytrpglugluglyvalalagln
595600605
metservalglyglnargalalysleuthrileserproasptyrala
610615620
tyrglyalathrglyhisproglyileileproprohisalathrleu
625630635640
valpheaspvalgluleuleulysleugluser
645650
<210>5
<211>416
<212>prt
<213>人类
<400>5
metaspglualaaspargargleuleuargargcysargleuargleu
151015
valglugluleuglnvalaspglnleutrpaspalaleuleuserser
202530
gluleupheargprohismetilegluaspileglnargalaglyser
354045
glyserargargaspglnalaargglnleuileileaspleugluthr
505560
argglyserglnalaleuproleupheilesercysleugluaspthr
65707580
glyglnaspmetleualaserpheleuargthrasnargglnalaala
859095
lysleuserlysprothrleugluasnleuthrprovalvalleuarg
100105110
progluilearglysprogluvalleuargprogluthrproargpro
115120125
valaspileglyserglyglypheglyaspvalglyalaleugluser
130135140
leuargglyasnalaaspleualatyrileleusermetgluprocys
145150155160
glyhiscysleuileileasnasnvalasnphecysargglusergly
165170175
leuargthrargthrglyserasnileaspcysglulysleuargarg
180185190
argpheserserprohisphemetvalgluvallysglyaspleuthr
195200205
alalyslysmetvalleualaleuleugluleualaglnglnasphis
210215220
glyalaleuaspcyscysvalvalvalileleuserhisglycysgln
225230235240
alaserhisleuglnpheproglyalavaltyrglythraspglycys
245250255
provalservalglulysilevalasnilepheasnglythrsercys
260265270
proserleuglyglylysprolysleuphepheileglnalacysgly
275280285
glygluglnlysasphisglyphegluvalalaserthrserproglu
290295300
aspgluserproglyserasnprogluproaspalathrprophegln
305310315320
gluglyleuargthrpheaspglnleuaspalaileserserleupro
325330335
thrproseraspilephevalsertyrserthrpheproglypheval
340345350
sertrpargaspprolysserglysertrptyrvalgluthrleuasp
355360365
aspilephegluglntrpalahissergluaspleuglnserleuleu
370375380
leuargvalalaasnalavalservallysglyiletyrlysglnmet
385390395400
proglycyspheasnpheleuarglyslysleuphephelysthrser
405410415
<210>6
<211>282
<212>prt
<213>人类
<400>6
glypheglyaspvalglyalaleugluserleuargglyasnalaasp
151015
leualatyrileleusermetgluprocysglyhiscysleuileile
202530
asnasnvalasnphecysarggluserglyleuargthrargthrgly
354045
serasnileaspcysglulysleuargargargpheserserleuhis
505560
phemetvalgluvallysglyaspleuthralalyslysmetvalleu
65707580
alaleuleugluleualaglnglnasphisglyalaleuaspcyscys
859095
valvalvalileleuserhisglycysglnalaserhisleuglnphe
100105110
proglyalavaltyrglythraspglycysprovalservalglulys
115120125
ilevalasnilepheasnglythrsercysproserleuglyglylys
130135140
prolysleuphepheileglnalacysglyglygluglnlysasphis
145150155160
glyphegluvalalaserthrserprogluaspgluserproglyser
165170175
asnprogluproaspalathrpropheglngluglyleuargthrphe
180185190
aspglnleuaspalaileserserleuprothrproseraspilephe
195200205
valsertyrserthrpheproglyphevalsertrpargaspprolys
210215220
serglysertrptyrvalgluthrleuaspaspilephegluglntrp
225230235240
alahissergluaspleuglnserleuleuleuargvalalaasnala
245250255
valservallysglyiletyrlysglnmetproglycyspheasnphe
260265270
leuarglyslysleuphephelysthrser
275280
<210>7
<211>108
<212>prt
<213>人工序列
<220>
<223>fkbp12domain
<400>7
metglyvalglnvalgluthrileserproglyaspglyargthrphe
151015
prolysargglyglnthrcysvalvalhistyrthrglymetleuglu
202530
aspglylyslyspheaspserserargaspargasnlysprophelys
354045
phemetleuglylysglngluvalileargglytrpglugluglyval
505560
alaglnmetservalglyglnargalalysleuthrileserproasp
65707580
tyralatyrglyalathrglyhisproglyileileproprohisala
859095
thrleuvalpheaspvalgluleuleulysleuglu
100105
<210>8
<211>100
<212>prt
<213>人工序列
<220>
<223>mtor的野生型frb区段
<400>8
metalaserargileleutrphisglumettrphisgluglyleuglu
151015
glualaserargleutyrpheglygluargasnvallysglymetphe
202530
gluvalleugluproleuhisalametmetgluargglyproglnthr
354045
leulysgluthrserpheasnglnalatyrglyargaspleumetglu
505560
alaglnglutrpcysarglystyrmetlysserglyasnvallysasp
65707580
leuthrglnalatrpaspleutyrtyrhisvalpheargargileser
859095
lysleugluser
100
<210>9
<211>100
<212>prt
<213>人工序列
<220>
<223>具有允许与ap21967结合的2098处t至l取代的frb
<400>9
metalaserargileleutrphisglumettrphisgluglyleuglu
151015
glualaserargleutyrpheglygluargasnvallysglymetphe
202530
gluvalleugluproleuhisalametmetgluargglyproglnthr
354045
leulysgluthrserpheasnglnalatyrglyargaspleumetglu
505560
alaglnglutrpcysarglystyrmetlysserglyasnvallysasp
65707580
leuleuglnalatrpaspleutyrtyrhisvalpheargargileser
859095
lysleugluser
100
<210>10
<211>100
<212>prt
<213>人工序列
<220>
<223>在全长mtor的2098处具有t至h取代和残基2101处的f取代为w的mtor的frb区段
<400>10
metalaserargileleutrphisglumettrphisgluglyleuglu
151015
glualaserargleutyrpheglygluargasnvallysglymetphe
202530
gluvalleugluproleuhisalametmetgluargglyproglnthr
354045
leulysgluthrserpheasnglnalatyrglyargaspleumetglu
505560
alaglnglutrpcysarglystyrmetlysserglyasnvallysasp
65707580
leuhisglnalapheaspleutyrtyrhisvalpheargargileser
859095
lysleugluser
100
<210>11
<211>100
<212>prt
<213>人工序列
<220>
<223>在全长mtor的残基2095处具有k至p取代的mtor的frb区段
<400>11
metalaserargileleutrphisglumettrphisgluglyleuglu
151015
glualaserargleutyrpheglygluargasnvallysglymetphe
202530
gluvalleugluproleuhisalametmetgluargglyproglnthr
354045
leulysgluthrserpheasnglnalatyrglyargaspleumetglu
505560
alaglnglutrpcysarglystyrmetlysserglyasnvalproasp
65707580
leuthrglnalatrpaspleutyrtyrhisvalpheargargileser
859095
lysleugluser
100
<210>12
<211>1554
<212>dna
<213>人工序列
<220>
<223>frb-fkbp12-l3-casp9核苷酸序列
<400>12
atggcttctagaatcctctggcatgagatgtggcatgaaggcctggaagaggcatctcgt60
ttgtactttggggaaaggaacgtgaaaggcatgtttgaggtgctggagcccttgcatgct120
atgatggaacggggcccccagactctgaaggaaacatcctttaatcaggcctatggtcga180
gatttaatggaggcccaagagtggtgcaggaagtacatgaaatcagggaatgtcaaggac240
ctcctccaagcctgggacctctattatcatgtgttccgacgaatctcaaagctcgagtat300
agcggcggcggcagcctggagggcgtgcaggtggagaccatcagcccaggcgacggcaga360
accttccccaagagaggccagacctgcgtggtgcactataccggcatgctggaggacggc420
aagaagttcgacagcagccgcgaccgcaataagcccttcaagttcatgctgggcaagcag480
gaggtgatcagaggctgggaggagggcgtggcccagatgagcgtgggccagagagccaag540
ctgaccatcagccccgactacgcctatggcgccaccggccaccccggcatcatcccaccc600
cacgccaccctggtgtttgatgtggagctgctgaagctggagtccggcggaggcgggtct660
ggaggaggcggcagcggcggcggcgggtcaggcgtggatggcttcggcgacgtgggagcc720
ctggagagcctgagaggcaacgccgatctggcctacatcctgagcatggagccctgtggc780
cactgcctgatcatcaacaacgtgaacttctgccgggagagcggcctgcggacccggacc840
ggcagcaacatcgactgcgagaagctgaggaggcgcttctcctccctgcactttatggtg900
gaggtgaaaggcgatctgactgccaagaaaatggtgctggccctgctggagctggcccag960
caggaccacggagccctggattgctgtgtggtggtgatcctgtcccacggctgccaggcc1020
agccacctgcagttccccggagccgtgtacggcaccgacggctgtcccgtgtccgtggag1080
aagatcgtgaacatcttcaacggcacctcctgcccctccctgggcggcaagcccaagctg1140
ttctttatccaggcctgtggcggcgagcagaaggaccacggctttgaggtggccagcacc1200
tcccccgaggacgagagcccaggcagcaaccccgagcccgacgccacccccttccaggag1260
ggcctgcgcaccttcgaccagctggacgccatcagcagcctgcccacccccagcgacatc1320
ttcgtgagctacagcacctttcccggcttcgtgagctggcgcgatcccaagtccggctct1380
tggtatgtggagaccctggacgacatctttgagcagtgggctcatagcgaggacctgcag1440
agcctgctgctgcgcgtggccaatgccgtgagcgtgaagggcatctacaagcagatgcca1500
ggctgcttcaacttcctgcggaagaagctgttcttcaagaccagcgcctcctga1554
<210>13
<211>1209
<212>dna
<213>人工序列
<220>
<223>fkbp12-dcasp9核苷酸序列
<400>13
atgctggagggcgtgcaggtggagaccatcagcccaggcgacggcagaaccttccccaag60
agaggccagacctgcgtggtgcactataccggcatgctggaggacggcaagaagttcgac120
agcagccgcgaccgcaataagcccttcaagttcatgctgggcaagcaggaggtgatcaga180
ggctgggaggagggcgtggcccagatgagcgtgggccagagagccaagctgaccatcagc240
cccgactacgcctatggcgccaccggccaccccggcatcatcccaccccacgccaccctg300
gtgtttgatgtggagctgctgaagctggagtccggaggcggctccggcgtggatggcttc360
ggcgacgtgggagccctggagagcctgagaggcaacgccgatctggcctacatcctgagc420
atggagccctgtggccactgcctgatcatcaacaacgtgaacttctgccgggagagcggc480
ctgcggacccggaccggcagcaacatcgactgcgagaagctgaggaggcgcttctcctcc540
ctgcactttatggtggaggtgaaaggcgatctgactgccaagaaaatggtgctggccctg600
ctggagctggcccagcaggaccacggagccctggattgctgtgtggtggtgatcctgtcc660
cacggctgccaggccagccacctgcagttccccggagccgtgtacggcaccgacggctgt720
cccgtgtccgtggagaagatcgtgaacatcttcaacggcacctcctgcccctccctgggc780
ggcaagcccaagctgttctttatccaggcctgtggcggcgagcagaaggaccacggcttt840
gaggtggccagcacctcccccgaggacgagagcccaggcagcaaccccgagcccgacgcc900
acccccttccaggagggcctgcgcaccttcgaccagctggacgccatcagcagcctgccc960
acccccagcgacatcttcgtgagctacagcacctttcccggcttcgtgagctggcgcgat1020
cccaagtccggctcttggtatgtggagaccctggacgacatctttgagcagtgggctcat1080
agcgaggacctgcagagcctgctgctgcgcgtggccaatgccgtgagcgtgaagggcatc1140
tacaagcagatgccaggctgcttcaacttcctgcggaagaagctgttcttcaagaccagc1200
gcctcctga1209
<210>14
<211>1179
<212>dna
<213>人工序列
<220>
<223>frb-dcasp9核苷酸序列
<400>14
atggcttctagaatcctctggcatgagatgtggcatgaaggcctggaagaggcatctcgt60
ttgtactttggggaaaggaacgtgaaaggcatgtttgaggtgctggagcccttgcatgct120
atgatggaacggggcccccagactctgaaggaaacatcctttaatcaggcctatggtcga180
gatttaatggaggcccaagagtggtgcaggaagtacatgaaatcagggaatgtcaaggac240
ctcctccaagcctgggacctctattatcatgtgttccgacgaatctcaaagctcgagtat300
agcggcggcggcagcggcgtggatggcttcggcgacgtgggagccctggagagcctgaga360
ggcaacgccgatctggcctacatcctgagcatggagccctgtggccactgcctgatcatc420
aacaacgtgaacttctgccgggagagcggcctgcggacccggaccggcagcaacatcgac480
tgcgagaagctgaggaggcgcttctcctccctgcactttatggtggaggtgaaaggcgat540
ctgactgccaagaaaatggtgctggccctgctggagctggcccagcaggaccacggagcc600
ctggattgctgtgtggtggtgatcctgtcccacggctgccaggccagccacctgcagttc660
cccggagccgtgtacggcaccgacggctgtcccgtgtccgtggagaagatcgtgaacatc720
ttcaacggcacctcctgcccctccctgggcggcaagcccaagctgttctttatccaggcc780
tgtggcggcgagcagaaggaccacggctttgaggtggccagcacctcccccgaggacgag840
agcccaggcagcaaccccgagcccgacgccacccccttccaggagggcctgcgcaccttc900
gaccagctggacgccatcagcagcctgcccacccccagcgacatcttcgtgagctacagc960
acctttcccggcttcgtgagctggcgcgatcccaagtccggctcttggtatgtggagacc1020
ctggacgacatctttgagcagtgggctcatagcgaggacctgcagagcctgctgctgcgc1080
gtggccaatgccgtgagcgtgaagggcatctacaagcagatgccaggctgcttcaacttc1140
ctgcggaagaagctgttcttcaagaccagcgcctcctga1179
<210>15
<211>1956
<212>dna
<213>人工序列
<220>
<223>fkbp12-casp9-2a-frb-frbw核苷酸序列
<400>15
atgctggagggcgtgcaggtggagaccatcagcccaggcgacggcagaaccttccccaag60
agaggccagacctgcgtggtgcactataccggcatgctggaggacggcaagaagttcgac120
agcagccgcgaccgcaataagcccttcaagttcatgctgggcaagcaggaggtgatcaga180
ggctgggaggagggcgtggcccagatgagcgtgggccagagagccaagctgaccatcagc240
cccgactacgcctatggcgccaccggccaccccggcatcatcccaccccacgccaccctg300
gtgtttgatgtggagctgctgaagctggagtccggaggcggctccggcgtggatggcttc360
ggcgacgtgggagccctggagagcctgagaggcaacgccgatctggcctacatcctgagc420
atggagccctgtggccactgcctgatcatcaacaacgtgaacttctgccgggagagcggc480
ctgcggacccggaccggcagcaacatcgactgcgagaagctgaggaggcgcttctcctcc540
ctgcactttatggtggaggtgaaaggcgatctgactgccaagaaaatggtgctggccctg600
ctggagctggcccagcaggaccacggagccctggattgctgtgtggtggtgatcctgtcc660
cacggctgccaggccagccacctgcagttccccggagccgtgtacggcaccgacggctgt720
cccgtgtccgtggagaagatcgtgaacatcttcaacggcacctcctgcccctccctgggc780
ggcaagcccaagctgttctttatccaggcctgtggcggcgagcagaaggaccacggcttt840
gaggtggccagcacctcccccgaggacgagagcccaggcagcaaccccgagcccgacgcc900
acccccttccaggagggcctgcgcaccttcgaccagctggacgccatcagcagcctgccc960
acccccagcgacatcttcgtgagctacagcacctttcccggcttcgtgagctggcgcgat1020
cccaagtccggctcttggtatgtggagaccctggacgacatctttgagcagtgggctcat1080
agcgaggacctgcagagcctgctgctgcgcgtggccaatgccgtgagcgtgaagggcatc1140
tacaagcagatgccaggctgcttcaacttcctgcggaagaagctgttcttcaagaccagc1200
gcctcccagtgcaccaattatgctttgcttaagctggcaggcgatgtggaatcaaacccg1260
ggtcctggggtacaggtggagaccatctctcctggcgacgggagaacatttcctaaaagg1320
ggccaaacatgcgtggttcactataccggtatgctggaggatggcaaaaaagtagactcc1380
agccgggatagaaacaaaccctttaagttcatgctgggtaagcaggaagttatacggggc1440
tgggaagagggagtagctcagatgtctgtgggccagagggccaagctgaccatctcaccg1500
gactacgcctacggcgctaccggccaccctggcattataccaccccatgcaactctcgtc1560
ttcgatgttgagttgctcaaactggaatcaggcggaggcgggtctggaggaggcggcagc1620
atgctggagggcgtgcaggtggagaccatcagcccaggcgacggcagaaccttccccaag1680
agaggccagacctgcgtggtgcactataccggcatgctggaggacggcaagaagttcgac1740
agcagccgcgaccgcaataagcccttcaagttcatgctgggcaagcaggaggtgatcaga1800
ggctgggaggagggcgtggcccagatgagcgtgggccagagagccaagctgaccatcagc1860
cccgactacgcctatggcgccaccggccaccccggcatcatcccaccccacgccaccctg1920
gtgtttgatgtggagctgctgaagctggagtcctga1956
<210>16
<211>20
<212>prt
<213>口蹄疫病毒
<400>16
argalagluglyargglyserleuleuthrcysglyaspvalgluglu
151015
asnproglypro
20
<210>17
<211>20
<212>prt
<213>口蹄疫病毒
<400>17
glncysthrasntyralaleuleulysleualaglyaspvalgluser
151015
asnproglypro
20
- 还没有人留言评论。精彩留言会获得点赞!